
and Fibronectin Addressin Cell Adhesion Molecule1 Lymphocyte Adhesion to Mucosal {alpha}4{beta}7...
11
of March 27, 2014. This information is current as and Fibronectin Mucosal Addressin Cell Adhesion Molecule-1 Integrin-Mediated Lymphocyte Adhesion to 7 β 4 α Modulates α Factor-1 The Chemokine Stromal Cell-Derived Parmo-Cabañas, Michael J. Briskin and Joaquin Teixidó Rodríguez-Frade, Silvia F. Soriano, Mario Mellado, Marisa Natalia Wright, Andrés Hidalgo, José Miguel http://www.jimmunol.org/content/168/10/5268 2002; 168:5268-5277; ; J Immunol References http://www.jimmunol.org/content/168/10/5268.full#ref-list-1 , 38 of which you can access for free at: cites 58 articles This article Subscriptions http://jimmunol.org/subscriptions is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/ji/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/cgi/alerts/etoc Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved. Copyright © 2002 by The American Association of 9650 Rockville Pike, Bethesda, MD 20814-3994. The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology by guest on March 27, 2014 http://www.jimmunol.org/ Downloaded from by guest on March 27, 2014 http://www.jimmunol.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of and Fibronectin Addressin Cell Adhesion Molecule1 Lymphocyte Adhesion to Mucosal {alpha}4{beta}7...

of March 27, 2014.This information is current as and Fibronectin
Mucosal Addressin Cell Adhesion Molecule-1Integrin-Mediated Lymphocyte Adhesion to
7β4α Modulates αFactor-1The Chemokine Stromal Cell-Derived
Parmo-Cabañas, Michael J. Briskin and Joaquin TeixidóRodríguez-Frade, Silvia F. Soriano, Mario Mellado, Marisa Natalia Wright, Andrés Hidalgo, José Miguel
http://www.jimmunol.org/content/168/10/52682002; 168:5268-5277; ;J Immunol
Referenceshttp://www.jimmunol.org/content/168/10/5268.full#ref-list-1
, 38 of which you can access for free at: cites 58 articlesThis article
Subscriptionshttp://jimmunol.org/subscriptions
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/ji/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/cgi/alerts/etocReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2002 by The American Association of9650 Rockville Pike, Bethesda, MD 20814-3994.The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on March 27, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on March 27, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from

The Chemokine Stromal Cell-Derived Factor-1� Modulates�4�7 Integrin-Mediated Lymphocyte Adhesion to MucosalAddressin Cell Adhesion Molecule-1 and Fibronectin1
Natalia Wright,* Andres Hidalgo, 2* Jose Miguel Rodrıguez-Frade,† Silvia F. Soriano,†
Mario Mellado, † Marisa Parmo-Cabanas,* Michael J. Briskin,‡ and Joaquin Teixido3*
The interaction between the integrin�4�7 and its ligand, mucosal addressin cell adhesion molecule-1, on high endothelial venulesrepresents a key adhesion event during lymphocyte homing to secondary lymphoid tissue. Stromal cell-derived factor-1� (SDF-1�)is a chemokine that attracts T and B lymphocytes and has been hypothesized to be involved in lymphocyte homing. In this workwe show that�4�7-mediated adhesion of CD4� T lymphocytes and the RPMI 8866 cell line to mucosal addressin cell adhesionmolecule-1 was up-regulated by SDF-1� in both static adhesion and cell detachment under shear stress assays. Both naive andmemory phenotype CD4� T cells were targets of SDF-1�-triggered increased adhesion. In addition, SDF-1� augmented�4�7-dependent adhesion of RPMI 8866 cells to connecting segment-1 of fibronectin. While pertussis toxin totally blocked chemotaxisof CD4� and RPMI 8866 cells to SDF-1�, enhanced�4�7-dependent adhesion triggered by this chemokine was partially inhibited,indicating the participation of G �i-dependent as well as G�i-independent signaling. Accordingly, we show that SDF-1� induceda rapid and transient association between its receptor CXCR4 and G�i, whereas association of pertussis toxin-insensitive G�13
with CXCR4 was slower and of a lesser extent. SDF-1� also activated the small GTPases RhoA and Rac1, and inhibition of RhoAactivation reduced the up-regulation of�4�7-mediated lymphocyte adhesion in response to SDF-1�, suggesting that activation ofRhoA could play an important role in the enhanced adhesion. These data indicate that up-regulation by SDF-1� of lymphocyteadhesion mediated by�4�7 could contribute to lymphocyte homing to secondary lymphoid tissues.The Journal of Immunology,2002, 168: 5268–5277.
L ymphocytes recirculate from blood to secondary lym-phoid tissues and back again into the blood circulation, aprocess that is of fundamental importance for normal im-
mune surveillance (1–3). Homing of lymphocytes to lymphoid tis-sues is mediated by selective interactions between adhesion recep-tors expressed on lymphocytes and their ligands displayed on highendothelial venules (HEV)4 located on postcapillary venules (4).The integrin�4�7 is a cell adhesion receptor expressed on differentT and B lymphocyte subsets that mediates their attachment to HEVin mucosa-associated lymphoid tissues by interacting with muco-sal vascular addressin mucosal addressin cell adhesion molecule-1(MAdCAM-1) (5–9). MAdCAM-1, a 60-kDa glycoprotein that be-
longs to the Ig superfamily, is expressed on HEV in Peyer’spatches, mesenteric lymph nodes, and lamina propria venuleswithin the gut, and its expression represents sites of lymphocyteextravasation into these intestine-associated lymphoid tissues (4, 9,10). Naive T lymphocytes (CD45RA� among CD4�) show rela-tively homogeneous intermediate levels of�4�7 expression,whereas memory B and T lymphocytes can be subdivided into�4�7
high and�4�7� populations (6, 11, 12). Both naive and mem-
ory T cells home efficiently to Peyer’s patches, but�4�7� lym-
phocytes are excluded (12, 13), indicating that�4�7 expression isassociated with homing to intestinal lymphoid tissues.
Apart from mediating cell adhesion to MAdCAM-1,�4�7 can alsointeract with the connecting segment-1 region of fibronectin (CS-1/FN) (14, 15). This interaction might play an important role in�4�7
high
lymphocyte homing within lymphoid tissues rather than in recruit-ment of lymphocytes from the blood (4). Several amino acids on both�4 and�7 subunits that are critical for�4�7-dependent cell adhesionto MAdCAM-1 and CS-1/FN have recently been identified (16–18).
After tethering and rolling of lymphocytes on HEV, they rapidlystick and arrest, a process involving�4�7/MAdCAM-1 interac-tions and favored by chemokine-triggered integrin activation (19).The stromal cell-derived factor-1� (SDF-1�; CXCL12) is a CXCchemokine that potently attracts lymphocytes (20–22) and exertschemoattractive and activating functions upon binding to its Gprotein-coupled receptor CXCR4, which is expressed on B and Tlymphocytes, including CD4� and CD8� cells (23–26). In addi-tion, CXCR4 acts as a coreceptor for T-tropic HIV, and SDF-1�
inhibits T tropic HIV infection (23, 25).Previous studies have shown that SDF-1� can modulate the ad-
hesive activity of the VLA-4 integrin on CD34� human bone mar-row hemopoietic progenitors, myeloma cells, and T lymphocytes
*Department of Immunology, Centro de Investigaciones Biologicas, and†Departmentof Immunology and Oncology, Centro Nacional de Biotecnologıa, Madrid, Spain; and‡Millenium Pharmaceuticals, Inc., Cambridge, MA 02139
Received for publication July 12, 2001. Accepted for publication March 5, 2002.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby markedadvertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by Grant SAF99-0057 from Ministerio de Ciencia y Tec-nologıa. N.W. is the recipient of a predoctoral fellowship from the Comunidad deMadrid.2 Current address: Department of Hematology, Mount Sinai School of Medicine, OneGustave L. Levy Place, New York, NY 10029.3 Address correspondence and reprint requests to Dr. Joaquin Teixido, Department ofImmunology, Centro de Investigaciones Biologicas, Velazquez 144, 28006 Madrid,Spain. E-mail address: [email protected] Abbreviations used in this paper: HEV, high endothelial venule; CS-1/FN, connectingsegment-1 of fibronectin; F-actin, polymerized actin; PTX, pertussis toxin; SDF-1�, stro-mal cell-derived factor-1�; rh, recombinant human; MAdCAM-1, mucosal addressin celladhesion molecule-1; sMAdCAM-1-IgG, soluble human MAdCAM-1-Ig fusion protein;CHO, Chinese hamster ovary; BCECF-AM, 2�,7�-bis-(2-carboxyethyl)-5-(and-6)-car-boxyfluorescein acetoxymethyl ester; SLC, secondary lymphoid tissue chemokine.
The Journal of Immunology
Copyright © 2002 by The American Association of Immunologists 0022-1767/02/$02.00
by guest on March 27, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from

(27–30). SDF-1� is constitutively expressed on many tissues, in-cluding secondary lymphoid tissues (20, 31), and is therefore apotential candidate to contribute to lymphocyte homing during re-circulation. In the present work we have investigated whether�4�7-dependent lymphocyte adhesion to MAdCAM-1 and CS-1/FN can be subjected to regulation by SDF-1�. Modulation of thisadhesion could contribute to lymphocyte homing to mucosa-asso-ciated lymphoid tissue.
