
Characterization of luteinizing hormone-releasing hormone receptor binding in rat pituitary cell...
12
Moleculur und Cellulur Endocrinology, 44 (1986) 147-l 58 Elsevier Scientific Publishers Ireland, Ltd. 147 MCE 01424 Characterization of luteinizing hormone-releasing hormone receptor binding in rat pituitary cell monolayer cultures; influence of intercellular communication Maria Andries and Carl Denef Laboratory of Cell Phurmacology, University of Leuven, School of Medicine, Campus Gasthuisherg B - 3000 Leuoen (Belgium) (Received 2 August 1985: accepted 28 October 1985) Key words: anterior pituitary: cell culture: luteinizing hormone-releasing hormone: receptors. Summary Receptors for the luteinizing hormone-releasing hormone (LHRH) were characterized in rat pituitary cells cultured for 3 days as monolayers on coverslips using ‘251-[o-Alah-Pro9-LHRH-NEt] as the labeled ligand. The monolayers were left intact during the binding assay. Specific binding displayed the various characteristics of binding to the physiological LHRH receptor. Various kinetic data corresponded to those reported previously. However, in these cultured cells, in which binding was tested in a physiological medium, the dose response of competition for binding by LHRH agonists ranged over a smaller concentration range (< 2 orders of magnitude) than that by LHRH antagonists. In a cation-free buffer competition curves of agonists and antagonists were parallel but the apparent dissociation constant was lower than in the physiological medium. In cultures of pituitary cell populations separated by unit gravity sedimentation, the specific binding increased with the proportional number of gonadotrophs in the various populations. However, when the gonadotroph-richest population ( = 70% gonadotrophs) was cultured after recombination with gonadotroph-poor populations, binding capacity significantly increased. Microscopic examinations suggested that this phenomenon was the consequence of disrupting cellular contacts among gonadotrophs. It is concluded that certain characteristics of LHRH receptors tested on cells in a tissue-like organization and in a physiological environment are different from those reported previously in disrupted cells or monodispersed cell suspensions and that intercellular communication is an important factor controlling LHRH receptors. Introduction Most in vitro studies on the luteinizing hor- mone-releasing hormone (LHRH) receptor in pituitary cells have been confined to membrane preparations or homogenates using non-physio- logical assay media (Spona, 1973a, b; Park et al., 1976; Theoleyre et al., 1976; Pedrosa et al., 1977; Heber and Odell, 1978,1979; Zolman and Valenta, 1978: Baumann and Kuhl, 1979; Clayton et al., 1979a,‘b? 1980; Conne et al., 1979; Clayton and Catt, 1980; Kuhl et al., 1980; Perrin et al., 1980; Chan et al., 1981; Dalkin et al., 1981; Hazum, 1981, 1982, 1983; Savoy-Moore et al., 1981; Marchetti and Labrie, 1982; Barkan et al., 1983). In some studies newly dispersed cells or cultured cells were used (Grant et al., 1973; Naor et al., 1980; Giguere et al., 1981; Meidan and Koch, 0303-7207/86/$03.50 0 1986 El sevier Scientific Publishers Ireland, Ltd.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Characterization of luteinizing hormone-releasing hormone receptor binding in rat pituitary cell...

Moleculur und Cellulur Endocrinology, 44 (1986) 147-l 58
Elsevier Scientific Publishers Ireland, Ltd.
147
MCE 01424
Characterization of luteinizing hormone-releasing hormone receptor binding in rat pituitary cell monolayer cultures; influence of intercellular
communication
Maria Andries and Carl Denef Laboratory of Cell Phurmacology, University of Leuven, School of Medicine, Campus Gasthuisherg B - 3000 Leuoen (Belgium)
(Received 2 August 1985: accepted 28 October 1985)
Key words: anterior pituitary: cell culture: luteinizing hormone-releasing hormone: receptors.
Summary
Receptors for the luteinizing hormone-releasing hormone (LHRH) were characterized in rat pituitary cells cultured for 3 days as monolayers on coverslips using ‘251-[o-Alah-Pro9-LHRH-NEt] as the labeled
ligand. The monolayers were left intact during the binding assay. Specific binding displayed the various characteristics of binding to the physiological LHRH receptor. Various kinetic data corresponded to those
reported previously. However, in these cultured cells, in which binding was tested in a physiological medium, the dose response of competition for binding by LHRH agonists ranged over a smaller concentration range (< 2 orders of magnitude) than that by LHRH antagonists. In a cation-free buffer competition curves of agonists and antagonists were parallel but the apparent dissociation constant was lower than in the physiological medium. In cultures of pituitary cell populations separated by unit gravity
sedimentation, the specific binding increased with the proportional number of gonadotrophs in the various populations. However, when the gonadotroph-richest population ( = 70% gonadotrophs) was cultured after
recombination with gonadotroph-poor populations, binding capacity significantly increased. Microscopic examinations suggested that this phenomenon was the consequence of disrupting cellular contacts among
gonadotrophs. It is concluded that certain characteristics of LHRH receptors tested on cells in a tissue-like organization and in a physiological environment are different from those reported previously in disrupted
cells or monodispersed cell suspensions and that intercellular communication is an important factor controlling LHRH receptors.
Introduction
Most in vitro studies on the luteinizing hor-
mone-releasing hormone (LHRH) receptor in pituitary cells have been confined to membrane preparations or homogenates using non-physio- logical assay media (Spona, 1973a, b; Park et al., 1976; Theoleyre et al., 1976; Pedrosa et al., 1977; Heber and Odell, 1978,1979; Zolman and Valenta,
1978: Baumann and Kuhl, 1979; Clayton et al., 1979a,‘b? 1980; Conne et al., 1979; Clayton and Catt, 1980; Kuhl et al., 1980; Perrin et al., 1980; Chan et al., 1981; Dalkin et al., 1981; Hazum, 1981, 1982, 1983; Savoy-Moore et al., 1981; Marchetti and Labrie, 1982; Barkan et al., 1983). In some studies newly dispersed cells or cultured cells were used (Grant et al., 1973; Naor et al., 1980; Giguere et al., 1981; Meidan and Koch,
0303-7207/86/$03.50 0 1986 El sevier Scientific Publishers Ireland, Ltd.

