
Comparison of Hyperspectral Versus Traditional Field ... - MDPI
Upload
independentCategory
view
3download
0
www.elsevier.com/locate/agrformet
Agricultural and Forest Meteorology 140 (2006) 287–307
Spatial modelling of the fraction of photosynthetically
active radiation absorbed by a boreal mixedwood
forest using a lidar–hyperspectral approach
V. Thomas a,*, D.A. Finch a, J.H. McCaughey a, T. Noland b, L. Rich b, P. Treitz a
a Department of Geography, Faculty of Arts and Science, Queen’s University, Kingston, Ont., Canada K7L 3N6b Ontario Ministry of Natural Resources, Ontario Forest Research Institute, 1235 Queen St. East,
Sault Ste. Marie, Ont., Canada P6A 2E5
Received 16 November 2005; accepted 17 April 2006
Abstract
The spatial variability of the fraction of photosynthetically active radiation absorbed by the canopy (fPAR) was characterized for
a heterogeneous boreal mixedwood forest site located in northern Ontario, Canada, based on relationships found between fPAR and
light detection and ranging (lidar) data over different canopy architectures. Estimates of fPAR were derived from radiation
measurements made above the canopy at a flux tower and below-canopy radiation was measured across a range of species
compositions and canopy architectures. Airborne lidar data were used to characterize spatial variability of canopy structure around
the flux tower and a map of mean canopy chlorophyll concentration was derived from airborne hyperspectral imagery. Different
volumes of lidar points for the locations directly above each photosynthetically active radiation (PAR) sensor were examined to
determine if there is an optimal method of relating lidar returns to estimated fPAR values.
The strongest correlations between mean lidar height and fPAR occurred when using points that fell within a theoretical cone
which originated at the PAR sensor having a solid angle a = 558. For diffuse conditions, the correlation (r) between mean lidar
height versus fPAR � chlorophyll was stronger than between mean lidar height versus fPAR by 8% for mean daily fPAR and from
10 to 20% for diurnal fPAR, depending on solar zenith angle. For direct light conditions, the relationship was improved by 12% for
mean daily fPAR and 12–41% for diurnal relationships.
Linear regression models of mean daily fPAR � chlorophyll versus mean lidar height were used in conjunction with gridded
lidar data and the canopy chlorophyll map to generate maps of mean daily fPAR for direct and diffuse sunlight conditions. Site
average fPAR calculated from these maps was 0.79 for direct light conditions and 0.78 for diffuse conditions. When compared to
point estimates of mean daily fPAR calculated on the tower, the average fPAR was significantly lower than the point estimate.
Subtracting the direct sunlight fPAR map from the diffuse sunlight fPAR map revealed a distinct spatial pattern showing that areas
with open canopies and relatively low chlorophyll (e.g., black spruce patches) have a higher fPAR under direct sunlight conditions,
while closed canopies with higher chlorophyll (e.g., deciduous species) absorb more PAR under diffuse conditions. These findings
have implications for scaling from point measurements at flux towers to larger resolution satellite imagery and addressing local
scale heterogeneity in flux tower footprints.
# 2006 Elsevier B.V. All rights reserved.
Keywords: fPAR; Canopy structure; Spatial variability; Lidar; Hyperspectral; Chlorophyll
* Corresponding author. Tel.: +1 613 533 6000x75101; fax: +1 613 533 6122.
E-mail addresses: [email protected] (V. Thomas), [email protected] (D.A. Finch),
[email protected] (J.H. McCaughey), [email protected] (T. Noland), [email protected] (P. Treitz).
0168-1923/$ – see front matter # 2006 Elsevier B.V. All rights reserved.
doi:10.1016/j.agrformet.2006.04.008

V. Thomas et al. / Agricultural and Forest Meteorology 140 (2006) 287–307288
1. Introduction
To better understand the exchanges of energy, water
and carbon dioxide between the forest and the
atmosphere, a detailed knowledge of the shortwave
radiation regime within the forest canopy is essential.
The architecture of the forest canopy strongly influ-
ences the spatial and temporal variability of shortwave
radiation transfer therein and depends upon tree species,
size and location of canopy gaps (Hardy et al., 2004).
FPAR is a key structural variable in national and global
models of net primary production, climate, hydrology,
biogeochemistry and ecology (e.g., Bonan, 1998;
Dickinson et al., 1986; Running and Coughlan, 1988;
Sellers et al., 1996).
The solar radiation regimes of various forest types
have been studied using sensors located at and near
meteorological towers (e.g., Baldocchi et al., 1984; Eck
and Deering, 1992; Chen et al., 1997; Hardy et al.,
2004). Meteorological measurements of total and
diffuse PAR from sensors located above and total
PAR below the canopy allow for the estimation of fPAR.
However, point estimates of fPAR are not necessarily
representative of forest canopies that exhibit a high
degree of spatial variability, such as boreal mixedwood
stands. In these environments, the spatial arrangement
of tree species and variability in canopy structure and
physiology will have a significant impact on the light
regime and the total capacity for photosynthesis in a
stand.
Remote sensing data can offer valuable perspectives
on the spatial variability of fPAR and photosynthetic
capacity across the landscape. Algorithms have been
developed for the retrieval of fPAR and LAI from
MODIS and other relatively low resolution satellite
platforms (Huemmerich et al., 2005; Knyazikhin et al.,
1998a,b). However, the pixel size of these systems is
very large (i.e., individual cell widths may range from
250 m to 1 km) and therefore do not address the spatial
variability that may exist within the footprint of a flux
tower. Further, in situ validation of coarse and moderate
resolution estimates is difficult, due spatial hetero-
geneity within the cell size of the models. Chen et al.
(2003) describe the traditional remote sensing
approaches to fPAR estimation as crude approximations
and discuss the need for alternative approaches that
consider the variability of clumping and LAI. Varia-
bility of fPAR is also highlighted by Shabanov et al.
(2003), who demonstrate that the spatial variability is at
least as large as the diurnal variability due to solar
zenith angle and stress the need to address ‘sub-pixel’
fPAR heterogeneity in current models.
High resolution hyperspectral and light detection and
ranging (lidar) sensors can provide considerable
information about canopy photosynthetic capacity
and structure, which may improve local estimates of
fPAR over point estimates derived from meteorological
towers or from large cell sizes available from traditional
remote sensing models. Lidar systems have the ability
to penetrate the forest canopy from above. This provides
a measure of canopy shape and structure, as opposed to
an assumption of simple crown shapes (i.e., conical or
spherical) (e.g., Arp et al., 1982; Aldred and Bonner,
1985; Nilsson, 1996; Næsset, 1997; Lefsky et al., 1997;
Magnussen and Boudewyn, 1998; Lefsky et al., 1999;
Thomas et al., 2006). Further, the ability to penetrate
into the canopy provides an opportunity to characterize
light transmittance through the canopy (e.g., Parker
et al., 2001; Todd et al., 2003). This, in turn, provides
information regarding the light available for photo-
synthesis and its variation throughout the depth of the
canopy (i.e., variations in fPAR).
The relationships between canopy photosynthetic
capacity and spectral reflectance have been observed in
the laboratory using spectroradiometers. There has been
some success developing hyperspectral indices that are
directly related to leaf and canopy chlorophyll content,
which has been shown to be related to photosynthetic
capacity (Waring et al., 1995). Very strong relationships
(r2 > 0.85) have been reported when comparing leaf
chlorophyll to leaf spectra measured with a spectro-
radiometer in laboratory or field conditions (Blackburn,
1998; Curran et al., 1990; Zarco-Tejada et al., 2000,
2001). Hyperspectral remote sensing potentially allows
for the expansion of these leaf relationships to the
canopy scale, by providing a more complete spectral
signature than what is possible with traditional remote
sensing platforms (e.g., Landsat or SPOT). Results of
canopy-scale studies of chlorophyll content of forests
and crops generally show moderate to strong correla-
tions of leaf samples to airborne hyperspectral data
(Yoder and Pettigrew-Crosby, 1995; Datt, 1998; Zarco-
Tejada et al., 2001; Blackburn, 2002).
In addition to the amount of PAR available
throughout the canopy and the concentration of
chlorophyll in the leaves, recent publications have
demonstrated that the ability to photosynthesize is also
influenced by the amount of diffuse versus direct PAR
available. Gu et al. (2003) demonstrated an increase in
diffuse PAR observed under cloudless conditions
immediately after the Pinatubo volcanic eruption which
translated into an increase in gross photosynthesis and a
resulting CO2 sink. Authors have also shown an
increase in CO2 uptake under cloudy conditions with