Materials and MethodsCells and Abs
The human B lymphoblastoid cell line RPMI 8866 was cultured in RPMI1640 medium (BioWhittaker, Verviers, Belgium) supplemented with 10%FBS (BioWhittaker) and antibiotics (complete medium). The Chinese ham-ster ovary (CHO)-MAdCAM-1 transfectants were maintained in �-MEM(BioWhittaker) supplemented with 10% FBS and containing 0.8 mg/mlG418 (Calbiochem, San Diego, CA). Human PBMC were isolated frombuffy coats using a Ficoll density gradient (Biochrom, Berlin, Germany).After attachment to plastic for 1.5 h at 37°C in complete medium, nonad-hered cells were recovered, and CD4� T lymphocytes were purified withthe CD4 Positive Isolation kit (Dynal Biotech, Oslo, Norway). Purity was�99% for each sample, as analyzed by flow cytometry (EPICS XL;Coulter, Hialeah, FL). The mAbs used in this study included anti-�1 Lia1/2.1, anti-CD4 T4, anti-CD45RA RP 2/2.1, P3X63 (all gifts from Dr. F.Sanchez-Madrid, Hospital de la Princesa, Madrid, Spain), anti-�4�7 Act-1(32), anti-CD45RO (Caltag Laboratories, Burlingame, CA), anti-CXCR444.717.111 (R&D Systems, London, U.K.), and CXCR4-01 (33). Anti-G�i
polyclonal Ab was purchased from Santa Cruz Biotechnology (Santa Cruz,CA), and anti-G�13 was a gift from Dr. P. C. Sternweis (University ofTexas, Dallas, TX).
Static adhesion assays
The generation of a soluble human MAdCAM-1-Ig fusion protein(sMAdCAM-1-IgG) has been previously described (18). The FN-H89 frag-ment of fibronectin, which contains the CS-1 site and lacks the RGD cen-tral binding domain, was generated as previously reported (34). Cells werelabeled for 20 min at 37°C with the fluorescent dye, 2�,7�-bis-(2-carboxy-ethyl)-5-(and-6)-carboxyfluorescein acetoxymethyl ester (BCECF-AM)(Molecular Probes, Leiden, The Netherlands), washed, resuspended in ad-hesion medium (RPMI/0.5% BSA), and preincubated with or without in-hibitors or Abs. In those experiments in which soluble recombinant human(rh)SDF-1� (R&D Systems) was used, cells were incubated with the che-mokine and added (6 � 104 RPMI 8866 or 105 CD4�) in triplicate to96-well dishes (High-binding; Costar, Cambridge, MA) coated with 5�g/ml FN-H89 as previously described (35) or containing a CHO-MAdCAM-1 monolayer. For adhesions to FN-H89, plates were spun for15 s to place cells in contact with the ligand and allowed to adhere for 5min at 37°C. For adhesions to CHO-MAdCAM-1, cells were incubated for10 min at 37°C with no previous centrifugation. Unbound cells were re-moved by three washes with RPMI 1640 medium. When rhSDF-1� wascoimmobilized with ligands, the chemokine was prepared at 0.5 �g/ml insodium bicarbonate buffer (0.1 M, pH 8.8) for FN-H89 (5 �g/ml) or incarbonate buffer (pH 9.5) for sMAdCAM-1-Ig (300 ng/well) adhesions.The mixtures (50 �l/well) were coated on 96-well dishes for 2 h at 37°C,and wells were finally blocked with 0.5% BSA in the same correspondingbuffers. For sMAdCAM-1-Ig, cells were allowed to settle for 5 min at 4°C,and then adhesion was conducted for 30 min at 37°C, followed by threegentle washes with RPMI 1640. Bound cells were lysed with 1% SDS inPBS, and the extent of adhesion was quantified using a fluorescence ana-lyzer (POLARstar Galaxy; BMG Labtechnologies, Offenburg, Germany).Inhibitors included cytochalasin D and pertussis toxin (PTX; Calbiochem).Recombinant C3 transferase was expressed and purified as previously de-scribed (36).
Flow chamber adhesion assays
Several 15-mm2 areas on petri dishes were coated for 2 h at 37°C with 15�l PBS containing sMAdCAM-1-IgG alone (4 �g/ml), sMAdCAM-1-IgGwith SDF-1� (2 �g/ml), or SDF-1� or PBS alone. The coated spots werewashed with PBS and blocked for 1 h at 37°C with FCS. Petri dishes wereincorporated as the lower wall of a parallel flow chamber (IQUUM, Bos-ton, MA) and mounted on an inverted microscope (IX-70, Olympus, To-kyo, Japan) connected to a CCD camera (Cohu, San Diego, CA). CD4� orRPMI 8866 cells (106/ml) preincubated in the absence or the presence ofPTX and/or anti-�4�7 Act-1 Ab (10 �g/ml) were infused at 37°C for 2 min
at a flow rate of 1 dyne/cm2. Flow was then stopped, and cells were allowedto settle for different times. Total cells from different fields were countedbefore flow was restored (0.5–2 dyne/cm2), and cells remaining tightlybound for �3 min were counted. Data are presented as the percentage ofcells remaining bound compared with total cells in each field before rees-tablishing the flow.
Actin polymerization and chemotactic assays
To determine the content of polymerized actin (F-actin), 105 cells per con-dition were permeabilized, fixed, and stained in a single step by addition ofa 2� solution containing 0.5 mg/ml L-�-lysophosphatidyl-choline (Sigma-Aldrich, St. Louis, MO), 8% formaldehyde, and 4 U/ml FITC-phalloidin(Molecular Probes, Eugene, OR). Cells were incubated at 22°C for 10 min,washed with PBS, and subjected to flow cytometry. For chemotactic as-says, RPMI 8866 (2 � 105) or CD4� cells (3 � 105) in 100 �l adhesionmedium were placed in the upper chamber of a Transwell (5-�m pore size;Costar). Then 600 �l adhesion medium with or without rhSDF-1� (200ng/ml) was added to the lower chamber, and cells were allowed to migratefor 3 h at 37°C. Viable migrated cells were counted in a flow cytometer,analyzing each sample in the same predetermined time and flow conditions.Where indicated, cells were treated with 500 ng/ml PTX for 2 h at 37°C.
GTPase activity assays
We followed essentially the method previously reported (37). The GST-C21 and GST-PAK-CD fusion proteins were generated as previously de-scribed (38). To determine the effect of SDF-1� on RhoA and Rac1 acti-vation, cells were treated with or without 150 ng/ml rhSDF-1�, washed inice-cold PBS, and incubated for 15 min at 4°C in lysis buffer (37). ForRhoA and Rac1 activation upon �4�7/sMAdCAM-1-Ig interaction, we per-formed soluble binding assays of RPMI 8866 cells with sMAdCAM-1-Ig(30–40 �g/ml, 30 min at 23°C), following the method previously de-scribed (16). Cells were then collected, washed, and lysed as describedabove. Lysates were centrifuged, 15 �l was kept for total lysate samples,and the remaining 185 �l was mixed with fusion proteins precoupled toglutathione-agarose beads. The beads and proteins bound to the fusionprotein were washed in an excess of lysis buffer, eluted in Laemmli samplebuffer, and analyzed for bound Rac1 or RhoA by Western blotting usingmAbs against human Rac1 (BD PharMingen/Transduction Laboratories,San Diego, CA) or RhoA (Santa Cruz Biotechnology).
Immunoprecipitation, SDS-PAGE, and Western blot
After SDF-1� stimulation, untreated or PTX-treated cells were solubilized(20 mM triethanolamine (pH 8), 300 mM NaCl, 2 mM EDTA, 20% glyc-erol, and 1% digitonin, with 10 �M sodium orthovanadate, 10 �g/ml leu-peptin, and 10 �g/ml aprotinin). Following centrifugation, lysates wereimmunoprecipitated as previously described (39) using CXCR4-01 Ab.Precipitates or protein extracts were separated in SDS-PAGE and trans-ferred to nitrocellulose membranes. Western blot analysis was performedas previously reported (39), using 5% nonfatty dry milk in TBS as theblocking agent. Protein loading was controlled using a protein detection kit(Pierce, Rockford, IL) as well as by reprobing the membrane with theimmunoprecipitating Ab.
Statistical analysis
The results are expressed as the mean � SD of data obtained from three ormore experiments performed in triplicate unless otherwise stated. Statisti-cal significance was determined using two-tailed Student’s t test.
