
Fatty Liver in Hormone Receptor- Positive Breast Cancer and ...
Upload
independentCategory
view
25download
0
Molecular and Cellular Endocrinology 172 (2001) 213–222
Expression and actions of both the follicle stimulating hormonereceptor and the luteinizing hormone receptor in normal ovarian
surface epithelium and ovarian cancer
Jeff A. Parrott 1, Vinayak Doraiswamy, Grace Kim, Rachel Mosher,Michael K. Skinner *
Center for Reproducti6e Biology, School of Molecular Biosciences, Washington State Uni6ersity, Pullman, WA 99163-4231, USA
Received 6 March 2000; accepted 7 August 2000
Abstract
The ability of gonadotropins to act on and regulate normal ovarian surface epithelial (OSE) cells and ovarian cancer cells wasinvestigated. Bovine OSE was used as a model to study normal OSE. Results demonstrate that follicle stimulating hormone (FSH)and the luteinizing hormone (LH) like molecule, human chorionic gonadotropin (hCG), can both stimulate (3H)-thymidineincorporation into DNA in normal OSE cells. Similar results were obtained using either purified hormones or recombinant humanhormones. A human ovarian cancer cell-line OCC1 was also stimulated to grow in response to FSH and hCG, but the growthof a different human ovarian cancer cell-line SKOV3 was not affected. In addition to effects on cell growth, gonadotropins alsostimulated growth factor expression. Both FSH and hCG stimulated steady state levels of keratinocyte growth factor (KGF),hepatocyte growth factor (HGF), and kit ligand (KL) mRNA in OSE cells. Previously, KGF, HGF, and KL have been shownto stimulate OSE growth. Both follicle stimulating hormone receptor (FSHR) and luteinizing hormone receptor (LHR) wereobserved in OSE cells by Northern blot analysis. Reverse transcription polymerase chain reaction (RT-PCR) analysis wasperformed on fresh and cultured OSE cells. Normal OSE was found to express FSHR and LHR both in vivo and in vitro. ThePCR reaction products were sequenced and found to provide a 100% homology with the bovine gonadotropin receptor sequencespreviously reported. FSHR and LHR transcripts were also detected in gonadotropin responsive OCC1 cells, but not in thegonadotropin insensitive SKOV3 cells. Observations support the hypothesis that gonadotropins may influence some ovariancancers. In summary, the current study demonstrates the novel observation that both the FSHR and LHR are expressed bybovine OSE and selected ovarian cancers. Interestingly, the actions of FSH and LH to promote OSE growth may in part bemediated indirectly through an elevation in the expression of autocrine growth factors (KGF, HGF, and KL). Ovarian cancer ismore common in conditions with elevated gonadotropins such as post-menopausal women. Therefore, gonadotropin actions onthe OSE are postulated to be a potential factor in the onset and progression of some ovarian cancers. © 2001 Elsevier ScienceIreland Ltd. All rights reserved.
Keywords: Gonadotropins; Follicle stimulating hormone receptor; FSH; Luteinizing hormone receptor; LH; Ovary; Ovarian cancer; Ovariansurface epithelium; OSE
www.elsevier.com/locate/mce
1. Introduction
The role of the ovarian surface epithelium (OSE) inthe disease referred to as ovarian cancer was first
recognized in the late 1800s by Spencer Wells (Hamil-ton, 1992). Greater than 95% of the ovarian cancersoriginate in the epithelial cells on the surface of theovary (Piver et al., 1991; Weiss et al., 1997). The OSEis a modified mesothelium covering the surface of theovary. It is a simple epithelium separated from underly-ing ovarian stromal tissue by a basal lamina (Nicosiaand Nicosia, 1988). One theory suggests that the hor-monal environment f the OSE may play a role in thedevelopment and progression of ovarian cancer by reg-
* Corresponding author. Tel.: +1-509-3351524; fax: +1-509-3352176.
E-mail address: [email protected] (M.K. Skinner).1 Present address: Atairgin Technologies, Inc., 101 Theory, Suite
150, Irvine, CA 92612, USA.
0303-7207/01/$ - see front matter © 2001 Elsevier Science Ireland Ltd. All rights reserved.PII: S 0 3 0 3 -7207 (00 )00340 -3

J.A. Parrott et al. / Molecular and Cellular Endocrinology 172 (2001) 213–222214
ulating the proliferative activity of OSE cells (Cramerand Welch, 1983). The hypothesis tested in the currentstudy is that gonadotropins, follicle stimulating hor-mone (FSH) and luteinizing hormone (LH), are in-volved in OSE proliferation that may lead to ovariancancer. Several previous observations have led to thishypothesis. Ovarian cancer is more common in condi-tions with elevated gonadotroipins such as post-menopausal women or women who have receivedtreatment for induction of ovulation (Rao and Slot-man, 1991; Shoham, 1994; Balen, 1995; Whittemore etal., 1995). Reduced risk for ovarian cancer is associatedwith multiple pregnancies, breast feeding, oral contra-ceptives, and estrogen replacement therapy which areassociated with lower levels and reduced exposure togonadotropins (Stadel, 1975; Rao and Slotman, 1991;Shoham, 1994; Rossing et al., 1994). FSH and LH maypromote the growth of some ovarian tumors by stimu-lating angiogenesis into the tumor mass (Schiffenbaueret al., 1997).
FSH and LH are glycoprotein hormones synthesizedin the anterior pituitary. Both hormones act in anendocrine manner and play pivotal roles in reproduc-tive physiology. The receptors, follicle stimulating hor-mone receptor (FSHR) and luteinizing hormonereceptor (LHR), are G-protein coupled receptors andare expressed in the target cells in the ovary. FSHR isexpressed by granulosa cells in developing ovarian folli-cles. LHR is expressed by theca cells in early developingfollicles and by both theca cells and granulosa cellsduring later stages of follicular development. The ac-tions of FSH and LH on granulosa cells and theca cellshave been well characterized and are essential for follic-ular development (Rao et al., 1978; Richards and Fa-rookhi, 1978).
The potential roles of FSH and LH in developmentand progression of ovarian cancer have been examinedpreviously. Expression of the FSHR has been observedin normal OSE cells in human ovaries (Zheng et al.,1996). Both FSH and LH have been shown to stimulateproliferation of normal OSE cells from rabbit ovaries invitro (Osterholzer et al., 1985). FSH and LH maystimulate proliferation of some human ovarian cancercell lines (Simon et al., 1983; Wimalasena et al., 1992;Feng et al., 1996). Many human ovarian tumors havebeen shown to contain binding sites for gonadotroipinsby biochemical studies (Wardlaw et al., 1975; Kammer-man et al., 1981; Rajaniemi et al., 1981; Nakano et al.,1989). Other studies have suggested that human ovariancarcinomas express the LHR mRNA (Mandai et al.,1997) and protein (Han, 1992; Kobayashi et al., 1996;Cui and Guo, 1997; Cui et al., 1997). Some humanovarian carcinomas proliferate in response to LH andothers do not respond to LH (Kurbacher et al., 1995).Despite these observations, the role that gonadotropinsmay play in normal OSE biology and ovarian tumori-genesis remain to be fully elucidated.
