
Small Nuclear RING Finger Protein Stimulates the Rat Luteinizing Hormone Promoter by Interacting...
14
Small Nuclear RING Finger Protein Stimulates the Rat Luteinizing Hormone- Promoter by Interacting with Sp1 and Steroidogenic Factor-1 and Protects from Androgen Suppression DENIS CURTIN, HEATHER A. FERRIS, MARIKA HA ¨ KLI, MATTHEW GIBSON, OLLI A. JA ¨ NNE, JORMA J. PALVIMO, AND MARGARET A. SHUPNIK Departments of Pharmacology (D.C.), Physiology (H.A.F.), and Internal Medicine (M.G., M.A.S.), University of Virginia, Charlottesville, Virginia 22908; and Institute of Biomedicine (M.H., O.A.J., J.J.P.) and Department of Clinical Chemistry (O.A.J.), University of Helsinki and Helsinki University Central Hospital, FIN-00014 Helsinki, Finland GnRH controls expression of the LH subunit genes, and LH, with the LH subunit regulated most dramatically. Two enhancer regions, distal and proximal, on the rat LH gene promoter cooperate for full basal expression and GnRH stimulation. It has been hypothesized that the transcription fac- tors binding to these regions, Sp1, Egr-1, and ste- roidogenic factor 1 (SF-1), may interact directly or indirectly via a coactivator. One such coactivator may be small nuclear RING finger protein (SNURF), which is expressed in pituitary tissue and the LT2 gonadotrope cell line. In transfection experiments in LT2 cells, SNURF stimulated basal expression of LH and increased overall GnRH stimulation. SNURF specifically stimulated LH, with no effect on the -subunit promoter. SNURF interacts with Sp1 and SF-1, but not Egr-1, in pull-down experi- ments. Point mutations or deletions of SNURF functional domains demonstrated that Sp1 and SF-1 interactions with SNURF are required for SNURF stimulatory effects on the LH promoter. Endogenous SNURF is associated with the LH promoter on native chromatin, suggesting that it plays a physiological role in LH gene expression. SNURF also binds the androgen receptor, and SNURF overexpression overcomes androgen suppression of GnRH-stimulated LH but not - subunit promoter activity. SNURF mutations that disrupt Sp1 or SF-1 binding eliminate rescue by SNURF. We conclude that SNURF may mediate interactions between the distal and proximal GnRH response regions of the LH promoter to stimulate transcription and can also protect the promoter from androgen suppression. (Molecular Endocrin- ology 18: 1263–1276, 2004) R EGULATION OF NORMAL mammalian sexual maturation and reproductive cycles are governed by the hypothalamic-pituitary-gonadal axis (1). Hypo- thalamic GnRH acts on the pituitary gland to regulate secretion and synthesis of the gonadotropins, FSH and LH. LH acts on the ovaries and testes to stimulate production of the sex steroids estrogen, testosterone, and progesterone. LH is regulated by both GnRH pulses and steroid feedback at the hypothalamus and pituitary. LH consists of two subunits, LH and -subunit, and expression of LH is limiting for overall LH syn- thesis. GnRH pulses tightly regulate expression of the LH subunit genes, and only intermediate frequency pulses stimulate LH expression (2). Two enhancer regions of the rat LH promoter are required for com- plete basal expression and GnRH stimulation, includ- ing a distal region comprised of an overlapping Sp1/ CArG box and second Sp1 site, and a proximal region with two bipartite binding sites for Egr-1 and the orphan nuclear receptor steroidogenic factor 1 (SF-1) separated by a homeodomain binding site that corre- sponds to a Ptx1 consensus site (3–9). GnRH treat- ment of clonal gonadotrope cell lines or rat pituitary cells increases expression of SF-1 and Egr-1 and may increase Egr-1 transcriptional activity (5, 10–13). Expression of Egr-1 and SF-1 are critical for fertility and development as well as for expression of LH, and a complicated relationship exists between the Sp1, SF-1, and Egr-1 sites on the LH promoter (3, 6, 14, 15). Mutations of one but not both Sp1 binding sites in the distal GnRH responsive region of the LH pro- moter severely reduces GnRH stimulation of the pro- moter, even though the proximal region is still intact (3). In addition, a change in the number of bases Abbreviations: AR, Androgen receptor; CBP, CREB-bind- ing protein; ChIP, chromatin immunoprecipitation; CMV, cy- tomegalovirus; CRE, cAMP response element; DBD, DNA- binding domain; DHT, dihydrotestosterone; ER, estrogen receptor; GST, glutathione-S-transferase; nt, nucleotide; SDS, sodium dodecyl sulfate; SF-1, steroidogenic factor 1; SNURF, small nuclear RING finger; TBP, TATA-binding pro- tein; Tc, threshold cycle. Molecular Endocrinology is published monthly by The Endocrine Society (http://www.endo-society.org), the foremost professional society serving the endocrine community. 0888-8809/04/$15.00/0 Molecular Endocrinology 18(5):1263–1276 Printed in U.S.A. Copyright © 2004 by The Endocrine Society doi: 10.1210/me.2003-0221 1263
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Small Nuclear RING Finger Protein Stimulates the Rat Luteinizing Hormone Promoter by Interacting...

Small Nuclear RING Finger Protein Stimulates theRat Luteinizing Hormone-� Promoter by Interactingwith Sp1 and Steroidogenic Factor-1 and Protectsfrom Androgen Suppression
DENIS CURTIN, HEATHER A. FERRIS, MARIKA HAKLI, MATTHEW GIBSON, OLLI A. JANNE,JORMA J. PALVIMO, AND MARGARET A. SHUPNIK
Departments of Pharmacology (D.C.), Physiology (H.A.F.), and Internal Medicine (M.G., M.A.S.),University of Virginia, Charlottesville, Virginia 22908; and Institute of Biomedicine (M.H., O.A.J.,J.J.P.) and Department of Clinical Chemistry (O.A.J.), University of Helsinki and Helsinki UniversityCentral Hospital, FIN-00014 Helsinki, Finland
GnRH controls expression of the LH subunit genes,� and LH�, with the LH� subunit regulated mostdramatically. Two enhancer regions, distal andproximal, on the rat LH� gene promoter cooperatefor full basal expression and GnRH stimulation. Ithas been hypothesized that the transcription fac-tors binding to these regions, Sp1, Egr-1, and ste-roidogenic factor 1 (SF-1), may interact directly orindirectly via a coactivator. One such coactivatormay be small nuclear RING finger protein (SNURF),which is expressed in pituitary tissue and the L�T2gonadotrope cell line. In transfection experimentsin L�T2 cells, SNURF stimulated basal expressionof LH� and increased overall GnRH stimulation.SNURF specifically stimulated LH�, with no effecton the �-subunit promoter. SNURF interacts withSp1 and SF-1, but not Egr-1, in pull-down experi-ments. Point mutations or deletions of SNURF
functional domains demonstrated that Sp1 andSF-1 interactions with SNURF are required forSNURF stimulatory effects on the LH� promoter.Endogenous SNURF is associated with the LH�promoter on native chromatin, suggesting that itplays a physiological role in LH� gene expression.SNURF also binds the androgen receptor, andSNURF overexpression overcomes androgensuppression of GnRH-stimulated LH� but not �-subunit promoter activity. SNURF mutations thatdisrupt Sp1 or SF-1 binding eliminate rescue bySNURF. We conclude that SNURF may mediateinteractions between the distal and proximal GnRHresponse regions of the LH� promoter to stimulatetranscription and can also protect the promoterfrom androgen suppression. (Molecular Endocrin-ology 18: 1263–1276, 2004)
REGULATION OF NORMAL mammalian sexualmaturation and reproductive cycles are governed
by the hypothalamic-pituitary-gonadal axis (1). Hypo-thalamic GnRH acts on the pituitary gland to regulatesecretion and synthesis of the gonadotropins, FSHand LH. LH acts on the ovaries and testes to stimulateproduction of the sex steroids estrogen, testosterone,and progesterone. LH is regulated by both GnRHpulses and steroid feedback at the hypothalamus andpituitary.
LH consists of two subunits, LH� and �-subunit,and expression of LH� is limiting for overall LH syn-
thesis. GnRH pulses tightly regulate expression of theLH subunit genes, and only intermediate frequencypulses stimulate LH� expression (2). Two enhancerregions of the rat LH� promoter are required for com-plete basal expression and GnRH stimulation, includ-ing a distal region comprised of an overlapping Sp1/CArG box and second Sp1 site, and a proximal regionwith two bipartite binding sites for Egr-1 and theorphan nuclear receptor steroidogenic factor 1 (SF-1)separated by a homeodomain binding site that corre-sponds to a Ptx1 consensus site (3–9). GnRH treat-ment of clonal gonadotrope cell lines or rat pituitarycells increases expression of SF-1 and Egr-1 and mayincrease Egr-1 transcriptional activity (5, 10–13).
Expression of Egr-1 and SF-1 are critical for fertilityand development as well as for expression of LH, anda complicated relationship exists between the Sp1,SF-1, and Egr-1 sites on the LH� promoter (3, 6, 14,15). Mutations of one but not both Sp1 binding sites inthe distal GnRH responsive region of the LH� pro-moter severely reduces GnRH stimulation of the pro-moter, even though the proximal region is still intact(3). In addition, a change in the number of bases
Abbreviations: AR, Androgen receptor; CBP, CREB-bind-ing protein; ChIP, chromatin immunoprecipitation; CMV, cy-tomegalovirus; CRE, cAMP response element; DBD, DNA-binding domain; DHT, dihydrotestosterone; ER, estrogenreceptor; GST, glutathione-S-transferase; nt, nucleotide;SDS, sodium dodecyl sulfate; SF-1, steroidogenic factor 1;SNURF, small nuclear RING finger; TBP, TATA-binding pro-tein; Tc, threshold cycle.
Molecular Endocrinology is published monthly by TheEndocrine Society (http://www.endo-society.org), theforemost professional society serving the endocrinecommunity.
0888-8809/04/$15.00/0 Molecular Endocrinology 18(5):1263–1276Printed in U.S.A. Copyright © 2004 by The Endocrine Society
doi: 10.1210/me.2003-0221
1263

between the distal and proximal regions attenuatesGnRH stimulation (6). These data have led to a hy-pothesis that the DNA between the two responsiveregions of the LH� promoter may enable either a directinteraction between the transcription factors bindingto these regions, or an indirect one mediated by acoactivator (3, 6, 16).
