
SPATIAL RELATIONSHIPS IN EARLY SIGNALING EVENTS OF FLOW-MEDIATED ENDOTHELIAL MECHANOTRANSDUCTION
23
Annu. Rev. Physiol. 1997. 59:527–49 Copyright c 1997 by Annual Reviews Inc. All rights reserved SPATIAL RELATIONSHIPS IN EARLY SIGNALING EVENTS OF FLOW-MEDIATED ENDOTHELIAL MECHANOTRANSDUCTION Peter F. Davies 1 , Kenneth A. Barbee 2 , MichaelV. Volin 3 , Andre Robotewskyj 3 , Jai Chen 4 , Loren Joseph 3 , Melvin L. Griem 5 , Miles N. Wernick 6 , Elizabeth Jacobs 7 , Denise C. Polacek 1 , Natacha DePaola 8 , and Abdul I. Barakat 3 1 Institute for Medicine and Engineering, University of Pennsylvania, Philadelphia, Pennsylvania 19104; 2 Department of Neurosurgery, Hahnemann Medical College of Pennsylvania, Philadelphia, Pennsylvania 19102; Departments of 3 Pathology, 4 Radiology, and 5 Radiation and Cellular Oncology, University of Chicago, Chicago, Illinois 60637; 6 Department of Electrical and Computer Engineering, Illinois Institute of Technology, Chicago, Illinois 60616; 7 Department of Medicine, Medical College of Wisconsin, Milwaukee, Wisconsin 53226; and 8 Department of Biomedical Engineering, Rensselaer Polytechnic Institute, Troy, New York 12181 KEY WORDS: endothelium, biomechanics, hemodynamic shear stress, blood flow, vasoregula- tion, atherosclerosis ABSTRACT Blood flow interactions with the vascular endothelium represent a specialized example of mechanical regulation of cell function that has important physiologi- cal and pathological cardiovascular consequences. The endothelial monolayer in vivo acts as a signal transduction interface for forces associated with flowing blood (hemodynamic forces) in the acute regulation of artery tone and chronic structural remodeling of arteries, including the pathology of atherosclerosis. Mechanisms related to spatial relationships at the cell surfaces and throughout the cell that influence flow-mediated endothelial mechanotransduction are discussed. In par- ticular, flow-mediated ion channel activation and cytoskeletal dynamics are con- sidered in relation to topographic analyses of the luminal and abluminal surfaces of living endothelial cells. 527 0066-4278/97/0315-0527$08.00
-
Upload
hms-harvard -
Category
Documents
-
view
1 -
download
0
Transcript of SPATIAL RELATIONSHIPS IN EARLY SIGNALING EVENTS OF FLOW-MEDIATED ENDOTHELIAL MECHANOTRANSDUCTION

P1: SBS/rsk P2: sbs/PLB QC: sbs
August 28, 1956 23:59 Annual Reviews DAVICHPT.TXT AR025-23
Annu. Rev. Physiol. 1997. 59:527–49Copyright c© 1997 by Annual Reviews Inc. All rights reserved
SPATIAL RELATIONSHIPSIN EARLY SIGNALING EVENTS OFFLOW-MEDIATED ENDOTHELIALMECHANOTRANSDUCTION
Peter F. Davies1, Kenneth A. Barbee2, Michael V. Volin3, AndreRobotewskyj3, Jai Chen4, Loren Joseph3, Melvin L. Griem5,Miles N. Wernick6, Elizabeth Jacobs7, Denise C. Polacek1,Natacha DePaola8, and Abdul I. Barakat31Institute for Medicine and Engineering, University of Pennsylvania, Philadelphia,Pennsylvania 19104;2Department of Neurosurgery, Hahnemann Medical Collegeof Pennsylvania, Philadelphia, Pennsylvania 19102; Departments of3Pathology,4Radiology, and5Radiation and Cellular Oncology, University of Chicago, Chicago,Illinois 60637;6Department of Electrical and Computer Engineering, Illinois Instituteof Technology, Chicago, Illinois 60616;7Department of Medicine, Medical Collegeof Wisconsin, Milwaukee, Wisconsin 53226; and8Department of BiomedicalEngineering, Rensselaer Polytechnic Institute, Troy, New York 12181
KEY WORDS: endothelium, biomechanics, hemodynamic shear stress, blood flow, vasoregula-tion, atherosclerosis
ABSTRACT
Blood flow interactions with the vascular endothelium represent a specializedexample of mechanical regulation of cell function that has important physiologi-cal and pathological cardiovascular consequences. The endothelial monolayer invivo acts as a signal transduction interface for forces associated with flowing blood(hemodynamic forces) in the acute regulation of artery tone and chronic structuralremodeling of arteries, including the pathology of atherosclerosis. Mechanismsrelated to spatial relationships at the cell surfaces and throughout the cell thatinfluence flow-mediated endothelial mechanotransduction are discussed. In par-ticular, flow-mediated ion channel activation and cytoskeletal dynamics are con-sidered in relation to topographic analyses of the luminal and abluminal surfacesof living endothelial cells.
5270066-4278/97/0315-0527$08.00

P1: SBS/rsk P2: sbs/PLB QC: sbs
August 28, 1956 23:59 Annual Reviews DAVICHPT.TXT AR025-23
528 DAVIES ET AL
INTRODUCTION
As blood flows through arteries, it imparts physical forces to the vascular wallthat regulate a number of important physiological responses in blood vesselsand are also implicated in the development of arterial wall pathologies. Changesin blood flow are responsible for the acute regulation of vessel tone, the de-velopment of blood vessel structure during embryogenesis and early growth,and the chronic remodeling of adult blood vessels following interventions thatalter the mechanical environment. Hemodynamic forces can be resolved intotwo principal vectors: first, shear stress, a frictional force acting at the inter-face between flowing blood and the vessel wall; and second, pressure actingnormal to the vessel wall, which imposes circumferential stretch to the tis-sue (24, 33). It is now well accepted that hemodynamic shear stress actsthrough the endothelium to regulate both acute vessel tone and chronic restruc-turing of blood vessels (19, 23). Thus the endothelium is a complex mechan-ical signal-transduction interface between flowing blood and the vessel wall(70).
In contrast to physiological regulation, the role of hemodynamics in patholog-ical events is less well understood. The most notable association is that regionsof disturbed flow correlate with the distribution of atherosclerotic lesions inlarge arteries, a relationship that has been recognized since the 1850s (83).Some regions of the arterial tree never succumb to atherosclerosis, whereasothers are highly susceptible. These areas usually coincide with regions ofseparation from unidirectional laminar flow that occur typically near branches,bifucations, regions of arterial narrowing, and curvatures in the arteries. The lo-cal hemodynamic forces generate complex gradients of shear stress and producehighly uneven stretch forces within the vessel wall. Consequently, theories ofhigh shear stress, low shear stress, particle residence time in the bulk fluid, anddifferential arterial compliance and stretch have all gained a degree of credibilityas causative factors of focal atherosclerosis (13, 32, 87). How the endotheliumplays a dominant hemodynamic role in such a complex environment is unclear,although at least one major endothelial dysfunction is associated with the de-veloping lesions: the failure of both flow-mediated and chemically regulatedvasodilation (37). Other vascular pathologies in which blood flow plays animportant role include the clotting and fibrinolytic cascades at the blood ves-sel surface, where critical concentrations of enzymes, substrates, and cofactorsresponsible for clot formation and dissolution are influenced by the local flowcharacteristics that regulate the convective delivery and removal of reagents(26, 35).
This article reviews spatial relationships that influence signaling responsesinitiated by hemodynamic shear stresses in endothelial cells. Cell imaging

P1: SBS/rsk P2: sbs/PLB QC: sbs
August 28, 1956 23:59 Annual Reviews DAVICHPT.TXT AR025-23
ENDOTHELIAL FLOW SIGNALING 529
techniques, in providing geometric information at cell surfaces, have prompteda reexamination of the localization of mechanotransduction sites. The readeris also referred to a recent review of flow-mediated endothelial mechanotrans-duction mechanisms (19).
Endothelial Bioresponses to Hemodynamic Forces:A HierarchyA sense of order can be created from the bewildering array of endothelialresponses to shear stress by arranging them in order of response times; a hi-erarchy of potential relationships can then be postulated. A prominent featureis that they range across a variety of biological disciplines, from electrophys-iology to biochemistry to cell biology, gene regulation and major changes ofcell structure (Figure 1). These diverse responses are often contemporaneous.For example, very early electrophysiological changes of membrane potential(seconds) and the activation of potent biochemical cascades appear to havesimilar time constants (8). The latter include G protein activation, the mobi-lization of phosphoinositide derivatives, release of intracellular free calcium,
Figure 1 Endothelial mechanotransduction: shear stress. IP3, inositol-1,4,5-trisphosphate; DAG,diacylglycerol; PG, prostaglandins; NO, nitric oxide; MAP, mitogen-activated protein; NFkB,nuclear factor kappa B; SSRE, shear stress response element; PDGF-B, platelet derived growthfactor B chain; bFGF, basic fibroblast growth factor; NOS, nitric oxide synthase; tPA, tissueplasminogen activator; TGFb, transforming growth factor beta; ICAM-1, intercellular adhesionmolecule-1; MCP-1, monocyte chemoattractant protein-1; HSP-70, 70-kDa heat shock protein;ET-1, endothelin-1; MTOC, microtubule-organizing center; Tm, thrombomodulin; Fn, fibronectin.