14X
1981; Loumaye et al., 1982; Meidan et al.. 1982). but in the fatter case ceils were scraped off the culture dishes or fysed by hypotonic medium be- fore binding assays were performed. In such pre- parations LHRH receptors may have suffered from
proteofytic and mechanical damage of the plasma membrane. Moreover, the possible influences of
intercellular communications are efiminated. Inter-
cellular communications in anterior pituitary ap-
pear to be functionally important. Indeed, in the intact pituitary of rats close associations have been
found between goi~adotrophs and lactotrophs but
not between gonadotrophs and other cell types (Nakane, 1970; Horvnth et al.. 1977: Sato, 1980). In previous work we have given evidence for the
existence of a paracrine interaction between
gonadotrophs and lactotrophs in monolayer (De- nef, 1981) and in reaggregate cell culture (Denef and Andries. 1983). Moreover, when a gonado-
troph-rich population separated by unit gravity sedimentation was recombined with a gonado-
troph-poor population, the LH response to LHRH was significantly enhanced (Denef, 1980).
The aim of the present study was (1) to investi-
gate whether the LHRH receptor in pituitary cells could be studied directly on cells in a tissue-like
organization without cells being disturbed, i.e. by
studying binding on intact monolayer cultures,
and (2) to study whether intercellular communica- tions among gonadotrophs or between gonado-
trophs and other cell types would have an in-
fluence on LHRH receptors.
Materials and methods
LHRH was provided through the National Hormone and Pituitary Program, NIADDK (Bal- timore, MD, U.S.A.). [~-Alan’-Prop’-LHRH-NEt]- and [~-Phe~-D-Alarm-LHRH were gifts of Dr. Al- fan Corbin (Wyeth Laboratories, Philadelphia, PA, U.S.A.). [r>-Leu’-Pro’-LHRH-NEt] and thyrot- ropin-releasing hormone (TRH) were from Bachem Fincllemi~als (Marina Del Rey, CA, U.S.A.). Gly- Leu-Arg-Pro-Gly-NH, ([6-IO]-LHRH) and p-en- dorphin were from U.C.B.-Christiaens (Brussels, Belgium). [D-Trp”-Pro’-LHRH-NEt] and [Prd- LHRH-NEtj were from Cambridge Research Bio- chemicals (Cambridgeshire, U.K.). [D-pGl~‘-D-
Phe’-o-Trp”.“]-LHRH was from Peninsula Laboratories (San Carlos, CA, U.S.A.). Vasoactive intestinal peptide (VIP) was from Aalto Biore-
agents (Dublin, Ireland) and somatostatin from Sigma Chemical Company (St. Louis, MO. U.S.A.). [D-Ala”-Pro’-LHRH-NEt] for radioiodination was
dissolved at a concentration of 1 mg/mI in 0.9%
NaCl and stored at -20°C in 10 ~1 fractions. All
other peptides were dissolved at concentrations of
1 or 0.1 mM in 0.9%, NaCl 1% crystalline bovine
serum albumin (BSA) (Miles Laboratories, U.K.) and stored in fractions at -20°C.
Fourteen-day-old female Wistar rats were used
in most experiments because the proportional number of gonadotrophs is higher than in adult
rats (Denef et al., 1982). In some experiments 14-day-old or adult male Wistar rats were used. The animals were housed in an environment of controlled temperature, humidity and day/night cycle. Adult rats had free access to a standard pellet diet and water.
Preparation of monodispersed pituitary cells
and separation of the cells according to size and
density by velocity gradient sedimentation at unit gravity were performed exactly as described previ-
ously (Denef et al., 1978a, b). The 1100 ml gradi-
ent was collected in 100 ml fractions. The cells from each fraction were collected by centrifuga-
tion (100 x g) and the pellet was resuspended in Dulbecco’s modified Eagle’s medium (DMEM) plus 0.3% BSA and counted with a Coulter coun- ter. The first 200 ml contained only cell debris. The following fractions were numbered from 1 to 9 (top to bottom of the gradient). Fraction 1 contained mainly red bfood cells. Usually fractions 6-9 were combined. The efficacy of cell sep- arations was checked for each gradient by histo- chemical staining (Denef et al., 1978a, b) of the different cell types and occasionally by immuno- staining (Denef et al., 1978a, b: Swennen et al.. 1985) or determination of cell size distribution by a Coulter automatic size distribution analyzer (Coulter Electronics. Hialeak, FL, U.S.A.).
To study LHRH binding sites, 2 X 10’ cells

149
(unless otherwise stated) in 50 ~1 DMEM plus
0.3% BSA were plated on poly-t_-lysine-treated coverslips (7.5 x 15 mm). Poly-L-lysine (type I-B; MW > 70000, Sigma Chemical Co., St. Louis,
MO, U.S.A.) was applied to acid-washed cover- slips at 0.1 mg/ml in distilled water for 30-60 min, after which the solution was removed to
dryness by suction. As described previously (De- nef et al., 1978a, b), this treatment allowed rapid
and firm attachment of virtually all the cells to the
coverslips. Cells were allowed to attach for about 20 min before culture medium was added. As
determined by microscopic examination the loss of
cells during culture was negligible. To determine the biological potency and intrin-
sic activity of LHRH and analogs, cells were cul-
tured in clustered plastic Petri dishes (Nunc, Roskilde, Denmark) of 15 mm diameter. Cells,
, suspended in 755100 ~1 DMEM plus 0.3% BSA at
a density of 2 X 10h cells/ml, were allowed to attach for about 30-40 min before additional cul- ture medium was added.
Culture medium consisted of DMEM supple- mented with 3.7 g/l NaHCO,, 10% horse serum or 10% newborn calf serum (Gibco, Grand Island,
NY, U.S.A.), penicillin (35 mg/l) and streptomy-
cin (50 mg/l). Cultures were maintained at 37°C in a water-saturated incubator in an atmosphere of
6.5% CO,/93.5% air. In several study sections serum-free defined medium was used. This culture
medium was composed of DMEM and Ham’s F12
medium (Gibco) supplemented with 1 g/l
NaHCO,, Hepes buffer, Tes buffer (Calbiochem- Behring, La Jolla, CA, U.S.A.), 0.5% BSA, trans-
ferrin, insulin, ethanolamine, sodium selenite, su- peroxide anion scavengers and antibiotics as de- scribed previously (Denef et al., 1984). These cul- tures were maintained in a water-saturated atmo- sphere of 1.2% C02/98.8% air.