Coactivators are generally divided into two catego-ries based on function: proteins that direct and recruitthe transcription complex, and factors, such as TATA-binding protein (TBP)-associated factors, and en-zymes that modify the structure of chromatin includingthose with histone acetyltransferase activity (17). Inconsidering possible coactivators that may play a rolein basal and GnRH-stimulated expression of the LH�
gene, we tested coactivators from these two generalclasses. We examined examples of chromatin remod-eling coactivators with histone acetyltransferase activ-ity, CREB binding protein (CBP) and p300, and thenovel coactivator small nuclear RING finger protein(SNURF) that may promote the assembly of the tran-scription complex (18).
SNURF was identified in yeast two-hybrid assaysusing the androgen receptor (AR) DNA binding domain(DBD) and part of the hinge region as bait (18). SNURFwas found to serve as a coactivator for steroid recep-tor-dependent and independent promoters, poten-tially through binding to TBP and acting as a bridgebetween transcription factors and the transcriptioncomplex. Overexpression of SNURF enhances notonly AR-dependent transcription but also that of othersteroid receptors, including the glucocorticoid, pro-gesterone, and estrogen receptors (18, 19). SNURFalso enhances transcription of promoters containingGC box elements (20, 21). The RING finger domain ofSNURF is critical for its interaction with Sp1 and ac-tivation of the latter promoters (18, 21). In addition, thepositively charged N-terminal amino acid residues ofSNURF that enable direct interaction with DNA arelinked with the ability of SNURF to enhance transcrip-tion of minimal promoters containing Sp1 binding sites(20). Because the Sp1 sites in the LH� promoter arecritical for both basal and GnRH-stimulated expres-sion, we investigated the role of SNURF in LH� ex-pression. The ability of SNURF to bind AR was alsointriguing, as androgens suppress GnRH stimulationof the rat and bovine LH� promoters (16, 22).
Steroid feedback has been implicated in the regu-lation of gonadotropins at the hypothalamic and pitu-itary levels (16, 22–27). Androgens can regulate GnRHexpression at the level of the hypothalamus (23) andsuppress transcription of the LH subunit genes, LH�
and �-subunit, in vivo (24). This suppression occurs, atleast in part, by direct androgen actions on the pitu-itary, but not necessarily by direct AR binding to DNA.For example, AR suppresses basal transcription of thehuman �-subunit by AR interference with binding ofactivating transcription factor 1 and c-Jun to the pro-
moter, rather than binding to the androgen responseelement (27, 28). Similarly, we and others have shownthat AR is required for androgen suppression of GnRHstimulation of the rat and bovine LH� genes (16, 22).AR interferes with transcription factors required for fullGnRH stimulation of LH� and thereby represses tran-scription of the LH� promoter (16, 22).
Here we investigated the role that SNURF plays inthe pituitary gonadotrope L�T2 cell line and, in partic-ular, how it activates the Sp1-driven rat LH� promoter.Our results indicate that SNURF is a coactivator for theLH� gene and interacts with transcription factorsfound in both GnRH-responsive regions of the LH�
promoter. Through these interactions, SNURF canalso prevent AR from disrupting the recruitment of thetranscription complex after androgen treatment.
RESULTS
The Coactivator Activity of SNURF Is Specific forthe LH� Promoter and Increases Both Basal andGnRH-Stimulated Promoter Activity
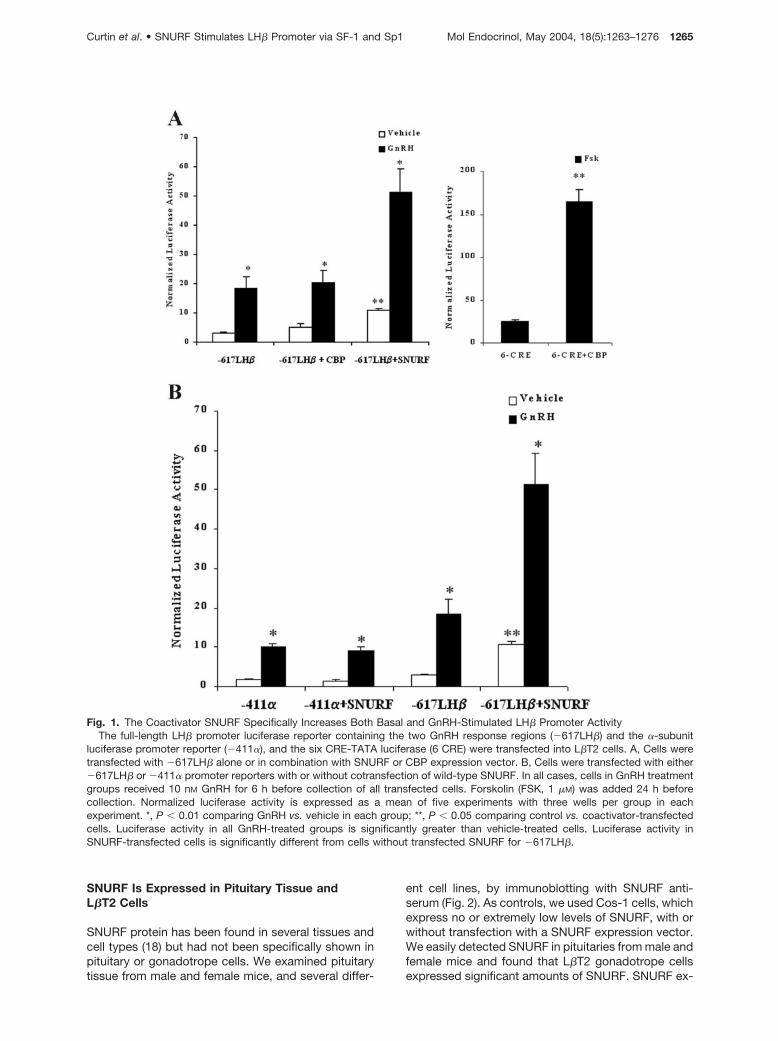
To investigate whether the coactivators CBP orSNURF could activate the promoter for the LH� geneand to determine the specificity of this activation, weperformed transfection studies using the mouse pitu-itary gonadotrope cell line, L�T2. These cells secreteLH and express both the LH� and �-subunit genes, aswell as receptors for GnRH, estrogen, and androgen(29). Cells were transfected with the coactivators CBPor SNURF and the rat LH� promoter construct con-taining the two GnRH-responsive regions of the LH�
promoter [�617 to �44 nucleotides (nt), hereaftertermed �617LH�] fused to luciferase. Overexpressionof SNURF increased both untreated (3.0-fold) andGnRH-treated (3.2-fold) LH� reporter activity whencompared with reporter alone (Fig. 1A). Overexpres-sion of CBP had no significant effect on basal orGnRH-stimulated LH� reporter activity in agreementwith previous studies (6), and similar experiments withp300 did not alter LH� promoter activity (data notshown). CBP was functional in these studies, however,as in parallel wells the forskolin-stimulated expressionof a cAMP response element (CRE)-containing re-porter was stimulated more than 5-fold (Fig. 1A).
To test the specificity of the SNURF response on theLH subunit genes, cells were transfected with a lucif-erase reporter construct containing the �411 to �77nt of the �-subunit promoter or �617LH� luciferasereporter construct alone or in combination with theSNURF expression plasmid. Both �-subunit and LH�
promoter reporters were stimulated more than 5-foldby GnRH treatment (Fig. 1B). However, only LH� pro-moter activity was stimulated for both basal and GnRHresponses when cotransfected with SNURF.
1264 Mol Endocrinol, May 2004, 18(5):1263–1276 Curtin et al. • SNURF Stimulates LH� Promoter via SF-1 and Sp1
SNURF Is Expressed in Pituitary Tissue andL�T2 Cells
SNURF protein has been found in several tissues andcell types (18) but had not been specifically shown inpituitary or gonadotrope cells. We examined pituitarytissue from male and female mice, and several differ-
ent cell lines, by immunoblotting with SNURF anti-serum (Fig. 2). As controls, we used Cos-1 cells, whichexpress no or extremely low levels of SNURF, with orwithout transfection with a SNURF expression vector.We easily detected SNURF in pituitaries from male andfemale mice and found that L�T2 gonadotrope cellsexpressed significant amounts of SNURF. SNURF ex-
Fig. 1. The Coactivator SNURF Specifically Increases Both Basal and GnRH-Stimulated LH� Promoter ActivityThe full-length LH� promoter luciferase reporter containing the two GnRH response regions (�617LH�) and the �-subunit
luciferase promoter reporter (�411�), and the six CRE-TATA luciferase (6 CRE) were transfected into L�T2 cells. A, Cells weretransfected with �617LH� alone or in combination with SNURF or CBP expression vector. B, Cells were transfected with either�617LH� or �411� promoter reporters with or without cotransfection of wild-type SNURF. In all cases, cells in GnRH treatmentgroups received 10 nM GnRH for 6 h before collection of all transfected cells. Forskolin (FSK, 1 �M) was added 24 h beforecollection. Normalized luciferase activity is expressed as a mean of five experiments with three wells per group in eachexperiment. *, P � 0.01 comparing GnRH vs. vehicle in each group; **, P � 0.05 comparing control vs. coactivator-transfectedcells. Luciferase activity in all GnRH-treated groups is significantly greater than vehicle-treated cells. Luciferase activity inSNURF-transfected cells is significantly different from cells without transfected SNURF for �617LH�.
Curtin et al. • SNURF Stimulates LH� Promoter via SF-1 and Sp1 Mol Endocrinol, May 2004, 18(5):1263–1276 1265

pression was not altered by GnRH treatment. GH3somatolactotrope cells expressed no or very low levelsof SNURF. Although there was some variation inSNURF expression levels among individual pituitariesexamined, there were no significant differences be-tween male and female glands. In L�T2 cells, andindividual pituitary glands with higher levels of SNURFexpression, there were often fainter bands above andbelow the main SNURF band. This could representsplice variants or other modifications of SNURF, butthese have not yet been identified. These data showthat SNURF is expressed in pituitary and the L�T2gonadotrope cells and could play a physiological rolein LH� expression.