P1: SBS/rsk P2: sbs/PLB QC: sbs
August 28, 1956 23:59 Annual Reviews DAVICHPT.TXT AR025-23
530 DAVIES ET AL
and cyclic nucleotide phosphorylation (9, 30). Structural changes, such as re-arrangements of actin and vimentin filaments, which are generally observedmuch later in the sequence of mechanotransduction events and are associatedwith changes of cell shape, are also detectable by sensitive techniques soonafter the initiation of a change of mechanical load (19, 31). The time constantsare similar to those associated with activation of some of the known early tran-scription factors (49). Although many responses support an ordered sequenceof initial signaling leading to transcription factor activation, gene regulation andcellular structural adaptation (associated with the synthesis of new proteins),there are also shorter signaling pathways. An example is shown in Figure 1 asrelease of the potent vasodilators nitric oxide (NO) and prostacyclin. By re-quiring only acute signaling, a vasoregulatory outcome can be stopped quicklyor reversed if the mechanical environment changes. Therefore, it is not neces-sary for the cells to undergo major restructuring in response to reversible acutemechanical events. On the other hand, if the new mechanical environmentis sustained, transcriptional, posttranscriptional, and posttranslational eventscause an appropriate functional and/or structural reorganization that may beconsidered an adaptive response. For example, the alignment of endothelialcells in the direction of flow is a usual outcome of exposure to directional flow(25, 60).
Between these acute and chronic responses are many interacting components.Two transcription factor families that are present in the cytoplasm of most cells,including endothelium, areRel-related nuclear factor kappa B (NFkB) and nu-clear factor activator protein-1 (AP1). Both are stimulated by shear stress(49), and NFkB p50-p65 complex binds to a recently described shear stressresponse element (SSRE) identified as a consensus sequence found in severalflow-responsive genes (68, 69). It has recently become apparent that there aremultiple interactive SSREs in the endothelium. Shyy et al (77, 78) have shownthat flow-induced expression of the endothelial early response gene, monocytechemoattractant protein-1 (MCP-1), is regulated through shear stress–sensitivecis-acting sequences in the 5′ promoter. A phorbol ester response element thatbinds the transcription factor AP1 was found to be essential for shear responses.Furthermore, recent studies of gap junction connexin gene expression in cul-tured endothelial cells subjected to flow demonstrate upregulation of connexin43 mRNA despite the absence of known SSREs in the connexin gene (DCPolacek et al, manuscript submitted). These experiments suggest that multiplecis response elements are involved in shear stress–induced gene regulation. Bydefining the temporal and spatial framework within which the bioresponsesoccur, a better understanding of the multiple interactions should follow.
There are four major areas currently under investigation within the generalpathways of flow-mediated mechanotransduction. The first is to address the

P1: SBS/rsk P2: sbs/PLB QC: sbs
August 28, 1956 23:59 Annual Reviews DAVICHPT.TXT AR025-23
ENDOTHELIAL FLOW SIGNALING 531
question: How is a mechanical force converted by the endothelium into an initialresponse? The second is to elucidate the pathways leading to rapid release ofNO and other vasodilators and to determine if these initial pathways are alsocommon to other downstream responses, particularly gene regulation. Studiesof transcription factors activated by shear stress and their interactions withDNA of (mechanoresponsive) genes in the nucleus constitute the third researchfocus. The fourth area is the study of alterations of cell structure, particularlythose involving the cytoskeleton. Following initial signaling, it is likely thatthese areas overlap and interact extensively, and cellular adaptation to flow, aslow process, may influence initial signaling. However, spatial relationshipsare currently most accessible for investigation of the first and fourth of theseareas, the initial cell response and the cytoskeleton.
IMAGING THE LUMINAL ENDOTHELIAL SURFACE
When hemodynamic shear stress acts at the endothelial cell surface, the stressdistribution at the surface and throughout the cell is determined by the three-dimensional shape of the luminal surface (Figure 2). Most studies have assumedthe shear stresses to be those calculated for a smooth surface without referenceto the individual cell topography. However, each cell responds to the forcesacting locally at its surface. Thus characterization of flow forces on a cellular
Figure 2 Schematic of an endothelial cell illustrating the importance of spatial considerations inrelation to hemodynamic shear stress. The three-dimensional surface geometry determines stressconcentratins at the cell surface that are transferred to the interior of the cell via the cytoskeleton.N, nucleus.

P1: SBS/rsk P2: sbs/PLB QC: sbs
August 28, 1956 23:59 Annual Reviews DAVICHPT.TXT AR025-23
532 DAVIES ET AL
Figure 3 AFM images of part of a confluent monolayer of bovine aortic endothelial cells in culture.PanelsA, D: Surface geometry before and after alignment with flow. PanelsB, E: Three-dimensionaldistribution of shear stress over the luminal endothelial surface calculated by finite element analysis(bulk fluid shear stress 12 dyn/cm2). PanelsC, F: Distribution of stress concentrations (shear stressgradients) at the luminal surface. (From Reference 6, reprinted with permission.)
scale, requiring detailed cell surface geometry, is essential. In a theoreticalstudy of flow over a sinusoidal undulating surface simulating the endothelium,Satcher et al (73) demonstrated variations in shear stress over the model cellsurface that were strongly dependent on the surface geometry. The model wasextended to living cells in near real-time by Barbee et al (5, 6) using atomic forcemicroscopy (AFM) to map out the real surface geometry. In the absence of flow,bovine aortic endothelial cells in confluent cultures were polygonal in shapewith quite smooth surfaces. Boundaries between cells were visible and theaverage differential from the highest point of the cells (over the nuclear region)to the lowest point at junctions between cells was severalµm (Figure 3A).After monolayers had been subjected to directional flow for 24 h, the cellsaligned with the flow and AFM measurements were repeated. There was asmall decrease in the average range of surface heights compared with controlcells, but a significant decrease in the amplitude of surface undulations i.e.streamlining of the cells. An 11◦ average slope in control cells was reducedto an 8◦ incline in the flow-aligned cells with significant implications for thedistribution of stresses.