V. Thomas et al. / Agricultural and Forest Meteorology 140 (2006) 287–307 289
diffuse PAR conditions as opposed to clear conditions
(e.g., Gu et al., 2002; Freedman et al., 2001). At a
minimum, models of photosynthesis must separate the
diffuse and direct components of PAR in their
calculations to assess the impacts of varying fractions
of diffuse and direct PAR conditions.
The objective of this research is to integrate lidar and
hyperspectral data, as well as above- and below-canopy
quantum measurements of direct and diffuse PAR and
field measures of leaf chlorophyll, to model the spatial
variability of fPAR for a boreal mixedwood forest
located in northern Ontario, Canada.
2. Background
Although lidar technology and data characteristics
have been well described in the literature, some
discussion of lidar data characteristics is helpful to
contextualize the research presented in this paper.
While traditional multispectral and hyperspectral
sensors are described as passive remote sensing
technology (i.e., they measure the intensity of the sun’s
reflectance off surfaces), lidar sensors are referred to as
active technology (similar in concept to radar and SAR
technologies). Here, high energy laser pulses emitted in
short intervals are directed towards the ground and the
time of pulse return is measured. Because the velocity
of light is a constant (i.e., 3 � 108 m/s), the elapsed time
between emission and receiving of the pulse can be used
to determine the distance between the sensor and object
or the ground, a process known as pulse ranging (Wehr
and Lohr, 1999; Lim et al., 2003b). The lidar system
used in this research is categorized as a discrete ‘‘first-
and last-return’’ system, which records the first and last
energy response above a specified noise threshold for
each laser pulse (this is in contrast to a full waveform
system, which would capture an amplitude versus time
waveform). Using this concept, x, y and z measures can
be obtained in the form of a three-dimensional point
cloud, with horizontal accuracies of 50 cm and vertical
accuracies of 10 cm (Baltsavias, 1999; Wehr and Lohr,
1999; Lim et al., 2003b).
There are some distinct differences between discrete
lidar datasets and the more traditional multispectral or
hyperspectral images that have a significant impact on the
way data are analyzed and the types of information that
can be extracted from the dataset. First, lidar datasets are
not images and the raw data cannot be easily visualized
from the traditional planar perspective. Rather, the first-
and last-return coordinates are most easily viewed as
points in a three-dimensional scatterplot or in a software
package capable of redendering point clouds. For
example, Fig. 1 illustrates the first-return points
contained within a circular white birch plot with an
11.3 m radius (400 m2). Most points are clustered
between 8 and 15 m above ground and are representative
of the forest canopy. A few points have penetrated
through gaps in the canopy and reached the ground (note
that if last returns were also displayed, the majority of
them would be at ground level). The ‘bumps’ along the
top of the scatterplot illustrate different tree crowns
within the plot. If one were to examine the entire lidar
dataset for the 1 km radius around the flux tower it would
look like a very large scatterplot, containing more than 16
million points. For the purpose of this research, the 1 km
radius dataset is referred to the ‘original point cloud’.
Points that fall within the area of the sample plots were
subset from the original point cloud and analyzed to
determine correlations with fPAR.
Another key difference between lidar datasets and
traditional multispectral and hyperspectral remotely
sensed images is that there is no measurement of solar
reflectance and therefore no relationship between a
coordinate point and solar zenith angle. For each lidar
data point, the scan angle from the sensor would have
some effect on how deep into the canopy the lidar pulse
would penetrate, mainly due to the spatial configuration
of leaves and branches at that specific location.
However, because the scan angle rapidly changes
within a 408 field of view and because there are multiple
flight lines over the study site to increase the point
density, it can be assumed that there is no consistent
directional bias to the dataset (i.e., each 11.3 m radius
plot contains over 1000 first returns; the canopy would
have been scanned from many angles).
To date, lidar forestry research has been largely
empirical, focusing on strong relationships between tree
height, biomass, volume and other structural metrics.
Lim et al. (2003a) reported success using lidar to predict
several structural variables, including canopy closure,
LAI, basal area, volume, above ground biomass and
height. The reader is referred to Baltsavias (1999), Wehr
and Lohr (1999), St-Onge et al. (2003) and Lim et al.
(2003b) for more detailed overviews of the technology
and its particular application to forestry.
3. Site description
The experiment was conducted during the growing
season of 2004 at the Groundhog River Flux Station
(GFRS), which is part of the Fluxnet-Canada Research
Network. The GRFS is a boreal mixedwood stand,
located approximately 80 km southwest of Timmins,
Ontario, Canada (Fig. 2). At the site, there is a 41-m-tall

V. Thomas et al. / Agricultural and Forest Meteorology 140 (2006) 287–307290
Fig. 1. Example of a lidar point cloud for an 11.3 m radius white birch plot. Lidar height above ground is presented on the vertical axis, with UTM
eastings and northings on the bottom.
micrometeorological flux tower where eddy covariance
(EC) measurements of turbulent fluxes (e.g., sensible
heat, latent heat, carbon and momentum), in addition to
climate data (e.g., air temperature, humidity, solar and
terrestrial radiation, and soil moisture) have been made
on a continuous basis since August 2003. The area of
the research site extends in a circular arrangement with
a 1 km radius centered on the flux tower.
The GRFS can be described as a heterogeneous
mixture of five primary tree species including:
trembling aspen [Populus tremuloides Michx.] (TA);
white birch [Betula papyrifera Marsh.] (WB); white
spruce [Picea glauca (Moench) Voss] (WS); black
spruce [Picea mariana (Mill.) BSP] (BS); and balsam
fir [Abies balsamea (L.) Mill.] (BF). There are also
distinct patches of northern white cedar [Thuja
occidentalis (L.)] (C) in a few wet locations within
the site. The canopy is characterized as under-mature,
with an average tree age of approximately 75 years.
There are 29 circular large-tree plots (radius = 11.3 m,
area = 400 m2) distributed across the range of species
associations, which were sampled in 2003 for structural
characteristics. Six of these plots were chosen for the
below-canopy PAR experiment, based on their species
composition. These included plots dominated by: (1)
white birch; (2) medium-sized aspen (i.e., tree
height < 25 m); (3) large aspen (i.e., tree height < 35 m)
m) with a balsam fir sub-canopy; (4) black spruce; (5)
cedar; (6) mixedwood (i.e., TA, WB, WS and BF)
(Fig. 2).
4. Methods
4.1. Mensurational data
Mensurational data for the 29 large tree plots
(including the 6 plots where under-canopy PAR was
measured) were collected during the summers of 2003
and 2004 in accordance with Fluxnet-Canada protocols,
which are derived from the Canadian National Forest
Inventory (NFI) protocols (Fluxnet-Canada, 2003).
Collected data includes diameter-at-breast height
(DBH), stem density, mean tree height, dominant tree
height, crown closure (%), species and leaf area index
(LAI) (for further details, the reader is referred to
Thomas et al., 2006). Plot coordinates were collected at

V. Thomas et al. / Agricultural and Forest Meteorology 140 (2006) 287–307 291
Fig. 2. Groundhog River Flux Site (80 km SW of Timmins, Ontario). Latitude/longitude: 48.217N, 82.156W.
decimeter-level accuracy using a survey-grade differ-
entially corrected global positioning system (DGPS).
4.2. PAR measurements
In this experiment, global PAR was measured as
photosynthetic photon flux density (PPFD) by quantum
PAR sensors (model LI-190SA, LI-COR, Lincoln, NE,
USA). Each sensor is composed of a silicon photodiode
that produces a linear current signal proportional to the
total photon flux density in the spectral waveband 400–
700 nm. A millivolt adapter (model 2290S, LI-COR)
converted each current signal to a voltage. The millivolt
signal is converted into PPFD (in mmol m�2 s�1) by use
of a sensor-specific calibration coefficient. The sensor is
cosine corrected to an 808 angle from vertical. In
addition to global PAR, downwelling diffuse PAR was
measured with a sunshine sensor (model BF2, Delta-T
Devices, Burwell, Cambridge, UK). This device uses an
array of silicon photodiodes and a unique shading
pattern on the radiometer dome to determine down-
welling diffuse PAR (as PPFD). The accuracies of these
sensors are �5 and �15% for the LI-190SA and BF2,
respectively (Delta-T Devices, 2005; LI-COR, 2005).
The calibrations of these sensors are checked once a
year using a factory-standard LI-190SA quantum sensor
calibrated by LI-COR Inc. (Fluxnet-Canada, 2003). The
GRFS calibration strategy also involves sensor inter-
comparison with a network ‘‘roving’’ standard LI-
190SA, which is part of Fluxnet-Canada’s site inter-
calibration program. This program entails the periodic
deployment of network standard meteorological and
eddy covariance sensors to evaluate the performance of
the site’s sensors and to ensure comparability of
measurements between stations in the network.
4.2.1. Above- and below-canopy PAR measured at
the flux tower
Above-canopy downwelling global PAR (ACP),
above-canopy upwelling global PAR (ACPup), above-
canopy downwelling diffuse PAR (ACPdiff) and below-
canopy downwelling global PAR (BCP) are measured
continuously at the flux tower as part of the Fluxnet-
Canada measurement program. The sensors are located
at 1.5 and 37 m above the ground, on south-facing
booms that are levelled and extended from the tower
approximately 5 m. Data from the above- and below-
canopy PAR sensors are recorded by dataloggers (i.e.,
model CR23X and CR7X for above- and below-canopy
PAR, respectively; Campbell Scientific, Logan, UT,
USA). Data are measured at a scan rate of 1 s, with
statistics (i.e., mean, maximum, minimum, standard

V. Thomas et al. / Agricultural and Forest Meteorology 140 (2006) 287–307292
deviation) calculated by the datalogger at 1- and 30-min
intervals for above-canopy PPFD and 5- and 30-min
intervals for BCP.
4.2.2. Spatially variable below-canopy PAR
measurements
In addition to the measurements of PAR at the tower, a
small, self-contained and portable system of PAR sensors
was serially deployed at the six large tree plots to measure
spatial variability in fPAR (from June 12, 2004 (after leaf-
out) to August 14, 2004). The system consisted of six LI-
190SAs (named PAR1–PAR6), a Campbell Scientific
CR10 datalogger, a Campbell Scientific SM4M storage
module and a 12 V dc battery power supply. The
datalogger, storage module and power supply were
housed in a Campbell Scientific ENC12/14 fiberglass
enclosure to protect them from the elements. The sensors
measured every second, with 1-min statistics calculated
by the datalogger. The output data were stored on the
storage module until it was downloaded manually to a
computer. This datalogger’s internal clock was synchro-
nized to on-tower datalogger clocks before each
deployment to ensure synchronization between measure-
ments and accuracy of the fPAR calculations.
Within the six large tree plots selected for the BCP
measurements, the six PAR sensors were distributed
with 10 m spacing along north–south and east–west
transects. The placement of the sensors coincided with
the locations of hemispherical photographs which have
been repeatedly taken at the site in 2003 and 2004. The
hemispherical photographs provide a second, indepen-
dent source of information on canopy structure and leaf
area index within the approximate field of view of the
Fig. 3. Below-canopy PAR sensor
PAR sensor. Depending on which sensor is considered,
the distance between the sensors is 10, 14, 20, 30 or
31 m (Fig. 3). Each sensor was mounted on a tripod and
levelled at a height of 1.2 m above the ground. This
placement ensured that the sensors were above the
understory vegetation. The sensors were left in place for
at least 1 week at each large tree plot to capture as much
variability in solar radiation conditions as possible. The
diffuser surfaces on the LI-190SAs were regularly
checked and cleaned of any debris that may have
accumulated during the deployment.
To minimize differences in the sensitivity between
each sensor, all of the sensors were compared to one
standard LI-190SA (PAR1), which had been calibrated
at the factory by LI-COR. The sensor inter-comparison
was performed before and after the field experiment, at
Queen’s University in Kingston, Ontario, using the
same measurement system as in the field. The sensors
were deployed on the rooftop of a building with a
largely unobstructed hemispherical view of the sky.
They were mounted side-by-side, to minimize sensor
separation as much as possible (ASTM D4430-00e1,
2000), atop a tripod and individually levelled. The
sensors were left for over 1 week to capture a range of
conditions. From the data used in this comparison,
statistics were calculated between each sensor and
PAR1 (i.e., mean absolute error (MAE), mean bias error
(MBE), root mean square error (RMSE), the standard
deviation of the difference (ESOD), linear regression
and the coefficient of determination (r2)). The results of
the comparison were used to evaluate the performance
of each sensor and to derive sensor-specific corrections
for those sensors that did not match that of PAR1.
layout at the GRFS in 2004.