Distal and Proximal GnRH-Responsive RegionsAre Required for the Full SNURF Response
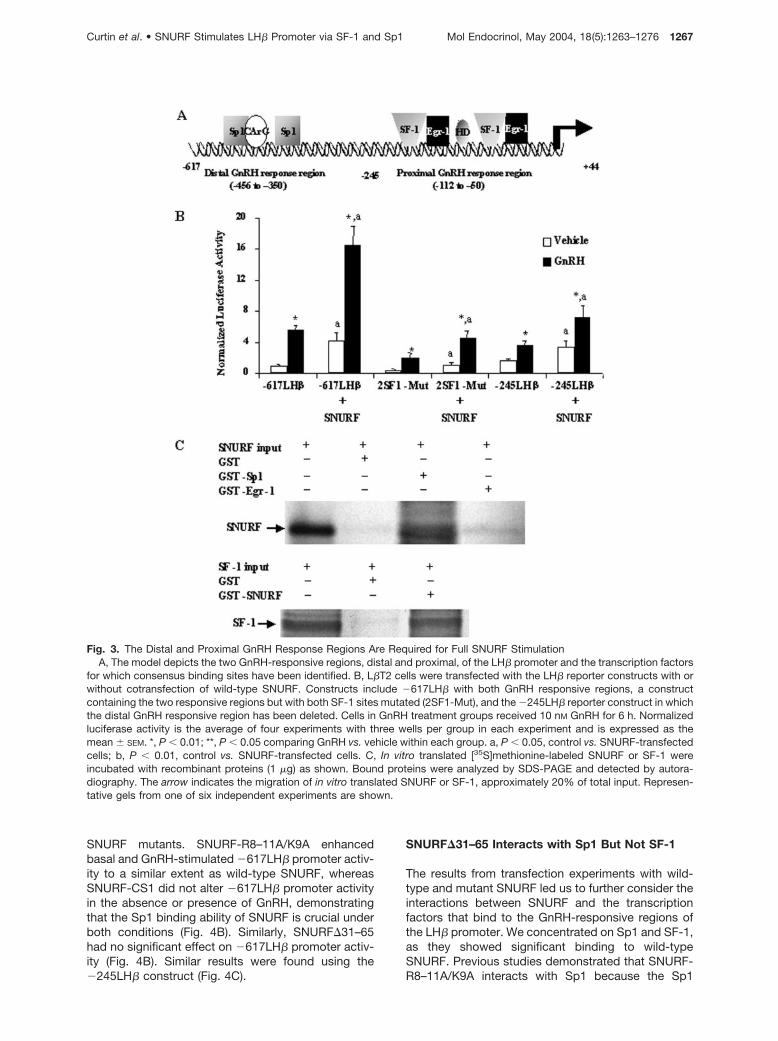
The LH� promoter has two cooperating enhancer re-gions that confer GnRH stimulation. These include thedistal GnRH-responsive region containing an overlap-ping Sp1/CArG site and an Sp1 consensus site, and aproximal GnRH-response region containing two bipar-tite Egr-1 and SF-1 binding sites separated by a ho-meodomain/Ptx-1 consensus site (Fig. 3A). These tworegions must cooperate for full GnRH stimulation (3,6). We performed transfection studies to identify therelationship between SNURF coactivation and the twoGnRH response regions. We used the �617LH� re-porter that contains both distal and proximal responseregions, and a truncated �245LH� reporter containingonly the proximal response region. To determine theimportance of the distal region in the SNURF re-sponse, a construct containing both response regionsbut with mutations in both SF-1 sites (2SF1-Mut) wasalso used. Transfection of SNURF leads to stimulationof each LH� promoter construct in both untreated and
GnRH-treated cells (Fig. 3B). As noted previously,GnRH stimulation of the �245LH� construct by GnRHis significantly less than stimulation of the larger con-struct (3). SNURF stimulation of basal (3.9-fold) andGnRH-stimulated (4-fold) activity of the �617LH� pro-moter containing both GnRH response regions wasgreater than SNURF stimulation of basal (2.1-fold) andGnRH-stimulated (2.0-fold) activity of the �245LH�promoter. SNURF stimulation of basal (2.5-fold) andGnRH-stimulated (2.3-fold) activity of the 2SF1-Mutconstruct was also less that that of the intact�617LH� promoter, suggesting that both Sp1 andSF-1 binding sites are required for the full response toSNURF.
SNURF Binds to Sp1 and SF-1
To address the possibility that SNURF may interactwith transcription factors that bind to the GnRH-re-sponsive regions of the LH� promoter, we performeda series of glutathione-S-transferase (GST) pull-downexperiments. SNURF has previously been shown tointeract with Sp1 by immunoprecipitation (21), and wedemonstrate this interaction using in vitro translated,35S-methionine-labeled SNURF and recombinantGST-Sp1 (Fig. 3C). Two transcription factors that bindto the proximal GnRH response region, Egr-1 andSF-1, were also used in pull-down experiments.SNURF interacted with GST-Egr-1 very weakly if at all,whereas GST-SNURF showed a strong interactionwith in vitro translated 35S-labeled SF-1 (Fig. 3C).Thus, SNURF interacts directly with transcription fac-tors that bind to both proximal and distal GnRH re-sponse regions of the LH� promoter.
SNURF Interaction with Sp1 Is Important forCoactivator Activity
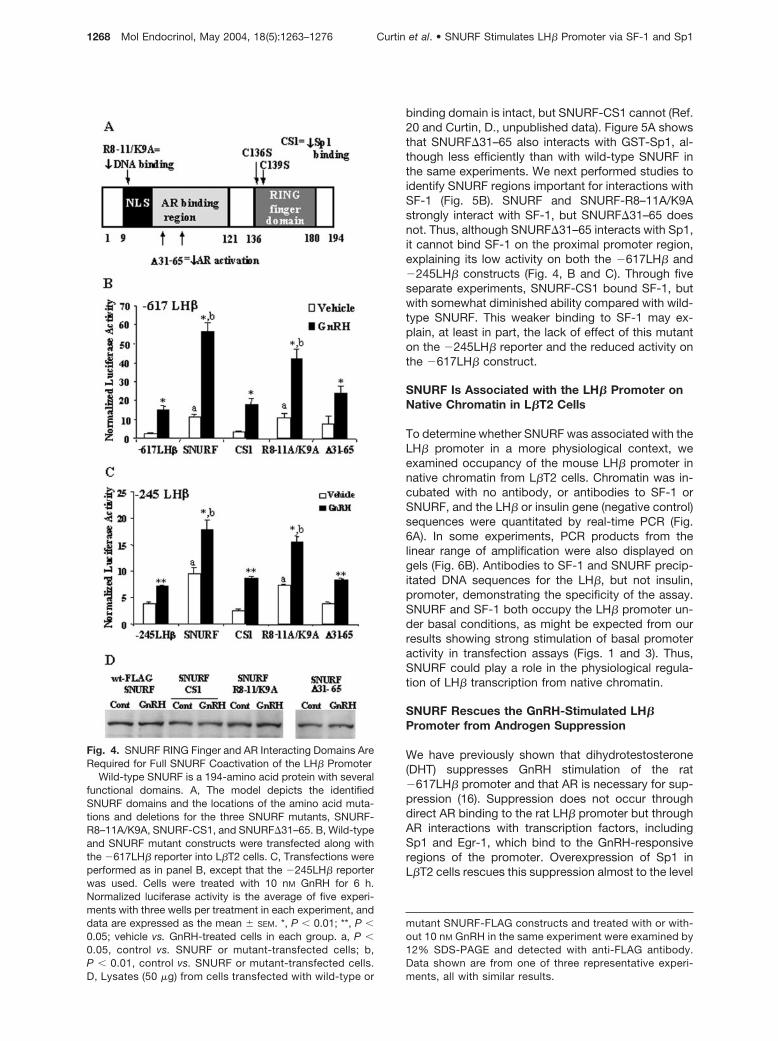
SNURF can activate steroid-dependent and -indepen-dent gene transcription through its interaction withsteroid receptors and Sp1 (18, 19, 21). It has also beenshown to bind DNA directly through several positivelycharged amino acid residues in its amino terminus(20). To test which of these characteristics of SNURFis most important in enhancing basal and GnRH-stim-ulated LH� promoter activity, we used several mutantsof SNURF (Fig. 4A) in transfection assays. These in-cluded SNURF-CS1, in which the RING finger domainand Sp1 binding has been disrupted by amino acidsubstitutions C136S and C139S; SNURF-R8–11A/K9A, in which the positively charged amino acids ofthe N terminus of SNURF have been changed to ala-nines to eliminate DNA binding; and SNURF�31–65, inwhich a portion of the AR interacting domain has beendeleted (18, 20, 21). All SNURF mutants were ex-pressed at levels equivalent to levels of wild-typeSNURF in transfected cells (Fig. 4D). The �617LH�and �245LH� promoter reporters were used in trans-fection experiments in L�T2 cells along with the
Fig. 2. Immunoblot of SNURF in Pituitary Tissue and ClonalCell Lines
Approximately 25 �g of total cell protein from female (F Pit)or male (M Pit) mouse pituitary, L�T2 cells treated for 2 h withvehicle (Con) or 100 nM GnRH (GnRH), GH3 cells, untrans-fected Cos-1 cells (Cos), or Cos-1 cells transfected with aSNURF expression vector (Cos �SNURF) were subjected toelectrophoresis on 10% SDS-PAGE. SNURF and �-actin (po-sition indicated by arrows on the right of each panel) weredetected sequentially, using the same blot. Migration of the29-kDa molecular mass marker is indicated by an arrow onthe left of the upper SNURF panel. A representative gel fromfour independent experiments is shown.
1266 Mol Endocrinol, May 2004, 18(5):1263–1276 Curtin et al. • SNURF Stimulates LH� Promoter via SF-1 and Sp1
SNURF mutants. SNURF-R8–11A/K9A enhancedbasal and GnRH-stimulated �617LH� promoter activ-ity to a similar extent as wild-type SNURF, whereasSNURF-CS1 did not alter �617LH� promoter activityin the absence or presence of GnRH, demonstratingthat the Sp1 binding ability of SNURF is crucial underboth conditions (Fig. 4B). Similarly, SNURF�31–65had no significant effect on �617LH� promoter activ-ity (Fig. 4B). Similar results were found using the�245LH� construct (Fig. 4C).