P1: SBS/rsk P2: sbs/PLB QC: sbs
August 28, 1956 23:59 Annual Reviews DAVICHPT.TXT AR025-23
ENDOTHELIAL FLOW SIGNALING 533
Stress Concentrations Determined by Surface GeometryThe AFM geometric data provided≈105 x, y,andzcoordinates for each cell. Byanalyzing AFM data from a representative region of the confluent monolayerin a computer simulation of flow, the distribution of shear stress on a cell-by-cell basis was calculated using the computational fluid dynamics programNEKTON (6). As shown in Figures 3BandC, in which gray scales representingshear stress and shear stress gradients, respectively, in the flow direction weresuperimposed on the cell contours, it can be seen that the stress distribution isdetermined by the topographic undulations. This is true for both control cellsand those aligned by flow. The average peak stress per cell was reduced in thealigned monolayers as was the peak shear stress gradient per cell. The relativeareas exposed to extremes of both shear stress and shear stress gradient weresmaller in aligned cells compared with non-aligned cells.
Heterogeneity of Endothelial Flow ResponsesA prominent feature of Figure 3, particularly in aligned cells, is the displayof cell-to-cell differences in stress concentrations that arise from the hetero-geneity of topography. The large variations in stress concentrations betweenneighboring cells may help explain the frequently observed differential endothe-lial responses to flow observed both in vivo and in vitro. Examples include theexpression of VCAM-1 protein in vivo (84) and in vitro (63), VCAM-1 mRNAin vivo (56), ICAM-1 protein expression in vivo (84) and in vitro (58), eleva-tion of intracellular calcium ([Ca2+]i ) measured in vitro (34, 76) and in vivo(27), induction of synthesis and nuclear relocalization of c-fos protein in vitro(67), expression of major histocompatibility complex (MHC) antigens in vitro(55), inhibition of endothelial cell division in vitro (88), and relocalization ofthe Golgi apparatus and microtubule-organizing center (MTOC) in vitro (16).In each of these cases, high levels of response in one or a group of cells isaccompanied by absent or diminished responses in adjacent cells of the sameendothelial monolayer despite exposure to an identical flow field in vitro orlocation in a predicted uniform hemodynamic environment in vivo. Becausethe responses are specifically flow-related, very local regulatory mechanismsmust be responsible for the differences. A plausible explanation is the hetero-geneous topography that exists in the monolayer. The sensitivity of a cell maybe determined by the fraction of cell surface exposed to stress concentrationsabove a critical threshold (20).
Critical Factors in the Location of Flow SensorsA similar argument can be made for individual flow sensors. In addition tocell-to-cell variations in stress distribution, variations in hemodynamic forceson a subcellular scale may also be important. Because the force distribution

P1: SBS/rsk P2: sbs/PLB QC: sbs
August 28, 1956 23:59 Annual Reviews DAVICHPT.TXT AR025-23
534 DAVIES ET AL
Figure 4 Proposed influence of endothelial surface topography upon the sensitivity of putativemechanosensors (open circle) at the lumenal cell surface following cell alignment by shear stress.At position 1, the shear stresses and stress gradients are similar, but at position 2, the stressconcentration (1τ ) is markedly reduced in the aligned cell. Cell responses may depend on both thedistribution of mechanosensors (or surface elements linked to them) and the altered topography.
is spatially variable, the precise location of any primary flow sensor at the lu-minal endothelial surface is relevant to that particular cell’s ability to respond.As shown simplistically in Figure 4, if there is a limited number of flow sen-sors at the luminal surface, those grouped at position 1 will be exposed tovirtually identical (maximal) stress concentration whether aligned or not. If,however, sensors are located where the subcellular geometries in the two cellconfigurations are very different (position 2), the stress concentrations in oneconfiguration may be above or below an activation threshold. The conceptcan be extended to any model of localized mechanical sensitivity involving amodest number of flow sensors per cell; e.g. flow-sensitive transmembrane pro-teins linked to the intracellular cytoskeleton that concentrate the stresses intoand throughout the cell body (85). Thus varying surface geometries in a flowenvironment can directly influence cell tension (43, 44).
Endothelial Surface Geometry Is Similar In Vitro and In SituTo explore the relevance of in vitro measurements of cell topography to in vivogeometry, the endothelium of a rabbit aorta was imaged by AFM (20). Atpresent, a limitation of this approach is the variable waviness of underlying

P1: SBS/rsk P2: sbs/PLB QC: sbs
August 28, 1956 23:59 Annual Reviews DAVICHPT.TXT AR025-23
ENDOTHELIAL FLOW SIGNALING 535
arterial wall structures that often exceeds the range of vertical movement of theAFM stylus. This results in reliable imaging of only relatively small regionsof the arterial surface; however, although limited in area, accurate detailedtopographic data was recorded. An AFM image obtained from the abdominalaorta is shown in Figure 5 (top panel). The luminal surfaces of five cells arevisible aligned in the direction of flow. They were of a size and shape thatresembled those of cells aligned by flow in vitro. A comparison of selectedlongitudinal height profiles (lower panels) demonstrates the similarity of boththe general shape of the profile (e.g. height-to-length ratio) and the relativesmoothness of the surface in both in vivo and in vitro images (20). These imagesare the first detailed measurements of endothelial surface geometry in livingarteries. They strongly suggest that the surface contours of endothelial cellsmaintained in culture are very similar to those observed in situ, a finding thatsupports the validity in vivo of conclusions derived from in vitro measurementsof cell surface geometry.
LOCATION OF FLOW-SENSITIVE K+ CHANNELS
An important early endothelial response to shear stress, now known to be en-tirely or partly localized to the luminal membrane, is activation of inwardlyrectifying K+ (IRK) channels, first identified by Olesen et al (65). When flowwas imposed, a membrane current developed as a function of shear stress withhalf-maximal activation at 0.7 dyn/cm2. Channel activity reached a plateaunear 20 dyn/cm2. On the basis of ion selectivity and reversal potential, thecurrent was identified as an inwardly rectifying K+ current and was designatedIKS. The current was rapidly activated in response to flow, only slowly desen-sitized (minutes), and was inactivated when flow was stopped. Neither atrialmyocytes nor vascular smooth muscle cells expressedIKS. Subsequent studiesof single IRK channel recordings in cell-attached patches (46) demonstratedflow-associated hyperpolarization and activation of large inward and small out-ward whole-cell currents, consistent with the previous whole-cell recordingsand confirming localization to the luminal cell surface. The absence of largeconductance single channel currents and insensitivity to charybdotoxin sug-gested that maxi-K channels did not contribute to flow responses. The openstate probability of individual IRK channels was increased within minutes ofexposure to shear stress, reflecting a delayed opening and closing response whencompared with shear stress-activated whole cell currents. Considering that therecordings were made from patches that were not directly exposed to the shearstress (being protected by the micropipette), the findings suggest that the chan-nel is activated secondarily to other signaling events initiated by shear stresselsewhere on the cell surface and transmitted to the channels in the micropipette.

P1: SBS/rsk P2: sbs/PLB QC: sbs
August 28, 1956 23:59 Annual Reviews DAVICHPT.TXT AR025-23
536 DAVIES ET AL
Figure 5 Upper panel. AFM raw image of the endothelium of rabbit aorta in situ. Cells arealigned with the longitudinal axis of the vessel in a straight segment with no branches.Lowerpanels. Longitudinal sections of a representative cell from an aligned culture (in vitro) and fromrabbit aortic endothelium (in situ). The sections are similar in height-to-length ratio and in theshape of the profile. (Reprinted from Reference 20, with permission.)

P1: SBS/rsk P2: sbs/PLB QC: sbs
August 28, 1956 23:59 Annual Reviews DAVICHPT.TXT AR025-23
ENDOTHELIAL FLOW SIGNALING 537
This could occur by a dependence upon membrane hyperpolarization effectedby IRK channels outside of the patch, or by mechanical (cytoskeletal) or chem-ical (e.g. G proteins, calcium) linking to primary mechanosensors elsewhere.Some evidence for a link to calcium was obtained by the finding that the openprobability of endothelial IRK channels in inside-out patches excised from thecell surface was increased twofold by exposure to micromolar concentrationsof calcium (46). Because these studies were limited to the luminal membrane,they confirm the existence of flow-sensitive IRK channels only at that site. Itremains unclear, however, whether the channels themselves are flow sensors;the delayed IRK responses in patches may occur by activation secondary to anupstream sensor.
These studies are consistent with measurements of unidirectional Rb+ ef-flux from endothelium that confirm a shear stress-dependent plasma membranepermeability to K+ (2), and recordings of endothelial hyperpolarization dur-ing flow using potentiometric dyes (59). Cooke and colleagues (17, 62) usedpharmacologic inhibitors to demonstrate the association of flow-sensitive en-dothelial K+ channel activity with the release of an endogenous nitrovasodilatorin arterial rings. They have also provided circumstantial evidence that activa-tion of a K+ channel is associated with G-protein coupling and the elevation ofendothelial cGMP. Although voltage-gated calcium channels are not present incultured endothelial cells (1, 64, 80), hyperpolarization can lead to increasedcalcium influx via a calcium/phosphatidylinositol/hyperpolarization-activatedCa2+-permeable channel (51, 52, 64), and hyperpolarization increases the elec-trochemical gradient for calcium, resulting in calcium influx (39). The oppositeeffect, depolarization, attenuates cellular functions that rely upon calcium in-flux, e.g. NO release, vasodilation (14, 52).
Endothelial hyperpolarization as a function of shear stress was also reportedusing membrane potential–sensitive fluorescent dyes (59). Recent studies (ABarakat & PF Davies, manuscript submitted) using the voltage-sensitive dyebisoxonol have revealed a simultaneous activation by flow of hyperpolarizingand depolarizing responses. A dominant initial hyperpolarization was reversedso that the cell reached a net depolarized state after several minutes. If IRKchannels were blocked with barium or cesium, immediate depolarization wasrecorded. Depolarization was blocked by chloride current inhibitors, suggestingthe involvement of flow-sensitive chloride channels. When chloride channelswere inhibited, a prolonged hyperpolarization resulted.
Olesen & Bundgaard (64) have demonstrated that an inwardly rectifying K+
channel in bovine aortic endothelial cells requires phosphorylation in order toremain open. Inside-out patches required administration of ATP to the cy-tosolic side in order to maintain ion channel activity. These findings suggestthat a prominent inwardly rectifying K+ channel in endothelial cells has some