(4) Determination of the biological potency and in- trinsic activity of LHRH and analogs
After 3 days the culture medium was removed and the dishes were rinced twice with DMEM, pH 7.5. Incubation with LHRH or analogs was done in 1 ml DMEM containing 1 g/l NaHCO,, 0.015 M Hepes and 0.015 M Tes buffer, pH 7.5 for 4 h at 37°C in an atmosphere of 1.2% CO, in air. The media were removed, BSA was added to a final
concentration of O.l%, and the mixture was
centrifuged. LH in the supernatants was estimated
by RIA, using the rat LH RIA kit distributed by the NIADDK National Hormone and Pituitary Program (Baltimore, MD, U.S.A.). The reliability of the assays has been shown previously (Denef et al., 1978a, b). The dose response curves of [D-Alah-
Pro’-LHRH-NEt] with cells cultured in Petri dishes and with cells cultured on coverslips were identi- cal. Thus, binding data in coverslips cultures could
be reliably compared with biological responses in Petri dish cultures. Bioactivity of the radio-
iodinated LHRH analog was tested similarly. Im-
munoprecipitated counts in the LH RIA were
corrected for non-specific counts from the labeled
LHRH analog. These corrections were accurate as
LH RIA gave the same results when LH secreted
in the bioassay was first separated from the labeled LHRH analog on Sephadex G-50.
(5) Radioiodination of [D-Al&-Pro’-LHRH-NEt]
“51-[D-Ala6-Proy-LHRH-NEt] was used in routine binding assays because, unlike “‘I-LHRH, it only labels high-affinity saturable sites (Clayton et al., 1979b). The analog was labeled with carrier-free “‘1 by the lactoperoxidase method (Marshall and Odell, 1975). The reaction mixture
was composed of 2 mCi (20 ~1) Na”“1 (specific
activity: 15517 mCi/pg iodide), 500 ng (10 ~1) lactoperoxidase, 2 pg (10 ~1) [D-Alah-Pro’-LHRH-
NEt], 80 ng (10 ~1) HzO,, 20 ~1 0.1 M sodium
acetate pH 5.6. After 4 min at room temperature
the reaction was terminated by dilution. The monoiodinated compound was separated from the
diiodinated and unlabeled compounds on a QAE- Sephadex column, eluted with 0.005 M borate buffer pH 9.2 containing 1% BSA. The purity of the monoiodinated peptide was further tested by gel filtration on Sephadex G-25 and by ion ex- change chromatography on QAE-Sephadex at pH 2 and 9.2. All systems gave one peak of radioactiv- ity containing more than 90% of the total radioac- tivity.
The specific activity was calculated by compar- ing the quantities of labeled and unlabeled ligand which cause the same decrease in bound radioac- tivity in competition experiments (Hollemans and Touber, 1974; Hollemans and Bertina, 1975). The specific activity was 78-96% of the theoretical

specific activity of monoiodinated ligand, being
IOOO- 1500 pCi/pg. Binding ahility of the tracer was deternlined as
follows. Coverslips (see Section 6) with 4 X 10’ cells were covered with 5 ~1 of the tracer (1.5 X 10 I” M) and put upside-down in a Petri dish to ensure homogenous exposure of the cells to the assay buffer. For technical reasons (overload) no
more than 4 x 10’ cells/coverslip could be used. Under the latter conditions 24% of the tracer was
specifically bound (for method see Section 6) after ’ 15 min of in~uh~~ti(~n (mean of 4 detern~inations).
It can be calculated from the equilibrium constant
(see further) that under these assay conditions
about 33% of the tracer can he theoretically bound.
Thus, the intrinsic binding ability of the labeled LHRH analog was estimated to be 75% of the
theoretical value. The intrinsic biological activity
(maximal effect) of the tracer was 60% of the unlabeled analog. The biological potency of the labeled ligand. as estimated from the concentra- tion at half-maximal stimulation of LH release from pituitary cell monolayer cultures was about 10 times lower than that of the unlabeled agonist.
Binding was measured on the third day in cul-
ture. ‘The assay buffer (phosphate-buffered saline,
PBS) consisted of 137 mM NaCI. 3 mM KCI, 6
mM NalHPO,. 1.5 mM KH,PO,. 0.5 mM MgC12. 0.5 mM CaCl,. 2% BSA, pH 7.5. The coverslips
were removed from the culture medium. rinsed and incubated at 25°C for various periods of time with different concentrations of “iI-]D-Ala(‘-Pro”-
LHRH-NEt] in 1 ml of the assay buffer. At the end of the incubation the bound radioactivity was separated from the unbound by transferring the
coverslips and washing them for 15 s in large volumes of the assay buffer. They were counted for bound radioactivity in a gamma counter with an efficiency of 70%, for “‘I and preset error at 1%. Non-specific binding was determined by ad- ding a lOO-fold excess of unlabeled peptide to each of the ~~~n~eI7tr~~ti(~I~s of ‘251-laheled ligand used. As far as tested over a concentration range of 5 x 10 I” to 10 h M of the “‘I-labeled ligand this lOO-fold excess was sufficient to saturate the high- affinity binding sites.
(7) Stutistid unu~vsis
Results are expressed as mean + SE (standard error). Single comparisons were tested for signifi-
cance by the paired or unpaired Student t-test. The paired Student t-test was used for results obtained with the same cell culture. Multiple com-
parisons were tested by analysis of variance fol- lowed by the multiple f-test procedure (Dunnet, 1970).
Results
Binding churucteristics of “‘I -(D-A iu”-Pro”-
LHR H-NEt] in pituituty monoluyer cultures
There was no measurable destruction of the
“‘I-labeled ligand during the incubations as in-
cubation of the labeled ligand with cultures for
1-3 h did not change subsequent binding to other cultures. There was a linear correlation between the amount of binding and the number of cells on the coverslips (correlation coefficient = 0.99).
Non-specific binding was IO-20% of total binding, half of which was due to non-specific binding to glass. Coverslips without cells, maintained for 1 day in culture, did not show any specific binding. Scatchard plot analysis of the binding data ob- tained with increasing concentrations of the labeled
ligand gave a straight line, suggesting the presence of a single class of high-affinity binding sites.
Pooled data of 4 separate experiments gave a
Kr, = 2.8 i 0.X x 10 -ii M. Binding capacity was
22.8 i 2.X x 10’ sites/gonadotroph (mean 7fr SE),
assuming that gonadotrophs from 14-day-old
female rats represent about 15% of the dispersed pituitary cell population (Denef et al.. 197Xa) and distribution of receptors is homogenous.
Specific binding was detectable within 30 s and reached equilibrium within 10 min (data not
shown). The association rate constant (k,,) was 6.2 x IO’ M ’ min-‘. The dissociation rate con- stant (k,) was 0.27 min-‘. The dissociation rate was the same after 2 min of incubation. The kinetically derived K,, = k,,/k,, = 4.4 X 10 ‘) M compares favorably with the value 2.8 & 0.8 x IO ’ M obtained from equilibrium analysis.
