
CD44 is required for two consecutive steps in HGF/c-Met signaling
Upload
independentCategory
view
4download
0
Role of c-Met/PI3k/Akt Signaling in HGF-mediated Lamellipodia Formation, ROS Generation and Motility of Lung Endothelial Cells
Peter V. Usatyuk1, Panfeng Fu1, Vijay Mohan1, Yulia Epshtein2,3, Jeffrey R. Jacobson2,3, Julian Gomez-
Cambronero4, Kishore K. Wary1, Vytas Bindokas5, Steven M. Dudek2,4, Ravi Salgia6, Joe G.N. Garcia2,3
and Viswanathan Natarajan1,3
1Department of Pharmacology, University of Illinois at Chicago, Chicago, IL; 2Section of Pulmonary and Critical Care Medicine, University of Illinois at Chicago, Chicago, IL; 3Institure for Personalized Respiratory Medicine, The University of Illinois at Chicago, Chicago, IL; 4Department of Biochemistry and Molecular Biology, Wright State University, Dayton, OH; 5Department of Pharmacological and Physiological Sciences, The University of Chicago, Chicago, IL; 6Department of Medicine, The University of Chicago, Chicago, IL Running Title: Hepatocyte growth factor in endothelial cells motility To whom correspondence should be addressed: V. Natarajan, PhD, Department of Pharmacology, University of Illinois at Chicago, COMRB, Room # 3137, 835 South Wolcott Avenue, Chicago, IL 60612, Tel.: 312-355-5896; Fax: 312-996-1225; E-mail: [email protected]
Key words: Lamellipodia, Cell motility, ROS, HGF, c-Met, PI3k, Akt, endothelial cells. Background: Lamellipodia structures provide a platform for the spatio-temporal localization of key components necessary for cell migration. Results: HGF activates c-Met/PI3k/Akt signaling axis, which is essential for the recruitment of actin, cortactin, p47phox and Rac1and ROS production in lamellipodia. Conclusion: HGF-induced spatio-temporal localization of cytoskeletal proteins and NADPH oxidase components regulate lamellipodial ROS and cell migration. Significance: This study identifies a novel role for lamellipodial ROS in cell motility.
http://www.jbc.org/cgi/doi/10.1074/jbc.M113.527556The latest version is at JBC Papers in Press. Published on March 14, 2014 as Manuscript M113.527556
Copyright 2014 by The American Society for Biochemistry and Molecular Biology, Inc.
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from

2
ABSTRACT Hepatocyte growth factor (HGF) mediated signaling promotes cell proliferation and migration in a variety of cell types and plays a key role in tumorigenesis. As cell migration is important to angiogenesis, we characterized HGF-mediated effects on the formation of lamellipodia, a pre-requisite for migration using human lung microvascular endothelial cells (HLMVECs). HGF, in a dose-dependent manner, induced c-Met phosphorylation (Y1234/1235, Y1349, S985, Y1003 and Y1313), activation of PI3k (phospho-Yp85) and Akt (phospho-Thr308 and phospho-Ser473) and potentiated lamellipodia formation and HLMVEC migration. Inhibition of c-Met kinase by SU11274 significantly attenuated c-Met, PI3k and Akt phosphorylation, suppressed lamellipodia formation and endothelial cell migration. LY294002, an inhibitor of PI3k, abolished HGF-induced PI3k (Y458), and Akt (T308 and S473) phosphorylation and suppressed lamellipodia formation. Furthermore, HGF stimulated p47phox/Cortactin/Rac1 translocation to lamellipodia and ROS generation. Moreover, inhibition of c-Met/PI3k/Akt signaling axis and NADPH oxidase attenuated HGF- induced lamellipodia formation, ROS generation and cell migration. Ex Vivo experiments with mouse aortic rings revealed a role for c-Met signaling in HGF-induced sprouting and lamellipodia formation. Taken together, these data provide evidence in support of a significant role for HGF-induced c-Met/PI3k/Akt signaling and NADPH oxidase activation in lamellipodia formation and motility of lung endothelial cells. INTRODUCTION Vascular endothelial cells provide a selective-structural barrier between
circulation and surrounding tissue, and regulate vascular permeability and blood flow (1,2). In addition, they play a major role in angiogenesis, a process required for the generation of new blood vessels and repair of damaged vasculature (3). Several pathological conditions, including tumor growth, ischemia/reperfusion, diabetes, rheumatoid arthritis, atherosclerosis, and inflammatory processes also require neo-angiogenesis. It is therefore not surprising that angiogenesis is therapeutically targeted in several human disorders (4-5). Several growth factors, including the hepatocyte growth factor (HGF), regulate angiogenesis. HGF, also known as scatter factor, is the natural ligand for the receptor tyrosine kinase, c-MET (6) that is expressed on a variety of cell types including vascular endothelial cells and is up-regulated during angiogenesis (7,8). HGF interacts with its cognate receptor c-Met and promotes angiogenesis by enhancing the expression of vascular endothelial growth factor, one of the principal mediators of angiogenesis. HGF binding to c-Met triggers a cascade of signal transduction pathways including activation of phosphatidylinositol-3-kinase (PI3k) and Akt, which have been implicated in cell migration (7-11). Cell migration is essential for embryonic development, wound healing, and angiogenesis (3,7,8,12-14). It is a multi-step process facilitated by highly coordinated cytoskeletal changes, including extracellular adhesion, plasma membrane protrusion at the leading edge (lamellipodia formation), formation of new adhesion sites under the protrusion and disruption of older adhesions at the rear and contraction, thereby drawing the cell forward (12-20). The lamellipodium is characterized by a dense network of short, branched actin and cortactin filaments, with elongated filaments that have barbed-end capping (13,15,16,18-24). At the edge of the lamellipodia are club like projections referred to as the filipodia that generate the adhesion sites. Recent studies suggested that the cytoskeletal proteins, particularly actin and cortactin, play a role in phagocytic and non-phagocytic activation of NADPH
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from

3
oxidase (25-29). Cortactin redistribution to cell periphery and its co-localization with p47phox, a NADPH oxidase subunit, is important in the assembly of NADPH oxidase components with the actin cytoskeleton during agonists-induced generation of reactive oxygen species (ROS) in human lung ECs (26-28). Thus, targeting NADPH oxidase components to focal complexes in lamellipodia may provide a mechanism to specifically generate ROS in localized structure such as lamellipodia, a requirement for stimulus-induced cell migration (10,30-34). During our investigations into cytoskeletal regulation of NADPH oxidase activation and endothelial barrier function, we observed that inhibition of HGF-induced NADPH oxidase activation in membrane protrusions attenuated lamellipodia formation and migration of HLMVECs. However, signaling mechanism(s) of HGF-induced, ROS-dependent lamellipodia and cell migration is not well understood. Here, we determined the role of HGF/c-Met/PI3k signaling axis in ROS-dependent lamellipodia formation and cell migration. Our results demonstrated that HGF-induced c-Met activation as well as phosphorylation of downstream targets such as PI3k/Akt kinases is required for lamellipodia formation and endothelial cell (EC) motility. In addition, HGF/c-Met signaling axis stimulated ROS formation at the lamellipodia, and blocking activation of PI3k, Akt and NADPH oxidase components (p47phox, and Rac1) attenuated HGF-induced lamellipodia formation and cell migration. MATERIALS AND METHODS Materials: HLMVECs and endothelial basal media (EBM-2) were obtained from Lonza (San Diego, CA). Phosphate-buffered saline (PBS) was from Biofluids Inc. (Rockville, MD). Glass bottom micro well dishes were purchased from MatTek (Ashland, MA). Gold antifade mounting media, DAPI, Hoechst and precast Tris-Glycine PAGE, Alexa Fluor 488, Alexa Fluor 568, Alexa Fluor Phalloidin 568, CellLight Actin-RFP
BacMam 2.0 were procured from Invitrogen (Eugene, OR). HGF was from R&D Systems (Minneapolis, MN) and PeproTech (Rocky Hill, NJ). SU11274 was purchased from Selleck Chemicals (Houston, TX). Anti-c-Met, anti-phospho-c-Met (Tyr1234/1235), anti-phospho-c-Met (Tyr1349), anti-phospho-c-Met (Ser985), anti-phospho-c-Met (Tyr1003), anti-Akt, anti-phospho-Akt (Thr308), anti-phospho-Akt (Ser473), LY294002 and cell lyses buffer were from Cell Signaling Technology (Danvers, MA). Scrambled siRNA and siRNA for c-Met, PI3k p85, and p47phox, antibodies for anti-Cortactin anti-phospho-c-Met (Tyr1313), and BSA were obtained from Santa Cruz Biotechnology (Santa Cruz, CA). Anti-actin (Sigma, St. Louis, MO), anti-p47phox (NIH, Bethesda), anti-phospho-PI3k (Y458) (Thermo Scientific, Rockford, IL), anti-Rac1 and anti-PI3k p85 (BD Biosciences, San Jose, CA), Fibrin Gel and In Vitro Angiogenesis Assay Kit (Millipore, Billerica, MA), IgG (H+L) HRP conjugates (BioRad, Hercules, CA) were all commercially obtained. Gene Silencer was from Gene Therapy System (San Diego, CA). pHyPer-cyto plasmid was purchased from Evrogen (Moscow, Russia). Actin-RFP and Cortactin-RFP plasmids were provided by Dr. Steven Dudek (University of Illinois at Chicago). FuGENE HD transfection reagent was from Promega (Madison, WI), ECIS electrodes 8W1E were procured from Applied Biophisics (Troy, NY). Immunobilon-P, 0.45 µm was procured from Millipore (Bedford, MA). Endothelial Cell Culture: HLMVECs cultured in complete media (EBM-2) (30), were maintained at 37o C and 5% CO2 and grown to contact-inhibited monolayers that revealed typical cobblestone morphology. Cells were then detached with 0.05% trypsin and resuspended in fresh medium and cultured on gold electrodes for electrical resistance determinations, on glass coverslips for fluorescent microscopy studies, or in 60-100 mm culture-dishes for preparation of cell lysates and Western blot analysis.
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from

4
Mouse aortic ring sprouting assay: Thoracic aortic rings from 2-month-old mice were prepared (20-25 g body weight; The Jackson Laboratory), housed under pathogen-free conditions at the University of Illinois at Chicago (UIC) Animal Care Vivarium and treated humanely in accordance with institutional guidelines (35). Briefly, thoracic aortae were isolated, dissected from connective tissues, and washed in sterile PBS extensively under aseptic conditions and the aortae were cut into rings ~1 mm in thickness. The rings were placed in the middle of the glass bottom 35 mm dishes, overlaid with 100 µl of Fibrinogen/Thrombin solution according to manufacturer protocol, and left to polymerize for 1 h at 37°C before the addition of EBM-2 media without serum- and growth factor, which was replaced each day with fresh media or media containing 20 ng/ml HGF. After 6 days of culture, emergent angiogeneic sprouts were examined using Zeiss Axiovert 40 phase contrast microscope (lens x10). Images were captured with the use of a digital camera. The area of angiogenic sprout outgrowth was quantified by using image acquisition and analysis software (Image J).
Lamellipodia Formation ex vivo in aortic sprouts: To determine lamellipodia formation ex vivo, aortic ring were grown in complete media for 4 days, infected for 24 h with CellLight Actin-RFP BacMam 2.0 according to protocol and living cells transiently expressed Actin-RFP were visualized by laser scanning microscope Zeiss 510 Meta (lens x40). Several cells were selected from each glass bottom dish and actin dynamics was monitored before and after HGF (20 ng/ml) stimulation with autofocus mode.
Preparation of Cell Lysates and Western Blotting: HLMVECs were cultured in 60-mm dishes to ~95% confluence and starved for 3 h in EBM-2 medium containing 0.1% FBS. Cells were stimulated with HGF (2-30 ng/ml) for 15 min, washed with PBS and lysed as described (28). Clarified cell lysates
were boiled with the sample buffer for 4 min, and the proteins (20-30 µg) were subjected to SDS-PAGE analysis, transferred to PVDF membranes, blocked in TBST containing 1% BSA prior to incubation with primary antibody (1:1000 dilution) overnight. After blocking and washing, membranes were incubated with appropriate secondary antibody (1:10,000 dilution), and blots were developed using an ECL chemiluminescence kit. Western blots were scanned by densitometry and integrated density of pixels in identified areas was quantified using ImageJ.
Immunofluorescence Microscopy HLMVECs grown in chamber slides were starved for 3 h in EBM-2 containing 0.1% FBS, pretreated with inhibitors or siRNA as indicated prior to treatment with HGF for 15 min. Cells were fixed in 3.7% formaldehyde in PBS for 10 min, washed three times with PBS, permeabilized with 0.25 % Triton X-100 or methanol for 4 min, blocked with 1 % BSA in TBST, incubated for 1 h with appropriate primary antibody (1:200 dilution), washed with TBST, and stained for 1 h with secondary antibody (1:200 dilution) in TBST containing 1 % BSA. Cells were visualized using a Nikon Eclipse TE2000-S immunofluorescence microscope and a Hamamatsu digital camera with x 60 oil immersion objective. siRNA Transfection of HLMVECs: Cells grown to ~50% confluence in 6-well plates or chamber slides were transfected with Gene Silencer® transfecting reagent containing scrambled siRNA (50 nM) or siRNA for target proteins (50 nM) in serum-free EBM-2 medium according to manufacturer’s recommendation. cDNA Transfection of HLMVECs: To determine intracellular hydrogen peroxide levels, HLMVECs grown to ~50% confluence were transiently transfected with 1 µg/ml of pHyPer-cyto plasmid (36) using FuGene HD (3 µg/ml) transfection reagent in serum-free EGM-2 medium according to manufacturer’s recommendation. Similarly,
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from