P1: SBS/rsk P2: sbs/PLB QC: sbs
August 28, 1956 23:59 Annual Reviews DAVICHPT.TXT AR025-23
538 DAVIES ET AL
similar characteristics to the rat kidney channel cloned by Ho et al (40) and forwhich a putative ATP-binding site implicated in phosphorylation was identi-fied. Evidence that flow-activated K+ channels may operate independently ofchemically sensitive K+ channels has recently been published (41).
Attempts to clone endothelial K+ channels on the basis of structural ho-mology to channels identified in other tissues have met with limited success.However, a IRK channel has recently been cloned from human umbilical veinendothelium that contains two transmembrane-spanning regions flanking a poreregion and an extraordinarily long extracellular loop. The channel has been des-ignated endothelial IRK (E-IRK; MV Volin et al, manuscript submitted). E-IRKmRNA is expressed strongly in Northern blots of cultured endothelial, but notsmooth muscle cells, and exclusively in endothelial cells of rabbit arterial tis-sue probed by in situ hybridization. E-IRK is the first K+ channel cloned fromendothelial cells (subfamily KIR 2.3); further studies will determine if it is flowsensitive (see note added in proof).
IMAGING THE ABLUMINAL ENDOTHELIALSURFACE ADHESION SITES
Focal Adhesion DynamicsWhatever the stress distribution induced by flow on the luminal endothelialsurface, resistance to cell detachment and rolling is localized to adhesion siteson the abluminal surface. We propose that some aspects of flow-mediated sig-naling in the endothelium could be located at adhesion sites, the stress forcesbeing transferred across the cell body through the cytoskeleton (19, 21–23,25). All anchorage-dependent cells, including the endothelial lining of bloodvessels, attach to the extracellular matrix at specialized regions called focaladhesion sites (12, 45, 79). In both in vivo and in vitro at such sites, the ex-tracellular domains of integral membrane proteins bind to adhesion proteins ofthe matrix or substratum. In vitro, the separation distances between a protein-coated glass surface and the cell approaches 10–15 nm, allowing imaging ofthe adhesion sites by interference reflection microscopy (IRM) made possibleby differences in refractive index of the components (36). Following initialobservations of cell-substratum adhesion sites in Swiss 3T3 cells by tandemscanning confocal microscopy (TSCM), the dynamic nature of these sites inliving endothelial cells was demonstrated using TSCM, digital image process-ing, and three-dimensional image reconstruction (22, 23) (Figure 6). In theraw image, TSCM combines IRM principles with confocal optical sectioningcapability. In an unperturbed confluent monolayer of endothelial cells adherentto a glass substratum coated with adhesion proteins, focal adhesion remodeling

P1: SBS/rsk P2: sbs/PLB QC: sbs
August 28, 1956 23:59 Annual Reviews DAVICHPT.TXT AR025-23
ENDOTHELIAL FLOW SIGNALING 539
Figure 6 Endothelial abluminal cell surface geometry observed in real-time by tandem scanningconfocal microscopy after image processing and computer enhancement. Reconstructed three-dimensional image of the abluminal surface of a living endothelial cell. The substratum has beenremoved from the image to expose the surface topography. Membrane projections represent focaladhesion sites where the cell is attached to the underlying (invisible) extracellular matrix.
occurred in a random fashion without any preferred orientation (21). Adhesionsites occasionally disappeared, sometimes in less than 1 min; in other cases, thearea of contact remained constant but changed considerably in shape and size.The appearance of new sites was observed where contact had not previouslyexisted between the cell and the adhesion proteins. Throughout these adhesiondynamics, the shape of individual cells in the monolayer remained unchanged,the overall effect being that of stationary cells constantly probing and reprobingthe substratum. Observations during directional flow showed complex changesof focal adhesion site dynamics, the net effect of which was site remodelingrelated to the direction of the luminal shear stress (22).
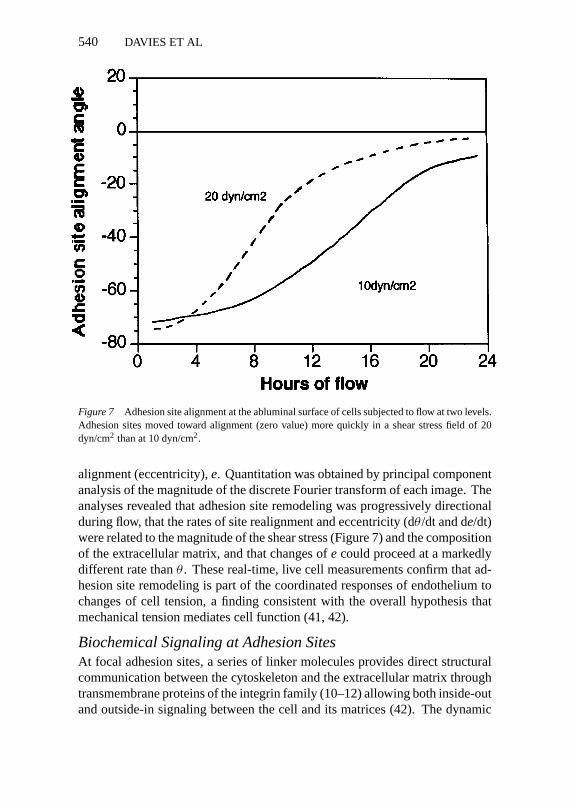
Recently, quantitative analyses of site orientation and alignment during flowhave been performed by expanding the use of the Fourier transform to quan-tify the complex nature of the changes in endothelial adhesion sites duringcontinuous exposure to flow forces (M Wernick et al, manuscript submitted).Two-dimensional directional remodeling of adhesions in cells exposed to flowwas analyzed for changes in the principal axis of alignment,θ , and the degree of
P1: SBS/rsk P2: sbs/PLB QC: sbs
August 28, 1956 23:59 Annual Reviews DAVICHPT.TXT AR025-23
540 DAVIES ET AL
Figure 7 Adhesion site alignment at the abluminal surface of cells subjected to flow at two levels.Adhesion sites moved toward alignment (zero value) more quickly in a shear stress field of 20dyn/cm2 than at 10 dyn/cm2.
alignment (eccentricity),e. Quantitation was obtained by principal componentanalysis of the magnitude of the discrete Fourier transform of each image. Theanalyses revealed that adhesion site remodeling was progressively directionalduring flow, that the rates of site realignment and eccentricity (dθ /dt and de/dt)were related to the magnitude of the shear stress (Figure 7) and the compositionof the extracellular matrix, and that changes ofe could proceed at a markedlydifferent rate thanθ . These real-time, live cell measurements confirm that ad-hesion site remodeling is part of the coordinated responses of endothelium tochanges of cell tension, a finding consistent with the overall hypothesis thatmechanical tension mediates cell function (41, 42).
Biochemical Signaling at Adhesion SitesAt focal adhesion sites, a series of linker molecules provides direct structuralcommunication between the cytoskeleton and the extracellular matrix throughtransmembrane proteins of the integrin family (10–12) allowing both inside-outand outside-in signaling between the cell and its matrices (42). The dynamic

