
Sonocatalytic degradation of tetracycline antibiotic in aqueous solution by sonocatalysis
Upload
independentCategory
view
2download
0
doi:10.1006/jmbi.2000.3840 available online at http://www.idealibrary.com on J. Mol. Biol. (2000) 300, 1309±1322
Tetracycline Affects Abnormal Properties of SyntheticPrP Peptides and PrPSc in vitro
Fabrizio Tagliavini1, Gianluigi Forloni2, Laura Colombo2
Giacomina Rossi1, Laura Girola2, Barbara Canciani1, Nadia Angeretti2
Lidia Giampaolo1, Elisa Peressini2, Tazeen Awan1, Luca De Gioia3
Enzio Ragg4, Orso Bugiani1 and Mario Salmona2*
Introduction
Prion diseases such as scrapie of sheep, spongi-form encephalopathy of cattle (BSE) and Creutz-feldt-Jakob disease (CJD), fatal familial insomniaand Gerstmann-StraÈussler-Scheinker disease (GSS)of humans are transmissible neurodegenerativedisorders that share a common pathogenic mech-
E-mail address of the corresponding author:[email protected]
Abbreviations used: BSE, bovine spongiformencephalopathy; CJD, Creutzfeldt-Jakob disease; vCJD,new variant of Creutzfeldt-Jakob disease; GSS,Gerstmann-StraÈussler-Scheinker disease; PrP, prionprotein; PrPC, cellular isoform of PrP; PrPSc, scrapieisoform of PrP; COSY-dQF, double quantum ®lteredcorrelated spectroscopy; TOCSY, total correlatedspectroscopy; NOE, nuclear Overhauser effect; NOESY,NOE spectroscopy.
0022-2836/00/051309±14 $35.00/0
anism, i.e. the conversion of the cellular prion pro-tein (PrPC) into disease-speci®c species termedPrPSc (Prusiner, 1991; Prusiner et al., 1998). Thisprocess involves a conformational change of PrPC
resulting in decreased a-helical structure and strik-ing increase in b-sheet content (Caughey et al.,1991; Pan et al., 1993; Safar et al., 1993). The PrPC toPrPSc transition is associated with the acquisitionof abnormal chemicophysical properties, includinginsolubility in non-denaturing detergents, partialresistance to protease digestion and high tendencyto form aggregates and amyloid ®brils (Prusiner,1991; Prusiner et al., 1998). The accumulation ofPrPSc and fragments thereof in the central nervoussystem is accompanied by activation of microglialcells, hypertrophy and proliferation of astrocytesand degeneration of neurons, suggesting a primaryrole of abnormal PrP peptides in the pathologicprocess (De Armond et al., 1988, 1993; Bruce et al.,1989; Williams et al., 1997). Evidence indicates that
1Istituto Nazionale NeurologicoCarlo Besta, Via Celoria 1120133 Milano, Italy2Department of Biochemistryand Molecular Pharmacologyand Department of NeurologyIstituto di RicercheFarmacologiche Mario NegriVia Eritrea 62, 20157 MilanoItaly3Dipartimento di Biotecnologiee Bioscienze, UniversitaÁ diMilano-Bicocca, Piazza DellaScienza 2, 20126 Milano, Italy4Dipartimento di ScienzeMolecolari Agroalimentari,FacoltaÁ di Agraria UniversitaÁ diMilano, Via Celoria 220133 Milano, Italy
Prion diseases are characterized by the accumulation of altered forms ofthe prion protein (termed PrPSc) in the brain. Unlike the normal protein,PrPSc isoforms have a high content of b-sheet secondary structure, areprotease-resistant, and form insoluble aggregates and amyloid ®brils. Evi-dence indicates that they are responsible for neuropathological changes(i.e. nerve cell degeneration and glial cell activation) and transmissibilityof the disease process. Here, we show that the antibiotic tetracycline: (i)binds to amyloid ®brils generated by synthetic peptides corresponding toresidues 106-126 and 82-146 of human PrP; (ii) hinders assembly of thesepeptides into amyloid ®brils; (iii) reverts the protease resistance of PrPpeptide aggregates and PrPSc extracted from brain tissue of patients withCreutzfeldt-Jakob disease; (iv) prevents neuronal death and astrocyte pro-liferation induced by PrP peptides in vitro. NMR spectroscopy revealedseveral through-space interactions between aromatic protons of tetra-cycline and side-chain protons of Ala117-119, Val121-122 and Leu125 of PrP106-126. These properties make tetracycline a prototype of compoundswith the potential of inactivating the pathogenic forms of PrP.
# 2000 Academic Press
Keywords: amyloidogenesis; cell cultures; NMR spectroscopy;PrP peptides; tetracycline
*Corresponding author# 2000 Academic Press

1310 Tetracycline Interacts with the Prion Protein
the neoformed PrPSc interacts with PrPC and con-verts the normal protein into a likeness of itself(Kocisko et al., 1994; Bessen et al., 1995). Accordingto this view, PrPSc is responsible for transmissibil-ity and progression of the disease as well as forbrain damage, thus representing a primary targetfor therapeutic strategies.
The urgency to identify and develop therapeuticcompounds has remarkably increased followingthe epidemic of BSE and the appearance of a newvariant of Creutzfeldt-Jakob disease (vCJD) thatseem to be causally linked (Will et al., 1996;Collinge et al., 1996; Bruce et al., 1997). Althoughthe number of vCJD cases observed so far is lim-ited, a future outbreak of this disease cannot beexcluded (Ghani et al., 1998).
A variety of compounds have been tested on cel-lular and animal models of prion disease. A few ofthese compounds, including sulphated polyanions(Kimberlin & Walker, 1983, 1986; Ehlers &Diringer, 1984; Farquhar & Dickinson, 1986;Caughey & Raymond, 1993), amphotericin B(Pocchiari et al., 1987; Xi et al., 1992; Adnjou et al.,1995), Congo red (Caughey & Race, 1992; Ingrossoet al., 1995), iododoxorubicin (Tagliavini et al.,1997), tetrapyrroles (Caughey et al., 1998; Priolaet al., 2000), branched polyamines (Supattaponeet al., 1999) and modi®ed PrP peptides (Soto et al.,2000), effectively reduce the accumulation of PrPSc
in scrapie-infected neuroblastoma cells and/ordelay the appearance of clinical symptoms andprolong the survival time of rodents experimen-tally infected with scrapie. This group of com-pounds is structurally heterogeneous and themechanism of action of individual molecules inexperimental prion disease is unknown. However,some of these molecules (i.e. Congo red, iododox-orubicin and tetrapyrroles) share common chemi-cophysical properties in that they contain anextended hydrophobic core formed by aromaticmoieties with a large number of hydrophilic substi-tuents conferring an amphiphilic character. Sincethe conversion of PrPC into PrPSc results in anincrease of hydrophobicity due to solvent exposureof lipophilic residues and in the ability to forminsoluble aggregates and amyloid ®brils, it is con-ceivable that the anti-prion activity of the com-pounds is related to their hydrophobic characterallowing for a strong interaction with PrPSc andfragments thereof. This interaction has been shownfor Congo red and iododoxorubicin, which bind toamyloid ®brils of different biochemical compo-sition, including PrP amyloid (Prusiner et al., 1983;Merlini et al., 1995; Tagliavini et al., 1997).
Based on structural analogies with Congo redand iododoxorubicin, we hypothesized that tetra-cyclines might be able to interact with PrPSc andPrP amyloid. An advantage of this class of drugs isthat several congeners with well characterizedpharmacological and pharmacokinetic propertiesare available. Further, tetracyclines have relativelylow toxicity and some derivatives cross the blood/brain barrier ef®ciently if an appropriate route of
treatment is used (Sande & Mandell, 1991). In thisstudy we show that tetracycline binds and disruptsPrP peptide aggregates, reverts the protease resist-ance of PrPSc extracted from brain tissue of patientswith CJD, and abolishes the neurotoxicity andastroglial proliferation induced by PrP peptidesin vitro. These properties make tetracycline a proto-type of compounds with the potential of inactivat-ing the pathogenic forms of PrP, impeding PrPC toPrPSc conversion, and hindering the developmentand progression of prion disease.
