
Physical, chemical and sensory properties of cassava ... - CORE
Upload
independentCategory
view
2download
0
Cpi
Wa
b
a
ARRAA
KPCPTI
1
wdpvn[tSipahs
sb4d
1h
Process Biochemistry 48 (2013) 1516–1523
Contents lists available at ScienceDirect
Process Biochemistry
jo u r n al homep age: www.elsev ier .com/ locate /procbio
assava (Manihot esculenta Crantz of cv. KU50) peroxidase and itsotential for the detection of some thiol compounds based on the
nhibitory effect of 3,3′,5,5′-tetramethylbenzidine oxidation
ischada Jongmevasnaa, Surachai Yaiyena, Manchumas Hengsakul Prousoontornb,∗
Program in Biotechnology, Faculty of Science, Chulalongkorn University, Bangkok 10330, ThailandDepartment of Biochemistry, Faculty of Science, Chulalongkorn University, Bangkok 10330, Thailand
r t i c l e i n f o
rticle history:eceived 25 March 2013eceived in revised form 26 June 2013ccepted 17 July 2013vailable online 7 August 2013
eywords:eroxidase
a b s t r a c t
Cassava peroxidase (CSP) isolated from cassava leaves (Manihot esculenta Crantz of cv. KU50) was purifiedby DEAE and concanavalin-A column chromatography. CSP was a haem-containing cationic glycoproteinwith the molecular weight of 38 and 44 kDa determined by MALDI-TOF-MS. Its kinetic catalysis in thepresence and absence of some thiols were investigated and compared to those of horseradish and soybeanperoxidases by using urea hydrogen peroxide (UHP) and 3,3′,5,5′-tetramethylbenzidine (TMB) as sub-strates. Inhibitory effects of some pesticides containing thiol groups such as thiosemicabazide, thioureaand thiophanate-methyl on TMB oxidation were graphically demonstrated their ability to be either com-
assavaurificationhiolsnhibition
petitive or noncompetitive inhibitors, depending on their properties. The degrees of inhibition wereexpressed as Ki and IC50 values. The applicable range for the detection of thiosemicabazide in solutionwas found to be in the range of 10–100 �M, whereas those of thiourea and thiophanate-methyl were40–400 �M and 10–460 �M, respectively. On the contrary, the inhibitory effects of some phenolic pol-lutants; 4-nitrophenol, 4-phenylphenol and pentachlorophenol on TMB oxidation were not significantlyobserved and the phenomenon was discussed.
. Introduction
Peroxidases are classified as oxidoreductases (EC 1.11.1.7)hich belong to a large family of enzymes that catalyze the oxi-ation of various organic and inorganic substances by hydrogeneroxide or related compounds. They are commonly found in aariety of isomers in plants and play an important role in mainte-ance of reduced state molecules in cell and in defense mechanism1,2]. These isoenzymes were differentially expressed in variousissues and organs depending on environmental conditions [3,4].everal oxidoreductases have been explored for both analytical andndustrial applications. However, the well-known classical planteroxidase is horseradish peroxidase (HRP) due to its relative high
ctivity and stability with a wide range of substrates available. Itas been found to be well suited in many analytical applicationsuch as immunohistology and enzyme amplified immunoassayAbbreviations: CSP, cassava peroxidase; HRP, horseradish peroxidase; SBP,oybean peroxidase; THU, thiourea; ETU, ethylenethiourea; TMB, tetramethyl-enzidine; TPM, thiophanate-methyl; TSC, thiosemicabazide; PHE, phenol; 4-PP,-phenylphenol; PCP, pentachlorophenol and 4-NP, 4-nitrophenol; DAB, 3,3′-iaminebenzidine; UHP, urea hydrogen peroxide.∗ Corresponding author. Tel.: +66 2 2185437; fax: +66 2 2185418.
E-mail address: [email protected] (M.H. Prousoontorn).
359-5113/$ – see front matter © 2013 Elsevier Ltd. All rights reserved.ttp://dx.doi.org/10.1016/j.procbio.2013.07.014
© 2013 Elsevier Ltd. All rights reserved.
system for the quantitative determination of trace substancesbased on ELISA (enzyme-linked immunosorbent assay) as a valu-able tool in biotechnology [5]. In electrochemical analysis, HRP isthe most widely used enzyme for the construction of biosensors forthe determination of hydrogen peroxide [6,7] as well as the detec-tion of peroxidase inhibitors such as sulfides [8] and thiols [9]. Inaddition, the HRP catalyzed reaction also has a potential applica-tion in the removal of phenolic pollutants from waste water [10,11].However, the main drawback for HRP application is its low sta-bility with respect to hydrogen peroxide and acid concentration[12]. Therefore, alternative plant peroxidases with higher stabilityand/or different specificity have been investigated to improve theirutilization in analytical kits and in the construction of biosensors.Peroxidase from various plant sources such as soybean peroxidase(SBP) from seed coat is commercially available and has been usedas an alternative source of available peroxidase [13]. Furthermoreperoxidase activities were also screened in a wild endemic hemi-parasitic plant (Viscum angulatum) [14], artichoke leaves [15] andtree legume stem [16].
Cassava (Manihot esculenta Crantz) is an economic crop of
Thailand which is the major supplier of cassava for feed, food, andindustrial uses to the world market [17]. There have been somereports describing that peroxidase and ATPase have a role in deteri-oration of cassava tuber during postharvest [18]. Peroxidases could
Bioch
blhctipfbw
2
2
ra(4fPSc(wR
2
2
p0dam
2
caD5tbawcaf2
2
csHbw((TnCbtsatTl
2
go4
W. Jongmevasna et al. / Process
e found in many parts of cassava such as root, stem, petiole andeaf which were considered as agricultural waste after cassava rootarvest. Therefore, our work focused on the extraction and purifi-ation of peroxidase from cassava leaves and the influence of somehiol pesticides and phenolic pollutants toward its catalytic activ-ty was investigated and compared to those of HRP and SBP. Theossible use of cassava peroxidase (CSP) in analytical applicationor the detection of some thiol pesticides and phenolic pollutantsased on the inhibition of 3,3′,5,5′-tetramethylbenzidine oxidationas also explored.
