
Osteogenesis of Mesenchymal Stem Cells by Nanoscale Mechanotransduction
10
NIKUKAR ET AL. VOL. 7 ' NO. 3 ' 27582767 ' 2013 www.acsnano.org 2758 February 27, 2013 C 2013 American Chemical Society Osteogenesis of Mesenchymal Stem Cells by Nanoscale Mechanotransduction Habib Nikukar, , Stuart Reid, , * P . Monica Tsimbouri, Mathis O. Riehle, Adam S. G. Curtis, and Matthew J. Dalby , * Centrefor Cell Engineering, Institutefor Molecular, Cell and SystemsB iology, Collegeof Medical, Veterinary and LifeSciences, University of G lasgow, G lasgow G128QQ , United K ingdom, SUPA, Thin F ilmCentre, University of theWest of Scotland, Paisley PA1 2BE , United K ingdom, and Shahid Sadoughi University of Medical Sciences, Yazd, I. R . Iran I t has been well-established that many cell types can respond to a variety of nanoscale cues, most notably with nano- topographies and nanoparticles. 1 8 Cell responses include signicant changes in adhesion,migration,morphology,and gene expression. More recently, research has demonstrated that nanotopography alone can stimulate targeted osteogenesis 9,10 and self-renewal 11 in mesenchymal stem cells (MSCs). MSCs are multipotent cells that can be isolated from the bone marrow 12 (as used in this study) or other tissues such as dental tissue, 13 adipose tissue, 14 or umbilical cord. 15 They have the potential to form cells of the bone, cartilage, adipose, and reticular tissues and are sometimes also implicated in smooth muscle 16 and even neural cell lineages. 17 Use of nanotopographieshasshown that the shift from stem cell to dierentiated osteoblast is regulated through changes in adhesion and intracellular tension. 18 This result is consistent with the fact that osteo- blasts, which possess larger cell morphol- ogy than, for example, adipocytes, bro- blasts, and MSCs, require large adhesions to support the tensile cytoskeletal scaolding. 18,19 Such eects have also been observed in MSCs cultured on hydrogels 20 or forced to conne to morphologies using microcontact printing of bronectin. 21 23 By modifying the gel rigidity, soft gels were observed to promote expression of neural markers as cells lost the ability to generate intracellular tension through deformation of the compliant gel substrate. Conversely on the harder gels, with similar stiness to precalcied bone at 40 kPa, the cells were able to retain intercellular tension asa result of large adhesion complexes and thus dif- ferentiate to osteoblasts. 20 An interesting study on MSC conne- ment used an array of small (1000 m 2 ) bronectin microcontact printed squares * Address correspondence to [email protected], [email protected]. Received for review January 14, 2013 and accepted February 27, 2013. Published online 10.1021/nn400202j ABSTRACT It is likely that mesenchymal stem cells will nd use in many autologous regenerative therapies. However, our ability to control cell stem growth and dierentiation ispresently limited, and thisisa major hurdle to the clinical useof thesemultipotent cellsespeciallywhenconsideringthedesirenot to use soluble factors or complex media formulations in culture. Also, the large number of cells required to be clinically useful is currently a hurdle to using materials-based (stiness, chemistry, nanotopography, etc.) culture substrates. Here we give a rst demonstration of using nanoscale sinusoidal mechanotransductive protocols(10 14 nmdisplacements at 1 kHz frequency), nanokicking , to promote osteoblastogenesisin human mesenchymal stemcell cultures. On thebasisof application of thereversepiezoeect, weuseinterferometry todevelop theoptimal stemcell stimulation conditions, allowingdeliveryof nanoscalecuesacrosstheentiresurfaceof thePetri dishesused. Acombinationof immunouorescence, PCR , andmicroarrayhasthen beenusedtodemonstrateosteoblastogenesis, andthearraysimplicateRhoAascentral toosteoblasticdierentiation inagreement withmaterials-based strategies. Wevalidatethiswithpharmacological inhibitionof RhoAkinase. It iseasytoenvisagesuchstimulationprotocolsbeingup-scaledtoformlarge- scale osteoblast bioreactorsasstandard cell culture platesand incubatorsare used in the protocol. KEYWORDS: nanoscale . mechanotransduction . osteogenesis . mesenchymal stem cells ARTICLE
Transcript of Osteogenesis of Mesenchymal Stem Cells by Nanoscale Mechanotransduction

NIKUKARETAL. VOL. 7 ' NO. 3 ' 2758�2767 ' 2013
www.acsnano.org2758
February 27, 2013
C 2013 American Chemical Society
Osteogenesis of MesenchymalStem Cells by NanoscaleMechanotransductionHabib Nikukar,�,� Stuart Reid,�,* P. Monica Tsimbouri,� Mathis O. Riehle,� Adam S. G. Curt is,� andMatthew J. Dalby�,*
�CentreforCell Engineering, InstituteforMolecular, Cell andSystemsBiology, Collegeof Medical, VeterinaryandLifeSciences, Universityof Glasgow,GlasgowG128QQ,UnitedKingdom, �SUPA, ThinFilmCentre, Universityof theWest of Scotland, PaisleyPA12BE,UnitedKingdom, and�ShahidSadoughi Universityof Medical Sciences, Yazd, I. R. Iran
It has been well-established that manycell types can respond to a variety ofnanoscalecues,most notablywith nano-
topographies and nanoparticles.1� 8 Cellresponses include signi�cant changes inadhesion,migration,morphology,and geneexpression. More recently, research hasdemonstrated that nanotopography alonecan stimulate targeted osteogenesis9,10 andself-renewal11 in mesenchymal stem cells(MSCs).MSCs are multipotent cells that can be
isolated from thebonemarrow12 (asused inthis study) or other tissues such as dentaltissue,13 adipose tissue,14 or umbilicalcord.15 Theyhave thepotential to form cellsof thebone,cartilage,adipose,and reticulartissues and are sometimes also implicatedin smooth muscle16 and even neural celllineages.17Useof nanotopographieshasshown that
the shift from stem cell to di�erentiatedosteoblast is regulated through changes in
adhesion and intracellular tension.18 Thisresult is consistent with the fact that osteo-blasts, which possess larger cell morphol-ogy than, for example, adipocytes, �bro-blasts, and MSCs, require large adhesionsto support the tensile cytoskeletalsca�olding.18,19 Such e�ectshavealso beenobserved in MSCs cultured on hydrogels20
or forced to con�ne to morphologiesusingmicrocontact printing of �bronectin.21� 23
By modifying the gel rigidity, soft gelswereobserved to promote expression of neuralmarkers as cells lost the ability to generateintracellular tension through deformationof the compliant gel substrate. Converselyon the harder gels, with similar sti�ness toprecalci�ed bone at 40 kPa, the cells wereable to retain intercellular tension asaresultof large adhesion complexes and thus dif-ferentiate to osteoblasts.20An interesting study on MSC con�ne-
ment used an array of small (1000 �m2)�bronectin microcontact printed squares
* Address correspondence [email protected],[email protected].
Received for review January 14, 2013and accepted February 27, 2013.