P1: SBS/rsk P2: sbs/PLB QC: sbs
August 28, 1956 23:59 Annual Reviews DAVICHPT.TXT AR025-23
ENDOTHELIAL FLOW SIGNALING 541
nature of adhesion sites in endothelial and other anchorage-dependent cellsis best explained as a rapid turnover of one or more key structural elementssuch that the sites constantly rearrange in area and position (19). This is aparticularly active mechanism in migrating cells. If adhesion sites are involvedin hemodynamic signaling, timing is important because a delayed responsemay imply that the remodeling is occurring secondary to overall cytoskeletalreorganization, whereas a rapid response may imply a primary signaling role inmechanotransduction. The TSCM studies demonstrated directional remodelingof adhesion sites with variable time constants ranging from almost immediateresponses to significant delays.
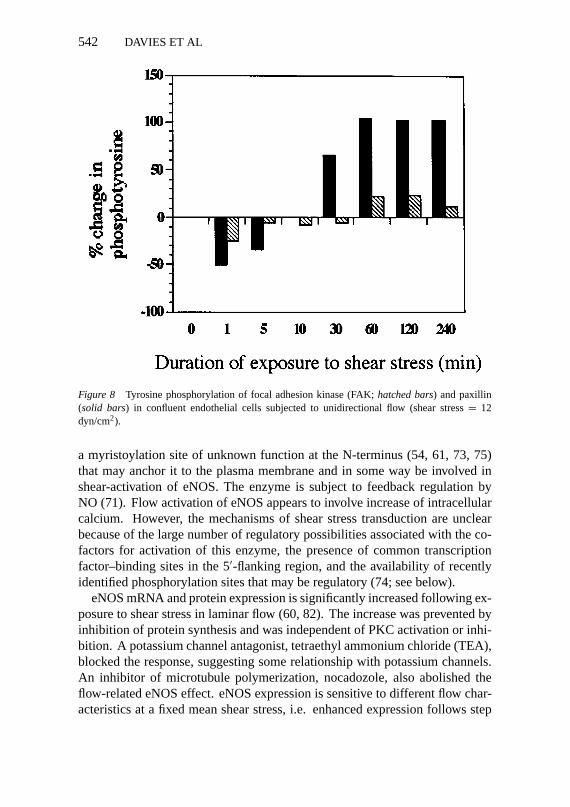
Potential flow-signaling molecules localized to focal adhesion sites includetyrosine, threonine, and serine kinases. The phosphorylation levels of intracel-lular proteins associated with adhesion sites, which are implicated in integrin-mediated cell adhesion, have been studied in endothelial cells subjected to flow.Anti-phosphotyrosine Western blots of abstracts of confluent endothelial cellsdemonstrated two prominent groups of protein at 120–130 kDa and 65–75 kDathat were identified as the proteins focal adhesion kinese (FAK) and paxillin,respectively. Both proteins were shown to be associated with endothelial focaladhesion sites by immunfluorescence. When compared with no-flow controlendothelial monolayers, phosphorylation of paxillin in bovine aortic endothelialcells increased several-fold within 2 h of exposure to unidirectional laminar flow(12 dyn/cm2 shear stress), whereas in contrast, FAK phosphorylation remainedwithin 20% of control levels (Figure 8). This was confirmed in Western blots ofimmunoprecipitated FAK and paxillin. The enhanced tyrosine phosphorylationof paxillin was preceded by a 50% decrease during the first minute, suggestingenhanced phosphatase activity. Measurements of FAK enzyme activity dur-ing flow revealed no changes. However, in human umbilical vein endothelialcells, Berk et al (B Berk, personal communication) have observed enhancedFAK tyrosine phosphorylation several minutes after initiation of flow withoutsignificant changes in paxillin phosphorylation levels. Although these data arepreliminary, they suggest that protein kinase activity at adhesion sites may beinvolved in the hemodynamic response of endothelial cells to shear stress.
FLOW-SENSITIVE SIGNALING PATHWAYS FOR NITRICOXIDE AND OTHER ACUTE VASOREGULATORS
The mechanism(s) of flow induced NO-mediated vasodilation are likely reg-ulated by endothelial nitric oxide synthase (eNOS) activity. The enzyme isconstitutively expressed at a basal level and its activity appears to be calcium/calmodulin-dependent (29, 50). Protoporphyrin IX, FAD, FMN, and tetrahy-drobiopterin (74) are required cofactors (75). A unique feature of eNOS is
P1: SBS/rsk P2: sbs/PLB QC: sbs
August 28, 1956 23:59 Annual Reviews DAVICHPT.TXT AR025-23
542 DAVIES ET AL
Figure 8 Tyrosine phosphorylation of focal adhesion kinase (FAK;hatched bars) and paxillin(solid bars) in confluent endothelial cells subjected to unidirectional flow (shear stress= 12dyn/cm2).
a myristoylation site of unknown function at the N-terminus (54, 61, 73, 75)that may anchor it to the plasma membrane and in some way be involved inshear-activation of eNOS. The enzyme is subject to feedback regulation byNO (71). Flow activation of eNOS appears to involve increase of intracellularcalcium. However, the mechanisms of shear stress transduction are unclearbecause of the large number of regulatory possibilities associated with the co-factors for activation of this enzyme, the presence of common transcriptionfactor–binding sites in the 5′-flanking region, and the availability of recentlyidentified phosphorylation sites that may be regulatory (74; see below).
eNOS mRNA and protein expression is significantly increased following ex-posure to shear stress in laminar flow (60, 82). The increase was prevented byinhibition of protein synthesis and was independent of PKC activation or inhi-bition. A potassium channel antagonist, tetraethyl ammonium chloride (TEA),blocked the response, suggesting some relationship with potassium channels.An inhibitor of microtubule polymerization, nocadozole, also abolished theflow-related eNOS effect. eNOS expression is sensitive to different flow char-acteristics at a fixed mean shear stress, i.e. enhanced expression follows step

P1: SBS/rsk P2: sbs/PLB QC: sbs
August 28, 1956 23:59 Annual Reviews DAVICHPT.TXT AR025-23
ENDOTHELIAL FLOW SIGNALING 543
changes in flow. eNOS expression in vivo correlates well with situations ofincreased or decreased flow (47).
eNOS, in common with other NOSs, contains consensus sequences for phos-phorylation by protein kinases A and C, and calmodulin kinase II (74). Rapidlyincreased phosphorylation of serine or threonine residues upon exposure toshear stress has been reported (18). In addition, there is a calcium/calmodulin-independent mechanism of activation of eNOS (66, 81). In cultured endothe-lium, flow-induced NO release is biphasic, with an initial rapid rise at the onsetof flow followed by slow development of a sustained increased level of produc-tion (7). The initial peak NO release is Ca2+ dependent but independent of shearstress magnitude. In contrast, the sustained phase is Ca2+ independent but de-pendent on shear stress magnitude. Flow appears to stimulate Ca2+-dependentNO release in periodic bursts at approximately 15 min intervals, whereas theCa2+-independent NO release is continuous and sustained. Recently, Ayajikiet al (4) reported that the Ca2+-dependent phase may be an in vitro artifact. Inexcised segments of rabbit iliac artery, they at first found both Ca2+-dependentand Ca2+-independent responses to be similar to those noted in vitro. How-ever, when the arterial segments were restored to their in vivo lengths, theCa2+-dependent phase was abolished and their data suggest that NO produc-tion proceeds principally through a Ca2+-independent mechanism in vivo. Ifthis is true, then the focus of eNOS control shifts toward other pathways. Aprominent candidate pathway is the phosphorylation of mitogen-activated pro-tein (MAP) kinases (81). MAP kineses are activated independently of Ca2+.Activation is inhibited by interference with the activity of protein kinase C. Alink to upstream G protein signaling was provided by the demonstration thatMAP kinase activation is preventable by exposing the cells to nonhydrolyzableGDP analogues. In vivo, inhibition of protein tyrosine kinase by herbimycinabolishes flow-dependent NO release in intact arteries (4). All these studiessuggest that tyrosine phosphorylation is required for the activation of eNOS butthe spatial relationships remain unclear.
THE ENDOTHELIAL CYTOSKELETONIN FLOW SIGNALING
Stress TransmissionCells rely upon a combination of structural and chemical elements for normalfunction; the two are closely intertwined. For example, cell shape determined bythe cytoskeleton mediates cell growth and differentiation (42). The cytoskeletondetermines the shape of anchorage-dependent cells and physically connectsmany cell components thereby generating cell tension. When endothelial cells