. Materials and methods
.1. Materials
Cassava leaves of cultivar KU50 were collected from Rayong field cropsesearch center at Rayong province, Thailand. Horseradish peroxidase (Type II)nd soybean peroxidase were commercial peroxidases purchased from SigmaU.S.A.). Concanavalin-A, 3,3′-diaminebenzidine (DAB), DEAE-cellulose, sepharoseB, 3,3′ ,5,5′-tetramethylbenzidine (TMB) and urea hydrogen peroxide (UHP) wererom Sigma (U.S.A.) and ammonium sulfate was from Fluka (Buchs, Switzerland).rotein molecular weight and pI markers were from GE healthcare (Uppsala,weden). Most of the chemical reagents used for inhibition study were of analyti-al grade. Thiourea (THU), ethylenethiourea (ETU), thiosemicabazide (TSC), phenolPHE), 4-phenylphenol (4-PP), 4-nitrophenol (4-NP) and pentachlorophenol (PCP)ere from Sigma (U.S.A.). Thiophanate-methyl (TPM) was a pesticide standard fromiëdel-dë Haën (Germany).
.2. Methods
.2.1. Preparation of cassava leaf extractFive hundred grams of fresh cassava leaves were washed and chopped into small
ieces before blending in cold 50 mM sodium phosphate buffer, pH 6.5 containing.1 M phenylmethylsulfonyl fluoride (PMSF) and 2% (w/v) of polyvinyl polypyrroli-one (PVPP) with the ratio of 1:1 (w/v). The homogenate was filtered and centrifugedt 12,000 × g for 30 min at 4 ◦C. The peroxidase activity in the supernatants waseasured and collected for further purification.
.2.2. Purification of cassava peroxidaseThe crude enzyme was fractionated by ammonium sulfate precipitation in the
oncentration range of 40–80% (w/v). The precipitate collected was then resolvednd dialyzed against 50 mM sodium phosphate buffer pH 6.5 before subjected toEAE-cellulose column chromatography. The column was pre-equilibrated with0 mM sodium phosphate buffer pH 6.5. The unbound proteins were washed withhe same buffer and the bound protein fractions were collected with the sameuffer containing 0.5 M NaCl. The fractions with peroxidase activity were poolednd further purified by Concanavalin-A column chromatography. The bound portionas completely eluted by the addition of 50 mM sodium phosphate buffer pH 6.5
ontaining 0.5 M �-d-methyl glucopyranoside over 10 column volume. Peroxidasectivity was investigated in all fractions using TMB and UHP as substrates and eachraction was tested for protein and haem content by measuring the absorbance at80 and 405 nm, respectively.
.2.3. Determination of molecular weightThe molecular weight of purified enzyme obtained from bound Concanavalin-A
olumn was estimated by sodium dodecylsulfate-polyacrylamide gel electrophore-is (SDS-PAGE) on a mini Protean III Electrophoresis Cell (Bio-Rad, Laboratories,ercules, MA, U.S.A.), with 12% resolving gel and 3% stacking gel usinguffer system according to Laemmli [19]. The molecular weight markers usedere �-galactosidase (116 kDa), bovine serum albumin (66.2 kDa), ovalbumin
45 kDa), lactate dehydrogenase (35 kDa), �-lactoglobulin (18.4 kDa) and lysozyme14.4 kDa). Both markers and enzyme were mixed with 5× sample buffer (0.3 mMris–HCl, 50% glycerol, 20% SDS, 5% 2-mercaptoethanol and 0.05% bromophe-ol blue) and boiled for 5 min. After electrophoresis, proteins were stained withoomasie brilliant blue. In addition, its molecular weight was also determinedy Matrix Assisted Laser Desorption Ionization-Time of Flight-Mass Spectrome-ry (MALDI-TOF-MS). One microliter of sample was mixed with 10 �L of saturatedinapinic acid in 0.1% TFA:ACN (70:30). Then 1 �L of the mixture was spotted on
stainless steel target and allowed to dry at room temperature. The dried mix-ure was measured on an AutoflexTOF/TOF (Bruker Daltonik, Bremem, Germany).he instrument was equipped with a N2-laser and all measurements were done ininear mode.
.2.4. Determination of iso-electric focusing point (IEF)The pI values of purified CSP, HRP and SBP were examined using a Bio-Rad mini
el IEF. The method and applied potential condition based on the instruction manualf mini IEF cell model 111 was carried out (100 V for 20 min, 200 V for 20 min and50 V for 90 min). After electrophoresis, the standard pI lane was cut and immersed
emistry 48 (2013) 1516–1523 1517
in the fixative solution for 30 min prior to protein staining. For activity staining, theperoxidase sample lane was cut and stained with the solution containing 0.1 mg/mLof DAB and 0.1 mg/mL of UHP as substrates in acetate-citric acid buffer, pH 6 [20]and isoelectric points were determined by comparing with IEF markers (IEF mix3.6–9.3, Sigma, St. Louis, USA).
2.2.5. Determination of peroxidase activity and total proteinPeroxidase activity was investigated by the use of TMB as a chromogenic sub-
strate. The method was slightly modified from Bos et al. [21]. Enzyme solution, 10 �L,was added into microtiter well, followed by the addition of 250 �L of substrate solu-tion containing 0.1 mg/mL of TMB and 0.1 mg/mL of UHP in 0.1 M acetate–citric acidbuffer, pH 6. The reaction was incubated for 3 min and stopped by the addition of40 �L of 2 M sulfuric acid. The absorbance at 450 nm was then measured by TitertekMultiscan plus plate reader (Labsystems Ins., Finland). One unit of enzyme is theamount of enzyme, which oxidizes 1 �mol of TMB per minute (The molar extinc-tion coefficient was 67,300 M−1 cm−1). Total protein content was determined byBradford [22], using bovine serum albumin as a standard.
2.2.6. Determination of pH optimumThe optimal pH of the purified CSP and both of commercial HRP and SBP for
the catalysis of substrates containing UHP and TMB in the pH range of 3–10 wereinvestigated. The assay was carried out at room temperature and the buffers usedwere 0.1 M acetatate–citric acid (pH 3.0–6.0), 0.1 M phosphate (5.0–8.0), Tris–HCl(8.0–9.0) and Tris–glycine (8.0–10.0). The enzyme activity was assayed as describedpreviously.
2.2.7. Determination of kinetic parametersThe Michaelis–Menten constant (KM) and maximum velocity (Vmax) were graph-
ically determined by Lineweaver–Burk and secondary plots [23]. Due to the fact thatthe catalysis of peroxidase is a bi-substrate enzymatic reaction, the KM for UHP wasfirst determined by using various concentration of UHP in the range of 28–228 �Munder 4 fixed concentrations of TMB in the range of 32–260 �M in the presence ofperoxidase (3 units) in the reaction. CSP activities in reaction buffer were carriedout and compared to those of HRP and SBP under the same condition. Enzyme solu-tion, 2 �L, was added into microtiter well and followed by the addition of 100 �Lof 0.1 M acetate–citric acid buffer, pH 6 and 100 �L of TMB solution. The reactionwas initiated by the addition of 50 �L UHP solution and the change in absorption at650 nm (�A/min) due to the oxidation of TMB was recorded during 3 min with 30 sinterval using the microtiter plate reader. The initial rate was the mean of triplicatedetermination and calculated using the molar absorption coefficient of its product,39,000 M−1 cm−1 [21].