V. Thomas et al. / Agricultural and Forest Meteorology 140 (2006) 287–307 293
4.2.3. Estimation of fPAR
For each experiment plot, fPAR was estimated on a
half-hourly basis using the above-canopy PAR sensors
located on the tower and the six below-canopy PAR
sensors mounted on tripods beneath the canopy
(described above). Because the PAR reflectance over
each of the plot areas was unknown, the above-canopy
upwelling PAR (ACPup) measured at the flux tower was
used as a general canopy reflectance correction for all
plots. The canopy around the tower is mixed, with white
birch, trembling aspen and black spruce trees present
and a canopy height of approximately 20 m. The canopy
may be more open near the tower than in other mixed-
species locations, due to some tree loss during tower
construction. This has an impact on the general
reflectance correction, by increasing the ground
component of upwelling PAR as compared to some
other locations across the site. Thus, the calculation of
fPAR is considered to be an estimation of the true value.
The half-hour fPAR for each below-canopy sensor
was estimated using the following formula:
fPARi ¼�
1� BCPi
ACP
���
ACPup
ACP
�(1)
where BCPi: below-canopy total PAR for sensor i (i = 1,
. . ., 6); ACP: above-canopy downwelling PAR at the
flux tower; ACPup: above-canopy upwelling PAR at the
flux tower.
Mean daily fPAR for each sensor was also calculated
by averaging the half-hourly fPAR values over daylight
hours (i.e., 1200–2400 UTC).
To determine if the spatial variability in fPAR at the
GRFS would cause a significant change in the average
fPAR for the site based on tower measurements, the
same method of fPAR calculation was also completed
for the below-canopy PAR sensor mounted on the
tower as part of the Fluxnet-Canada measurement
protocol.
4.2.4. Direct versus diffuse PAR
A classification scheme based on sky condition was
devised to examine the influence of varying solar
radiation conditions on fPAR. This was accomplished
by comparing the fraction of ACPdiff and ACP (ACPdiff/
ACP) for the timeframe of the experiment. Values
approaching 1 for the entire day indicate overcast
(diffuse sunlight) conditions (i.e., ratio > 0.8), while
those approaching zero indicate sunny (direct sunlight)
conditions (i.e., ratio < 0.3). For the experiment at each
plot, the most sunny day and the most overcast day were
selected for model development.
4.3. Lidar data
The discrete lidar data collection for this study was
completed by Airborne 1 Corp. (a commercial data
provider based in El Segundo, CA, USA) using the
ALTM 2050 (Optech Inc., Toronto, Ontario, Canada).
This sensor operates at a wavelength of 1067 nm, with a
pulse rate of 50 kHz, a beam divergence setting of
0.3 mrad and a variable scan angle which ranges from
nadir to �208. The data were collected during the first
week of August 2003 at an approximate flying altitude
of 800 ft (244 m). The point spacing of the data across
the site ranged from approximately 3–8 pulses/m2 (i.e.,
3–8 first returns and 3–8 last returns), with a mean point
spacing of 4 pulses/m2 for the study plots. The
estimated positional accuracy at this flight altitude is
approximately 12.5 cm in the x–y direction, with an
error of less than 15 cm in the z direction.
4.3.1. Lidar analysis
In theory, the entire sphere of the sky above the PAR
sensor can have some influence on its recorded values.
When the PAR sensor is placed below the forest canopy,
significant portions of the sky are obstructed by trees
and no longer contribute to the below-canopy PAR
values. If one is trying to relate the below-canopy PAR
values to some calculation derived from lidar data, the
question arises about which portion of the forest canopy
above the sensor is most influencing the below-canopy
PAR values. This will affect the subsetting of lidar
points from the original point cloud for the canopy
above the PAR sensor.
Although there is currently no standard procedure for
analyzing lidar data in this context, some guidance can
be gained by examining results of fPAR and leaf area
index calculated from hemispherical photographs
situated below the canopy. When examining these
photographs, taken using a fish-eye lens, the sky can be
divided into a series of rings based on the angle from the
zenith of the sensor (i.e., directly overhead is 08). By
examining different rings, relative light availability and
canopy gaps can be investigated at specific zenith
angles (e.g., Nicotra et al., 1999). A similar approach
can be adopted for lidar data analysis, by creating a
theoretical cone which originates from the below-
canopy PAR sensor and extends skywards. By creating
theoretical lidar cones with different solid angles (or
theoretical lidar cone angle ‘‘a’’; Fig. 4) it is possible to
determine whether there exists a volume of lidar points
located above the PAR sensor that is most closely
related to fPAR (i.e., which portion of the forest canopy
above the sensor is most influencing the below-canopy

V. Thomas et al. / Agricultural and Forest Meteorology 140 (2006) 287–307294
Fig. 4. Variable canopy volumes within a theoretical lidar cone located directly above a below-canopy PAR sensor and opening towards the sky.
fPAR values). The optimal a is unknown, and mainly a
function of canopy height and density in the vicinity of
the sensor.
To determine whether an optimal a exists, multiple
lidar cones of increasing a were extracted over each
PAR sensor location (i.e., a = 9.158, 158, 208, 258, 308,358, 408, 458, 508, 558, 608, 658, 708, 758 and 808).Analyzing each a separately, the average lidar height
was calculated within each cone for every PAR sensor
location. These lidar heights, while not a direct measure
of canopy height or density, are a positive function of
both (Thomas et al., 2006). The lidar heights were
correlated to measures of fPAR over each below-canopy
PAR sensor. Result of each a was compared to
determine the optimal cone.
It should be noted here that as a increases, the
potential for overlap between lidar cones also increases,
particularly in the upper canopy. Whether or not this
actually causes overlap in the cone lidar data depends on
the physical structure of the canopy at the sensor
locations (i.e., whether or not there are lidar returns in
the area of overlap). If there is significant overlap
between two lidar cones (particularly for sensors closest
together), it is not statistically valid to consider them as
independent data points in a correlation analysis.
Therefore, the amount of overlap between the lidar
cones was tested for each a. Where necessary (i.e., for
large a), the ‘‘middle’’ sensors were removed from the
analysis to increase the distance between sensors (i.e.,
Fig. 3) and make them statistically independent.
In addition to mean lidar height within the cone,
several other measures of canopy height and density
were calculated and correlated to fPAR. Several authors
have demonstrated the utility of examining different
lidar percentiles ( p), where the pth percentile is defined
as the lidar height above the ground such that at least p%
of the lidar returns are at or below this height (Miller
et al., 1990; Lim and Treitz, 2004; Næsset, 2004;
Thomas et al., 2006). Therefore, the higher the
percentile, the higher the value of mean lidar height
will be. Previous work at the GRFS, demonstrated that
the 50th and 75th percentiles of lidar data are strongly
related to mean dominant height, basal area, crown
closure and above-ground biomass (R2 = 0.89, 0.86,
0.75 and 0.79, respectively) (Thomas et al., 2006). For
this analysis, the mean, maximum, minimum and
median were calculated for the 25th, 50th and 75th
percentiles and correlated to fPAR. A canopy density