SNURF�31–65 Interacts with Sp1 But Not SF-1
The results from transfection experiments with wild-type and mutant SNURF led us to further consider theinteractions between SNURF and the transcriptionfactors that bind to the GnRH-responsive regions ofthe LH� promoter. We concentrated on Sp1 and SF-1,as they showed significant binding to wild-typeSNURF. Previous studies demonstrated that SNURF-R8–11A/K9A interacts with Sp1 because the Sp1
Fig. 3. The Distal and Proximal GnRH Response Regions Are Required for Full SNURF StimulationA, The model depicts the two GnRH-responsive regions, distal and proximal, of the LH� promoter and the transcription factors
for which consensus binding sites have been identified. B, L�T2 cells were transfected with the LH� reporter constructs with orwithout cotransfection of wild-type SNURF. Constructs include �617LH� with both GnRH responsive regions, a constructcontaining the two responsive regions but with both SF-1 sites mutated (2SF1-Mut), and the �245LH� reporter construct in whichthe distal GnRH responsive region has been deleted. Cells in GnRH treatment groups received 10 nM GnRH for 6 h. Normalizedluciferase activity is the average of four experiments with three wells per group in each experiment and is expressed as themean � SEM. *, P � 0.01; **, P � 0.05 comparing GnRH vs. vehicle within each group. a, P � 0.05, control vs. SNURF-transfectedcells; b, P � 0.01, control vs. SNURF-transfected cells. C, In vitro translated [35S]methionine-labeled SNURF or SF-1 wereincubated with recombinant proteins (1 �g) as shown. Bound proteins were analyzed by SDS-PAGE and detected by autora-diography. The arrow indicates the migration of in vitro translated SNURF or SF-1, approximately 20% of total input. Represen-tative gels from one of six independent experiments are shown.
Curtin et al. • SNURF Stimulates LH� Promoter via SF-1 and Sp1 Mol Endocrinol, May 2004, 18(5):1263–1276 1267
binding domain is intact, but SNURF-CS1 cannot (Ref.20 and Curtin, D., unpublished data). Figure 5A showsthat SNURF�31–65 also interacts with GST-Sp1, al-though less efficiently than with wild-type SNURF inthe same experiments. We next performed studies toidentify SNURF regions important for interactions withSF-1 (Fig. 5B). SNURF and SNURF-R8–11A/K9Astrongly interact with SF-1, but SNURF�31–65 doesnot. Thus, although SNURF�31–65 interacts with Sp1,it cannot bind SF-1 on the proximal promoter region,explaining its low activity on both the �617LH� and�245LH� constructs (Fig. 4, B and C). Through fiveseparate experiments, SNURF-CS1 bound SF-1, butwith somewhat diminished ability compared with wild-type SNURF. This weaker binding to SF-1 may ex-plain, at least in part, the lack of effect of this mutanton the �245LH� reporter and the reduced activity onthe �617LH� construct.
SNURF Is Associated with the LH� Promoter onNative Chromatin in L�T2 Cells
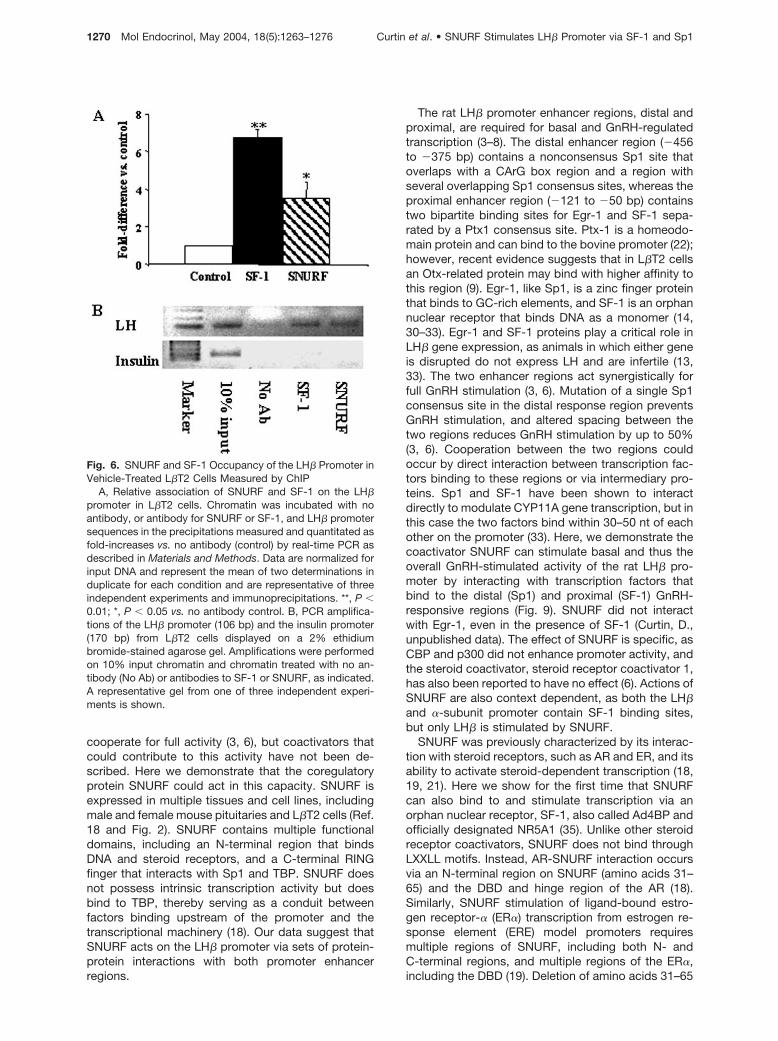
To determine whether SNURF was associated with theLH� promoter in a more physiological context, weexamined occupancy of the mouse LH� promoter innative chromatin from L�T2 cells. Chromatin was in-cubated with no antibody, or antibodies to SF-1 orSNURF, and the LH� or insulin gene (negative control)sequences were quantitated by real-time PCR (Fig.6A). In some experiments, PCR products from thelinear range of amplification were also displayed ongels (Fig. 6B). Antibodies to SF-1 and SNURF precip-itated DNA sequences for the LH�, but not insulin,promoter, demonstrating the specificity of the assay.SNURF and SF-1 both occupy the LH� promoter un-der basal conditions, as might be expected from ourresults showing strong stimulation of basal promoteractivity in transfection assays (Figs. 1 and 3). Thus,SNURF could play a role in the physiological regula-tion of LH� transcription from native chromatin.
SNURF Rescues the GnRH-Stimulated LH�Promoter from Androgen Suppression
We have previously shown that dihydrotestosterone(DHT) suppresses GnRH stimulation of the rat�617LH� promoter and that AR is necessary for sup-pression (16). Suppression does not occur throughdirect AR binding to the rat LH� promoter but throughAR interactions with transcription factors, includingSp1 and Egr-1, which bind to the GnRH-responsiveregions of the promoter. Overexpression of Sp1 inL�T2 cells rescues this suppression almost to the level
Fig. 4. SNURF RING Finger and AR Interacting Domains AreRequired for Full SNURF Coactivation of the LH� Promoter
Wild-type SNURF is a 194-amino acid protein with severalfunctional domains. A, The model depicts the identifiedSNURF domains and the locations of the amino acid muta-tions and deletions for the three SNURF mutants, SNURF-R8–11A/K9A, SNURF-CS1, and SNURF�31–65. B, Wild-typeand SNURF mutant constructs were transfected along withthe �617LH� reporter into L�T2 cells. C, Transfections wereperformed as in panel B, except that the �245LH� reporterwas used. Cells were treated with 10 nM GnRH for 6 h.Normalized luciferase activity is the average of five experi-ments with three wells per treatment in each experiment, anddata are expressed as the mean � SEM. *, P � 0.01; **, P �0.05; vehicle vs. GnRH-treated cells in each group. a, P �0.05, control vs. SNURF or mutant-transfected cells; b,P � 0.01, control vs. SNURF or mutant-transfected cells.D, Lysates (50 �g) from cells transfected with wild-type or
mutant SNURF-FLAG constructs and treated with or with-out 10 nM GnRH in the same experiment were examined by12% SDS-PAGE and detected with anti-FLAG antibody.Data shown are from one of three representative experi-ments, all with similar results.
1268 Mol Endocrinol, May 2004, 18(5):1263–1276 Curtin et al. • SNURF Stimulates LH� Promoter via SF-1 and Sp1

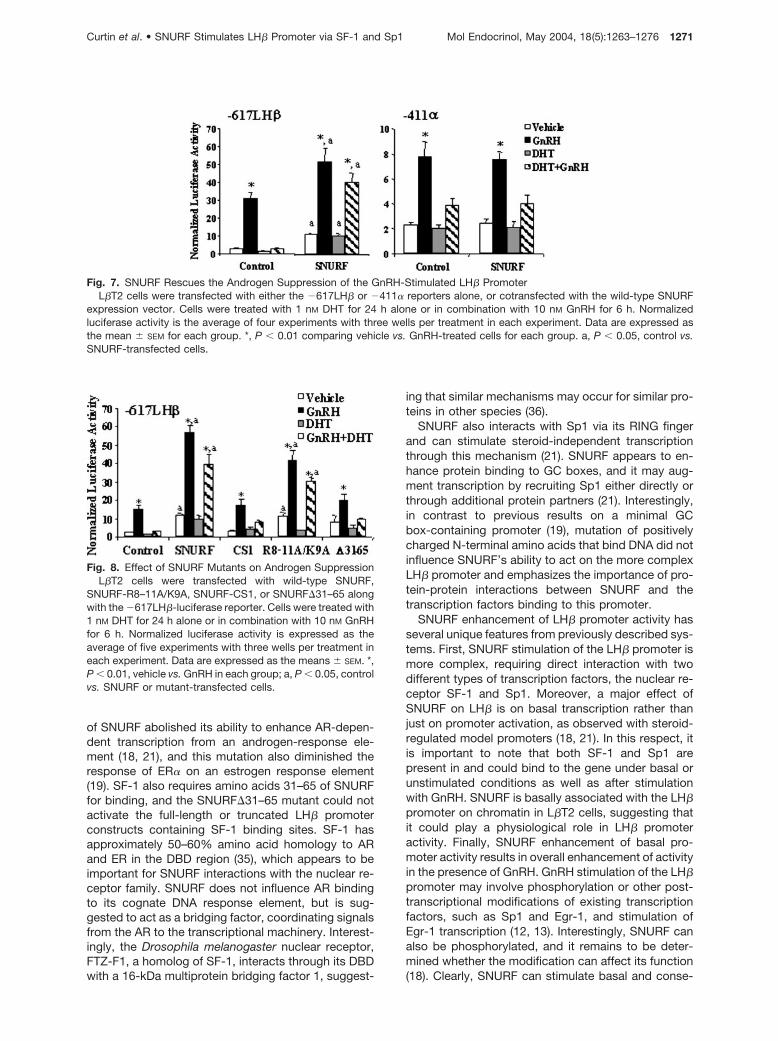
of GnRH treatment alone (16). Because SNURF inter-acts with AR via yeast two-hybrid and GST pull-downassays (18, 21), we tested its influence in androgenmodulation of the promoter. Interestingly, transfectionof SNURF into cells treated with GnRH and DHT re-stored �617LH� promoter activity to a level that ap-proximated GnRH treatment alone (Fig. 7). The relativelevels of AR and SNURF were important, as overex-pression of AR resulted in lower GnRH stimulationeven in the presence of SNURF (data not shown).GnRH stimulation of the rat �-subunit promoter is alsosuppressed by androgens, but SNURF cannot rescuethe promoter from androgen suppression, suggestingthat simple SNURF titration of AR does not completelyexplain the results with the LH� promoter.