Published online10.1021/nn400202j
ABSTRACT It is likely that mesenchymal stem cells will �nd use in manyautologous regenerative therapies. However, our ability to control cell stemgrowthanddi�erentiation ispresently limited, and thisisamajor hurdleto theclinical useof thesemultipotent cellsespeciallywhenconsideringthedesirenot touse soluble factors or complex media formulations in culture. Also, the largenumber of cells required to be clinically useful is currently a hurdle to usingmaterials-based (sti�ness, chemistry, nanotopography, etc.) culture substrates. Here we give a �rst demonstration of using nanoscale sinusoidalmechanotransductiveprotocols(10� 14nmdisplacementsat 1kHzfrequency), �nanokicking�, topromoteosteoblastogenesisinhumanmesenchymalstemcell cultures.Onthebasisof applicationof thereversepiezoe�ect,weuseinterferometrytodeveloptheoptimal stemcell stimulationconditions,allowingdeliveryof nanoscalecuesacrosstheentiresurfaceof thePetri dishesused.Acombinationof immuno�uorescence,PCR,andmicroarrayhasthenbeenusedtodemonstrateosteoblastogenesis, andthearraysimplicateRhoAascentral toosteoblasticdi�erentiationinagreement withmaterials-basedstrategies.Wevalidatethiswithpharmacological inhibitionofRhoAkinase. It iseasytoenvisagesuchstimulationprotocolsbeingup-scaledtoformlarge-scaleosteoblast bioreactorsasstandardcell cultureplatesand incubatorsareused in theprotocol.
KEYWORDS: nanoscale . mechanotransduction . osteogenesis . mesenchymal stem cells
ARTICLE

NIKUKARETAL. VOL. 7 ' NO. 3 ' 2758�2767 ' 2013www.acsnano.org
2759
(among a low adhesion, PEGbackground), preventingspreading of cells, therefore, inhibiting intracellulartension and ultimately leading to the formation ofadipose (fat) tissue. However, when spreading wasactively promoted on larger, 10000 �m2, �bronectinsquares, cells formed large adhesion complexeswith awell-organized cytoskeleton, resulting in di�erentia-tion to osteoblasts.21 An elegant update on this studyused printed �bronectin star and �ower shapes todemonstrate that even when MSCs are dimensionallycon�ned, features that promote adhesion (such asthe sharp corners of the star shapes) promote intra-cellular tension and thus osteogenesis. Conversely,features that reduce adhesion (such as the softercontours of the �ower pattern) signi�cantly reduceintracellular tension and thereforeprevent osteoblasticdi�erentiation.23The commonality of all of the above studies is
investigation of the RhoA kinase pathway (ROCK) as akeymodulator of osteogenesis.Thisresult isexplicablesince RhoA is a small G-protein involved in the activa-tion of actin/myosin contraction. Cells require thiscytoskeletal contraction mechanism in order to spreadand to migrate. The intracellular actin micro�lamentsare located onto focal adhesions on the cell mem-brane, against which (with myosin) they pull. As dis-cussed, osteoblasts are a large progeny of MSCs andare typically very well spread, thusahighly contractedcytoskeleton is required to support this phenotype,and this generates high intracellular tension. In addi-tion, focal adhesionsand thecytoskeleton network arein a force� balance relationship such that a highlycontractile cytoskeleton requires large focal adhesionsand large focal adhesions require force to gatherintegrins and hence to grow.24� 28 It is notable thatosteoblastshave ahigh proportion of supermature, or�brilar, adhesions (adhesions>5 �m in length).29,30It appears that, as cells form adhesions, the plasma
membrane creates nanoscale vibrations, which helpdeterminehow thecell interactswith thesurface.24 It isthe aim of this study to see if we can feedback thesevibrations to the cells to target MSC di�erentiation.That is, if we settle the cells on surfaces that vibrateon the nanoscale, does this a�ect MSCadhesion andultimately phenotype?At themacro- and microscale,awidevariety of cells,
including MSCs,havebeen shown to respond to stressand strain.31� 34 In fact, mechanical stimulation is acommonstrategy in tissueengineeringof cartilageandligament, for example.35� 38Thisisbecausemechanicalalterations in cell morphology lead to changes in cellsignaling and gene transcription. Indeed, not onlymechanical load patterns but also electrical and elec-tromagnetic stimulation patternshave been shown tobe e� cacious in neural and bone regeneration.39� 41Focusing on focal adhesionsasamechanoreceptor,
but being mindful there are others, alterations in
adhesion formation lead to changes in cell tensionandG-protein signaling.However, thismayalso impactfocal adhesion kinase (FAK) and extracellular signal-related kinase (ERK) signaling cascades,32,42,43 whichare intimately linked to activation of transcriptionfactors, such as runt-related transcription factor 2(RUNX2), responsible for transcription and thusexpres-sion of bone-related genessuch asosteocalcin.44� 46In this report, the reverse piezo e�ect is used to
stimulate MSCs. A potential is applied to a piezoceramic actuator causing expansion. The actuator canbedriven so that both the frequency and amplitudeofthe displacement can be accurately and reproduciblycontrolled to deliver nanoscale �kicks�to MSCs.
RESULTSExperimental Setup. Through use of laser interfero-
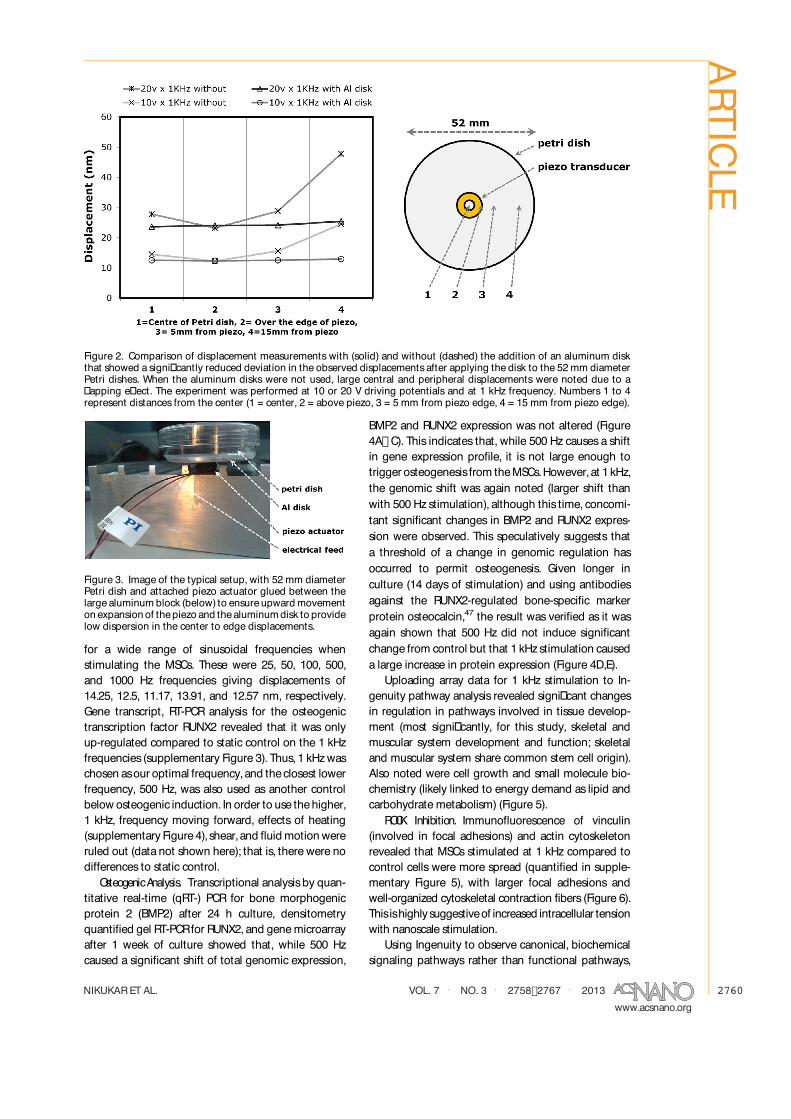
metry, it was demonstrated that reproducible nano-scaleexcursionscould beproduced at different drivingfrequencies, up to 1 kHz. These were in the range of5� 30 nm depending on applied potential (Figure 1).An aluminum block,of approximatedimensions320�100 � 50mm,wasused to provide a stable, rigid basein order to direct the majority of the displacementupward to thePetri dish.In addition,an aluminumdisk,of 50 mm diameter and 3 mm thickness, was rigidlyglued between the piezo ceramic and the polystyrenetissue culture Petri dish (a 60 mm diameter dish, butthe usable bottom surface is 52 mm in diameter) tominimize thedeviation in thedisplacementsthat wereobserved acrossthesurfaceof thedish (Figures2and 3).Displacements were typically observed to reduce asthe driving frequency increased. For the stimulationfrequencies that will be used for promoting osteogen-esis here, the average decrease in the displacementwas observed to be 5.3% and therefore assumed tobe negligible. Further schematics of the experimentalsetup are provided in the Supporting InformationFigures1 and 2.