There was no dramatic effect of the batch of serum added to the culture medium on the number of LHRH binding sites. When horse serum was used, occasionally a higher number of binding
sites/gonadotroph could be found (69 f 11 x 10’
vs. 19+4X lo”, 20+6X10” and 20+3X lo3
with 4 different batches; r-test: P -c 0.005). When the serum was extracted with 0.1% Dextran 70 and 1% charcoal, a procedure commonly used to re-
move endogenous steroid hormones from serum (Drouin et al., 1976) the number of receptors fell to the values commonly found. In serum-free
chemically defined medium the number of LHRH receptors exceeded that found in medium supple- mented with extracted serum (50 i 7 X 10” sites/gonadotroph; n = 5; t-test: P < 0.01).
There was no marked difference in the receptor
affinity among the various cultures tested in these
different culture conditions, K,, ranging around 2.4 x lo-” M with a variation coefficient of 20%.
Competition experiments und correlation of binding
with biological responses
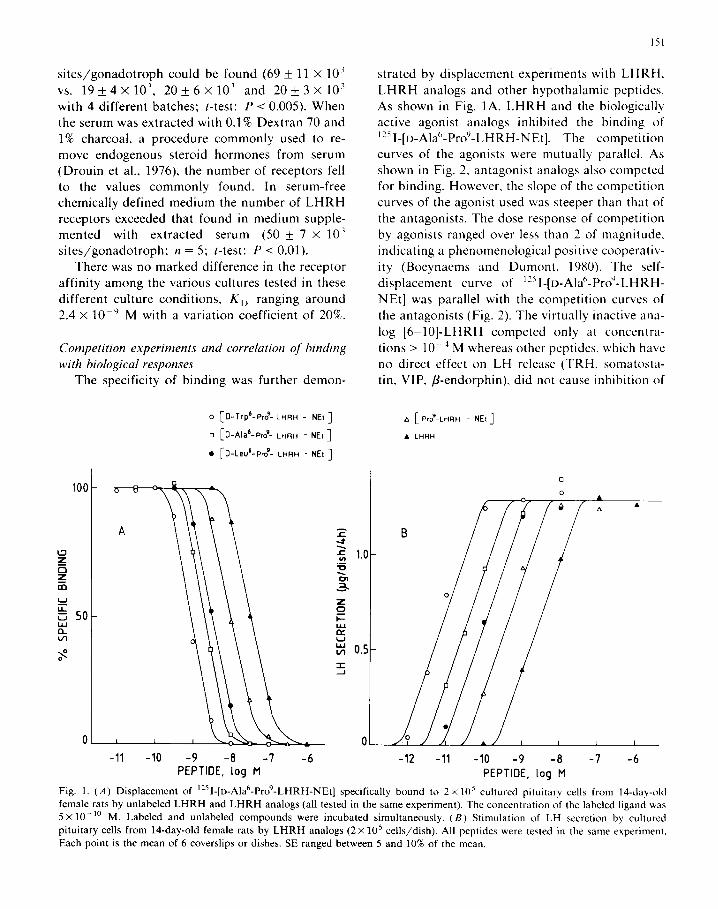
The specificity of binding was further demon-
-11 -10 -9 -0 -1 -6 PEPTIDE, log M
o [D-Trp6-~rr?- LHRH - NE1 ]
q [D-Ala6-~roQ- LHRH NEt ]
. [D-Ld-~rz- LHRH - NEt ]
IS1
strated by displacement experiments with LHRH,
LHRH analogs and other hypothalamic peptides. As shown in Fig. lA, LHRH and the biologically active agonist analogs inhibited the binding of ““I-[D-Ala”-Pro’-LHRH-NEt]. The competition
curves of the agonists were mutually parallel. As shown in Fig. 2, antagonist analogs also competed for binding. However, the slope of the competition
curves of the agonist used was steeper than that of the antagonists. The dose response of competition by agonists ranged over less than 2 of magnitude,
indicating a phenomenological positive cooperativ-
ity (Boeynaems and Dumont, 1980). The self-
displacement curve of “51-[D-AIa’-Pro’1-LHRH-
NEt] was parallel with the competition curves of
the antagonists (Fig. 2). The virtually inactive ana-
log [6-IO]-LHRH competed only at concentra-
tions > 10 ’ M whereas other peptides. which have
no direct effect on LH release (TRH. somatosta- tin, VIP, /3-endorphin). did not cause inhibition of
A [ P~CI-LHRH NEI]
. LHRH
1.0 -
).5-
O_
-12 -11 -10 -9 -0 -1 -6 PEPTIDE, log M
Fig. 1. (A) Displacement of ‘251-[D-Alah-Pro9-LHRH-NEt] specifically bound to 2 x 10’ cultured pituitary cells from 14-day-old female rats by unlabeled LHRH and LHRH analogs (all tested in the same experiment). The concentration of the labeled ligand was
5 x lo-” M. Labeled and unlabeled compounds were incubated simultaneously. (B) Stimulation of LH secretion by cultured
pituitary cells from 14-day-old female rats by LHRH analogs (2 X 10’ cells/dish). All peptides were tested in the same experiment.
Each point is the mean of 6 coverslips or dishes. SE ranged between 5 and 10% of the mean.

152
\
0 1 ’ 4_
-10 -9 -.a -1 -6 -5 -L
PEPTIOE. log M
Fig. 2. Displacement of ‘251-[~-Alah-Proy-LHRH-NEt] specifi-
cally bound to 2 x lo5 cultured pituitary cells from lCday-old
females by an LHRH agonist, [D-Ala’-Pro’-LHRH-NEt], com-
pared with the displacement by LHRH antagonist. [D-Phe’-D- Alah]-LHRH and [D-pGIu’-o-Phe2-D-Trp’,h]-LHRH, inactive
compounds and with the self-displacement of the labeled ligand.
The concentration of the labeled ligand was 5 X IO- ” M. Each
point is the mean of 3-6 coverslips. SE ranged between 5 and
10% of the mean.
binding (Fig. 2). The rank order of potency of agonists in competing for binding corresponded with the rank order of LH-releasing potency of the
respective analogs in monolayer cultures (Fig. 1 B). In order to exclude participation of internaliza-
tion in the apparent binding measured, cells were incubated for 15 min at 25°C with labeled ligand
alone and subsequently a 100-fold excess of un- labeled ligand was added for another 15 min.
Competition of labeled ligand binding by the un-
labeled analog was the same as that measured
after 30 min of incubation of labeled and un- labeled ligand together, indicating that no labeled ligand is internalized to a measurable extent dur-
ing the 15 min of routine incubations at 25°C (data not shown).