5
to estimate ROS production in lamellipodia ECs were double transfected with 0.5/0.5 µg/ml of pHyPer-cyto/Actin-RFP or pHyPer-cyto/Cortactin-RFP plasmids. After 3 h, the media of transfected cells were replaced by complete EGM-2 containing 10% fetal bovine serum, and the cells were incubated for 72 h. Determination of intracellular hydrogen peroxide in lamellipodia: Intracellular hydrogen peroxide levels were determined by a mammalian expression vector encoding a fluorescent hydrogen peroxide sensor Hyper (36). Cells were grown on glass bottom 35-mm dishes, transfected with 1 µg/ml of pHyPer-cyto plasmid and treated as described above, cells were washed twice with Phenol Red free basal EBM-2 and fluorescence of pHyPer-cyto in living cells was examined under a Nikon Eclipse TE 2000-S fluorescence microscope with a Hamamatsu digital CCD camera (Japan), using a 60 X objective lens. To calculate pHyPer-cyto fluorescence intensity (pixels) similar rectangular cell periphery regions were selected in control and treated cells using MetaVue software (Universal Imaging Corp., PA) and values are expressed as % of Control. At least 6-10 cells were evaluated for each treatment in triplicate, averaged to three independent experiments and background fluorescence from empty dish were subtracted. Confocal micrographs of pHyPer images were acquired every minute on a Marianas Yokogawa CSU-X1 spinning disk confocal system (Intelligent Imaging Innovations, Denver) on a Zeiss Observer Z1 platform using a 40X NA1.3 oil objective under control of SlideBook 5.5 software. pHyPer-cyto emission (525/50 nm) was excited using 405 and 488 nm lasers, mCherry was excited with 561 nm laser and emission collected at 607/43 nm. Temperature was maintained at 37o C and cells were kept in 5% CO2 (OKO Labs). Reporter response was calculated as the ratio of background-subtracted 488 and 405 signals. pHyPer-cyto/Actin-RFP and pHyPer-cyto/Cortactin-RFP fluorescent intensity profiles (3-pixel-
wide average over 10 µm) across lamellipodia were estimated using ImageJ (Wayne Rasband, NIH). Quantification of lamellipodia: Quantification of lamellipodia was performed as described earlier (37). Briefly, for each image, background signal was subtracted by drawing a region of interest around the cell periphery of individual cells. All areas outside the cell were cleared to best visualize the leading edges including cell periphery and the fluorescence intensity within the entire cell was summed by MBF ImageJ bundle (Tony Collins, McMaster University, http://www.macbiophotonics.ca/imageJ/ and Wayne Rasband, NIH, http://rsb.info.nih.gov/ij/). Electrical Cell Substrate Impedance Sensing (ECIS) Assay: HLMVEC were cultured in 8-well ECIS electrode arrays (8W1E, Applied Biophysics, NY) to ~95% confluence and starved in the EBM-2 medium with 0.1% BSA for 1-3 h. An elevated field (3 V at 40,000 Hz for 10 sec) was applied to wound the cells on the electrode. Inhibitors and HGF were reconstituted in EBM-2 medium containing 0.1% BSA and endothelial wound healing was monitored for 12-16 h by measuring the trans endothelial electrical resistance using the ECIS equipment (38). Statistical Analysis ANOVA with Student-Newman-Keul’s test was used to compare means of clearance rates of two or more different treatment groups. The level of significance is p < 0.05 unless otherwise stated. Data are expressed as mean ± SEM. RESULTS
HGF stimulates lamellipodia formation in lung ECs: Cell motility plays a central role in migration, wound healing and angiogenesis. The driving force for cell migration is lamellipodia formation
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from

6
propelled by the reorganization of the actin and cortactin cytoskeleton at the cell front and the retraction of the cell at the rear (13-15,18,19,21-24,39). We therefore determined the effect of HGF on lamellipodia formation. HLMVECs were either treated with vehicle or vehicle containing HGF (2, 5, 10, 20 and 30 ng/ml for 15 min). Cells were immunostained for actin and cortactin co-localization, an index of lamellipodia formation (19, 24, 25). The vehicle treated cells revealed typical F-actin staining with a few stress fibers in the central area of the cell and diffused cortactin staining; however, HGF in a dose-dependent manner, induced F-actin stress fiber formation (red) and cortactin (green) redistribution to cell periphery, which were co-localized in lamellipodia (merge, yellow) (Fig. 1 A and B). Despite extensive actin polymerization at the cell periphery, stress fibers remained stable. These results suggest a key role for HGF in lamellipodia formation in HLMVECs. HGF enhances hydrogen peroxide formation in lamellipodia of lung ECs: The cytoskeletal proteins, actin and cortactin, are known to play a role in NADPH-oxidase assembly, activation and ROS generation (27-29). HGF activated NADPH oxidase and increased ROS generation (10,40,41); however, the role of HGF in lamellipodial ROS generation is unclear. Therefore, we hypothesized that HGF may stimulate ROS accumulation in lamellipodia. HLMVECs transfected with pHyPer-cyto (36) were challenged with varying concentrations of HGF (2-30 ng/ml) and hydrogen peroxide generated in living cells was analyzed using fluorescent microscopy. As shown in Fig. 2 A and B, HGF, in a dose-dependent manner, induced pHyPer-cyto green fluorescence (a measure of hydrogen peroxide accumulation, in the cytosol, peri-nuclear and in cell periphery), which was confirmed by confocal microscopy (Fig. 2 C and D). To further elucidate HGF-induced ROS accumulation in lamellipodia, ECs were double transfected with pHyPer-cyto/Actin-RFP and pHyPer-
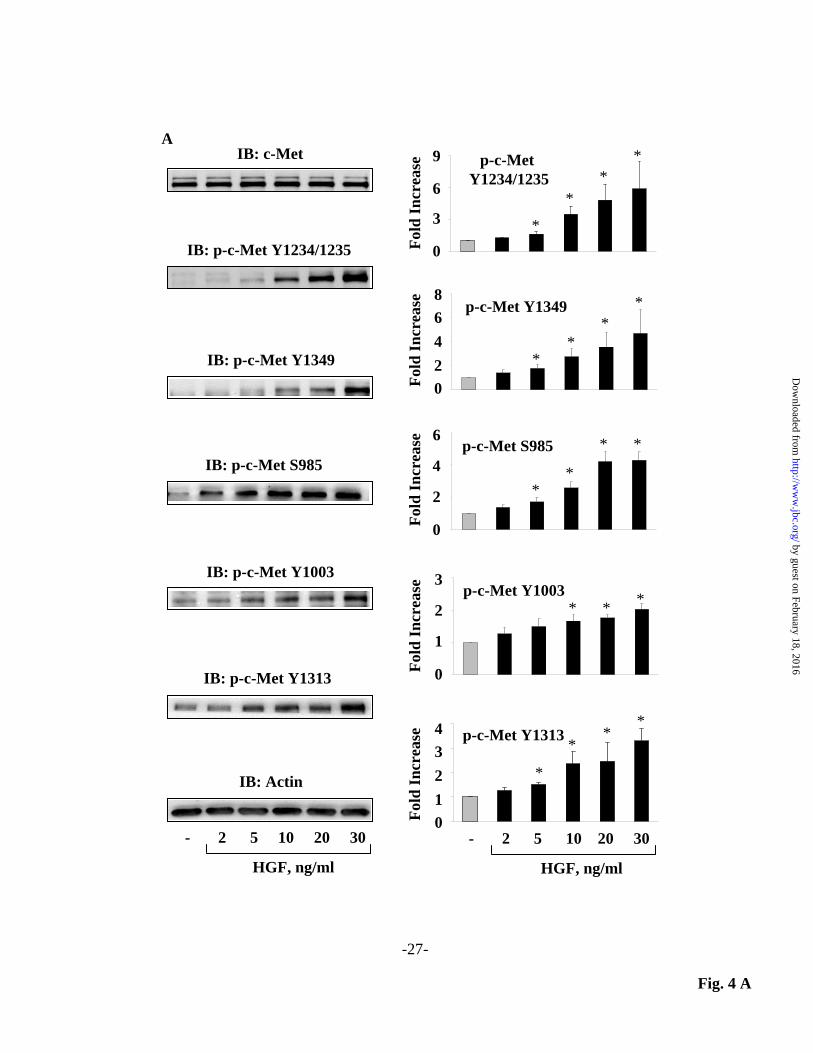
cyto/Cortactin-RFP and living cells were analyzed under confocal microscopy. As shown in Fig. 3 A and B, HGF stimulated hydrogen peroxide accumulation in cell periphery which were co-localized with actin and cortactin in lamellipodia. pHyPer-cyto/Actin-RFP and pHyPer-cyto/Cortactin-RFP fluorescent intensity profiles (3-pixel-wide average over 10 µm) across lamellipodia demonstrated colocalization of signals and significant increases in both red and green fluorescence after HGF exposure (Fig. 3 C and D). Taken together, these data support accumulation of hydrogen peroxide in lamellipodia in response to HGF in human lung ECs. Down-regulation of c-Met or inhibition of c-Met phosphorylation attenuates HGF stimulated phosphorylation of c-Met, PI3k and Akt in lung ECs: Binding of HGF to its receptor, c-Met, induces dimerization, and auto phosphorylation of specific tyrosine residues resulting in enhanced kinase activity and activation of diverse intracellular signaling pathways including MAPKs, STAT3, Rac1 and Akt (7,10,41-46), which promote EC barrier function (47,48). To further characterize HGF/c-Met signaling axis in EC function, we used SU11274 (49) to inhibit HGF mediated c-Met phosphorylation. As expected, HGF, in a dose-dependent manner, induced c-Met phosphorylation at residues Y1234/1235, Y1349, Y1003, and Y1313 as well as S985 in HLMVECs (Fig. 4 A). Pretreatment of cells with SU11274 (1 µM) for 1 h, prior to HGF treatment (20 ng/ml, 15 min) attenuated HGF-induced c-Met phosphorylation at various tyrosine residues (Fig. 4 B). Down-regulation of c-Met with siRNA or SU11274 also inhibited HGF enhanced phosphorylation of PI3k (Y548) and Akt (T308 and S473) (Fig. 5 A and B). These results show that HGF mediated activation of PI3k/Akt pathway is via c-Met phosphorylation in HLMVECs. Inhibition of c-Met tyrosine phosphorylation by SU11274 attenuates HGF-induced lamellipodia formation,
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from