V. Thomas et al. / Agricultural and Forest Meteorology 140 (2006) 287–307 295
function described in Næsset (2004) was also calcu-
lated, which involved dividing the height into equal
intervals and determining the proportions of lidar hits
above this height interval to the total number of pulses.
4.3.2. Estimation of vertical foliage profile
To facilitate interpretation of lidar relationships to
fPAR, a vertical foliage profile was estimated for each
species. This was done by generating vertical histo-
grams of all lidar returns contained in the 11.3 m radius
plots (e.g., the vertical histogram derived from the data
presented in Fig. 1 would be the representative
histogram for white birch) (Thomas et al., 2006).
4.4. Chlorophyll analysis
The ability of a canopy to absorb PAR is not only a
function of canopy architecture, but also of the inherent
capacity of the leaf to photosynthesize. Many factors
contribute to this, including the physiological status and
the chlorophyll concentration in the leaf (Waring et al.,
1995). Consider the scenario of a large black spruce tree
compared to a large white spruce tree. If the trees are of
a similar physical structure (i.e., height, shape, branch
architecture, etc.), they will appear very similar in the
lidar data. However, their inherent capacity to photo-
synthesize may be quite different, depending on the
chlorophyll concentration within the needles for each
species. This difference will affect the amount of PAR
absorbed by the canopy, but will not be represented by
the lidar data alone. To address variability in
chlorophyll concentration between species, field mea-
surements of leaf total chlorophyll (a + b) (mg/cm2)
were made and incorporated into the statistical
correlation analysis. The fPAR values were multiplied
by average leaf total chlorophyll (a + b) concentrations
(i.e., fPAR � chlorophyll) and compared with the lidar
data within the theoretical cones. Scatterplots and
regression analyses were used to determine the
correlation between fPAR � chlorophyll versus mean
lidar height and the predictive linear regression
equations for direct and diffuse sunlight. A map of
chlorophyll was generated to enable the application of
the linear regression model to the entire site.
4.4.1. Field measures of leaf chlorophyll
Leaf samples were collected from the top of the
canopy (by shooting the upper branches with a shotgun)
for analysis of canopy chlorophyll (e.g., O’Neill et al.,
2002; Zarco-Tejada et al., 2001). Thirty trees (five trees
per each of the six species) were sampled four times
throughout the growing season (i.e., leaf-out, just after
leaf-out, leaf flush and senescence). For trembling
aspen, white birch and cedar, branches from the north
and south side of the trees were sampled (with each
sample consisting of multiple leaves). Black spruce,
white spruce and balsam fir were further stratified into
old and new leaf samples. Mean leaf total chlorophyll
(Chl(a + b)) (mg/cm2) was determined using standard
laboratory spectrophotometric analysis of pigments
(e.g., Zarco-Tejada et al., 2000).
Average values of leaf total chlorophyll (a + b)
concentration (mg/cm2) were determined for each of the
major species at the site. An interspecies comparison
was completed using the difference of means test (i.e., F
test, based on mean, standard deviation and number of
samples) to demonstrate statistical significance in the
chlorophyll concentration differences between species.
4.4.2. Mapping chlorophyll with airborne
hyperspectral data
Hyperspectral data collection was completed over
the GRFS on August 15, 2004, using a Compact
Airborne Spectrographic Imager (CASI, Itres Research
Limited, Calgary, Alberta) at an altitude of approxi-
mately 1900 m. Seventy-two channels with an approx-
imate bandwidth of 7.5 nm were collected over the
wavelength range of 400–940 nm. The CAM5S atmo-
spheric model was used to pre-process the data to at-
sensor radiance and then to ground reflectance (Zarco-
Tejada et al., 2001; Freemantle, 2005). The 1 km radius
around the flux tower was imaged in three flight swaths,
which were georeferenced using a sub-metre digital
elevation model of the site and validated with
differentially corrected GPS data collected by a
survey-grade receiver. The pixel size of the georefer-
enced reflectance image was 4 m � 4 m.
Many authors have discussed the potential utility of
the ‘red edge’ of the spectral reflectance curve (i.e., the
wavelengths between maximum chlorophyll reflectance
in the red and high reflectance in the near infrared,
approximately 680–750 nm) for the prediction of
chlorophyll and other pigments (e.g., Filella et al.,
1995; Gitelson et al., 1996; Boochs et al., 1990; Curran
et al., 1990; Danson and Plummer, 1995; Munden et al.,
1994; Pinar and Curran, 1996). The first and second
derivatives of the spectral reflectance curve (dR and
ddR), particularly along the red edge, are thought to
mitigate the confounding variability of background
reflectance and illumination (Curran et al., 1991;
Elvidge and Chen, 1995; Zarco-Tejada et al., 2000,
2001). This is particularly important when moving from
laboratory measurement of spectral reflectance to
airborne and satellite hyperspectral imagery.

V. Thomas et al. / Agricultural and Forest Meteorology 140 (2006) 287–307296
For the work reported here, the maximum first
derivative over the red edge (i.e., max[dR680–750]) was
calculated from the casi hyperspectral data. Average
values were then extracted from 11 large tree plots
which were composed of either a single dominant
species or a relatively simple species mixture. These
values were compared to the average chlorophyll
concentration for the species present in the plot based
on the field measurements of leaf chlorophyll. The
linear regression model generated from these two
variables was applied to the entire image to produce a
map of total chlorophyll (a + b) for the site.
4.5. Effect of solar zenith angle
To determine the effect of variations in solar zenith
angle on fPAR and corresponding model performance,
the analysis was completed on both a diurnal and mean
daily basis. Diurnal variations in fPAR were examined
for each species for both direct and diffuse conditions.
The effect of increasing solar zenith angle on cor-
relations between fPAR (and fPAR � Chl) and mean
lidar height was also calculated.
4.6. Spatial modelling of mean daily fPAR
The best relationships between below-canopy PAR
and lidar under direct and diffuse sunlight conditions
were used to generate raster base maps showing the
spatial distribution of mean daily fPAR for the entire
GRFS. The map cell size was determined by calculating
the average H tan a (Fig. 4, where H is the canopy
height and a is optimal theoretical lidar cone angle). By
imposing a grid over the lidar data set, a cone of lidar
points was extracted over the centre of each grid cell and
the mean lidar height above the ground within that cone
was calculated for each cell. This resulting map was
used in conjunction with the chlorophyll map to solve
for fPAR in the linear regression model. Once maps of
fPAR were generated for the month of August for direct
Table 1
Plot mensuration data
TA/BF TA (medium) WB
DBH(quadratic) (cm) 25.7 21.8 15.6
Stem density (#/ha) 1400 1350 122
Mean height (m) 18.2 16.8 13.9
Mean dominant height (m) 32 25.2 16.6
Crown closure (m2/m2) 2.06 2.61 1.89
Species (%) TA-86, BF-10 TA-85, WB-13 WB
LAI (m2/m2) 2.27 1.72 2.42
Species codes—TA: trembling aspen; BF: balsam fir; WB: white birch; M
and diffuse conditions, descriptive statistics were then
generated for the entire GRFS and compared to
estimates of fPAR generated from the tower sensors
alone.
5. Results and discussion
A summary of mensurational data for the experiment
plots is provided in Table 1. It is evident that each plot is
quite distinct in terms of its physical structure and
species composition. The largest trees are found the
in TA/BF plot, which has two layers of canopy. The
upper layer is composed of large trembling aspen
(height > 30 m) with overlapping crowns. The lower
layer is composed mainly of smaller balsam fir, which
greatly increases the stem density of the plot, but does
not contribute to the upper canopy. The smallest trees
are located in the C/BS plot, which could be described
as a cedar thicket. In contrast, the BS plot can be
described as a thin black spruce bog, which extends well
beyond the plot boundary and can be seen as a large
distinct patch within the overall study site. WB, the
most commonly occurring species at the site, are
medium-sized, with less crown overlap than exists with
the TA. However, the crowns are fuller, with larger
leaves, than those in the aspen trees, resulting in a higher
LAI for the plot.
5.1. PAR and fPAR results
The calibration statistics for the below-canopy PAR
sensors are shown in Table 2 (where the calibration
sensor is the independent variable (x) and the other PAR
sensors are the dependent variables (y)). The high
correlation and minimal scatter for all sensors, as well
as the very low MAE, MBE, RMSE and ESOD
(Table 1), suggests that the only significant source of
error requiring correction is a systematic underestima-
tion of PAR for some sensors. Sensors 3 and 4 are within
the stated accuracy of the sensor (<5%). PAR2, PAR5
MW C/BS BS
17.1 16.0 12.1
5 1400 1725 575
16.3 8.8 9.7
22.6 18.1 13
1.63 0.97 0.16
-99 WB-52, TA-25, WS-15, BS-7 C-66, BS-29 BS-100
2.49 3.03 2.41
W: mixedwood; C: cedar; BS: black spruce.

V. Thomas et al. / Agricultural and Forest Meteorology 140 (2006) 287–307 297
Table 2
Pre-experiment calibration statistics for below-canopy PAR sensors
Sensor Regression equation R2 MAE (mV) MBE (mV) RMSE (mV) ESOD (mV)
Pre-experiment
2 Y = 0.92x + 0.002 0.99 0.23 0 0.04 0.04
3 Y = 0.97x + 2e�05 0.99 0.01 0 0.03 0.03
4 Y = 0.98x + 0.002 0.99 0.02 0 0.03 0.03
5 Y = 0.74x + 0.001 0.99 0.01 0 0.02 0.02
6 Y = 0.81x + 0.004 0.99 0.02 0 0.02 0.02
Post-experiment
2 Y = 1.01x + 0.001 0.99
3 Y = 0.98x + 0.001 1
4 Y = 0.99x + 0.002 0.99
5 Y = 0.97x + 3e�05 0.99
6 Y = 1.07x + 0.009 0.99
and PAR6 were corrected using the slope of the
regression equation as the calibration factor. Post-
experiment calibration statistics were also calculated to
identify any sensor drift throughout the experiment.
Table 2 demonstrates slight drift by the end of the
experiment for PAR6 (i.e., 2% beyond the accuracy of
the sensor). Therefore, values from this sensor were
ignored for August dates. High correlations and
accuracies are demonstrated for all other sensors as
compared to the original pre-experiment calibration
standard.
5.1.1. Direct versus diffuse PAR
Examination of the half-hour average above-canopy
direct and diffuse PAR measurements provides insight
Fig. 5. Half-hour incident above-canopy total and diffuse PAR (lower plots) a
experiment.
into the sunlight conditions throughout the various
experiments. Fig. 5 shows a sample time series of the
daily PAR regime above the canopy for the medium-
sized trembling aspen experiment, along with the fPAR
estimated from each of the below-canopy sensors. From
these data, it can be seen how sky conditions (i.e.,
cloudless or overcast) affects the radiation regime above
and below the canopy. For example, day of year (DOY)
164 (i.e., June 12, 2004) is a cloudless day, with a mean
daily ratio of ACPdiff/ACP of 0.27 (i.e., direct sunlight
conditions). There is a high degree of spatial and
temporal variability in fPAR between the six sensors,
with some low half-hourly values of fPAR (<0.4). This
is because during certain half-hourly periods (especially
around midday), the direct component of ACP was able
nd estimated fPAR (upper plots) for the medium-sized trembling aspen