2.2.8. Inhibitory effects of some thiol and phenol compoundsInfluences of some thiols (THU, ETU, TPM and TSC) and phenols (PHE, 4-PP, 4-NP
and PCP) on initial rates of CSP, HRP and SBP were examined. The concentration ofUHP and TMB were kept constant at their KM and the inhibitor concentrations werevaried from 1 to 500 �M. All inhibitor compounds were prepared in methanol asstock solution and diluted to have certain concentration with 0.1 M acetate–citricacid buffer, pH 6 containing 10% of methanol. The assay was carried out as thedetermination of kinetic parameters except that 100 �L of inhibitor solution wasused instead of 100 �L of 0.1 M acetate–citric acid buffer. The decrease in activity (%)was then plotted against inhibitor concentration in �M. Their dissociation constants(Ki) and inhibition mechanisms were graphically determined by secondary plot ofappKM . Half-maximal inhibitory concentration (IC50) values were also determined bydose–response curve [23].
3. Results and discussion
3.1. Preparation and characteristics of CSP
Peroxidase was successfully extracted from cassava leaves ascrude with approximately 190 units/g of leaf and the purificationtable of CSP is given in Table 1. After ammonium sulfate fraction-ation and DEAE-cellulose column chromatography, the unboundportion with peroxidase activity was observed as a major proteinwith the specific activity of 1200 units/mg protein. It was furtherpurified by Concanavalin-A affinity column which specificallybinds molecules containing �-d-mannopyranosyl and �-d-glucopyranosyl residues [24]. The bound portion was desorbedwith buffer containing �-d-glucopyranoside. After sequentialpurifications, CSP was purified to 15 folds with the overall protein
yield of 4% and the specific activity was found to be 8200 units/mgprotein. Comparative study of biochemical characteristics ofall three peroxidases (CSP, HRP and SBP) were investigated andsummarized in Table 2. Their RZ (Reinheitszahl) values which were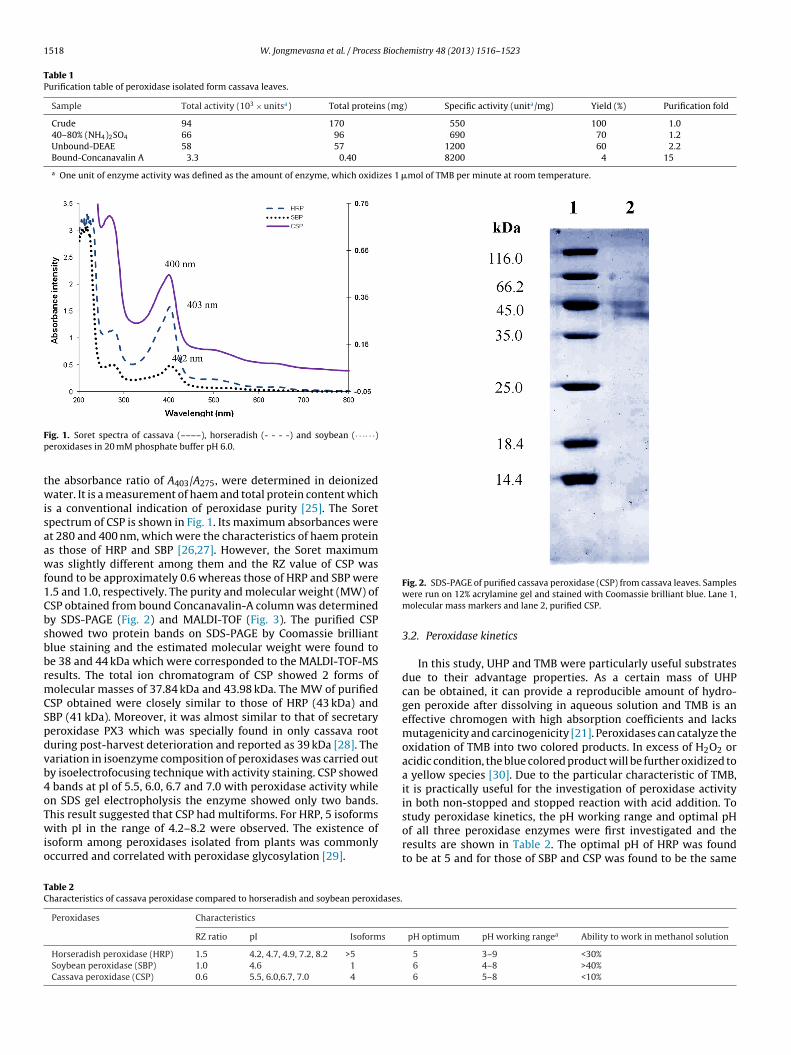
1518 W. Jongmevasna et al. / Process Biochemistry 48 (2013) 1516–1523
Table 1Purification table of peroxidase isolated form cassava leaves.
Sample Total activity (103 × unitsa) Total proteins (mg) Specific activity (unita/mg) Yield (%) Purification fold
Crude 94 170 550 100 1.040–80% (NH4)2SO4 66 96 690 70 1.2Unbound-DEAE 58 57 1200 60 2.2Bound-Concanavalin A 3.3 0.40 8200 4 15
a One unit of enzyme activity was defined as the amount of enzyme, which oxidizes 1 �mol of TMB per minute at room temperature.
