
Periventricular Preoptic Area Neurons Coactivated with Luteinizing Hormone (LH)-Releasing Hormone...
10
Periventricular Preoptic Area Neurons Coactivated with Luteinizing Hormone (LH)-Releasing Hormone (LHRH) Neurons at the Time of the LH Surge Are LHRH Afferents* WEI WEI LE, KATHIE A. BERGHORN, STEFANIE RASSNICK, AND GLORIA E. HOFFMAN Department of Anatomy and Neurobiology, University of Maryland School of Medicine (W.W.L., G.H.), Baltimore, Maryland 21201; the Laboratory for Pregnancy and Newborn Research, Cornell University (K.A.B.), Ithaca, New York 14853; and the Department of Neuroscience, University of Pittsburgh (S.R.), Pittsburgh, Pennsylvania 15217 ABSTRACT Earlier studies demonstrated coactivation of the periventricular preoptic area (pePOA) with LHRH neurons at the time of an induced or spontaneous LH surge, suggesting that the pePOA might regulate LHRH neurons. To investigate this hypothesis, studies were con- ducted to determine the temporal pattern of pePOA Fos activation during the rat estrous cycle and establish the connections of the pePOA neurons with LHRH neurons. Fos activation within LHRH and pePOA neurons showed the same temporal pattern. Both were absent during diestrous I, diestrous II, and the morning of proestrus. Fos was induced in the pePOA and LHRH neurons beginning on the afternoon of proestrus (4 h before lights off), with a decline 8 h later on proestrous evening. Tract-tracing studies then established the relationship between LHRH and pePOA neurons. Retrograde label- ing with fluorogold determined that a portion of the Fos-positive pePOA neurons present at the time of the LH surge sent a projection to regions that contain LHRH cells. Anterograde tracer (neurobiotin) injections established that the pePOA neurons sent axons to the LHRH cells. Taken together, these data indicate that the pePOA provides direct input to LHRH neurons that is likely to stimulate LHRH neurons at the time of the LH surge. (Endocrinology 140: 510 –519, 1999) T HE PERIVENTRICULAR zone of the preoptic area (pePOA) is one of a number of sexually dimorphic nuclear regions, with neuronal density greater in the female than in the male (1). Calculations of cell number for any of the phenotypically identified neurons found in that region [e.g. dopamine (2), atrial natriuretic peptide (3), cholecysto- kinin (4), and neurotensin (5)] show greater numbers of neu- rons in females than in males. A number of sexually dimorphic functions, including sex- ual posturing (6, 7), maternal behavior (8, 9), and the ability to display cyclic LH surges (10, 11, 12), are associated with subdivisions of the preoptic area. Studies with the use of Fos as an activity marker in preoptic area neurons during sexual behavior (13–16) have led to the conclusion that the medial preoptic nucleus, centrally located within the POA, is a part of the circuitry subserving reproductive behaviors. A similar role for the pePOA in these same behaviors has not been demonstrated. Previous studies noted that after estrogen (17) or estrogen plus progesterone (16) treatment, neurons along the POA’s periventricular zone strongly expressed Fos-like proteins. The times that Fos proteins were induced coincided with the times LH surges would be evoked, suggesting that activation of the pePOA might be linked to stimulation of LHRH neu- rons. This feature is further supported by our recent studies demonstrating that immature animals treated with estrogen and progesterone so as to induce a LH surge had simulta- neous activation of both LHRH and pePOA neurons; by contrast, animals treated with steroids in a paradigm where LH surges were inhibited failed to show either LHRH or pePOA Fos activation (18). That the pePOA might direct activity in LHRH neurons is further suggested by anatomical studies in which degenerating axon terminals synapsing onto LHRH neurons were observed after neurotoxin (6-hy- droxydopamine) lesions of the preoptic area (19). As most of the dopamine neurons of the POA are located within the pePOA (2), these data raise the likelihood that the pePOA participates directly in LHRH control. The literature cited above supports the idea that the pePOA actively participates in the LH surge mechanism, but fails to address the question of whether pePOA activation is synchronized to LHRH activation and whether there exist direct connections of activated pePOA neurons with LHRH neurons. To address these questions, we conducted two stud- ies. One examined the temporal pattern of Fos induction in neurons of the pePOA in cycling rats and compared those results to LHRH cell activation patterns to test the hypothesis that the pePOA activation was synchronized to that of LHRH neurons. A second series of experiments used tract tracing to determine whether neurons retrogradely labeled from re- gions that contain LHRH neurons were activated at the time Received May 4, 1998. Address all correspondence and requests for reprints to: Gloria E. Hoffman, Ph.D., Department of Anatomy and Neurobiology, University of Maryland, Room 222 HSF, 685 West Baltimore Street, Baltimore, Maryland 21201. E-mail: [email protected]. * This work was supported by Grant NS-28730. 0013-7227/99/$03.00/0 Vol. 140, No. 1 Endocrinology Printed in U.S.A. Copyright © 1999 by The Endocrine Society 510
-
Upload
independent -
Category
Documents
-
view
5 -
download
0
Transcript of Periventricular Preoptic Area Neurons Coactivated with Luteinizing Hormone (LH)-Releasing Hormone...

Periventricular Preoptic Area Neurons Coactivated withLuteinizing Hormone (LH)-Releasing Hormone (LHRH)Neurons at the Time of the LH Surge Are LHRHAfferents*
WEI WEI LE, KATHIE A. BERGHORN, STEFANIE RASSNICK, AND
GLORIA E. HOFFMAN
Department of Anatomy and Neurobiology, University of Maryland School of Medicine (W.W.L., G.H.),Baltimore, Maryland 21201; the Laboratory for Pregnancy and Newborn Research, Cornell University(K.A.B.), Ithaca, New York 14853; and the Department of Neuroscience, University of Pittsburgh (S.R.),Pittsburgh, Pennsylvania 15217
ABSTRACTEarlier studies demonstrated coactivation of the periventricular
preoptic area (pePOA) with LHRH neurons at the time of an inducedor spontaneous LH surge, suggesting that the pePOA might regulateLHRH neurons. To investigate this hypothesis, studies were con-ducted to determine the temporal pattern of pePOA Fos activationduring the rat estrous cycle and establish the connections of thepePOA neurons with LHRH neurons. Fos activation within LHRHand pePOA neurons showed the same temporal pattern. Both wereabsent during diestrous I, diestrous II, and the morning of proestrus.Fos was induced in the pePOA and LHRH neurons beginning on the
afternoon of proestrus (4 h before lights off), with a decline 8 h lateron proestrous evening. Tract-tracing studies then established therelationship between LHRH and pePOA neurons. Retrograde label-ing with fluorogold determined that a portion of the Fos-positivepePOA neurons present at the time of the LH surge sent a projectionto regions that contain LHRH cells. Anterograde tracer (neurobiotin)injections established that the pePOA neurons sent axons to theLHRH cells. Taken together, these data indicate that the pePOAprovides direct input to LHRH neurons that is likely to stimulateLHRH neurons at the time of the LH surge. (Endocrinology 140:510–519, 1999)
THE PERIVENTRICULAR zone of the preoptic area(pePOA) is one of a number of sexually dimorphic
nuclear regions, with neuronal density greater in the femalethan in the male (1). Calculations of cell number for any ofthe phenotypically identified neurons found in that region[e.g. dopamine (2), atrial natriuretic peptide (3), cholecysto-kinin (4), and neurotensin (5)] show greater numbers of neu-rons in females than in males.