V. Thomas et al. / Agricultural and Forest Meteorology 140 (2006) 287–307298
to penetrate the canopy through gaps and reach the
below-canopy sensors. The increase in fPAR and the
reduction in variability at the beginning and end of the
day can be explained by the fact that direct solar
radiation cannot penetrate as far into the canopy from an
oblique angle, and thus the radiation reaching the senor
is mainly diffuse during these times.
In contrast, DOY 169 (June 17) is mainly overcast,
with a mean daily ratio of ACPdiff/ACP of 0.97 (i.e.,
diffuse sunlight conditions). The even distribution of
PAR throughout the canopy causes a reduction in the
variability in fPAR throughout the day. DOYs 165–168
show mixed conditions and consequently mixed
variability in fPAR and ACPdiff/ACP.
5.1.2. Influence of solar zenith angle, species and
canopy architecture on fPAR
Differences in estimated fPAR for the various species
configurations are shown for both direct (Fig. 6) and
diffuse (Fig. 7) sunlight conditions. The highest
estimated fPAR occurs in the large trembling aspen plot
(with balsam fir understory) for both cloudless and
overcast conditions. This plot demonstrates an insensi-
Fig. 6. Estimated fPAR and incoming total PAR for each of the experim
Trembling aspen (medium size); (B) black spruce; (C) mixedwood; (D) tre
tivity of fPAR to solar zenith angle, with a very slight
decrease in fPAR at around solar noon under cloudless
conditions. The maximum estimated fPAR (0.99) occurs
in the morning and late afternoon (i.e., large zenith
angles), with a minimum value (0.96) at around solar
noon (Fig. 6D). The vertical foliage profile calculated
from lidar data (Fig. 8) demonstrates high amounts of
lidar returns from the upper canopy. This suggests that the
large branches and foliage of the trembling aspen canopy
are preventing lidar penetration of these trees. A similar
effect would occur with the direct sunlight, causing
multiple scattering and reflection from the canopy. This
would increase the amount of diffuse light within the
canopy, reducing the sensitivity of fPAR to solar zenith
angle under cloudless conditions. Further contributing to
the higher estimated fPAR values is the larger volume of
canopy that is visible to the PAR sensors as a result of the
taller trees (Fig. 4), providing more opportunities for
foliage to absorb the PAR during photosynthesis.
The black spruce bog has the lowest value of fPAR
and the highest degree of sensitivity to solar zenith
angle shows a high degree of sensitivity to solar zenith
angle for both direct and diffuse conditions (Figs. 6B
ent plots under direct sunlight conditions at the GRFS in 2004. (A)
mbling aspen and balsam fir; (E) white birch; (F) cedar/black spruce.

V. Thomas et al. / Agricultural and Forest Meteorology 140 (2006) 287–307 299
Fig. 7. Estimated fPAR and incoming total PAR for each of the experiment plots under diffuse sunlight conditions at the GRFS in 2004. (A)
Trembling aspen (medium size); (B) black spruce; (C) mixedwood; (D) trembling aspen and balsam fir; (E) white birch; (F) cedar/black spruce.
and 7B). The vertical foliage profile (Fig. 8) is markedly
different for these species, showing three heights of
significant lidar returns, all lower than 10 m above
ground. The mean tree height for this plot is 10 m and
Fig. 8. Vertical foliage profiles estimated from lidar for PAR experi-
ment plots at the GRFS in 2004.
the maximum height is 16 m. The highest of the three
lobes of lidar returns corresponds to slightly less than
the mean height, likely because of the improbability of
the lidar pulse striking the apex of the crown. The lower
lobe indicates that many of the lidar returns reached the
ground (which suggests a significant component of the
direct sunlight would also reach the ground, lowering
fPAR for the plot under cloudless conditions). Lower
fPAR for diffuse conditions may result from a relatively
small amount of canopy falling within the field of view,
meaning considerably less foliage available for photo-
synthesis (note the very low value for canopy closure,
0.16; Table 1).
Low lidar heights (<10 m) are also evident for the
cedar/black spruce plot (Fig. 8). Once again, the cedar/
black spruce plot has a relatively open canopy (canopy
closure is 0.97; Table 1), although there is a much higher
stem density than exists for the black spruce bog
(1725 ha�1 as compared to 575 ha�1; Table 1). Although
there is not a significant lobe of lidar returns near the
ground level, the foliage profile demonstrates lidar
returns for the entire vertical distribution of the canopy,

V. Thomas et al. / Agricultural and Forest Meteorology 140 (2006) 287–307300
Fig. 9. Seasonal mean total chlorophyll by species at the GRFS in
2004.
including the lower trunks. Again, this suggests a high
probability for direct sunlight to penetrate the canopy.
At first glance, the medium-sized TA, mixedwood
and WB plots show similar foliage distributions to the
large aspen/balsam fir plot and to each other. However,
upon consideration of the true canopy structure and tree
height for these plots, it becomes evident that the lidar
pulses are behaving differently for each case (Note:
mean height = 17, 16 and 14 m for medium-sized TA,
mixedwood and WB, respectively). In the case of the
medium-sized aspen, most of the lidar pulses are
returned from dominant trees (height > 20 m), whereas
the white birch allows greater penetration into the
canopy, with the maximum lidar returns occurring at
approximately 11 m. The white birch also has a slightly
lower fPAR for diffuse conditions, suggesting a
relationship between tree height and fPAR. The
mixedwood plot falls in the middle of these two cases,
with the height of maximum lidar return being
approximately the same as the mean tree height. There
is no notable difference in fPAR between the medium-
sized aspen and mixedwood plots under diffuse
conditions, but there is a slight decrease in mean fPAR
for the medium-sized aspen under direct conditions.
5.2. Lidar and fPAR
All of the lidar metrics calculated for this analysis are
a function of canopy height and density and were found
to be moderately-to-strongly correlated to fPAR and
fPAR � chlorophyll. Given that the strongest and most
consistent correlation to fPAR and fPAR � chlorophyll
is with mean lidar height, the discussion will focus on
this variable. Across all of the tested theoretical lidar
cone angles (‘‘a’’), the linear correlation coefficient
between mean daily fPAR and mean lidar height for the
cone of extraction ranges from r = 0.53 to 0.79 for direct
sunlight conditions and from r = 0.69 to 0.87 for diffuse
conditions (Fig. 11). A lidar cone with a = 558 clearly
showed the best correlation under both direct and
diffuse conditions. At this angle, data is included from
16 sensors (i.e., no overlap between lidar cones for these
16 sensors). Examination of the mean daily fPAR
scatterplots (Fig. 13A and B) reveals stronger relation-
ships and less variability for the diffuse model (for a
simple linear relationship, R2 = 0.75 and 0.62 for direct
and diffuse models, respectively).
Using a = 558, the diurnal relationships between
lidar and fPAR are demonstrated for diffuse light
(Fig. 12A) and direct light (Fig. 12B). For diffuse
conditions, there is a relatively small effect of solar
zenith angle on the performance of the fPAR–lidar
relationships (i.e., Spearman r correlation values range
from 0.74 to 0.85). The relationship is non-linear, which
peaks at 558 and tapers off for higher solar zenith angles.
Under clear sky conditions, there is more variability
with increasing solar zenith angle. This is due to the
relatively open canopy of the large homogenous black
spruce patch, which allows for a high amount of light
penetration through gaps. This causes a decrease in
correlation with fPAR for certain sensor locations,
depending on the relative position of neighbouring trees
and the sun. At very large zenith angles (i.e., >558),trees along the periphery of the sensor reduce the effect
of large gaps and the result is a relatively constant
relationship between lidar and fPAR (r > 0.70).
Because the lidar volume is constant, there is a
decrease in the strength of the relationship between
lidar and fPAR (as well as an increase it the variability
of the relationship) for smaller zenith angles.
5.3. Measured chlorophyll
Seasonal mean chlorophyll per species is shown in
Fig. 9. For most dates, TA has the highest chlorophyll
concentration. As expected, the chlorophyll concentra-
tions of the deciduous species increase throughout the
growing season and decrease as senescence begins. There
is no consistent trend for the coniferous species, although
WS, BS and BF do show variability throughout the
growing season. This variability is mainly a function of
the new needles, which begin to grow at approximately
mid-June and increase in chlorophyll throughout the
season. The old needles were found to have a relatively
constant chlorophyll concentration throughout the
season. Although the new needles cause an apparent
decrease in chlorophyll concentration for the spruce
trees, they are important to consider because they are the