To determine which region of SNURF was requiredto rescue the LH� promoter from androgen suppres-sion, we examined the effects of SNURF mutants inL�T2 cells treated with GnRH or GnRH plus DHT.Transfection of L�T2 cells with either wild-typeSNURF or SNURF-R8–11A/K9A rescued GnRH-stim-ulated LH� promoter activity from DHT (Fig. 8). Neither
SNURF-CS1 nor SNURF�31–65 was able to signifi-cantly stimulate promoter activity in cells treated withboth GnRH and DHT (Fig. 8), suggesting that both Sp1and SF-1 binding were required for this effect. On thebasis of these data, a model for the relationships be-tween SNURF, transcription factor binding, and AR onthe LH� promoter is presented in Fig. 9.
DISCUSSION
Coactivators serve a variety of functions in mediatingtranscription and include proteins that are compo-nents of the basal transcription machinery, those thatcontact the basal transcription machinery, and thosethat modify chromatin. TBP-associated factors, ATP-dependent nucleosome remodeling complexes (SWI/SNF), and histone acetyltransferases (CBP/p300,p160 family coactivators) represent the diversity ofcoactivator activity (reviewed in Ref. 17). The rat LH�promoter contains two enhancer regions that must
Fig. 5. SNURF�31–65 Interacts with Sp1 But Not SF-1A, In vitro translated [35S]methionine-labeled wild-type SNURF and SNURF�31–65 were incubated with 1 �g of recombinant
GST or GST-Sp1. Bound proteins are shown on the autoradiogram. The arrow indicates the migration of input proteins SNURFor SNURF�31–65, approximately 20% of total input. B, In vitro translated [35S]methionine-labeled wild-type SNURF, SNURF-CS1, SNURF-R8–11A/K9A, and SNURF�31–65 were incubated with 1 �g of recombinant GST or GST-SF-1. Bound proteins areshown on the autoradiogram. The upper arrow indicates the migration of input proteins SNURF, SNURF-CS1, and SNURF-R8–11A/K9A. The lower arrow indicates input lane for SNURF�31–65. Representative gels from one of eight independent experimentsare shown.
Curtin et al. • SNURF Stimulates LH� Promoter via SF-1 and Sp1 Mol Endocrinol, May 2004, 18(5):1263–1276 1269
cooperate for full activity (3, 6), but coactivators thatcould contribute to this activity have not been de-scribed. Here we demonstrate that the coregulatoryprotein SNURF could act in this capacity. SNURF isexpressed in multiple tissues and cell lines, includingmale and female mouse pituitaries and L�T2 cells (Ref.18 and Fig. 2). SNURF contains multiple functionaldomains, including an N-terminal region that bindsDNA and steroid receptors, and a C-terminal RINGfinger that interacts with Sp1 and TBP. SNURF doesnot possess intrinsic transcription activity but doesbind to TBP, thereby serving as a conduit betweenfactors binding upstream of the promoter and thetranscriptional machinery (18). Our data suggest thatSNURF acts on the LH� promoter via sets of protein-protein interactions with both promoter enhancerregions.
The rat LH� promoter enhancer regions, distal andproximal, are required for basal and GnRH-regulatedtranscription (3–8). The distal enhancer region (�456to �375 bp) contains a nonconsensus Sp1 site thatoverlaps with a CArG box region and a region withseveral overlapping Sp1 consensus sites, whereas theproximal enhancer region (�121 to �50 bp) containstwo bipartite binding sites for Egr-1 and SF-1 sepa-rated by a Ptx1 consensus site. Ptx-1 is a homeodo-main protein and can bind to the bovine promoter (22);however, recent evidence suggests that in L�T2 cellsan Otx-related protein may bind with higher affinity tothis region (9). Egr-1, like Sp1, is a zinc finger proteinthat binds to GC-rich elements, and SF-1 is an orphannuclear receptor that binds DNA as a monomer (14,30–33). Egr-1 and SF-1 proteins play a critical role inLH� gene expression, as animals in which either geneis disrupted do not express LH and are infertile (13,33). The two enhancer regions act synergistically forfull GnRH stimulation (3, 6). Mutation of a single Sp1consensus site in the distal response region preventsGnRH stimulation, and altered spacing between thetwo regions reduces GnRH stimulation by up to 50%(3, 6). Cooperation between the two regions couldoccur by direct interaction between transcription fac-tors binding to these regions or via intermediary pro-teins. Sp1 and SF-1 have been shown to interactdirectly to modulate CYP11A gene transcription, but inthis case the two factors bind within 30–50 nt of eachother on the promoter (33). Here, we demonstrate thecoactivator SNURF can stimulate basal and thus theoverall GnRH-stimulated activity of the rat LH� pro-moter by interacting with transcription factors thatbind to the distal (Sp1) and proximal (SF-1) GnRH-responsive regions (Fig. 9). SNURF did not interactwith Egr-1, even in the presence of SF-1 (Curtin, D.,unpublished data). The effect of SNURF is specific, asCBP and p300 did not enhance promoter activity, andthe steroid coactivator, steroid receptor coactivator 1,has also been reported to have no effect (6). Actions ofSNURF are also context dependent, as both the LH�and �-subunit promoter contain SF-1 binding sites,but only LH� is stimulated by SNURF.
SNURF was previously characterized by its interac-tion with steroid receptors, such as AR and ER, and itsability to activate steroid-dependent transcription (18,19, 21). Here we show for the first time that SNURFcan also bind to and stimulate transcription via anorphan nuclear receptor, SF-1, also called Ad4BP andofficially designated NR5A1 (35). Unlike other steroidreceptor coactivators, SNURF does not bind throughLXXLL motifs. Instead, AR-SNURF interaction occursvia an N-terminal region on SNURF (amino acids 31–65) and the DBD and hinge region of the AR (18).Similarly, SNURF stimulation of ligand-bound estro-gen receptor-� (ER�) transcription from estrogen re-sponse element (ERE) model promoters requiresmultiple regions of SNURF, including both N- andC-terminal regions, and multiple regions of the ER�,including the DBD (19). Deletion of amino acids 31–65
Fig. 6. SNURF and SF-1 Occupancy of the LH� Promoter inVehicle-Treated L�T2 Cells Measured by ChIP
A, Relative association of SNURF and SF-1 on the LH�promoter in L�T2 cells. Chromatin was incubated with noantibody, or antibody for SNURF or SF-1, and LH� promotersequences in the precipitations measured and quantitated asfold-increases vs. no antibody (control) by real-time PCR asdescribed in Materials and Methods. Data are normalized forinput DNA and represent the mean of two determinations induplicate for each condition and are representative of threeindependent experiments and immunoprecipitations. **, P �0.01; *, P � 0.05 vs. no antibody control. B, PCR amplifica-tions of the LH� promoter (106 bp) and the insulin promoter(170 bp) from L�T2 cells displayed on a 2% ethidiumbromide-stained agarose gel. Amplifications were performedon 10% input chromatin and chromatin treated with no an-tibody (No Ab) or antibodies to SF-1 or SNURF, as indicated.A representative gel from one of three independent experi-ments is shown.
1270 Mol Endocrinol, May 2004, 18(5):1263–1276 Curtin et al. • SNURF Stimulates LH� Promoter via SF-1 and Sp1
of SNURF abolished its ability to enhance AR-depen-dent transcription from an androgen-response ele-ment (18, 21), and this mutation also diminished theresponse of ER� on an estrogen response element(19). SF-1 also requires amino acids 31–65 of SNURFfor binding, and the SNURF�31–65 mutant could notactivate the full-length or truncated LH� promoterconstructs containing SF-1 binding sites. SF-1 hasapproximately 50–60% amino acid homology to ARand ER in the DBD region (35), which appears to beimportant for SNURF interactions with the nuclear re-ceptor family. SNURF does not influence AR bindingto its cognate DNA response element, but is sug-gested to act as a bridging factor, coordinating signalsfrom the AR to the transcriptional machinery. Interest-ingly, the Drosophila melanogaster nuclear receptor,FTZ-F1, a homolog of SF-1, interacts through its DBDwith a 16-kDa multiprotein bridging factor 1, suggest-
ing that similar mechanisms may occur for similar pro-teins in other species (36).
SNURF also interacts with Sp1 via its RING fingerand can stimulate steroid-independent transcriptionthrough this mechanism (21). SNURF appears to en-hance protein binding to GC boxes, and it may aug-ment transcription by recruiting Sp1 either directly orthrough additional protein partners (21). Interestingly,in contrast to previous results on a minimal GCbox-containing promoter (19), mutation of positivelycharged N-terminal amino acids that bind DNA did notinfluence SNURF’s ability to act on the more complexLH� promoter and emphasizes the importance of pro-tein-protein interactions between SNURF and thetranscription factors binding to this promoter.
SNURF enhancement of LH� promoter activity hasseveral unique features from previously described sys-tems. First, SNURF stimulation of the LH� promoter ismore complex, requiring direct interaction with twodifferent types of transcription factors, the nuclear re-ceptor SF-1 and Sp1. Moreover, a major effect ofSNURF on LH� is on basal transcription rather thanjust on promoter activation, as observed with steroid-regulated model promoters (18, 21). In this respect, itis important to note that both SF-1 and Sp1 arepresent in and could bind to the gene under basal orunstimulated conditions as well as after stimulationwith GnRH. SNURF is basally associated with the LH�promoter on chromatin in L�T2 cells, suggesting thatit could play a physiological role in LH� promoteractivity. Finally, SNURF enhancement of basal pro-moter activity results in overall enhancement of activityin the presence of GnRH. GnRH stimulation of the LH�promoter may involve phosphorylation or other post-transcriptional modifications of existing transcriptionfactors, such as Sp1 and Egr-1, and stimulation ofEgr-1 transcription (12, 13). Interestingly, SNURF canalso be phosphorylated, and it remains to be deter-mined whether the modification can affect its function(18). Clearly, SNURF can stimulate basal and conse-
Fig. 7. SNURF Rescues the Androgen Suppression of the GnRH-Stimulated LH� PromoterL�T2 cells were transfected with either the �617LH� or �411� reporters alone, or cotransfected with the wild-type SNURF
expression vector. Cells were treated with 1 nM DHT for 24 h alone or in combination with 10 nM GnRH for 6 h. Normalizedluciferase activity is the average of four experiments with three wells per treatment in each experiment. Data are expressed asthe mean � SEM for each group. *, P � 0.01 comparing vehicle vs. GnRH-treated cells for each group. a, P � 0.05, control vs.SNURF-transfected cells.