A number of sexually dimorphic functions, including sex-ual posturing (6, 7), maternal behavior (8, 9), and the abilityto display cyclic LH surges (10, 11, 12), are associated withsubdivisions of the preoptic area. Studies with the use of Fosas an activity marker in preoptic area neurons during sexualbehavior (13–16) have led to the conclusion that the medialpreoptic nucleus, centrally located within the POA, is a partof the circuitry subserving reproductive behaviors. A similarrole for the pePOA in these same behaviors has not beendemonstrated.
Previous studies noted that after estrogen (17) or estrogenplus progesterone (16) treatment, neurons along the POA’speriventricular zone strongly expressed Fos-like proteins.The times that Fos proteins were induced coincided with thetimes LH surges would be evoked, suggesting that activation
of the pePOA might be linked to stimulation of LHRH neu-rons. This feature is further supported by our recent studiesdemonstrating that immature animals treated with estrogenand progesterone so as to induce a LH surge had simulta-neous activation of both LHRH and pePOA neurons; bycontrast, animals treated with steroids in a paradigm whereLH surges were inhibited failed to show either LHRH orpePOA Fos activation (18). That the pePOA might directactivity in LHRH neurons is further suggested by anatomicalstudies in which degenerating axon terminals synapsingonto LHRH neurons were observed after neurotoxin (6-hy-droxydopamine) lesions of the preoptic area (19). As most ofthe dopamine neurons of the POA are located within thepePOA (2), these data raise the likelihood that the pePOAparticipates directly in LHRH control.
The literature cited above supports the idea that thepePOA actively participates in the LH surge mechanism, butfails to address the question of whether pePOA activation issynchronized to LHRH activation and whether there existdirect connections of activated pePOA neurons with LHRHneurons. To address these questions, we conducted two stud-ies. One examined the temporal pattern of Fos induction inneurons of the pePOA in cycling rats and compared thoseresults to LHRH cell activation patterns to test the hypothesisthat the pePOA activation was synchronized to that of LHRHneurons. A second series of experiments used tract tracing todetermine whether neurons retrogradely labeled from re-gions that contain LHRH neurons were activated at the time
Received May 4, 1998.Address all correspondence and requests for reprints to: Gloria E.
Hoffman, Ph.D., Department of Anatomy and Neurobiology, Universityof Maryland, Room 222 HSF, 685 West Baltimore Street, Baltimore,Maryland 21201. E-mail: [email protected].
* This work was supported by Grant NS-28730.
0013-7227/99/$03.00/0 Vol. 140, No. 1Endocrinology Printed in U.S.A.Copyright © 1999 by The Endocrine Society
510

of the LHRH surge, and whether anterograde labeling ofpePOA axons came into close contact with LHRH neurons.
Materials and MethodsAnimals
Adult female Sprague-Dawley rats purchased from Zivic-Miller Lab-oratories, Inc. (Zelienople, PA) were kept under standard laboratoryconditions (12-h light, 12-h dark cycles, with lights on at 0400 h), withfood and water available ad libitum. The University Committee on An-imal Research approved all protocols. Daily vaginal smears performedat 1000 h determined the estrous cycle stage.
For the experiments examining the coordination of LHRH andpePOA neuronal Fos activation, 29 rats exhibiting 3 consecutive 4-dayestrous cycles were chosen for further study. Animals were killed atvarious times during the estrous cycle, as indicated in Table 1, after beinganesthetized with an overdose of pentobarbital (100 mg/kg). All animalswere administered 1000 U heparin, and a blood sample was takendirectly from the heart for determination of plasma LH values. Subse-quently, each rat was perfused transcardially with saline containing 2%sodium nitrite followed by 2.5% acrolein in buffered 4% paraformal-dehyde (20). All brains were sunk in 25% aqueous sucrose and sectionedon a Reichert AO freezing microtome (AO Instruments, Buffalo, NY) intoa 1 in 12 series of 25-mm sections. Sections were stored in cryoprotectant(21) until staining for Fos and LHRH was initiated.
For determining whether activated pePOA neurons could be retro-gradely labeled from collections of LHRH neurons, a group of cyclingrats (n 5 15) was injected with the retrograde tracer fluorogold (FAu,Fluorochrome, Englewood, CO) placed into the POA at sites whereLHRH neurons are most numerous [adjacent to the organum vasculo-sum of the lamina terminalis (OVLT) or within 400 mm caudal to theOVLT along the borders of the medial and lateral preoptic areas] (22).These rats were anesthetized with Equithesin anesthesia (0.33 ml/100 gbody weight ip of a solution of sodium pentobarbital 0.98 g/dl, chloralhydrate 4.25 g/dl, and MgSO4 2.12 g/dl) and placed in a stereotaxicframe (David Kopf Instruments). Glass capillaries with a tip od of 20–40mm were filled with 0.9% NaCl solution containing 4% FAu and loweredstereotaxically into LHRH cell-containing regions (23) aided by the Paxi-nos and Watson atlas (24). Iontophoretic injections were made into thePOA using a constant positive current of approximately 3.9–4.0 mAthrough the capillary. Injection times were varied between 30 sec and 2min to achieve a minimal degree of dye spread (,200 mm) into theinjected area. Animals were allowed to recover for a minimum of 14days, which is sufficient to allow retrograde transport of the dye to allcell bodies with terminals within the injection site. Two or 3 days aftertracer application, vaginal smears were again monitored. After three
consecutive cycles, animals were selected on proestrus at the time ofexpected peak LH secretion, anesthetized, and killed.
A second series of cycling female rats (n 5 17) received stereotaxicinjections of the anterograde tracer neurobiotin (Vector Laboratories,Inc., Burlingame, CA) placed into the pePOA. Glass capillaries with a tipof 20–40 mm od were filled with 0.9% NaCl solution containing 5%neurobiotin and lowered stereotaxically into the pePOA, 300–600 mmcaudal from the OVLT [aided by use of a stereotaxic atlas (24)] underpentobarbital anesthesia. The capillaries were connected in a fashionidentical to that used for FAu injection; injection times are varied be-tween 5–10 min (with current delivered 10 sec on, 10 sec off) to achievea minimal degree of dye spread (,100 mm) into the injected area. Aftera 24-h survival time, the animals were anesthetized and perfused witha 4% paraformaldehyde-2.5% acrolein solution. The brains were re-moved, sunk in 25% aqueous sucrose, and stored in cryoprotectantsolution until they were stained for double labeling of LHRH andneurobiotin.