Studies involving OSE have been limited by the lackof a suitable experimental model. Normal OSE of therat, rabbit, and human have been isolated and cultured(Adams and Auersperg, 1981; Seimens and Auersperg,1988; Piquette and Timms, 1990), but the size andavailability of tissue often limit the use of these models.Bovine ovaries present a useful model for OSE andovarian stromal interactions. The bovine ovary hasessentially the same physiology and size as the humanovary. The cow is a mono-ovulator and has an ovariancycle similar to that of the human. Bovine ovariancancer has been reported (Marchant, 1980), but thefrequency remains to be assessed. Observations suggestthat bovine OSE has similar tumorigenic potential ashuman OSE (Marchant, 1980). Large amounts of freshbovine ovaries can be obtained and large numbers offresh OSE can be isolated from them (Vigne et al.,1994). Therefore, the bovine ovary provides a usefulmodel for examining the actions of gonadotropins onnormal OSE cells.
The current study was designed to examine the ac-tions of gonadotropins on ovarian surface epitheliumand selected ovarian cancer cells. The hypothesis istested that FSH and LH may directly stimulate normalOSE cells and ovarian cancer cells (i.e. OCC1 andSKOV3). Information regarding the hormonal regula-tion of normal OSE provides insight into the mecha-nisms that influence the onset and progression ofovarian cancer.
2. Materials and methods
2.1. Tissue Isolation and cells culture
Bovine ovaries were obtained from young non-preg-nant cycling heifers less than 10 min after slaughter anddelivered on ice. Generally, 50 ovaries were used for anindividual cell preparation for a set of experiments.Bovine OSE cells were isolated fresh or cultured invitro. OSE were scraped from the surface of the ovarywith a rubber policeman as previously described (Kruket al., 1990). Sheets of epithelial cells were suspended inHanks’ buffered salt solution and then pelleted andwashed prior to suspension for plating. Cells wereplated with an initial density of approximately 106 cellsper 2 cm2, and were maintained at 37°C in 5% CO2
atmosphere in Ham’s F-12 (Gibco, Grand Island, NY),supplemented with 10% calf serum. Medium waschanged every 48–72 h. Once the cells had grown toconfluence, the cells were tryupsinized and split intoappropriate plates. For isolation of RNA from culturedcells, OSE cells were plated in 100 mm large cultureplates (Nunc) and maintained in Ham’s F-12 supple-mented with 10% calf serum. For growth assays, OSEcells were plated in 24-well plates in DMEM (Gibco)

J.A. Parrott et al. / Molecular and Cellular Endocrinology 172 (2001) 213–222 215
supplemented with 10% calf serum. When cells achieved50–70% confluency, cells were washed in DMEM con-taining 0.1% calf serum for growth assays. For analysisof gonadotropin treatment on keratinocyte growth fac-tor (KGF), hepatocyte growth factor (HGF), and kitligand (KL) expression OSE cells were plated in 6-wellplates containing Ham’s F-12 supplemented with 10%calf serum. When cells achieved confluency, cells werewashed with serum-free Ham’s F-12. OSE cells weretreated with gonadotropins or growth factors for 48 hand harvested for total RNA. The purity of OSEisolated by this procedure is greater than 98% bykeratin staining with no detectable stromal contamina-tion or follicle cells (e.g. granulosa) (Vigne et al., 1994).The human ovarian cancer cell line SKOV3 was ob-tained from American Type Culture Collection (ATTC,Rockville, MD) and OCC1 cells (Wong et al., 1990)were generously provided by Dr Gordon Mills (MDAnderson Cancer Center, Houston, TX) and culturedunder recommended conditions.
2.2. Growth assays
Cell growth was analyzed by quantifying (3H)-thymidine incorporation into newly synthesized DNA.Normal bovine OSE, SKOV3, and OCC1 cells wereplated (approximately 1 million cells per cm2 providing50% confluency) in 0.5 ml DMEM medium containing0.1% calf serum to synchronize the cell population.After 48 h, cells were treated with no factor (control),100 ng/ml follicle stimulating hormone (ovine FSHS-20, 100 ng/ml; National Pituitary Agency, Baltimore,MD), or 100 ng/ml human chorionic gonadotropin(hCG, 4010 IU/mg, Calbiochem). Optimal dose wasdetermined with dose response curves which demon-strated similar actions for gonadotropins at 50 ng/ml.Cells were plated for 48 h in 0.1% calf serum with notreatment and then treated with gonadotropins orgrowth factors for 20 h. After treatment, 0.5 mlDMEM containing 2.5 mCi (3H)-thymidine was addedto each well, and the cells were incubated for 4 h at37°C and then sonicated. The quantity of (3H)-thymidine incorporated into DNA was determined, aspreviously described (Vigne et al., 1994). Data werenormalized to total DNA per well using an ethidiumbromide procedure previously described. Subsequentstudies confirmed results on OSE cells using recombi-nant human FSH (rhFSH) and hCG (rhCG) obtainedfrom NIH which lacks contaminants such as thyroidstimulating hormone.
2.3. DNA assays
DNA was measured fluorometrically with ethidiumbromide as previously described (Vigne et al., 1994). Analiquot of the sonicated cell suspension was added to an
equal volume of ethidium bromide solution (0.25 mMethidium bromide, 100 U/ml heparin in ethidium bro-mide buffer (EBB, 20 mM sodium chloride, 5 mMethylene diamine tetracetate, 10 mM Tris, pH 7.8);Sigma), was diluted 1:2 with EBB, and was allowed toincubate at room temperature for 30 min. Fluorescentemission at 585 with 350 nm excitation was then moni-tored. A standard curve with calf thymus DNA wasused to quantify DNA levels in the culture wells. Thisassay has a sensitivity of approximately 0.1 mg DNA.
2.4. RNA preparation
Total RNA was prepared from freshly isolated orcultured OSE cells using Trizol reagent (Gibco). Trizolwas added directly to freshly isolated cells or to theculture plate to prevent RNA degradation. Total RNAwas utilized for quantitative reverse transcription-poly-merase chain reaction (RT-PCR) analysis or used topurify mRNA using the FastTrack 2.0 mRNA Isola-tion Kit (Invitrogen, Carlsbad, CA). RNA was storedat −70°C until use.