ResultsSDF-1� up-regulates �4�7-mediated lymphocyte adhesion toMAdCAM-1
Expression of CXCR4 on human CD4� T lymphocytes and that onthe �4�7
��4�1� B lymphoblastoid cell line RPMI 8866 have been
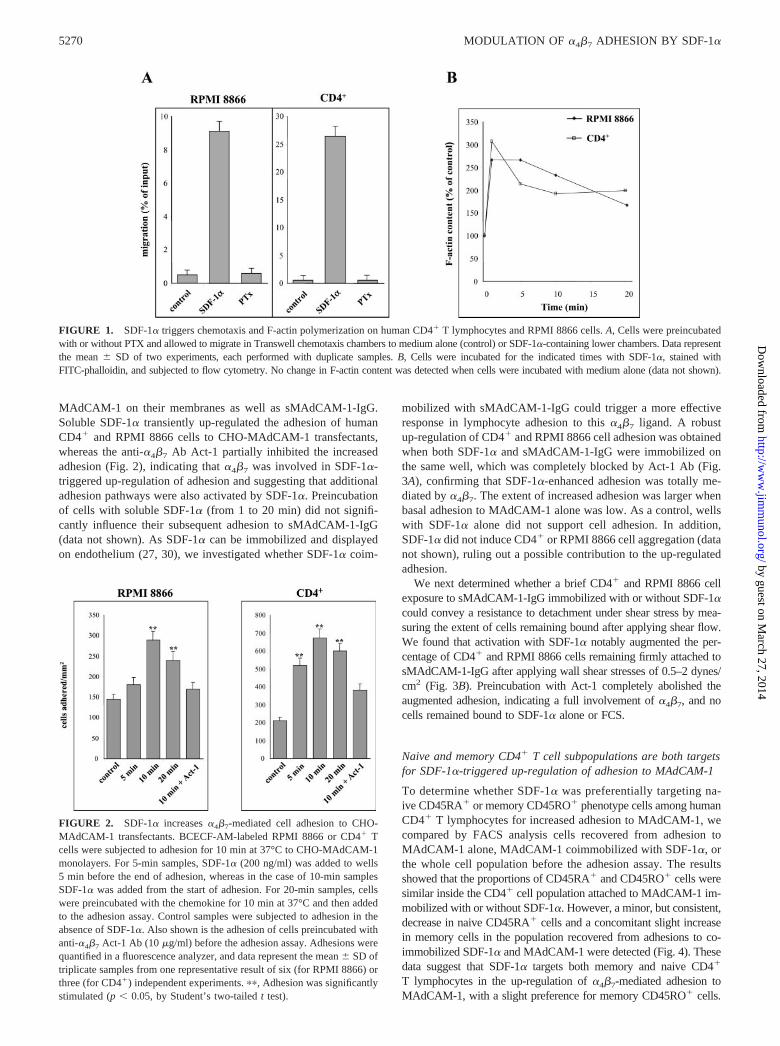
previously described (24, 26). CXCR4 expression conveyed a che-motactic response of CD4� and RPMI 8866 cells to SDF-1� thatwas abolished by PTX, indicating the participation of heterotri-meric G proteins of the Gi family (Fig. 1A). In addition, SDF-1�triggered a rapid and transient increase in F-actin polymerizationin both cell types (Fig. 1B). These results indicated that CXCR4 onCD4� and RPMI 8866 cells mediated cellular responses leading tocell motility and changes in the organization of actin cytoskeleton.
To investigate whether SDF-1� could influence lymphocyte ad-hesion to MAdCAM-1, we used CHO transfectant cells expressing
5269The Journal of Immunology
by guest on March 27, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from
MAdCAM-1 on their membranes as well as sMAdCAM-1-IgG.Soluble SDF-1� transiently up-regulated the adhesion of humanCD4� and RPMI 8866 cells to CHO-MAdCAM-1 transfectants,whereas the anti-�4�7 Ab Act-1 partially inhibited the increasedadhesion (Fig. 2), indicating that �4�7 was involved in SDF-1�-triggered up-regulation of adhesion and suggesting that additionaladhesion pathways were also activated by SDF-1�. Preincubationof cells with soluble SDF-1� (from 1 to 20 min) did not signifi-cantly influence their subsequent adhesion to sMAdCAM-1-IgG(data not shown). As SDF-1� can be immobilized and displayedon endothelium (27, 30), we investigated whether SDF-1� coim-
mobilized with sMAdCAM-1-IgG could trigger a more effectiveresponse in lymphocyte adhesion to this �4�7 ligand. A robustup-regulation of CD4� and RPMI 8866 cell adhesion was obtainedwhen both SDF-1� and sMAdCAM-1-IgG were immobilized onthe same well, which was completely blocked by Act-1 Ab (Fig.3A), confirming that SDF-1�-enhanced adhesion was totally me-diated by �4�7. The extent of increased adhesion was larger whenbasal adhesion to MAdCAM-1 alone was low. As a control, wellswith SDF-1� alone did not support cell adhesion. In addition,SDF-1� did not induce CD4� or RPMI 8866 cell aggregation (datanot shown), ruling out a possible contribution to the up-regulatedadhesion.
We next determined whether a brief CD4� and RPMI 8866 cellexposure to sMAdCAM-1-IgG immobilized with or without SDF-1�could convey a resistance to detachment under shear stress by mea-suring the extent of cells remaining bound after applying shear flow.We found that activation with SDF-1� notably augmented the per-centage of CD4� and RPMI 8866 cells remaining firmly attached tosMAdCAM-1-IgG after applying wall shear stresses of 0.5–2 dynes/cm2 (Fig. 3B). Preincubation with Act-1 completely abolished theaugmented adhesion, indicating a full involvement of �4�7, and nocells remained bound to SDF-1� alone or FCS.
Naive and memory CD4� T cell subpopulations are both targetsfor SDF-1�-triggered up-regulation of adhesion to MAdCAM-1
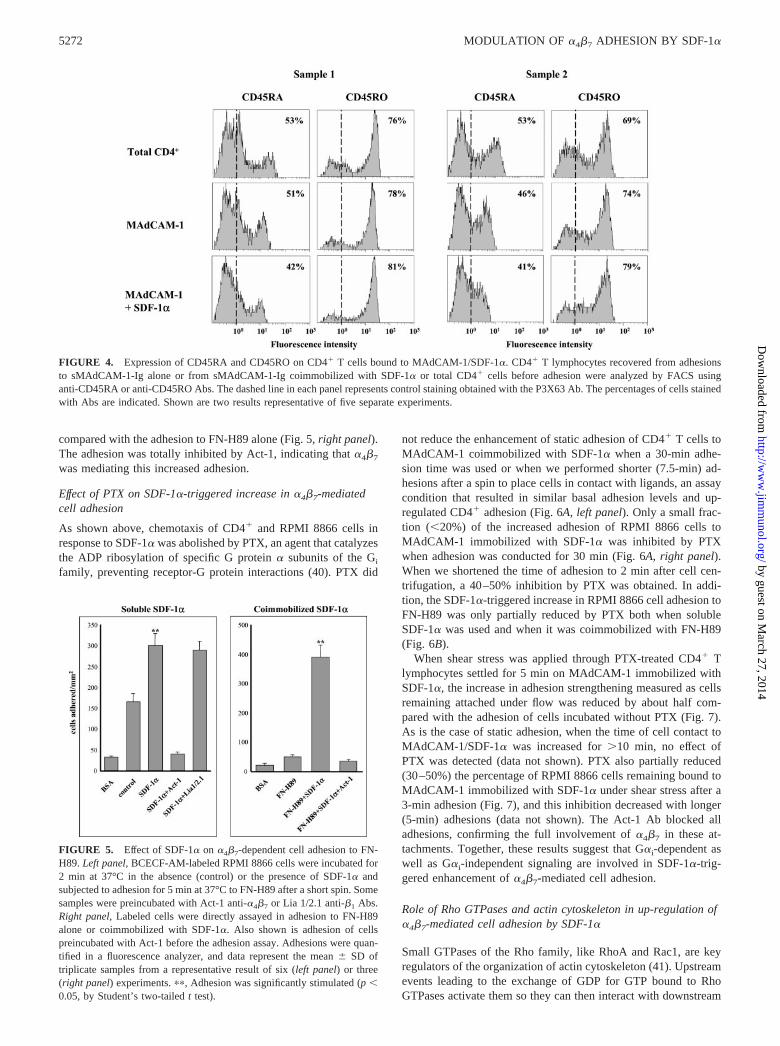
To determine whether SDF-1� was preferentially targeting na-ive CD45RA� or memory CD45RO� phenotype cells among humanCD4� T lymphocytes for increased adhesion to MAdCAM-1, wecompared by FACS analysis cells recovered from adhesion toMAdCAM-1 alone, MAdCAM-1 coimmobilized with SDF-1�, orthe whole cell population before the adhesion assay. The resultsshowed that the proportions of CD45RA� and CD45RO� cells weresimilar inside the CD4� cell population attached to MAdCAM-1 im-mobilized with or without SDF-1�. However, a minor, but consistent,decrease in naive CD45RA� cells and a concomitant slight increasein memory cells in the population recovered from adhesions to co-immobilized SDF-1� and MAdCAM-1 were detected (Fig. 4). Thesedata suggest that SDF-1� targets both memory and naive CD4�
T lymphocytes in the up-regulation of �4�7-mediated adhesion toMAdCAM-1, with a slight preference for memory CD45RO� cells.
FIGURE 1. SDF-1� triggers chemotaxis and F-actin polymerization on human CD4� T lymphocytes and RPMI 8866 cells. A, Cells were preincubatedwith or without PTX and allowed to migrate in Transwell chemotaxis chambers to medium alone (control) or SDF-1�-containing lower chambers. Data representthe mean � SD of two experiments, each performed with duplicate samples. B, Cells were incubated for the indicated times with SDF-1�, stained withFITC-phalloidin, and subjected to flow cytometry. No change in F-actin content was detected when cells were incubated with medium alone (data not shown).