Immunocytochemistry
Fos and LHRH. Staining for Fos in combination with LHRH used es-sentially the same strategy as that previously reported (20, 25–31).Briefly, the sections (from a 1 in 12 series) for immunocytochemicalstaining were removed from the cryoprotectant, rinsed in PBS, treatedwith a 1% NaBH4 solution (Sigma Chemical Co., St. Louis, MO), rinsed,and then incubated with anti-Fos antibody (Oncogene Science, Inc.,Tarzana, CA; AB-2; 1:50,000, or 0.02 mg/ml) in PBS with 0.4% TritonX-100 for 48 h at 4 C. After rinsing, the tissue was incubated for 1 h atroom temperature in biotinylated goat antirabbit IgG (heavy and lightchains; Vector Laboratories, Inc.) at a concentration of 1:600 in PBS with0.4% Triton X-100, rinsed, and incubated for 1 h in avidin-biotin complexsolution (ELITE ABC kit, Vector Laboratories, Inc.; 4.5 ml each/ml in-cubation mixture). After rinsing first in PBS and then in 0.175 m sodiumacetate (NaOAc), the Fos antibody-peroxidase complex was visualizedwith a solution of NiSO4 (25 mg/ml), 3,3-diaminobenzidine HCl(NiDAB; 0.2 mg/ml), H2O2 (0.83 ml of a 3% solution/ml final mixture)in aqueous 0.175 m NaOAc that yielded a blue-black reaction product.After approximately 15–20 min, the tissue was transferred into theacetate solution to stop the reaction, rinsed in PBS, and then placed intoanti-LHRH (LR-1, gift from Drs. Benoit and Guillemin, 1:100,000 forfluorescence double labeling using amplified biotin techniques or1:150,000 for conventional double immunoperoxidase labeling) usingthe protocols described by Lee et al. (31) and Hoffman et al. (20), re-spectively. The former provided a means by which the pePOA andLHRH activation patterns could be independently determined; the latteroffered a means of retaining permanently stained sections. After com-
TABLE 1. Relationship between LHRH and pePOA activation during the estrous cycle
Cycle stage nTimea
Plasma LH (ng/ml RP-3) LHRH/Fos pePOA/FosTime relative to lights off Clock time
Diestrous I 1 6 h before 1000 h 1.06 0 2Diestrous I 4 1–2 h after 1700–1800 h 0.73 6 0.08 0 2
Diestrous II 2 6 h before 1000 h 0.70 1 0.09 0 2Diestrous II 2 1–2 h after 1700–1800 h 0.64b 0 2
Proestrus 9 10 h after (DII) to 6 h before (Pro) 0200–1000 h 1.01 1 0.12 0 to (1) 2Proestrus 4 4 h before 1200 h 2.76 1 0.7 0 to 11 1 to 11Proestrus 4 2 h before 1400 13.82 1 2.71 11 to 111 11 to 111Proestrus 4 Time of lights off 1600 16.66 1 5.37 111 11 to 111
Proestrus 4 1–2 h after 1700–1800 12.35 1 3.93 111 111Proestrus 2 4 h after 2000 6.28 1 3.25 1 to 111 0 to 11
This table shows the temporal relationship of patterns of Fos activation in LHRH and pePOA neurons at the times indicated during the estrouscycle. Times are expressed both in clock time and hours relative to the time of lights off.
Scales for Fos activation were: 0, no cells activated; 2, occasional cell activated; (1), only one cell activated; 1, few cells consistently activated(up to one third the maximum); 11, one to two thirds of the maximal activation; 111, two thirds of maximal activation.
a Lights were on from 0400–1600 h (EST).b LH values could not be determined for one of the rats.
pePOA CONNECTIONS WITH LHRH NEURONS 511

pletion of staining, sections were rinsed in normal saline, mounted ontosubbed slides, dried overnight, dehydrated through alcohols, cleared,and coverslipped.
Determination of the FAu injection site. Staining for FAu was accomplishedusing single immunoperoxidase methods on a 1 in 12 series of sections.The strategy was essentially the same as that outlined above for Fos, butwith anti-FAu antibody rather than anti-Fos. Anti- FAu, a gift from Dr.H. T. Chang, University of Tennessee (Memphis, TN) (32) and laterpurchased from Chemicon (Temecula, CA), was used at a concentrationof 1:50,000, in PBS with 0.4% Triton X-100, for 48 h 4 C. After stainingfor approximately 15–20 min in NiDAB solution, the tissue was trans-ferred into the acetate solution to stop the reaction, rinsed in normalsaline, and mounted onto subbed glass slides. The sections were dehy-drated through alcohols, cleared in Histoclear, and coverslipped withHistomount. Some sections were counterstained with neutral red to aidin cytoarchitectonic nuclear identification or were double labeled withLHRH (using the same strategy as the Fos/LHRH studies) to betterdetermine whether the LHRH population was encompassed by theinjection.
Fos and retrograde tracing. To determine whether retrogradely labeledpePOA neurons were activated at the time of a LH surge, sections (froma 1 in 6 series of sections) were double stained for Fos and FAu. In thisinstance, Fos was stained first using the immunoperoxidase proceduredescribed previously for Fos and LHRH, with NiDAB as the chromogen.Next, FAu was stained with immunoperoxidase procedures and dia-minobenzidene (DAB) as the chromogen (as described above for LHRH)or with biotin-amplified immunofluorescence (33) using a streptavidin-bodipy fluorophore. The latter method consisted of the following steps:after reacting the sections for Fos using NiDAB, the tissue was rinsed inacetate solution followed by a few rinses in PBS and incubated in anti-FAu antibody (1:70,000) for 48 h at 4 C. The tissue was rinsed in PBS andincubated in biotinylated goat antirabbit serum (Vector Laboratories,Inc.; 1:5,000 in PBS with 0.4% Triton X-100) for 1 h at room temperature.After rinsing, the tissue was incubated for 30 min with ELITE avidin-biotin complex reagents (1.125 ml each/ml mixture in PBS with 0.4%Triton X-100), rinsed again for 30 min, and incubated for 15 min inbiotinylated tyramine [prepared as previously described (34)] to whichH2O2 was added to achieve a final concentration of 0.005%. The tissuewas rinsed for 30 min in PBS and then incubated in streptavidin-Bodipy(5 ml streptavidin-Bodipy/ml PBS with 0.4% Triton X-100) for 2.5 h at37–40 C. The tissue was rinsed in saline, mounted, air-dried overnight,cleared in xylenes, and coverslipped in Histomount (Fisher Scientific,Rockford, IL).