Selectionof Frequency. By continuoussinewavedrive,10 Vwaschosen as the peak-to-peak driving potential
Figure 1. Displacement (nm) as a function of electricalpotential (V) at di�erent frequencies measured at the sur-face of the piezo actuator by interferometry.
ARTICLE
NIKUKARETAL. VOL. 7 ' NO. 3 ' 2758�2767 ' 2013www.acsnano.org
2760
for a wide range of sinusoidal frequencies whenstimulating the MSCs. These were 25, 50, 100, 500,and 1000 Hz frequencies giving displacements of14.25, 12.5, 11.17, 13.91, and 12.57 nm, respectively.Gene transcript, RT-PCR analysis for the osteogenictranscription factor RUNX2 revealed that it was onlyup-regulated compared to static control on the 1 kHzfrequencies (supplementary Figure 3). Thus, 1 kHzwaschosen asour optimal frequency,and theclosest lowerfrequency, 500 Hz, was also used as another controlbelow osteogenic induction. In order to use thehigher,1 kHz, frequency moving forward, effects of heating(supplementary Figure4),shear,and fluid motion wereruled out (data not shown here); that is, there were nodifferences to static control.
OsteogenicAnalysis. Transcriptional analysisby quan-titative real-time (qRT-) PCR for bone morphogenicprotein 2 (BMP2) after 24 h culture, densitometryquantified gel RT-PCRfor RUNX2,and genemicroarrayafter 1 week of culture showed that, while 500 Hzcaused a significant shift of total genomic expression,
BMP2 and RUNX2 expression was not altered (Figure4A�C). This indicates that, while 500 Hz causesa shiftin gene expression profile, it is not large enough totrigger osteogenesisfrom theMSCs.However,at 1kHz,the genomic shift was again noted (larger shift thanwith 500Hzstimulation), although this time, concomi-tant significant changes in BMP2 and RUNX2 expres-sion were observed. This speculatively suggests thata threshold of a change in genomic regulation hasoccurred to permit osteogenesis. Given longer inculture (14 days of stimulation) and using antibodiesagainst the RUNX2-regulated bone-specific markerprotein osteocalcin,47 the result was verified as it wasagain shown that 500 Hz did not induce significantchange from control but that 1 kHzstimulation causeda large increase in protein expression (Figure 4D,E).
Uploading array data for 1 kHz stimulation to In-genuity pathway analysis revealed signi�cant changesin regulation in pathways involved in tissue develop-ment (most signi�cantly, for this study, skeletal andmuscular system development and function; skeletaland muscular system share common stem cell origin).Also noted were cell growth and small molecule bio-chemistry (likely linked to energy demand as lipid andcarbohydratemetabolism) (Figure 5).
ROCK Inhibition. Immunofluorescence of vinculin(involved in focal adhesions) and actin cytoskeletonrevealed that MSCs stimulated at 1 kHz compared tocontrol cells were more spread (quantified in supple-mentary Figure 5), with larger focal adhesions andwell-organized cytoskeletal contraction fibers (Figure 6).Thisishighlysuggestiveof increased intracellular tensionwith nanoscale stimulation.
Using Ingenuity to observe canonical, biochemicalsignaling pathways rather than functional pathways,
Figure 2. Comparison of displacement measurements with (solid) and without (dashed) the addition of an aluminum diskthat showed a signi�cantly reduced deviation in the observed displacementsafter applying the disk to the 52mm diameterPetri dishes. When the aluminum disks were not used, large central and peripheral displacements were noted due to a�apping e�ect. The experiment was performed at 10 or 20 V driving potentials and at 1 kHz frequency. Numbers 1 to 4represent distances from the center (1 = center, 2 = above piezo, 3 = 5 mm from piezo edge, 4 = 15 mm from piezo edge).
Figure 3. Image of the typical setup, with 52 mm diameterPetri dish and attached piezo actuator glued between thelargealuminum block (below) to ensureupward movementon expansion of thepiezo and thealuminumdisk to providelow dispersion in the center to edge displacements.
ARTICLE

NIKUKARETAL. VOL. 7 ' NO. 3 ' 2758�2767 ' 2013www.acsnano.org
2761
it was seen that ROCK signaling was highlighted assigni�cantly changed with stimulation (Figure 7).Furthermore, network analysis implicated roles foradhesion, FAK, ERK, actin, and sonic hedgehog signal-ing as well as RhoA signaling (see canonical pathway(CP) tags linked into the network) (Figure 8). It isnoteworthy that this network illustrates the input ofthesebiochemical pathwaysinto genescontrolling cell
growth, proliferation, and division. In the classicalStein and Lian48 osteogenic timelines, day 7 is whenmaximal proliferation is seen in cells undergoing os-teogenesiscompared to control cultures; thearraywasperformed at day 7, and it is thus sensible that theseregulatory geneswere seen to be up-regulated.
That RhoA signaling is implicated (Figures7 and 8)isclearly interesting asit hasbeen highlighted asbeing
Figure 4. Transcriptional analysis: (A)Microarray principle component analysisshowing large shiftsof gene regulation fromMSCson control after 1 week of culture. The changeswere greatest (from control) for MSCs stimulated at 1 kHz. The graphshows that the main component (PC1) accounts for 25.7% of di�erences and the secondary component (PC2) accounts for17.5%of di�erences. (B) qRT-PCRof BMP2expression showing increased expression in response to 1 kHzstimulation but not500Hzafter 24h of culture. (C)Densitometry fromRT-PCRgelsshowing no change in RUNX2expression between control and500 Hz stimulation, but up-regulation after 1 kHz stimulation for 1 week. Looking at protein expression of the osteoblastmarker,osteocalcin,after 14daysshowed increased expression in responseto 1kHzstimulation (F)compared to control (D)of500 Hz (E) (green = osteocalcin, blue = nucleus). For PCR, n = 3 (one experiment with three materials replicates for qPCR,and another experiment with three materials replicates for RT-PCR); results are mean ( standard deviation, *p < 0.05 byANOVA, � = individual data points used to calculate mean and standard deviation; arrow shows control mean.
Figure 5. Functional pathway analysis. Among the highlighted pathways di�erentially regulated after 1 week with 1 kHzstimulation are cell growth, organ, and tissue development and skeletal and muscular system development and function.N = 3, threshold = p < 0.05 compared to unstimulated control. * Denotes skeletal system development.