Others have shown that cations, such as K’, Na’, Ca” and Mg’+ inhibit binding of LHRH
and analogs (Marian and Conn, 1980; Conn et al., 1981; Hazum, 1981, 1982, 1983). As we measured receptor binding in PBS containing the main phys- iological ions, whereas in many other studies bind- ing assays were performed in cation-free buffers (see Introduction), we compared the binding char- acteristics of “51-[~-Ala“-Proy-LHRH-NEt] in PBS
with those in 0.01 M Tris-HC1/0.25 M sucrose.
The dissociation constants of all LHRH analogs
tested were lower in Tris-HCl/sucrose than in PBS
(Table 1) (Student’s t-test: P -c 0.005). In contrast to those in PBS competition curves
of an agonist and an antagonist LHRH analog were parallel to each other in Tris-HCl/sucrose
and the dose response of competition ranged over 2 orders of magnitude (Fig. 3). The self-displace- ment curve of “51-[~-Alah-Proy-LHRH-NEt]
coincided with the competition curve of unlabeled [D-Ala’-Pro’-LHRH-NEt] (Fig. 3).
TABLE 1
COMPARISON OF APPARENT DISSOCIATION CONSTANTS* OF LHRH AGONISTS AND ANTAGONISTS IN 2
DIFFERENT ASSAY BUFFERS
Values are mean k SE; the number of independent experiments is given in parentheses.
Analog K,, m PBS K,, in Tris-HCl/sucrose
[D-Alah-Pro’-LHRH-NEt] ~’ 1.6kO.l x10_’ M (5) 4.2kO.lxlO~“‘M(3)
‘L51-[~-Alah-Proy-LHRH-NEt] 2.7+0.3x10-’ M(4) 6.0~0.8~10~“’ M(3)
[D-pGIu’-o-PheZ-D-Trp3.h]-LHRH ’ 1.8+0.1xlO~‘M(4) 1.0+0.2xlO~“’ M (2) _
” Agonist.
h Antagonist.
*K,,= dissociation constant: K,, = IC,,,
1 + PI/K,,,, ) where ICs,, is the concentration displacing 50% of specific binding of ‘“‘I-ligand.
K I),Lj the dissociation constant of “sI-ligand, and [L] the concentration of “51-ligand.

153
[D-Ala’-Pro’- LHRH-NEt ]
lb1 [D-Al& Pro’- LHRH - NEt ]
[D-p Glu’-O-Phe’-D-Trp’,‘] H-W
01 I I
-10 -9 -8 -1
PEPTIDE, log M
Fig. 3. Displacement of ‘251-[~-Alah-Pro9-LHRH-NEt] specific
binding by labeled. unlabeled [o-Alah-Pro’-LHRH-NEt] and by
[D-pGlu’-u-Phe2-D-Trp7.h)-LHRH, an LHRH antagonist in
Tris-HCl/sucrose buffer. Two X lo5 cells from 14-day-old
female rat pituitary were incubated with 5 X 1OK”’ M tracer.
Binding to enriched populations of gonudotrophs,
obtained by velocity gradient sedimentation at unit
gravity
As shown in Fig. 4, there was a good correla- tion between the amount of specific binding of ‘251-[D-Ala6-Pro9-LHRH-NEt] to 2 X lo5 cells and the proportional number of gonadotrophs in the
different populations separated by gradient sedi- mentation of pituitary cells derived from 3 animal models (16day-old male and female, and adult
male rats). Due to the limited number of cells in gradient-
separated cell populations, only approximations of affinity constants could be calculated. Using 3 concentrations (4 x lO_” M, lop9 M and 4 x
lop9 M) of the 125 I-ligand, no obvious differences in K, among gradient fractions and animal mod-
els were found (K, = 1.9-4.0 x lop9 M).
Influence of co-culturing gonadotrophs with other cell types
When cells of the gonadotroph-rich gradient fraction 6-9 (60-70s gonadotrophs) from 14-day- old female rats were co-cultured in serum-free
fractions
d adult
Fig. 4. Specific binding of “’ I-[D-Alah-Pro’-LHRH-NEt] (10 ‘)
M) to subpopulations of cultured pituitary cells obtained by
unit gravity gradient sedimentation. (II) Proportional number
of gonadotrophs as determined by immunostaining in the
different gradient fractions. (h) Specific binding of ‘2s1-[~-
Alah-Pro’-LHRH-NEt] (lo-’ M) in the different gradient frac-
tions as expressed per 2 x 10’ cells/coverslip. The same shape
of curve was obtained with 2 other concentrations of the ligand (4XlO_‘O M and 4 x lOmy M). Values are means of 3 (frac-
tions 5. 6-9) or 6 (fractions 2. 3, 4) coverslips. SE was 5-10s
of the mean.
defined medium with the gonadotroph-poor frac- tion 2 (containing mainly lactotrophs and chro- mophobes; see Swennen et al., 1985) in a 1 : 3
ratio, binding was significantly higher than could be expected from the sum of binding values in the separate fractions. This increase in binding was
seen under all culture conditions tested, i.e. in the presence or absence of T3, dexamethasone and estradiol (Fig. 5). When medium supplemented with extracted serum was used, the effect was only seen when estradiol (1 nM) was added to the culture medium (Fig. 5). In contrast, when fraction 4 was co-cultured with fraction 2 no effect on the binding was seen (Fig. 5). An increase in binding was also seen when fraction 6-9 was co-cultured

154
. separate frartlon b-9 0 to-culture fractm b-9 + fractm 2
OL - l. 1 I ,
“One 1 nM E, M”P lnME,
NBCS HS
t 1 I I 1 1 , I I , 1 I
none IlIME, 5nfl T, 80 ntl Oex 5nM 1) 5nM T3
80 nM Dex 80 nM Dex
1nH E,
serum-free defined medium
Fig. 5. Specific binding of “‘1 -( o-Alah-Pro’-LHRH-NEt] to co-cultures of cells from gradient fraction 6-9 or of cells from fraction 4
with cells from gradient fraction 2 obtained by sedimentation at unit gravity of pituitary cells from 14-day-old female rats. Cells were
cultured either in medium supptemented with 10% extracted newborn calf serum (NBCS) or with 10% extracted horse serum (HS) or
in serum-free defined medium with or without T3 (5 nM), dexamethasone (80 nM) (Dex) and estradiol(1 nM) (E,). The concentration
of the labeled tigand was lOmy M. Values are the mean&SE of 3-5 independent experiments. ’ P c 0.05: ++ P (0.025; +'+ P < 0.01; ++++P cc 0.005 (paired f-test)
with a somatotroph-rich fraction obtained from adult male rats or with human fibroblasts (Table
As examined by light-microscopic examination, there was no difference in plating efficiency with cells from fraction 6-9 and cells from fraction 6-9 mixed and co-cultured with the other populations. In our routine experiments plating density is ap- proximately 2 x 10” cells/mm2. When cells from fraction 6-9 were cultured at lower densities (15, 10 or 5 X lo2 celIs/mm2), no effect on the binding per gonadotroph was seen (data not shown). There was perfect linear correlation between binding and number of cells. At densities lower than 5 X 10’ cells/mm2 there was a considerable loss of cells,
as estimated by light-microscopic examination.