P1: SBS/rsk P2: sbs/PLB QC: sbs
August 28, 1956 23:59 Annual Reviews DAVICHPT.TXT AR025-23
544 DAVIES ET AL
are subjected to flow, the cytoskeletal tension is likely to change. This hasnot been directly measured (although it is now feasible), but is supported by anumber of experiments.
Change in cell shape and cytoskeletal distribution was one of the earliestobservations in cultured endothelium subjected to directional flow, resultingin cell alignment (24, 60). The reorganization of adhesion sites during flow isbest explained by transfer of stress across the cell from the luminal to abluminalsurfaces. At the luminal surface, Wang et al (85) have demonstrated the transferof twisting forces across the luminal endothelial membrane via transmembraneintegrins, resisted by cytoskeletal components. At adhesion sites on the ablumi-nal surface, cytoskeletal elements are linked to integrins that bind extracellularadhesion proteins (11). Thus there is continuity of structural elements acrossthe cell that are involved in mechanotransduction.
The cytoskeleton might be considered to be involved in shear stress mechan-otransduction at every level. It determines cell geometry including the luminaltopography. It maintains cell tension at (a) adhesion sites, critical locationsregulating many aspects of cell function; (b) cell junctions, which regulate con-tact inhibition of endothelial growth and cell-cell communication; and (c) thenuclear membrane, which may be physically important for gene regulation. Itinteracts with biochemical pathways throughout the cell including those reg-ulating its own turnover, and it is implicated in mechanosensitive ion channelactivation (72). The three principal cytoskeletal groups of proteins—actin mi-crofilaments, intermediate filaments, and microtubules—participate in poorlyunderstood ways in flow-mediated mechanotransduction.
Actin has been most widely studied (24, 31, 38, 86). Microfilament reor-ganization in flow results in the assembly of bundles oriented in the directionof flow (38), possibly associated with new actin synthesis (31). Changes inactin distribution are detectable within minutes of the onset of flow (PF Davies,unpublished observations) and are probably faster, observations being limitedonly by technical constraints.
Endothelial intermediate filaments have recently been shown to respond toflow (28). The cells contain a dense, vimentin-containing intermediate filamentnetwork that extends throughout the cytoplasm and provides mechanical con-tinuity between luminal and abluminal surfaces and the nucleus. Within 2 hof flow, the network loses some continuity and short discontinuous strands andglobular foci of vimentin are present in many cases. With continued flow, thisintermediate state resolved to a reorganized, less dense network aligned withthe long axis of the cell (and flow direction). Two additional findings in thisstudy were the association of intermediate filaments with focal adhesion sitesand the rapid redistribution of an intermediate filament-binding protein. Thelatter may also associate with adhesion sites and be a link to actin filaments andother proteins located there (FW Flitney et al, unpublished studies).

P1: SBS/rsk P2: sbs/PLB QC: sbs
August 28, 1956 23:59 Annual Reviews DAVICHPT.TXT AR025-23
ENDOTHELIAL FLOW SIGNALING 545
Microtubules have not been extensively studied in mechanotransduction re-search. Recently, however, Malek & Izumo (53) have shown that microtubuledisruption by nocodazole blocked cell alignment by flow and the induction ofactin stress fibers. They extended these findings to show that cell shape changewas dependent on tyrosine kinase activity and intracellular calcium concen-tration, although the precise link to the cytoskeleton was not identified. Cellalignment was not dependent on intermediate filaments.
Three-Dimensional ImagingProgress in understanding the role of the cytoskeleton in mechanotransduc-tion is enhanced by direct observation of the spatial relationships within livingendothelial cells, an approach applied to surfaces by AFM and TSCM tech-niques, as described above. Improvements in laser confocal microscopy andgreater computational power applied to conventional fluorescence microscopeimages promises to revolutionize real-time visualization of spatial elements(15). Deconvolution of stacked images obtained by optical sectioning of indi-vidual cells removes out-of-focus information. Extension of these approachesto living cells looks promising with the introduction of green fluorescent proteingene constructs that provide endogenous fluorescence to the cytoskeleton.
FLOW-MEDIATED CHEMICAL SIGNALING
The spatial relationships discussed throughout this review are most relevant toshear stress forces that result in displacement of specific localized membranesensors and/or multiple connected components extending throughout the cell.However, labile chemicals at the cell surface may activate endothelial flow re-sponses independently of physical displacement of the cell (reviewed in 19).Thus when high local concentrations of labile agonists are released close tothe endothelial surface, the probability of interaction with endothelial receptorsis greatly influenced by flow; increased flow improves convective delivery tothe receptors, whereas decreased flow slows convection. When the removalrate exceeds convective and diffusive delivery rates from the bulk fluid, a steepconcentration gradient exists between the fluid and the cell surface, and this inturn will be influenced by the flow characteristics. It appears that flow-mediatedchemical responses and shear stress displacement responses may occur inde-pendently of each other. When shear stress was greatly increased by changingthe fluid viscosity, with only small changes in mass transport, flow-mediatedrelaxation of intact arteries was enhanced, indicating that the physical forcewas the principal effector (48, 57). Ando et al (3) have also provided evidencefor a direct mechanism of force transduction in evoking [Ca2+]i responses thatis additional to the effects of flow upon mass transport. Thus consideration ofthe potency of the local chemical environment should be included whenever

P1: SBS/rsk P2: sbs/PLB QC: sbs
August 28, 1956 23:59 Annual Reviews DAVICHPT.TXT AR025-23
546 DAVIES ET AL
possible in the interpretation of flow experiments, and the spatial relationshipswithin chemical gradients near the cell surface can become a complex factor inflow-mediated cellular responses. Both physical displacement of the cell andchemical concentrations may interact to evoke a mechano-chemical transduc-tion response.
ACKNOWLEDGMENTS
This work is supported by SCOR grant HL15062 and HL36049 MERIT Award(PFD) from the National Heart Lung and Blood Institute of the National Instituteof Health, and Grant-in-Aid 91-1557 from the American Heart Association.
Literature Cited
1. Adams DJ, Barakeh J, Laskey R, vanBreemen C. 1989. Ion channels and reg-ulation of intracellular calcium in vascularendothelial cells.FASEB J.3:2389–400
2. Alevriadou BR, Eskin SG, McIntire SV,Schilling WP. 1993. Effect of shear stresson86Rb+ efflux from calf pulmonary arteryendothelial cells.Ann. Biomed. Eng.21:1–7
3. Ando J, Ohtsuka A, Korenaga R, Kawa-mura T, Kamiya A. 1993. Wall shear stressrather than shear rate regulates cytoplas-mic Ca2+ responses to flow in vascularendothelial cells.Biochem. Biophys. Res.Commun.190:716–23
4. Ayajiki K, Hindermann M, Hecker M,Fleming I, Busse R. 1996. Intracellular pHand tyrosine phosphorylation but not cal-cium determine shear stress-induced ni-tric oxide production in native endothelialcells.Circ. Res.78:750–58
5. Barbee KA, Davies PF, Lal R. 1994. Shearstress induced reorganization of the sur-face topography of living endothelial cellsimaged by atomic force microscopy.Circ.Res.74:163–71
6. Barbee KA, Mundel T, Lal R, DaviesPF. 1995. Subcellular distribution of shearstress at the surface of flow aligned andnon-aligned endothelial monolayers.Am.J. Physiol.268:1765–72
7. Berk BC, Corson MA, Peterson TE, TsengH. 1995. Protein kinases as mediators offluid shear stress stimulated signal trans-duction in endothelial cells: a hypothesisfor calcium-independent events activatedby flow. J. Biomech.28:1439–50
8. Berthiaume F, Frangos JA. 1992. Flow-induced prostacyclin production is medi-ated by a pertussis toxin-sensitive G pro-tein.FEBS Lett.308:277–79
9. Bhagyalakshmi A, Berthiaume F, Re-ich KM, Frangos JA. 1992. Fluid shearstress stimulates membrane phospholipidmetabolism in cultured human endothelialcells.J. Vasc. Res.29:443–49
10. Buck C, Horwitz A. 1987. Integrin, a trans-membrane glycoprotein complex mediat-ing cell-substratum adhesion.J. Cell Sci.Suppl.8:231–50
11. Burridge K. 1986. Substrate adhesions innormal and transformed fibroblasts: or-ganization and regulation of cytoskeletal,membrane, and extracellular matrix com-ponents at focal contacts.Cancer Res.4:18–78
12. Burridge K, Fath K, Kelly T, Nuckolls G,Turner C. 1988. Focal adhesions: trans-membrane junctions between the extracel-lular matrix and the cytoskeleton.Annu.Rev. Cell Biol.4:487–525
13. Caro CG, Fitzgerald JM, Schroter RC.1969. Arterial wall shear and distribution ofearly atheroma in man.Nature223:1159–61
14. Chand N, Altura MB. 1981. Acetylcholineand bradykinin relax intrapulmonary arter-ies by acting on endothelial cells: role inlung vascular disease.Science213:1376–79
15. Chen J, Aarsvold J, Chen CT, GriemML, Davies PF, et al. 1996. High perfor-mance image analysis and visualization forthree-dimensional light microscopy.Proc.IASTED Signal Image Process.In press
16. Coan DE, Wechezak AR, Viggers RF,Sauvage LR. 1993. Effect of shear stressupon localization of the Golgi apparatusand microtubule organizing center in iso-lated cultured endothelial cells.J. Cell Sci.104:1145–53
17. Cooke JP, Rossitch E, Andon NA, Loscalzo