Results
Tetracycline binds to PrP peptides
The ability of tetracycline to interact with PrPaggregates was investigated using amyloid ®brilsgenerated by synthetic peptides homologous toresidues 82 to 146 and 106 to 126 of human PrP(PrP 82-146 and PrP 106-126). The former corre-sponds to the amyloid protein puri®ed from braintissue of GSS patients (Tagliavini et al., 1991, 1994)while the latter is a fragment thereof which shareschemicophysical and biological properties with PrP82-146 as well as with the protease-resistant core ofPrPSc. In particular, PrP 106-126 has a high propen-sity to adopt b-sheet secondary structure and formamyloid ®brils, it is neurotoxic and induces glialcell activation in vitro (Forloni et al., 1993, 1994;Selvaggini et al., 1993; Tagliavini et al., 1993; Brownet al., 1996). The PrP 82-146 and PrP 106-126 assem-blies were incubated either with the amyloid-bind-ing ¯uorochrome thio¯avine S or with tetracyclinewhich has an intrinsic ¯uorescence (l excitation422 nm, l emission 530 nm). Microscopic analysisshowed that the PrP 106-126 and PrP 82-146 aggre-gates revealed by thio¯avine S were labeled bytetracycline (Figure 1(a)-(d)). This ®nding was veri-®ed by ¯uorescence spectroscopic analysis of tetra-cycline in the absence or presence of PrP 82-146and PrP 106-126 at a peptide-to-tetracycline molarratio ranging from 1:0.16 to 1:8 (Figure 1(e)-(l)). Inthe presence of the peptides, tetracycline showed ashift in the emission spectra with a new maximumat 472 nm, con®rming the binding of the com-pound to the PrP peptides. Conversely, the exci-tation spectra did not differ qualitatively even if amodest shift in the maxima was apparent.
Tetracycline prevents aggregation andacquisition of protease resistance ofPrP peptides
These ®ndings prompted us to investigatewhether the interaction between tetracycline andPrP peptides affected the peptide's ability to aggre-gate. PrP 106-126 and PrP 82-146 were dissolved indeionized water in the absence or presence of equi-molar concentration of tetracycline, and incubatedat 37 �C for one and four hours, respectively. Phos-phate buffer was then added to the samples to trig-ger peptide aggregation. Following 0-96 hours

Figure 1. Binding of tetracyclineto PrP peptides. (a)-(d) Fluor-escence microscopy of (a) and (b)PrP 106-126, and (c) and (d) PrP82-146 aggregates after incubationwith (a) and (c) the amyloid-bind-ing ¯uorochrome thio¯avine S and(b) and (d) tetracycline. The peptideaggregates possess staining proper-ties of (a) and (c) amyloid and bind(b) and (d) the antibiotic tetra-cycline. The scale bar represents10 mm. (e)-(l) Fluorescence spec-troscopy of tetracycline either in (e)and (f) the absence or (g) and (h)the presence of PrP 106-126, and (i)and (l) PrP 82-146. The excitationand emission spectra of 100, 250and 500 mM tetracycline in deio-nized water showed maxima at 422and 530 nm, (e) and (f) respect-ively. The addition of (g) and (h)PrP 106-126 and (i) and (l) PrP 82-146 to tetracycline showed achange in the emission spectra witha new maximum at 472 nm, indi-cating the binding of the com-pound to PrP. The spectra shownin (g)-(l) result from the analysis of100 mM tetracycline solutions con-taining 16, 32, 64, 128, 256, 512 and800 mM PrP peptides, after subtrac-tion of the spectrum of tetracyclinealone.
Tetracycline Interacts with the Prion Protein 1311
incubation, the samples were centrifuged and thesupernatants were analyzed by HPLC to determinethe percentage of non-sedimentable peptide. In theabsence of tetracycline, both PrP peptides readilyformed macroaggregates although with differentkinetics. Over 60 % of PrP 106-126 was sedimenta-ble after four hours and only 10 % was still presentin the supernatant after two days. On the otherhand, a signi®cant decrease in the amount of sol-uble PrP 82-146 was ®rst observed after 72 hours;nevertheless, 90 % of this peptide was alsosedimented by centrifugation after 96 hours
(Figure 2(a)). Tetracycline largely prevented theformation of macroaggregates, since more than80 % of PrP 106-126 and PrP 82-146 was in thesupernatant fraction after 96 hours (Figure 2(a)).
We have shown that the aggregation of PrP 106-126 is accompanied by the acquisition of partialresistance to protease digestion (Selvaggini et al.,1993). To investigate whether the ability of tetra-cycline to hinder PrP peptide assembly resulted inincreased susceptibility to proteolysis, PrP 106-126and PrP 82-146 incubated in the presence orabsence of the compound were subjected to pro-

Figure 2. Tetracycline hindersaggregation and abolishes protease-resistance of PrP peptides.(a) Time-course of PrP peptideaggregation in the absence or pre-sence of tetracycline. The peptidesPrP 106-126 and PrP 82-146 wereincubated with the compound atequimolar concentration for0-96 hours. Following centrifu-gation, the supernatant was ana-lyzed by HPLC. The non-sedimentable peptide is expressedas the percentage of the totalamount of peptide at time 0. Eachvalue is the mean � SD of at least®ve experiments. (b) and (c) Pro-tease-resistance of (b) PrP 106-126and (c) PrP 82-146 in either theabsence or presence of tetracycline.Solutions of peptides were incu-bated with the compound at equi-molar concentration for 18 hoursand then subjected to digestionwith proteinase K at 1:100 (w/w)enzyme-to-substrate ratio. Follow-ing centrifugation, the pellet wasdissolved in 10 % formic acid con-taining 0.1 % tri¯uoroacetic acid,and analyzed by HPLC. The extentof proteolysis was calculated as thepercentage of peptide present inthe pellet. Each value is the mean� SD of at least ®ve experiments.
1312 Tetracycline Interacts with the Prion Protein
teinase K digestion. Without tetracycline, theenzyme hydrolyzed �50 % of either peptide, asdeduced by HPLC analysis; conversely, when PrP106-126 and PrP 82-146 were preincubated withthe drug, the peptides were almost completelydigested by proteinase K (Figure 2(b) and (c)).
Tetracycline disrupts PrP peptide aggregates
To establish the effects of tetracycline on thestructure of peptide assemblies, PrP 106-126 wasdissolved in Tris buffer in the presence or absenceof equimolar concentration of the compound andanalyzed by electron microscopy after 1, 24 and72 hours incubation. In the absence of tetracycline,PrP 106-126 aggregates consisted of dense mesh-works of straight, unbranched ®brils, �8 nm indiameter and up to 2 mm in length (Figure 3(a)), asdescribed previously (Tagliavini et al., 1993); nosigni®cant differences were observed amongsamples following 1, 24 and 72 hours incubation,except for increasing ®bril density. Tetracyclineaffected ®brillogenesis of PrP106-126 and the stab-
ility of peptide assemblies. The incubation with thecompound for one hour yielded a change in theappearance of the ®brils without obvious modi®-cation of their density as compared to that of con-trols. In particular, the ®brillary structures wereshorter and more variable in diameter, which ran-ged from �3 to �8 nm. A more prolonged incu-bation with tetracycline (24 and 72 hours) resultedin disarrangement and fragmentation of the ®brilsand ultimately in disappearance of ®brillary struc-tures (Figure 3(b) and (c)).