Fig. 8. Effect of SNURF Mutants on Androgen SuppressionL�T2 cells were transfected with wild-type SNURF,
SNURF-R8–11A/K9A, SNURF-CS1, or SNURF�31–65 alongwith the �617LH�-luciferase reporter. Cells were treated with1 nM DHT for 24 h alone or in combination with 10 nM GnRHfor 6 h. Normalized luciferase activity is expressed as theaverage of five experiments with three wells per treatment ineach experiment. Data are expressed as the means � SEM. *,P � 0.01, vehicle vs. GnRH in each group; a, P � 0.05, controlvs. SNURF or mutant-transfected cells.
Curtin et al. • SNURF Stimulates LH� Promoter via SF-1 and Sp1 Mol Endocrinol, May 2004, 18(5):1263–1276 1271

quently the overall GnRH-stimulated LH� promoteractivity, and this effect is greatest on the �617LH�construct that contains both GnRH-responsive re-gions and thus binds both Sp1 and SF-1.
We cannot definitively deduce from these studieswhether more than one SNURF molecule operateswith the LH� promoter at the same time, or whetherone SNURF molecule binds to both Sp1 and SF-1 onthe promoter simultaneously. Because SNURF canbind to Sp1 via its RING finger domain and to SF-1 viaits N-terminal region, simultaneous binding is possi-ble. Only SNURF forms that bind both Sp1 and SF-1have full stimulatory activity on the LH� promoter.SNURF and the DNA-binding mutant SNURF-R8–11A/K9A, both of which bind Sp1 and SF-1, stimulate basaland GnRH-stimulated transcription of the LH� pro-moter. In contrast, even though the SF-1 binding mu-tant SNURF�31–65 can interact with Sp1, it is unableto enhance basal or GnRH-stimulated transcription(Fig. 4B). Similarly, the Sp1 binding mutant, SNURF-CS1, which still retains some SF-1 binding ability,does not enhance activity of the �617LH� promoter(Fig. 4B). This mutant also lacks TBP binding ability.The failure of CS1 to activate the �617 and �245LH�promoter constructs may be due, in part, to the loss ofthis property, as the ability to act as a bridging factorto TBP appears to be critical for SNURF action on ARand ER and is likely to be important for SF-1 (18–21).Looping of chromatin enabling two distantly spacedpromoter regions to interact has been postulated forinteractions between AR and Sp1 on the cyclin-dependent kinase inhibitor p21 gene (37) and has beendemonstrated in chromatin for Pit-1 and ER bindingenhancer regions of the rat PRL gene after estrogen
treatment (38–40). Other coactivator proteins that canbind multiple proteins and have been shown to pos-sess multiple different functions include E6-AP, an E3ubiquitin protein ligase (41), high mobility group-1/2,chromatin high mobility group proteins (42), Zac1, azinc finger protein (43), and SRA, a steroid receptorcoactivator that is an RNA molecule (44). Thus,SNURF is one of several coactivators that may inte-grate several transcription functions, with potentiallyimportant tissue and gene-specific roles.
Given that SNURF was originally identified throughits interaction with AR, and androgens suppress GnRHstimulation of LH�, we tested the effects of SNURF onthis suppression. Previously, we demonstrated thatAR represses the promoter, not via direct binding toDNA but by interference with transcription factor bind-ing to the promoter (16) and that holo-AR inhibitsbinding of Sp1 to the LH� distal enhancer region.Other investigators have found that AR prevents SF-1binding to the bovine promoter, and that the degree ofthis interaction is influenced by Egr-1 and Pitx1 (23).We show in this work that overexpression of SNURFrescues LH� promoter activity from androgen sup-pression. SNURF interacting with Sp1 as well as withSF-1 may protect these factors bound to the promoterfrom AR, or SNURF may alternatively bind to AR andsequester it (Fig. 9). Two findings argue against this asthe primary mechanism of action. First, the SNURF-CS1 mutant can still bind AR, but did not restoreGnRH stimulation of the promoter in the presence ofDHT. Second, the �-subunit promoter is suppressedby DHT, and titration of AR by SNURF should thenprevent or rescue suppression by DHT. These datasuggest that SNURF interaction with Sp1 and SF-1 on
Fig. 9. Model for SNURF Interaction with the Two GnRH-Responsive Regions of LH� PromoterThe top panel illustrates the distal and proximal GnRH-responsive regions of the LH� promoter and transcription factors
proposed to bind to these regions. SNURF has been shown to interact with Sp1 and SF-1. It is proposed that the DNA betweenthe distal and proximal regions forms a loop enabling the interaction between these two regions. SNURF binding with factors inboth regions may enable it to mediate the interaction between the regions, thereby increasing transcription under basal andstimulated conditions and preventing interference by AR.
1272 Mol Endocrinol, May 2004, 18(5):1263–1276 Curtin et al. • SNURF Stimulates LH� Promoter via SF-1 and Sp1

DNA, either by steric hindrance or changes in protein/chromatin conformation, blocks the access of AR tothe transcription factors. The mechanism that is mostimportant in the cell may depend on relative amountsof each factor or relative affinity of SNURF for AR,SF-1, and Sp1 bound to chromatin.
Overall, our results indicate that SNURF can func-tion as a bridging factor on a complex physiologicalpromoter. Enhancement of LH� promoter activity re-quires coordinate interaction of SNURF with two sep-arate transcription factors, Sp1 and the nuclear orphanreceptor SF-1, which bind to different specific SNURFdomains. These transcription factors bind the pro-moter on enhancer regions positioned over 250 ntapart, suggesting a novel mechanism by whichSNURF may act as an integrator for enhancercooperation.
MATERIALS AND METHODS
Cell Culture and Transfection Studies
The clonal gonadotrope cell line, L�T2, secretes LH andexpresses LH� and �-subunit as well as GnRH, estrogen,and androgen receptors (29), and was originally obtainedfrom Dr. Pamela Mellon (University of California San Diego).Cells were treated in phenol red-free DMEM with charcoal-stripped 5% newborn calf serum and antibiotics as described(16). Cells were plated in 12-well plates at a density of 5 � 105
cells per well 16–20 h before CaPO4 transfection. Each wellwas transfected with 1.5 �g of reporter vector as indicated for16 h, washed, and treated with 10 nM GnRH for 6 h beforecollection and measurement of luciferase activity. Steroids orforskolin (1 �M; Sigma Chemical Co., St. Louis, MO) wereincluded as indicated during the transfection period and theGnRH treatment period, for a total treatment of 24 h. Inseveral experiments, a cytomegalovirus (CMV)-�-galactosi-dase vector (0.3 �g/well) was included to normalize for lucif-erase activity, and in all cases luciferase activity was alsonormalized for protein. For studies with mutant LH� promoterconstructs, normalized luciferase activity was expressed rel-ative to the full-length �617LH� construct, which was set at1.0. After transfection, cells were washed with PBS and ly-sates were collected in 150 �l lysis buffer (Promega Corp.,Madison, WI), vortexed, centrifuged for 1 min, and assayed ina Turner 20e luminometer (Turner Designs, Mountain View,CA). Protein concentrations were determined by the colori-metric assay from Bio-Rad Laboratories (Hercules, CA). Hor-mones were obtained from Sigma Chemical Co.
LH�-luciferase reporter vectors have been described else-where (3). For most studies, the construct �617LH� contain-ing the promoter region from �617 to �44 nt relative to therat LH� gene transcriptional start site, and both GnRH-re-sponsive elements, was used. To test the relative importanceof the distal region in the SNURF response, a mutant con-struct containing both GnRH-response regions, but with mu-tations in both SF-1 sites in the proximal response regions(2SF1-Mut) was used. In some experiments the deletion con-struct �245LH� corresponding to nt �45 to �44 and con-taining only the proximal GnRH response element was alsotested. The �-subunit promoter luciferase reporter vector(�411�) contains �411 to �77 nt of the �-subunit promoter(3, 45). The pcDNA3.1(�)-FLAG-SNURF constructs encodingthe following SNURF forms were used: wild-type SNURF,SNURF-CS1 in which the cysteines 136 and 139 are substi-tuted with serines creating a mutant unable to bind Sp1 (21),SNURF-R8–11A/K9A in which Arg8, Lys9, Arg10, and Arg11
were changed to alanines yielding a DNA binding-deficientmutant (20), and SNURF�31–65 in which amino acid residues31–65 have been deleted, creating a mutant that cannotactivate AR-dependent transcription (21). CMV-hAR (46) andpcDNA-CBP (47) expression vectors have been described.For studies using wild-type or mutant SNURF, total DNA ineach transfection reaction was balanced with empty vector.A SNURF dose-response curve was performed to test max-imally effective concentrations and, unless otherwise indi-cated, SNURF constructs were added at 1.5 �g/500,000cells. To test CBP function, we used a CRE-luciferase re-porter construct containing six tandem CRE elements fusedto the TATA box of the rat PRL gene promoter, kindly pro-vided by Dr. Richard N. Day (University of Virginia, Char-lottesville, VA).
Immunoblotting
Expression of transfected wild-type and mutant SNURF-FLAG constructs was evaluated in L�T2 cells 24 h posttrans-fection by immunoblotting. Approximately 50 �g of trans-fected cell lysate were examined on 12% SDS-PAGE,followed by transfer to a nylon membrane, incubation withM5 FLAG primary antibody (1:2000; Sigma Chemical Co.),horseradish peroxidase-labeled goat antimouse IgG second-ary antibody (1:10,000; Jackson ImmunoResearch Laborato-ries, Inc., West Grove, PA) followed by detection with PicoW-est (Pierce Chemical Co., Rockford, IL). Expression of nativeSNURF protein was examined in pituitary tissue from maturemale and female mice and in clonal cell lines. GH3 cells werecultured as described (48). Cos-1 cells were transfected asdescribed (49) with 1.5 �g of SNURF expression vector andcollected after 24 h. L�T2 cells were treated with and without100 nM GnRH for 2 h, which stimulates Egr-1 expression.Tissue or cell lysates (25 �g) were examined on 10% SDS-PAGE and transferred to a nylon membrane. Polyclonal anti-SNURF antibody was generated in rabbits against full-lengthSNURF protein produced in insect cells (18). SNURF wasdetected with this antiserum at a dilution of 1:20,000, fol-lowed by incubation with horseradish peroxidase-labeleddonkey antirabbit IgG secondary antibody (1:20,000, Amer-sham Biosciences, Buckinghamshire, UK) followed by detec-tion with PicoWest. For both types of studies, �-actin wasused as a loading control, as previously described (48). Du-plicate samples were tested from three to four independentexperiments, and samples from the same experiment wererun simultaneously on parallel gels in the same apparatus.