FIGURE 2. SDF-1� increases �4�7-mediated cell adhesion to CHO-MAdCAM-1 transfectants. BCECF-AM-labeled RPMI 8866 or CD4� Tcells were subjected to adhesion for 10 min at 37°C to CHO-MAdCAM-1monolayers. For 5-min samples, SDF-1� (200 ng/ml) was added to wells5 min before the end of adhesion, whereas in the case of 10-min samplesSDF-1� was added from the start of adhesion. For 20-min samples, cellswere preincubated with the chemokine for 10 min at 37°C and then addedto the adhesion assay. Control samples were subjected to adhesion in theabsence of SDF-1�. Also shown is the adhesion of cells preincubated withanti-�4�7 Act-1 Ab (10 �g/ml) before the adhesion assay. Adhesions werequantified in a fluorescence analyzer, and data represent the mean � SD oftriplicate samples from one representative result of six (for RPMI 8866) orthree (for CD4�) independent experiments. ��, Adhesion was significantlystimulated (p � 0.05, by Student’s two-tailed t test).
5270 MODULATION OF �4�7 ADHESION BY SDF-1�
by guest on March 27, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from

SDF-1� increases �4�7-mediated adhesion to CS-1/fibronectin
The CS-1 region of fibronectin represents an additional bindingsite for �4�7 (14, 15). We used RPMI 8866 cells to test the effectof SDF-1� on adhesion to FN-H89, a CS-1-containing fragment offibronectin. A short (2-min) incubation of RPMI 8866 cells with
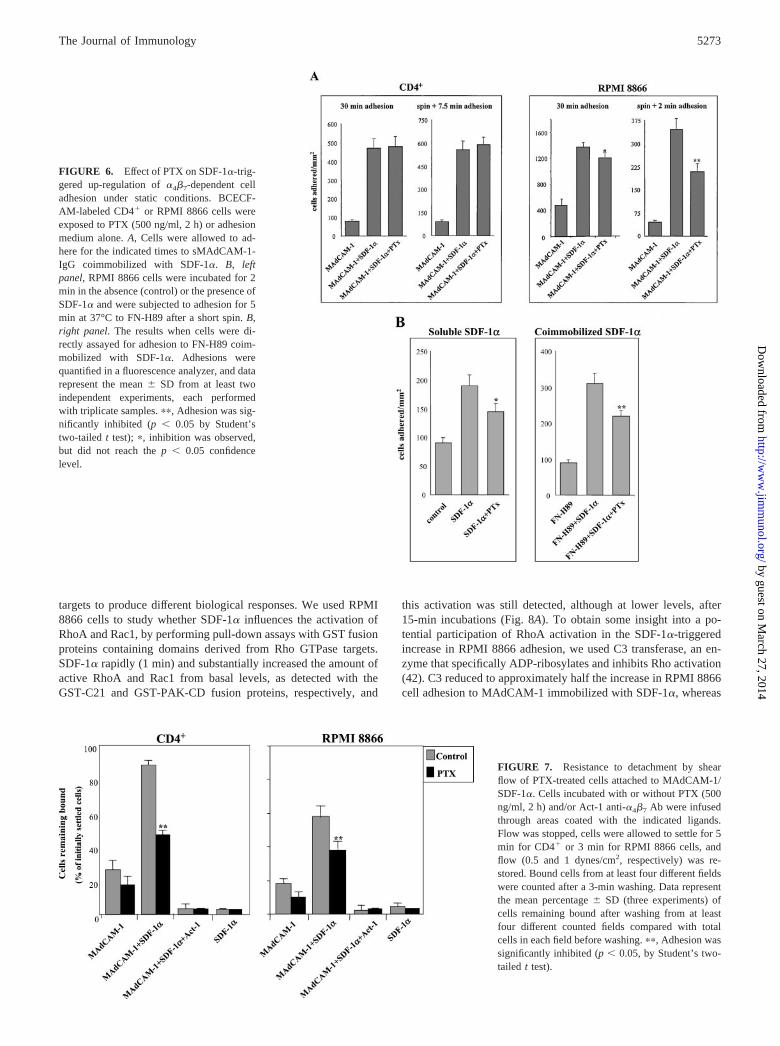
soluble SDF-1� augmented the adhesion to FN-H89, which wasblocked by Act-1, but not by Lia 1/2.1 anti-�1 Ab (Fig. 5, leftpanel). Similar to sMAdCAM-1-IgG, a substantially higher up-regulation of adhesion of RPMI 8866 cells was obtained whenboth FN-H89 and SDF-1� were coimmobilized in the same well
FIGURE 3. SDF-1� coimmobilized with sMAdCAM-1-IgG up-regulates �4�7-dependent cell adhesion. A, Static cell adhesion. SDF-1�, sMAdCAM-1-IgG, or both proteins together were immobilized in adhesion wells as described in Materials and Methods. BCECF-AM-labeled CD4� T lymphocytesor RPMI 8866 cells were subjected to adhesion for 30 min at 37°C. Some samples were preincubated with anti-�4�7 Act-1 (10 �g/ml) Ab before theadhesion assay. Adhesions were quantified in a fluorescence analyzer, and data shown are from two independent experiments. Basal adhesion to wellscoated with BSA alone is also shown. B, Adhesion under flow conditions. CD4� or RPMI 8866 cells incubated with or without Act-1 were infused undershear flow over immobilized SDF-1�, sMAdCAM-1-IgG, or both proteins together. Flow was then stopped, and cells were allowed to settle for 5 min forCD4� or for 3 min for RPMI 8866 cells. Nonbound cells were washed off with adhesion medium at flow rates of 0.5 or 2 dynes/cm2, as indicated by thenumbers in parentheses. Resistance to detachment by shear flow is represented as the mean percentage � SD (three experiments) of cells remaining boundafter 3-min washing from at least four different counted fields compared with the total cells in each field before washing. The percentage of cells remainingbound to FCS is also shown. ��, Adhesion was significantly stimulated (p � 0.05, by Student’s two-tailed t test).
5271The Journal of Immunology
by guest on March 27, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from
compared with the adhesion to FN-H89 alone (Fig. 5, right panel).The adhesion was totally inhibited by Act-1, indicating that �4�7
was mediating this increased adhesion.
Effect of PTX on SDF-1�-triggered increase in �4�7-mediatedcell adhesion
As shown above, chemotaxis of CD4� and RPMI 8866 cells inresponse to SDF-1� was abolished by PTX, an agent that catalyzesthe ADP ribosylation of specific G protein � subunits of the Gi
family, preventing receptor-G protein interactions (40). PTX did
not reduce the enhancement of static adhesion of CD4� T cells toMAdCAM-1 coimmobilized with SDF-1� when a 30-min adhe-sion time was used or when we performed shorter (7.5-min) ad-hesions after a spin to place cells in contact with ligands, an assaycondition that resulted in similar basal adhesion levels and up-regulated CD4� adhesion (Fig. 6A, left panel). Only a small frac-tion (�20%) of the increased adhesion of RPMI 8866 cells toMAdCAM-1 immobilized with SDF-1� was inhibited by PTXwhen adhesion was conducted for 30 min (Fig. 6A, right panel).When we shortened the time of adhesion to 2 min after cell cen-trifugation, a 40–50% inhibition by PTX was obtained. In addi-tion, the SDF-1�-triggered increase in RPMI 8866 cell adhesion toFN-H89 was only partially reduced by PTX both when solubleSDF-1� was used and when it was coimmobilized with FN-H89(Fig. 6B).
When shear stress was applied through PTX-treated CD4� Tlymphocytes settled for 5 min on MAdCAM-1 immobilized withSDF-1�, the increase in adhesion strengthening measured as cellsremaining attached under flow was reduced by about half com-pared with the adhesion of cells incubated without PTX (Fig. 7).As is the case of static adhesion, when the time of cell contact toMAdCAM-1/SDF-1� was increased for �10 min, no effect ofPTX was detected (data not shown). PTX also partially reduced(30–50%) the percentage of RPMI 8866 cells remaining bound toMAdCAM-1 immobilized with SDF-1� under shear stress after a3-min adhesion (Fig. 7), and this inhibition decreased with longer(5-min) adhesions (data not shown). The Act-1 Ab blocked alladhesions, confirming the full involvement of �4�7 in these at-tachments. Together, these results suggest that G�i-dependent aswell as G�i-independent signaling are involved in SDF-1�-trig-gered enhancement of �4�7-mediated cell adhesion.
Role of Rho GTPases and actin cytoskeleton in up-regulation of�4�7-mediated cell adhesion by SDF-1�
Small GTPases of the Rho family, like RhoA and Rac1, are keyregulators of the organization of actin cytoskeleton (41). Upstreamevents leading to the exchange of GDP for GTP bound to RhoGTPases activate them so they can then interact with downstream
FIGURE 5. Effect of SDF-1� on �4�7-dependent cell adhesion to FN-H89. Left panel, BCECF-AM-labeled RPMI 8866 cells were incubated for2 min at 37°C in the absence (control) or the presence of SDF-1� andsubjected to adhesion for 5 min at 37°C to FN-H89 after a short spin. Somesamples were preincubated with Act-1 anti-�4�7 or Lia 1/2.1 anti-�1 Abs.Right panel, Labeled cells were directly assayed in adhesion to FN-H89alone or coimmobilized with SDF-1�. Also shown is adhesion of cellspreincubated with Act-1 before the adhesion assay. Adhesions were quan-tified in a fluorescence analyzer, and data represent the mean � SD oftriplicate samples from a representative result of six (left panel) or three(right panel) experiments. ��, Adhesion was significantly stimulated (p �0.05, by Student’s two-tailed t test).