7
accumulation of ROS in lamellipodia and lung EC migration: Having established that HGF stimulates lamellipodia formation in HLMVECs via c-Met signaling, we next determined the potential link between c-Met inhibition and HGF-induced lamellipodia formation, ROS localization in lamellipodia and cell migration. As shown in Fig. 6 A, SU11274 significantly attenuated HGF- induced actin and cortactin redistribution to cell periphery and their co-localization in lamellipodia. Further, inhibition of c-Met phosphorylation with SU11274 reduced HGF-induced hydrogen peroxide accumulation in lamellipodia of HLMVECs (Fig. 6 B) as measured by pHyPer-cyto fluorescence. We also determined the role of c-Met on EC motility using a wound repair ECIS assay. Local area of cell culture monolayer, grown on gold microelectrodes, was mechanically disrupted (3 V at 40,000 Hz for 10 sec) that resulted in a very rapid drop in transendothelial resistance (TER) followed by an increase due to the migration of cells from the perimeter of the electrode to replace the wounded cells (Fig. 6 C). Restoration of TER in the presence of HGF (20 ng/ml) was more pronounced compared to control cells suggesting that HGF is a potent inducer of wound repair processes in lung ECs, and pre-treatment of cells with SU11274 resulted in a dramatic suppression of HGF mediated wound healing as determined by decreased cell migration (Fig. 6 C). Interestingly, SU11274 alone caused a significant reduction in endothelial wound repair in the absence of HGF suggesting the importance of basal c-Met activation in EC motility. Taken together, these results establish a key role for HGF/c-Met signaling in lamellipodia formation, ROS in lamellipodia and EC motility. Role of PI3k/Akt signaling in HGF-mediated lamellipodia formation, ROS in lamellipodia and lung EC migration: To further investigate the role of PI3k/Akt signaling in HGF mediated lamellipodia formation, we used LY294002, a PI3K specific inhibitor (50) and also down-regulated PI3k using specific siRNA. Both
LY294002 and siRNA against PI3k significantly reduced HGF-induced PI3k (Y458) and Akt (T308 and S473) phosphorylation (Fig. 7 A and B). Furthermore, HGF (20 ng/ml, 15 min) induced co-localization of actin and cortactin at the lamellipodia was inhibited by pre-treatment of cells with the PI3k inhibitor LY294002 (Fig. 7 C). Similar experiments carried out in PI3k siRNA transfected cells revealed comparable results to those obtained with LY 294002 (Fig. 7 D). In parallel experiments we also demonstrated that down regulation of PI3k/Akt signaling pathways attenuated HGF-induced formation of hydrogen peroxide accumulation in lamellipodia (Fig. 7 E and F) and wound repair in HLMVECs (Fig. 7 G and H). The above results demonstrated a key role for HGF-mediated PI3k/Akt activation in lamellipodia formation, ROS accumulation in lamellipodia and cell migration. c-Met/PI3k/Akt signaling is essential for HGF-mediated p47phox, cortactin and Rac1 redistribution to lamellipodia in lung ECs: Recent studies indicate that PI3k and Akt play an important role in p47phox activation and production of ROS (10,40,51-53). Also, we have demonstrated previously that hyperoxia-induced activation of lung endothelial NADPH oxidase that results in ROS generation is dependent on p47phox and cortactin translocation to cell periphery (26-28,54). Further, cortactin acts as a scaffold protein for NADPH oxidase assembly and is essential for agonist-induced p47phox
translocation to cell periphery, cortactin/p47phox co-localization and O2
.-
/ROS generation in lung ECs (27,29). To determine the role of c-Met/PI3k/Akt signal transduction in translocation of p47phox , cortactin and Rac1 to lamellipodia, HLMVECs were pretreated with c-Met inhibitor SU112744, PI3k inhibitor LY294002, or transfected with PI3k siRNA (50 nM, 72 h) followed by HGF (20 ng/ml) challenge for 15 min. As shown in Fig. 8 A HGF enhanced co-localization of p47phox and cortactin in lamellipodia (merged color
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from

8
yellow) as compared to control cells challenged with vehicle alone. Treatment of cells with either SU11274 or LY294002 or knockdown of PI3k with siRNA blocked the redistribution and co-localization of p47phox and cortactin in lamellipodia (Fig. 8 A). Small GTPase Rac1 also plays an important role in lamellipodia formation, cortactin redistribution to cell periphery and in the activation of NADPH oxidase and ROS generation (26,44,55-57). We therefore examined the localization of Rac1 in lamellipodia using the above three approaches. Clearly, HGF also induced Rac1 translocation to lamellipodia in HLMVECs, while downregulation of c-Met or PI3k/Akt signaling attenuated the process (Fig. 8 B). Statistical analyses of the data, revealed a strong role for c-Met/PI3k/Akt signaling in p47phox/cortactin/Rac1 localization to the cell periphery (Fig. 8 C and D). Inhibition of NADPH oxidase and downregulation of cortactin impairs lamellipodia formation, and ROS accumulation in lung ECs: To further evaluate the role of ROS in HGF-mediated cytoskeletal rearrangement and lamellipodia formation, NADPH oxidase dependent ROS was blocked by pretreatment with apocynin (a general ROS inhibitor) or by silencing p47phox with siRNA. As shown in Fig. 9 A, apocynin treatment or p47phox siRNA transfection of HLMVECs resulted in a significant loss in HGF-induced lamellipodia formation compared to control cells. In parallel experiments, a concomitant decrease in ROS generation in apocynin or p47phox siRNA transfected cells was observed (Fig. 9 B). In addition, knock down of cortactin using specific siRNA significantly attenuated HGF-induced lamellipodia formation, ROS accumulation in lamellipodia and p47phox and Rac1 localization in lamellipodia (Fig. 9 A-D). Similarly, inhibition of NADPH oxidase by apocynin, or down regulation of p47phox or cortactin with siRNA attenuated HGF-induced cell migration as measured by wound-repair assay (Fig. 10 A-C). These
results suggest a potential role for p47phox, and cortactin in ROS formation in lamellipodia by HGF in lung ECs. HGF-induced endothelial sprouting and lamellipodia formation ex vivo Having established a role for HGF stimulated lamellipodia formation via c-Met in HLMVECs, we next determined the role of HGF/c-Met signaling axis in an ex vivo system using aortic ring angiogenesis assay (58). As shown in Fig. 11 A isolated aortic rings in fibrin matrigel formed endothelial sprouts and tube elongation into fibrin matrix in the presence of serum. Addition of HGF stimulated mouse aortic sprouting density (~1.9 fold) compare to control aortic rings without HGF. To further elucidate the role of c-Met in HGF mediated lamellipodia formation in the sprouts, aortic rings embedded in fibrin matrigels were transfected with Actin-RFP construct and lamellipodia formation was visualized in living cells under confocal microscope. As shown in Fig. 11 B and C, HGF enhanced actin accumulation in lamellipodial structures of the aortic endothelial sprouts, which was blocked by the c-Met tyrosine kinase inhibitor, SU11274. Thus, our aortic ring endothelial sprout assay further substantiates that HGF stimulates endothelial sprouting and lamellipodia formation ex vivo. DISCUSSION Cell migration, a complex process that involves protrusion, adhesion, contraction and retraction, is essential for a variety of cellular functions such as cell growth, motility, angiogenesis and morphogenesis (12-15,19,21,59). ECs like many other eukaryotic cells also undergo directed cell migration mediated by lamellipodial protrusions for the formation of adhesions at leading edge of the cells (59-61). While the participation of cytoskeleton, mainly composed of actin microfilaments, microtubules and intermediary filaments, is essential in the polarized cell migration, mechanisms regulating the generation of
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from

9
spatially and temporally restricted signaling molecules in lamellipodial protrusions are yet to be completely defined. Here, we report that in human lung ECs, the HGF-mediated activation of c-Met/PI3k/Akt signaling axis regulates cell migration, which requires formation of lamellipodial protrusions. Further, we demonstrate that the lamellipodia are not only enriched in actin and cortactin but also with components of NAPH oxidase such as p47phox and Rac1 and hydrogen peroxide generated via NADPH oxidase, which require c-Met/PI3k/Akt signaling axis. All of the above, triggered by HGF/c-Met signaling are required for lamellipodia formation in ECs and their migration. HGF is the same as scatter factor that was originally shown to be fibroblast derived cell-motility factor for epithelial cells (6). Through its natural receptor c-Met, HGF is now known to promote matrix invasion, angiogenesis, tumorigenesis and tissue regeneration (7,8,62-64). HGF has been previously shown to promote cell motility in several normal and cancer cell lines (8,10,44,48,63,65). The driving force for cell migration is provided by continuous growth of actin filaments and cortactin rearrangement in the lamellipodial protrusions in front end of the cell (13-15,18-22,24,59,66). The formation of lamellipodia is controlled by Rho family G proteins particularly Rac and Cdc42 which regulate the Arp2/3 complex through the Wiskott-Aldrich syndrome protein (WASP) to promote lamellipodia formation (23,24,31,44,56,57), while Rho regulates the formation of stress fibers and focal adhesions and induces actomyosin contractility (14,67). Our results in lung ECs demonstrated that HGF-induced activation of c-Met mimicked the classic c-Met phosphorylation at Y1234/1235, Y1349, S985, Y1003 and 1313 (Fig. 4), which regulate the tyrosine kinase activity and recruit other key signaling molecules (6,63,68). Phosphorylation of c-Met at Y1313 is known to recruit PI3k to the cell periphery (63) and inhibition of c-Met tyrosine kinase activity using SU11274 not
only suppressed the activation of c-Met and the immediate downstream targets PI3k/Akt, but also the ensuing accumulation and co-localization of actin and cortactin, NADPH oxidase components at the lamellipodia (Figs. 4, 5, 6 and 7). The regulatory p85 subunit of PI3k can bind either directly to c-Met at phosphorylated Y1313 or indirectly through GAB1, which then signals through Akt/protein kinase B (6). Although we have not shown the direct binding of PI3k to c-Met, our studies revealed clear activation of both PI3k and Akt, as evidenced by specific phosphorylation of Akt (T308, S373), and blocking PI3k with the inhibitor LY294002 or the knockdown of PI3k by siRNA attenuated HGF mediated EC motility (Fig. 7 G and F). Involvement of PI3k/Akt signaling pathway in HGF-mediated migration of cancer cells has been described. In Uveal melanoma cells, HGF was found to enhance cell migration, and that HGF-induced migration was dependent on PI3K/Akt pathway. The activation of PI3K/Akt pathway induced by the HGF/c-Met axis is involved in the down-regulation of cell adhesion molecules E-cadherin and β-catenin, contributing to the attenuation of cell-cell adhesion and promoting the enhanced motility (69). Although localization of PI3k in membrane ruffles and lamellipodia is unclear, the product of PI3k, namely phosphatidylinositol 3,4,5-trisphospahte (PIP3) is highly localized at the site of cell edges and lamellipodia and the spatial and temporal generation PIP3 has been implicated in directed cell migration (69-71). Results presented here clearly demonstrated a role for c-Met/PI3k/Akt signaling pathway in the accumulation of ROS at the lamellipodia and cell migration as well as endothelial barrier function. In addition to actin and cortactin, Rac1 and p47phox components of NADPH oxidase are targeted to the lamellipodia and blocking Rac1 and p47phox attenuated lamellipodia formation and cell migration (31). These results suggested that targeting of NADPH oxidase to lamellipodia represents a novel mechanism for a cell to promote directed migration to external stimuli such as HGF
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from