Statistical Analysis
Data for normalized luciferase activity are presented as themean � SEM for six wells per group, compared with untreatedcontrols, and each experiment was performed between threeand six times with equivalent results. Statistical significancewas evaluated by two-way ANOVA comparing control andGnRH treatment groups, and wild-type and mutant SNURF orCBP expression, and analyzed post hoc by Tukey’s whollysignificant difference test. Statistical significance was takento be at least P � 0.05, or as shown in the figures.
GST Pull-Down Experiments
BL21 bacterial cells were transformed with constructs ex-pressing GST, GST-Sp1, GST-Egr-1, GST-SF-1, or GST-SNURF. Luria Broth (100 ml) containing 50 �g/ml ampicillinwas inoculated with 1 ml of bacteria and incubated in anorbital shaker at 37 C. Bacteria were grown to A600 � 0.5,induced with 0.1 mM isopropyl �-thiogalactopyranoside, andshaken overnight at room temperature. The bacterial pelletwas resuspended in 5 ml of buffer containing 50 mM Tris (pH7.5), 0.5 mM EDTA, 300 mM NaCl, 10 mg/ml lysozyme, and 1
Curtin et al. • SNURF Stimulates LH� Promoter via SF-1 and Sp1 Mol Endocrinol, May 2004, 18(5):1263–1276 1273

mM dithiothreitol. One hundred microliters of 10% NonidetP-40 were added, and after 10 min, the lysate was frozen at�70 C in an ethanol bath. Lysate was thawed at room tem-perature and then incubated for 1 h in 5 ml of buffer contain-ing 1.5 M NaCl, 12 mM MgCl2, 5 �g DNase I, 10 �g/mlleupeptin, 1 �g/ml pepstatin A, and 0.1 mM phenylmethylsul-fonylfluoride. Lysates were passed through a 20-gauge nee-dle and centrifuged for 30 min at 7500 � g. Supernatantswere incubated with glutathione beads (Sigma) overnight at 4C. Beads were extensively washed with PBS, and boundprotein concentrations were analyzed by electrophoresis on12% SDS-PAGE followed by Coomassie Blue staining andimmunoblotting with appropriate antibodies [Sp1 and Egr-1antibody from Santa Cruz Biotechnology, Inc., Santa Cruz,CA; SF-1 antibody from Upstate Biotechnology, Lake Placid,NY; and anti-SNURF antiserum (17)]. For pull-down experi-ments, approximately 1 �g of GST fusion protein was used ineach sample incubation. BSA (20 �g/ml) was added to eachincubation containing [35S]methionine-labeled (0.04 mCi/50-�l reaction) in vitro translated proteins (TNT Rabbit Re-ticulocyte Transcription/Translation Kit; Promega). Labeledproteins included SNURF, SNURF-CS1, SNURF R8–11A/K9A, SNURF �31–65, Egr-1, SF-1, and Sp1. For SNURFstudies, approximately equal molar quantities of SNURF orSNURF mutant proteins were used in reactions. Total volumewas adjusted to 150 �l with GST wash buffer (10 mM MgCl2;150 mM KCl; 20 mM HEPES, pH 7.6; 10% glycerol; and 0.12%Nonidet P-40). Beads and proteins were incubated for 1.5 hat 4 C including GST alone with labeled proteins for controlexperiments, and then centrifuged and washed four times inGST wash buffer. Beads were resuspended in 10 �l of so-dium dodecyl sulfate (SDS) loading buffer and boiled for 5min. Proteins were electrophoresed on SDS containing 10%acrylamide gels at 150 V, along with standard molecularweight markers (Benchmark, Life Technologies, Inc., Gaith-ersburg, MD). Gels containing [35S]methionine-labeled pro-teins were dried and exposed to film for 24–72 h at �70 C.Films were scanned and subjected to densitometric analysis.Each experiment was performed a minimum of five to sixtimes.
Chromatin Immunoprecipitation (ChIP) Assays
ChIP assays were performed essentially as described byChakarabarti et al. (50). L�T2 proteins were cross-linked togenomic DNA by addition of formaldehyde to a final concen-tration of 1%, in PBS. Chromatin was sonicated to an aver-age length of 1 kb in 1% Triton X-100, 0.1% deoxycholate, 50mM Tris, pH 8.1, 150 mM NaCl, 5 mM EDTA. After the lysateswas clarified, 1 �g of control CMV-luciferase plasmid and 100�g of BSA were added. Chromatin lysates were then dilutedwith sonication buffer plus protease inhibitors (0.2 mg/mlleupeptin and aprotinin, 20 mM phenylmethylsulfonylfluoride),divided into aliquots, and incubated at 4 C overnight with noantibody, or antibodies to SNURF (5 �l) or SF-1 (5 �l), andthen precipitated with protein A beads (Santa Cruz Biotech-nology, Inc.). Protein DNA complexes were washed threetimes in buffer without protease inhibitors and once in TE,and cross-linking was reversed by addition of NaCl to 100 mM
and incubation at 65 C. Proteins were eluted from the beadsby incubation with elution buffer (1% SDS, 0.1 M NaHCO3,0.01 mg/ml herring sperm DNA) and were digested withProteinase K. DNA was extracted with phenol-chloroform,and mouse LH� promoter sequences were detected withprimers that flank the sequence between �102 bp (5�-CT-GTGTCTCGCCCCCAAAGAGATTA-3�) and �1 bp (5�-CCT-GGCTTTATACCTGCGGGGTT-3�). Sequences were de-tected by real-time PCR (iCycler, Bio-Rad Laboratories,Hercules, CA), incorporating SYBR green, as previously de-scribed (50). Insulin gene promoter primers were used as aspecificity control, and luciferase gene primers were used asan internal control for sample handling (50). For some studies,PCR was performed for 28 cycles, within the linear range of
the curve, and the products were subjected to electrophore-sis. For PCR quantification, previously described protocolswere followed (50), using a total number of 40 cycles, withcontinuous SYBR green monitoring. Briefly, relative propor-tions of immunoprecipitated promoter fragments were deter-mined based on the threshold cycle (Tc) value for each PCRreaction, which is determined automatically and is the cycleat which fluorescence rises 10 times above the mean SD ofbackground in all reaction wells. The �Tc is calculated bycorrection of immunoprecipitated sample fluorescence forluciferase plasmid and for signal with control samples (noantibody). All samples were normalized for input chromatin.Values are calculated for each treatment, and fold changeswere then calculated by raising 2 to the �Tc power. Calcu-lations are summarized as the fold-difference (SNURF ChIPvs. Control ChIP) � 2[Tc(control)-Tc(SNURF)], where Tc �Tc(immunoprecipitated sample)Tc(input). Each promoterfragment was analyzed in duplicate on at least two separateoccasions and from three independent immunoprecipita-tions.
Acknowledgments
We thank Dr. Raghu Mirmira at the University of Virgina, forhis help in establishing the ChIP assays, and many helpfuldiscussions.
Received June 7, 2003. Accepted February 17, 2004.Address all correspondence and requests for reprints to:
Margaret A. Shupnik, Ph.D, Department of Internal Medicine/Endocrinology, PO Box 800578 Health Sciences Center, Uni-versity of Virginia, Charlottesville, Virginia 22908. E-mail:[email protected].
This work was supported by National Institutes of ChildHealth and Human Development/National Institutes of Healththrough cooperative agreement (U54 HD28934) as part of theSpecialized Cooperative Centers Program in ReproductionResearch though both an individual research project (M.A.S.)and the Molecular Core at the University of Virginia, and theAcademy of Finland. This work was also supported by thegenerous gift of reagents from Dr. Elizabeth M. Wilson andthe Specialized Cooperative Centers Program in Reproduc-tion Research Center at the University of North Carolina atChapel Hill (U54-HD35041), through an individual researchproject (to E.M.W).