Site of interaction between tetracyclineand PrP 106-126 as deduced byNMR spectroscopy
The site of interaction between tetracycline andPrP peptide was investigated by 2D-NMR. Reson-ance assignments were achieved by standard tech-niques involving the analysis of the individualspin-systems (double quantum ®ltered correlatedspectroscopy (COSY-dQF) and total correlatedspectroscopy (TOCSY) experiments) and, in
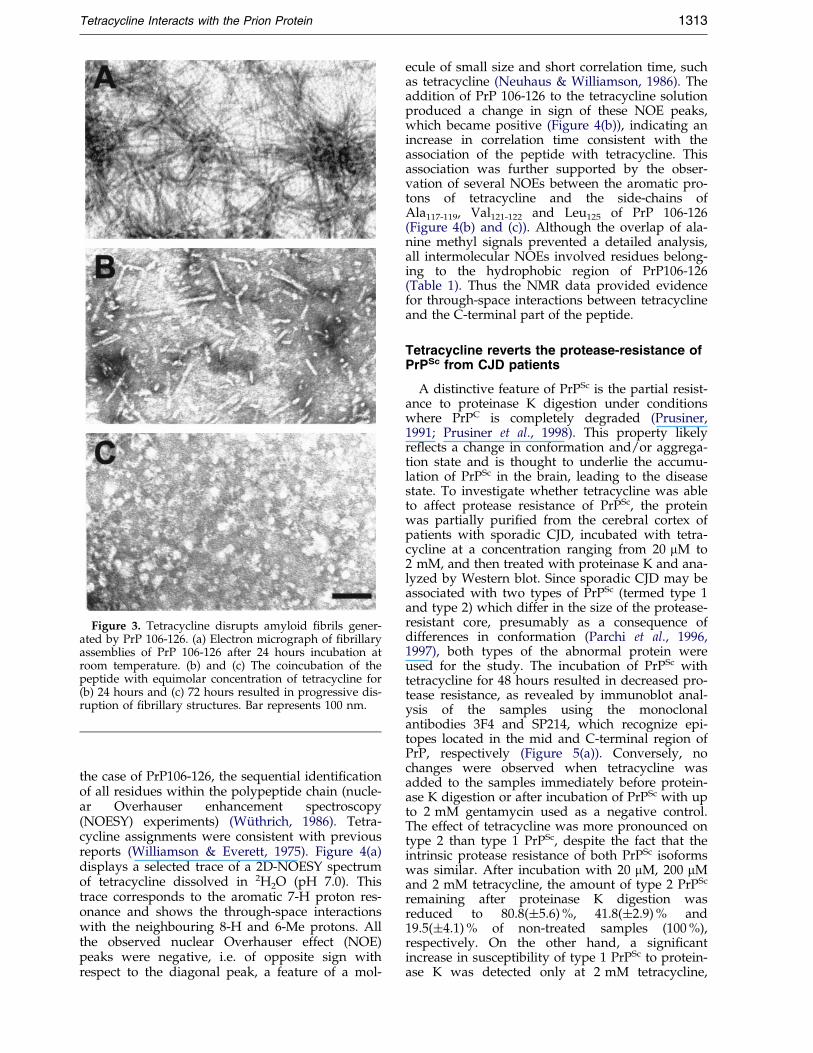
Figure 3. Tetracycline disrupts amyloid ®brils gener-ated by PrP 106-126. (a) Electron micrograph of ®brillaryassemblies of PrP 106-126 after 24 hours incubation atroom temperature. (b) and (c) The coincubation of thepeptide with equimolar concentration of tetracycline for(b) 24 hours and (c) 72 hours resulted in progressive dis-ruption of ®brillary structures. Bar represents 100 nm.
Tetracycline Interacts with the Prion Protein 1313
the case of PrP106-126, the sequential identi®cationof all residues within the polypeptide chain (nucle-ar Overhauser enhancement spectroscopy(NOESY) experiments) (WuÈ thrich, 1986). Tetra-cycline assignments were consistent with previousreports (Williamson & Everett, 1975). Figure 4(a)displays a selected trace of a 2D-NOESY spectrumof tetracycline dissolved in 2H2O (pH 7.0). Thistrace corresponds to the aromatic 7-H proton res-onance and shows the through-space interactionswith the neighbouring 8-H and 6-Me protons. Allthe observed nuclear Overhauser effect (NOE)peaks were negative, i.e. of opposite sign withrespect to the diagonal peak, a feature of a mol-
ecule of small size and short correlation time, suchas tetracycline (Neuhaus & Williamson, 1986). Theaddition of PrP 106-126 to the tetracycline solutionproduced a change in sign of these NOE peaks,which became positive (Figure 4(b)), indicating anincrease in correlation time consistent with theassociation of the peptide with tetracycline. Thisassociation was further supported by the obser-vation of several NOEs between the aromatic pro-tons of tetracycline and the side-chains ofAla117-119, Val121-122 and Leu125 of PrP 106-126(Figure 4(b) and (c)). Although the overlap of ala-nine methyl signals prevented a detailed analysis,all intermolecular NOEs involved residues belong-ing to the hydrophobic region of PrP106-126(Table 1). Thus the NMR data provided evidencefor through-space interactions between tetracyclineand the C-terminal part of the peptide.
Tetracycline reverts the protease-resistance ofPrPSc from CJD patients
A distinctive feature of PrPSc is the partial resist-ance to proteinase K digestion under conditionswhere PrPC is completely degraded (Prusiner,1991; Prusiner et al., 1998). This property likelyre¯ects a change in conformation and/or aggrega-tion state and is thought to underlie the accumu-lation of PrPSc in the brain, leading to the diseasestate. To investigate whether tetracycline was ableto affect protease resistance of PrPSc, the proteinwas partially puri®ed from the cerebral cortex ofpatients with sporadic CJD, incubated with tetra-cycline at a concentration ranging from 20 mM to2 mM, and then treated with proteinase K and ana-lyzed by Western blot. Since sporadic CJD may beassociated with two types of PrPSc (termed type 1and type 2) which differ in the size of the protease-resistant core, presumably as a consequence ofdifferences in conformation (Parchi et al., 1996,1997), both types of the abnormal protein wereused for the study. The incubation of PrPSc withtetracycline for 48 hours resulted in decreased pro-tease resistance, as revealed by immunoblot anal-ysis of the samples using the monoclonalantibodies 3F4 and SP214, which recognize epi-topes located in the mid and C-terminal region ofPrP, respectively (Figure 5(a)). Conversely, nochanges were observed when tetracycline wasadded to the samples immediately before protein-ase K digestion or after incubation of PrPSc with upto 2 mM gentamycin used as a negative control.The effect of tetracycline was more pronounced ontype 2 than type 1 PrPSc, despite the fact that theintrinsic protease resistance of both PrPSc isoformswas similar. After incubation with 20 mM, 200 mMand 2 mM tetracycline, the amount of type 2 PrPSc
remaining after proteinase K digestion wasreduced to 80.8(�5.6) %, 41.8(�2.9) % and19.5(�4.1) % of non-treated samples (100 %),respectively. On the other hand, a signi®cantincrease in susceptibility of type 1 PrPSc to protein-ase K was detected only at 2 mM tetracycline,

Figure 4. Interaction betweentetracycline and PrP 106-126 asdeduced by NMR. (a) Trace of a2D-NOESY spectrum of 3 mMtetracycline-HCl in 2H2O (pH 7.0),corresponding to 7-H proton.(b) and (c) Traces of 2D-NOESYspectra of 3 mM tetracycline-HCl in2H2O (pH 7.0), after addition of1.5 mM PrP 106-126. The tracescorrespond to tetracycline (b) 7-Hand (c) 8-H. (a) Note the change insign of the NOE peaks in compari-son to trace. The symbol * indicatesthe 2D-NOESY diagonal peaks.(d) Reference 1D-NMR spectrum ofthe mixture used in (b) and (c). Theassignment of protons involved inintermolecular interactions is dis-played. The inset shows the chemi-cal structure of tetracycline.
1314 Tetracycline Interacts with the Prion Protein
which caused a reduction of protease-resistant PrPto 45.0(�8.1) % of control values. To rule out thepossibility that these ®ndings could be due to over-stabilization of PrPSc aggregates impeding theentry of the protein into the gel as observed withCongo red (Caspi et al., 1998), some samples were
Table 1. NOE interactions observed for PrP10
PrP106-126
Val121/Val122 Meg 7-Val121/Val122 Hb 7-Ala117-119 Meb a 7-Leu125 Meg b 7-Leu125 Med 6-
Classi®ed on the basis of the observed intensity aa Residue not assigned due to the large overlappib Observed at pH 3.5.
treated with 3 M guanidine thiocyanate beforeSDS/polyacrylamide gel electrophoresis. Theresults were comparable to those observed withoutguanidine denaturation.
To investigate whether tetracycline could bemore effective when the drug-to-protein interaction
6-126 in the presence of tetracycline
Tetracycline
H,8-H,9-H,5a-H(w),6-H,4-NMe2(s), H4(w)H, 8-H, 9-H, 5a-H(w), 4-NMe2, H4(w)H, 8-H, 9-H, 6-H, 5a-H(w) , 4-NMe2(s), 4-H(w)H,8-HH,4-NMe2(w)
s strong (s), medium, weak (w).ng of Me signals.