2.5. Quantitati6e RT-PCR assays
Steady state levels of KGF, HGF, KL, and cy-clophilin (i.e. IB15) mRNA’s were analyzed using aspecific quantitative RT-PCR assay for each gene.These quantitative RT-PCR assays have previouslybeen described in detail (Parrott and Skinner, 1997,1998a,b). The primers used were for conserved regionsbetween several species including bovine. The PCRproducts generated were confirmed with sequencing.The primers used in this quantitative analysis of KGF,HGF, KL, and 1B15 were, KGF, 5%-ATA CTG ACATGG ATC CTG CCA AGT TTG CTC TAC AGATCA TGC TC-3% (5% primer, 45-mer) and 5%-TCC AACTGC CAC GGT CCT GAT-3% (3% primer, 21-mer)which generated a specific 306-bp KGF PCR product;HGF, 5%-ACA GCT TTT TGC CTT CGA GCT ATCGGG GTA AAG ACC TAC AGG-3% (5% primer, 42-mer) and 5%-CAT CAA AGC CCT TGT CGG CATA-3% (3% primer, 22-mer) which generated a specific292-bp HGF PCR product; KL, 5%-GGA CAA GTTTTC GAA TAT TTC TGA AGG CTT GAG TAATTA TTG-3% (5% primer, 30-mer) which generated aspecific 452-bp KL PCR product; and 1B15, 5%-ACACGC CAT AAT GGC ACT GGT GGC AAG TCCATC-3% (5%-primer, 33-mer) and 5%-ATT TGC CATGGA CAA GAT GCC AGG ACC TGT ATG-3% (3%primer, 33-mer) which generated a specific 105-bpproduct from all cell types. The cycleophilin (1B15) wasused to normalize the data and correct for the integrityand amount of the RNA in the samples, as previouslydescribed (Parrott and Skinner, 1997, 1998a,b). BeforeRT, tubes containing total RNA and specific 3%-primers

J.A. Parrott et al. / Molecular and Cellular Endocrinology 172 (2001) 213–222216
were heated to 65°C for 10 min to facilitate denatur-ing and cooled to room temperature to facilitate an-nealing. Total RNA (1 ug) was RT for 1 h at 37°Cusing the following conditions, 1 ug total RNA, 1 uMspecific 3%-primers of interest (up to four differentprimers including 1B15), 0.1 mM dNTP’s, 10 mMDTT, 40 U RNAse inhibitor (Promega, Madison,WI), and 200 U M-MLV RT (Gibco BRL, Gaithers-burg, MD) in 40 ul RT buffer (50 mM Tris–HCl pH8.3, 75 mM KCl,3 mM MgCl2). After 1 h, sampleswere heated to 95°C for 5 min to inactivate the RTenzyme. Samples were immediately diluted 2.5 foldand carrier DNA (Bluescript plasmid, Stratagene) wasadded to a final concentration of 10 ng/ul. This con-centration of Bluescript carrier DNA (10 ng/ul) wasincluded in all subsequent dilutions of samples andstandards. Immediately before amplification, each un-known sample was further diluted 1:10 in order toimprove the fidelity of the PCR reaction. PlasmidDNA’s (e.g. Bluescript) containing bovine KGF,HGF, KL, or 1B15 subclones were used to generatestandard curves from 1 ag/ul (10−15 g/ul) to 10 pg/ul(101×0−9 g/ul) each containing 10 ng/ul Bluescriptcarrier DNA. Identical 10 ul aliquots of each sampleand standard were pipetted in duplicate into a 96 wellreaction plate (Marsh Biomedical Products,Rochester, NY) and sealed with adhesive film (MarshBiomedical Products) for PCR amplification. By thisdesign it was possible to simultaneously assay fiveknown standard concentrations and 40 unknown sam-ples for each gene. Amplification was performed in aPerkin Elmer 9600 equipped with a heated lid usingthe following conditions, 0.4 uM each primer; 16 uMdNTP’s; and 1.25 U AmpliTaq polymerase in 50 ulGeneAmp PCR buffer (containing 1.5 mM MgCl2,Perkin Elmer). Each PCR amplification consisted ofan initial denaturing reaction (5 min, 95°C); 25–31cycles of denaturing (30 s, 95°C), annealing (1 min,60°C), and elongation (2 min, 72°C) reactions; and afinal elongation reaction (10 min, 72°C). At least 0.25uCi of 32P-labeled DCTP (Redivue, Amersham LifeSciences, Arlington Heights, IL) was included in eachsample during amplification for detection purposes.Specific PCR products were quantitated by elec-trophoresing all samples on 4–5% polyacrylamidegels, simultaneously exposing the gels to a phosphorscreen for 8–24 h, followed by quantitating the spe-cific bands on a Storm PhorphorImager (MolecularDynamics, Sunnyvale, CA). Each gene was assayed inseparate PCR reactions from the same RT samples.Equivalent steady state mRNA levels for each genewere determined by comparing each sample to theappropriate standard curve. All KGF, HGF, and KLdata were normalized for 1B15, as previously de-scribed (Parrott and Skinner, 1997, 1998a,b, 2000a).
Optimal cycle number for amplification was deter-mined for each assay in order to achieve maximumsensitivity while maintaining linearity (i.e. logarithmicphase of PCR reactions). KGF quantitative PCRproducts were amplified for 28 cycles, HGF and KLPCR products were amplified for 31 cycles, and 1B15PCR products were amplified for 25 cycles. The sensi-tivity of each quantitative PCR assay was below 1 fgwhich corresponds to less than 125 fg target mRNAper ug total RNA. For each assay, all samples weresimultaneously measured in duplicate resulting in in-tra-assay variabilities of 11.3% (KGF), 13.6% (HGF),8.9% (KL), and 6.5% (1B15).
The RT-PCR for the FSH receptor and LH recep-tor involved similar procedures as listed above withboth 3% primer and oligo-dT RT with 40 cycles ofPCR. The primers for the reactions were FSHR, 5%AACCTTCCCAACCTCCGATATCT-3% (5% PRIMER,23 mer) and 5% ACTGGGAAGCTTTTT-TAAGCGGT-3% (3% primer, 23 mer) which generateda 480 bp product and the LHR, 5% TGTTCTC-CTGACCAGTCGTTACAA-3% (5% primer, 24 mer)and 5% AATGCCTTTGTGAAAATTGCGTAC-3% (3%primer, 24 mer) which generated a 840 bp product.The PCR products were sequenced and found to behomologous to bovine FSHR and LHR by the Centerfor Reproductive Biology, Molecular Biology CoreLaboratory.
2.6. Northern blot analysis
Total RNA and mRNA from OSE cells andovarian stromal cells were isolated as described above.Approximately 6 ug of total RNA and 10 ug mRNAwere fractionated on a 1% formaldehyde agarose gel.Following fractionation, the RNA in the gel wastransferred into Nylon membrane (Hybond N+ ,Amersham) in 1× MOPS buffer and UV crosslinked. The membranes were then prehybridized (500mM phosphate buffer pH 7.2, 1 mM EDTA, 1%BSA, 7% SDS) for 2 h at 60°C. The hybridizationwas carried out at 60°C over night with 32P-labeledFSHR and LHR probes obtained by random primerextension (Prime-It II, Stratagene) of rat cDNA’s. Themembrane was washed in wash buffer (0.2× SSC,0.1% SDS) at room temperature at 60°C for 20 min.Membranes were exposed to X-ray film (X-OMAT,Kodak) overnight at −70°C using an intensifyingscreen. The membrane was subsequently stripped andrehybridized with bovine cyclophilin using a similarprocedure. Rat cDNA’s for FSHR and LHR wereused and found to be cross reactive with the cow andgenerously contributed by Dr Walter Tribley and DrMichael Griswold in the Department of Biochemistryand Biophysics at Washington State University.