ARTICLE

NIKUKARETAL. VOL. 7 ' NO. 3 ' 2758�2767 ' 2013www.acsnano.org
2762
central to osteogenesis in seminal studies.21,23 UsingY27632 to inhibit ROCKactivity, it wasseen that MSCs�ability to expressosteocalcin was reduced with inhibi-tion, con�rming its role in our nanostimulation system(supplementary Figure 6).
DISCUSSIONFor the�rst time,it isshown that MSCsareresponsive
to vertical nanoscale mechanical excursions. Further-more, it was demonstrated that this osteoblastogenicresponse was ROCK-dependent, in agreement withprevious materials-based strategies for osteoinduc-tion.11,20� 23 That ROCKinhibition blocked osteoblasto-genesis demonstrates that the stimulation is a reale�ect. Furthermore, that BMP2 is activated by 24 hand RUNX2 by 7 days is logical, as RUNX2 activity
is mediated by BMP2 signaling,49,50 this would befollowed by increased osteocalcin expression by day14 according to the Stein and Lian osteoblastogenesistimelines.38 Ingenuity canonical analysisalso highlightspurine and nicotinamide metabolism (involved in en-ergy demand) and sonic hedgehog signaling (involvedis stem cell di�erentiation)51 as well as adhesion, FAK,ERK, and actin-mediated biochemical signaling (allknown tobeimplicated inRUNX2activation)19,29,45,46,52being central to nanomechanotransduction.53,54Useof an open hypothesisfor input frequency led to
use of 1 kHz. This appears high for the time scale ofmany cell actions.Pierreset al.55 providean interestingperspective article on cell membrane undulationsbased on their original data31 and the literature. They�rst note that many familiar, conventional membrane
Figure 6. Morphology, focal adhesion, and actin cytoskeleton staining. (A) On unstimulated control, cellswere smaller withactin stress �bers terminating at focal adhesion sites. (B) MSCs were seen to have a standard morphology. (C) With 1 kHzstimulation, cellswere larger,morepolygonal,and tended to haveboth larger andmoreadhesionswith well-organized actinstress �bers and long cellular processes. (D) Electron micrograph of a typical MSC stimulated at 1 kHz with long cellularprojections. For �uorescent images red = actin, green = vinculin, and blue = nucleus.
Figure 7. Canonical pathway analysis. A signi�cantly altered canonical pathway is RhoA signaling (asterisk). Alsosigni�cantly altered from unstimulated control are sonic hedgehog signaling, purine and nicotinamide metabolism. N = 3,threshold = p < 0.05.
ARTICLE

NIKUKARETAL. VOL. 7 ' NO. 3 ' 2758�2767 ' 2013www.acsnano.org
2763
deformations occur at the tens of seconds time scale,such as �lopodial probing (�lopodia are actin-drivenprojections used to �probe�the extracellular environ-ment for adhesion points).However, they next note that many cell membrane
interactions occur much faster than this. For example,neutrophils completely ingest pathogens withinseconds56 and endothelial arrest of �owing leukocytesinvolves subsecond integrin activation.57 The authorspoint out that subsecond membrane undulationsmaybe key to these phenomena. Interestingly, erythrocytemembranes display high-frequency undulations ashigh as 1 kHz in the tens of nanometers range called�ickering58,59; this is very similar scale to the stimula-tion we supply the cells with in this report. However,it is noted that such movements have been noted asbeing slower in nucleated cells such as lymphocytesand monocytes from blood displaying 20� 30 nm
undulations at up to 30 Hz and �broblasts(descendants of MSCs) displaying 1�4 nm displace-ments at up to 0.5 Hz.60� 62Over a number of years, there has been discussion
of piezoelectic e�ect in bone. This is because, whilecollagen ispiezoelectricwhen dry, for e�ect to beseenin hydrated bone, kilohertz range stimulus is required,that is, beyond that considered physiological.63Furthermore, it is known that electricity can be usedto guide cells (galvanotaxis)64,65 and stimulate bonerepair.66 It has been postulated that bones�piezoelec-trical propertiesfacilitateosteoblast activity asthey areattracted by the electrical dipoles produced by piezo-electricity or deformation of thebone,67 and modelinghas been used to show that piezo e�ects could beimportant in bone homeostasiswith mechanical loadsleading to polarization of bone, suggesting that nega-tive charges accumulate at sites of bone removal and
Figure 8. Network analysis showing involvement of focal adhesion, FAK, ERK, actin, sonic hedgehog, and RhoA signaling inresponse to 1 kHznanomechanostimulation.N= 3.Note that CP= canonical pathway and the lines into the network indicatewhere thesebiochemical pathwaysare involved in thenetwork.Thenetwork illustratesgene interactionsinvolved in growth,proliferation,and division (solid line= direct interaction,dashed lines= indirect interaction).Genesarecompartmentalized asexternal, membrane-bound, cytoplasmic, and nuclear (green = down-regulation, red = up-regulation, white = not tested(below quality threshold or not on the array)).
ARTICLE

NIKUKARETAL. VOL. 7 ' NO. 3 ' 2758�2767 ' 2013www.acsnano.org
2764
positivechargesat sitesof bone formation.68However,we note that while it is tempting to try and drawparallels between the valuesobtained for piezoelectice�ect in bone, thebene�t to boneof electrical stimula-tion, and the 1 kHz osteogenic stimulation we haveused here,it isfar toospeculative,at thisstage,to doso.In addition to describing the details of the applied
motion in terms of frequency and amplitude, it mayalso beof interest to consider the forcesbeing appliedto each cell. During the MSC stimulation presentedhere, a periodic force is imposed on each cell throughthePetri dish substrate.Since the cellsare adherent tothe substrate surface prior to stimulation, this forceis transmitted through each cell as it supports thecolumn of aqueous solution directly above. Takingapproximations for the cell shape (adhered surfacearea typically a few tens of micrometer � a few tensof micrometers) and height of solution (3f 5mm), theaccelerative force on each cell can be calculated to bein the range of 1f 10 pN. This can be calculated fromNewton�ssecond law,Facc=ma,wheretheaccelerativeforce, Facc, driving the motion of the column of waterwith acceleration, a, and with mass, m,must be trans-mitted through the cell to the oscillating substratesurface. Note that these forces are of a similar orderof magnitude to the forces exerted by �broblast
undulations, observed in the range of 20f 80 pN.61Further investigations are required to evaluate theprecise mechanisms responsible for how MSCs sensethese applied mechanical forces.
CONCLUSIONWe introduce a new nanoscale method of MSC
stimulation for targeted osteoblastogenesis. Thisdoesnot rely on novel materials, complex chemistry, orelectronic clean room facilities. Rather, it is based ontraditional cell culture plasticswith simple addition ofpiezo ceramics.Up-scale to bioreactors that can primeautologousMSCsto form osteoblastswithout recourseto soluble factors can be easily envisaged. Further-more, we note that whole body vibration isbecomingadopted in the clinic to help with musculoskeletalregeneration following, for example, spinal trauma,osteoporosis, and stress fracture repair.69,70 It couldthus be envisaged that in vitro experiments could beused to inform therapy with the noted caveat that themodeling from cell to whole body is nontrivial andmuch research aimed at practical/theoretical scalingbetween nanoscale cell cultureand thehuman body isrequired.Such techniquescould becomplementary toexisting external stimuli for musculoskeletal regenera-tion such asextracorporeal shock wave treatment.71
MATERIALS ANDMETHODSSetup. The piezo actuators were driven by continuous sine
wave using an Agilent 33210A function generator in combina-tion with a DC offset circuit for maintaining positive-onlyvoltages applied with the same polarity as the poling voltageused in the piezo manufacture. The piezo actuators used inall of the reported investigations were supplied by PhysikInstrumente GmbH (Germany), model number 010-05H ring-typepiezo stacks.Thisexperimental setup,asshown in Figure3and supplementary Figures 1 and 2, was used for supplyingvertical displacements to MSCcultures.