Triple labeling of Fos, LHRH, and FAu. In a few cases we performed triplelabeling of sections to verify that the FAu injections encompassed re-gions containing activated LHRH neurons, and that the activatedpePOA neurons projected to the vicinity of LHRH neurons, as deter-mined by the presence of FAu within Fos-positive pePOA neurons. Forthis procedure (35) the nuclear antigen Fos is first stained using immu-noperoxidase methods and NiDAB as the chromogen, as describedabove. Transmitter (in this case, LHRH) is next stained with an immun-ofluorescence-alkaline phosphatase procedure described by van derLoos (36). Briefly, after staining for Fos and incubating in anti-LHRH(1:70,000), the tissue is rinsed and then incubated with goat-antirabbitIgG conjugated to alkaline phosphatase (Vector Laboratories, Inc.; 1:500for 2 h at room temperature), rinsed in Tris buffer (pH 8.0) for 15 min,and incubated with CAS Red solution (Cell Analytical Systems,Elmhurst, IL) containing levamisole (1 mm) according to manufacturerinserts. [It should be noted that Cell Analytical Systems is no longermaking CAS Red; Vector Red (Vector Laboratories, Inc.) prepared ac-cording to product inserts can be substituted.] After a 1- to 3-h incubationin the staining solution, the tissue is rinsed in distilled water followedby PBS, incubated in anti-FAu (1:70,000), and processed in the fashiondescribed for biotin-amplified immunofluorescence above (usingstreptavidin-Bodipy as the fluorophore). After the final reaction, thetissue was rinsed in PBS mounted from saline, air-dried overnight,dehydrated through ascending alcohols, cleared in xylenes, and cover-slipped with Histomount. The presence of FAu was evident as a greenfluorescent product; LHRH immunoreactivity within the cell cytoplasmwas barely detected as pink reaction product with brightfield optics, butwas brightly red/orange fluorescent under fluorescence optics; Fos was
stained blue-black. LHRH neurons were considered within the area ofFAu uptake if diffuse FAu product surrounded the LHRH cells.
LHRH and anterograde tracer, neurobiotin. Labeling of neurobiotin wasaccomplished immunocytochemically with antibiotin antibodies.Briefly, after rinsing the cryoprotectant from the tissue, the sections(from a 1 in 6 series of sections) were treated with sodium borohydride,rinsed, and incubated with antibiotin (1:60,000; Vector Laboratories,Inc.) for 48 h at 4 C. After incubation with the antibiotin, the tissue wasrinsed in PBS, and the sections were incubated in biotinylated rabbitantigoat IgG and avidin-biotin complex reagents as described above.Sections were then rinsed in PBS followed by NaOAc solution. Thestaining of neurobiotin was accomplished by using a mixture of H2O2(0.83 ml of a 3% solution/ml reaction solution) and NiDAB. After stain-ing for 7–10 min, the tissue was transferred to acetate solution to stopthe reaction, rinsed in PBS, incubated in anti-LHRH (1:150,000), andstained with either DAB or amplified fluorescent labeling as describedfor the Fos and LHRH double labeling above. Axons labeled for neu-robiotin appeared blue-black; LHRH was stained either brown whenDAB was used or fluoresced red with the fluorophore, Texas Red.
Tissue analysis
Retrogradely labeled cells or cells stained for Fos and/or LHRH andthe extent of the tracer injection were plotted using stage-mounted X-Ypotentiometers mounted on a Nikon fluorescence microscope (Melville,NY) linked to a Macintosh II computer (37).
Analyses of the interactions of neurobiotin-containing axons andLHRH were qualitative. The criteria for considering a LHRH neuroncontacted by a neurobiotin-positive axon were those established in thereport by Fitzsimmons et al. (38) and required that no visible space bepresent interposed between the biotin-immunoreactive axon and theLHRH neuron at a magnification of 3600.
LH RIA
The RIA protocol we used for plasma LH has been previously de-scribed (39, 40). Plasma volumes of 20 and 5 ml from each animal wereassayed for each sample. For the LH assay, the rat LH CSU-120 antibodywas used. Standard curves were constructed using the rat LH RP-3standard from the National Hormone and Pituitary Program of theNIDDK.
ResultsFos expression in LHRH neurons and that in pePOAneurons were coordinated during the LH surge
No increased pePOA Fos or LHRH-Fos activation wasnoted at any time on diestrous I or II or during the morningof proestrus when LH values were low (Table 1 and Figure1). As expected (27–29), animals killed on the afternoon ofproestrus showed marked expression of Fos within LHRHneurons that first appeared approximately 8 h after lights onwhen the rising phase of the LH surge was initiated. Eachanimal displaying an increase in plasma LH values showedactivation of LHRH neurons, and there was a coordinatedincrease in pePOA Fos staining as well. The pePOA Fospattern at the peak of the LH surge lay within the region100–150 mm lateral to the third ventricular surface that wasmaximal 300–450 mm caudal to the OVLT (Figure 2). At therostral pole of the third ventricle (where LHRH neuron num-bers are maximal) few Fos-positive cells were found in theperiventricular zone (Fig. 2A). At the more caudal pePOAlevels (Fig. 2D), the numbers of Fos-positive neurons de-clined in the pePOA. By the beginning of the hypothalamus,about 150 mm caudal to level D, Fos-positive cells in theperiventricular zone were no longer present (not shown).
512 pePOA CONNECTIONS WITH LHRH NEURONS Endo • 1999Vol 140 • No 1

Retrograde tracer injections into regions containing LHRHneurons labeled pePOA neurons, some of which were Fosactivated at the time of a LH surge
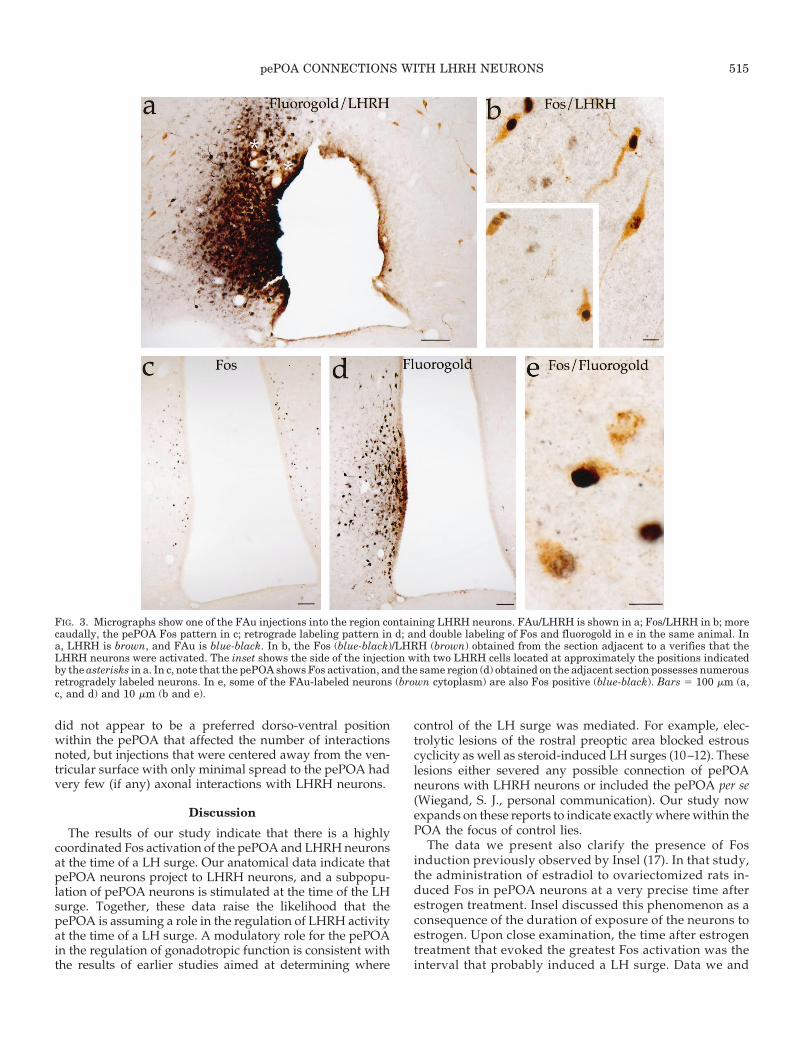
To determine the likelihood that pePOA neurons activatedat the time of a LH surge innervated LHRH neurons, we firstmade small iontophoretic injections of FAu into the sites ofLHRH neurons and then determined whether the retro-gradely labeled neurons expressed Fos at the time of a LHsurge. Of the 15 animals injected, 7 possessed sites of injec-tion that encompassed LHRH neurons but did not extendinto the pePOA. The 7 injection sites were approximately200–400 mm in diameter. Most were spheres, but in 1 casewhere the injection lay in the ventrolateral cell field of LHRHneurons, the injection spread caudally with the LHRH pop-ulation for about 600 mm but did not spread dorsally ormedially. In each case where LHRH neurons were present inthe injection site, a collection of retrogradely labeled neuronswas noted in the pePOA. An example of a typical injectionis shown in Fig. 3a. Staining of the adjacent section (Fig. 3b)for Fos and LHRH revealed that Fos-activated LHRH neu-rons were present within the injection site and that the
pePOA neurons expressed Fos, as expected (Fig. 3c). Withtriple labeling (Fig. 4) we could verify that the injection sitespossessed activated LHRH neurons.