V. Thomas et al. / Agricultural and Forest Meteorology 140 (2006) 287–307 301
Fig. 10. Mean canopy chlorophyll for PAR experiment plots at the
GRFS in 2004. Note: WS—white spruce.
Fig. 12. Effects of solar zenith angle on fPAR and fPAR � Chl
relationships with lidar for: (A) overcast (diffuse light) conditions
and (B) sunny (direct light) conditions.
most visible needles to the hyperspectral sensor. BS has
the lowest leaf chlorophyll of all species tested.
Mean chlorophyll per plot is presented in Fig. 10.
Chlorophyll samples from the sampling date closest to
the date of the below-canopy PAR experiment at each
plot were used for the calculations. The mixedwood
value was determined by averaging the chlorophyll
values for TA, WB, WS and BF. The two values
reported for the TA/BF plot represent the species
composition surrounding each of the sensors (i.e.,
sensors 2, 4, 5 and 6 are surrounded by an even mixture
of TA/BF, while sensors 1 and 3 are surrounded by
mainly BF). As expected, BS has the lowest chlorophyll
value, while TA/BF has the highest. Shapiro–Wilks
statistics of the plot chlorophyll concentrations demon-
strate normality of the distribution (n = 7, W = 95,
p = 0.75), meaning that parametric statistics can be
applied with confidence (e.g., linear regression statis-
tics, etc.) (Shapiro et al., 1968).
Fig. 11. Correlation curves for mean daily fPAR and fPAR � chlorophyll
theoretical lidar cone angles.
5.3.1. Influence of chlorophyll on fPAR–lidar
relationships
Both the mean daily (Fig. 11) and diurnal (Fig. 12A
and B) analysis show an increase in the strength and a
decrease in the variability of the relationships between
(Chl) vs. lidar return for direct and diffuse conditions with varying
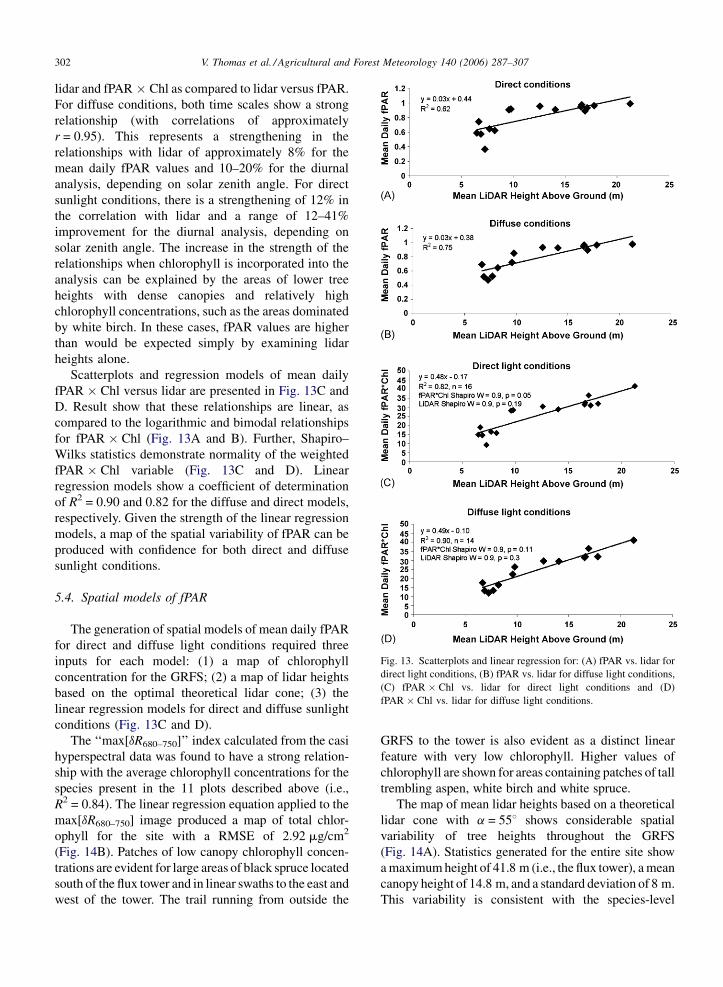
V. Thomas et al. / Agricultural and Forest Meteorology 140 (2006) 287–307302
Fig. 13. Scatterplots and linear regression for: (A) fPAR vs. lidar for
direct light conditions, (B) fPAR vs. lidar for diffuse light conditions,
(C) fPAR � Chl vs. lidar for direct light conditions and (D)
fPAR � Chl vs. lidar for diffuse light conditions.
lidar and fPAR � Chl as compared to lidar versus fPAR.
For diffuse conditions, both time scales show a strong
relationship (with correlations of approximately
r = 0.95). This represents a strengthening in the
relationships with lidar of approximately 8% for the
mean daily fPAR values and 10–20% for the diurnal
analysis, depending on solar zenith angle. For direct
sunlight conditions, there is a strengthening of 12% in
the correlation with lidar and a range of 12–41%
improvement for the diurnal analysis, depending on
solar zenith angle. The increase in the strength of the
relationships when chlorophyll is incorporated into the
analysis can be explained by the areas of lower tree
heights with dense canopies and relatively high
chlorophyll concentrations, such as the areas dominated
by white birch. In these cases, fPAR values are higher
than would be expected simply by examining lidar
heights alone.
Scatterplots and regression models of mean daily
fPAR � Chl versus lidar are presented in Fig. 13C and
D. Result show that these relationships are linear, as
compared to the logarithmic and bimodal relationships
for fPAR � Chl (Fig. 13A and B). Further, Shapiro–
Wilks statistics demonstrate normality of the weighted
fPAR � Chl variable (Fig. 13C and D). Linear
regression models show a coefficient of determination
of R2 = 0.90 and 0.82 for the diffuse and direct models,
respectively. Given the strength of the linear regression
models, a map of the spatial variability of fPAR can be
produced with confidence for both direct and diffuse
sunlight conditions.
5.4. Spatial models of fPAR
The generation of spatial models of mean daily fPAR
for direct and diffuse light conditions required three
inputs for each model: (1) a map of chlorophyll
concentration for the GRFS; (2) a map of lidar heights
based on the optimal theoretical lidar cone; (3) the
linear regression models for direct and diffuse sunlight
conditions (Fig. 13C and D).
The ‘‘max[dR680–750]’’ index calculated from the casi
hyperspectral data was found to have a strong relation-
ship with the average chlorophyll concentrations for the
species present in the 11 plots described above (i.e.,
R2 = 0.84). The linear regression equation applied to the
max[dR680–750] image produced a map of total chlor-
ophyll for the site with a RMSE of 2.92 mg/cm2
(Fig. 14B). Patches of low canopy chlorophyll concen-
trations are evident for large areas of black spruce located
south of the flux tower and in linear swaths to the east and
west of the tower. The trail running from outside the
GRFS to the tower is also evident as a distinct linear
feature with very low chlorophyll. Higher values of
chlorophyll are shown for areas containing patches of tall
trembling aspen, white birch and white spruce.
The map of mean lidar heights based on a theoretical
lidar cone with a = 558 shows considerable spatial
variability of tree heights throughout the GRFS
(Fig. 14A). Statistics generated for the entire site show
a maximum height of 41.8 m (i.e., the flux tower), a mean
canopy height of 14.8 m, and a standard deviation of 8 m.
This variability is consistent with the species-level

V. Thomas et al. / Agricultural and Forest Meteorology 140 (2006) 287–307 303
Fig. 14. Characteristics of the GRFS in 2004. (A) Mean lidar heights optimal theoretical lidar cone of a = 558 (20 m cell size), (B) mean canopy total
chlorophyll (a + b) modelled from casi hyperspectral data (4 m cell size), (C) estimated fPAR for August for sunny (direct light) conditions, (D)
estimated fPAR for August for overcast (diffuse light) conditions and (E) spatial differences between fPAR for direct and diffuse light conditions.
mensuration data (Table 1) which show a range of mean
dominant heights from 13 to 32 m.
Integration of the lidar and chlorophyll maps with
the linear regression models result in a map of the
spatial variability of fPAR for direct and diffuse
conditions for the month of August (Fig. 14C and
D). Considerable spatial variability is evident through-
out the site, although the spatial patterns are not as

V. Thomas et al. / Agricultural and Forest Meteorology 140 (2006) 287–307304
Table 3
Spatially modelled mean daily fPAR data compared with tower calculated fPAR
Reflectance up Tower data Modelled data
Fraction diffuse Mean daily fPAR S.D. (temporal) Mean daily fPAR S.D. (spatial)
Direct (cloudless) 0.034 0.243 0.882 0.008 0.787 0.131
Diffuse (overcast) 0.033 1.066 0.846 0.009 0.783 0.136
distinct. Statistics were generated for the entire site and
compared to values of fPAR estimated for the month of
August (Table 3).
Examination of the standard deviations of the tower
estimate fPAR (Table 3) reveals low temporal variability
in fPAR throughout the month of August for both direct
and diffuse conditions (i.e., standard deviation < 1%).
However, no information about the spatial variability of
fPAR can be gleaned from the tower results. In contrast,
the modelled data standard deviations show significant
variability in fPAR within the 1 km radius (i.e., standard
deviation > 13% with a range of more than 0.8 for both
sunny and overcast conditions). Depending on the wind
speed and direction (and resulting tower footprint), this
could have a significant impact on modelled photo-
synthesis as compared to estimates of fPAR from the
tower. Further, when comparing tower-measured versus
modelled mean fPAR, the tower results are approxi-
mately 9% higher for direct conditions and 6% higher for
diffuse conditions. This is most likely because of to the
large areas of black spruce within the GRFS with
relatively open canopies (resulting in a lower lidar height
estimate for the cone of extraction) and with relatively
low canopy chlorophyll, which is a very different canopy
configuration than what exists at the tower.
The modelled results of mean fPAR suggest very
little sensitivity to sunny versus diffuse light conditions.
However, examination of the spatial distribution of
these differences (Fig. 14E) reveals a distinct spatial
pattern. Direct sunlight conditions show a greater fPAR
than diffuse conditions for the large areas of black
spruce and other areas with relatively low chlorophyll or
open canopies. However, more PAR is absorbed under
diffuse conditions for areas containing more deciduous
trees, which have higher canopy chlorophyll concen-
trations and fewer gaps. This is likely because under
diffuse conditions, light reaches all parts of the canopy
equally well because it is coming from all directions,
whereas under direct sunlight conditions, some portions
of the closed canopy will be in shade (e.g., Gu et al.,
2003, 2002; Freedman et al., 2001).
The models described above are based on the
assumption that canopy structure and physiology (i.e.,
chlorophyll available for photosynthesis) are two of the
main factors affecting fPAR. Canopy structure, parti-
cularly lidar metrics based on height, will not change
significantly throughout the growing season for a
mature stand (unless there is significant structural
damage caused by a major event, such as a tornado or
fire). Therefore, the major temporal differences in fPAR
are likely to be caused by changes in canopy
physiology. The very low variability in fPAR over
the month of August, calculated from the tower sensors
(Table 3), suggests that a single base map for the month
of August is sufficient (i.e., there is not a significant
change in canopy physiology for the month of August).
Examination of the months of June and July show that
fPAR increases from 0.80 to 0.89, while the standard
deviation decreases from 0.03 to 0.01. This suggests
that most of the temporal variability in fPAR occurs in
June, during leaf-out and the period directly after leaf-
out when the leaves are maturing. Therefore, capturing
the change in canopy chlorophyll during leaf-out and
after leaf-out, as well as monthly throughout the rest of
the growing season, should be sufficient to model the
temporal variability in fPAR. This information, in
conjunction with continuous measurements of the
temperature and total incoming direct/diffuse PAR
(measured above the canopy at the tower) will allow for
the modelling of photosynthesis at the site throughout
the growing season which incorporates the spatial
variability at the site.
6. Conclusions
The strong relationship between lidar and fPAR once
species chlorophyll concentrations are included suggest
that if lidar data and chlorophyll maps are available, local
spatial variability in fPAR can be mapped for hetero-
geneous forests. This has important implications for at
least two related streams of research: (1) these data may
help fill the ‘‘missing link’’ in scaling efforts from point
measurements at flux towers to larger resolution satellite
imagery such as MODIS (e.g., Huemmerich et al., 2005;
Chen et al., 2003; Knyazikhin et al., 1998a,b), and (2)
addressing local scale heterogeneity for mixedwoods
and its impact on light absorption and photosynthesis
(e.g., Shabanov et al., 2003).