FIGURE 4. Expression of CD45RA and CD45RO on CD4� T cells bound to MAdCAM-1/SDF-1�. CD4� T lymphocytes recovered from adhesionsto sMAdCAM-1-Ig alone or from sMAdCAM-1-Ig coimmobilized with SDF-1� or total CD4� cells before adhesion were analyzed by FACS usinganti-CD45RA or anti-CD45RO Abs. The dashed line in each panel represents control staining obtained with the P3X63 Ab. The percentages of cells stainedwith Abs are indicated. Shown are two results representative of five separate experiments.
5272 MODULATION OF �4�7 ADHESION BY SDF-1�
by guest on March 27, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from
targets to produce different biological responses. We used RPMI8866 cells to study whether SDF-1� influences the activation ofRhoA and Rac1, by performing pull-down assays with GST fusionproteins containing domains derived from Rho GTPase targets.SDF-1� rapidly (1 min) and substantially increased the amount ofactive RhoA and Rac1 from basal levels, as detected with theGST-C21 and GST-PAK-CD fusion proteins, respectively, and
this activation was still detected, although at lower levels, after15-min incubations (Fig. 8A). To obtain some insight into a po-tential participation of RhoA activation in the SDF-1�-triggeredincrease in RPMI 8866 adhesion, we used C3 transferase, an en-zyme that specifically ADP-ribosylates and inhibits Rho activation(42). C3 reduced to approximately half the increase in RPMI 8866cell adhesion to MAdCAM-1 immobilized with SDF-1�, whereas
FIGURE 7. Resistance to detachment by shearflow of PTX-treated cells attached to MAdCAM-1/SDF-1�. Cells incubated with or without PTX (500ng/ml, 2 h) and/or Act-1 anti-�4�7 Ab were infusedthrough areas coated with the indicated ligands.Flow was stopped, cells were allowed to settle for 5min for CD4� or 3 min for RPMI 8866 cells, andflow (0.5 and 1 dynes/cm2, respectively) was re-stored. Bound cells from at least four different fieldswere counted after a 3-min washing. Data representthe mean percentage � SD (three experiments) ofcells remaining bound after washing from at leastfour different counted fields compared with totalcells in each field before washing. ��, Adhesion wassignificantly inhibited (p � 0.05, by Student’s two-tailed t test).
FIGURE 6. Effect of PTX on SDF-1�-trig-gered up-regulation of �4�7-dependent celladhesion under static conditions. BCECF-AM-labeled CD4� or RPMI 8866 cells wereexposed to PTX (500 ng/ml, 2 h) or adhesionmedium alone. A, Cells were allowed to ad-here for the indicated times to sMAdCAM-1-IgG coimmobilized with SDF-1�. B, leftpanel, RPMI 8866 cells were incubated for 2min in the absence (control) or the presence ofSDF-1� and were subjected to adhesion for 5min at 37°C to FN-H89 after a short spin. B,right panel. The results when cells were di-rectly assayed for adhesion to FN-H89 coim-mobilized with SDF-1�. Adhesions werequantified in a fluorescence analyzer, and datarepresent the mean � SD from at least twoindependent experiments, each performedwith triplicate samples. ��, Adhesion was sig-nificantly inhibited (p � 0.05 by Student’stwo-tailed t test); �, inhibition was observed,but did not reach the p � 0.05 confidencelevel.
5273The Journal of Immunology
by guest on March 27, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from

it totally blocked their up-regulated adhesion to FN-H89 (Fig. 8B),suggesting that activation of RhoA could be involved in the in-creased adhesion. Cytochalasin D, an agent that disrupts actin fil-aments, blocked the up-regulation of F-actin polymerization trig-gered by SDF-1� on CD4� and RPMI 8866 cells (data not shown).Preincubation with cytochalasin D at concentrations of up to 5�g/ml substantially inhibited the enhanced adhesion of CD4� Tlymphocytes without affecting their basal adhesion to MAdCAM-1alone (Fig. 8C). This same concentration partially inhibited (30%)the increase in adhesion of RPMI 8866 cells, and we did not usehigher concentrations because basal adhesion was affected. Theseresults suggest that the actin cytoskeleton plays a relevant role inSDF-1�-induced enhancement of cell adhesion to MAdCAM-1mediated by �4�7.
We also analyzed whether the �4�7/sMAdCAM-1-IgG inter-action itself could trigger additional activation of RhoA and/or
Rac1. In a soluble binding assay (16), RPMI 8866 cells boundsMAdCAM-1-IgG, as detected by flow cytometry using a PE-conjugated anti-human IgG Fc-specific Ab, and the Act-1 Ababolished the binding (data not shown). Pull-down assays per-formed with GST-PAK-CD and GST-C21 fusion proteins usingRPMI 8866 cells incubated with sMAdCAM-1-IgG showed amodest (2-fold) increase in Rac1 activation that was inhibited byblocking �4�7/sMAdCAM-1-IgG binding with Act-1, whereas nosignificant alterations in RhoA activation were detected (Fig. 9).
SDF-1� induces association of CXCR4 with heterotrimeric G�i
and G�13 proteins
Western blot analysis of RPMI 8866 lysates using Abs to sev-eral heterotrimeric G� proteins revealed significant expression ofG�i as well as PTX-resistant G�13 and G�q/11 (data not shown).To characterize G protein association with CXCR4 upon SDF-1�
FIGURE 8. Role of Rho GTPases andactin cytoskeleton in up-regulation of�4�7-mediated cell adhesion by SDF-1�.A, SDF-1� activates RhoA and Rac1 onRPMI 8866 cells. Cells were treated withSDF-1� for the stated times and solubi-lized, and lysates were incubated with glu-tathione agarose-bound fusion proteins todetect active RhoA and Rac1. Beads andproteins bound to fusion proteins wereeluted in Laemmli sample buffer and ana-lyzed for bound RhoA and Rac1 by West-ern blot using anti-RhoA and anti-Rac1Abs. Aliquots of the respective lysatesserved as controls for analyzing the totalamounts of RhoA and Rac1. B, Effect ofC3 on the up-regulation of �4�7-dependentcell adhesion by SDF-1�. RPMI 8866 cellswere exposed to C3 (50 �g/ml, 16 h) oradhesion medium alone (control). After la-beling with BCECF-AM, cells were al-lowed to adhere for 30 min at 37°C tosMAdCAM-1-IgG alone or coimmobi-lized with SDF-1� (left panel) or were in-cubated for 2 min with or without SDF-1�and assayed for 5 min at 37°C in adhesionto FN-H89 (right panel). C, Role of actincytoskeleton in SDF-1�-triggered increasein �4�7-mediated adhesion to MAdCAM-1.BCECF-AM-labeled CD4� or RPMI 8866cells were incubated for 30 min at 37°C inadhesion medium alone (control) or with theindicated amounts of cytochalasin D (Cyt D)and allowed to adhere to sMAdCAM-1-IgGalone or coimmobilized with SDF-1�. Ad-hesions were quantified in a fluorescence an-alyzer, and data represent the mean � SDfrom at least three independent experiments,each performed with triplicate samples. ��,Adhesion was significantly inhibited (p �0.05, by Student’s two-tailed t test); �, inhi-bition was observed but did not reach thep � 0.05 confidence level.
5274 MODULATION OF �4�7 ADHESION BY SDF-1�
by guest on March 27, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from

stimulation, we incubated RPMI 8866 cells with SDF-1�, and celllysates were immunoprecipitated with anti-CXCR4 Abs, followedby Western blotting using anti-G�i and G�13 Abs. The resultsshowed that G�i rapidly and transiently interacted with CXCR4upon SDF-1� activation, with a maximum association detectedbetween 1 and 3 min (Fig. 10). G�13 also associated with CXCR4after SDF-1� stimulation, but with slower kinetics (maximum at�5 min) and to a lesser extent compared with CXCR4/G�i asso-ciation, as longer gel exposures were needed to sufficiently detectG�13. G�i was barely detected associated with CXCR4 when theseexperiments were conducted with PTX-treated cells. Interestingly,higher amounts of G�13 were found associated with CXCR4, andtime kinetics revealed a faster CXCR4/G�13 interaction comparedwith nontreated RPMI 8866 cells (Fig. 10). These data indicatethat G�i and G�13 are involved in SDF-1�-induced RPMI 8866cell activation.
DiscussionIn the present study we have investigated the role of SDF-1� in�4�7-mediated lymphocyte adhesion using human CD4� T lym-phocytes and the lymphoblastoid cell line RPMI 8866 as models.The results indicated that SDF-1� strongly up-regulated their�4�7-dependent adhesion to sMAdCAM-1-Ig as well as their ad-hesion to membrane-bound MAdCAM-1 expressed on CHO trans-fectants. In addition, SDF-1� increased �4�7-mediated RPMI8866 cell attachment to the CS-1-containing FN-H89 fragment offibronectin.