The pattern of retrograde labeling within the pePOA fromanimals with injections that included LHRH neurons invari-ably revealed a population of neurons located within thepePOA (Figs. 3d and 4, E and G). Although in some animalsthe ventricular surface was infused with FAu, the ependymallining was generally not torn. The labeling within the pePOAwas probably not the result of disruption of the ependymallining or leakage into the ventricle, as the locations of theretrogradely labeled neurons indicated that the projectionsfrom the pePOA to the LHRH neurons were essentially un-crossed and if derived from the ventricle would have beenbilateral. In some injections, staining of FAu did not saturatethe ependymal cells, indicating that the tracer did not reachthe ventricle, and in those animals, the general pePOA FAulabeling pattern was not different from that in animals thatdid show ependymal FAu saturation.
Measurement of LH from plasma obtained at the time ofdeath and examination of Fos and LHRH staining verified
FIG. 1. Micrographs of Fos expressionwithin pePOA neurons (brightfield im-ages) and LHRH neurons (insets) before(a–c) and during (d) a LH surge. In therats killed before the LH surge, theLHRH neurons are devoid of Fos, andthe pePOA displays few Fos-positivenuclei, whereas at the time of the LHsurge (d), LHRH neurons (inset) andnumerous neurons of the pePOA (ar-rows) are activated to express Fos.Bar 5 100 mm (brightfield images) and10 mm (fluorescence images).
pePOA CONNECTIONS WITH LHRH NEURONS 513

that each of the FAu-injected animals displayed a proestrousLH surge, LHRH Fos activation, and pePOA Fos activation.The presence of the tracer did not hinder the animal’s abilityto show Fos activation in either LHRH neurons or the pePOA(as determined by symmetry of staining). Double labeling ofFos and FAu (Figs. 3e and 4G) established that a subset of theretrogradely labeled pePOA neurons was Fos activated. Al-though as many as 25–50% of the pePOA retrogradely la-beled neurons expressed Fos, the numbers of pePOA neu-rons that were Fos positive after any one injectioncomprised only a small fraction of the total Fos-activatedpePOA population. In considering this feature, one mustbe aware that the LHRH neurons are so widely scatteredthat injections encompassing large numbers of LHRH neu-rons could not be made without the FAu extending into thepePOA (which lies only 300 mm from the largest collectionof LHRH neurons). Thus, as is shown in Fig. 4, individualinjections, although including regions where LHRH neu-rons were present, encompassed only small numbers ofLHRH neurons, and the double labeled pePOA neuronsconstituted only a small portion of the total Fos-activatedpePOA population (Fig. 4, E and G). Nonetheless, the
presence of even a few activated retrogradely labeledpePOA neurons demonstrates that sites of LHRH neuronsreceive projections from the pePOA.
Anterograde labeling verified that projections from thepePOA extended to LHRH neurons
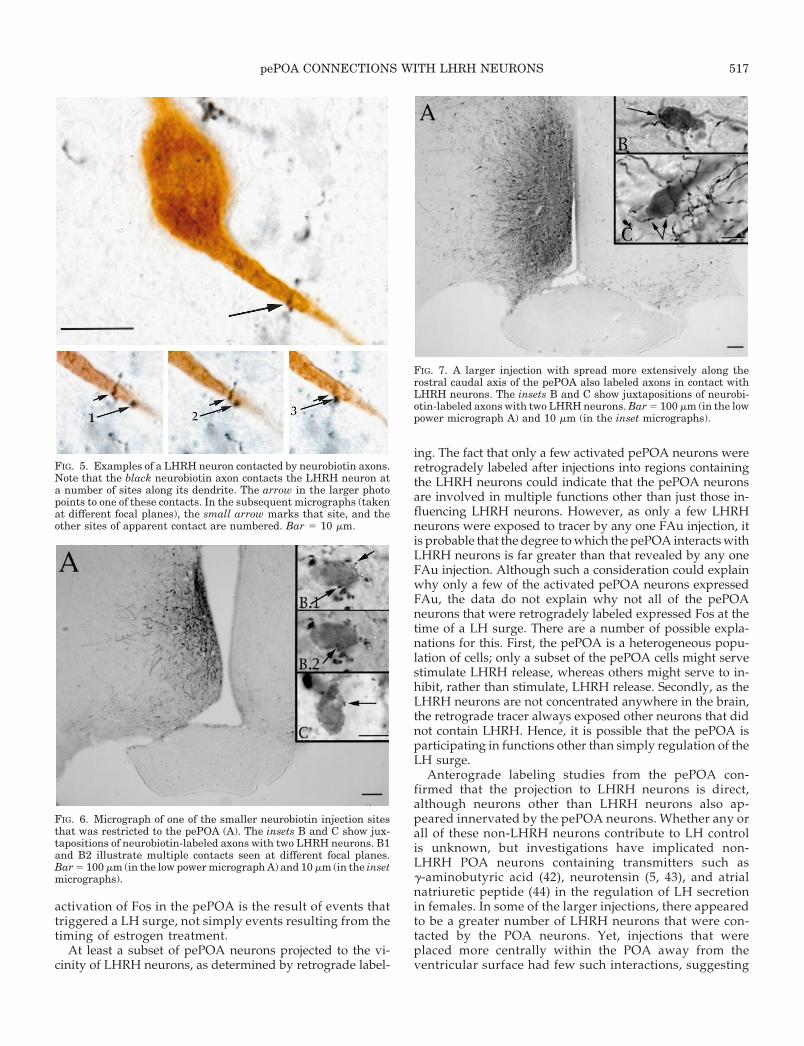
Experiments in which neurobiotin was injected into thepePOA determined that pePOA neurons extended axons tothe LHRH neurons. Of the 17 animals injected with neuro-biotin, 7 had injections that encompassed the pePOA withlittle spread into the adjacent POA. Tissue from these animalsshowed neurobiotin-positive axons arising from the pePOAin close contact with LHRH neurons (Figs. 5-7 and coverphoto). These encompassed LHRH neurons around theOVLT and more caudal sites, but generally did not involvethe rostrally located LHRH neurons. A single axon mademultiple contacts on the same LHRH neuron (Fig. 5). Twoinjection sites of neurobiotin into the pePOA are illustratedin Figs. 6 and 7 to show the range of the injection sizes weobtained. For each of these, examples of biotin-labeled fibersin contact with LHRH neurons are included (insets). There
FIG. 2. Rostral to caudal brain plots (A)at the level of the OVLT where LHRHneurons are most numerous and 300(B), 450 (C) and 600 (D) mm caudal tothe OVLT depict LHRH neurons andFos-positive non-LHRH neurons froman animal killed at the time of the LHsurge. Fos-only neurons (small dots) areplotted for a distance of approximately250 mm from the midline to include theentire pePOA, but not the lateral POA.Open squares indicate LHRH neuronsdevoid of Fos; filled squares show thoseLHRH neurons that expressed Fos.Note that the pePOA Fos activation ismost marked at levels 300–450 mmfrom the OVLT.