Fp
twisaawf1CbsbbrmCSpdvb4oTwio
TC
ig. 1. Soret spectra of cassava (––––), horseradish (- - - -) and soybean (· · ·· · ·)eroxidases in 20 mM phosphate buffer pH 6.0.
he absorbance ratio of A403/A275, were determined in deionizedater. It is a measurement of haem and total protein content which
s a conventional indication of peroxidase purity [25]. The Soretpectrum of CSP is shown in Fig. 1. Its maximum absorbances weret 280 and 400 nm, which were the characteristics of haem proteins those of HRP and SBP [26,27]. However, the Soret maximumas slightly different among them and the RZ value of CSP was
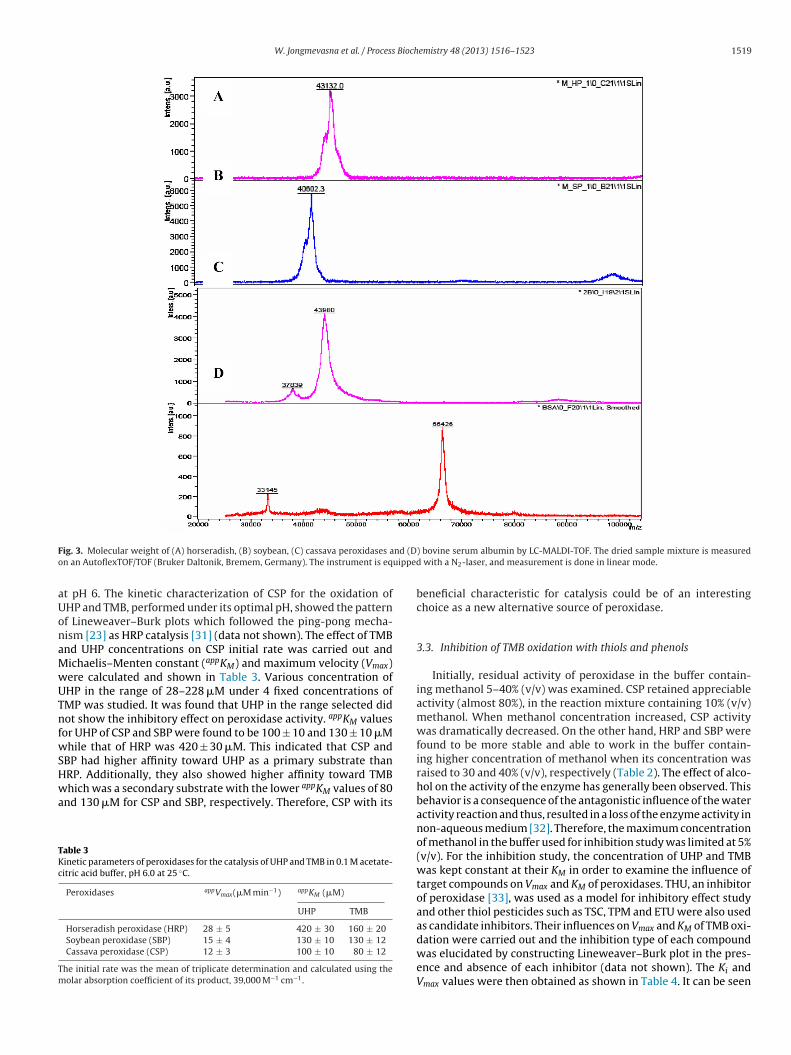
ound to be approximately 0.6 whereas those of HRP and SBP were.5 and 1.0, respectively. The purity and molecular weight (MW) ofSP obtained from bound Concanavalin-A column was determinedy SDS-PAGE (Fig. 2) and MALDI-TOF (Fig. 3). The purified CSPhowed two protein bands on SDS-PAGE by Coomassie brilliantlue staining and the estimated molecular weight were found toe 38 and 44 kDa which were corresponded to the MALDI-TOF-MSesults. The total ion chromatogram of CSP showed 2 forms ofolecular masses of 37.84 kDa and 43.98 kDa. The MW of purified
SP obtained were closely similar to those of HRP (43 kDa) andBP (41 kDa). Moreover, it was almost similar to that of secretaryeroxidase PX3 which was specially found in only cassava rooturing post-harvest deterioration and reported as 39 kDa [28]. Theariation in isoenzyme composition of peroxidases was carried outy isoelectrofocusing technique with activity staining. CSP showed
bands at pI of 5.5, 6.0, 6.7 and 7.0 with peroxidase activity whilen SDS gel electropholysis the enzyme showed only two bands.
his result suggested that CSP had multiforms. For HRP, 5 isoformsith pI in the range of 4.2–8.2 were observed. The existence ofsoform among peroxidases isolated from plants was commonlyccurred and correlated with peroxidase glycosylation [29].
able 2haracteristics of cassava peroxidase compared to horseradish and soybean peroxidases.
Peroxidases Characteristics
RZ ratio pI Isoforms
Horseradish peroxidase (HRP) 1.5 4.2, 4.7, 4.9, 7.2, 8.2 >5
Soybean peroxidase (SBP) 1.0 4.6 1
Cassava peroxidase (CSP) 0.6 5.5, 6.0,6.7, 7.0 4
Fig. 2. SDS-PAGE of purified cassava peroxidase (CSP) from cassava leaves. Sampleswere run on 12% acrylamine gel and stained with Coomassie brilliant blue. Lane 1,molecular mass markers and lane 2, purified CSP.
3.2. Peroxidase kinetics
In this study, UHP and TMB were particularly useful substratesdue to their advantage properties. As a certain mass of UHPcan be obtained, it can provide a reproducible amount of hydro-gen peroxide after dissolving in aqueous solution and TMB is aneffective chromogen with high absorption coefficients and lacksmutagenicity and carcinogenicity [21]. Peroxidases can catalyze theoxidation of TMB into two colored products. In excess of H2O2 oracidic condition, the blue colored product will be further oxidized toa yellow species [30]. Due to the particular characteristic of TMB,it is practically useful for the investigation of peroxidase activityin both non-stopped and stopped reaction with acid addition. To
study peroxidase kinetics, the pH working range and optimal pHof all three peroxidase enzymes were first investigated and theresults are shown in Table 2. The optimal pH of HRP was foundto be at 5 and for those of SBP and CSP was found to be the samepH optimum pH working rangea Ability to work in methanol solution
5 3–9 <30%6 4–8 >40%6 5–8 <10%
W. Jongmevasna et al. / Process Biochemistry 48 (2013) 1516–1523 1519
F nd (Do uipped
aUonaMwUTnfwSHwa
TKc
Tm
ig. 3. Molecular weight of (A) horseradish, (B) soybean, (C) cassava peroxidases an an AutoflexTOF/TOF (Bruker Daltonik, Bremem, Germany). The instrument is eq
t pH 6. The kinetic characterization of CSP for the oxidation ofHP and TMB, performed under its optimal pH, showed the patternf Lineweaver–Burk plots which followed the ping-pong mecha-ism [23] as HRP catalysis [31] (data not shown). The effect of TMBnd UHP concentrations on CSP initial rate was carried out andichaelis–Menten constant (appKM) and maximum velocity (Vmax)ere calculated and shown in Table 3. Various concentration ofHP in the range of 28–228 �M under 4 fixed concentrations ofMP was studied. It was found that UHP in the range selected didot show the inhibitory effect on peroxidase activity. appKM values
or UHP of CSP and SBP were found to be 100 ± 10 and 130 ± 10 �Mhile that of HRP was 420 ± 30 �M. This indicated that CSP and
BP had higher affinity toward UHP as a primary substrate than
RP. Additionally, they also showed higher affinity toward TMBhich was a secondary substrate with the lower appKM values of 80nd 130 �M for CSP and SBP, respectively. Therefore, CSP with its
able 3inetic parameters of peroxidases for the catalysis of UHP and TMB in 0.1 M acetate-itric acid buffer, pH 6.0 at 25 ◦C.