V. Thomas et al. / Agricultural and Forest Meteorology 140 (2006) 287–307 305
The multiple species present at the GRFS provide the
ideal scenario to characterize species structural differ-
ences and their impact on the spatial pattern of mean
daily fPAR. The analysis reveals a number of key
findings in this regard:
1. T
here are distinct structural differences betweenthe major species present at the GRFS. These
differences are evident in the apparent foliage
profiles estimated from airborne lidar measure-
ments.
2. S
trong correlations (r, R2) exist between fPAR andmean lidar heights (a function of canopy height and
density). Solar zenith angle has an impact on the
relationship between fPAR and mean lidar heights,
particularly under direct sunlight conditions. This is
most prevalent in more open canopies, such as the
homogenous black spruce bog. In this patch,
correlations decrease for certain sensor locations,
depending on the relative position of the sensor,
neighbouring trees, and the sun.
3. H
yperspectral remote sensing data and fieldsampling indicate that there are significant differ-
ences in canopy chlorophyll concentrations
between the major species at the GRFS.
4. f
PAR � chlorophyll has stronger correlations withlidar than fPAR alone. For diffuse conditions,
correlation (r) is stronger by 8% for mean daily
fPAR and from 10 to 20% for diurnal fPAR,
depending on solar zenith angle. For sunny (direct
light) conditions, correlation to lidar is improved by
12% for mean daily fPAR and 12–41% for diurnal
relationships, depending on solar zenith angle.
Chlorophyll also reduces diurnal variability caused
by the solar zenith angle and makes the relation-
ships to lidar more linear.
5. S
imulation of multiple theoretical lidar coneslocated above the below-canopy PAR sensor reveals
consistently stronger relationships with fPAR when
using lidar points falling within a cone defined by
a = 558.
6. W ith a theoretical lidar cone angle of a = 558, linearregression models were developed with 90% of the
variance explained for diffuse sunlight conditions
and 82% of the variance explained for direct
conditions.
7. f
PAR calculated from above and below-canopyPAR sensors mounted on the tower shows very low
variation over the month of August. Average fPAR
was 0.88 for direct sunlight conditions and 0.85 for
diffuse conditions with standard deviation less than
1% for both direct and diffuse conditions. No
information about the spatial variability can be
gleaned from these data.
8. T
he fPAR maps developed for the month of Augustfor the entire GRFS show a site average fPAR of
0.79 for direct light conditions and 0.78 for diffuse
conditions. This is lower than the point estimate of
fPAR derived by tower measures of above and
below-canopy PAR by 9% for direct conditions and
7% for diffuse conditions.
9. S
tatistics calculated over the spatial extent of thesite demonstrate standard deviation of fPAR of
approximately 0.13 and a range of approximately
0.80 for both direct and diffuse sunlight conditions.
This represents a spatial variability that is much
larger than the temporal variability for the month of
August.
10. M
apping of fPAR for direct and diffuse sunlightconditions allows for a spatial comparison (i.e.,
subtraction of the maps) that reveals a distinct
spatial pattern showing that areas with open
canopies and relatively low chlorophyll (e.g., black
spruce patches) have a higher fPAR under direct
sunlight conditions, while closed canopies with
higher chlorophyll (e.g., deciduous species) absorb
more PAR under diffuse conditions.
11. D
epending on the wind direction, wind speed andspatial extent of the flux footprint (i.e., the source-
area for scalar fluxes with respect to the eddy
covariance system), modelled photosynthesis based
on fPAR derived from tower-mounted PAR sensors
could deviate significantly from those derived from
fPAR calculated from the area within the tower
footprint.
Future work evolving from the analysis described in
this paper will involve the expansion of the model to
incorporate changes in canopy chlorophyll over the
growing season. These will be used in conjunction with
total incoming direct and diffuse PAR, along with
continuous measurements of temperature, wind speed,
wind direction, etc., to model photosynthesis within the
variable tower footprint throughout the entire year of
2004. Modelled results will be compared to estimates of
gross ecosystem productivity calculated from tower
measurements.
Acknowledgements
The authors would like to thank the anonymous
reviewers for their insights and helpful comments
which, we feel, have greatly strengthened this manu-
script. We also gratefully acknowledge the field efforts

V. Thomas et al. / Agricultural and Forest Meteorology 140 (2006) 287–307306
of Denzil Irving, Bob Oliver, David Atkinson, BjOrn
Prenzel, Chris Hopkinson, Laura Chasmer, Brock
McLeod, Lauren MacLean and Adam Thompson. We
would also like to thank Al Cameron and Lincoln
Rowlinson for establishing cruise-line trail and on-site
assistance with tree species identification. Garry
Koteles provided valuable information regarding tree
and understory species and did all of the field
measurements for the NFI validation plots. The Ontario
Forest Research Institute (OFRI) provided technical
expertise for sampling and processing of leaves, use of
laboratory facilities, and performed the pigment
concentration analysis. Financial assistance for this
work was provided by the Natural Sciences and
Engineering Research Council of Canada (NSERC),
Fluxnet-Canada (NSERC, BIOCAP Canada, and the
Canadian Foundation for Climate and Atmospheric
Sciences (CFCAS)), the Ontario Graduate Scholarship
Program (OGS), the Centre for Research in Earth and
Space Technology (CRESTech), and Queen’s Univer-
sity.
References
Aldred, A., Bonner, M., 1985. Application of Airborne Lasers to
Forest Surveys. Canadian Forestry Service, Petawawa National
For. Inst. Inf. Rep. PI-X-51, 62 pp.
Arp, H.J., Griesbach, J., Burns, J., 1982. Mapping in tropical forests: a
new approach using the laser APR. ISPRS J. Photogram. Rem.
Sens. 48, 91–100.
ASTM D4430-00e1, 2000. Standard Practice for Determining the
Operational Comparability of Meteorological Measurements.
ASTM International, www.ASTM.org.
Baldocchi, D.D., Hutchinson, B.A., Matt, D.R., McMillen, R.T., 1984.
Solar radiation in an oak–hickory forest: an evaluation of extinc-
tion coefficients for several radiation components during full-
leafed and leafless periods. Agric. For. Meteorol. 32, 317–322.
Baltsavias, E.P., 1999. A comparison of between photogrammetry and
laser scanning. ISPRS J. Photogram. Rem. Sens. 54, 83–94.
Blackburn, G.A., 2002. Remote sensing of forest pigments using
airborne imaging spectrometer and LIDAR imagery. Rem. Sens.
Environ. 82, 311–321.
Blackburn, G.A., 1998. Quantifying chlorophylls and caroteniods at
leaf and canopy scales: an evaluation of some hyperspectral
approaches. Rem. Sens. Environ. 66, 273–285.
Bonan, G.B., 1998. The land surface climatology of the NCAR land
surface model coupled the NCAR community climate model. J.
Clim. 11, 1307–1327.
Boochs, F., Dockter, K., Kupfer, G., Kuhbauch, W., 1990. Shape of the
red edge as a vitality indicator for plants. Int. J. Rem. Sens. 11,
1741–1754.
Chen, J.M., Blanken, P., Black, T.A., Guilbeault, M., Chen, S., 1997.
Radiation regime and canopy architecture of a boreal aspen forest.
Agric. For. Meteorol. 86, 107–125.
Chen, J.M., Liu, J., Leblanc, S.G., Lacaze, R., Roujean, J.-L., 2003.
Multi-angular optical remote sensing for assessing vegetation
structure and carbon absorption. Rem. Sens. Environ. 84, 516–
525.
Curran, P.J., Dungan, J.L., Golz, H.L., 1990. Exploring the relation-
ship between reflectance red edge and chlorophyll content in slash
pine. Tree Physiol. 7, 33–48.
Curran, P.J., Dungan, J.L., Macler, B.A., Plummer, S.E., 1991. The
effect of a red leaf pigment on the relationship between red-
edge and chlorophyll concentration. Rem. Sens. Environ. 35,
69–75.
Danson, F.M., Plummer, S.E., 1995. Red-edge response to forest leaf
area index. Int. J. Rem. Sens. 16, 183–188.
Datt, B., 1998. Remote sensing of chlorophyll a, chlorophyll b,
chlorophyll a + b and total carotenoid content in eucalyptus
leaves. Rem. Sens. Environ. 66, 111–121.
Delta-T Devices, 2005. Product News, Light Measurement and
Canopy Analysis, Sunshine Sensor Type BF-2. Delta-T Devices,
Ltd., Burwell, Cambridge, England. Available from http://
www.delta-t.co.uk [cited 3 October 2005].
Dickinson, R.E., Henderson-Sellers, A., Kennedy, P.J., Wilson, M.F.,
1986. Biosphere–Atmosphere Transfer Scheme (BATS) for the
NCAR CCM, NCAR/TN-275-STR. NCAR Research, Boulder,
CO.
Eck, T.F., Deering, D.W., 1992. Canopy albedo and transmittance in
spruce-hemlock forest in mid-September. Agric. For. Meteorol.
59, 237–248.
Elvidge, C.D., Chen, Z., 1995. Comparison of broad-band and nar-
rowband red and near-infrared vegetation indices. Rem. Sens.
Environ. 54, 38–48.
Filella, I., Serrano, L., Serra, J., Penuelas, J., 1995. Evaluating wheat
nitrogen status with canopy reflectance indices and discriminant
analysis. Crop Sci. 35, 1400–1405.
Fluxnet-Canada, 2003. Fluxnet-Canada Measurement Protocols,
Working Draft Version 1.3 [online]. Fluxnet Canada Network
Management Office, Laval, Quebec. Available from http://
www.fluxnet-canada.ca [cited 3 February 2005].
Freedman, J.M., Fitzjarrald, D.R., Moore, K.E., Sakai, R.K., 2001.
Boundary layer clouds and vegetation–atmosphere feedbacks. J.
Clim. 14, 180–197.
Freemantle, J., 2005. CASI Data Collection Report, Sudbury 2004,
Groundhog River. York University, Unpublished document.
Gitelson, A.A., Kaufman, Y.J., Merzlyak, M.N., 1996. Use of a green
channel in remote sensing of vegetation from EOS-MODIS. Rem.
Sens. Environ. 58, 289–298.
Gu, L., Baldocchi, D., Verma, S.B., Black, T.A., Vesala, T., Falge,
E.M., Dowty, P.R., 2002. Advantages of diffuse radiation for
terrestrial ecosystem productivity. J. Geophys. Res. 107, 1–23.
Gu, L., Baldocchi, D.B., Wofsy, S.C., Munger, J.W., Michalsky, J.J.,
Urbanski, S.P., Boden, T.A., 2003. Response of a deciduous forest
to the mount Pinatubo eruption: enhanced photosynthesis. Science
299, 2035–2038.
Hardy, J.P., Melloh, R., Koenig, G., Marks, D., Winstral, A., Pomeroy,
J.W., Link, T., 2004. Solar radiation transmission through conifer
canopies. Agric. For. Meteorol. 126, 257–270.
Huemmerich, K.F., Privette, J.L., Mukelabai, M., Myneni, R.B.,
Knyazikhin, Y., 2005. Time-series validation of MODIS land
biophysical products in a Kalahari Woodland, Africa. Int. J.
Rem. Sens. 26, 4381–4398.
Knyazikhin, Y., Martonchik, J.V., Diner, D.J., Myneni, R.B., Ver-
straete, M., Pinty, B., Gobron, N., 1998a. Estimation of vegetation
canopy leaf area index and fraction of absorbed photosynthetically
active radiation from atmosphere-corrected MISR data. J. Geo-
phys. Res. 103, 32239–32257.