514 pePOA CONNECTIONS WITH LHRH NEURONS Endo • 1999Vol 140 • No 1
did not appear to be a preferred dorso-ventral positionwithin the pePOA that affected the number of interactionsnoted, but injections that were centered away from the ven-tricular surface with only minimal spread to the pePOA hadvery few (if any) axonal interactions with LHRH neurons.
Discussion
The results of our study indicate that there is a highlycoordinated Fos activation of the pePOA and LHRH neuronsat the time of a LH surge. Our anatomical data indicate thatpePOA neurons project to LHRH neurons, and a subpopu-lation of pePOA neurons is stimulated at the time of the LHsurge. Together, these data raise the likelihood that thepePOA is assuming a role in the regulation of LHRH activityat the time of a LH surge. A modulatory role for the pePOAin the regulation of gonadotropic function is consistent withthe results of earlier studies aimed at determining where
control of the LH surge was mediated. For example, elec-trolytic lesions of the rostral preoptic area blocked estrouscyclicity as well as steroid-induced LH surges (10–12). Theselesions either severed any possible connection of pePOAneurons with LHRH neurons or included the pePOA per se(Wiegand, S. J., personal communication). Our study nowexpands on these reports to indicate exactly where within thePOA the focus of control lies.
The data we present also clarify the presence of Fosinduction previously observed by Insel (17). In that study,the administration of estradiol to ovariectomized rats in-duced Fos in pePOA neurons at a very precise time afterestrogen treatment. Insel discussed this phenomenon as aconsequence of the duration of exposure of the neurons toestrogen. Upon close examination, the time after estrogentreatment that evoked the greatest Fos activation was theinterval that probably induced a LH surge. Data we and
FIG. 3. Micrographs show one of the FAu injections into the region containing LHRH neurons. FAu/LHRH is shown in a; Fos/LHRH in b; morecaudally, the pePOA Fos pattern in c; retrograde labeling pattern in d; and double labeling of Fos and fluorogold in e in the same animal. Ina, LHRH is brown, and FAu is blue-black. In b, the Fos (blue-black)/LHRH (brown) obtained from the section adjacent to a verifies that theLHRH neurons were activated. The inset shows the side of the injection with two LHRH cells located at approximately the positions indicatedby the asterisks in a. In c, note that the pePOA shows Fos activation, and the same region (d) obtained on the adjacent section possesses numerousretrogradely labeled neurons. In e, some of the FAu-labeled neurons (brown cytoplasm) are also Fos positive (blue-black). Bars 5 100 mm (a,c, and d) and 10 mm (b and e).
pePOA CONNECTIONS WITH LHRH NEURONS 515

others obtained earlier indicated that the administration ofestrogen with progesterone to immature rats (18) or toovariectomized adult rats (16, 41) evokes a pattern ofpePOA activation when the LH surge was induced that issimilar to that seen at the time of a spontaneous LH surge.
This feature is true whether estrogen is administered in themorning (immature rats) or at later times before the surge(adults), but is absent if estrogen is given with progester-one in a paradigm that blocks, rather than evokes, the LHsurge (18). Taken together, these results indicate that the
FIG. 4. Triple labeling of FAu, LHRH,and Fos. FAu is revealed with biotin-amplified fluorescence (green, Bodipyfluorescence); LHRH is stained with ared fluorophore (CAS red); Fos isstained with NiDAB (black). A, Theboundaries of the FAu injection are in-dicated by white arrowheads in a sec-tion taken at a level of the OVLT, justrostral to the center of the injection site.The arrow in A indicates one of theLHRH neurons within the injectionsite. This cell is magnified in B and C.The LHRH cell viewed under fluores-cence optics (B) shows the strong im-munoreactivity of LHRH. C, Brightfieldimage of same cell shows the presence ofFos activation (black nucleus) in theLHRH neuron. Bars 5 100 mm (A) and10 mm (B). D, Plot showing the injectionsite of the same animal depicted in A–C,including plots of the LHRH neurons(circles) and Fos1 neurons that do notexpress LHRH (small dots). LHRH neu-rons that are Fos1 are filled circles;LHRH neurons not expressing Fos areopen circles. No retrograde labeled cellswere seen in that region. E, Retrogradelabeling of POA neurons after the in-jection of FAu into the LHRH cell-con-taining region is depicted. Squares in-dicate the FAu cells; open squares lackFos staining, and closed squares wereFos1. Fos only in the pePOA is shownwith small dots. F, Injection site andLHRH activation patterns in anotherrat (no. 7423). G, Retrograde labelingpattern in the pePOA from rat 7423.The red box shows the region magnifiedin the inset illustrating one Fos1 retro-gradely labeled neuron (white arrow)and above it one of the retrogradely la-beled neurons that did not express Fos.Red structures are portions of LHRHneurons (dendrites and axons) in thesame region. Bar 5 100 mm (A) and 10mm (B, C, and G).
516 pePOA CONNECTIONS WITH LHRH NEURONS Endo • 1999Vol 140 • No 1
activation of Fos in the pePOA is the result of events thattriggered a LH surge, not simply events resulting from thetiming of estrogen treatment.
At least a subset of pePOA neurons projected to the vi-cinity of LHRH neurons, as determined by retrograde label-
ing. The fact that only a few activated pePOA neurons wereretrogradely labeled after injections into regions containingthe LHRH neurons could indicate that the pePOA neuronsare involved in multiple functions other than just those in-fluencing LHRH neurons. However, as only a few LHRHneurons were exposed to tracer by any one FAu injection, itis probable that the degree to which the pePOA interacts withLHRH neurons is far greater than that revealed by any oneFAu injection. Although such a consideration could explainwhy only a few of the activated pePOA neurons expressedFAu, the data do not explain why not all of the pePOAneurons that were retrogradely labeled expressed Fos at thetime of a LH surge. There are a number of possible expla-nations for this. First, the pePOA is a heterogeneous popu-lation of cells; only a subset of the pePOA cells might servestimulate LHRH release, whereas others might serve to in-hibit, rather than stimulate, LHRH release. Secondly, as theLHRH neurons are not concentrated anywhere in the brain,the retrograde tracer always exposed other neurons that didnot contain LHRH. Hence, it is possible that the pePOA isparticipating in functions other than simply regulation of theLH surge.