Peroxidases appVmax(�M min−1) appKM (�M)
UHP TMB
Horseradish peroxidase (HRP) 28 ± 5 420 ± 30 160 ± 20Soybean peroxidase (SBP) 15 ± 4 130 ± 10 130 ± 12Cassava peroxidase (CSP) 12 ± 3 100 ± 10 80 ± 12
he initial rate was the mean of triplicate determination and calculated using theolar absorption coefficient of its product, 39,000 M−1 cm−1.
) bovine serum albumin by LC-MALDI-TOF. The dried sample mixture is measured with a N2-laser, and measurement is done in linear mode.
beneficial characteristic for catalysis could be of an interestingchoice as a new alternative source of peroxidase.
3.3. Inhibition of TMB oxidation with thiols and phenols
Initially, residual activity of peroxidase in the buffer contain-ing methanol 5–40% (v/v) was examined. CSP retained appreciableactivity (almost 80%), in the reaction mixture containing 10% (v/v)methanol. When methanol concentration increased, CSP activitywas dramatically decreased. On the other hand, HRP and SBP werefound to be more stable and able to work in the buffer contain-ing higher concentration of methanol when its concentration wasraised to 30 and 40% (v/v), respectively (Table 2). The effect of alco-hol on the activity of the enzyme has generally been observed. Thisbehavior is a consequence of the antagonistic influence of the wateractivity reaction and thus, resulted in a loss of the enzyme activity innon-aqueous medium [32]. Therefore, the maximum concentrationof methanol in the buffer used for inhibition study was limited at 5%(v/v). For the inhibition study, the concentration of UHP and TMBwas kept constant at their KM in order to examine the influence oftarget compounds on Vmax and KM of peroxidases. THU, an inhibitorof peroxidase [33], was used as a model for inhibitory effect studyand other thiol pesticides such as TSC, TPM and ETU were also usedas candidate inhibitors. Their influences on Vmax and KM of TMB oxi-
dation were carried out and the inhibition type of each compoundwas elucidated by constructing Lineweaver–Burk plot in the pres-ence and absence of each inhibitor (data not shown). The Ki andVmax values were then obtained as shown in Table 4. It can be seen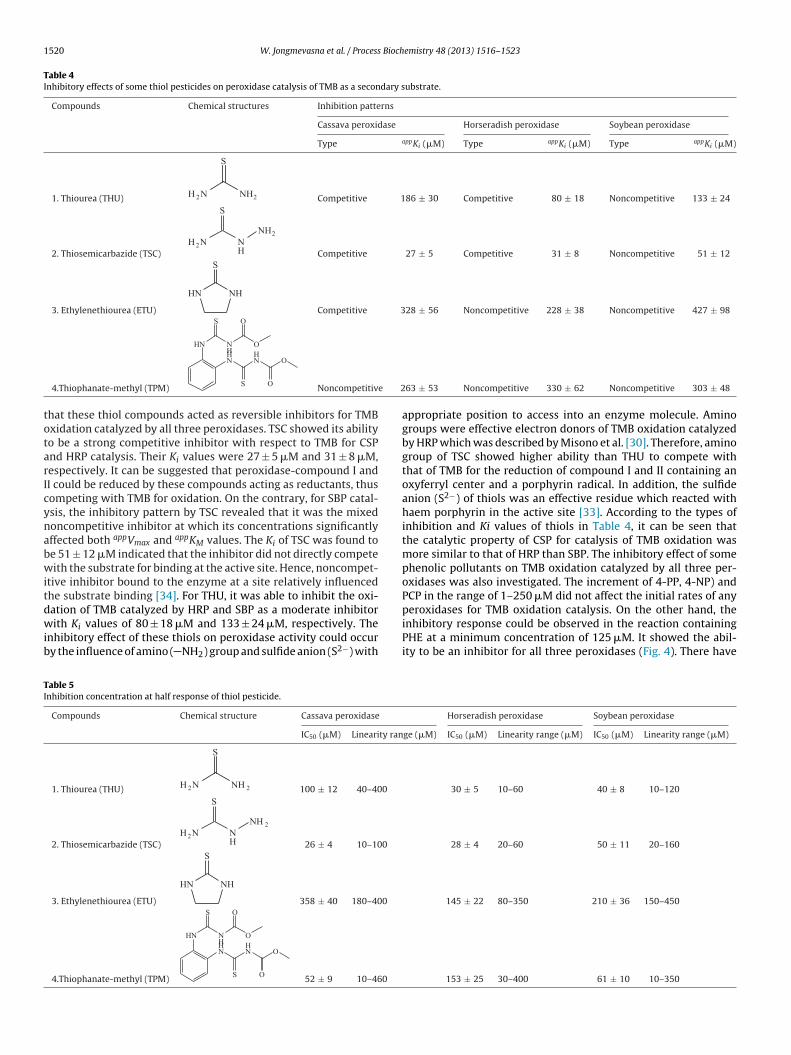
1520 W. Jongmevasna et al. / Process Biochemistry 48 (2013) 1516–1523
Table 4Inhibitory effects of some thiol pesticides on peroxidase catalysis of TMB as a secondary substrate.