J.A. Parrott et al. / Molecular and Cellular Endocrinology 172 (2001) 213–222 217
2.7. Statistical analysis
All data were analyzed by a JMP 3.1 statisticalanalysis program (SAS Institute Inc., Cary, NC). Ef-fects on (3H)-thymidine incorporation into DNA andgrowth factor expression were analyzed by a one-wayanalysis of variance (ANOVA). Observed significanceprobabilities of 0.05 (P\F) or less were consideredevidence that an ANOVA model fits the data. Signifi-cant differences between treated cells and control (un-treated) cells were determined using the Dunnett’s testwhich guards against the high a-size (Type I) error rateacross the hypothesis (Dunnett, 1955). This multiplecomparison test is recommended for multiple compari-sons with control (Hsu, 1996).
3. Results
The ability of gonadotropins to influence the growthof bovine OSE cells was investigated. OSE were platedat 50% confluence and maintained in 0.1% calf serumfor 48 h to synchronize the cell population. Cells werethen treated with 100 ng/ml follicle stimulating (FSH)or 100 ng/ml human chorionic gonadotropin (hCG, anLH agonist) for 20 h followed by a 4 h incubation in(3H)-thymidine. Optimum dose was determined with adose response curve (data not shown). Cells were alsotreated with 10−7 M estradiol (E-7) and 40 ng/mlepidermal growth factor (EGF) or 10% bovine serum(Fig. 1A). Estradiol had no effect on cell growth whileEGF and serum both stimulated OSE cell growth.Purified FSH and hCG were found to stimulate DNAsynthesis in bovine OSE (Fig. 1A). Recombinant hu-man FSH and hCG were also tested to rule out poten-tial effect of minor contaminants in the FSH and hCGpreparations. Both recombinant hormones had similareffects on the growth of bovine OSE cells (Fig. 1B).Analysis of cell number after a 72 h treatment confi-rmed proliferation of the cells (data not shown). There-fore, FSH and hCG (i.e. LH) can act as growth factorsfor normal OSE cells. A combined treatment with FSHand hCG gave the same results of either alone and wasnot additive (data not shown). The ability of go-nadotropins to promote the growth of OSE cells sug-gests that gonadotropins may be involved in the normalgrowth function of the OSE.
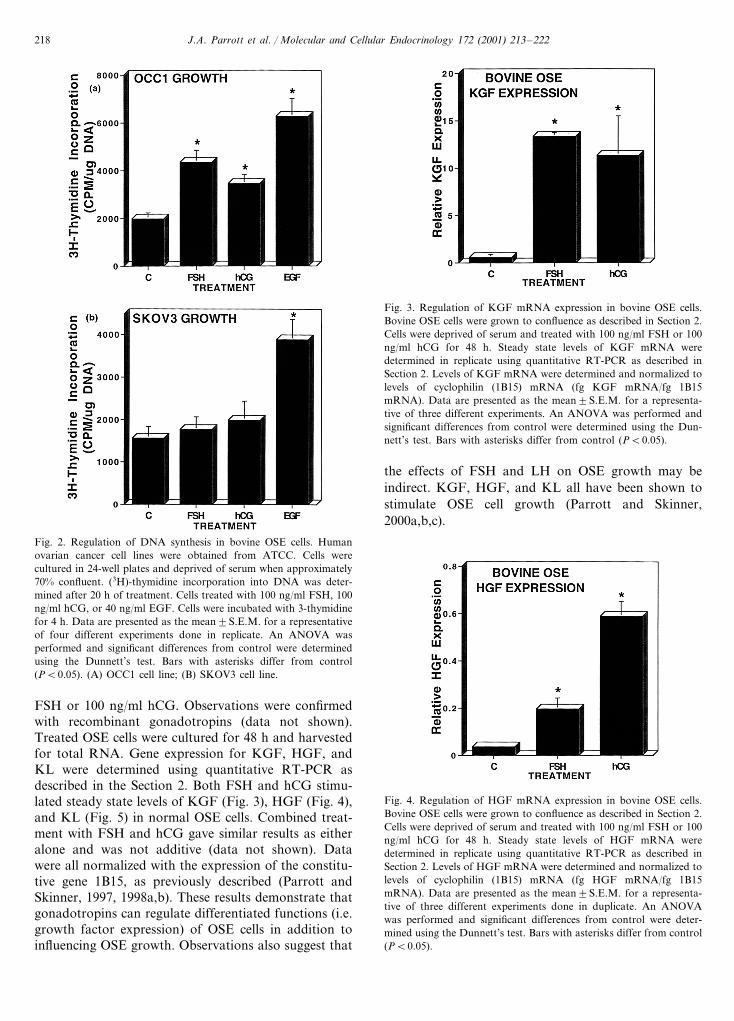
Similar growth experiments were performed usingtwo human ovarian cancer cell lines. SKOV3 andOCC1 (Wong et al., 1990). SKOV3 and OCC1 cells areestablished cell lines that were derived from humanovarian tumors. Both FSH and hCG were found tostimulate DNA synthesis in OCC1 cells (Fig. 2A), butneither hormone affected DNA synthesis in SKOV3cells (Fig. 3B). An increased dose of gonadotropin (i.e.200 ng/ml) also had no effect on SKOV3 growth (data
not shown). EGF was used as a positive control. Obser-vations were confirmed with recombinant go-nadotropins (data not shown). These resultsdemonstrate that gonadotropins can regulate thegrowth of some human ovarian cancer cell-lines.
Potential effects of gonadotropins on growth factorexpression in normal OSE were examined. Normal OSEcells express a number of growth factors includingKGF, HGF, and KL (Parrott and Skinner, 2000a,b,c).Expression of KGF, HGF, and KL were examinedusing sensitive quantitative RT-PCR assays (Parrottand Skinner, 1997, 1998a,b, 2000a,b). OSE cells weregrown to confluency in 6-well plates and maintained inHam’s F-12 in the absence of calf serum. OSE cellswere treated with no growth factor (control), 100 ng/ml
Fig. 1. Regulation of DNA synthesis in bovine OSE cells. BovineOSE cells were isolated as described in Section 2. Cells were deprivedof serum when approximately 70% confluent. (3H)-thymidine incor-poration into DNA was determined after 20 h of treatment. (A) OSEcells treated with 100 ng/ml FSH, 100 ng/ml hCG, 10–7 M estradiol(E-7), 40 ng/ml EGF or 10% bovine calf serum. (B) OSE cells treatedwith 100 ng/ml recombinant hFSH, 100 ng/ml recombinant hCG, or40 ng/ml EGF. Cells were incubated with 3H-thymidine for 4 h. Dataare presented as the mean9S.E.M. for a representative of fourdifferent experiments done in replicate. An ANOVA was performedand significant differences from control were determined using theDunnett’s test. Bars with asterisks differ from control (PB0.05).