Figure 5. Tetracycline reverts protease-resistance ofPrPSc from CJD brains. (a) Western blot analysis of theprotease-resistant core of PrPSc from a CJD patient. Thesample, corresponding to type 2 PrPSc, was incubatedalone (Ctr) or with increasing concentration of eithertetracycline or gentamycin, used as a negative control,for 48 hours and then subjected to proteinase K diges-tion. (b) Western blot analysis of type 1 PrPSc from aCJD patient. After reversible denaturation with guani-dine HCl, the sample was renatured in the absence orpresence of an increasing concentration of tetracycline,and then digested with proteinase K.
Tetracycline Interacts with the Prion Protein 1315
occurred during PrPSc folding, type 1 PrPSc,selected for being less susceptible to the com-pound, was subjected to reversible denaturationwith 2 M guanidine-HCl followed by renaturationin the presence of tetracycline at concentrationsranging from 20 nM to 20 mM. The samples werethen digested with proteinase K and analyzed byWestern blot with the antibodies 3F4 and SP214.The study showed that under these conditions thecompound was effective at nM concentration, thefraction of protease-resistant PrP being reducedto 78.1(�1.8) %, 62.6(�1.8) %, 50.4(�2.4) % and20.0(�2.8) % at 20 nM, 200 nM, 2 mM and 20 mMtetracycline, respectively (Figure 5(b)).
Tetracycline inhibits neuronal death andastroglial proliferation induced by PrP peptides
It has been shown that the peptide PrP 106-126 is toxic to neurons and stimulates astroglialproliferation in vitro (Forloni et al., 1993, 1994),similar to the protease-resistant core of PrPSc (DeArmond et al., 1988; MuÈ ller et al., 1993). Toevaluate whether these effects were antagonizedby tetracycline, primary cultures of rat hippo-campal or cortical neurons and astrocytes were
exposed for seven days to micromolar concen-tration of PrP 106-126 in the absence or presenceof the compound at 1:0.5 molar ratio. As shownin Figure 6, tetracycline alone did not affect theviability of neuronal cultures, which wasreduced by 50 % following prolonged exposureto PrP 106-126. The neurotoxicity of the peptidewas abolished by co-treatment of cultures withtetracycline. To verify the speci®city of this ®nd-ing, we assessed the effectiveness of the com-pound against the excitotoxic activity of kainicacid, an analog of glutamate (Forloni et al.,1997). Kainic acid reduced nerve cell viability by60 % in either the absence or presence of tetra-cycline, suggesting that the neuroprotectionobserved in the previous experiments was linkedto the interaction of the compound with PrP106-126 (Figure 6(f)). Tetracycline also abolishedthe effects of the peptide on astrocytes. Whilethe viability of primary astroglial cultures wasincreased by 35 % following long-term treatmentwith PrP 106-126, no signi®cant differences fromcontrols were observed when tetracycline wasadded to the culture medium with the peptide(Figure 7).
Discussion
The present study shows that tetracycline inter-acts with a synthetic peptide homologous to theamyloid protein puri®ed from GSS brains(Tagliavini et al., 1994) and a fragment thereofwhich is highly ®brillogenic, neurotoxic and glio-trophic in vitro (Forloni et al., 1993, 1994; Tagliaviniet al., 1993; Brown et al., 1996).
This interaction affects the aggregation proper-ties of the peptides and antagonizes their deleter-ious effects on nerve and glial cells. It is mostremarkable that tetracycline is able to interact withPrPSc extracted from brain tissue of CJD patientsand to revert the abnormal protein into a protease-sensitive form. Since PrPSc is thought to be theessential and possibly only component of the infec-tious prion particle (Prusiner, 1991; Prusiner et al.,1998), tetracycline can be regarded as a putativeanti-prion agent as well as a useful molecule toinvestigate the pathogenesis of prion diseases.
Previous studies showed that amyloid-bindingmolecules such as Congo red and iododoxorubicinare effective in cellular and/or animal models ofprion disease (Caughey & Race, 1992; Ingrossoet al., 1995; Tagliavini et al., 1997). Although themolecular basis of this activity is not well under-stood, it is possibly related to the propensity ofthese drugs to bind conformationally modi®edforms of the prion protein. The selection of tetra-cycline as test compound was based on structuralanalogies with Congo red, iododoxorubicin as wellas tetrapyrroles, a recently identi®ed class ofinhibitors of PrPSc formation (Caughey et al., 1998;Priola et al., 2000). All these molecules share a com-mon feature in that they contain an extended

Figure 6. Tetracycline hindersneurotoxicity induced by PrP106-126. (a)-(d) Photomicrographs ofprimary cultures of hippocampalneurons after 7 day exposure to (a)vehicle solution, (b) 40 mM tetra-cycline, (c) 80 mM PrP 106-126 or(d) 80 mM PrP 106-126 in the pre-sence of 40 mM tetracycline. Barrepresents 50 mm. (e) Viability ofprimary cultures of cortical neuronsafter 7 day exposure to vehicle sol-ution, 20 mM tetracycline, 40 mMPrP 106-126 or 40 mM PrP 106-126in the presence of 20 mM tetra-cycline. Values are the mean � SEof ®ve experiments. F interac-tion � p < 0.01 (two-way ANOVA).* p < 0.01 versus control group(Tukey's test). (f) Viability of pri-mary cultures of cortical neuronsafter 24 hour exposure to vehiclesolution, 20 mM tetracycline,250 mM kainic acid, or 250 mM kai-nic acid in the presence of 20 mMtetracycline. Values are the mean� SD of ®ve experiments. F inter-action � p not signi®cant (two-wayANOVA). * p < 0.01 versus controlgroup (Tukey's test).
1316 Tetracycline Interacts with the Prion Protein
hydrophobic core formed by aromatic moietieswhich enables interactions with lipophilic residuesof PrPSc. A second characteristic of these com-pounds is the presence of a variety of polar substi-tuents that can form hydrogen bonds with speci®cresidues of PrPSc, strengthening the drug-to-proteininteraction. The polar substituents have beenfound to play an important role in the in vitro inhi-bition of PrPSc formation by porphyrins and phtha-locyanines (Caughey et al., 1998). Preliminarystudies with tetracycline analogs suggest that boththe nature and position of polar substituents havesigni®cant effects on the ef®cacy of the compoundsto impede PrP peptide aggregation.
The ability of tetracycline to interact with hydro-phobic domains of PrP was veri®ed by NMR spec-troscopy, using a synthetic peptide homologous toresidues 106 to 126 of the human protein. Previousstudies showed that this peptide is highly suscep-
tible to structural transitions, although it tends toadopt a stable b-sheet conformation in bufferedsolutions (De Gioia et al., 1994). Noteworthily, PrP106-126 corresponds to a highly conserved regionof the protein located in the ¯exibly disordered N-terminal domain which is thought to undergo aprofound conformational change in PrPC to PrPSc
conversion (Riek et al., 1996, 1997; Donne et al.,1997). Binding of tetracycline to this or otherhydrophobic parts of the protein might interferewith the conversion process and/or PrPSc assem-bly. The observation that tetracycline hinders rena-turation of reversibly denatured PrPSc extractedfrom CJD brains and renders the protein suscep-tible to proteolysis supports the view that the com-pound acts on protein-folding intermediatesduring the conversion reaction (Weissmann, 1991;Safar et al., 1993). On the other hand, the strikingdecrease of protease resistance following direct

Figure 7. Tetracycline abolishes astroglial proliferationinduced by PrP 106-126. Viability of primary cultures ofastrocytes after seven day exposure to vehicle solution,40 mM tetracycline, 80 mM PrP 106-126 or 80 mM PrP106-126 in the presence of 40 mM tetracycline. Values arethe mean � SD of ®ve experiments. F interac-tion � p < 0.01 (two-way ANOVA). * p < 0.01 versus con-trol group (Tukey's test).
Tetracycline Interacts with the Prion Protein 1317
incubation of PrPSc with the drug, though at highconcentrations, suggests that tetracycline can alsodisrupt PrPSc aggregates and/or revert the confor-mation of the abnormal protein. The compoundwas more effective on type 2 than type 1 PrPSc.The basis of this variation in not known, but maybe a result of differences in conformation betweenthese PrPSc isoforms (Parchi et al., 1996). The poten-tial to affect macromolecular assemblies of PrPSc issustained by the ®nding of a progressive fragmen-tation and eventually disappearance of amyloid-like ®brils generated by PrP 106 to 126 followingprolonged exposure to the compound. Thus, thetype of interaction of tetracycline with PrPSc differsfrom that of Congo red, which causes overstabili-zation rather than destabilization of PrPSc mol-ecules (Caspi et al., 1998).