Compounds Chemical structures Inhibition patterns
Cassava peroxidase Horseradish peroxidase Soybean peroxidase
Type appKi (�M) Type appKi (�M) Type appKi (�M)
1. Thiourea (THU) NH2
H2N
S
Competitive 186 ± 30 Competitive 80 ± 18 Noncompetitive 133 ± 24
2. Thiosemicarbazide (TSC)N
H
H2N
S
NH2
Competitive 27 ± 5 Competitive 31 ± 8 Noncompetitive 51 ± 12
3. Ethylenethiourea (ETU)
NHHN
S
Competitive 328 ± 56 Noncompetitive 228 ± 38 Noncompetitive 427 ± 98
HN
H
NH
S
O
O
H
e 2
totarIcynabwitdwib
TI
4.Thiophanate-methyl (TPM)
N N
S
O
O Noncompetitiv
hat these thiol compounds acted as reversible inhibitors for TMBxidation catalyzed by all three peroxidases. TSC showed its abilityo be a strong competitive inhibitor with respect to TMB for CSPnd HRP catalysis. Their Ki values were 27 ± 5 �M and 31 ± 8 �M,espectively. It can be suggested that peroxidase-compound I andI could be reduced by these compounds acting as reductants, thusompeting with TMB for oxidation. On the contrary, for SBP catal-sis, the inhibitory pattern by TSC revealed that it was the mixedoncompetitive inhibitor at which its concentrations significantlyffected both appVmax and appKM values. The Ki of TSC was found toe 51 ± 12 �M indicated that the inhibitor did not directly competeith the substrate for binding at the active site. Hence, noncompet-
tive inhibitor bound to the enzyme at a site relatively influencedhe substrate binding [34]. For THU, it was able to inhibit the oxi-
ation of TMB catalyzed by HRP and SBP as a moderate inhibitorith Ki values of 80 ± 18 �M and 133 ± 24 �M, respectively. Thenhibitory effect of these thiols on peroxidase activity could occury the influence of amino ( NH2) group and sulfide anion (S2−) with
able 5nhibition concentration at half response of thiol pesticide.
Compounds Chemical structure Cassava peroxidase
IC50 (�M) Linearity ran
1. Thiourea (THU) NH2
H2N
S
100 ± 12 40–400
2. Thiosemicarbazide (TSC)N
H
H2N
S
NH2
26 ± 4 10–100
3. Ethylenethiourea (ETU)
NHHN
S
358 ± 40 180–400
4.Thiophanate-methyl (TPM)
HN
HN
NH
S
O
O
HN
S
O
O 52 ± 9 10–460
63 ± 53 Noncompetitive 330 ± 62 Noncompetitive 303 ± 48
appropriate position to access into an enzyme molecule. Aminogroups were effective electron donors of TMB oxidation catalyzedby HRP which was described by Misono et al. [30]. Therefore, aminogroup of TSC showed higher ability than THU to compete withthat of TMB for the reduction of compound I and II containing anoxyferryl center and a porphyrin radical. In addition, the sulfideanion (S2−) of thiols was an effective residue which reacted withhaem porphyrin in the active site [33]. According to the types ofinhibition and Ki values of thiols in Table 4, it can be seen thatthe catalytic property of CSP for catalysis of TMB oxidation wasmore similar to that of HRP than SBP. The inhibitory effect of somephenolic pollutants on TMB oxidation catalyzed by all three per-oxidases was also investigated. The increment of 4-PP, 4-NP) andPCP in the range of 1–250 �M did not affect the initial rates of any
peroxidases for TMB oxidation catalysis. On the other hand, theinhibitory response could be observed in the reaction containingPHE at a minimum concentration of 125 �M. It showed the abil-ity to be an inhibitor for all three peroxidases (Fig. 4). There haveHorseradish peroxidase Soybean peroxidase
ge (�M) IC50 (�M) Linearity range (�M) IC50 (�M) Linearity range (�M)
30 ± 5 10–60 40 ± 8 10–120
28 ± 4 20–60 50 ± 11 20–160
145 ± 22 80–350 210 ± 36 150–450
153 ± 25 30–400 61 ± 10 10–350

W. Jongmevasna et al. / Process Biochemistry 48 (2013) 1516–1523 1521
0.0 0. 5 1. 0 1. 5 2. 0 2.5 3.00
20
40
60
80
100
120
4-PP
PHEPCP4-NP
log[Conc entration, µM]
% F
ract
iona
l act
ivity
0.0 0.5 1.0 1. 5 2.0 2.5 3. 00
20
40
60
80
100
120
ETU
TPMTSCTHU
log [Conc entration , µM]
% F
ract
iona
l act
ivity
0.0 0.5 1.0 1. 5 2.0 2.5 3. 00
20
40
60
80
100
120
ETU
TPMTSCTHU
log [Conc entration , µM]
% F
ract
iona
l act
ivity
0.0 0. 5 1.0 1.5 2.0 2. 5 3.00
20
40
60
80
100
120
ETU
TPMTSCTHU
log[Con centration , µM]
% F
ract
iona
l act
ivity
0.0 0.5 1.0 1.5 2.0 2.5 3.00
20
40
60
80
100
120
4-PP
PHEPCP4-NP
log[Conce ntr ation , µM]
% F
ract
iona
l act
ivity
0.0 0. 5 1. 0 1. 5 2. 0 2.5 3.00
20
40
60
80
100
120
4-PP
PHEPCP4-NP
log[Conc entration, µM]
% F
ract
ion
activ
ity
2.A 1.A
2.B 1.B
2.C 1.C
Fig. 4. Dose–response curves of thiols (A1-C1: ETU, THU; TSC and TPM) and phenols (A2-C2: 4-PP, 4-NP; PCP and PHE) toward activities of (A) horseradish, (B) soybean and(C) cassava peroxidases.
bdtranrwowaoptwnB
Dose–response plots are widely used for comparing the relativeinhibitor potencies of multiple compounds for the same enzyme
een many studies focused on the investigation of phenol and itserivative properties toward peroxidase catalysis. They suggestedhat phenols were usually considered as inhibitors or alternativeeducing substrates for peroxidases. Their certain abilities to acts inhibitors were dependent on the oxidation potential of phe-ols compared to that of substrate used for monitoring peroxidaseeaction [35–37]. According to the report of Bagirova et al. [38], itas found that phenol acted as a weak inhibitor for peanut per-
xidase catalysis of TMB oxidation. They proposed that phenolsith higher redox potentials than the secondary substrate were
ble to act as peroxidase inhibitor and their influences on per-xidase catalysis also correlated with other factors such as size,osition of their substitutes and the accessibility of hydroxyl groupo the enzyme molecule. Moreover, the degree of inhibitory effectas also reliant on phenol concentration and the ratio of phe-
ol and hydrogen peroxide was shown as a summary reported inaynton et al. [39].3.4. Dose–response curve of thiols toward peroxidase activity
The half maximal inhibitory concentration (IC50) values of somethiol compounds were determined by dose–response curve. Thefinding might be useful for the development of a method for thedetection of thiol pesticide residue based on inhibitory effect. Thiolpesticide such as TSC has been used as a rodenticide and is effectivefor the control of bacterial leaf blight of rice. TPM is an effec-tive fungicide against a wide range of fungal pathogens of cerealsand fruits [40] and ETU, a metabolite of ethylenebisdithiocarba-mate fungicides degradation [41] also showed the ability to bean inhibitor of peroxidases. Therefore, the potentially detectablerange of those compounds was examined by dose response curve.