J.A. Parrott et al. / Molecular and Cellular Endocrinology 172 (2001) 213–222218
Fig. 2. Regulation of DNA synthesis in bovine OSE cells. Humanovarian cancer cell lines were obtained from ATCC. Cells werecultured in 24-well plates and deprived of serum when approximately70% confluent. (3H)-thymidine incorporation into DNA was deter-mined after 20 h of treatment. Cells treated with 100 ng/ml FSH, 100ng/ml hCG, or 40 ng/ml EGF. Cells were incubated with 3-thymidinefor 4 h. Data are presented as the mean9S.E.M. for a representativeof four different experiments done in replicate. An ANOVA wasperformed and significant differences from control were determinedusing the Dunnett’s test. Bars with asterisks differ from control(PB0.05). (A) OCC1 cell line; (B) SKOV3 cell line.
Fig. 3. Regulation of KGF mRNA expression in bovine OSE cells.Bovine OSE cells were grown to confluence as described in Section 2.Cells were deprived of serum and treated with 100 ng/ml FSH or 100ng/ml hCG for 48 h. Steady state levels of KGF mRNA weredetermined in replicate using quantitative RT-PCR as described inSection 2. Levels of KGF mRNA were determined and normalized tolevels of cyclophilin (1B15) mRNA (fg KGF mRNA/fg 1B15mRNA). Data are presented as the mean9S.E.M. for a representa-tive of three different experiments. An ANOVA was performed andsignificant differences from control were determined using the Dun-nett’s test. Bars with asterisks differ from control (PB0.05).
the effects of FSH and LH on OSE growth may beindirect. KGF, HGF, and KL all have been shown tostimulate OSE cell growth (Parrott and Skinner,2000a,b,c).
Fig. 4. Regulation of HGF mRNA expression in bovine OSE cells.Bovine OSE cells were grown to confluence as described in Section 2.Cells were deprived of serum and treated with 100 ng/ml FSH or 100ng/ml hCG for 48 h. Steady state levels of HGF mRNA weredetermined in replicate using quantitative RT-PCR as described inSection 2. Levels of HGF mRNA were determined and normalized tolevels of cyclophilin (1B15) mRNA (fg HGF mRNA/fg 1B15mRNA). Data are presented as the mean9S.E.M. for a representa-tive of three different experiments done in duplicate. An ANOVAwas performed and significant differences from control were deter-mined using the Dunnett’s test. Bars with asterisks differ from control(PB0.05).
FSH or 100 ng/ml hCG. Observations were confirmedwith recombinant gonadotropins (data not shown).Treated OSE cells were cultured for 48 h and harvestedfor total RNA. Gene expression for KGF, HGF, andKL were determined using quantitative RT-PCR asdescribed in the Section 2. Both FSH and hCG stimu-lated steady state levels of KGF (Fig. 3), HGF (Fig. 4),and KL (Fig. 5) in normal OSE cells. Combined treat-ment with FSH and hCG gave similar results as eitheralone and was not additive (data not shown). Datawere all normalized with the expression of the constitu-tive gene 1B15, as previously described (Parrott andSkinner, 1997, 1998a,b). These results demonstrate thatgonadotropins can regulate differentiated functions (i.e.growth factor expression) of OSE cells in addition toinfluencing OSE growth. Observations also suggest that
J.A. Parrott et al. / Molecular and Cellular Endocrinology 172 (2001) 213–222 219
Fig. 5. Regulation of KL mRNA expression in bovine OSE cells.Bovine OSE cells were grown to confluence as described in Section 2.Cells were deprived of serum and treated with 100 ng/ml FSH or 100ng/ml hCG for 48 h. Steady state levels of KL mRNA were deter-mined in replicate using quantitative RT-PCR as described in Section2. Levels of KL mRNA were determined and normalized to levels ofcyclophilin (1B15) mRNA (fg KL mRNA/fg 1B15 mRNA). Date arepresented as the mean9S.E.M. for a representative of three differentexperiments done in replicate. An ANOVA was performed andsignificant differences from control were determined using the Dun-nett’s test. Bars with asterisks differ from control (PB0.05).
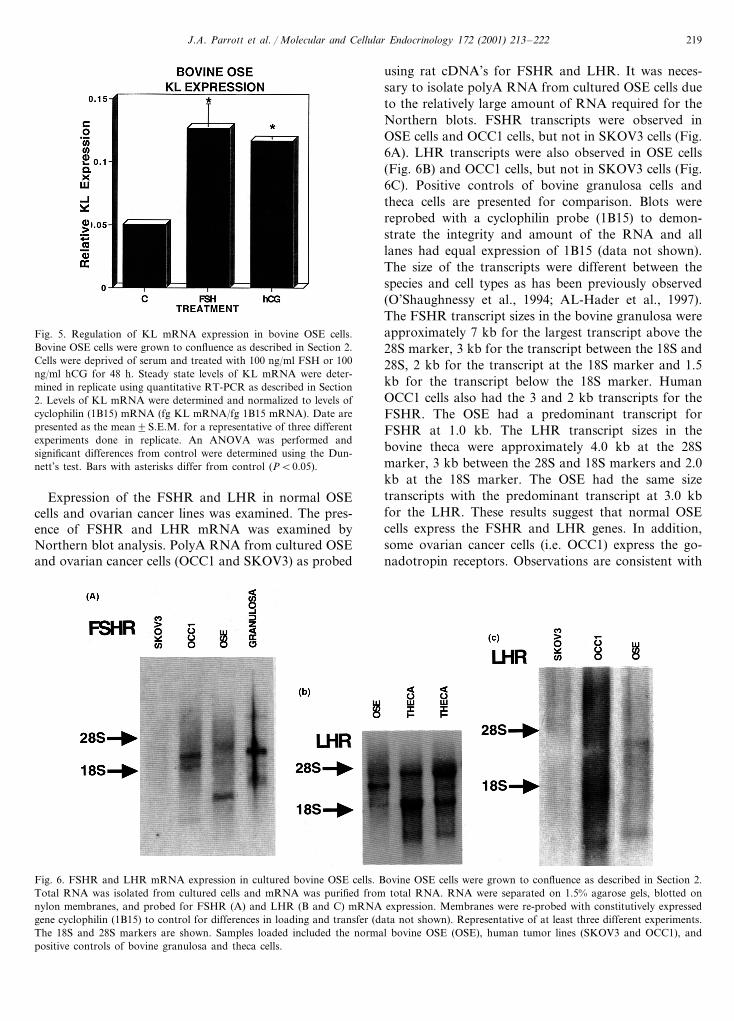
using rat cDNA’s for FSHR and LHR. It was neces-sary to isolate polyA RNA from cultured OSE cells dueto the relatively large amount of RNA required for theNorthern blots. FSHR transcripts were observed inOSE cells and OCC1 cells, but not in SKOV3 cells (Fig.6A). LHR transcripts were also observed in OSE cells(Fig. 6B) and OCC1 cells, but not in SKOV3 cells (Fig.6C). Positive controls of bovine granulosa cells andtheca cells are presented for comparison. Blots werereprobed with a cyclophilin probe (1B15) to demon-strate the integrity and amount of the RNA and alllanes had equal expression of 1B15 (data not shown).The size of the transcripts were different between thespecies and cell types as has been previously observed(O’Shaughnessy et al., 1994; AL-Hader et al., 1997).The FSHR transcript sizes in the bovine granulosa wereapproximately 7 kb for the largest transcript above the28S marker, 3 kb for the transcript between the 18S and28S, 2 kb for the transcript at the 18S marker and 1.5kb for the transcript below the 18S marker. HumanOCC1 cells also had the 3 and 2 kb transcripts for theFSHR. The OSE had a predominant transcript forFSHR at 1.0 kb. The LHR transcript sizes in thebovine theca were approximately 4.0 kb at the 28Smarker, 3 kb between the 28S and 18S markers and 2.0kb at the 18S marker. The OSE had the same sizetranscripts with the predominant transcript at 3.0 kbfor the LHR. These results suggest that normal OSEcells express the FSHR and LHR genes. In addition,some ovarian cancer cells (i.e. OCC1) express the go-nadotropin receptors. Observations are consistent with
Expression of the FSHR and LHR in normal OSEcells and ovarian cancer lines was examined. The pres-ence of FSHR and LHR mRNA was examined byNorthern blot analysis. PolyA RNA from cultured OSEand ovarian cancer cells (OCC1 and SKOV3) as probed