Calibration. The calibration of individual piezo devicesand the displacement measurements across the surface of thePetri disheswereperformed using aSIOSinterferometer,modelSP-S120 laser interferometric vibrometer. The response of theindividual piezo deviceswasobserved to be linear with drivingvoltage, with deviations typically less than a few percent.The response was also observed to be linear with respect tofrequency, with the displacement being observed to decreaseby typically 10� 15% from 50 Hz to 1 kHz. The measureddisplacements across empty Petri dishes were observed totypically vary by 10� 15%, around a 3-fold decrease in thevariability ascompared to Petri dishes that were not supportedon their underside by a 3 mm thick aluminum disk.
Thermal Check. Since the piezo actuators have to do workagainst the Petri dish assembly, it can be assumed that heat isgenerated. If the power dissipated within the piezo ceramicmaterial transfers through conduction to the aluminum disk,the polystyrene Petri dish, and ultimately to the culture solution,then there exists the possibility of thermal shock to the cells.To evaluate the possibility of thermal shock to the cell cultures,a thermal imaging was used to estimate the degree of heatingduring stimulation.Asshown in supplementaryFigure4,theeffectof heatingcanbeassumed tobenegligiblein theseinvestigations.
ViscousandShear Forces. Aspreviously detailed in theCalibra-tion section,carewasundertaken to ensure that thestimulation
setup acted as a rigid body, transmitting equal displacementsacross the surface of the culture substrate. Therefore, displace-ments are expected to be closely confined to the verticaldirection and thusminimize the possibility of fluid flow, whichcould promote largeviscousforceson thecells.Shear forcesarealso expected to remain negligible.
Cell Culture. MSCswerepurchased fromPromocell (Germany)andusedat passages1� 3.MSCsweremaintained inbasalmedia(aMEM (PAA)) supplemented with 10% FBS (PAA), 1% (v/v)200 mM L-glutamine (Gibco), and antibiotics (6.74 U/mL peni-cillin-streptomycin,0.2�g/mLfungizone) (PAA)at 37�Cwith 5%CO2 in an incubator. Cells were seeded into the Petri dishesat 1� 104 cells/mL,and mediumwaschanged every 3 days.Forinhibition studies,Y27632 (Sigma),aROCKinhibitor,wasused at10 �M concentration throughout each experiment.
Immunofluorescence. MSCswere fixed in a10%formaldehydesolution, permeabilized and blocked in 1% (w/v) BSA/PBS, andstained with anti-osteocalcin and anti-RUNX2 (1:50) (SantaCruzBiotechnology) or anti-vinculin (1:200) (Clone hVin-1, Sigma) in1%(w/v)BSA/PBSand incubated at 37�Cfor 1h, in conjunctionwith phalloidin-rhodamine (1:200) (Molecular Probes). Cellswere washed 3 � 5 min in 1� PBS/0.5% Tween-20 and a bio-tinylated secondary anti-mouse antibody (Vector Laboratories)wasadded at 1:50 in 1%(w/v) BSA/PBSand incubated at 37 �Cfor 1 h. After being washed, the samples were incubated withFITC-conjugated streptavidin (1:50, Vector Laboratories) for30 min at 4 �C, washed, and mounted using Vectashieldmountant with DAPI nuclear stain (Vector Laboratories).
Scanning Electron Microscopy. MSCs were cultured and stimu-lated for 7daysprior to fixing.After 1h incubation of samplesat4 �C in a mixture of 1.5% glutaraldehyde � 0.1 M sodiumcacodylate (buffer fix), the disheswere rinsed by 0.1 M sodiumcacodylate 3 � 5 min. Then 1% osmium tetroxide was addedfor 1 h in room temperature, followed by distilled water (DW)wash (3 � 10 min each). Uranyl acetate was then added for1 h incubation in the dark, followed by a quick DW rinse and
ARTICLE

NIKUKARETAL. VOL. 7 ' NO. 3 ' 2758�2767 ' 2013www.acsnano.org
2765
ethanol dehydration series (30, 50, 70, 90, 100% followed by adried absoluteethanol wash).After being washed two timesfor5min each by hexamethyl disilazane (HMDS), thesampleswereleft overnight for gradual evaporation prior to coating. UsingaPolaron SC515 SEM coating system, the sampleswere coatedby gold palladium (about 18 nm coverage thickness) and thenthey were cut into circles (15 mm diameter) and stuck ontoaluminum stubs.ACarl ZeissSigmavariable-pressure analyticalSEMwith Oxford Microanalysiswasused to image the samples.
RT-PCR(Agarose Gel Electrophoresis). MSCs were cultured andtreated in thePetri dishesfor 7 dayswith three replicasfor eachcondition. Total RNA was extracted using a Qiagen RNeasymicro kit. Reverse transcription of extracted RNA to cDNAwas performed using the Omniscript reverse transcription kit(Qiagen) according to themanufacturer�sprotocol.For efficientcDNAamplification, theGoTaq Hot Start Green Master Mix wasused in combination with gene-specific primers (Table 1) and athermal cycler program for 30 cycles(2min 95�Chold followedby cycles30 sin 95,55,and 72�Ceach and 5min in 72�Casthefinal stage). Amplified cDNAs from three replicas in a controlgroup and stimulated groups were run on a 1% agarose gelbeside a 100 bp ladder, and the expression of RUNX2 wascompared between groupsof samples.
Quantitative Real-Time (qRT-) PCR. Total RNA was extractedfrom three replicas for every condition using a Qiagen RNeasymicro kit. Real-time qPCR was performed and analyzed aspreviously described to assess the expression of RUNX2 andBMP2 (Table 1). RNA samples were reverse transcribed usingtheOmniscript First Strand synthesis kit (Qiagen). qRT-PCRwascarried out using the 7500 Real Time PCRsystem from AppliedBiosystems. GAPDH served as the housekeeping gene to nor-malize expression of the tested genes. With the SYBRGreenmethod primer, sequences for the genes were validated bydissociation curve/melt curve analysis. The 2���Ct method wasused for analysis of gene expression. Statistical analysis wascarried out using the Tukey�Kramer multiple comparisonspost-test analysis of variance (ANOVA). The relative transcriptlevelswere expressed as themean ( standard deviation (n = 3for each group) for graph representation.
Microarray. Briefly, the cellswere cultured in the Petri dishes(stimulated and control, three replicates of each) for 7 days.The cells were then lysed, and total RNA was extracted usinga Qiagen RNeasy kit (Qiagen, UK). Gene expression changeswere detected by hybridization of mRNA to Affymetrix HuGene1.0 ST human arrays according to the manufacturer�s instruc-tions. Initial bioinformaticanalysiswasbased on rankproduct,72and a false discovery rate of 20%was used to upload selectedgene changes to the Ingenuity Pathway Analysis (IPA) serverto identify (1) canonical signaling pathways, (2) functionalpathways, and (3) to produce networks. IPA uses pathwaylibraries derived from the scientific literature. Statistics forfunctional analysis were carried out by Fischer�s exact test (asdone automatically by the software).
Con�ict of Interest: The authors declare no competing�nancial interest.