10
and other growth factors. Although details on targeting NADPH oxidase components to lamellipodia are largely unknown, targeting p47phox to focal complexes seems to be mediated via adaptor protein TRAF4-Hic5 complex (72). In a recent study Martinelli et al. showed that ventral lamellipodia of ECs were enriched in and functionally dependent on cortactin, IQGAP1 and p47phox (17). Additionally, IQGAP1 may also regulate directional migration by binding Rac1 and recruitment of p47phox and other components to leading edge of the cell required for directed cell migration. We previously showed the involvement of phospholipase D 2 (PLD2) mediated activation of IQGAP1 through Rac1 in tyrosine phosphorylation of Src and cortactin (26), and a potential role for cytoskeletal proteins and dynamin 2 in the assembly and activation of NADPH oxidase in lipid rafts in response to hyperoxia in human lung endothelial cells (44). Interestingly, the hyperoxia-induced IQGAP1 activation and redistribution to cell periphery was attenuated in Rac1 down-regulated human lung ECs suggesting Rac1 activation up-stream of IQGAP1 (26). Although there is considerable evidence for the localization of NADPH oxidase components and generation of hydrogen peroxide in lamellipodia, the physiological relevance for this spatio-temporal enrichment in cell migration or wound healing is unclear. Blocking NADPH oxidase activation either with apocynin or p47phox siRNA not only significantly reduced lamellipodia formation and hydrogen peroxide production in the lamellipodia (Fig. 9), but also attenuated ventral lamellipodia micro-wound healing (17). Mechanism(s) of regulation of cell migration or wound healing by localized lamellipodial ROS is unclear; however, it may be involved in controlling the cofilin
activity (73-75) or in the regulation of temporal and spatial concentrations of PI(3,4,5)P3 (70,76), a critical component in determining actin polymerization and lamellipodia projections. Other possible mechanism may involve SeIR/Mical-mediated oxidation of actin to regulate F-actin dynamics (77,78). Recent studuies demonstrate that endogenous hydrogen peroxide play an important role in angiogenesis in vitro and neovascularization in vivo (79-81). Aortic ring ex vivo model also demonstrates impaired vascular formation, vessel sprouting and tube elongation by endothelial-specific catalase overexpression (79), further emphasizing a critical role for endogenous ROS in angiogenesis. In our ex vivo experiments, using mouse aortic rings, HGF enhanced capillary sprouting and tube elongation into fibrin matrix (Fig. 11 A). Further, HGF-induced lamellipodia formation in aortic ring sprouts was significantly attenuated by inhibition of c-Met tyrosine phosphorylation (Fig. 11 B and C). In summary, we have demonstrated a novel role for c-Met/PI3k/Akt signaling in HGF induced lamellipodia formation in lung endothelium, which is dependent on recruitment of cortactin, Rac1 and p47phox components and localized ROS production (Fig. 12). We also show that localized ROS production in lamellipodia to be involved in EC migration, an important component of angio- and vasculo-genesis. Further understanding of lamellipodial ROS accumulation and the role of this localized ROS in regulation of signal transduction pathways in lamellipodia is necessary to define the physiological relevance of localized ROS signal, as in lamellipodia, in the directional migration and wound healing.
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from

11
REFERENCES 1. Komarova, Y., and Malik, A. B. (2010) Regulation of endothelial permeability via
paracellular and transcellular transport pathways. Annu Rev Physiol 72, 463-493 2. Dudek, S. M., and Garcia, J. G. (2001) Cytoskeletal regulation of pulmonary
vascular permeability. J Appl Physiol 91, 1487-1500 3. Marcelo, K. L., Goldie, L. C., and Hirschi, K. K. (2013) Regulation of endothelial
cell differentiation and specification. Circ Res 112, 1272-1287 4. You, W. K., and McDonald, D. M. (2008) The hepatocyte growth factor/c-Met
signaling pathway as a therapeutic target to inhibit angiogenesis. BMB Rep 41, 833-839
5. Sala, V., and Crepaldi, T. (2011) Novel therapy for myocardial infarction: can HGF/Met be beneficial? Cell Mol Life Sci 68, 1703-1717
6. Nakamura, T., Sakai, K., Nakamura, T., and Matsumoto, K. (2011) Hepatocyte growth factor twenty years on: Much more than a growth factor. J Gastroenterol Hepatol 26 Suppl 1, 188-202
7. McKinnon, H., Gherardi, E., Reidy, M., and Bowyer, D. (2006) Hepatocyte growth factor/scatter factor and MET are involved in arterial repair and atherogenesis. Am J Pathol 168, 340-348
8. Kuhlmann, C. R., Schaefer, C. A., Fehsecke, A., Most, A. K., Tillmanns, H., and Erdogan, A. (2005) A new signaling mechanism of hepatocyte growth factor-induced endothelial proliferation. J Thromb Haemost 3, 2089-2095
9. Graupera, M., and Potente, M. (2013) Regulation of angiogenesis by PI3K signaling networks. Exp Cell Res 319, 1348-1355
10. Jagadeeswaran, R., Jagadeeswaran, S., Bindokas, V. P., and Salgia, R. (2007) Activation of HGF/c-Met pathway contributes to the reactive oxygen species generation and motility of small cell lung cancer cells. Am J Physiol Lung Cell Mol Physiol 292, L1488-1494
11. Schroder, K., Schutz, S., Schloffel, I., Batz, S., Takac, I., Weissmann, N., Michaelis, U. R., Koyanagi, M., and Brandes, R. P. (2011) Hepatocyte growth factor induces a proangiogenic phenotype and mobilizes endothelial progenitor cells by activating Nox2. Antioxid Redox Signal 15, 915-923
12. Hoelzle, M. K., and Svitkina, T. (2012) The cytoskeletal mechanisms of cell-cell junction formation in endothelial cells. Mol Biol Cell 23, 310-323
13. Insall, R. H., and Machesky, L. M. (2009) Actin dynamics at the leading edge: from simple machinery to complex networks. Dev Cell 17, 310-322
14. Le Clainche, C., and Carlier, M. F. (2008) Regulation of actin assembly associated with protrusion and adhesion in cell migration. Physiol Rev 88, 489-513
15. Burnette, D. T., Manley, S., Sengupta, P., Sougrat, R., Davidson, M. W., Kachar, B., and Lippincott-Schwartz, J. (2011) A role for actin arcs in the leading-edge advance of migrating cells. Nature cell biology 13, 371-381
16. Chichili, G. R., and Rodgers, W. (2009) Cytoskeleton-membrane interactions in membrane raft structure. Cell Mol Life Sci 66, 2319-2328
17. Martinelli, R., Kamei, M., Sage, P. T., Massol, R., Varghese, L., Sciuto, T., Toporsian, M., Dvorak, A. M., Kirchhausen, T., Springer, T. A., and Carman, C. V. (2013) Release of cellular tension signals self-restorative ventral lamellipodia to heal barrier micro-wounds. J Cell Biol 201, 449-465
18. Sung, B. H., Zhu, X., Kaverina, I., and Weaver, A. M. (2011) Cortactin controls cell motility and lamellipodial dynamics by regulating ECM secretion. Curr Biol 21, 1460-1469
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from

12
19. Vinzenz, M., Nemethova, M., Schur, F., Mueller, J., Narita, A., Urban, E., Winkler, C., Schmeiser, C., Koestler, S. A., Rottner, K., Resch, G. P., Maeda, Y., and Small, J. V. (2012) Actin branching in the initiation and maintenance of lamellipodia. J Cell Sci 125, 2775-2785
20. Viola, A., and Gupta, N. (2007) Tether and trap: regulation of membrane-raft dynamics by actin-binding proteins. Nat Rev Immunol 7, 889-896
21. Bryce, N. S., Clark, E. S., Leysath, J. L., Currie, J. D., Webb, D. J., and Weaver, A. M. (2005) Cortactin promotes cell motility by enhancing lamellipodial persistence. Curr Biol 15, 1276-1285
22. Koestler, S. A., Rottner, K., Lai, F., Block, J., Vinzenz, M., and Small, J. V. (2009) F- and G-actin concentrations in lamellipodia of moving cells. PloS one 4, e4810
23. Lai, F. P., Szczodrak, M., Oelkers, J. M., Ladwein, M., Acconcia, F., Benesch, S., Auinger, S., Faix, J., Small, J. V., Polo, S., Stradal, T. E., and Rottner, K. (2009) Cortactin promotes migration and platelet-derived growth factor-induced actin reorganization by signaling to Rho-GTPases. Mol Biol Cell 20, 3209-3223
24. Weed, S. A., Karginov, A. V., Schafer, D. A., Weaver, A. M., Kinley, A. W., Cooper, J. A., and Parsons, J. T. (2000) Cortactin localization to sites of actin assembly in lamellipodia requires interactions with F-actin and the Arp2/3 complex. J Cell Biol 151, 29-40
25. Pendyala, S., Usatyuk, P. V., Gorshkova, I. A., Garcia, J. G., and Natarajan, V. (2009) Regulation of NADPH oxidase in vascular endothelium: the role of phospholipases, protein kinases, and cytoskeletal proteins. Antioxid Redox Signal 11, 841-860
26. Usatyuk, P. V., Gorshkova, I. A., He, D., Zhao, Y., Kalari, S. K., Garcia, J. G., and Natarajan, V. (2009) Phospholipase D-mediated activation of IQGAP1 through Rac1 regulates hyperoxia-induced p47phox translocation and reactive oxygen species generation in lung endothelial cells. J Biol Chem 284, 15339-15352
27. Usatyuk, P. V., Romer, L. H., He, D., Parinandi, N. L., Kleinberg, M. E., Zhan, S., Jacobson, J. R., Dudek, S. M., Pendyala, S., Garcia, J. G., and Natarajan, V. (2007) Regulation of hyperoxia-induced NADPH oxidase activation in human lung endothelial cells by the actin cytoskeleton and cortactin. J Biol Chem 282, 23284-23295
28. Usatyuk, P. V., Singleton, P. A., Pendyala, S., Kalari, S. K., He, D., Gorshkova, I. A., Camp, S. M., Moitra, J., Dudek, S. M., Garcia, J. G., and Natarajan, V. (2012) Novel role for non-muscle myosin light chain kinase (MLCK) in hyperoxia-induced recruitment of cytoskeletal proteins, NADPH oxidase activation, and reactive oxygen species generation in lung endothelium. J Biol Chem 287, 9360-9375
29. Touyz, R. M., Yao, G., Quinn, M. T., Pagano, P. J., and Schiffrin, E. L. (2005) p47phox associates with the cytoskeleton through cortactin in human vascular smooth muscle cells: role in NAD(P)H oxidase regulation by angiotensin II. Arteriosclerosis, thrombosis, and vascular biology 25, 512-518
30. Mishina, N. M., Tyurin-Kuzmin, P. A., Markvicheva, K. N., Vorotnikov, A. V., Tkachuk, V. A., Laketa, V., Schultz, C., Lukyanov, S., and Belousov, V. V. (2011) Does cellular hydrogen peroxide diffuse or act locally? Antioxid Redox Signal 14, 1-7
31. Moldovan, L., Mythreye, K., Goldschmidt-Clermont, P. J., and Satterwhite, L. L. (2006) Reactive oxygen species in vascular endothelial cell motility. Roles of NAD(P)H oxidase and Rac1. Cardiovasc Res 71, 236-246
32. Pendyala, S., Gorshkova, I. A., Usatyuk, P. V., He, D., Pennathur, A., Lambeth, J. D., Thannickal, V. J., and Natarajan, V. (2009) Role of Nox4 and Nox2 in hyperoxia-induced reactive oxygen species generation and migration of human lung endothelial cells. Antioxid Redox Signal 11, 747-764
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from