Enhancement of cell adhesion to sMAdCAM-1-Ig was detectedby coimmobilizing SDF-1� with this �4�7 ligand and wasachieved in both static adhesion assays as well as under shearstress by measuring the resistance to cell detachment after apply-ing shear flow. The data suggested that the effect of SDF-1� on�4�7-mediated adhesion to sMAdCAM-1 probably involvedstrengthened cell adhesion to this ligand, leading to firm attach-ment. The rational for using immobilized SDF-1� was based onthe observation that it can be found attached to bone marrow en-dothelium (27) and can be displayed on the surface of HUVEC(30). Although we do not have experimental evidence, it is possi-ble that up-regulation of �4�7-mediated lymphocyte adhesion toCHO-MAdCAM-1 induced by soluble SDF-1� might be due to itsattachment to the surface of CHO-MAdCAM-1 monolayers, whichcould stabilize its binding to CXCR4 on lymphocytes.
The increase in �4�7-mediated CD4� cell adhesion to MAdCAM-1by SDF-1� targeted both naive CD45RA� and memory CD45RO�
subset cells. Memory cells express lower levels of CXCR4 than naivecells (24, 26, 43), but they respond to SDF-1� with equal or evenslightly higher efficacy in up-regulation of adhesion to MAdCAM-1.This could be due to the fact that �4�7
� memory cells express higherlevels of �4�7 than naive cells (11, 12, 44), which might allow themto attach with more strength to MAdCAM-1, enhancing the likelihoodof SDF-1�/CXCR4 binding. These results suggest that the pattern of�4�7 expression among memory and naive subset cells and the sub-sequent �4�7/MAdCAM-1 interaction might be dominant over thesignaling originated upon SDF-1�/CXCR4 binding, which triggersthe increase in cell adhesion.
Chemotaxis of CD4� and RPMI 8866 cells in response toSDF-1� was abolished by PTX, indicating an involvement of het-erotrimeric G proteins of the Gi family. However, their up-regu-lated adhesion to sMAdCAM-1-Ig triggered by SDF-1� was onlypartially affected by PTX when tested on resistance to cell detach-ment under shear flow or in static adhesion assays. These dataindicate that G�i-dependent signaling is involved in the subse-quent increase in �4�7 adhesive activity and suggest that addi-tional, G�i-independent signaling plays a role in SDF-1�-triggeredenhanced adhesion. Reinforcing the involvement of G�i in themodulation of �4�7-mediated cell adhesion, we detected a rapidand transient association with CXCR4 in RPMI 8866 cells incu-bated with SDF-1�.
PTX-insensitive heterotrimeric G proteins include members ofthe G12 and Gq families (45). We show in this work that in RPMI8866 cells SDF-1� stimulated the interaction of CXCR4 withG�13, a G�12 family member, although to a lesser degree and withslower kinetics than CXCR4/G�i association. Interestingly, theamount of CXCR4-associated G�13 upon SDF-1� stimulation wasnotably enhanced in cells treated with PTX, and, as expected, therewas a large reduction in G�i/CXCR4 interaction, raising the pos-sibility that both G�i and G�13 interact with the same or closelylocated binding sites on CXCR4 and that G�13/CXCR4 associa-tion increases after G�i blockade. Alternatively, G�i might influ-ence G�13 binding at a different site on CXCR4, which again couldbe increased upon inhibition of G�i binding. Therefore, the PTX-resistant increase in cell adhesion to MAdCAM-1 in response toSDF-1� could be the result of enhanced G�13 activation and itsinvolvement in this adhesion. A known downstream effector targetof G�13 is p115 RhoGEF (46), a Rho-activating guanine nucleo-tide exchange factor, and it has been reported that activation ofG�13 mimicked the effects of activated forms of Rho on stress fiberformation that was inhibited by C3 (47, 48). In the present workwe show that SDF-1� activated RhoA and Rac1 on RPMI 8866cells, and inhibition of RhoA activation by C3 considerably re-duced the up-regulation of �4�7-dependent adhesion, suggesting
FIGURE 9. Activation of RhoA and Rac1 upon �4�7/MAdCAM-1 in-teraction. RPMI 8866 cells were incubated with medium alone (Control) orwith soluble sMAdCAM-1-Ig in the presence or the absence of the Act-1anti-�4�7 Ab. Cells were solubilized, lysates were incubated with gluta-thione agarose-bound fusion proteins to detect active RhoA and Rac1, andsamples were processed and analyzed as described in Fig. 8.
FIGURE 10. SDF-1� induces association of CXCR4 with G�i andG�13. Untreated (Control) or PTX-treated (500 ng/ml, 2 h) RPMI 8866cells were stimulated with SDF-1� for the indicated times. Cells weresolubilized, and lysates were immunoprecipitated with anti-CXCR4–01Ab and tested in a Western blot with anti-G�i or G�13 Ab as indicated.Equivalent receptor loading was controlled by reprobing each membranewith CXCR4–01 Ab. As a positive control, unprecipitated RPMI 8866 celllysates were tested in a Western blot with G�i or G�13 Ab (lysate). Arepresentative experiment of three performed is shown.
5275The Journal of Immunology
by guest on March 27, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from

that activation of RhoA represents an important point in the signalsleading to increased adhesion. Previous work reported that expres-sion of activated forms of RhoA caused an increase in �4�7-de-pendent T lymphoma adhesion to MAdCAM-1 (49), an observa-tion in the same line as the present findings on the proposed roleof RhoA activation by SDF-1� in the up-regulation of �4�7-me-diated cell adhesion.
Additionally, we found that blockade of SDF-1�-triggered F-actin polymerization in CD4� and RPMI 8866 cells by cytocha-lasin D interfered with their response by increased adhesion toMAdCAM-1. As RhoA and Rac1 are key regulators of actin cy-toskeleton organization, and their activity influences cell motility(41), it is possible that a functional link exists between SDF-1�-induced activation of RhoA and Rac1 and reorganization of actincytoskeleton, which could be associated with the modulation of�4�7-dependent cell adhesion.
Together these data indicate that G�i-dependent signalingplays an important role in the up-regulation of �4�7-dependentcell adhesion to MAdCAM-1 in response to SDF-1� and raisesthe possibility that activation of G�13 could also contribute tothe up-regulated adhesion. The kinetics of CXCR4 associationwith G�i and G�13 suggest that the faster signaling wave leadingto increased adhesion originates from G�i activation. Activation ofG�i results in inhibition of adenylyl cyclases (45), and it has beensuggested that changes in cAMP levels due to modulation ofadenylyl cyclase activity by chemoattractants could representan important point in the regulation of integrin-dependent leu-kocyte adhesion (50). Therefore, alterations in cAMP levels byG�i-activated downstream signaling could also be involved in theSDF-1�-triggered increase in �4�7 adhesive activity. Several stud-ies focused on �4 integrin-mediated adhesion have also determinedthat Gi-dependent signaling is needed to activate this adhesion,based on PTX inhibition (19, 30, 51, 52). A second signaling wave,G�I independent and potentially arising from G�13, could activatedownstream effectors, such as RhoA and perhaps also Rac1, whichcould result in an additional increase in and/or maintenance of�4�7-mediated adhesion, favoring lymphocyte adhesion strength-ening and resistance to detachment under shear stress. Studiesaddressing G�i and G�13 involvement in modulation of �4�7-de-pendent lymphocyte adhesion using mutant forms of these G pro-teins will be of key importance to characterize their participation inthis process. In addition, it has been recently reported that SDF-1�activates Gq, which mediates LFA-1 activation during in vivo mi-gration of T cell hybridoma cells (53). A possible activation of Gq
by SDF-1� in lymphocytes could represent an additional candidatemechanism contributing to the increase in �4�7 adhesive activity.
Reorganization of actin cytoskeleton induced by SDF-1� mightcluster �4�7 on the cell membrane, which might influence theavidity for its ligands. In this regard, it has been proposed thatintegrin clustering plays important roles in changes in the avidityof �4�1-mediated adhesion (30, 54). Additionally, affinity modu-lation of �4�7-dependent adhesion by SDF-1� is not excluded, asthis chemokine can influence �4�1 affinity for VCAM-1 (55). Wealso show here that �4�7/MAdCAM-1 interaction activates Rac1,which could contribute to the maintenance of enhanced lympho-cyte adhesion and resistance to detachment.
The interaction of �4�7 with MAdCAM-1 on Peyer’s patchesand lamina propria HEV represents a key adhesion pathway inlymphocyte homing during recirculation (4). SDF-1� is expressedon most tissues, including secondary lymphoid tissues such as Pey-er’s patches (31), and it has been proposed to be involved in lym-phocyte recirculation (20, 31). SDF-1� displayed by HEV couldbind to CXCR4 on lymphocytes, and our present data indicate thata functional consequence of this interaction is the up-regulation of
�4�7 adhesive activity, which might lead to lymphocyte adhesionstrengthening to MAdCAM-1, representing a mechanism contrib-uting to lymphocyte homing to secondary lymphoid tissues.