Anterograde labeling studies from the pePOA con-firmed that the projection to LHRH neurons is direct,although neurons other than LHRH neurons also ap-peared innervated by the pePOA neurons. Whether any orall of these non-LHRH neurons contribute to LH controlis unknown, but investigations have implicated non-LHRH POA neurons containing transmitters such asg-aminobutyric acid (42), neurotensin (5, 43), and atrialnatriuretic peptide (44) in the regulation of LH secretionin females. In some of the larger injections, there appearedto be a greater number of LHRH neurons that were con-tacted by the POA neurons. Yet, injections that wereplaced more centrally within the POA away from theventricular surface had few such interactions, suggesting
FIG. 5. Examples of a LHRH neuron contacted by neurobiotin axons.Note that the black neurobiotin axon contacts the LHRH neuron ata number of sites along its dendrite. The arrow in the larger photopoints to one of these contacts. In the subsequent micrographs (takenat different focal planes), the small arrow marks that site, and theother sites of apparent contact are numbered. Bar 5 10 mm.
FIG. 6. Micrograph of one of the smaller neurobiotin injection sitesthat was restricted to the pePOA (A). The insets B and C show jux-tapositions of neurobiotin-labeled axons with two LHRH neurons. B1and B2 illustrate multiple contacts seen at different focal planes.Bar 5 100 mm (in the low power micrograph A) and 10 mm (in the insetmicrographs).
FIG. 7. A larger injection with spread more extensively along therostral caudal axis of the pePOA also labeled axons in contact withLHRH neurons. The insets B and C show juxtapositions of neurobi-otin-labeled axons with two LHRH neurons. Bar 5 100 mm (in the lowpower micrograph A) and 10 mm (in the inset micrographs).
pePOA CONNECTIONS WITH LHRH NEURONS 517

that indeed the pePOA was providing the significant inputto LHRH neurons. As it was not possible to label the entirepePOA without the injection extending more laterally, weare not certain of the true magnitude of the projection toLHRH neurons, and it is unlikely that tract-tracing studieswill definitively resolve the issue of the magnitude ofcontrol of LHRH neurons by the pePOA. We also acknowl-edge that the use of anterograde tracing does not provethat the same neurons providing the labeled fibers are thepePOA neurons that expressed Fos at the time of the surge,but it raises confidence that these neurons could providesuch input. In a preliminary study we observed that chem-ical lesion of the pePOA neurons prevents Fos activationof LHRH neurons at the time of a LH surge, further sug-gesting that the anatomical link described has functionalsignificance.
Although synchronization of pePOA activation withLHRH activation and anatomical connectivity suggest thatthe pePOA has a role in modulating LHRH activity, whatthat role must still be defined. Studies from our laboratoryindicate that the pePOA neurons activated at the time of a LHsurge contain both progesterone receptors (18) and estrogen(a) receptors (unpublished data). The fact that LH surges canbe induced by local exposure of the POA to these steroidhormones suggests that the pePOA is the transduction sitefor steroidal effects on LHRH activity.
What now remains an important next step is character-ization of the phenotype of the pePOA neurons. Preliminarystudies (41, 45) have suggested that at least some of theactivated pePOA neurons contain dopamine (;10%),whereas others express galanin (; 40%) and both of thesetransmitters are implicated in LHRH control (46, 47). Deter-mining the transmitter of the remaining pePOA neuronsrequires further study.
Acknowledgments
The authors thank Drs. Anne Murphy and Lique Coolen for theircareful reading of the manuscript and their constructive editorialcomments.
References
1. Bleier R, Byne W, Siggelkow I 1982 Cytoarchitectonic sexual dimorphisms ofthe medial preoptic and anterior hypothalamic areas in guinea pig, rat, ham-ster, and mouse. J Comp Neurol 212:118–130
2. Simerly R 1989 Hormonal control of the development and regulation oftyrosine hydroxylase expression within a sexually dimorphic population ofdopaminergic neurons in the hypothalamus. Mol Brain Res 6:297–310
3. Watson RE, Hutchinson RK, Langub MC, Landis JW, Seksaria S, RaineyDM, Keil LC 1994 Colocalization of natriuretic peptide and estrogen receptorimmunoreactivities in preoptic nuclei in the female rat. J Neuroendocrinol6:79–87
4. Fox CA, Adam DE, Watson Jr RE, Hoffman GE, Jacobson CD l990 Immu-nohistochemical localization of cholecystokinin in the medial preoptic area andanterior hypothalamus of the Brazilian gray short-tailed opossum: a sex dif-ference. J Neurobiol 21:705–718
5. Alexander M, Kiraly Z, Leeman S 1991 Sexually dimorphic distribution ofneurotensin/neuromedin N mRNA in the rat preoptic area. J Comp Neurol311:84–96
6. Lisk R 1966 Inhibitory centers in sexual behavior in the male rat. Science152:669–670
7. Powers B, Valenstein E 1972 Sexual receptivity: facilitation by medial preopticlesions in female rats. Science 175:1003–1005
8. Numan M, Callahan EC 1980 The connections of the medial preoptic regionand maternal behavior in the rat. Physiol Behav 25:653–665
9. Numan M 1988 Maternal behavior. In: Knobil E, Neill JD (eds) The Physiologyof Reproduction. Raven Press, New York, pp 1569–1649
10. Wiegand SJ, Terasawa E, Bridson WE, Goy RW 1980 Effects of discrete lesionsof preoptic and suprachiasmatic structures in the female rat. Alterations in thefeedback regulation of gonadotropin secretion. Neuroendocrinology31:147–157
11. Wiegand SJ, Terasawa E 1982 Discrete lesions reveal functional heterogeneityof suprachiasmatic structures in regulation of gonadotropin secretion in thefemale rat. Neuroendocrinology 34:395–404
12. Wiegand SJ, Terasawa E, Bridson WE 1978 Persistent estrus and blockade ofprogesterone-induced LH release follows lesions which do not damage thesuprachiasmatic nucleus. Endocrinology 102:1645–1648
13. Wersinger S, Baum M, Erskine M 1993 Mating induced Fos-like immunore-activity in the rat forebrain: a sex comparison and a dimorphic effect of pelvictransection. J Neuroendocrinol 5:557–568
14. Pfaus J, Kleopoulos S, Mobbs C, Gibbs R, Pfaff D 1993 Sexual stimulationactivates c-fos within estrogen-concentrating regions of the female rat fore-brain. Brain Res 624:253–267