To evaluate the mutual arrangement of gona- dotrophs under co-culture conditions, we ex- amined the distribution of acidophils and basophils in 3-day-old cell cultures after histochemical stain- ing. Acidophils represent somatotrophs and lactotrophs whereas basophils include the gona- dotrophs and thyrotrophs. In fraction 6-9 from 14-day-old female rats 85-95s of the basophils are gonadotrophs as determined by immuno- staining (Denef et al., 1978a, b). First we com- pared the gonadotroph-rich fraction (fraction 6-9 from 14-day-old female rats) with a co-culture of this fraction and the somatotroph fraction 7-9 from adult males in a 1 : 3 ratio. Each basophif

155
TABLE 2
EFFECT OF CO-CULTURING FRACTION 6-9 ISOLATED
BY GRADIENT SEDIMENTATION OF 14-DAY-OLD
FEMALE RAT PITUITARY CELLS WITH OTHER CELL
TYPES
Co-culture values are significantly higher than the values of
pure fraction 6-9.
Cell culture o.h cpm bound/lo5 cells
of fraction 6-9 ’
Experiment I
Fraction 6-9 0 1850 f 200
Fraction 6-9 ? + fraction 2 P 3950*400 **
Fraction 6-9 0 + human fibroblasts 4300 + 40 **
Experiment 2
Fraction 6-9 0 3 520 f 250
Fraction 6-9 0 + fraction 2 0 582Ok540 *
Fraction 6-9 ? + fraction 7-9 $ 5869k320 **
E.rperiment 3
Fraction 6-9 0 2600* 20
Fraction 669 0 + fraction 2 0 348Ok160 **
Fraction 6-9 ? + fraction 7-9 6 362Ok280 *
The number of cells of the respective co-culture partner was
3 times that of fraction 6-9.
Culture medium of Experiment 1 was supplemented with
10% extracted horse serum and 1 nM estradiol; culture
medium of Experiments 2 and 3 was supplemented with
10% newborn calf serum and 1 nM estradiol.
Human fibroblasts did not give any specific binding. Frac-
tion 2 from 14-day-old females gave a specific binding of
2-558 of the specific binding of fraction 6-9. whereas
specific binding in fraction 7-9 from adult males was
IO-15% of the specific binding of fraction 6-9. This specific
binding was subtracted from the specific binding of the
co-cultures.
** P < 0.005; * P < 0.01 (Student’s t-test).
was classified as making contact with other basophils, with basophils and acidophils, with only acidophils or with no cells at all. As shown in Fig.
6 most basophils were in contact with other basophils when fraction 6-9 from lCday-old
female rats was cultured alone. When the same fraction was co-cultured with somatotrophs from adult male rats in a 1 : 3 ratio most basophils were in contact with only acidophils.
We also explored the gonadotroph-rich fraction 6-9 at low plating density. We classified each basophil as being in contact with one other basophil, with more basophils or as making no cell contacts at all. The number of basophils in contact
100
80
:
U
% 0
>
60 I
40
20
0 n GON GON + SOM
1 L
++ ++ ;L
I
++ z
+ + +I,
++ r _
ab cdef ab cdef
Fig. 6. Microscopic examination of the gonadotroph-rich frac-
tion 6-9 of lCday-old females (GON) with co-cultures of the
latter fraction with the somatotroph-rich fraction 7-9 of adult
males (SOM). a, basophils; b, acidophils; c, basophils in con-
tact with only basophils; d, basophils in contact with basophils
and acidophils; e, basophils in contact with only acidophils; f.
single basophils. Values are significantly different (Student’s
t-test) from values obtained with pure fraction 6-9. + P <
0.025; ++ P i 0.005.
with more than one other basophil decreased slightly but the number of single cells did not
augment by decreasing the plating density of the cells (data not shown).
It should be mentioned that K, values in the
co-culture conditions could not be measured due to the limited number of cells in fraction 6-9.
Discussion
“51-[D-Alu”-ProY-LHRH-NEt] labels LHRH re-
ceptors on living pituitary cells in a tissue-like
organization
The present investigation demonstrates that monolayer cultures of rat pituitary cells on cover-
slips represent a relatively easy system to study presumptive LHRH receptors not only under bio- chemically reliable conditions but also under cell physiological conditions, since binding assays were performed directly on the monolayers.
Binding of ‘251-[D-Alah-Proy-LHRH-NEt] was highly specific since more than 80% of total bind-

156
ing represented binding to saturable high-affinity sites. Non-specific binding to glass was very low.
Specific binding to glass, as reported for certain
other peptides (Cuatrecasas and Hollenberg, 1975) did not occur. There was no evidence for degrada-
tion of the “51-labeled ligand during the assays.
Internalization of LHRH agonists has been re- ported to occur only after 30 min incubation at 25°C (Naor et al., 1981). Indirect evidence was found that internalization did not occur under the conditions of the present study (15 min incubation at 25°C). Thus, apparent binding reflected high-
affinity sites at the external surface of the plasma membrane. All these characteristics were found
under conditions preserving the morphological and functional integrity of the cells.
There is strong evidence that the specific bind- ing of “SI-[D-Alah-Proy-LHRH-NEt] to cultured
cells represents binding to a biologically relevant LHRH receptor. First, only LHRH, its biologi-
cally active analogs and antagonists effectively competed for binding. Inactive LHRH fragments
or other peptides did not. Second, the rank order of biological potency of the analogs correlated well with the rank order of binding affinity. Third, in experiments with gradient-separated cell popula- tions with low and high proportional numbers of gonadotrophs, a correlation was found between the amount of binding and the proportional num-
ber of gonadotrophs in these populations. The latter data extend similar findings by Naor et al.