P1: SBS/rsk P2: sbs/PLB QC: sbs
August 28, 1956 23:59 Annual Reviews DAVICHPT.TXT AR025-23
ENDOTHELIAL FLOW SIGNALING 547
J, Dzau V. 1991. Flow activates an endothe-lial potassium channel to release an en-dogenous nitrovasodilator.J. Clin. Invest.88:1663–71
18. Corson MA, Berk BC, Navas JP, HarrisonDG. 1993. Phosphorylation of endothelialnitric oxide synthase in response to shearstress.Circulation88:1–183 (Abstr.)
19. Davies PF. 1995. Flow-mediated endothe-lial mechanotransduction.Physiol. Rev.75:519–60
20. Davies PF, Mundel T, Barbee KA. 1995. Amechanism for heterogeneous endothelialresponses to flow in vivo and in vitro.J.Biomech.28:1553–60
21. Davies PF, Robotewskyj A, Griem ML.1993. Endothelial cell adhesion in real-time. Measurements in vitro by tandemscanning confocal image analysis.J. Clin.Invest.91:2640–52
22. Davies PF, Robotewskyj A, Griem ML.1994. Quantitative studies of endothelialcell adhesion: directional remodeling offocal adhesion sites in response to flowforces.J. Clin. Invest.93:2031–38
23. Davies PF, Tripathi SC. 1993. Mechani-cal stress mechanisms and the cell. An en-dothelial paradigm.Circ. Res.72:239–45
24. Dewey CF. 1979. Dynamics of arterialflow. Adv. Exp. Med. Biol.115:55–89
25. Dewey CF Jr, Bussolari SR, Gimbrone MAJr, Davies PF. 1981. The dynamic responseof vascular endothelial cells to fluid shearstress.J. Biomech. Eng.103:177–88
26. Diamond SL, Sharefkin JB, DieffenbachC, Frazier-Scott K, McIntire LV, Eskin SG.1990. Tissue plasminogen activator mRNAlevels increase in cultured human endothe-lial cells exposed to laminar shear stress.J.Cell. Physiol.143:364–71
27. Falcone JC, Kuo L, Meininger GA. 1993.Endothelial cell calcium increases duringflow-induced dilation in isolated arterioles.Am. J. Physiol.264:H653–59
28. Flitney FW, Goldman RD, Skalli O, Mer-curius K, Davies PF. 1996. Dynamic prop-erties of intermediate filaments in culturedendothelial cells: effects of controlled fluidshear stress. InThe Biology of Nitric Ox-ide,ed. S Moncada, J Stamler, S Gross, EAHiggs, p. 251. London: Portland
29. Forstermann U, Pollock JS, Schmidt HH,Heller M, Murad F. 1991. Calmodulin-dependent endothelium-derived relaxingfactor synthase activity is present in the par-ticulate and cytosolic fractions of bovineaortic endothelial cells.Proc. Natl. Acad.Sci. USA88:1788–92
30. Frangos JA, Eskin SG, McIntire LV, IvesCL. 1985. Flow effects on prostacyclinproduction by cultured human endothelial
cells.Science227:1477–7931. Franke RP, Grafe M, Schnittler H, Seiffge
D, Mittermayer C, Drenckhahn D. 1984.Induction of human vascular endothelialstress fibers by fluid shear stress.Nature307:648–50
32. Fry DL. 1973. Atherogenesis: initiatingfactors. InCiba Found. Symp. 12,pp. 96–118. London: Ciba Found.
33. Fung YC, Liu SQ. 1993. Elementary me-chanics of the endothelium of blood ves-sels.J. Biomech. Eng.115:1–12
34. Geiger RV, Berk BC, Alexander RW,Nerem RM. 1992. Flow-induced calciumtransients in single endothelial cells: spa-tial and temporal analysis.Am. J. Physiol.262:C1411–17
35. Grabowski EF, Lam FP. 1995. Endothe-lial cell function, including tissue factorexpression, under flow.Thromb. Hemost.74:123–28
36. Heath JP, Dunn GA. 1978. Cell to substra-tum contacts of chick fibroblasts and theirrelation to the microfilament system. Acorrelated interference reflection and highvoltage electronmicroscope study.J. CellSci.29:197–12
37. Henry PD, Cabello OA, Chen CH. 1995.Hypercholesterolemia and endothelial dys-function.Curr. Opin. Lipidol.6:190–95
38. Herman IM, Brant AM, Warty VS, Bonac-corso J, Klein EC, et al. 1987. Hemody-namics and the vascular endothelial cy-toskeleton.J. Cell Biol.105:291–302
39. Himmel HM, Whorton AR, Strauss HC.1993. Intracellular calcium, current, andstimulus-response coupling in endothelialcells.Hypertension21:112–27
40. Ho K, Nichols CG, Lederer WJ, Lyt-ton J, Vassilev PM, et al. 1993. Cloningand expression of an inwardly rectifyingATP-regulated potassium channel.Nature362:31–38
41. Hutcheson IR, Griffith TM. 1994. Hetero-geneous populations of K+ channels medi-ate EDRF release to flow but not agonists inrabbit aorta.Am. J. Physiol.266:H590–96
42. Hynes RO. 1992. Integrins: versatility,modulation, and signalling and cell adhe-sion.Cell 69:11–25
43. Ingber D. 1991. Integrins as mecha-nochemical transducers.Curr. Opin. CellBiol. 3:841–48
44. Ingber DE, Jamieson JD. 1985. Cells astensegrity structures: architectural regu-lation of histodifferentiation by physicalforces transduced over basement mem-brane. InGene Expression During Nor-mal and Malignant Differentiation,ed. LCAndersson, CG Gahmberg, P Ekblom, pp.13–32. Orlando FL: Academic