Acknowledgment. We would like to thank the Ministry ofHealth andMedical Education of I.R.Iran for funding thisproject.Wewould like to thank Linn Products Ltd, Prof. Rein Ulijn, Prof.James Hough, Prof. James Faller, and Dr. Fiona Henriquez foruseful discussionsand their interest in thiswork.Wearegratefulfor the additional �nancial support provided by BBSRC, STFC,the University of Glasgow, and the University of the West ofScotland.Wealso thank theRoyal Society of Edinburgh and theScottishGovernment for providing aresearch fellowship for S.R.
We also thank Carol-Anne Smith, Peter Chung, and MargaretMullin for laboratory assistance.
Supporting Information Available: Additional �gures. Thismaterial is available free of charge via the Internet at http://pubs.acs.org.
REFERENCES AND NOTES1. Dalby,M.J.;Yarwood,S.J.;Riehle,M.O.;Johnstone,H.J.H.;
A�rossman,S.;Curtis,A.S.G.IncreasingFibroblast Responseto Materials Using Nanotopography: Morphological andGenetic Measurements of Cell Response to 13-nm-HighPolymer Demixed Islands.Exp.Cell Res. 2002, 276, 1�9.
2. Dalby, M. J.; Riehle, M. O.; Johnstone, H.; A�rossman, S.;Curtis, A. S. G. In Vitro Reaction of Endothelial Cells toPolymer Demixed Nanotopography. Biomaterials 2002,23, 2945�2954.
3. Andersson, A. S.; Backhed, F.; von Euler, A.; Richter-Dahlfors,A.;Sutherland,D.;Kasemo,B.NanoscaleFeaturesIn�uence Epithelial Cell Morphology and Cytokine Pro-duction. Biomaterials2003, 24, 3427�3436.
4. Yim, E. K.; Reano, R. M.; Pang, S.W.; Yee, A. F.; Chen, C. S.;Leong,K.W.Nanopattern-InducedChangesinMorphologyandMotility of SmoothMuscleCells.Biomaterials2005,26,5405�5413.
5. Child,H.W.;Del Pino,P.A.;DeLaFuente,J.M.;Hursthouse,A. S.; Stirling, D.; Mullen, M.; McPhee, G. M.; Nixon, C.;Jayawarna, V.; Berry, C. C. Working Together: The Com-bined Application of a Magnetic Field and Penetratinfor the Delivery of Magnetic Nanoparticles to Cells in 3D.ACSNano 2011, 5, 7910�7919.
6. Conde, J.; Ambrosone, A.; Sanz, V.; Hernandez, Y.;Marchesano, V.; Tian, F.; Child, H.; Berry, C. C.; Ibarra,M. R.; Baptista, P. V.; et al. Design of Multifunctional GoldNanoparticles for In Vitro and In Vivo Gene Silencing. ACSNano 2012, 6, 8316�8324.
7. Yim, E. K.; Pang, S. W.; Leong, K. W. Synthetic Nanostruc-tures Inducing Di�erentiation of Human MesenchymalStem Cells into Neuronal Lineage. Exp. Cell Res. 2007,313, 1820�1829.
8. Yim, E. K.; Darling, E. M.; Kulangara, K.; Guilak, F.; Leong,K. W. Nanotopography-Induced Changes in Focal Adhe-sions, Cytoskeletal Organization, and Mechanical Proper-ties of Human Mesenchymal Stem Cells. Biomaterials2010, 31, 1299�1306.
9. Oh,S.;Brammer,K.S.; Li, Y.S.; Teng,D.;Engler, A. J.;Chien,S.; Jin, S. Stem Cell Fate Dictated Solely by AlteredNanotube Dimension. Proc. Natl. Acad. Sci. U.S.A. 2009,106, 2130�2135.
10. Dalby,M.J.;Gadegaard,N.;Tare,R.;Andar,A.;Riehle,M.O.;Herzyk, P.;Wilkinson, C. D.W.; Ore�o, R.O. C. The Controlof Human Mesenchymal Cell Di�erentiation Using Nano-scale Symmetry and Disorder. Nat. Mater. 2007, 6, 997�1003.
11. McMurray, R. J.; Gadegaard, N.; Tsimbouri, P. M.; Burgess,K. V.; McNamara, L. E.; Tare, R.; Murawski, K.; Kingham, E.;Ore�o,R.O.;Dalby,M. J.Nanoscale Surfaces for the Long-Term Maintenance of Mesenchymal Stem Cell Phenotypeand Multipotency. Nat. Mater. 2011, 10, 637�644.
12. Tare, R. S.; Babister, J. C.; Kanczler, J.; Ore�o, R. O. SkeletalStemCells:Phenotype,Biology and Environmental NichesInformingTissueRegeneration.Mol.Cell.Endocrinol.2008,288, 11�21.
13. Huang,G.T.;Gronthos, S.; Shi, S.Mesenchymal Stem CellsDerived fromDental Tissuesvs.Those fromOther Sources:Their Biology and Role in Regenerative Medicine. J. Dent.Res. 2009, 88, 792�806.
14. Scha� er, A.; Buchler, C. Concise Review: Adipose Tissue-Derived Stromal Cells; Basic and Clinical Implications forNovel Cell-Based Therapies.Stem Cells2007, 25, 818�827.
15. Longo, U. G.; Loppini, M.; Berton, A.; La Verde, L.; Khan,W. S.; Denaro, V. Stem Cells from Umbilical Cord andPlacenta for Musculoskeletal Tissue Engineering. Curr.Stem Cell Res. Ther. 2012, 7, 272�281.
TABLE 1. Primer Details for PCR
gene forwardprimer reverseprimer
RUNX2 CAGACCAGCAGCACTCCATA CAGCGTCAACACCATCATTCBMP2 ATGGATTCGTGGTGGAAGTG GTGGAGTTCAGATGATCAGCGAPDH GTCAGTGGTGGACCTGACCT ACCTGGTGCTCAGTGTAGCC
ARTICLE

NIKUKARETAL. VOL. 7 ' NO. 3 ' 2758�2767 ' 2013www.acsnano.org
2766
16. Vater, C.; Kasten, P.; Stiehler, M. Culture Media forthe Di�erentiation of Mesenchymal Stromal Cells. ActaBiomater. 2011, 7, 463�477.
17. Liu, J.; Song, L.; Jiang, C.; Liu, Y.; George, J.; Ye, H.; Cui, Z.Electrophysiological Properties and Synaptic Function ofMesenchymal Stem Cells During Neurogenic Di�erentia-tion: A Mini-Review. Int. J. Artif. Organs 2012, 35, 323�337.
18. Tsimbouri,P.M.;McMurray,R.J.;Burgess,K.V.;Alakpa,E.V.;Reynolds, P. M.; Murawski, K.; Kingham, E.; Ore�o, R. O.;Gadegaard, N.; Dalby, M. J. Using Nanotopography andMetabolomics To Identify Biochemical E�ectors of Multi-potency. ACSNano 2012, 6, 10239�10249.
19. Biggs, M. J. P.; Richards, R. G.; Gadegaard, N.; Wilkinson,C.D.W.;Ore�o,R.O.C.; Dalby,M. J. TheUse of NanoscaleTopography To Modulate the Dynamics of AdhesionFormation in PrimaryOsteoblastsand Erk/MapkSignallingin Stro-1�Enriched Skeletal StemCells.Biomaterials2009,30, 5094�5103.