13
33. Reddy, M. M., Fernandes, M. S., Salgia, R., Levine, R. L., Griffin, J. D., and Sattler, M. (2011) NADPH oxidases regulate cell growth and migration in myeloid cells transformed by oncogenic tyrosine kinases. Leukemia 25, 281-289
34. Ushio-Fukai, M. (2006) Localizing NADPH oxidase-derived ROS. Sci STKE 2006, re8
35. Kohler, E. E., Cowan, C. E., Chatterjee, I., Malik, A. B., and Wary, K. K. (2011) NANOG induction of fetal liver kinase-1 (FLK1) transcription regulates endothelial cell proliferation and angiogenesis. Blood 117, 1761-1769
36. Mishina, N. M., Markvicheva, K. N., Bilan, D. S., Matlashov, M. E., Shirmanova, M. V., Liebl, D., Schultz, C., Lukyanov, S., and Belousov, V. V. (2013) Visualization of Intracellular Hydrogen Peroxide with HyPer, a Genetically Encoded Fluorescent Probe. Methods Enzymol 526, 45-59
37. Brown, M., Adyshev, D., Bindokas, V., Moitra, J., Garcia, J. G., and Dudek, S. M. (2010) Quantitative distribution and colocalization of non-muscle myosin light chain kinase isoforms and cortactin in human lung endothelium. Microvasc Res 80, 75-88
38. Keese, C. R., Wegener, J., Walker, S. R., and Giaever, I. (2004) Electrical wound-healing assay for cells in vitro. Proc Natl Acad Sci U S A 101, 1554-1559
39. Kelley, L. C., Hayes, K. E., Ammer, A. G., Martin, K. H., and Weed, S. A. (2010) Cortactin phosphorylated by ERK1/2 localizes to sites of dynamic actin regulation and is required for carcinoma lamellipodia persistence. PloS one 5, e13847
40. Clavijo-Cornejo, D., Enriquez-Cortina, C., Lopez-Reyes, A., Dominguez-Perez, M., Nuno, N., Dominguez-Meraz, M., Bucio, L., Souza, V., Factor, V. M., Thorgeirsson, S. S., Gutierrez-Ruiz, M. C., and Gomez-Quiroz, L. E. (2013) Biphasic regulation of the NADPH oxidase by HGF/c-Met signaling pathway in primary mouse hepatocytes. Biochimie 95, 1177-1184
41. Zhou, Y. J., Yang, H. W., Wang, X. G., and Zhang, H. (2009) Hepatocyte growth factor prevents advanced glycation end products-induced injury and oxidative stress through a PI3K/Akt-dependent pathway in human endothelial cells. Life Sci 85, 670-677
42. Madonna, R., Bolli, R., Rokosh, G., and De Caterina, R. (2013) Targeting phosphatidylinositol 3-kinase-Akt through hepatocyte growth factor for cardioprotection. J Cardiovasc Med (Hagerstown) 14, 249-253
43. Ruggeri, R. M., Vitarelli, E., Barresi, G., Trimarchi, F., Benvenga, S., and Trovato, M. (2010) The tyrosine kinase receptor c-met, its cognate ligand HGF and the tyrosine kinase receptor trasducers STAT3, PI3K and RHO in thyroid nodules associated with Hashimoto's thyroiditis: an immunohistochemical characterization. Eur J Histochem 54, e24
44. Singleton, P. A., Salgia, R., Moreno-Vinasco, L., Moitra, J., Sammani, S., Mirzapoiazova, T., and Garcia, J. G. (2007) CD44 regulates hepatocyte growth factor-mediated vascular integrity. Role of c-Met, Tiam1/Rac1, dynamin 2, and cortactin. J Biol Chem 282, 30643-30657
45. Zhao, Y., He, D., Stern, R., Usatyuk, P. V., Spannhake, E. W., Salgia, R., and Natarajan, V. (2007) Lysophosphatidic acid modulates c-Met redistribution and hepatocyte growth factor/c-Met signaling in human bronchial epithelial cells through PKC delta and E-cadherin. Cell Signal 19, 2329-2338
46. Martinez-Palacian, A., del Castillo, G., Suarez-Causado, A., Garcia-Alvaro, M., de Morena-Frutos, D., Fernandez, M., Roncero, C., Fabregat, I., Herrera, B., and Sanchez, A. (2013) Mouse hepatic oval cells require Met-dependent PI3K to impair TGF-beta-induced oxidative stress and apoptosis. PloS one 8, e53108
47. Dossumbekova, A., Berdyshev, E. V., Gorshkova, I., Shao, Z., Li, C., Long, P., Joshi, A., Natarajan, V., and Vanden Hoek, T. L. (2008) Akt activates NOS3 and
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from

14
separately restores barrier integrity in H2O2-stressed human cardiac microvascular endothelium. Am J Physiol Heart Circ Physiol 295, H2417-2426
48. Ephstein, Y., Singleton, P. A., Chen, W., Wang, L., Salgia, R., Kanteti, P., Dudek, S. M., Garcia, J. G., and Jacobson, J. R. (2013) Critical role of S1PR1 and integrin beta4 in HGF/c-Met-mediated increases in vascular integrity. J Biol Chem 288, 2191-2200
49. Berthou, S., Aebersold, D. M., Schmidt, L. S., Stroka, D., Heigl, C., Streit, B., Stalder, D., Gruber, G., Liang, C., Howlett, A. R., Candinas, D., Greiner, R. H., Lipson, K. E., and Zimmer, Y. (2004) The Met kinase inhibitor SU11274 exhibits a selective inhibition pattern toward different receptor mutated variants. Oncogene 23, 5387-5393
50. Vlahos, C. J., Matter, W. F., Hui, K. Y., and Brown, R. F. (1994) A specific inhibitor of phosphatidylinositol 3-kinase, 2-(4-morpholinyl)-8-phenyl-4H-1-benzopyran-4-one (LY294002). J Biol Chem 269, 5241-5248
51. Baumer, A. T., Ten Freyhaus, H., Sauer, H., Wartenberg, M., Kappert, K., Schnabel, P., Konkol, C., Hescheler, J., Vantler, M., and Rosenkranz, S. (2008) Phosphatidylinositol 3-kinase-dependent membrane recruitment of Rac-1 and p47phox is critical for alpha-platelet-derived growth factor receptor-induced production of reactive oxygen species. J Biol Chem 283, 7864-7876
52. Chen, Q., Powell, D. W., Rane, M. J., Singh, S., Butt, W., Klein, J. B., and McLeish, K. R. (2003) Akt phosphorylates p47phox and mediates respiratory burst activity in human neutrophils. J Immunol 170, 5302-5308
53. Kim, J. H., Chu, S. C., Gramlich, J. L., Pride, Y. B., Babendreier, E., Chauhan, D., Salgia, R., Podar, K., Griffin, J. D., and Sattler, M. (2005) Activation of the PI3K/mTOR pathway by BCR-ABL contributes to increased production of reactive oxygen species. Blood 105, 1717-1723
54. Chowdhury, A. K., Watkins, T., Parinandi, N. L., Saatian, B., Kleinberg, M. E., Usatyuk, P. V., and Natarajan, V. (2005) Src-mediated tyrosine phosphorylation of p47phox in hyperoxia-induced activation of NADPH oxidase and generation of reactive oxygen species in lung endothelial cells. J Biol Chem 280, 20700-20711
55. Ohsawa, M., Kobayashi, T., Okura, H., Igarashi, T., Mizuguchi, M., and Hino, O. (2013) TSC1 controls distribution of actin fibers through its effect on function of Rho family of small GTPases and regulates cell migration and polarity. PloS one 8, e54503
56. Nobes, C. D., and Hall, A. (1995) Rho, rac, and cdc42 GTPases regulate the assembly of multimolecular focal complexes associated with actin stress fibers, lamellipodia, and filopodia. Cell 81, 53-62
57. Wozniak, M. A., Kwong, L., Chodniewicz, D., Klemke, R. L., and Keely, P. J. (2005) R-Ras controls membrane protrusion and cell migration through the spatial regulation of Rac and Rho. Mol Biol Cell 16, 84-96
58. Baker, M., Robinson, S. D., Lechertier, T., Barber, P. R., Tavora, B., D'Amico, G., Jones, D. T., Vojnovic, B., and Hodivala-Dilke, K. (2012) Use of the mouse aortic ring assay to study angiogenesis. Nature protocols 7, 89-104
59. Ryan, G. L., Petroccia, H. M., Watanabe, N., and Vavylonis, D. (2012) Excitable actin dynamics in lamellipodial protrusion and retraction. Biophys J 102, 1493-1502
60. Lamalice, L., Le Boeuf, F., and Huot, J. (2007) Endothelial cell migration during angiogenesis. Circ Res 100, 782-794
61. Schmidt, A., Brixius, K., and Bloch, W. (2007) Endothelial precursor cell migration during vasculogenesis. Circ Res 101, 125-136
62. Fountoulaki, K., and Parissis, J. (2012) Hepatocyte Growth Factor as a Prognostic Marker in Heart Failure: Promise and Challenges. Cardiology 121, 237-239
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from

15
63. Lawrence, R. E., and Salgia, R. (2010) MET molecular mechanisms and therapies in lung cancer. Cell Adh Migr 4, 146-152
64. Madonna, R., Cevik, C., Nasser, M., and De Caterina, R. (2012) Hepatocyte growth factor: molecular biomarker and player in cardioprotection and cardiovascular regeneration. Thromb Haemost 107, 656-661
65. Wondergem, R., Ecay, T. W., Mahieu, F., Owsianik, G., and Nilius, B. (2008) HGF/SF and menthol increase human glioblastoma cell calcium and migration. Biochem Biophys Res Commun 372, 210-215
66. Ammer, A. G., and Weed, S. A. (2008) Cortactin branches out: roles in regulating protrusive actin dynamics. Cell Motil Cytoskeleton 65, 687-707
67. Raftopoulou, M., and Hall, A. (2004) Cell migration: Rho GTPases lead the way. Dev Biol 265, 23-32
68. Sattler, M., Reddy, M. M., Hasina, R., Gangadhar, T., and Salgia, R. (2011) The role of the c-Met pathway in lung cancer and the potential for targeted therapy. Ther Adv Med Oncol 3, 171-184
69. Ye, M., Hu, D., Tu, L., Zhou, X., Lu, F., Wen, B., Wu, W., Lin, Y., Zhou, Z., and Qu, J. (2008) Involvement of PI3K/Akt signaling pathway in hepatocyte growth factor-induced migration of uveal melanoma cells. Invest Ophthalmol Vis Sci 49, 497-504
70. Hilpela, P., Vartiainen, M. K., and Lappalainen, P. (2004) Regulation of the actin cytoskeleton by PI(4,5)P2 and PI(3,4,5)P3. Curr Top Microbiol Immunol 282, 117-163
71. Stampoulis, P., Ueda, T., Matsumoto, M., Terasawa, H., Miyano, K., Sumimoto, H., and Shimada, I. (2012) Atypical membrane-embedded phosphatidylinositol 3,4-bisphosphate (PI(3,4)P2)-binding site on p47(phox) Phox homology (PX) domain revealed by NMR. J Biol Chem 287, 17848-17859
72. Wu, R. F., Xu, Y. C., Ma, Z., Nwariaku, F. E., Sarosi, G. A., Jr., and Terada, L. S. (2005) Subcellular targeting of oxidants during endothelial cell migration. J Cell Biol 171, 893-904
73. Taulet, N., Delorme-Walker, V. D., and DerMardirossian, C. (2012) Reactive oxygen species regulate protrusion efficiency by controlling actin dynamics. PloS one 7, e41342
74. Delorme, V., Machacek, M., DerMardirossian, C., Anderson, K. L., Wittmann, T., Hanein, D., Waterman-Storer, C., Danuser, G., and Bokoch, G. M. (2007) Cofilin activity downstream of Pak1 regulates cell protrusion efficiency by organizing lamellipodium and lamella actin networks. Dev Cell 13, 646-662
75. Kim, J. S., Huang, T. Y., and Bokoch, G. M. (2009) Reactive oxygen species regulate a slingshot-cofilin activation pathway. Mol Biol Cell 20, 2650-2660
76. Frantz, C., Barreiro, G., Dominguez, L., Chen, X., Eddy, R., Condeelis, J., Kelly, M. J., Jacobson, M. P., and Barber, D. L. (2008) Cofilin is a pH sensor for actin free barbed end formation: role of phosphoinositide binding. J Cell Biol 183, 865-879
77. Hung, R. J., Spaeth, C. S., Yesilyurt, H. G., and Terman, J. R. (2013) SelR reverses Mical-mediated oxidation of actin to regulate F-actin dynamics. Nature cell biology 15, 1445-1454
78. Hung, R. J., Pak, C. W., and Terman, J. R. (2011) Direct redox regulation of F-actin assembly and disassembly by Mical. Science (New York, N.Y 334, 1710-1713
79. Urao, N., Sudhahar, V., Kim, S. J., Chen, G. F., McKinney, R. D., Kojda, G., Fukai, T., and Ushio-Fukai, M. (2013) Critical role of endothelial hydrogen peroxide in post-ischemic neovascularization. PloS one 8, e57618
80. Hodara, R., Weiss, D., Joseph, G., Velasquez-Castano, J. C., Landazuri, N., Han, J. W., Yoon, Y. S., and Taylor, W. R. (2011) Overexpression of catalase in myeloid
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from