The CC chemokine secondary lymphoid tissue chemokine(SLC) (CXCL21) has also been reported to up-regulate �4�7-de-pendent adhesion of human PBL and T cell lines to MAdCAM-1in both static and flow chamber adhesion assays (51). It was foundthat SLC had a greater effect on naive than on memory CD4� Tcells, consistent with a preferential role of this chemokine for thehoming of naive T cells, which has been further demonstrated withthe use of plt mice that lack SLC expression (56–58). BothSDF-1� and SLC are probably involved in lymphocyte homing tosecondary lymphoid tissues from the blood, but the present resultstogether with SLC modulation of �4�7-dependent lymphocyte ad-hesion suggest that SDF-1� might have a lesser role than SLC indefining the specificity of lymphocyte subset targeting.
�4�7 binding to CS-1/FN might be important in gut-hominglymphocyte-matrix interactions within lymphoid tissues (4). In ad-dition, �4�7/FN interaction has been suggested to play importantroles in lymphoma dissemination (59). Consequently, modulationof this adhesion by SDF-1� could contribute to lymphocyte traf-ficking in normal immune surveillance as well as in lymphomainvasion.
Further research directed at characterizing the mechanisms un-derlying chemokine-triggered modulation of �4�7-dependent lym-phocyte adhesion, including the study of G�i and G�13 involve-ment, is required for a better characterization of the molecularevents implicated in lymphocyte homing during recirculation.
AcknowledgmentsWe thank Drs. Francisco Sanchez-Madrid, Angel Corbı, M. Luisa Toribio,Alan Hall, Paul C. Sternweis, and John Collard for reagents. We also thankDrs. Francisco Sanz-Rodrıguez and Jens Stein for their contribution toparts of this work, and Pedro Lastres for help with flow cytometry.
References1. Picker, L. J., and E. C. Butcher. 1992. Physiological and molecular mechanisms
of lymphocyte homing. Annu. Rev. Immunol. 10:561.2. Mackay, C. R. 1993. Homing of naive, memory and effector lymphocytes. Curr.
Opin. Immunol. 5:423.3. Springer, T. A. 1995. Traffic signals on endothelium for lymphocyte recirculation
and leukocyte emigration. Annu. Rev. Physiol. 57:827.4. Butcher, E. C., M. Williams, K. Youngman, L. Rott, and M. J. Briskin. 1999.
Lymphocyte trafficking and regional immunity. Adv. Immunol. 72:209.5. Holzmann, B., B. W. McIntyre, and I. L. Weissman. 1989. Identification of a
murine Peyer’s patch-specific lymphocyte homing receptor as an integrin mole-cule which an � chain homologous to human VLA-4�. Cell 56:37.
6. Erle, D. J., C. Ruegg, D. Sheppard, and R. Pytela. 1991. Complete amino acidsequence of an integrin � subunit (�7) identified in leukocytes. J. Biol. Chem.266:11009.
7. Yuan, Q., W.-M. Jiang, E. Leung, D. Hollander, J. D. Watson, andG. W. Krissansen. 1992. Molecular cloning of the mouse integrin �7 subunit.J. Biol. Chem. 267:7352.
8. Hu, M. C.-T., D. T. Crowe, I. L. Weissman, and B. Holzmann. 1992. Cloning andexpression of mouse integrin �p(�7): a functional role in Peyer’s patch-specificlymphocyte homing. Proc. Natl. Acad. Sci. USA 89:8254.
9. Berlin, C., E. L. Berg, M. J. Briskin, D. P. Andrew, P. J. Kilshaw, B. Holzmann,I. L. Weissman, A. Hamann, and E. C. Butcher. 1993. �4�7 integrin mediateslymphocyte binding to the mucosal vascular addressin MAdCAM-1. Cell 74:185.
10. Streeter, P. R., E. L. Berg, B. T. N. Rouse, R. F. Bargatze, and E. C. Butcher.1988. A tissue-specific endothelial cell molecule involved in lymphocyte homing.Nature 331:41.
11. Schweighoffer, T., Y. Tanaka, M. Tidswell, D. J. Erle, K. J. Horgan,G. E. Ginther, A. I. Lazarovits, D. Buck, and S. Shaw. 1993. Selective expressionof integrin �4�7 on a subset of human CD4� memory T cells with hallmarks ofgut-tropism. J. Immunol. 151:717.
12. Rott, L. S., M. J. Briskin, D. P. Andrew, E. L. Berg, and E. C. Butcher. 1996. Afundamental subdividision of circulating lymphocytes defined by adhesion tomucosal addressin cell adhesion molecule-1. J. Immunol. 156:3727.
13. Williams, M. B., and E. C. Butcher. 1997. Selective homing of naive and memoryT lymphocyte subsets to Peyer’s patches, lymph nodes and spleen. J. Immunol.59:1746.
14. Chan, B. M. C., M. J. Elices, E. Murphy, and M. E. Hemler. 1992. Adhesion tovascular cell adhesion molecule 1 and fibronectin: comparison of �4�1 (VLA-4)and �4�7 on the human B cell line JY. J. Biol. Chem. 267:8366.
5276 MODULATION OF �4�7 ADHESION BY SDF-1�
by guest on March 27, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from

15. Ruegg, C., A. A. Postigo, E. E. Sikorski, E. C. Butcher, R. Pytela, and D. J. Erle.1992. Role of integrin �4�7/�4�P in lymphocyte adherence to fibronectin andVCAM-1 and homotypic cell clustering. J. Cell Biol. 117:179.
16. Ruiz-Velasco, N., M. Guerrero-Esteo, M. J. Briskin, and J. Teixido. 2000. The �4
integrin subunit Tyr187 has a key role in �4�7-dependent cell adhesion. J. Biol.Chem. 275:7052.
17. Higgins, J. M., M. Cernadas, K. Tan, A. Irie, J. Wang, Y. Takada, andM. B. Brenner. 2000. The role of � and � chains in ligand recognition by �7
integrins. J. Biol. Chem. 275:25652.18. Tidswell, M., R. Pachynski, S. W. Wu, S.-Q. Qiu, E. Dunham, N. Cochran,
M. J. Briskin, P. J. Kilshaw, A. I. Lazarovits, D. P. Andrew, et al. 1997. Structure-function analysis of the integrin �7 subunit. J. Immunol. 159:1497.
19. Berlin, C., R. F. Bargatze, J. J. Campbell, U. H. von Andrian, M. C. Szabo,S. R. Hasslen, R. D. Nelson, E. L. Berg, S. L. Erlandsen, and E. C. Butcher. 1995.�4 integrins mediate lymphocyte attachment and rolling under physiologic flow.Cell 80:413.
20. Bleul, C. C., R. Fuhlbrigge, J. M. Casanovas, A. Aiuti, and T. A. Springer. 1996.A highly efficacious lymphocyte chemoattractant, stromal cell-derived factor 1(SDF-1). J. Exp. Med. 184:1101.
21. Bleul, C. C., J. L. Schultze, and T. A. Springer. 1998. B lymphocyte chemotaxisregulated in association with microanatomic localization, differentiation state andB cell receptor engagement. J. Exp. Med. 187:753.
22. D’Apuzzo, M., A. Rolink, M. Loetscher, J. A. Hoxie, I. Clark-Lewis,F. Melchers, M. Baggliolini, and B. Moser. 1997. The chemokine SDF-1, stromalcell-derived factor-1, attracts early stage B cell precursors via the chemokinereceptor CXCR4. Eur. J. Immunol. 27:1788.
23. Bleul, C. C., M. Farzan, H. Choe, C. Parolin, I. Clark-Lewis, J. Sodroski, andT. A. Springer. 1996. The lymphocyte chemoattractant SDF-1 is a ligand forLESTR/fusin and blocks HIV-1 entry. Nature 382:829.
24. Bleul, C. C., L. Wu, J. A. Hoxie, T. A. Springer, and C. R. Mackay. 1997. TheHIV receptors CXCR4 and CCR5 are differentially expressed and regulated onhuman T lymphocytes. Proc. Natl. Acad. Sci. USA 94:1925.
25. Oberlin, E., A. Amara, F. Bachelerie, C. Bessia, J. L. Virelizier,F. Arenzana-Seisdedos, O. Schwartz, J. M. Heard, I. Clark-Lewis, D. F. Legler,et al. 1996. The CXC chemokine SDF-1 is the ligand for LESTR/fusin and pre-vents infection by T-cell-line-adapted HIV-1. Nature 382:833.
26. Forster, R., E. Kremmer, A. Schubel, D. Breitfeld, A. Kleinschmidt, C. Nerl, G.,Bernhardt, and M. Lipp. 1998. Intracellular and surface expression of the HIV-1correceptor CXCR4/fusin on various leukocyte subsets: rapid internalization andrecycling upon activation. J. Immunol. 160:1522.
27. Peled, A., V. Grabovsky, L. Habler, J. Sandbank, F. Arenzana-Seisdedos, I. Petit,H. Ben-Hur, T. Lapidot, and R. Alon. 1999. The chemokine SDF-1 stimulatesintegrin-mediated arrest of CD34� cells on vascular endothelium under shearflow. J. Clin. Invest. 104:1199.
28. Hidalgo, A., F. Sanz-Rodrıguez, J. L. Rodrıguez-Fernandez, R. Albella, C. Blaya,N. Wright, F. Prosper, C. Cabanas, J. C. Gutierrez-Ramos, and J. Teixido. 2001.The chemokine stromal cell-derived factor-1� modulates VLA-4 integrin-depen-dent adhesion to fibronectin and VCAM-1 on bone marrow hematopoietic pro-genitor cells. Exp. Hematol. 29:345.