V. Thomas et al. / Agricultural and Forest Meteorology 140 (2006) 287–307 307
Knyazikhin, Y., Martonchik, J.V., Myneni, R.B., Diner, D.J., Running,
S.W., 1998b. Synergistic algorithm for estimating vegetation
canopy leaf area index and fraction of absorbed photosynthetically
active radiationfrom MODIS and MISR data. J. Geophys. Res.
103, 32257–32277.
Lefsky, M.A., Cohen, W.B., Acker, S.A., Spies, T.A., Parker, G.G.,
Harding, D., 1997. Lidar Remote Sensing of Forest Canopy
Structure and Related Biophysical Parameters at the H.J. Andrews
Experimental Forest, Oregon, USA. In Natural Resources Man-
agement Using Remote Sensing and GIS. ASPRS, Washington,
DC, pp. 79–91.
Lefsky, M.A., Cohen, W.B., Acker, S.A., Parker, G.G., Spies, T.A.,
Harding, D., 1999. Lidar remote sensing of the canopy structure
and biophysical properties of Douglas-fir western hemlock forests.
Rem. Sens. Environ. 70, 339–361.
LI-COR, 2005. LI-190 Specifications. LI-COR Biosciences 4421
Superior St. Lincoln, NE 68504, USA. Available from http://
www.licor.com [cited 3 October 2005].
Lim, K.S., Treitz, P.M., 2004. Estimation of above ground forest
biomass from airborne discrete return laser scanner data using
canopy-based quantile estimators. Scand. J. For. Res. 19, 558–570.
Lim, K., Treitz, P., Baldwin, K., Morrison, I., Green, J., 2003a. LiDAR
remote sensing of biophysical properties of tolerant northern
hardwood forests. Can. J. Rem. Sens. 29, 658–678.
Lim, K., Treitz, P., Wulder, M., St-Onge, B., Flood, M., 2003b. Lidar
remote sensing of forest structure. Prog. Phys. Geogr. 27, 88–106.
Magnussen, S., Boudewyn, P., 1998. Derivations of stand heights from
airborne laser scanner data with canopy-based quantile estimators.
Can. J. For. Res. 28, 1016–1031.
Miller, I., Freund, J.E., Johnson, R.A., 1990. Probability and Statistics
for Engineers, fourth ed. Prentice Hall, Englewood Cliffs.
Munden, R., Curran, P.J., Catt, J.A., 1994. The relationship between
red edge and chlorophyll concentration in the Broadbalk winter
wheat experiment at Rothamsted. Int. J. Rem. Sens. 15, 705–709.
Næsset, E., 1997. Determination of mean tree height of forest stands
using airborne laser scanner data. ISPRS J. Photogram. Rem. Sens.
52, 49–56.
Næsset, E., 2004. Effects of different flying altitudes on biophysical
stand properties estimated from canopy height and density mea-
sured with a small-footprint airborne scanning laser. Rem. Sens.
Environ. 91, 243–255.
Nicotra, A.B., Chazdon, R.L., Iriarte, S.V.B., 1999. Spatial hetero-
geneity of light and woody seedling regeneration in tropical wet
forests. Ecology 80, 1908–1926.
Nilsson, M., 1996. Estimation of tree heights and stand volume using
an airborne lidar system. Rem. Sens. Environ. 56, 1–7.
O’Neill, A.L., Kupiec, J.A., Curran, P.J., 2002. Biochemical and
reflectance variation throughout a Sitka spruce canopy. Rem.
Sens. Environ. 80, 134–142.
Parker, G.G., Lefsky, M.A., Harding, D.J., 2001. Light transmittance
in forest canopies determined using airborne laser altimetry and
in-canopy quantum measurements. Rem. Sens. Environ. 76, 298–
309.
Pinar, A., Curran, P.J., 1996. Grass chlorophyll and the reflectance red
edge. Int. J. Rem. Sens. 17, 351–357.
Running, S.W., Coughlan, J.C., 1988. A general model of forest
ecosystem processes for regional applications. Ecol. Model. 42,
124–154.
Sellers, P.J., Randall, D.A., Collatz, G.J., Berry, J.A., Field, C.B.,
Dazlich, D.A., Zhang, C., Collelo, G.D., Bounoua, L., 1996. A
revised land surface parameterization (SIB2) for atmospheric
GCMs: Part I. Model formulation. J. Clim. 9, 676–705.
Shabanov, N.V., Wang, Y., Buermann, W., Dong, J., Hoffman, S.,
Smith, G.R., Tian, Y., Knyazikhin, Y., Myneni, R.B., 2003. Effect
of foliage spatial heterogeneity in the MODIS LAI and FPAR
algorithm over broadleaf forests. Rem. Sens. Environ. 85, 410–
423.
Shapiro, S.S., Wilk, M.B., Chen, H.J., 1968. A comparative study of
various tests of normality. J. Am. Stat. Assoc. 63, 1343–1372.
St-Onge, B., Treitz, P., Wulder, M., 2003. Tree and canopy height
estimation with scanning lidar (light detection and ranging). In:
Franklin, S., Wulder, M. (Eds.), Remote Sensing of Forest Envir-
onments: Concepts and Case Studies. Kluwar Academic
Publishers, pp. 489–509.
Thomas, V., Treitz, P., McCaughey, J.H., Morrison, I., 2006. Mapping
stand-level forest biophysical variables for a mixedwood boreal
forest using lidar: an examination of scanning density. Can. J. For.
Res. 36, 34–47.
Todd, K.W., Csillag, F., Atkinson, P.M., 2003. Three-dimensional
mapping of light transmittance and foliage distribution using lidar.
Can. J. Rem. Sens. 29, 544–555.
Waring, R.H., Law, B.E., Goulden, M.L., Bassow, S.L., McCreight,
R.W., Wofsy, S.C., Bazzaz, F.A., 1995. Scaling gross ecosystem
production at Harvard Forest with remote sensing: a comparison of
estimates from a constrained quantum use efficiency model and
eddy correlation. Plant Cell Environ. 18, 1201–1213.
Wehr, A., Lohr, U., 1999. Airborne laser scanning—an introduction
and overview. ISPRS J. Photogram. Rem. Sens. 54, 68–82.
Yoder, B.J., Pettigrew-Crosby, R.E., 1995. Predicting nitrogen and
chlorophyll content and concentrations from reflectance spectra
(400–2500 nm) at leaf and canopy scales. Rem. Sens. Environ. 53,
199–211.
Zarco-Tejada, P.J., Miller, J.R., Noland, T.L., Mohammad, G.H.,
Sampson, P.H., 2000. Chlorophyll fluorescence effects on vegeta-
tion apparent reflectance: II laboratory and airborne canopy-level
measurements with hyperspectral data. Rem. Sens. Environ. 74,
596–608.
Zarco-Tejada, P.J., Miller, J.R., Noland, T.L., Mohammad, G.H.,
Sampson, P.H., 2001. Scaling-up and model inversion methods
with narrowband optical indices for chlorophyll estimation in
closed forest canopies with hyperspectral data. IEEE Trans.
Geosci. Rem. Sens. 39, 1491–1507.
Copyright © 2022 FDOKUMEN



