16
cells causes impaired postischemic neovascularization. Arteriosclerosis, thrombosis, and vascular biology 31, 2203-2209
81. Oshikawa, J., Urao, N., Kim, H. W., Kaplan, N., Razvi, M., McKinney, R., Poole, L. B., Fukai, T., and Ushio-Fukai, M. (2010) Extracellular SOD-derived H2O2 promotes VEGF signaling in caveolae/lipid rafts and post-ischemic angiogenesis in mice. PloS one 5, e10189
Acknowledgments: This work was supported by grants from National Institutes of Health PO1 HL058064 to V.N.; K.K.W. was supported by AHA grant (12GRNT12070159) and NIH (2R01HL079356). The abbreviations used are: ECs, endothelial cells; HGF, hepatocyte growth factor; HLMVECs, human lung microvascular endothelial cells; ROS, reactive oxygen species; TER, trans endothelial electrical resistance. Conflict of interest: The authors declare that no conflicting interests exist.
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from

17
FIGURE LEGENDS Fig. 1. HGF modulates lamellipodia formation in HLMVECs. HLMVECs grown on slide chambers were treated with HGF for 15 min, probed with anti-actin and anti-cortactin antibodies, and examined by immunofluorescence microscopy. (A) Actin and cortactin reorganization to lamellipodia was visualized by immunocytochemistry as described in Material and Methods. Nuclei were stained with DAPI. Shown are representative immunofluorescence images from three independent experiments. (B) Actin and cortactin co-localization in lamellipodia shown in Panel A was quantified using an image analysis as described under Materials and Methods. Values are means + S.D. from three independent experiments. *, p<0.05 compared to cells without HGF.
Fig. 2. HGF enhances hydrogen peroxide formation in HLMVECs. (A) HLMVECs grown on glass bottom dishes to ~70% confluence were transfected with pHyPer-cyto plasmid for 72 h, which responses with enhanced green fluorescence to hydrogen peroxide changes. Cells were treated for 15 min with different concentrations of HGF and living cells were analyzed by Nikon immunofluorescence microscope (lens x60) as described in Material and Methods. Nuclei were stained with Hoechst. Shown are representative images from three independent experiments. (B) pHyPer-cyto immunofluorescence in cell periphery was quantified from panel A using an image analysis. Values are means + S.D. from three independent experiments. . * Significantly different from untreated-control (p < 0.05). (C) Time-course of HGF-induced pHyPer-cyto fluorescence in response to HGF (15 min, 20 ng/ml) was analyzed by confocal microscopy (lens x60) as described in Material and Methods. (D) ROS production in non treated (Control) and HGF-treated (15 min) are shown. Bars, 20 µm. Fig. 3. HGF induces hydrogen peroxide formation in lamellipodia of HLMVECs. (A) HLMVECs grown on glass bottom dishes to ~70% confluence were double transfected with pHyPer-cyto/Actin-RFP (A) or pHyPer-cyto/Cotactin-RFP (B) plasmid (0.5/0.5 µg/ml ) for 72 h. Living cells were analyzed by confocal fluorescence microscopy (lens x40) as described in Material and Methods. Shown are representative images from three independent experiments. Distribution of pHyPer-cyto immunofluorescence in lamellipodia (10 µm) with Actin-RFP (C) and Cortactin-RFP (D) was quantified from upper panels using an image analysis software and expressed as Relative Intensity Units. Bars, 20 µm. Fig. 4. HGF stimulates c-Met phosphorylation in HLMVECs. HLMVECs grown to ~95% confluence on 100-mm dishes were treated with varying concentrations of HGF for 15 min (A) or pretreated with 1 µM SU11274 (c-Met inhibitor) (B) for 1h, prior to challenge with HGF (20 ng/ml) for 15 min. Cell lysates (20-40 µg of protein) were subjected to 10 % SDS-PAGE, probed with total- or anti-phospho-c-Met (Y1234/1235, Y1349, Y1003, Y1313, S985), and anti-actin antibodies as indicated. Shown are representative blots from three independent experiments. Western blots were scanned and quantified by automated digitizing system. Values are means + S.E.M. * Significantly different from untreated-control (p < 0.05). Fig. 5. Downregulation of c-Met phosphorylation attenuates PI3k and Akt activation in HLMVECs. HLMVECs were transfected with c-Met siRNA (50 nM, 72 h) (A) or pretreated with 1 µM SU11274 (c-Met inhibitor) (B) for 1h, prior to challenge with HGF (20 ng/ml) for 15 min. Cell lysates (20-40 µg of protein) were subjected to 10 % SDS-PAGE, and probed with total- and anti-phospho PI3k (p85), total- and anti-phospho Akt (T308, S473), and anti-actin antibodies as described under Materials and Methods. Shown are representative blots from three independent experiments. Western blots were scanned and quantified by automated digitizing
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from

18
system. Values are means + S.E.M. * Significantly different from untreated-control cells (p < 0.05); ** Significantly different from HGF-treated cells (p<0.05). Fig. 6. HGF-mediated c-Met phosphorylation is essential for lamellipodia formation, hydrogen peroxide accumulation in lamellipodia and cell migration in HLMVECs. (A) HLMVECs grown to ~95% confluence on slide chambers were pre-treated with 1 µM SU11274 for 1 h prior to stimulation with HGF (20 ng/ml) for 15 min. Cells were washed, fixed and stained for actin and cortactin co-localization in lamellipodia as described under Material and Methods. Nuclei were stained with DAPI. Shown are representative immunofluorescence images from three independent experiments. Actin and cortactin co-localization in lamellipodia was quantified using an image analysis as described under Materials and Methods. Values are means + S.E.M from three independent experiments. * Significantly different from untreated-control cells (p < 0.05); ** Significantly different from HGF-treated cells (p<0.05). (B) HLMVECs grown on glass bottom dishes to ~60 % confluence were transfected with pHyPer-cyto plasmid for 72 h, as described in Fig. 1. Cells were pre-treated with SU11274 (1 µM) for 1 h prior to HGF (20 ng/ml) challenge for 15 min. Live cells were analyzed by Nikon immunofluorescence microscope (lens x60) as described in Material and Methods. pHyPer-cyto immunofluorescence in lamellipodia was quantified using an image analysis. Values are means + S.E.M. from three independent experiments. . * Significantly different from untreated-control (p < 0.05); ** Significantly different from HGF-treated cells without SU11274 (p<0.05). (C) HLMVECs grown on gold electrodes to ~95% confluence were pre-treated with 1 µM SU11274 for 1 h prior to HGF (20 ng/ml) stimulation. Transendothelial electrical resistance (TER) was recorded using an electrical cell substrate impedance-sensing system (ECIS) for 12 h, and HGF-mediated wound repair (% control) was calculated from TER tracings as described under Materials and Methods. Values are means + S.E.M. from three independent experiments. * Significantly different from untreated-control (p < 0.05); ** Significantly different from HGF-treated cells (p<0.05).
Fig. 7. Inhibition of HGF-induced PI3k signaling attenuates Akt activation, hydrogen peroxide accumulation in lamellipodia and cell migration in HLMVECs. (A) HLMVECs grown to ~95% confluence in 100-mm dishes were pretreated with 10 µM LY294002 (PI3k inhibitor) for 1 h, prior to challenge with HGF (20 ng/ml). Cell lysates (20-40 µgs of protein) were subjected to 10 % SDS-PAGE, probed with anti-phospho-PI3k, anti-phospho-Akt (T308, S473), anti-PI3k and anti-actin antibodies as indicated and fold changes were determined from the respective Western blots by image analysis. Shown are representative blots from 3 independent experiments. Values are means ± S.E.M. Significantly different from untreated-control (p< 0.05); ** Significantly different from HGF-treated cells without LY294002 (p<0.05). (B) HLMVECs grown to ~60% confluence in 60-mm dishes were transfected with scrambled (sc) or PI3k siRNA (50 nM, 72 h) prior to stimulation with HGF (20 ng/ml) for 15 min. Cell lysates were subjected to 10 % SDS-PAGE and Western blotting with antibodies as described under Fig. 6A. Western blots were scanned and quantified by image analysis. Shown are representative bots from three independent experiments and values are means ± S.E.M. Significantly different from untreated-control (p< 0.05); ** Significantly different from scrambled siRNA transfected and HGF-treated cells (p<0.05). (C) HLMVECs grown on slide chambers to ~90% confluence were pretreated with PI3k inhibitor LY294002 (10 µM) for 1 h prior to stimulation with HGF (20 ng/ml) for 15 min. (D) HLMVECs grown on slide chambers to ~60% confluence were transfected with scrambled or PI3k siRNA (50 nM, 72 h) prior to stimulation with HGF (20 ng/ml) for 15 min. In (C) and (D), cells were analyzed for co-localization of actin and cortactin by Nikon immunofluorescence microscope (lens x60) as described in Material and Methods. Shown are representative immunofluorescence images from three independent experiments, and values are means + S.E.M. Nuclei were stained with DAPI. * Significantly different from untreated or scrambled siRNA transfected control cells (p < 0.05); ** Significantly different from
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from

19
HGF-treated cells without LY294002 or PI3k siRNA (p<0.05). (E) HLMVECs grown on glass bottom dishes to ~90% confluence were pretreated with PI3k inhibitor LY294002 (10 µM) for 1 h prior to stimulation with HGF (20 ng/ml) for 15 min. (F) HLMVECs grown on glass bottom dishes to ~60% confluence were transfected with scrambled or PI3k siRNA (50 nM, 72 h) prior to stimulation with HGF (20 ng/ml) for 15 min. In (E) and (F), live cells were analyzed by Nikon immunofluorescence microscope (lens x60) for pHyPer-cyto immunofluorescence (index of hydrogen peroxide formation) in lamellipodia and was quantified using an image analysis. Values are means + S.E.M. from three independent experiments. * Significantly different from untreated-control (p < 0.05); ** Significantly different from HGF-treated cells without LY2094002 or PI3k siRNA (p<0.05). (G) HLMVECs grown on gold electrodes to ~95% confluence were pre-treated with PI3k inhibitor LY294002 (10 µM) for 1 h prior to HGF (20 ng/ml) stimulation. (H) HLMVECs grown on gold electrodes to ~60% confluence were transfected with scrambled or PI3k siRNA (50 nM, 72 h) prior to HGF (20 ng/ml) stimulation. Transendothelial electrical resistance (TER) was recorded using an electrical cell substrate impedance-sensing system (ECIS) for 12 h. HGF-mediated wound repair (% control) was calculated from the tracings as described under Materials and Methods. Shown is a representative tracing from three independent experiments. Values are means + SEM. * Significantly different from untreated-control cells (p < 0.05); ** Significantly different from HGF-treated cells without LY2094002 or PI3k siRNA control cells (p<0.05). Fig. 8. Inhibition of c-Met/PI3k/Akt signaling attenuates p47phox, cortactin and Rac1 redistribution to lamellipodia in HLMVECs. HLMVECs grown to ~95% confluence on slide chambers were pretreated with 1 µM SU11274, a c-Met inhibitor, or 10 µM LY294002, a PI3k inhibitor for 1 h. For down-regulation of PI3k with siRNA, HLMVECs were transfected with scrambled or PI3k siRNA (50 nM, 72 h). Cells were challenged with HGF (20 ng/ml). p47phox, and cortactin (A) and Rac1 (B) redistribution to lamellipodia were analyzed by fluorescent microscopy (lens x60). Nuclei were stained with DAPI. Shown are representative immunofluorescence images from three independent experiments. Co-localization of p47phox/cortactin (C) and Rac1 (D) in lamellipodia was quantified as described in Material and Methods. Values are means + S.E.M. * Significantly different from untreated-control (p< 0.05); ** Significantly different from HGF-treated cells without SU11274, LY294002 or PI3k siRNA (p<0.05).
Fig. 9. Down-regulation of p47phox and cortactin or inhibition of NADPH oxidase attenuates lamellipodia formation, and hydrogen peroxide accumulation in lamellipodia of HLMVECs. HLMVECs grown on slide chambers to ~60% confluence were transfected with scrambled or siRNA for p47phox and cortactin (50 nM, 72 h). In experiments with apocynin, HLMVECs grown to ~95% confluence on slide chambers were pre-treated with 30 µM Apocynin, a NADPH oxidase inhibitor for 1 h. Cells were challenged with HGF (20 ng/ml) for 15 min. (A) Actin and cortactin co-localization in lamellipodia was visualized by immunocytochemistry and quantified using an image analysis as described under Materials and Methods. Nuclei were stained with DAPI. Values are means + S.E.M. from three independent experiments. * Significantly different from untreated-control (p< 0.05); ** Significantly different from HGF-treated cells without apocynin, p47phox siRNA or cortactin siRNA (p<0.05). (B) HLMVECs grown on glass bottom dishes to ~60% confluence were transfected with pHyPer-cyto plasmid as described in Fig. 1. Additionally, some cells, as indicated, were also transfected with scrambled siRNA (50 nM), p47phox siRNA (50 nM), cortactin siRNA (50 nM) for 72 h or treated with apocynin (30 µM) for 1 h. Cells were challenged with HGF (20 ng/ml) for 15 min and living cells were analyzed by fluorescent microscopy (lens x60) and accumulation of hydrogen peroxide, as measured by pHyPer-cyto fluorescence, in lamellipodia was estimated as described in Material and Methods. (C and D) p47phox (C) and Rac1 (D) redistribution to lamellipodia was
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from