29. Sanz-Rodrıguez, F., A. Hidalgo, and J. Teixido. 2001. Chemokine stromal cell-derived factor-1� modulates VLA-4 integrin-mediated multiple myeloma celladhesion to CS-1/fibronectin and VCAM-1. Blood 97:346.
30. Grabovsky, V., S. Feigelson, C. Chen, D. A. Bleijs, A. Peled, G. Cinamon,F. Baleux, F. Arenzana-Seisdedos, T. Lapidot, I. van Kooyk, et al. 2000. Sub-second induction of �4 clustering by immobilized chemokines stimulates leuko-cyte tethering and rolling on endothelial vascular cell adhesion molecule 1 underflow conditions. J. Exp. Med. 192:495.
31. Nagasawa, T., K. Tachibana, and K. Kawabata. 1999. A CXC chemokine SDF-1/PBSF: a ligand for a HIV coreceptor, CXCR4. Adv. Immunol. 71:211.
32. Lazarovits, A. I., R. A. Moscicki, J. T. Kurnick, D. Camerini, A. K. Bhan,L. G. Baird, M. Erikson, and R. B. Colvin. 1984. Lymphocyte activation antigens.I. Monoclonal Ab, anti-Act I, defines a new late lymphocyte activation antigen.J. Immunol. 133:1857.
33. Vila-Coro, A., J. M. Rodriguez-Frade, A. Martin de Ana, M. C. Moreno-Ortiz, C.Martınez-A, and M. Mellado. 1999. The chemokine SDF-1� triggers CXCR4receptor dimerization and activates the JAK/STAT pathway. FASEB J. 13:1699.
34. Mould, A. P., J. A. Askari, S. E. Craig, A. N. Garrat, J. Clements, andM. J. Humphries. 1994. Integrin �4�1-mediated melanoma cell adhesion andmigration on vascular cell adhesion molecule-1 (VCAM-1) and the alternativelyspliced IIICS region of fibronectin. J. Biol. Chem. 269:27224.
35. Robledo, M. M., F. Sanz-Rodrıguez, A. Hidalgo, and J. Teixido. 1998. Differ-ential use of VLA-4 and VLA-5 integrins by hematopoietic precursors and my-eloma cells to adhere to TGF-�1-treated bone marrow stroma. J. Biol. Chem.273:12056.
36. Nobes, C. D., and A. Hall. 1995. Rho, rac and cdc42 regulate the assembly ofmulti-molecular focal complexes associated with actin stress fibers, lamellipodiaand filopodia. Cell 81:53.
37. Robledo, M. M., R. A. Bartolome, N. Longo, J. M. Rodrıguez-Frade, M. Mellado,I. Longo, G. N. P. van Muijen, P. Sanchez-Mateos, and J. Teixido. 2001. Ex-pression of functional chemokine receptors CXCR3 and CXCR4 on human mel-anoma cells. J. Biol. Chem. 276:45098.
38. Sander, E. E., S. van Delft, J. P. ten Klooster, T. Reid, R. A. van der Kamen,F. Michiels, and J. G. Collard. 1998. Matrix-dependent Tiam1/Rac signaling inepithelial cells promotes either cell-cell adhesion or cell migration and is regu-lated by phosphatidylinositol 3-kinase. J. Cell Biol. 143:1385.
39. Mellado, M., J. M. Rodriguez-Frade, A. M. Aragay, G. del Real, A. Vila-Coro,A. Martin de Ana, A. Serrano, F. Mayor, Jr., and C. Martınez-A. 1998. Thechemokine MCP-1 triggers tyrosine phosphorylation of the CCR2B receptor andthe JAK2/STAT3 pathway. J. Immunol. 161:805.
40. Fields, T. A., and P. J. Casey. 1997. Signaling functions and biochemical prop-erties of pertussis toxin-resistant G-proteins. Biochem. J. 321:561.
41. Hall, A. 1998. Rho GTPases and the actin cytoskeleton. Science 279:509.42. Aktories, K., U. Braunn, S. Rosener, I. Just, and A. Hall. 1989. The rho gene
product expressed in E. coli is a substrate of botulinum ADP-ribosyltransferaseC3. Biochem. Biophys. Res. Commun. 158:209.
43. Sallusto, F., D. Lenig, C. R. Mackay, and A. Lanzavecchia. 1998. Flexible pro-grams of chemokine receptor expression on human polarized T helper 1 and 2lymphocytes. J. Exp. Med. 187:875.
44. Erle, D. J., M. J. Briskin, E. C. Butcher, A. Garcia-Pardo, A. I. Lazarovits, andM. Tidswell. 1994. Expression and function of the MAdCAM-1 receptor, integrin�4�7, on human leukocytes. J. Immunol. 153:517.
45. Kehrl, J. H. 1998. Heterotrimeric G protein signaling: roles in immune functionand fine-tuning by RGS proteins. Immunity 8:1.
46. Kozasa, T., X. Jiang, M. J. Hart, P. M. Sternweis, W. D. Singer, A. G. Gilman,G. Bollag and P. C. Sternweis. 1998. p115 RhoGEF, a GTPase activating proteinfor G�12 and G�13. Science 280:2074.
47. Buhl, A. M., N. L. Johnson, N. Dhanasekaran, and G. L. Johnson. 1995. G�12 andG�13 stimulate Rho-dependent stress fiber formation and focal adhesion assem-bly. J. Biol. Chem. 270:24631.
48. Fromm, C., O. A. Coso, S. Montaner, N. Xu, and J. S. Gutkind. 1997. The smallGTP-binding protein Rho links G protein-coupled receptors and G�12 to theserum response element and to cellular transformation. Proc. Natl. Acad. Sci.USA 94:10098.
49. Zhang, W. V., Y. Yang, R. W. Berg, E. Leung, and G. W. Krissansen. 1999. Thesmall GTP-binding proteins Rho and Rac induce T cell adhesion to the mucosaladdressin MAdCAM-1 in a hierarchical fashion. Eur. J. Immunol. 29:2875.
50. Laudanna, C., J. J. Campbell and E. C. Butcher. 1997. Elevation of intracellularcAMP inhibits RhoA activation and integrin-dependent leukocyte adhesion in-duced by chemoattractants. J. Biol. Chem. 272:24141.
51. Pachynski, R. K., S. W. Wu, M. D. Gunn, and D. J. Erle. 1998. Secondarylymphoid-tissue chemokine (SLC) stimulates integrin �4�7-mediated adhesion oflymphocytes to mucosal addressin cell adhesion molecule-1 (MAdCAM-1) underflow. J. Immunol. 161:952.
52. Gerszten, R. E., E. A. Garcıa-Zepeda, Y.-C. Lim, M. Yoshida, H. A. Ding,M. A. Gimbrone, Jr., A. D. Luster, F. W. Luscinskas, and A. Rosenzweig. 1999.MCP-1 and IL-8 trigger firm adhesion of monocytes to vascular endotheliumunder flow conditions. Nature 398:718.
53. Soede, R. D. M., I. S. Zeelenberg, Y. M. Wijnands, M. Kamp, and E. Roos. 2001.Stromal cell-derived factor-1-induced LFA-1 activation during in vivo migrationof T cell hybridoma cells requires Gq/11, RhoA, and myosin, as well as Gi andcdc42. J. Immunol. 166:4293.
54. D’Souza-Schorey, C., B. Boettner, and L. Van Aelst. 1998. Rac regulates inte-grin-mediated spreading and increased adhesion of T lymphocytes. Mol. Cell.Biol. 18:3936.
55. Chan, J. R., S. J. Hyduk, and M. I. Cybulsky. 2001. Chemoattractants induce arapid and transient upregulation of monocyte �4 integrin affinity for vascular celladhesion molecule 1 which mediates arrest: an early step in the process of em-igration. J. Exp. Med. 193:1149.
56. Gunn, M. D., S. Kyuwa, C. Tam, T. Kakiuchi, A. Matsuzawa, L. T. Williams, andH. Nakano. 1999. Mice lacking expression of secondary lymphoid organ che-mokine have defects in lymphocyte homing and dendritic cell localization.J. Exp. Med. 189:451.
57. Mori, S., H. Nakano, K. Aritomi, C. R. Wang, M. D. Gunn, and T. Kakiuchi.2001. Mice lacking expression of the chemokines CCL21-ser and CCL19 ( pltmice) demonstrate delayed but enhanced T cell immune responses. J. Exp. Med.193:207.
58. Vassileva, G., H. Soto, A. Zlotnik, H. Nakano, T. Kakiuchi, J. A. Hedrick, andS. A. Lira. 1999. The reduced expression of 6Ckine in the plt mouse results fromthe deletion of one of two 6Ckine genes. J. Exp. Med. 190:1183.
59. Yin, Z., E. Giacomello, E. Gabriele, L. Zardi, S. Aota, K. M. Yamada,B. Skerlavaji, R. Doliana, A. Colombatti, and R. Perris. 1999. Cooperative ac-tivity of �4�1 and �4�7 integrins in mediating human B-cell lymphoma adhesionand chemotaxis on fibronectin through recognition of multiple synergizing bind-ing sites within the central cell-binding domain. Blood 93:1221.
5277The Journal of Immunology
by guest on March 27, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from




