15. Erskine M 1993 Mating-induced increases in Fos protein in preoptic area andmedial amygdala of cycling female rats. Brain Res Rev 32:447–451
16. Coolen L, Peters HPW, Veening JG 1996 Fos immunoreactivity in the rat brainfollowing consumatory elements of sexual behavior: a sex comparison. BrainRes 738:67–82
17. Insel TR 1990 Regional induction of c-Fos-like protein in rat brain followingestradiol administration. Endocrinology 126:1849–1853
18. Le WW, Attardi B, Berghorn KA, Blaustein J, Hoffman GE 1997 Progesteroneblockade of a luteinizing hormone surge blocks luteinizing hormone-releasinghormone Fos activation and activation of its preoptic area afferents. Brain Res778:272–280
19. Horvath TL, Naftolin F, Leranth C 1993 Luteinizing hormone-releasing hor-mone and gamma-aminobutyric acid neurons in the medial preoptic area aresynaptic targets of dopamine axons originating in anterior periventricularareas. J Neuroendocrinol 5:71–79
20. Hoffman GE, Smith MS, Fitzsimmons MD 1992 Detecting steroidal effects onimmediate early gene expression in the hypothalamus. Neuroprotocols1:52–66
21. Watson RE, Wiegand SJ, Clough RW, Hoffman GE 1986 Use of cryoprotectantto maintain longterm peptide immunoreactivity and tissue morphology. Pep-tides 7:155–159
22. Wray S, Hoffman GE 1986 A developmental study of the quantitative dis-tribution of LHRH neurons in postnatal male and female rats. J Comp Neurol252:522–531
23. Barry J, Hoffman G, Wray S 1984 LHRH systems. In: Bkorklund A, HokfeltT (eds) Handbook of Chemical Neuroanatomy. Elsevier/North Holland, Am-sterdam, pp 166–215
24. Paxinos G, Watson C 1986 The Rat Brain in Stereotaxic Coordinates. AcademicPress, Sydney
25. Hoffman GE, Lee WS, Attardi B, Yann V, Fitzsimmons MD 1990 LHRHneurons express c-fos after steroid activation. Endocrinology 126:1736 –1741
26. Hoffman GE, Smith MS, Verbalis JG 1993 c-Fos and related immediate earlygene products as markers for neuronal activity in neuroendocrine systems.Front Neuroendocrinol 14:173–213
27. Lee WS, Smith MS, Hoffman GE 1990 Luteinizing hormone releasing hor-mone (LHRH) neurons express c-Fos during the proestrous LH surge. ProcNatl Acad Sci USA 87:5163–5167
28. Lee WS, Smith MS, Hoffman GE 1990 Progesterone enhances the surge ofluteinizing hormone by increasing the activation of luteinizing hormone-releasing hormone neurons. Endocrinology 127:2604–2606
29. Lee WS, Smith MS, Hoffman GE 1992 cFos activity identifies recruitment ofLHRH neurons during the ascending phase of the proestrous luteinizinghormone surge. J Neuroendocrinol 4:161–166
30. Lee WS, Smith MS, Hoffman GE 1992 LHRH neurons express cJun during theproestrous surge of luteinizing hormone. Endocrinology 130:3101–3103
31. Lee WS, Abbud R, Hoffman GE, Smith MS 1993 Effects of n-methyl-d-aspartate receptor activation on cFos expression in luteinizing hormone-releasing hormone neurons in female rats. Endocrinology 133:2248 –2254
32. Chang HT, Kuo H, Whittaker JA, Cooper NGF 1990 Light and electronmicroscopic analysis of projection neurons retrogradely labeled with fluoro-gold: notes on the application of antibodies to fluorogold. J Neurosci Methods35:31–37
33. Berghorn KA, Bonnet JH, Hoffman GE 1994 cFos immunoreactivity is en-hanced with biotin amplification. J Histochem Cytochem 42:1635–1642
34. Adams JC 1992 Biotin amplification of biotin and horseradish peroxidasesignals in histochemical stains. J Histochem Cytochem 40:1457–1463
35. Rinaman L, Hoffman GE, Dohanics J, Stricker EM, Verbalis JG 1995 Cho-lecystokinin activates catecholaminergic neurons in the caudal medulla thatinnervate the paraventricular nucleus of the hypothalamus in rats. J CompNeurol 360:246–256
36. van der Loos C, Becker A 1994 Double epi-illumination microscopy withseparate visualization of two antigens: a combination of epi-polarization forimmunogold-silver staining and epifluorescence for alkaline phosphatasestaining. J Histochem Cytochem 42:289–296
37. Olson BR, Freilino M, Hoffman GE, Stricker EM, Sved AF, Verbalis JG 1993
518 pePOA CONNECTIONS WITH LHRH NEURONS Endo • 1999Vol 140 • No 1

c-Fos expression in rat brain and brainstem nuclei in response to treatmentsthat alter food intake and gastric motility. Mol Cell Neurosci 4:238–248
38. Fitzsimmons MD, Olschowka JA, Wiegand SJ, Hoffman GE 1992 Interactionof opioid peptide-containing terminals with dopaminergic perikarya in the rathypothalamus. Brain Res 581:10–18
39. Fox SR, Smith MS 1984 The suppression of pulsatile luteinizing hormonesecretion during lactation in the rat. Endocrinology 115:2045–2051
40. Fox SR, Smith MS 1985 Changes in the pulsatile pattern of luteinizinghormone secretion during the rat estrous cycle. Endocrinology 116:1485–1492
41. Le WW, Berghorn KA, Wang HJ, Hoffman GE 1995 Phenotypic character-ization of a population of preoptic area neurons activated during an LH surge.Soc Neurosci Abstr 21:1889 (Abstract 742.5)
42. Wuttke W, Jarry H, Flugge G 1987 GABA is a neurotransmitter of estrogen-receptive neurons in the central nervous system. In: McCann SM, Weiner RI(eds) Integrative Neuroendocrinology: Molecular, Cellular and Clinical As-pects. Karger, Basel, pp 70–79
43. Alexander MJ, Mahoney PD, Ferris CF, Carraway RE, Leeman SE 1989Evidence that neurotensin participates in the central regulation of thepreovulatory surge of luteinizing hormone in the rat. Endocrinology124:783–788
44. Steele M 1990 Additive effects of atrial natriuretic peptide and angiotensin IIon luteinizing hormone and prolactin release in female rats. Neuroendocri-nology 51:345–350
45. Berghorn KA, Smith MS, Watson RE, Langub MC, Hoffman GE, Fos acti-vation of estrogen receptive dopamine neurons in the preoptic area: coordi-nation with LHRH activation. 75th Annual Meeting of The Endocrine Society,Anaheim, CA, 1994 (Abstract 709) p 379
46. Kalra SP 1986 Neural circuitry involved in the control of LHRH secretion: amodel for preovulatory LH release. Front Neuroendocrinol 9:31–75
47. Lopez FJ, Vegro-Vilar A 1990 Galanin stimulates luteinizing hormone-releas-ing hormone secretion from arcuate nucleus-median eminence fragments invitro. Involvement of an a-adrenergic mechanism. Endocrinology 127:2431–2436
pePOA CONNECTIONS WITH LHRH NEURONS 519




