20. Engler, A. J.; Sen, S.; Sweeney, H. L.; Discher, D. E. MatrixElasticity Directs Stem Cell Lineage Speci�cation. Cell2006, 126, 677�689.
21. McBeath, R.; Pirone, D. M.; Nelson, C. M.; Bhadriraju, K.;Chen, C. S. Cell Shape, Cytoskeletal Tension, and RhoARegulate Stem Cell Lineage Commitment. Dev. Cell 2004,6, 483�495.
22. Gao,L.;McBeath,R.;Chen,C.S.Stem Cell ShapeRegulatesa Chondrogenic versusMyogenic Fate through Rac1 andN-Cadherin. Stem Cells2010, 28, 564�572.
23. Kilian, K.A.; Bugarija, B.; Lahn,B. T.;Mrksich,M.GeometricCues for Directing the Di�erentiation of MesenchymalStem Cells. Proc. Natl. Acad. Sci. U.S.A. 2010, 107, 4872�4877.
24. Sawada, Y.; Sheetz, M. P. Force Transduction by TritonCytoskeletons. J. Cell Biol. 2002, 156, 609�615.
25. Galbraith, C.G.; Yamada, K.M.; Sheetz,M. P. The Relation-ship between Force and Focal Complex Development.J. Cell Biol. 2002, 159, 695�705.
26. Vogel, V.; Sheetz, M. Local Force and Geometry SensingRegulate Cell Functions. Nat. Rev. Mol. Cell. Biol. 2006, 7,265�275.
27. Sawada,Y.;Tamada,M.;Dubin-Thaler,B.J.;Cherniavskaya,O.; Sakai, R.; Tanaka, S.; Sheetz, M. P. Force Sensing byMechanical Extension of the Src Family Kinase SubstrateP130cas. Cell 2006, 127, 1015�1026.
28. del Rio, A.; Perez-Jimenez, R.; Liu, R.; Roca-Cusachs, P.;Fernandez, J.M.; Sheetz,M. P. Stretching Single Talin RodMolecules Activates Vinculin Binding. Science 2009, 323,638�641.
29. Biggs, M. J. P.; Richards, R. G.; McFarlane, S.; Wilkinson,C.D.W.;Ore�o,R.O.C.;Dalby,M.J.Adhesion Formation ofPrimary Human Osteoblasts and the Functional ResponseofMesenchymal StemCellsto 330nmDeepMicrogrooves.J. R. Soc. Interface2008, 5, 1231�1242.
30. Biggs, M. J. P.; Richards, R. G.; Gadegaard, N.; McMurray,R. J.; A�rossman, S.; Wilkinson, C. D. W.; Ore�o, R. O. C.;Dalby, M. J. Interactions with Nanoscale Topography:Adhesion Quanti�cation and Signal Transduction in Cellsof Osteogenic and Multipotent Lineage. J. Biomed.Mater.Res., Part A 2009, 91A, 195�208.
31. Pierres,A.; Benoliel, A.M.; Touchard,D.; Bongrand,P.HowCellsTiptoeonAdhesiveSurfacesbeforeSticking.Biophys.J. 2008, 94, 4114�4122.
32. Guilak, F.; Cohen, D. M.; Estes, B. T.; Gimble, J. M.; Liedtke,W.; Chen, C. S. Control of Stem Cell Fate by PhysicalInteractions with the Extracellular Matrix. Cell Stem Cell2009, 5, 17�26.
33. Park,S.H.;Sim,W.Y.;Min,B.H.;Yang,S.S.;Khademhosseini,A.;Kaplan,D.L.Chip-BasedComparisonof theOsteogenesisof Human Bone Marrow- and Adipose Tissue-DerivedMesenchymal Stem Cells under Mechanical Stimulation.PLoSONE2012, 7, e46689.
34. Shi, Y.; Fu, Y.; Tong, W.; Geng, Y.; Lui, P. P.; Tang, T.;Zhang, X.; Dai, K. Uniaxial Mechanical Tension PromotedOsteogenic Di�erentiation of Rat Tendon-Derived Stem
Cells(Rtdscs)via theWnt5a-RhoAPathway.J.Cell.Biochem.2012, 113, 3133�3142.
35. Pingguan-Murphy,B.; Lee,D.A.;Bader,D. L.; Knight,M.M.Activation of ChondrocytesCalciumSignalling byDynamicCompression Is Independent of Number of Cycles. Arch.Biochem. Biophys. 2005, 444, 45�51.
36. Knight, M. M.; van de Breevaart Bravenboer, J.; Lee, D. A.;van Osch, G. J.; Weinans, H.; Bader, D. L. Cell and NucleusDeformation in Compressed Chondrocyte-Alginate Con-structs:Temporal ChangesandCalculation ofCellModulus.Biochim. Biophys. Acta 2002, 1570, 1�8.
37. Cheema, U.; Chuo, C.-P.; Sarathchandra, P.; Nazhat, S. N.;Brown, R. A. Engineering Functional Collagen Sca�olds:Cyclical Loading Increases Material Strength and FibrilAggregation. Adv. Funct. Mater. 2007, 17, 2426�2431.
38. Eastwood, M.; McGrouther, D. A.; Brown, R. A. FibroblastResponses to Mechanical Forces. Proc. Inst. Mech. Eng.,Part H 1998, 212, 85�92.
39. Kanaka, T. S.; Kumar, M. M. Neural Stimulation for SpinalSpasticity. Paraplegia 1990, 28, 399�405.
40. Chen, I. I.; Saha, S. Analysis of the Current Distribution inBoneProduced by Pulsed Electro-Magnetic Field Stimula-tion of Bone. Biomater. Artif. Cells Artif. Organs 1987, 15,737�744.
41. Pompeiano, O.; Wand, P.; Sontag, K. H. The Sensitivity ofRenshaw Cells to Velocity of Sinusoidal Stretches of theTricepsSurae Muscle. Arch. Ital. Biol. 1975, 113, 280�294.
42. Ingber, D. E. Tensegrity and Mechanotransduction.J. Bodyw. Mov. Ther. 2008, 12, 198�200.
43. Ward,D.F.,Jr.;Salasznyk,R.M.;Klees,R.F.;Backiel,J.;Agius,P.; Bennett, K.; Boskey, A.; Plopper,G. E.Mechanical StrainEnhances Extracellular Matrix-Induced Gene Focusing andPromotesOsteogenicDi�erentiationof HumanMesenchymalStemCellsthroughanExtracellular-RelatedKinase-DependentPathway.Stem CellsDev. 2007, 16, 467�480.
44. Hamilton, D.W.; Brunette, D. M. The E�ect of SubstratumTopography on Osteoblast Adhesion Mediated SignalTransduction and Phosphorylation. Biomaterials 2007,28, 1806�1819.
45. Ge, C.; Xiao, G.; Jiang, D.; Franceschi, R. T. Critical Role ofthe Extracellular Signal-Regulated Kinase-Mapk Pathwayin Osteoblast Di�erentiation and Skeletal Development.J. Cell Biol. 2007, 176, 709�718.
46. Xiao, G.; Jiang, D.; Gopalakrishnan, R.; Franceschi, R. T.Fibroblast Growth Factor 2 Induction of the OsteocalcinGene Requires Mapk Activity and Phosphorylation ofthe Osteoblast Transcription Factor, Cbfa1/Runx2. J. Biol.Chem. 2002, 277, 36181�36187.
47. Willis,D.M.;Loewy,A.P.;Charlton-Kachigian,N.;Shao,J.-S.;Ornitz,D.M.;Towler,D.A.Regulation of Osteocalcin GeneExpression by a Novel Ku Antigen Transcription FactorComplex. J. Biol. Chem. 2002, 277, 37280�37291.