REFERENCES
1. Karsch FJ 1987 Central actions of ovarian steroids in thefeedback regulation of pulsatile secretion of luteinizinghormone. Annu Rev Physiol 49:365–382
2. Haisenleder DJ, Ortolano GA, Dalkin AC, Marshall JC,Shupnik MA 1991 A pulsatile GnRH stimulus is requiredto increase transcription of the gonadotropin subunitgenes: evidence for differential stimulation of transcrip-tion by pulse frequency in vivo. Endocrinology 128:509–517
3. Weck J, Anderson AC, Jenkins S, Fallest PC, ShupnikMA 2000 Divergent and composite gonadotropin-releas-ing hormone-responsive elements in the rat luteinizinghormone subunit genes. Mol Endocrinol 14:472–485
4. Kaiser UB, Sabbagh E, Chen MT, Chin WW, SaundersBD 1998 Sp1 binds to the rat luteinizing hormone beta(LH�) gene promoter and mediates gonadotropin-releas-ing hormone-stimulated expression of the LH� subunitgene. J Biol Chem 273:12943–12951
5. Tremblay JJ, Drouin J 1999 Egr-1 is a downstream ef-fector of GnRH and synergizes by direct interaction with
1274 Mol Endocrinol, May 2004, 18(5):1263–1276 Curtin et al. • SNURF Stimulates LH� Promoter via SF-1 and Sp1

Ptx1 and SF-1 to enhance luteinizing hormone � genetranscription. Mol Cell Biol 19:2567–2576
6. Kaiser UB, Halvorson LM, Chen MT 2000 Sp1, steroido-genic factor 1 (SF-1), and early growth response protein1 (egr-1) binding sites form a tripartite gonadotropin-releasing hormone response element in the rat luteinizinghormone-� gene promoter: an integral role for SF-1. MolEndocrinol 14:1235–1245
7. Dorn C, Ou Q, Svaren J, Crawford PA, Sadovsky Y 1999Activation of luteinizing hormone � gene by gonado-tropin-releasing hormone requires the synergy of earlygrowth response-1 and steroidogenic factor-1. J BiolChem 274:13870–13876
8. Halvorson LM, Kaiser UB, Chin WW 1996 Stimulation ofluteinizing hormone � gene promoter activity by the or-phan nuclear receptor, steroidogenic factor-1. J BiolChem 271:6645–6650
9. Rosenberg SB, Mellon PL 2002 An Otx-related home-odomain protein binds an LH� promoter element impor-tant for activation during gonadotrope maturation. MolEndocrinol 16:1280–1298
10. Haisenleder DJ, Yasin M, Dalkin AC, Gilrain J, MarshallJC 1996 GnRH regulates steroidogenic factor-1 (SF-1)gene expression in the rat pituitary. Endocrinology 137:5719–5722
11. Wolfe MW, Call GB 1999 Early growth response protein1 binds to the luteinizing hormone � promoter and me-diates gonadotropin-releasing hormone stimulated geneexpression. Mol Endocrinol 13:752–763
12. Duan WR, Ito M, Park Y, Maizels ET, Hunzicker-Dunn M,Jameson JL 2002 GnRH regulates early growth responseprotein 1 transcription through multiple promoter ele-ments. Mol Endocrinol 16:221–233
13. Zhang T, Wolfe MW, Roberson MS 2001 An early growthresponse protein (Egr) 1 cis-element is required forgonadotropin-releasing hormone-induced mitogen-activated protein kinase phosphatase 2 gene expression.J Biol Chem 276:45604–45613
14. Hanley NA, Ikeda Y, Luo X, Parker KL 2000 Steroido-genic factor 1 (SF-1) is essential for ovarian developmentand function. Mol Cell Endocrinol 163:27–32
15. Call GB Wolfe MW 2002 Species differences in GnRHactivation of the LH� promoter: role of Egr1 and Sp1. MolCell Endocrinol 189:85–96
16. Curtin D, Jenkins S, Farmer N, Anderson AC, HaisenlederDJ, Rissman E, Wilson EM, Shupnik MA 2001 Androgensuppression of GnRH-stimulated rat LH� gene transcrip-tion occurs through Sp1 sites in the distal GnRH-respon-sive promoter region. Mol Endocrinol 15:1906–1917
17. Naar AM, Lemon BD, Tjian R 2001 Transcriptional coac-tivator complexes. Annu Rev Biochem 70:475–501
18. Moilanen AM, Poukka H, Karvonen U, Hakli M, JanneOA, Palvimo JJ 1998 Identification of a novel RING fingerprotein as a coregulator in steroid receptor-mediatedgene transcription. Mol Cell Biol 18:5128–5139
19. Saville B, Poukka H, Wormke M, Janne OA, Palvimo JJ,Stoner M, Samudio I, Safe S 2002 Cooperative coacti-vation of estrogen receptor � in ZR-75 human breastcancer cells by SNURF and TATA-binding protein. J BiolChem 277:2485–2497
20. Hakli M, Karvonen U, Janne OA, Palvimo JJ 2001 TheRING finger protein SNURF is a bifunctional proteinpossessing DNA binding activity. J Biol Chem 276:23653–23660
21. Poukka H, Aarnisalo P, Santti H, Janne OA, Palvimo JJ2000 Coregulator small nuclear RING finger protein(SNURF) enhances Sp1- and steroid receptor-mediatedtranscription by different mechanisms. J Biol Chem 275:571–579
22. Jorgensen JS, Nilson JH 2001 AR suppresses transcrip-tion of the LH� subunit by interacting with steroidogenicfactor-1. Mol Endocrinol 15:1505–1516
23. Spratt DP, Herbison AE 1997 Regulation of preoptic areagonadotrophin-releasing hormone (GnRH) mRNA ex-pression by gonadal steroids in the long-term gonadec-tomized rat. Brain Res Mol Brain Res 47:125–133
24. Gharib SD, Wierman ME, Shupnik MA, Chin WW 1990Molecular biology of the pituitary gonadotropins. EndocrRev 11:177–199
25. Midgley Jr AR, Jaffe RB 1971 Regulation of human go-nadotropins. X. Episodic fluctuation of LH during themenstrual cycle. J Clin Endocrinol Metab 33:962–969
26. Yasin M, Dalkin AC, Haisenleder DJ, Kerrigan JR, Mar-shall JC 1995 Gonadotropin-releasing hormone (GnRH)pulse pattern regulates GnRH receptor gene expres-sion: augmentation by estradiol. Endocrinology 136:1559–1564
27. Jorgensen JS, Nilson, JH 2001 AR suppresses transcrip-tion of the � glycoprotein hormone subunit gene throughprotein-protein interactions with cJun and activationtranscription factor 2. Mol Endocrinol 15:1496–1504
28. Heckert LL, Wilson EM, Nilson JN 1997 Transcriptionalrepression of the �-subunit gene by androgen receptoroccurs independently of DNA binding but requires theDNA-binding and ligand-binding domains of the recep-tor. Mol Endocrinol 11:1497–1506
29. Windle JJ, Weiner RI, Mellon PL 1990 Cell lines of thepituitary gonadotrope lineage derived by targeted onco-genesis in transgenic mice. Mol Endocrinol 4:597–603
30. Cook T, Gebelein B, Urrutia R 1999 Sp1 and its likes:biochemical and functional predictions for a growingfamily of zinc finger transcription factors. Ann NY AcadSci 880:94–102
31. Liu C, Calogero A, Ragona G, Adamson E, Mercola D1996 EGR-1, the reluctant suppression factor: Crit RevOncog 7:101–125
32. Tremblay JJ, Lanctot C, Drouin J 1998 The pan-pituitaryactivator of transcription, Ptx1 (pituitary homeobox 1),acts in synergy with SF-1 and Pit1 and is an upstreamregulator of the Lim-homeodomain gene LIM3/Lhx3. MolEndocrinol 12:428–441
33. Liu Z, Simpson ER 1997 Steroidogenic factor 1 (SF-1)and Sp1 are required for regulation of bovine CYP11Agene expression in bovine luteal cells and adrenal Y1cells. Mol Endocrinol 11:127–137
34. Topilko P, Schneider-Maurnory S, Levi G, Trebleau A,Gourdji D, Driancourt M-A, Rao CV, Charnay P 1998Multiple pituitary, ovarian defects in Krox-24 (NGFI-A,Egr-1)-targeted mice. Mol Endocrinol 12:107–122
35. Honda SI, Morohashi KI, Nomura M, Takeya H, KitajimaM, Omura T 1993 Ad4BP regulating steroidogenic P-450gene is a member of steroid hormone receptor super-family. J Biol Chem 268:7494–7502
36. Takemaru K-I, Li F-Q, Ueda H, Hirose S 1997 Multipro-tein bridging factor (MBF1) is an evolutionary conservedtranscriptional coactivator that connects a regulatoryfactor and TATA element binding protein. Proc Natl AcadSci USA 94:7251–7256
37. Lu S, Jenster G, Epner DE 2000 Androgen induction ofcyclin-dependent kinase inhibitor p21 gene: role of an-drogen receptor and transcription factor Sp1 complex.Mol Endocrinol 14:753–760
38. Gothard LQ, Hibbard, JC, Seyfred MA 1996 Estrogenmediated induction of rat prolactin gene transcriptionrequires the formation of a chromatin loop between thedistal enhancer and proximal promoter regions. Mol En-docrinol 10:185–195
39. Cullen KE, Kladde MP, Seyfred MA 1993 Interaction be-tween transcription regulatory regions of prolactin chro-matin. Science 261:203–206
40. Seyfred MA, Gorski, J 1990 An interaction between the 5�flanking distal and proximal regulatory domains of the ratprolactin gene is required for transcriptional activation byestrogens. Mol Endocrinol 4:1226–1234
Curtin et al. • SNURF Stimulates LH� Promoter via SF-1 and Sp1 Mol Endocrinol, May 2004, 18(5):1263–1276 1275

41. Nawaz Z, Lonard DM, Smith CL, Lev-Lehman E, Tsai SY,Tsai MJ, O’Malley BW 1999 The Angelman syndrome-associated protein, E6-AP, is a coactivator for the nu-clear hormone receptor superfamily. Mol Cell Biol 19:1182–1189
42. Melvin VS, Edwards DP 1999 Coregulatory proteins insteroid hormone receptor action: the role of chromatinhigh mobility group proteins HMG-1 and -2. Steroids64:576–586
43. Huang S, Stallcup MR 2000 Mouse Zac1, a transcrip-tional coactivator and repressor for nuclear receptors.Mol Cell Biol 20:1855–1867
44. Lanz RB, McKenna NJ, Onate SA, Albrecht U, Wong J,Tsai SY, Tsai MJ, O’Malley BW 1999 A steroid receptorcoactivator, SRA, functions as an RNA and is present inan SRC-1 complex. Cell 97:17–27
45. Weck J, Fallest PC, Pitt LK, Shupnik MA 1998 Differentialgonadotropin-releasing hormone stimulation of rat lu-teinizing hormone subunit gene transcription by calciuminflux and mitogen-activated protein kinase-signalingpathways. Mol Endocrinol 12:451–457
46. He B, Kempainen JA, Wilson EM 2000 FXXLF andWXXLF sequences mediate the NH2-terminal interactionwith the ligand binding domain of the androgen receptor.J Biol Chem 275:22986–22994
47. Chrivia JC, Kwok RP, Lamb N, Hagiwara MR, GoodmanRH 1993 Phosphorylated CREB binds specifically to thenuclear protein CBP. Nature 365:855–859
48. Schreihofer DA, Stoler M, Shupnik MA 2000 Cell-specificexpression and regulation of estrogen receptor isoformsin rat pituitary: estrogen treatment specifically decreasesER� protein levels and estrogen responsiveness. Endo-crinology 141:2174–2184
49. Lin VY, Resnick EM, Shupnik MA 2003 Stimulation ofestrogen receptor activity by truncated estrogen recep-tor product-1 occurs by titration of repressors. J BiolChem 278:38125–38131
50. Chakrabarti SK, James JC, Mirmira RG 2002 Quantita-tive assessment of gene targeting in vitro and in vivoby pancreatic transcription factor, Pdx1. J Biol Chem277:13286–13293
Molecular Endocrinology is published monthly by The Endocrine Society (http://www.endo-society.org), the foremostprofessional society serving the endocrine community.
1276 Mol Endocrinol, May 2004, 18(5):1263–1276 Curtin et al. • SNURF Stimulates LH� Promoter via SF-1 and Sp1




