(1982) who used only one animal model. Further- more, specific binding was detectable within 30 s
of incubation which agrees with the onset of action
of LHRH (Hopkins, 1977). As cells were cultured for 3 days before the
binding studies were done, it could be expected
that the substances of the culture medium might influence the affinity and quantity of LHRH re- ceptors. Variations in binding characteristics as a function of the batch of serum used were only occasionally found and could be reduced by ex- tracting the serum (steroids) with dextran-coated charcoal. However, when cultured in serum-free chemically defined medium, which is an enriched medium supplemented with transferrin. insulin and other growth-promoting substances, the number of LHRH binding sites, but not the dissociation con- stant, was higher than in the traditional serum-
supplemented medium. The reason for this re- mains to be determined.
Adashi et al. (1981) reported that insulin
(0.1-1000 mg/l) enhances LHRH-stimulated LH
and FSH release by cultured pituitary cells. It is therefore possible that the larger number of LHRH
receptors found in serum-free defined medium is due to the presence of 5 mg/l insulin which is lo- to lOO-fold higher than the value one should ex- pect in medium supplemented with 10% serum (Adashi et al., 1981).
By comparing binding in Tris-HCl/sucrose with that in PBS, a medium containing cations in
physiological concentrations, we found that ap- parent K,, values were decreased by the cations present in PBS. An interesting new finding was
that in PBS but not in cation-free buffer, the slope of the competition curve of an antagonist was not
parallel to that of an agonist. Competition for
binding by agonists ranged over less than 2 orders of magnitude which suggests that agonist binding displays phenomenological positive cooperativity
(Boeynaems and Dumont. 1980). Competition curves, both ranging concentrations approaching 2 orders of magnitude, for agonists and antagonists
were parallel in Tris-HCl/sucrose, suggesting that positive cooperativity does not occur in the latter buffer (Boeynaems and Dumont, 1980). As the above characteristics were not observed by others using membranes or cell suspensions in cation-
containing media, these findings indicate that the LHRH receptor displays a higher level of specific-
ity when tested in reassociated cells in monolayer culture. It should be noted, however, that dif- ferences in agonist and antagonist binding have
also been mentioned by others on the basis of other criteria. Hazum (1981, 1982, 1983; Loumaye et al., 1982) reported that binding of agonists and
antagonists is differently influenced by cations and by petreatment of the membranes with proteo lytic enzymes, sulfhydryl blocking agents, neuro- aminidase and wheat germ agglutinin. Agonist binding was more decreased than antagonist bind- ing. Conn et al. (1982) found evidence that stimu- lation of LH release requires microaggregation of LHRH receptors and that agonists trigger this microaggregation of receptors while antagonists do not. However, the phenomenon of differential binding of agonists and antagonists remains con-

157
troversial, as others did not find an effect of
cations on binding (Perrin et al., 1983). As far as “51-[D-Alah-Pro’-LHRH-NEt] is con-
cerned, saturation experiments gave linear Scat-
chard plots in both assay buffers. The self-dis-
placement curve of this iodinated LHRH analog
was parallel with the displacement curves of the antagonists suggesting this iodinated analog be- haves as an antagonist. However, in release experi- ments ‘251-[D-Alah-Proy-LHRH-NEt] had LH-re- leasing activity. Although it was 10 times less potent than [D-Ala’-Pro’)-LHRH-NEt] the binding affinities of these compounds were almost the
same. The maximal stimulation of LH release by
the labeled analog also did not reach the maximal stimulation by other analogs. All this indicates
that iodination of [D-Ala’-Pro’-LHRH-NEt] con- verts this agonist into a partial agonist. In fact,
this is not surprising because labeling occurs on
the TyrS residue which is situated on the active site of the molecule (Humphries et al,, 1974).
Intercellular communication affects LHRH recep- tors
In the present experiments we found evidence that the LHRH receptor is influenced by mutual contacts between the gonadotrophs. When the gonadotrophs were in contact with each other,
LHRH agonist binding was lower than when they were separated from each other by co-culturing them with other glandular cell types (lactotrophs or somatotrophs) or with fibroblasts. The latter
modulation of binding was highly reproducible as it was seen under several hormonal conditions. Because the effect of increased binding under co- culture conditions was not induced by a specific
cell type as it was seen in co-cultures with
lactotrophs, somatotrophs and even with non- glandular cells (fibroblasts) we propose that the phenomenon is the consequence of a disruption of intercellular communication among the gonado- trophs. This was confirmed by microscopic evalua- tion of cell contacts: the gonadotrophs were un- able to make contacts among each other when an excess of other cells was present. Dilutions of the gonadotrophs to low population density, on the other hand, did not result in an increase of LHRH agonist binding but microscopic examination showed that, although gonadotrophs were in con-
tact with less cells, the number of single gonadotrophs in the culture did not increase.
It is highly unlikely that the increased binding
to LHRH receptors in the co-culture conditions is based on a trivial mechanism such as selective
difference in plating efficiency of the gonado-
trophs. Microscopic examination of the cells dur-
ing the culture period would have revealed such
events. There is also little reason to suspect a multiplication of gonadotrophs as it is well estab- lished that mitosis in pituitary monolayer cultures is very slow (see Denef, 1980).
The physiological significance of the present
findings remains to be studied but could be related to the fact that the topographical distribution of
gonadotrophs (see Denef, 1980) and their affinity for other cell types (Sato, 1980) in the intact
pituitary gland changes during development and the estrous cycle in the rat and that these changes
parallel variations in hormone release. Further-
more, in previous work we have shown that LHRH-stimulated LH release in monolayer cul-
tures of a highly enriched population of gonado-
trophs is considerably lower than in monolayer cultures of the latter population recombined with a lactotroph-enriched population (Denef, 1980). The importance of intercellular communication in pituitary cell physiology (Denef and Andries, 1983) has also become evident since we recently found
that gonadotrophs secrete a substance capable of stimulating the lactotrophs.
Finally, it is likely that the non-parallel compe- tition curves by agonists and antagonists which we
found in our assay system is also caused by an effect of intercellular communication on LHRH receptors. Indeed, others (Meidan and Koch, 1981)
using mono-dispersed cells, between which com- munication is disrupted, did not find this dif-
ference between agonists and antagonists, even using a physiological assay buffer.
References
Adashi, E.Y.. Hsueh, A.J.W. and Yen. S.S.C. (1981) Endo- crinology 108, 144-1449.
Barkan, A.L., Regiani, S.R.. Duncan, J.A. and Marshall. J.C. (1983) Endocrinology 112, 1042-1048.
Baumann. R. and Kuhl, H. (1979) Acta Endocrinol. 92. 228-241.