and screening several compounds as potential inhibitors of a par-ticular target enzyme [23]. The IC50 value is a practical readout of

1 Bioch
taiaecaaaT2tabae0(dw[to
4
Tioetrica
A
tMfigC
A
b1
R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
522 W. Jongmevasna et al. / Process
he relative effects on enzyme activity of different substances under specific set of solutions. In Fig. 4, TSC was considered to be a strongnhibitor which gave the lowest IC50 value for TMB oxidation cat-lyzed by CSP and HRP. In the case of THU, it was also found to be anffective inhibitor for HRP and SBP catalysis. Moreover, the appli-able range for thiol detection by enzymatic inhibition was defineds shown in Table 5. From these results, CSP-catalyzed reaction wasble to detect TSC and TPM in solution in the range of 10–100 �Mnd 10–460 �M, respectively. In HRP-catalyzed reaction, THU andSC could be detected in the applicable range of 10–60 �M and0–60 �M, respectively, whereas SBP showed the ability to detecthem in the range of 10–120 �M and 20–160 �M, respectively. Thepplicable uses of peroxidase for target compounds determinationased on the inhibitory effects were also reported in many liter-tures. THU in methanol could be detected by amperometric HRPlectrode in the range of 0.05–0.6 mM [33], chloramine in the range.026–1.04 mM could be determined by flow injection analysisFIA) with enzyme-reactor [42]. In addition, the concentration-ependent plots of 3-phenylcoumarins in the range of 1–50 �Mere assessed by luminal-enhanced chemiluminescence assay
43]. Therefore, these preliminary results indicated a good perspec-ive for the practical use of CSP for the detection of some thiols andther related compounds.
. Conclusions
Peroxidase from cassava leaves, an agricultural waste inhailand, could be an alternative source of cationic haem perox-dase. Its catalytic ability for urea hydrogen peroxide and TMBxidation was in the pH range of 5–8. CSP-catalyzed reaction underxperimentally designed condition had a potential for the detec-ion of thiosemicarbazide, thiourea and thiophanate methyl as itseversible inhibitors. The catalytic activity of CSP was particularlynfluenced by some thiol pesticides and phenolic pollutants inomparison with HRP and SBP which can be an advantage to thepplicable use of the enzyme in chemical analysis.
cknowledgments
The authors would like to thank Dr. Jaran Jainhuknan for theechnical assistance of the molecular weight determination by
ADI-TOF and gratefully acknowledge the laboratory facilities andnancial support provided by Department of Biochemistry, Pro-ram in Biotechnology, Faculty of Science and Graduate School ofhulalongkorn University.
ppendix A. Supplementary data
Supplementary material related to this article cane found, in the online version, at http://dx.doi.org/0.1016/j.procbio.2013.07.014.
eferences
[1] Song F, Song G, Dong A, Kong K. Regulatory mechanisms of host plant defenseresponses to arbuscular mycorrhiza. Acta Ecologica Sinica 2011;31(6):322–7.
[2] Nikraftar F, Taheri P, Rastegar MF, Tarighi S. Tomato partial resistance to Rhi-zoctonia solani involves antioxidative defense mechanisms. Physiology andMolecular Biology of Plants 2013;81:74–83.
[3] Bogdanovic J, Milosavic N, Prodanovic R, Ducic T, Radotic K. Variability ofantioxidant enzyme activity and isoenzyme profile in needles of Serbianspruce (Picea omorika (Panc.) Purkinye). Biochemical Systematics and Ecology2007;35(5):263–73.
[4] Talano MA, Agostini E, Medina MI, Reinoso H, Tordable MC, Tigier HA, et al.
Changes in ligno-suberization of cell walls of tomato hairy roots produced bysalt treatment: the relationship with the release of a basic peroxidase. Journalof Plant Physiology 2006;163(7):740–9.[5] Hamid M, Rehman K. Potential applications of peroxidases. Food Chemistry2009;115(4):1177–86.
[
emistry 48 (2013) 1516–1523
[6] Kafi AKM, Wu G, Chen A. A novel hydrogen peroxide biosensor based on theimmobilization of horseradish peroxidase onto Au-modified titanium dioxidenanotube arrays. Biosensors and Bioelectronics 2008;24(4):566–71.
[7] ElKaoutit M, Rodriguez IN, Domínguez M, Artiga MPH, Milla DB,Hidalgo de Cisneros JLH. A third-generation hydrogen peroxide biosen-sor based on Horseradish Peroxidase (HRP) enzyme immobilized in aNafion–Sonogel–Carbon composite. Electrochimica Acta 2008;53(24):7131–7.
[8] Liu L, Chen Z, Yang S, Jin X, Lin X. A novel inhibition biosensor constructed bylayer-by-layer technique based on biospecific affinity for the determination ofsulfide. Sensors and Actuators B: Chemical 2008;129(1):218–24.
[9] Yu D, Blankert B, Kauffmann JM. Development of amperometric horseradishperoxidase based biosensors for clozapine and for the screening of thiol com-pounds. Biosensors and Bioelectronics 2007;22:2707–11.
10] Dalal S, Gupta MN. Treatment of phenolic wastewater by horseradish peroxi-dase immobilized by bioaffinity layering. Chemosphere 2007;67(4):741–7.
11] Cheng J, Yu SM, Zuo P. Horseradish peroxidase immobilized on aluminumpillared interlayered clay for the catalytic oxidation of phenolic wastewater.Water Research 2006;40:283–90.
12] Wright PJ, English AM. Buffer-anion-dependent Ca2+ leaching from horseradishperoxidase at low pH. Journal of Biological Inorganic Chemistry 2001;6:348–58.
13] Kamal1 JKA, Behere DV. Kinetic stabilities of soybean and horseradish peroxi-dases. Biochemical Engineering Journal 2008;38(1):110–4.
14] Das MK, Sharma RS, Mishra V. A novel cationic peroxidase (VanPrx) from ahemi-parasitic plant (Viscum angulatum) of Western Ghats (India): purifica-tion, characterization and kinetic properties. Journal of Molecular Catalysis B:Enzymatic 2011;71(1–2):63–70.
15] Cardinali A, Tursi N, Ligorio A, Giuffrida MG, Napolitano L, Caliandro R,et al. Purification, biochemical characterization and cloning of a new cationicperoxidase isoenzyme from artichoke. Plant Physiology and Biochemistry2011;49:395–403.