48. Stein, G. S.; Lian, J. B. Molecular Mechanisms MediatingProliferation/Di�erentiation Interrelationshipsduring Pro-gressive Development of the Osteoblast Phenotype.Endocr. Rev. 1993, 14, 424�442.
49. Huang, Y. F.; Lin, J. J.; Lin, C. H.; Su, Y.; Hung, S. C. C-JunN-Terminal Kinase 1 Negatively Regulates OsteoblasticDi�erentiation Induced by Bmp2 via Phosphorylation ofRunx2 at Ser104. J. BoneMiner. Res. 2012, 27, 1093�1105.
50. Jang,W.G.;Kim,E.J.;Kim,D.K.;Ryoo,H.M.; Lee,K.B.;Kim,S. H.; Choi, H. S.; Koh, J. T. Bmp2 Protein RegulatesOsteocalcin Expression via Runx2-Mediated Atf6 GeneTranscription. J. Biol. Chem. 2012, 287, 905�915.
51. Kim,W.K.;Meliton,V.;Bourquard,N.;Hahn,T.J.;Parhami,F.Hedgehog Signaling and Osteogenic Di�erentiation inMultipotent Bone Marrow Stromal Cells Are Inhibited byOxidative Stress. J. Cell. Biochem. 2010, 111, 1199�1209.
52. Dai,Z.; Li,Y.;Quarles,L.D.;Song,T.;Pan,W.;Zhou,H.;Xiao,Z. Resveratrol Enhances Proliferation and OsteoblasticDi�erentiation in Human Mesenchymal Stem Cells viaER-Dependent ERK1/2 Activation. Phytomedicine 2007,14, 806�814.
53. James, A.W.; Leucht, P.; Levi, B.; Carre, A. L.; Xu, Y.; Helms,J. A.; Longaker, M. T. Sonic Hedgehog In�uences the
ARTICLE

NIKUKARETAL. VOL. 7 ' NO. 3 ' 2758�2767 ' 2013www.acsnano.org
2767
Balance of Osteogenesis and Adipogenesis in MouseAdipose-Derived Stromal Cells. Tissue Eng., Part A 2010,16, 2605�2616.
54. Day,T.F.;Yang,Y.Wnt and Hedgehog Signaling Pathwaysin Bone Development. J. Bone Joint Surg. Am. 2008, 90Suppl 1, 19�24.
55. Pierres, A.; Monnet-Corti, V.; Benoliel, A. M.; Bongrand, P.Do Membrane Undulations Help Cells Probe the World?TrendsCell. Biol. 2009, 19, 428�433.
56. Evans, E. Kinetics of Granulocyte Phagocytosis: RateLimited by Cytoplasmic Viscosity and Constrained by CellSize. Cell Motil. Cytoskeleton 1989, 14, 544�551.
57. Shamri, R.; Grabovsky, V.; Gauguet, J. M.; Feigelson, S.;Manevich, E.; Kolanus,W.; Robinson,M.K.; Staunton,D.E.;von Andrian, U. H.; Alon, R. Lymphocyte Arrest RequiresInstantaneous Induction of an Extended Lfa-1 Conforma-tion Mediated by Endothelium-Bound Chemokines. Nat.Immunol. 2005, 6, 497�506.
58. Rappaz,B.;Barbul,A.;Ho�mann,A.;Boss,D.;Korenstein,R.;Depeursinge, C.; Magistretti, P. J.; Marquet, P. SpatialAnalysisof Erythrocyte Membrane Fluctuationsby DigitalHolographic Microscopy. Blood Cells, Mol., Dis. 2009, 42,228�232.
59. Evans, J.; Gratzer, W.; Mohandas, N.; Parker, K.; Sleep, J.Fluctuations of the Red Blood Cell Membrane: Relationto Mechanical Properties and Lack of ATP Dependence.Biophys. J. 2008, 94, 4134�4144.
60. Krol, A.; Grinfeldt, M. G.; Levin, S. V.; Smilgavichus, A. D.Local Mechanical Oscillations of the Cell Surface withinthe Range 0.2� 30 Hz. Eur. Biophys. J. 1990, 19, 93�99.
61. Pelling, A. E.; Veraitch, F. S.; Pui-Kei Chu, C.; Nicholls, B.M.;Hemsley, A. L.; Mason, C.; Horton, M. A. Mapping Corre-lated Membrane Pulsations and Fluctuations in HumanCells. J. Mol. Recognit. 2007, 20, 467�475.
62. Szabo,B.;Selmeczi,D.;Kornyei,Z.;Madarasz,E.;Rozlosnik,N. Atomic Force Microscopy of Height Fluctuations ofFibroblast Cells. Phys. Rev. E2002, 65, 041910.
63. Reinish,G.B.;Nowick,A.S.PiezoelectricPropertiesof BoneasFunctionsof Moisture Content.Nature1975, 253, 626�627.
64. Finkelstein, E. I.; Chao, P. H.; Hung, C. T.; Bulinski, J. C.Electric Field-Induced Polarization of Charged CellSurface Proteins Does Not Determine the Direction ofGalvanotaxis. Cell Motil. Cytoskeleton 2007, 64, 833�846.
65. Mycielska, M. E.; Djamgoz, M. B. Cellular Mechanisms ofDirect-Current Electric Field E�ects: Galvanotaxis andMetastatic Disease. J. Cell. Sci. 2004, 117, 1631�1639.
66. Goldstein, C.; Sprague, S.; Petrisor, B. A. Electrical Stimula-tion for Fracture Healing: Current Evidence. J. Orthop.Trauma 2010, 24 Suppl 1, S62�S65.
67. Noris-Suarez,K.;Lira-Olivares,J.;Ferreira,A.M.;Feijoo,J.L.;Suarez,N.;Hernandez,M.C.; Barrios, E. In Vitro Depositionof HydroxyapatiteonCortical BoneCollagenStimulated byDeformation-Induced Piezoelectricity. Biomacromolecules2007, 8, 941�948.
68. Fernandez, J. R.; Garcia-Aznar, J. M.; Martinez, R. Piezo-electricity Could Predict Sites of Formation/Resorption inBoneRemodelling andModelling.J.Theor.Biol.2012,292,86�92.
69. Wysocki, A.; Butler, M.; Shamliyan, T.; Kane, R. L. Whole-Body Vibration Therapy for Osteoporosis: State of theScience. Ann. Intern. Med. 2011, 155, 680�686W206-613.
70. Alizadeh-Meghrazi, M.; Masani, K.; Popovic, M. R.; Craven,B. C. Whole-Body Vibration During Passive Standing inIndividualswith Spinal Cord Injury:E�ectsof PlateChoice,Frequency,Amplitude,and Subject�sPosture on VibrationPropagation. PM&R2012, 4, 963�975.
71. Tamma, R.; dell�Endice, S.; Notarnicola, A.; Moretti, L.;Patella,S.;Patella,V.;Zallone,A.;Moretti,B.ExtracorporealShock Waves Stimulate Osteoblast Activities. UltrasoundMed. Biol. 2009, 35, 2093�2100.
72. Breitling, R.; Armengaud,P.; Amtmann,A.; Herzyk, P.RankProducts: A Simple, Yet Powerful, New Method To DetectDi�erentially Regulated Genes in Replicated MicroarrayExperiments. FEBSLett. 2004, 573, 83�92.
ARTICLE




