The ability of tetracycline to abolish neuronaldeath and astroglial proliferation induced by PrP106-126 in vitro was likely a result of its interactionwith the peptide. Previous studies showed that theneurotoxicity of PrP 106-126 is dependent upon theexpression of endogenous PrP, as neuronal culturesof PrP null mice survive peptide treatment (Brownet al., 1994). Further, the toxic effect is not strictlyrelated to the ®brillogenic properties of PrP 106-
126, since amidation of the C terminus remarkablyreduces amyloid ®bril formation but not neurotoxi-city (Salmona et al., 1999). This would imply thattetracycline acts not only on PrP 106-126 aggre-gates but also on oligomeric-monomeric forms ofthe peptide. Due to the complexity of the cell cul-ture system, other possible mechanisms of neuro-protection can be foreseen ranging from a directinteraction of tetracycline with endogenous PrP toindirect effects on the biology of the cell. On theother hand, the inhibition of PrP-induced astroglialproliferation probably relies on the anti-®brillo-genic capacity of tetracycline, since the effects ofPrP 106-126 on astrocytes is dependent on theaggregation state of the peptide (Peyrin et al., 1999;Salmona et al., 1999).
The need to develop therapies for prion dis-eases has remarkably increased following theemergence of the vCJD that seems to be causallylinked to BSE (Will et al., 1996; Collinge et al.,1996; Bruce et al., 1997). Although several com-pounds have been found to antagonize prionpropagation in vitro or in vivo, their suitabilityfor therapy is limited, primarily due to poorpassage of the blood/brain barrier and/or severetoxicity. Our data indicate that tetracycline caninterfere with PrPSc formation and protect fromPrP-induced neurotoxicity and glial activation. Itis conceivable that these effects are due to itsability to interact with both PrP aggregates andPrP monomers/oligomers, as indicated by ¯uor-escence microscopy and NMR spectroscopy, thusreducing the availability of altered PrP peptidesto act as a template for the conversion of thenormal protein. Tetracycline has been shown toreach plasma levels of 2-2.5 mg/l in man after asingle oral dose of 250 mg (Kapusnik-Uner et al.,1996). This value corresponds to a plasma con-centration of about 5 mM which we found to beeffective on PrPSc from CJD patients under con-ditions of refolding. Such action suggests thattetracycline could be regarded as a prophylacticagent which hinders formation of new PrPSc fol-lowing peripheral exposure to exogenous PrPSc,rather than a treatment once disease has pro-gressed. Thus, a potential interest of the com-pound could be conditions such as iatrogenicand vCJD. Preliminary studies show that tetra-cycline derivatives, such as doxycycline and min-ocycline, share similar activities but, at variancewith tetracycline, have the ability to cross theblood/brain barrier ef®ciently. Accordingly, thesetetracycline congeners represent good candidatesfor in vivo studies.
Materials and Methods
Peptide synthesis and purification
The peptides PrP 106-126 (KTNMKHMAGAAAA-GAVVGGLG), PrP 106-126 scrambled (NGAKALMGGHGATKVMVGAAA) and PrP 82-146(GQPHGGGWGQ GGGTHSQWNKPSKPKTNMKAG

1318 Tetracycline Interacts with the Prion Protein
AAAAGAVVGGLGGYMLGSAMSRPIIHFGSDYE) weresynthesized by solid phase chemistry on an Applied Bio-system 430A synthesiser as described (Forloni et al.,1993; Tagliavini et al., 1993). The peptides were cleavedfrom the resin with phenol/thioanisole/tri¯uoroaceticacid, precipitated and washed several times with colddiethylether, and puri®ed by preparative reverse-phaseHPLC (model 243, Beckman Instruments).The purity and identity of peptides were determined byanalytical reverse-phase HPLC, capillary electrophoresis(Quanta 4000, Millipore), amino acid sequencing (6600Prosequencer, Milligen) and mass spectrometry using asingle quadrupole mass spectrometer equipped with anelectro-spray interface (Hewlett-Packard). The purity ofthe peptides was greater than 95 %.
Binding of tetracycline to PrP peptides
Fluorescence microscopy
To generate amyloid ®brils, the peptides PrP 106-126and PrP 82-146 were suspended in 200 mM phosphatebuffer (pH 7.0) at a concentration of 0.5 mM, and incu-bated for one week at room temperature. Aqueous sol-ution of tetracycline hydrochloride (Sigma) or theamyloid-binding ¯uorochrome thio¯avine S was addedto suspensions of peptide aggregates to a ®nal concen-tration of 0.5 mM. After one hour incubation at roomtemperature, the samples were centrifuged, and the pel-let was washed several times with phosphate buffer andapplied to gelatin-coated slides. The samples were thenexamined by ¯uorescence microscopy using ®lters selec-tive for tetracycline or thio¯avine S (Zeiss).
Fluorescence spectroscopy
Tetracycline was dissolved in deionized water at con-centrations of 100, 250 and 500 mM. To investigate thebinding of the compound to PrP 106-126 and PrP 82-146,500 ml of peptide solution in deionized water was addedto the same volume of 200 mM tetracycline to ®nal pep-tide concentrations ranging from 16 to 800 mM. Thesamples were then incubated at 37 �C for one hour (PrP106-126) or four hours (PrP 82-146) and the excitationand emission spectra were determined with a PerkinElmer LS 50B spectrophoto¯uorimeter in the range of350-600 nm. The excitation and emission wavelengthsused to obtain the spectra of tetracycline alone were 422and 530 nm, and tetracycline with PrP peptides 410 and472 nm.
Sedimentation experiments
Peptides were dissolved in deionized water at a con-centration of 1 mM: 100 ml of 1 mM tetracycline solutionin deionized water was added to 100 ml of PrP 106-126or PrP 82-146, and the samples were incubated at 37 �Cfor one or four hours, respectively (solutions A and B).Then, 800 ml of 200 mM phosphate buffer (pH 7.4) wereadded to the samples which were kept at 37 �C for 0, 1,2, 4, 18, 24, 36, 48, 72 and 96 hours. After centrifugationat 13,000 g for ten minutes at 4 �C, 20 ml of supernatantwas analyzed by reverse-phase HPLC and the relativeamount of non-sedimentable peptide was calculated asthe percentage of peptides in solutions A and B.
Proteinase K digestion
PrP 106-126 and PrP 82-146 were dissolved in deio-nized water at a concentration of 2 mM: 20 ml of 2 mMtetracycline was added to the same volume of peptidesolution, and the samples were incubated at 37 �C forone or four hours, respectively. After addition of 40 ml of20 mM Tris-HCl (pH 7.4), containing 1 mM CaCl2, thesamples were further incubated at 37 �C for 18 hours,and then digested with proteinase K (37 �C, 30 minutes)at 1:100 (w/w) enzyme-to-substrate ratio. Proteolysiswas terminated by the addition of EGTA (5 mM ®nalconcentration). After centrifugation at 13,000 g for tenminutes at 4 �C, the pellets were dissolved in 80 ml of10 % formic acid containing 0.1 % tri¯uoroacetic acid and20 ml was analyzed by reverse-phase HPLC. Parallelsamples were run in the absence of tetracycline and ana-lyzed as speci®ed above with or without proteinase Kdigestion. The extent of proteolysis was calculated as thepercentage of peptide present in the pellet compared tothe total amount originally present.
Electron microscopy
The synthetic peptide PrP 106-126 was suspendedin 20 mM Tris-HCl (pH 7.0), at a concentration of0.5 mM either in the absence or in the presence ofequimolar concentration of tetracycline. Following 1,24 and 72 hours incubation at 37 �C, 5 ml aliquots ofpeptide suspension were applied to Formvar-coatednickel grids, negatively stained with 5 % (w/v) uranylacetate, and observed in an electron microscope (EM109, Zeiss) at 80 kV.