16] Pandey VP, Dwivedi UN. Purification and characterization of peroxidase fromLeucaena leucocephala, a tree legume. Journal of Molecular Catalysis B: Enzy-matic 2011;68:168–73.
17] The Thai Tapioca Trade Association. TTTA NEWS. Issue No.24/2010/31December; 2010.
18] Gómez-Vasquez R, Day R, Buschmann H, Randles S, Beeching JR, Cooper RM.Phenylpropanoids, phenylalanine ammonialyase and peroxidases in elicitor-challenged cassava (Manihot esculenta) suspension cells and leaves. Annals ofBotany 2004;94(1):87–97.
19] Laemmli UK. Cleavage of structural proteins during the assembly of the headof bacteriophage T4. Nature 1970;227:680–5.
20] Fernandes CF, Moraes VCP, Vasconcelos IM, Silveira JAG, Oliveira JTA. Inductionof an anionic peroxidase in cowpea leaves by exogenous salicylic acid. Journalof Plant Physiology 2006;163:1040–8.
21] Bos ES, Doelen AA, Rooy N, Shuurs AHWM. 3,3′ ,5,5′-Tetramethylbenzidineas an Ames test negative chromogen for horseradish peroxidase in enzymeimmunoassay. Journal of Immunoassay 1981;2:187–204.
22] Bradford MM. A rapid and sensitive method for the quantitation of microgramsquantities for proteins utilizing the principle of protein-dye binding. AnalyticalBiochemistry 1976;72:248–54.
23] Copeland RA. Enzyme: a practical introduction to structure, mechanism, anddata analysis. 2nd ed. USA: Wiley-VCH, Inc.; 2000.
24] Miranda MV, Magri ML, Navarro del Canizo AA, Cascone O. Study ofvariables involved in horseradish and soybean peroxidase purification byaffinity chromatography on concanavalin-A-agrarose. Process Biochemistry2002;38:537–43.
25] Das MK, Sharma RS, Mishra V. A novel cationic peroxidase (VanPrx) from ahemi-parasitic plant (Viscum angulatum) of Western Ghats (India): purifica-tion, characterization and kinetic properties. Journal of Molecular Catalysis B:Enzymatic 2011;71:63–70.
26] Welinder KG. Amino acid sequence studies of horseradish peroxidase: aminoand carboxyl termini, cyanogen bromide and tryptic fragments, the completesequence, and some structural characteristics of horseradish peroxidase C.European Journal of Biochemistry 1979;96:483–502.
27] Welinder KG, Larsen YB. Covalent structure of soybean seed coat peroxidase.Biochimica et Biophysica Acta 2004;1698:121–6.
28] Reilly K, Bernal D, Corte’s DF, Tohme RJ, Beeching JR. Towards identifying the fullset of genes expressed during cassava post-harvest physiological deterioration.Plant Molecular Biology 2007;64:187–203.
29] Gobaldon C, Gomez-ros LV, Nunez-flores MJL, Esteban-Carrason A, Barcelo AR.Post-translational modifications of the basic peroxidase isoenzyme from Zinniaelegans. Plant Molecular Biology 2007;65:43–61.
30] Misono Y, Ohkata Y, Morikawa T, Itoh K. Resonance Raman and adsorp-tion spectroscopic studies on the electronchemical oxidation processesof 3,3′ ,5,5′-tetramethylbenzidine. Journal of Electroanalytical Chemistry1997;436:203–12.
31] Banci L. Structural properties of peroxidases. Journal of Biotechnology1997;53:253–63.
32] Dordick JS. Enzymatic catalysis in organic media: fundamentals and selectedapplications. ASGSB Bulletin 1991;4:125–32.
33] Adeyoju O, Iwuoha EI, Smyth MR. Reactivities of amperometric organic
phase peroxidase-modified electrodes in the presence and absence ofthiourea and ethylenethiourea as inhibitors. Analytica Chimica Acta 1995;305:57–64.34] Sariri R, Sajedi RH, Jafarian V. Inhibition of horseradish peroxidase activity bythiol type inhibitors. Journal of Molecular Liquids 2006;123:20–3.

Bioch
[
[
[
[
[
[
[
[
Chimica Acta 1999;390:141–6.[43] Kabeya LM, Marchi AA, KanashiroA, Lopes NP, Silva CHTP, Pupob MT, et al. Inhi-
W. Jongmevasna et al. / Process
35] Riso AD, Gullotti M, Casell L, Monzani E, Profumo A, Gianelli L, et al. Selectivityin the peroxidase catalyzed oxidation of phenolic sulfides. Journal of MolecularCatalysis A: Chemical 2003;204–205:391–400.
36] Job D, Dunford HB. Substituent effect of the oxidation of phenols and aromaticamines by horseradish peroxidase compound I. European Journal of Biochem-istry 1976;66:607–14.
37] Metelitza DI, Karasyova EI, Grintsevich EE, Thorneley RNF. Peroxidase-catalyzed co-oxidation of 3,3′ ,5,5′-tetramethylbenzidine in the presence ofsubstituted phenols and their polydisulfides. Journal of Inorganic Biochemistry2004;98:1–9.
38] Bagirova NA, Shekhovtsova TN, Huystee RB. Enzymatic determination of phe-nols using peanut peroxidase. Talanta 2001;55:1151–64.
39] Baynton KJ, Brewtra JK, Biswas N, Taylor KE. Inactivation of horseradish perox-idase by phenol and hydrogen peroxide: a kinetic investigation. Biochimica etBiophysica Acta 1994;1206:272–8.
emistry 48 (2013) 1516–1523 1523
40] EPA RED. Facts thiophanate-methyl. In: Prevention Pesticides and Toxic Sub-stances. United States Environmental Protection Agency; 2004 (7508C).
41] Engst R, Nash RG, Newsome WH, Morley HV, Schnaak W, Turtle E, et al.International union of pure and applied chemistry, applied chemistry divi-sion commission on terminal pesticide residues ethyleneurea. Pure & AppliedChemistry 1977;49:675–89.
42] Buck S, Stein K, Schwedt G. Use of a peroxidase reactor in flow injection anal-ysis for the determination of chloramine and the inhibition kinetics. Analytica
bition of horseradish peroxidase catalytic activity by new 3-phenylcoumarinderivatives: synthesis and structure–activity relationships. Bioorganic andMedicinal Chemistry 2007;15:1516–24.
Copyright © 2022 FDOKUMEN




