Fig. 6. FSHR and LHR mRNA expression in cultured bovine OSE cells. Bovine OSE cells were grown to confluence as described in Section 2.Total RNA was isolated from cultured cells and mRNA was purified from total RNA. RNA were separated on 1.5% agarose gels, blotted onnylon membranes, and probed for FSHR (A) and LHR (B and C) mRNA expression. Membranes were re-probed with constitutively expressedgene cyclophilin (1B15) to control for differences in loading and transfer (data not shown). Representative of at least three different experiments.The 18S and 28S markers are shown. Samples loaded included the normal bovine OSE (OSE), human tumor lines (SKOV3 and OCC1), andpositive controls of bovine granulosa and theca cells.

J.A. Parrott et al. / Molecular and Cellular Endocrinology 172 (2001) 213–222220
Fig. 7. RT-PCR for FSHR and LHR. RT-PCR for the FSHR andLHR were performed on total RNA from freshly isolated bovineOSE (lane 2 and 5) and positive controls of bovine granulosa cells(lane 3) and theca cells (lane 6) and compared with blank lanes withno RT (lanes 1 and 4). The size of the PCR product is listed and thedata is representative of three different experiments.
following mechanism for the role of gonadotropins inthe progression of ovarian tumors. When the levels ofgonadotropins are low, tumors will progress slowly orwill remain dormant as small avascular microtumors.When the levels of gonadotropins rise, as occurs duringmenopause or after ovariectomy, vascularization inthese small microtumors may accelerate. The early de-velopment of ovarian tumors may not only be pro-moted by gonadotropins, but may also be dependent onthem. It was previously shown that development ofhuman ovarian tumors in mice could be inhibited bytreatment with gonadotropin-releasing hormone antag-onists (Yano et al., 1994). Current results extend thistheory through the novel observations that go-nadotropins stimulate growth and growth factor ex-pression of the early ovarian tumor cells. In addition,both gonadotropins FSH and LH appear to be in-volved. Conditions with elevated gonadotropins mayhave profound effects on OSE cells and early ovariancancer microtumors by regulating cellular proliferation,growth factor expression, and angiogenesis.
Several observations support the gonadotropin the-ory of ovarian carcinogenesis. Previous studies haveshown that some human ovarian tumors express thegonadotropin receptors (Zheng et al., 2000). The novelobservation that the normal bovine OSE express boththe FSHR and LHR is presented in the current study.The analysis of the LHR and FSHR by Northernanalysis revealed different sized transcripts in thebovine OSE and OCC1 cells as compared with thepositive controls of gonadal cells. Previously, alterna-tive splicing and transcript truncation has been shownto generate different size transcripts between cell typesand between species (O’Shaughnessy et al., 1994; AL-Hader et al., 1997). The growth of normal bovine OSEcells and a selected cell line (OCC1) derived from ahuman ovarian tumor were shown for the first time tobe stimulated by gonadotropins in the current study.This supports previous studies on other species withFSH or LH alone (Simon et al., 1983; Kurbacher et al.,1995; Feng et al., 1996; Wimalasena et al., 1992).Ovarian cancer is more common in conditions withelevated gonadotropins such as post-menopausalwomen or women who have received treatment forinduction of ovulation (Rao and Slotman, 1991;Shoham, 1994; Balen, 1995; Whittemore et al., 1995).Reduced risk for ovarian cancer is associated withmultiple pregnancies, breast feeding, oral contracep-tives, and estrogen replacement therapy which is associ-ated with lower levels and reduced exposure togonadotropins (Stadel, 1975; Rao and Slotman, 1991;Shoham, 1994; Rossing et al., 1994). In addition,ovarian tumors develop in some animal models afterprolonged treatment with exogenous gonadotropins orelevated levels of endogenous gonadotropins (Biskindet al., 1953; Kammerman et al., 1977; Marchant, 1980).
the abilities of FSH and hCG to stimulate growth andgrowth factor expression in these cells.
The above examination of FSHR and LHR expres-sion in normal OSE cells was performed on relativelylarge amounts of cultured OSE cells. In order to exam-ine FSHR and LHR expression in freshly isolated OSEcells, RT-PCR analysis was performed. Total RNA wasisolated from different preparations of fresh and cul-tured OSE cells and examined for FSHR and LHRexpression using non-quantitative RT-PCR analysis.FSHR and LHR expression was observed in bothfreshly isolated and cultured OSE cells (Fig. 7 and datanot shown). The RT-PCR products obtained were sub-cloned and sequenced to confirm the identity of thePCR product as either the FSHR or LHR. The se-quence obtained was an exact match with the bovineFSHR and LHR sequences in Genebank (data notshown). These results further demonstrate that the re-ceptors for FSH and LH are expressed by normal OSEcells in vivo.
4. Discussion
Elevated levels and prolonged exposure to go-nadotropins are common to almost all ovarian cancerpatients. Many women who develop ovarian cancer arepost-menopausal and most are treated by removing theovaries. Therefore, an understanding of the effects ofgonadotropins on OSE cells and ovarian cancer cellsmay be important for understanding the developmentand progression of the disease. The gonadotropin the-ory of ovarian carcinogenesis is potentially critical tocertain cases and may be acting by many differentmechanisms. Gonadotropins may promote progressionof ovarian carcinoma by accelerating angiogenesis inthe tumor (Schiffenbauer et al., 1997). In the currentstudy, observations suggest that gonadotropins maypromote ovarian tumor development and progressionon at least two levels, direct stimulation of cellulargrowth factor expression.