P1: SBS/rsk P2: sbs/PLB QC: sbs
August 28, 1956 23:59 Annual Reviews DAVICHPT.TXT AR025-23
548 DAVIES ET AL
45. Izzard CS, Lochner LR. 1980. Formation ofcell-to-substrate contacts during fibroblastmotility: an interference-reflexion study.J.Cell Sci.42:81–116
46. Jacobs ER, Cheliakine C, Gebremedhin D,Birks EK, Davies PF, Harder DR. 1995.Shear activated channels in cell-attachedpatches of cultured bovine aortic endothe-lial cells.Pflugers Arch.431:129–31
47. Kaiser L, Spickard RC, Oliver NB. 1989.Heart failure depresses endothelium de-pendent responses in canine femoral artery.Am. J. Physiol.259:H962–67
48. Koller A, Sun D, Kaley G. 1993. Role ofshear stress and endothelial prostaglandinsin flow- and viscosity-induced dilation ofarterioles in vivo. Circ. Res. 72:1276–84
49. Lan QX, Mercurius KO, Davies PF. 1994.Stimulation of transcription factors NF-κ-B and AP1 in endothelial-cells subjectedto shear-stress.Biochem. Biophys. Res.Commun.201:950–56
50. Lopez-Jaramillo P, Gonzalez MC, PalmerRM, Moncada S. 1990. The crucial roleof physiological calcium concentrations inthe production of endothelial nitric oxideand the control of vascular tone.Br. J. Phar-macol.101:489–93
51. Luckoff A, Busse R. 1990. Calcium influxinto endothelial cells and formation of en-dothelium derived relaxing factor is con-trolled by the membrane potential.PflugersArch.416:305–11
52. Luckhoff A, Clapham DE. 1992. Inositol1, 3, 4,5-tetrakisphosphate activates an en-dothelial Ca2+-permeable channel.Nature355:356–58
53. Malek AM, Izumo S. 1996. Mechanismof endothelial cell shape change and cy-toskeletal remodeling in response to fluidshear stress.J. Cell Sci.109:713–26
54. Marsden PA, Heng HH, Scherer SW, Stew-art RJ, Hall AV, Shi XM, et al. 1993. Struc-ture and chromosomal localization of thehuman constitutive endothelial nitric oxidesynthase.J. Biol. Chem.268:17478–88
55. Martin-Mondiere CF, Caprani A, Des-granges PC, Loisance DY, Charron DJ.1989. Shear stress affects expression ofmajor histocompatibility complex antigenson human endothelial cells.ASAIO Trans.35:288–90
56. McKinsey J, DePaola N, Cybulsky M,Davies PF, Polacek D. 1995. Increased ve-nous endothelial VCAM-1 expression indisturbed flow regions associated with ar-teriovenous fistula in normal and hyper-cholesterolemic rabbits.FASEB J.9:A343(Abstr.)
57. Melkumyants AM, Balashov SA, KhayutinVM. 1989. Endothelium dependent controlof arterial dismeter by blood viscosity.Car-diovasc. Res.23:741–47
58. Nagel T, Resnick N, Atkinson WJ, DeweyCF Jr, Gimbrone MA Jr. 1994. Shear stressselectively up regulates ICAM-1 expres-sion in cultured human vascular endothelialcells.J. Clin. Invest.94:885–91
59. Nakache M, Gaub HE. 1988. Hydro-dynamic hyperpolarization of endothelialcells.Proc. Natl. Acad. Sci. USA85:1841–43
60. Nerem RM, Levesque MJ, Cornhill JF.1981. Vascular endothelial morphology asan indicator of the pattern of blood flow.J.Biomech. Eng.103:172–77
61. Nishida K, Harrison DG, Navas JP, FisherAA, Dockery SP, et al. 1992. Molecularcloning and characterization of the consti-tutive bovine aortic endothelial cell nitricoxide synthase.J. Clin. Invest.90:2092–96
62. Ohno M, Gibbons GH, Dzau V, CookeJP. 1993. Shear stress elevates endothelialcGMP. Role of a potassium channel and G-protein coupling.Circulation88:193–97
63. Ohtsuka A, Ando J, Korenaga R, KamiyaA, Toyama-Sorimaci N, Miyasaka M.1993. The effect of flow on the expres-sion of vascular adhesion molecule-1 bycultured mouse endothelial cells.Biochem.Biophys. Res. Commun.193:303–10
64. Olesen SP, Bundgaard M. 1993. ATP-dependent closure and reactivation of in-ward rectifier K+ channels in endothelialcells.Circ. Res.73:492–95
65. Olesen SP, Clapham DE, Davies PF. 1988.Hemodynamic shear stress activates a K+current in vascular endothelial cells.Nature331:168–70
66. Perry PB, O’Neill WC. 1993. Flow stim-ulates nitric oxide synthesis in endothelialcells through a calcium-independent mech-anism.Circulation88:1–134 (Abstr.)
67. Ranjan V, Diamond SL. 1993. Fluid shearstress induces synthesis and nuclear local-ization of c-fosin cultured human endothe-lial cells.Biochem. Biophys. Res. Commun.196:79–84
68. Resnick N, Collins T, Atkinson W, Bon-thron DT, Dewey CF Jr, GimbroneMA Jr. 1993. Platelet-derived growthfactor-B chain promoter contains acis-acting fluid shear-stress-responsive ele-ment.Proc. Natl. Acad. Sci. USA90:4591–95
69. Resnick N, Gimbrone MA Jr. 1995. Hemo-dynamic forces are complex regulatorsof endothelial gene expression.FASEB J.9:874–82

P1: SBS/rsk P2: sbs/PLB QC: sbs
August 28, 1956 23:59 Annual Reviews DAVICHPT.TXT AR025-23
ENDOTHELIAL FLOW SIGNALING 549
70. Rodbard S. 1970. Negative feedback mech-anisms in the architecture and function ofthe connective and cardiovascular tissues.Perspect. Biol. Med.13:507–27
71. Rogers NE, Ignarro LJ. 1992. Constitutivenitric oxide synthase from cerebellum isreversibly inhibited by nitric oxide formedfrom L-arginine.Biochem. Biophys. Res.Commun.189:242–49
72. Sachs F. 1988. Mechanical transduction inbiological systems.CRC Crit. Rev. Biomed.Eng.16:141–69
73. Satcher RL, Bussolari SR, Gimbrone MA,Dewey CF. 1992. The distribution of fluidforces on model arterial endothelium usingcomputational fluid dynamics.J. Biomech.Eng.114:309–16
74. Sessa WC. 1994. The nitric oxide synthasefamily of proteins.J. Vasc. Res.31:131–43
75. Sessa WC, Pritchard K, Seyedi N, WangJ, Hintze TH. 1994. Chronic exercise in-creases coronary vascular nitric oxide pro-duction and endothelial cell nitric ox-ide synthase gene expression.Circ. Res.74:349–53
76. Shen J, Luscinskas FW, Connolly A,Dewey CF Jr, Gimbrone MA Jr. 1992. Fluidshear stress modulates cytosolic free cal-cium in vascular endothelial cells.Am. J.Physiol.262:C384–90
77. Shyy YJ, Hsieh HJ, Usami S, Chien S.1994. Fluid shear stress induces a bipha-sic response of human monocyte chemo-tactic protein-1 gene expression in vascu-lar endothelium.Proc. Natl. Acad. Sci. USA90:4678–82
78. Shyy YJ, Li YS, Lin MC, Chen W, YuanS, et al. 1995. Multipleciselements medi-ate shear stress-induced gene expression.J.Biomech.28:1451–57
79. Stehbens WE. 1966. The basal attachmentof endothelial cells.J. Ultrastruct. Res.15:388–400
80. Takeda K, Schini V, Stoeckel H. 1987.Voltage-activated potassium, but not cal-cium currents in cultured bovine aortic en-dothelial cells.Pflugers Arch.410:385–93
81. Tseng H, Peterson TE, Berk BC. 1995.Fluid shear stress stimulates mitogen-activated protein kinase in endothelialcells.Circ. Res.77:869–78
82. Uematsu M, Navas JP, Nishida K, Ohara Y,Murphy TJ, et al. 1995. Regulation of en-dothelial cell nitric oxide synthase mRNAexpression by shear stress.Am. J. Physiol.269:C1371–78
83. Virchow R. 1860. InCellular Pathology asBased Upon Physiological and Patholog-ical Histology.Transl. F Clance. London:Churchill (From German)
84. Walpola PL, Gotlieb AI, Cybulsky MI,Langille BL. 1995. Expression of ICAM-1 and VCAM-1 and monocyte adherencein arteries exposed to altered shear stress.Arterioscler. Thromb. Vasc. Biol.15:2–10
85. Wang N, Butler JP, Ingber DE. 1993.Mechanotransduction across the cell sur-face and through the cytoskeleton.Science260:1124–27
86. Wechezak AR, Viggers RF, Sauvage LR.1985. Fibronectin and F-actin redistribu-tion in cultured endothelial cells exposedto shear stress.Lab. Invest.53:639–47
87. Zarins CK, Zatina MA, Giddens DP, KuDN, Glagov S. 1987. Shear stress regula-tion of artery lumen diameter in experimen-tal atherogenesis.J. Vasc. Surg.5:413–20
88. Ziegler T, Nerem RM. 1994. Effect of flowon the process of endothelial cell division.Arterioscler. Thromb.14:636–43
NOTE ADDED IN PROOF
An additional K+ channel cDNA cloned from endothelial cells has been reported(subfamily KIR 2.1) by Forsyth S, Hoger A, Hoger J. 1996.FASEB J.10:A627(Abst.)




