20
quantified as described in Material and Methods. Down regulation of p47phox or cortactin with siRNA was confirmed by Western blotting (inserts). Values are means + S.E.M. of three independent experiments. * Significantly different from untreated-control (p< 0.05); **Significantly different from HGF-treated cells without apocynin, p47phox siRNA or cortactin siRNA (p<0.01). Fig. 10. Down-regulation of p47phox and cortactin or inhibition of NADPH oxidase attenuates cell migration of HLMVECs. (A) In experiments with apocynin, HLMVECs grown on gold electrodes to ~90% confluence were pre-treated with apocynin (30 µM) for 1 h prior to HGF (20 ng/ml) stimulation. (B and C) HLMVECs grown on gold electrodes to ~60% confluence were transfected with scrambled, p47phox siRNA (50 nM, 72 h) (B) or cortactin siRNA (50 nM, 72 h) (C) prior to HGF (20 ng/ml) stimulation. Transendothelial electrical resistance (TER) was recorded using an electrical cell substrate impedance-sensing system (ECIS) for 12 h. HGF-mediated wound repair (% control) was calculated from the tracings as described under Materials and Methods. Values are means + S.E.M. from three independent experiments. * Significantly different from untreated-control (P < 0.05); ** Significantly different from HGF-treated cells without apocynin, p47phox siRNA or cortactin siRNA (p<0.05). Fig. 11. HGF potentiates endothelial sprouting formation in mouse aortic rings. Aortic rings (1 mm) isolated from mouse thoracic aortae were grown in fibrin matrix for 6 days without (Control) and in the presence of 20 ng/ml HGF and sprouts outgrowth (A) was examined using Zeiss Axiovert 40 phase contrast microscope (lens x10) as described under Materials and Methods. (B) Aortic rings grown in fibrin matrix were infected for 24 h with CellLight Actin-RFP BacMam 2.0 according to manufacturer’s protocol, pretreated with 1 µM SU11274 (c-Met inhibitor) for 1h prior to challenge with HGF (20 ng/ml). Living cells transiently expressing Actin-RFP were visualized by laser scanning microscope Zeiss 510 Meta (lens x40). Actin dynamics were monitored before and after HGF stimulation. Insert depicts enhanced Actin-RFP accumulation in lamellipodia due to HGF treatment which was significantly abrogated by c-Met inhibitor (C). Values are means + S.E.M. * Significantly different from untreated-control cells (p < 0.05); ** Significantly different from HGF-treated cells (p<0.05). Bars, 20 µm. Fig. 12. Proposed model on the role for c-Met/PI3k/Akt signaling in HGF-induced lamellipodia formation and cell motility in lung endothelium. As depicted in the model, c-Met/PI3k/Akt signaling cascade modulates: i) Actin and Cortactin, redistribution to cell periphery; ii) translocation of p47phox, Rac1, assembly of NADPH oxidase and ROS production in cell periphery; iii) lamellipodia formation and cell motility; iv) downregulation of Met/PI3k/Akt or NADPH oxidase abolished described effects suggesting the role of HGF in EC migration, an important component of angio- and vasculo-genesis.
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from
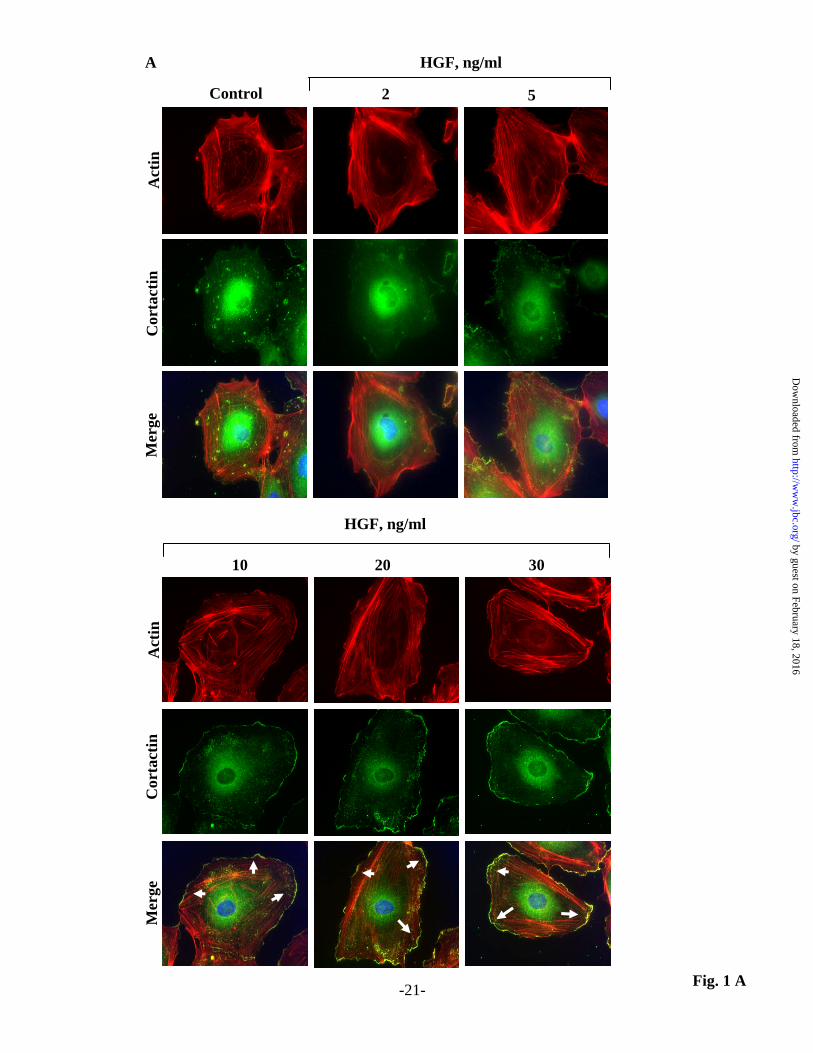
Fig. 1 A
Control 2 5
HGF, ng/ml
Act
inC
orta
ctin
Mer
ge
10 20 30
Act
inC
orta
ctin
Mer
ge
HGF, ng/ml
A
-21-
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from

Fig. 1 B
Control 2 5
HGF, ng/ml
10 20 30
0
600
200
400
Lam
ellip
odia
For
mat
ion,
%
Con
trol
*
*
*
**
B
-22-
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from

Fig. 2 AB
Control 2 ng/ml 5 ng/ml
10 ng/ml 20 ng/ml 30 ng/ml
A
0
800
200
400
pHyP
er F
luor
esce
nce,
%
Con
trol 600 *
*
*
HGF, ng/ml
B
Control 2 5 10 20 30
-23-
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from

C
Fig. 2 CD
Control HGFD
-24-
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from

Con
trol
HG
FActin-RFP pHyPer-cyto Merge
Cortactin-RFP pHyPer-cyto
Con
trol
HG
F
Merge
A
B
Fig. 3 AB-25-
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from

D pHyPer-cyto
Cortactin-RFP
0 2 4 6 8 10
Distance, µm
Control
2
4
6
8
10
12
14
Rel
ativ
e In
tens
ity, (
A.U
. x 1
03 )
0 2 4 6 8 10
Distance, µm
HGF
2
4
6
8
10
12
14
Rel
ativ
e In
tens
ity, (
A.U
. x 1
03 )
Fig. 3 CD-26-
C Actin-RFP
0 2 4 6 8 10
Distance, µm
Rel
ativ
e In
tens
ity, (
A.U
. x 1
03 )
20
40
60
0 2 4 6 8 10
Distance, µm
Rel
ativ
e In
tens
ity, (
A.U
. x 1
03 )
20
40
60
pHyPer-cyto
HGFControl
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from
IB: p-c-Met Y1003
- 2 5 10 20 30
HGF, ng/ml
IB: Actin
IB: c-Met
IB: p-c-Met Y1234/1235
IB: p-c-Met Y1349
Fig. 4 A
IB: p-c-Met S985
3
8
0
6
9
0246
0
2
4
6
p-c-MetY1234/1235
p-c-Met Y1349
p-c-Met S985
*
**
*
**
**
**
* *
Fold
Incr
ease
Fold
Incr
ease
Fold
Incr
ease
0
1
2
3p-c-Met Y1003
* * *
Fold
Incr
ease
IB: p-c-Met Y1313
4
0123
Fold
Incr
ease
*
**
*
- 2 5 10 20 30
HGF, ng/ml
p-c-Met Y1313
A
-27-
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from

SU
HGF - + - +
IB: p-c-Met Y1234/1235
IB: p-c-Met S985
IB: p-c-Met Y1349
Fig. 4 B
IB: c-Met
IB: p-c-Met Y1003
Control
IB: p-c-Met Y1313
Actin
p-c-MetY1234/1235
30
6
9 *
Fold
Incr
ease
**
8
0246
Fold
Incr
ease
*
**
p-c-Met Y1349
02
46
Fold
Incr
ease *
**
p-c-Met S985
p-c-Met Y1003p-c-Met Y1003*
**
Fold
Incr
ease
0
12
3
p-c-Met Y1313*
**
4
0123
Fold
Incr
ease
SU
HGF - + - +
Control
-28-
B
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from

SU
HGF - + - +
IB: Actin
Control
IB: p-Akt S473
IB: p-Akt T308
IB: Akt
IB: p-PI3k
Actin
IB: PI3k
SUControl*
**
8
0246
Fold
Incr
ease p-Akt T308
SU
HGF - + - +
p-Akt S473*
**
8
0246
Control
Fold
Incr
ease
4
0123
Fold
Incr
ease *
**p-PI3k
Fig. 5 A
A
-29-
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from

Fig. 5 B-30-
IB: p-Akt S473
IB: p-Akt T308
IB: Actin
IB: p-PI3k
8
0246
Fold
Incr
ease
si c-Met
HGF - + - +
scsi c-Met
HGF - + - +
sc
12
0369
Fold
Incr
ease
IB: Akt
IB: c-Met
Fold
Incr
ease
0
1
2
IB: PI3k
*
**
p-Akt T308
B
*
**
p-PI3k
p-Akt S473
**
*
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from

Fig. 6 AB
Control
HGF SU + HGF
SU
Control HGF
*
**
SU + HGFSU0
300
200
400
Lam
ellip
odia
For
mat
ion,
%
Con
trol
100
A
Control HGF SU + HGFSU0
200
400
600
pHyP
er F
luor
esce
nce,
%
Con
trol
B*
**
-31-
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from

Fig. 6 C
0
SU
Control
SU+HGF
HGF
HGF
Wound
42 86 1210Time, h
1.0
0.5
1.5
Wou
nd R
epai
r, %
Con
trol
200
100
300
Control
HGF - + - +
SU
*
**
0
C
-32-
Nor
m. T
ER
, Ohm
s x10
3
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from