The role that hormones play in ovarian carcinogene-sis may be important stages of tumor development andprogression. Schiffenbauer et al. (1997) proposed the

J.A. Parrott et al. / Molecular and Cellular Endocrinology 172 (2001) 213–222 221
The current study supports the theory that go-nadotropins play an important role in human epithelialovarian cancer. Observations suggest that FSH and LHmay help regulate normal OSE biology and selectedovarian tumors. Significantly, results demonstrated thatgonadotropins may stimulate cellular proliferation andthe expression of locally produced autocrine growthfactors. Previously, the bovine OSE have been shown toexpress KGF, HGF, and KL that can act as autocrinegrowth factors and stimulate OSE (Parrott and Skin-ner, 2000a,b,c). Therefore, the ability of FSH and LHto stimulate KGF, HGF, and KL expression supportthe proposal that the effects of gonadotropins on OSEcell growth may in part be indirectly mediated throughthese growth factors. Further studies are required tofully elucidate these potential indirect growth effects ofthe gonadotropins. The current data on mRNA levelswill also need to be confirmed with the analysis ofprotein levels. The concept that hormones can act astumor promoters by stimulating cell proliferation hasbeen proposed (Henderson et al., 1982; Cramer andWelch, 1983; Hamilton and Davies, 1983). The currentfinding may help explain the epidemiological observa-tion of increased risk of ovarian cancer associated withelevated levels of gonadotropins (Zheng et al., 2000).By gaining a better understanding of the actions ofgonadotropins on OSE cells and ovarian cancer, insightwill be gained into the clinical implications of condi-tions such as menopause, fertility treatments, and estro-gen replacement therapy.
The current study demonstrates a novel site for theexpression of both the FSHR and LHR in the ovary.The ability of the OSE to express and respond to FSHand LH provides insight into OSE biology. As FSHand LH promote the growth of the ovarian follicle, theOSE must also proliferate. In addition, after ovulationthe OSE must proliferate to cover the site of ovulation.The current study suggests this growth of the OSE maybe under the control of gonadotropins (Zheng et al.,2000). Since super-ovulation and excessive folliclegrowth have been shown to promote ovarian tumordevelopment, these actions of gonadotropin on theOSE may be a factor in the early onset of the ovariantumor. The subsequent response of selected ovariantumors to gonadotropins also suggest a role for go-nadotropins in tumor progression. Future studies char-acterizing the gonadotropin receptors in OSE andovarian tumors will require an investigation of bindingcapacity, affinity, and unique splice variants. Alter-ations in the gene amplification or deletion in ovariancancers will also be informative.
Acknowledgements
We thank Gene Herrington and Dr Eric Nilsson for
technical assistance and Susan Cobb for assistance inpreparation of the manuscript. This work was sup-ported by a grant from the National Institutes ofHealth (NIH) to Michael K. Skinner involving an NCIOvarian Cancer Program project.
References
Adams, A.T., Auersperg, N., 1981. Transformation of cultured ratovarian surface epithelial cells by Kirsten murine sarcoma virus.Cancer Res. 41, 2063–2072.
Al-Hader, A.A., Lei, S.M., Rao, C.V., 1997. Neurons from fetal ratbrains contain functional luteinizing hormone/chroionic go-nadotropin receptors. Biol. Reprod. 56, 1071–1076.
Balen, A., 1995. The effects of ovulation induction with go-nadotropins on the ovary and uterus and implications for assistedreproduction. Hum. Reprod. 10, 2233–2237.
Biskind, G.R., Berstein, D.E., Gospe, S.M., 1953. The effect ofexogenous gonadotropins on the development of experimentalovarian tumors in rats. Cancer Res. 13, 216–222.
Cramer, D.W., Welch, W.R., 1983. Determinants of ovarian cancerrisk II. Inferences regarding pathogenesis. J. Natl. Cancer Inst. 71,717–721.
Cui, J., Guo, Y., 1997. Relation between ovarian cancer and go-nadotropin and its receptor. Chung Hua Fu Chan Ko Tsa Chih32, 118–120.
Cui, J., Guo, Y., You, Z., 1997. LH-CG receptor protein expressionin epithelial ovarian cancer. Chung Hua Fu Chan Ko Tsa Chih32, 742–745.
Dunnett, C.W., 1955. A multiple comparison procedure of comparingseveral treatments with control. J. Am. Stat. Assoc. 50, 1096–1121.
Feng, Y., Zhang, X., Ge, B., 1996. Gonadotropins stimulate theproliferation of human epithelial ovarian cancer cell. Chung HuaFu Chan Ko Tsa Chih 31, 166–168.
Hamilton, T.C., 1992. Ovarian cancer, part I: biology. Curr. Probl.Cancer 16, 1–57.
Hamilton, T.C., Davies, P., 1983. Hormonal relationships in ovariancancer. Endocr.-Relat. Cancer 14, 19–22.
Han, S., 1992. Characteristics of monoclonal antibodies against cho-rionic gonadotropin receptor. Chung Hau Fu Chan Ko Tsa Chih(Taipei) 72, 74–76.
Henderson, B.E., Ross, R.K., Pike, M.C., Casagrande, J.T., 1982.Endogeous hormones as a major factor in human cancer. CancerRes. 42, 3232–3239.
Hsu, J.C., 1996. Multiple Comparisons: Theory and Methods. Chap-man & Hall, New York, pp. 1–277.
Kammerman, S., Demopoulos, R.I., Ross, J., 1977. Gonadotropinreceptors in experimentally induced ovarian tumors in mice. Can-cer Res. 37, 2578–2582.
Kammerman, S., Demopoulos, R.I., Raphael, C., Ross, J., 1981.Gonadotropic hormone binding to human ovarian tumors. Hum.Pathol. 12, 886–890.
Kobayashi, F., Monma, C., Nanbu, K., Konishi, I., Saawa, N., Mori,T., 1996. Rapid growth of an ovarian clear cell carcinoma ex-pressing LH/hCG receptor arising from endometriosis duringearly pregnancy. Gynecol. Oncol. 62, 309–313.
Kruk, P.A., Maines-Bandiera, S.L., Auersperg, N.A., 1990. A sim-plified method to culture human ovarian surface epithelium. Lab.Invest. 63, 132–136.
Kurbacher, C.M., Jager, W., Kurbacher, J.A., Bittl, A., Wildt, L.,Lang, N., 1995. Influence of human luteinizing hormone on cellgrowth and CA 125 secretion of primary epithelial ovarian car-cinomas in vitro. Tumor Biol. 16, 374–384.

J.A. Parrott et al. / Molecular and Cellular Endocrinology 172 (2001) 213–222222
Mandai, M., Konishi, I., Kuroda, H., Fukumoto, M., Komatsu, T.,Yamamoto, S., Nanbu, K., Rao, C.V., Mori, T., 1997. Messengerribonucleic acid expression of LH/hCG receptor gene in humanovarian carcinomas. Eur. J. Cancer 33, 1501–1507.
Marchant, J., 1980. Animal models for tumors of the ovary. In:Murphy, E.D., Beamer, W.G. (Eds.), Biology of Ovarian Neo-plasia, vol. 50. UICC Tech Rep Ser, pp. 50–65.