NMR spectroscopy
NMR samples were prepared by dissolving tetra-cycline and PrP 106-126 in 0.6 ml of H2O/2H2O mixture(9:1, v/v) (pH 3.5), or 0.01 M phosphate buffer (pH 7.0)in 2H2O. Final tetracycline and PrP concentrations wererespectively 1.5 and 3 mM. Deuterated solvents with99.9 % isotopic purity were purchased from Isotech. Sol-utions were immediately transferred into a 5 mm ODNMR tube. All experiments were performed at 600.13MHz on a Bruker AMX 600 NMR spectrometer at 298�K,using a 5 mm reverse-detection probe equipped withgradients. In the case of 2H2O solutions, the water signalwas suppressed by means of a NOESY-type pulsesequence with presaturation. In the case of H2O/2H2Osolutions, spectra were measured using gradient-basedpulse programs, capable of suppressing the water signaland minimize magnetization losses due to saturationtransfer. Chemical shifts were measured in d (ppm) andreferenced to a DSS solution sealed in a capillary, coaxialwith the NMR tube. Resonance assignments for tetra-cycline and PrP 106-126 were performed using well-established procedures (WuÈ thrich, 1986). Homonuclearcorrelation spectra, 2D COSY-dQF and 2D TOCSY wereobtained in phase-sensitive mode using standard pulseprograms.
Conversion of protease-resistant PrP from CJD to aprotease-sensitive form
Type 1 and type 2 PrPSc were partially puri®ed fromthe cerebral cortex of four patients with CJD following aprocedure described previously (Caughey et al., 1997)but with minor modi®cations. Sample aliquots contain-

Tetracycline Interacts with the Prion Protein 1319
ing approximately 2 ng of PrPSc were incubated at 37 �Cfor 48 hours either in the absence or in the presence oftetracycline (0.02-2 mM concentration) and then treatedwith proteinase K (100 mg/ml, 37 �C, one hour). Theamount of PrP remaining after proteolysis was assessedby Western blot analysis using the monoclonal anti-bodies 3F4 (1:50,000) and SP214 (1:200) which recognizeepitopes located in the mid and C-terminal regions ofPrP, respectively (Kascsak et al., 1987; Rogers et al., 1991;JimeÂnez-Huete et al., 1998). PrP immunoreactive bandswere visualized by ECL and their average signal inten-sity was quanti®ed by densitometry. Values wereexpressed as the percentage of signal intensity ofsamples not treated with tetracycline. Control exper-iments included: (i) the incubation of samples with0.02-2 mM gentamicin instead of tetracycline; (ii) theaddition of tetracycline to the samples immediatelybefore proteinase K digestion; and (iii) the treatment ofproteinase K-digested samples with 3 M guanidinethiocyanate (pH 2.5), for ten minutes, followed bythe addition of bovine serum albumin (BSA) to a ®nalconcentration of 200 mg/ml and methanol-precipitation,prior to SDS/polyacrylamide gel electrophoresis andimmunoblot (Caspi et al., 1998).
To investigate whether tetracycline could be moreeffective during PrPSc folding, type 1 PrPSc was subjectedto reversible denaturation with guanidine-hydrochloride(GdnHCl, 2 M ®nal concentration) for 16 hours at 37 �C.The mixture was then diluted to 0.75 M GdnHCl usingTBS (10 mM Tris-HCl (pH 7.5), 150 mM NaCl) sup-plemented with cetylpyridinium chloride (1.5 mM ®nalconcentration) and tetracycline was added to ®nal con-centrations of 0.02-20 mM. The samples were incubatedat 37 �C for 48 hours, further diluted to 0.375 M GdnHClwith TBS and digested with proteinase K (50 (mg/ml,37 �C, one hour). The digestion was terminated by theaddition of phenylmethylsulfonyl ¯uoride (1 mM ®nalconcentration); the proteins were then precipitated withten volumes of methanol after addition of BSA, and ana-lyzed by Western blot.
Neuronal and astroglial cultures
Neurons
Neuronal cultures were prepared and characterized asreported previously (Forloni et al., 1993). In brief, brainswere removed from fetal rats on embryonic day 17. Hip-pocampal and cortical cells were dissociated in serum-free medium containing 0.1 % (w/v) trypsin (DIFCO)and 25 mg/ml deoxyribonuclease for ®ve minutes atroom temperature, and plated (5 � 105 cells/ml) in 24 or96-well dishes pre-coated with poly-D-lysine (50 mg/ml;SIGMA). The cells were cultured in basal medium Eagle(BME-Hanks' salt, GIBCO) supplemented with 10 %(w/v) fetal calf serum (FCS, GIBCO) and 2 mM gluta-mine. Cultures were kept at 37 �C in a water-saturatedatmosphere of 95 % air, 5 % CO2. After ®ve days in vitro,non-neuronal cell division was halted by exposure to10ÿ5 M cytosine arabinoside, an inhibitor of mitosis, toprevent overgrowth of glial cells.
Astrocytes
Glial cell cultures were prepared from newborn ratpups. The brain tissue was separated from meninges anddissociated by trituration with a Pasteur pipette. Glialcells were grown in Dulbecco's modi®ed minimal essen-tial medium (DMEM; GIBCO) supplemented with 10 %
FCS and 2 mM glutamine, and cultured at 37 �C in awater-saturated atmosphere of 95 % air, 5 % CO2. Afterten days of culture, ¯asks containing mixed glial cultureswere given fresh medium and placed on a shaker at37 �C for 12-16 hours to separate astrocytes from micro-glia and oligodendrocytes (Yao et al., 1992). The adherentcells (astrocytes) were then exposed for ®ve minutes to0.25 % trypsin, followed by the addition of an equalvolume of DMEM/FCS 10 %. The suspension was centri-fuged and the pellet was resuspended in medium con-taining 10 % FCS. Cells were plated at a density of5 � 104 cells/ml (Forloni et al., 1997).
Cell treatments
Neuronal cultures were exposed for 7 days to PrP106-126 alone or in combination with tetracycline(1:0.5 molar ratio) starting from the ®rst plating day.PrP peptide concentrations were 40 and 20 mM forhippocampal and cortical neurons, respectively. Toverify the speci®city of the protective effect of tetra-cycline against PrP neurotoxicity, cortical neuronswere treated with 250 mM kainic acid alone or in com-bination with 20 mM tetracycline for 24 hours at day 6of culture (Forloni et al., 1997). The astrocytes wereexposed for seven days to 40 mM PrP 106-126 aloneor in combination with tetracycline (1:0.5 molar ratio)starting seven days after plating. Control cultureswere treated with vehicle solution, a scrambledsequence of PrP 106-126 or tetracycline alone. Follow-ing treatment, neuronal cell death and astroglial pro-liferation were quantitatively assessed using crystalviolet staining (0.5 % in 20 % (v/v) methanol) andcomputer-assisted densitometric analysis (Andreoniet al., 1998).
Acknowledgments
This work was supported, in part, by the Italian Min-istry of Health, Department of Social Services, Telethon-Italy (Grant E.574) and the European Community(BMH4 CT-98-6011 and CT-98-6051). We are grateful toR.J. Kascsak and F. Prelli for providing us with the anti-bodies 3F4 and SP214.
References
Adnjou, K. T., Demaimay, R., Lasmezas, C., Deslys, J. P.,Seman, M. & Dormont, D. (1995). MS-8209, a newamphotericin B derivative, provides enhanced ef®-cacy in delaying hamster scrapie. Antimicrob. AgentsChemother. 39, 2810-2812.
Andreoni, G., Angeretti, N., Lucca, E. & Forloni, G.(1998). Densitometric quanti®cation of neural viabi-lity by computerized image analysis. Expt. Neurol.148, 281-287.
Bessen, R. A., Kocisko, D. A., Raymond, G. J., Nandan,S., Lansbury, P. T., Jr & Caughey, B. (1995). Non-genetic propagation of strain-speci®c properties ofscrapie prion protein. Nature, 375, 698-700.
Brown, D. R., Herms, J. & Kretzschmar, H. A. (1994).Mouse cortical cells lacking cellular PrP survive inculture with a neurotoxic PrP fragment. Neuroreport,5, 2057-2060.

1320 Tetracycline Interacts with the Prion Protein
Brown, D. R., Schmidt, B. & Kretzschmar, H. A. (1996).Role of microglia and host prion protein in neuro-toxicity of a prion protein fragment. Nature, 380,345-347.
Bruce, M. E., McBride, P. A. & Farquhar, C. F. (1989).Precise targeting of the pathology of the sialoglyco-protein PrP and vacuolar degeneration in mousescrapie. Neurosci. Letters, 102, 1-6.