Fig. 7 AB
B
IB: p-Akt S473
IB: p-Akt T308
IB: Actin
IB: p-PI3k
*
**
8
0246
Fold
Incr
ease p-Akt T308
siPI3k
HGF - + - +
scsiPI3k
HGF - + - +
sc
8
0246
Fold
Incr
ease p-Akt S473
***
IB: Akt
IB: p-Akt S473
IB: p-Akt T308
IB: Akt
IB: Actin
A
LY
HGF - + - +
*
**
8
0246
Fold
Incr
ease
p-Akt T308
8
0246
Control
Fold
Incr
ease p-Akt S473*
**
LY
HGF - + - +
Control
IB: p-PI3k
IB: PI3k
p-PI3k
p-PI3k
Fold
Incr
ease
Fold
Incr
ease
4
0123
4
0123
***
*
**
IB: PI3k
-33-
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from

Fig. 7 C
Control
HGF
LY
LY + HGF
Control HGF0
400
600
200
LY+HGFLYLam
ellip
odia
For
mat
ion,
%
Con
trol
*
**
C
-34-
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from

sc
sc + HGF siPI3k + HGF
siPI3k
Lam
ellip
odia
For
mat
ion,
%
Con
trol
0
400
600
200
sc sc + HGF
*
**
siPI3k + HGFsiPI3k
D
Fig. 7 D-35-
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from

Fig. 7 EF
0
200
400
600
*
**
E
0
200
400
pHyP
er F
luor
esce
nce,
% C
ontr
ol 600
*
**
F
siPI3k
HGF - + - +
scLY
HGF - + - +
Control
pHyP
er F
luor
esce
nce,
% C
ontr
ol
-36-
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from

LY
Control
LY+HGF
HGF
HGF
Wound
si
sc
si + HGF
sc + HGF
HGF
Wound
H
Wou
nd R
epai
r, %
Con
trol
200
100
0
*
**
300
Control LY
Wou
nd R
epai
r, %
Con
trol
200
100
0
300
*
**
HGF - + - +
siPI3ksc
0
Nor
m. T
ER
, Ohm
s x10
3
G
Fig 7 GH
42 86 1210Time, h
HGF - + - +
0
1.0
0.5
1.5
42 86 1210Time, h
-37-
Nor
m. T
ER
, Ohm
s x10
3
1.0
0.5
1.5
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from

Fig 8 A
p47phox/Cortactin
Control HGF
SU SU + HGF
LY LY + HGF
si si + HGF
A
-38-
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from

Fig. 8 B
Control HGFRac1
Control HGF
SU SU + HGF
LY LY + HGF
si si + HGF
B
-39-
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from

Fig. 8 C D
Control0
100
500
400
300
200
SU LY siPI3k
p47ph
ox/C
orta
ctin
, (%
Con
trol
)VehHGF
**
C
Control0
100
400
300
200
SU LY siPI3k
Rac
1,
(% C
ontr
ol)
VehHGF
D
-40-
*
*
** **
** ****
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from

0
100
200
300
400
*
****
**
VehHGF
100
200
300
400
0
Lam
ellip
odia
, (%
Con
trol
)
Control Apocynin sip47 siCortactin
A
0
100
200
300
400
500
600
*
****
**
VehHGF
100
200300
400
0HyP
er-C
yto,
(%
Con
trol
)
Control Apocynin sip47 siCortactin
500600
B
0
100
200
300
400
500
600
*
** ****
VehHGF
100
200300
400
0
p47,
(%
Con
trol
)
Control Apocynin sip47 siCortactin
500
600C
0
100
200
300
400
*
**
VehHGF
100
200
300
400
0
Rac
1,
(% C
ontr
ol)
Control Apocynin sip47 siCortactin
D
Fig. 9 A-D
sc si
IB: p47phox
IB: Actin
sc si
IB: Actin
IB: Cortactin
-41-
**
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from

Fig. 10-42-
0
1.0
0.5
1.5
42 6 1210
Time, h
Control
HGF
Apocynin
Apocynin + HGF
Wound
42 6 1210
Time, h
0
1.0
0.5
1.5
Control
HGF
sip47
sip47 + HGF
Wound
42 6 1210
Time, h0
1.0
0.5
1.5
Control
HGF
siCortactin
siCortactin + HGF
Wound
Wou
nd R
epai
r, %
Con
trol
200
100
0
Control Apocynin
HGF - + - +
300
Control sip47
Wou
nd R
epai
r, %
Con
trol
200
100
0
300
*
**
Control siCortactin
Wou
nd R
epai
r, %
Con
trol
200
100
0
300*
**
*
**
A
B
C
Nor
m. T
ER
, Ohm
s x10
3N
orm
. TE
R, O
hms x
103
Nor
m. T
ER
, Ohm
s x10
3
HGF
HGF
HGF
8
8
8
HGF - + - +
HGF - + - +
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from
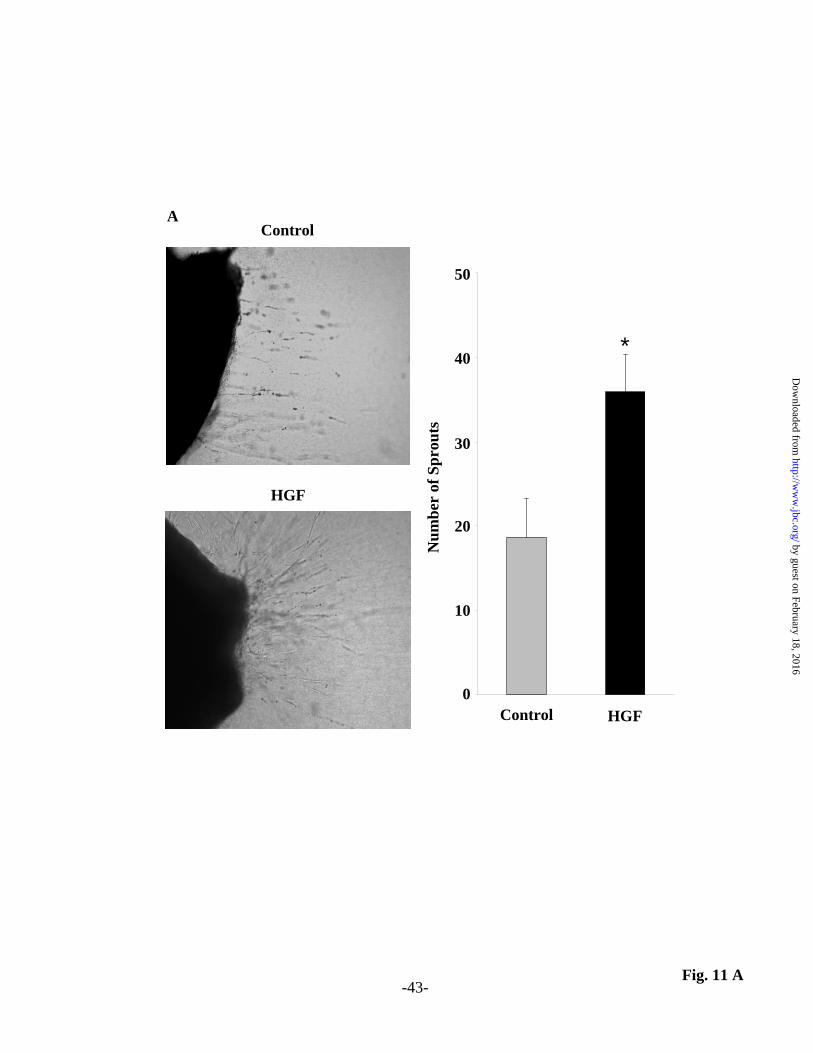
Control
HGF
Control HGF
*
Num
ber
of S
prou
ts
0
10
20
30
40
50
Fig. 11 A
A
-43-
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from
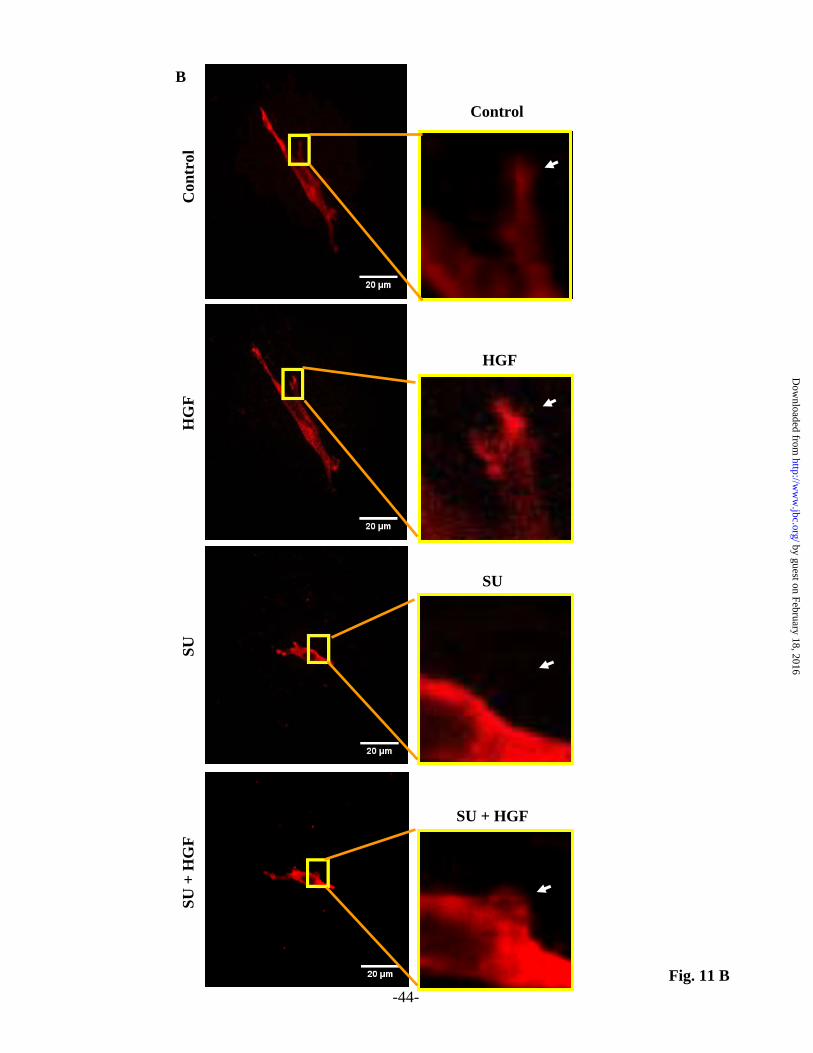
Con
trol
HG
FSU
+ H
GF
SU
Fig. 11 B
B
Control
HGF
SU + HGF
SU
-44-
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from

Fig. 11 C
4
1
3
SU
HGF - + - +
Control
0
2
*
**
Mea
n L
amel
lipod
iaIn
tens
ity, (
Fold
Incr
ease
)
-45-
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from

p47
Cortactin
c-Met
PI3k
Akt
ActinRac1
c-Met
P
P
P PI3k
HGF
AktP
NADPH oxidase
O2-.
p47Rac1
LeadingLeadingEdgeEdge
NADPH oxidase
SU, siRNA
LY, siRNA
Apo, siRNA
Fig. 12 -46-
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from

G. N. Garcia and Viswanathan NatarajanGomez-Cambronero, Kishore K Wary, Vytas Bindokas, Steven M. Dudek, Ravi Salgia, Joe
Peter V. Usatyuk, Panfeng Fu, Vijay Mohan, Yulia Epshtein, Jeffrey R. Jacobson, JulianGeneration and Motility of Lung Endothelial Cells
Role of c-Met/PI3k/Akt Signaling in HGF-mediated Lamellipodia Formation, ROS
published online March 14, 2014J. Biol. Chem.
10.1074/jbc.M113.527556Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/early/2014/03/14/jbc.M113.527556.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on February 18, 2016http://w
ww
.jbc.org/D
ownloaded from
Copyright © 2022 FDOKUMEN




