Nakano, R., Kitayama, S., Yamoto, M., Shima, K., Ooshima, A.,1989. Localization of gonadotropin binding sites in humanovarian neoplasms. Am. J. Obstet. Gynecol. 161, 905–910.
Nicosia, S.V., Nicosia, R.F., 1988. Pathology of Human Neoplasms.Raven Press, New York, pp. 435–486.
O’Shaughnessy, P.J., Marsh, P., Dudley, K., 1994. Follicle-stimulat-ing hormone receptor mRNA in the mouse ovary during post-na-tal development in the normal mouse and in the adulthypogonadal (hpg) mouse: structure of alternate transcripts. Mol.Cell. Endocrinol. 101, 197–201.
Osterholzer, H.O., Streibel, E.J., Nicosia, S.V., 1985. Growth effectsof protein hormones on cultured rabbit ovarian surface epithelialcells. Biol. Reprod. 33, 247–258.
Parrott, J.A., Skinner, M.K., 1997. Direct actions of kit-ligand ontheca cell growth and differentiation during follicle development.Endocrinology 138, 3819–3827.
Parrott, J.A., Skinner, M.K., 1998a. Developmental and hormonalregulation of hepatocyte growth factor (HGF) expression andaction in the ovarian follicle. Biol. Reprod. 59, 553–560.
Parrott, J.A., Skinner, M.K., 1998b. Developmental and hormonalregulation of keratinocyte growth factor expression and action inthe ovarian follicle. Endocrinology 139, 228–235.
Parrott, J., Skinner M.K., 2000. Expression and action of kerati-nocyte growth factor (KGF) in normal ovarian surface epitheliumand ovarian cancer. Mol. Cell. Endocrinol., 167, 77–87.
Parrott, J., Skinner, M.K., 2000b. Expression and action of hepato-cyte growth factor in normal ovarian surface epithelium andovarian cancer. Biol. Reprod. 62, 491–500.
Parrott, J., Skinner, M.K., 2000c. Expression and action of kitligand/stem cell factor in normal ovarian surface epithelium andovarian cancer. Biol. Reprod. 62, 1600–1609.
Piquette, G.N., Timms, B.G., 1990. Isolation and characterization ofrabbit ovarian surface epithelium, granulosa cells, and peritonealmesothelium in primary culture. In Vitro Cell Dev. Biol. 26,471–481.
Piver, M.S., Baker, T.R., Piedmonte, M., Sandecki, A.M., 1991.Epidemiology and etiology of ovarian cancer. Semin. Oncol. 18,177–185.
Rajaniemi, H., Kauppila, A., Ronnberg, L., Selander, K., Pystynen,P., 1981. LH/hCG receptor in benign and malignant tumors ofhuman ovary. Acta Obstet. Gynecol. Scand. Suppl. 101, 83–86.
Rao, B.R., Slotman, B.J., 1991. Endocrine factors in common epithe-lial ovarian cancer. Endocr. Rev. 12, 14–26.
Rao, M.C., Midgley, A.R., Jr, Richards, J.S., 1978. Hormonal regu-lation of ovarian cellular proliferation. Cell 14, 71–78.
Richards, J.S., Farookhi, R., 1978. Gonadotrophins and ovarian-fol-licular growth. Clin. Obstet. Gynocol. 5, 363–373.
Rossing, M.A., Daling, J.R., Weiss, N.S., Moore, D.E., Self, S.G.,1994. Ovarian tumors in a cohort of infertile women. New Engl.J. Med. 331, 771–771.
Schiffenbauer, Y.S., Abramovitch, R., Meir, G., Nevo, N., Holzinger,M., Itin, A., Keshet, E., Neeman, M., 1997. Loss of ovarianfunction promotes angiogenesis in human ovarian carcinoma.Proc. Natl. Acad. Sci. USA 94, 13203–13208.
Seimens, C.H., Auersperg, N., 1988. Serial propagation of humanovarian surface epithelium in tissue culture. J. Cell Physiol. 134,347–356.
Shoham, Z., 1994. Epidemiology, etiology, and fertility drugs inovarian epithelial carcinoma: where are we today. Fertil. Steril.62, 433–448.
Simon, W.E., Albrecht, M., Hansel, M., Dietel, M., Holzel, F., 1983.Cell lines derived from human ovarian carcinomas: growth stimu-lation by gonadotropic and steroid hormones. J. Natl. CancerInst. 70, 839–845.
Stadel, B.V., 1975. Letter: the etiology and prevention of ovariancancer. Am. J. Obstet. Gynecol. 123, 772–774.
Vigne, J.L., Halburnt, L.L., Skinner, M.K., 1994. Characterization ofbovine ovarian surface epithelium and stromal cells: identificationof secreted proteins. Biol. Reprod. 51, 1213–1221.
Wardlaw, S., Laursen, N.H., Saxena, B.B., 1975. The lh-hcg receptorof human ovary at various stages of the menstral cycle. ActaEndocrinol (Copenhagen) 79, 568–576.
Weiss, N.S., Homochuk, T., Young, J.L., Jr, 1997. Incidence ofhistologic types of ovarian cancer: the US third national cancersurvey, 1969–1971. Gynecol. Oncol. 5, 161–167.
Whittemore, A.S., Harris, R., Itnyre, J., 1995. Characteristics relatingto ovarian cancer risk: collaborative analysis of 12 US case-con-trol studies. II. Invasive epithelial ovarian cancers in whitewomen. Collaborative ovarian cancer group. Am. J. Epidemiol.136, 772–774.
Wimalasena, J., Dostal, R., Meehan, D., 1992. Gonadotropins, estra-diol, and growth factors regulate epithelial ovarian cancer cellgrowth. Gynecol. Oncol. 46, 345–350.
Wong, W.S., Wong, Y.F., Ng, Y.T., Huang, P.D., Chew, E.C., Ho,T.H., Chang, M.Z., 1990. Establishment and characterization of anew human cell line derived from ovarian clear cell carcinoma.Gynecol. Oncol. 38, 37–45.
Yano, T., Pinski, J., Halmos, G., Szepeshazi, K., Groot, K., Schally,A.V., 1994. Inhibition of growth of OV-1063 human-releasinghormone antagonist SB-75. Proc. Natl. Acad. Sci. USA 91,7090–7094.
Zheng, W., Magid, M.S., Kramer, E.E., Chen, Y.T., 1996. Follicle-stimulating hormone receptor is expressed in human ovariansurface epithelium and fallopian tube. Am. J. Pathol. 148, 47–53.
Zheng, W., Lu, J.J., Luo, F., Zheng, Y., Feng, Y.J., Felix, J.C.,Lauchlan, S.C., Pike, M.C., 2000. Ovarian epithelial tumorgrowth promotion by follicle-stimulating hormone and inhibitionof the effect by leutenizing hormone. Gynecol. Oncol. 76, 80–88.
.
Copyright © 2022 FDOKUMEN




