Bruce, M. E., Will, R. G., Ironside, J. W., McConnell, I.,Drummond, D., Suttle, A., McCardle, L., Chree, A.,Hope, J., Birkett, C., Cousens, S., Fraser, H. &Bostock, C. J. (1997). Transmission to mice indicatethat ``new variant'' CJD is caused by the BSE agent.Nature, 389, 498-501.
Caspi, S., Halimi, M., Yanai, A., Ben Sasson, S.,Taraboulos, A. & Gabizon, R. (1998). The anti-prionactivity of Congo red. J. Biol. Chem. 273, 3484-3489.
Caughey, B. & Race, R. E. (1992). Potent inhibition ofscrapie-associated PrP accumulation by Congo red.J. Neurochem. 59, 768-771.
Caughey, B. & Raymond, G. J. (1993). Sulfated poly-anion inhibition of scrapie-associated PrP accumu-lation in cultured cells. J. Virol. 67, 643-650.
Caughey, B. W., Dong, A., Bhat, K. S., Ernst, D., Hayes,S. F. & Caughey, W. S. (1991). Secondary structureanalysis of the scrapie-associated protein PrP 27-30in water by infrared spectroscopy. Biochemistry, 30,7672-7680.
Caughey, B., Kocisko, D. A., Priola, S. A., Raymond,G. J., Race, R. E., Bessen, R. A., Lansbury, P. T., Jr& Chesebro, B. (1997). Methods for studying prionprotein (PrP) metabolism and the formation ofprotease-resistant PrP in cell culture and cell-freesystems. In Methods in Molecular Medicine: Prion Dis-eases (Baker, H. & Ridley, R. M., eds), pp. 285-300,Humana Press, Totowa.
Caughey, W. S., Raymond, L. D., Horiuchi, M. &Caughey, B. (1998). Inhibition of protease-resistantprion protein formation by porphyrins and phtalo-cyanines. Proc. Natl Acad. Sci. USA, 95, 12117-12122.
Collinge, J., Sidle, K. C. L., Heads, J., Ironside, J. & Hill,A. F. (1996). Molecular analysis of prion strain vari-ation and the aetiology of new variant CJD. Nature,383, 685-680.
De Armond, S. J., Gonzales, M., Mobley, W. C., Kon,A. A., Sern, H. & Prusiner, S. B. (1988). PrPSc inscrapie-infected hamster brain is spatially and tem-porally related to histopathology and infectivitytiter. In Alzheimer's Disease and Related Disorders.Progress in Clinical and Biological Research (Ikbal, K.,Wisniewski, H. M. & Winblad, B., eds), pp. 601-618,Alan R. Liss, New York.
De Armond, S. J., Yang, S.-L., Lee, A., Bowler, R.,Taraboulos, A., Groth, D. & Prusiner, S. B. (1993).Three scrapie prion isolates exhibit differentaccumulation patterns of the prion protein scrapieisoform. Proc. Natl Acad. Sci. USA, 90, 6449-6453.
De Gioia, L., Selvaggini, C., Ghibaudi, E., Diomede, L.,Bugiani, O., Forloni, G., Tagliavini, F. & Salmona,M. (1994). Conformational polymorphism of theamyloidogenic and neurotoxic peptide homologousto residues 106-126 of the prion protein. J. Biol.Chem. 269, 7859-7862.
Diringer, H. & Ehlers, B. (1991). Chemoprophylaxis ofscrapie in mice. J. Gen. Virol. 72, 457-460.
Donne, D. G., Viles, J. H., Groth, D., Mehlhorn, I.,James, T. L., Cohen, F. E., Prusiner, S. B., Wright,P. E. & Dyson, H. J. (1997). Structure of the recom-binant full-length hamster prion protein PrP (29-
231): the N terminus is highly ¯exible. Proc. NatlAcad. Sci. USA, 94, 13452-13457.
Ehlers, B. & Diringer, H. (1984). Dextran sulphate 500delays and prevents mouse scrapie by impairmentof agent replication in spleen. J. Gen. Virol. 65, 1325-1330.
Farquhar, C. F. & Dickinson, A. G. (1986). Prolongationof scrapie incubation period by an injection of dex-tran sulphate 500 within the month before or afterinjection. J. Gen. Virol. 67, 463-473.
Forloni, G., Angeretti, N., Chiesa, R., Monzani, E.,Salmona, M., Bugiani, O. & Tagliavini, F. (1993).Neurotoxicity of a prion protein fragment. Nature,362, 543-546.
Forloni, G., Del, Bo R., Angeretti, N., Chiesa, R.,Smiroldo, S., Doni, R., Ghibaudi, E., Salmona, M.,Porro, M., Verga, L., Giaccone, G., Bugiani, O. &Tagliavini, F. (1994). A neurotoxic prion proteinfragment induces rat astroglial proliferation andhypertrophy. Eur. J. Neurosci. 6, 1415-1422.
Forloni, G., Lucca, E., Angeretti, N., Chiesa, R. &Vezzani, A. (1997). Neuroprotective effect ofsomatostatin on non-apoptotic NMDA-inducedneuronal death: role of cGMP. J. Neurochem. 68, 319-327.
Ghani, A. C., Ferguson, N. M., Donnelly, C. A.,Hagenaars, T. J. & Anderson, R. M. (1998). Epide-miological determinants of the pattern and magni-tude of the vCJD epidemic in Great Britain. Proc.Roy. Soc. London, 265, 2443-2452.
JimeÂnez-Huete, A., Lievens, P. M. J., Vidal, R., Piccardo,P., Ghetti, B., Tagliavini, F., Frangione, B. & Prelli,F. (1998). Endogenous proteolytic cleavage of nor-mal and disease-associated isoforms of the humanprion protein in neural and non-neural tissues. Am.J. Pathol. 153, 1561-1572.
Ingrosso, L., Ladogana, A. & Pocchiari, M. (1995).Congo red prolongs the incubation period inscrapie-infected hamsters. J. Virol. 69, 506-508.
Kapusnik-Uner, J. E., Sande, M. A. & Chambers, F.(1996). Antimicrobial agents. In The PharmacologicalBasis of Therapeutics (Harman, J. G., Limbird, L. E.,Molinof, P. B. & Ruddon, R. W., eds), 9th edit., pp.1123-1130, MaGraw-Hill, New York.
Kascsak, R. J., Rubenstein, R., Merz, P. A., Tonna-DeMasi, M., Fersko, R., Carp, R. I., Wisniewski,H. M. & Diringer, H. (1987). Mouse polyclonal andmonoclonal antibody to scrapie-associated ®brilproteins. J. Virol. 61, 3688-3693.
Kimberlin, R. H. & Walker, C. A. (1983). The antiviralcompound HPA-23 can prevent scrapie whenadministered at the time of infection. Arch. Virol.78, 9-18.
Kimberlin, R. H. & Walker, C. A. (1986). Suppression ofscrapie infection in mice by heteropolyanion 23,dextran sulfate, and some other polyanions. Antimi-crob. Agents Chemother. 30, 409-413.
Kocisko, D. A., Come, J. H., Priola, S. A., Chesebro, B.,Raymond, G. J., Lansbury, P. T., Jr & Caughey, B.(1994). Cell-free formation of protease-resistantprion protein. Nature, 370, 471-474.
Merlini, G., Ascari, E., Amboldi, N., Bellotti, V.,Arbustini, E., Perfetti, V., Ferrari, M., Zorzoli, I.,Marinone, M. G., Garini, P., Diegoli, M., Trizio, D.& Ballinari, D. (1995). Interaction of the anthra-cycline 40-iodo-40-deoxydoxorubicin with amyloid®brils: inhibition of amyloidogenesis. Proc. NatlAcad. Sci. USA, 92, 2959-2963.

Tetracycline Interacts with the Prion Protein 1321
MuÈ ller, W. E. G., Ushijima, H., SchroÈder, H. C., Forrest,J. M. S., Shatton, W. F. H., Rytik, P. G. & Heffner-Lauc, M. (1993). Cytoprotective effects of NMDAreceptor antagonists on prion protein (PrionSc)-induced toxicity in rat cortical cell cultures. Eur. J.Pharmacol. 246, 261-267.
Neuhaus, D. & Williamson, M. P. (1986). The kinetics ofthe NOE. In The Nuclear Overhauser Effect in Struc-tural and Conformational Analysis (Neuhaus, D. &Williamson, M. P., eds), pp. 103-140, VCHPublishers Inc., New York.
Pan, K. M., Baldwin, M., Nguyen, J., Gasset, M., Serban,A., Groth, D., Mehlhron, I., Huang, Z., Fletterick,R. J., Cohen, F. E. & Prusiner, S. B. (1993). Conver-sion of a-helices into b-sheets features in the for-mation of the scrapie prion proteins. Proc. NatlAcad. Sci. USA, 90, 10962-10966.
Parchi, P., Castellani, R., Capellari, S., Ghetti, B., Young,K., Chen, S. G., Farlow, M., Dickson, D. W., Sima,A. A. F., Trojanowski, J. Q., Petersen, R. B. &Gambetti, P. (1996). Molecular basis of phenotypicvariability in sporadic Creutzfeldt-Jacob disease.Ann. Neurol. 39, 767-778.
Parchi, P., Capellari, S., Chen, S. G., Petersen, R. B. &Gambetti, P. (1997). Typing prion isoforms. Nature,386, 232-234.
Peyrin, J.-M., Lasmezas, C. I., Haik, S., Tagliavini, F.,Salmona, M., Willians, A., Richie, D., Deslys, J.-P.& Dormont, D. (1999). Microglial cells respond toamyloidogenic PrP peptides by the production ofin¯ammatory cytokines. Neuroreport, 10, 723-729.
Pocchiari, M., Schmittinger, S. & Masullo, C. (1987).Amphotericin B delays the incubation period ofscrapie in intracerebrally inoculated hamsters. J. Gen.Virol. 68, 219-223.
Priola, S. A., Raines, A. & Caughey, W. S. (2000). Por-phyrin and phthalocyanine antiscrapie compounds.Science, 25, 1503-1506.
Prusiner, S. B., McKinley, M. P., Bowman, K. A., Bolton,D. C., Bendheim, P. E., Groth, D. F. & Glenner,G. G. (1983). Scrapie prions aggregate to form amy-loid-like birefringent rods. Cell, 35, 349-358.
Prusiner, S. B. (1991). Molecular biology of prion dis-eases. Science, 252, 1515-1522.
Prusiner, S. B., Scott, M. R., De Armond, S. J. &Cohen, F. E. (1998). Prion protein biology. Cell,93, 337-348.
Riek, R., Hornemann, S., Wider, G., Billeter, M.,Glockshuber, R. & WuÈ thrich, K. (1996). NMR struc-ture of the mouse prion protein domain PrP (121-231). Nature, 382, 180-182.
Riek, R., Hornemann, S., Wider, G., Glockshutber, R. &WuÈ thrich, K. (1997). NMR characterization of thefull-length recombinant murine prion protein, mPrP(23-231). FEBS Letters, 413, 282-288.
Rogers, M., Serban, D., Gyuris, T., Scott, M., Torchia, T.& Prusiner, S. B. (1991). Epitope mapping of theSyrian hamster prion protein utilizing chimeric andmutant genes in a vaccinia virus expression system.J. Immunol. 147, 3568-3574.
Safar, J., Roller, P. P., Gajdusek, D. C. & Gibbs, C. J.(1993). Conformational transitions, dissociation andunfolding of scrapie amyloid (prion) protein. J. Biol.Chem. 268, 20276-20284.
Salmona, M., Malesani, P., De Gioia, L., Gorla, S.,Bruschi, M., Molinari, A., Della, Vedova F.,Pedrotti, B., Marrari, A. M., Awan, T., Bugiani, O.,Forloni, G. & Tagliavini, F. (1999). Molecular deter-minants of the physicochemical properties of a
critical prion protein region comprising residues106-126. Biochem. J. 342, 207-214.
Sande, M. A. & Mandell, G. L. (1991). Antimicrobialagents. In The Pharmacological Basis of Therapeutics(Gilman, A. G., Rall, T. W., Nies, A. S. & Taylor, P.,eds), pp. 1117-1145, Pergamon Press, New York.
Selvaggini, C., De Gioia, L., CantuÁ , L., Ghibaudi, E.,Diomede, L., Passerini, F., Forloni, G., Bugiani, O.,Tagliavini, F. & Salmona, M. (1993). Molecularcharacteristics of a protease-resistant, amyloidogenicand neurotoxic peptide homologous to residues106-126 of the prion protein. Biochem. Biophys. Res.Commun. 194, 1380-1386.
Soto, C., Kascsak, R. J., Saborio, G. P., Aucouturier, P.,Wisniewski, T., Prelli, F., Kascsak, R., Mendez, E.,Harris, D. A., Ironside, J., Tagliavini, F., Carp, R. I.& Frangione, B. (2000). Reversion of prion proteinconformational changes by synthetic b-sheet breakerpeptides. Lancet, 355, 192-197.
Supattapone, S., Nguyen, H. O., Cohen, F. E.,Prusiner, S. B. & Scott, M. R. (1999). Eliminationof prions by branched polyamines and impli-cations for therapeutics. Proc. Natl Accad. Sci.USA, 96, 14529-14534.
Tagliavini, F., Prelli, F., Ghiso, J., Bugiani, O., Serban,D., Prusiner, S. B., Farlow, M. R., Ghetti, B. &Frangione, B. (1991). Amyloid protein of Gerst-mann-StraÈussler-Scheinker disease (Indiana kindred)is an 11 kd fragment of prion protein with an N-terminal glycine at codon 58. EMBO J. 10, 513-519.
Tagliavini, F., Prelli, F., Verga, L., Giaccone, G., Sarma,R., Gorevic, P., Ghetti, B., Passerini, F., Ghibaudi,E., Forloni, G., Salmona, M., Bugiani, O. &Frangione, B. (1993). Synthetic peptides homologousto prion protein residues 106-147 form amyloid-like®brils in vitro. Proc. Natl Acad. Sci. USA, 90, 9678-9682.
Tagliavini, F., Prelli, F., Porro, M., Rossi, G., Giaccone,G., Farlow, M. R., Dlouhy, S. R., Ghetti, B., Bugiani,O. & Frangione, B. (1994). Amyloid ®brils in Gerst-mann-StraÈussler-Scheinker disease (Indiana andSwedish kindreds) express only PrP peptidesencoded by the mutant allele. Cell, 79, 695-703.
Tagliavini, F., McArthur, R. A., Canciani, B., Giaccone,G., Porro, M., Bugiani, M., Lievens, P. M.-J.,Bugiani, O., Peri, E., Dall'Ara, P., Rocchi, M., Poli,G., Forloni, G., Bandiera, T. & Varasi, M., et al.(1997). Effectiveness of anthracycline against exper-imental prion disease in syrian hamsters. Science,276, 1119-1122.
Weissmann, C. (1991). A uni®ed theory of prion propa-gation. Nature, 352, 679-683.
Will, R. G., Ironside, J. W., Zeidler, M., Cousens, S. N.,Estibeiro, K., Alperovitch, A., Poser, S., Pocchiari,M., Hofman, A. & Smith, P. G. (1996). A new var-iant of Creutzfeldt-Jakob disease in the UK. Lancet,347, 921-925.
Williams, A., Lucassen, P. J., Ritchie, D. & Bruce, M.(1997). PrP deposition, microglial activation andneuronal apoptosis in murine scrapie. Expt. Neurol.14, 433-438.
Williamson, D. E. & Everett, G. W., Jr (1975). A protonnuclear magnetic resonance study of the site ofmetal binding in tetracycline. J. Am. Chem. Soc. 97,2397-2407.
WuÈ thrich, K. (1986). Sequence-speci®c resonance assign-ments in proteins. In NMR of Proteins and NucleicAcids (WuÈ thrich, K., ed.), pp. 130-161, John Wileyand Sons, New York.

1322 Tetracycline Interacts with the Prion Protein
Xi, Y. G., Ingrosso, L., Ladogana, A., Masullo, C. &Pocchiari, M. (1992). Amphotericin B treatment dis-sociates in vivo replication of the scrapie agent fromPrP accumulation. Nature, 356, 598-601.
Yao, J., Keri, J. E., Taffs, R. E. & Colton, C. A.(1992). Characterization of ineterleukin-1 product-ion by microglia in colture. Brain Res. 591, 88-93.
Edited by J. Karn
(Received 7 February 2000; received in revised form 8 May 2000; accepted 9 May 2000)
Copyright © 2022 FDOKUMEN




