Boeynaems. J.M. and Dumont. J.E. (Eds.) (1980) In: Outlines

158
of Receptor Theory (Elsevier/North Holland Biomedical
Press, Amsterdam) pp. 33-37.
Chan, V.. Clayton, R.N., Knox, G. and Catt, K.J. (1981)
Endocrinology 108. 208662092.
Clayton, R.N. and Catt, K.J. (1980) Endocrinology 106,
115441159.
Clayton, R.N.. Shakespear, R.A. and Marshal, J.C. (1978) Mol.
Cell. Endocrinol. 11, 63-78.
Clayton, R.N., Shakespear. R.A.. Duncan, J.A. and Marshall.
J.C. (1979a) Endocrinology 104, 148441494.
Clayton, R.N., Shakespear. R.A., Duncan, J.A. and Marshall.
J.C. [with appendix by Munsun, P.J. and Rodbard. D.]
(1979b) Endocrinology 105, 1369-l 381.
Clayton, R.N.. Solano, A.R., Garcia-Vela. A., Dufau, M.L. and
Catt, K.J. (1980) Endocrinology 107, 699-706.
Conn, P.M.. Marian, J., McMillian, M., Stern, J.. Rogers, D.,
Hamby. M.. Penna. A. and Grant, E. (1981) Endocr. Rev. 2,
174-185.
Conn. P.M., Rogers, D.C., Stewart. J.M., Niedel. J. and Schef-
field, T. (1982) Nature (London) 296, 653-655.
Conne, B.S., Aubert, M.L. and Sizonenko. P.C. (1979) Bio-
them. Biophys. Res. Commun. 90. 124991256.
Cuatrecasas, P. and Hollenberg. M.D. (1975) Biochem. Bio-
phys. Res. Commun. 62, 31-41.
Dalkin. A.C.. Bourne, G.A.. Pieper. D.R.. Regiani. S. and
Marshall, J.C. (1981) Endocrinology 108. 16581664.
Denef. C. (1980) In: Synthesis and Release of Adenohypophy-
seal Hormones, Eds.: M. Jutisz and K.W. McKerns (Plenum
Press. New York) pp. 659-676.
Denef. C. (1981) Ann. Endocrinol. 42. 65-66.
Denef, C. and Andries, M. (1983) Endocrinology 112, 813-822.
Denef, C., Hautekeete. E., De Wolf. A. and Vanderschueren. B.
(1978a) Endocrinology 103, 7244735.
Denef, C.. Hautekeete, E. and Dewals. R. (1978b) Endocrinol-
ogy 103. 736-747.
Denef, C.. Swennen. L. and Andries, M. (1982) Int. Rev. Cytol.
76. 225-244.
Denef, C., Baes, M. and Schramme, C. (1984) Endocrinology
114, 1371-1378.
Drouin, J., Lagace. L. and Labrie. F. (1976) Endocrinology 99,
1477-1481.
Dunnet, C.W. (1970) In: Statistics in Endocrinology. Eds.: J.W.
McArthur and T. Colton (MIT Press, Cambridge) p. 79.
Giguere. V., Lefebvre. F.A. and Labrie, F. (1981) Endocrinol-
ogy 108, 350-352.
Grant. G., Vale, W. and Rivier, J. (1973) Biochem. Biophys.
Res. Commun. 50, 771-778.
Hazum. E. (1981) Mol. Cell. Endocrinol. 23. 2755281. Hazum. E. (1982) Mol. Cell. Endocrinol. 26, 217-222.
Hazum. E. (1983) Biochem. Biophys. Res. Commun. 112, 306-312.
Heber, D. and Odell, W.D. (1978) Biochem. Biophys. Res. Commun. 82. 67-73.
Heber, D. and Odell. W.D. (1979) Am. J. Physiol. 237.
E1366E141.
Hollemans. H.J.G. and Touber, J.L. (1974) Clin. Chim. Acta
56, 3055306.
Hollemans, H.J.G. and Bertina, R.M. (1975) Clin. Chem. 21.
1769-1773.
Hopkins, C.R. (1977) J. Cell Biol. 73. 685-695.
Horvath. E.. Kovaca, K. and Ezrin, C. (1977) IRCS Med. Sci. 5.
511.
Humphries, J.. Wan. Y.-P., Fisher, G., Folkers. G. and Bowers.
C.Y. (1974) Biochem. Biophys. Res. Commun. 57,6755682.
Kuhl. H.. Baumann. R., Sandow. J. and Zaubert, H.D. (1980)
Mol. Cell. Endocrinol. 17. 61-70.
Loumaye, E.. Naor, Z. and Catt. K.J. (1982) Endocrinology
111, 730-736.
Marchetti. B. and Labrie. F. (1982) Endocrinology 111.
1209-1216.
Marian, J. and Conn, P.M. (1980) Life Sci. 27, 87-92.
Marian, J. and Conn, P.M. (1983) Endocrinology 112. 1044112.
Marshall, J.C. and Odell, W.D. (1975) Proc. Sot. Exp. Biol.
Med. 149. 351-355.
Meidan, R. and Koch, Y. (1981) Life Sci. 28. 1961-1967.
Meidan. R.. Aroya. N.B. and Koch, Y. (1982) Life Sci. 30.
535-541.
Nakane, P.K. (1970) J. Histochem. Cytochem. 18, 9920.
Naor. Z., Clayton. R.N. and Catt, K.J. (1980) Endocrinology
107. 1144-1152.
Naor, Z.. Atlas. D.. Clayton, R.N., Forman. D.S., Amsterdam.
A. and Catt. K. (1981) .I. Biol. Chem. 256. 3049-3052.
Naor, Z.. Childs. G.V.. Leifer. A.M., Clayton. R.N.. Amater-
dam. A. and Catt. K.J. (1982) Mol. Cell. Endocrinol. 25.
85597.
Park. K.R.. Saxena. B.B. and Gandy. H.M. (1976) Acta Endo-
crinol. 82. 62-70.
Pedrosa, E., Vilchez-Martinez. J.A.. Fishbach, J.. Arimura. A.
and Schally, A.V. (1977) Biochem. Biophys. Res. Commun.
79, 234-238.
Perrin, M.H., Rivier. J.E. and Vale, W.W. (1980) Endocrinol-
ogy 106, 1289-1296.
Perrin. M.H., Haas. Y.. Rivier, J.E. and Vale, W. (1983) Mol.
Pharmacol. 23. 44-51.
Sato. S. (1980) Endocrinol. Jpn. 27, 573-583.
Savoy-Moore, R.T., Schwartz, N.B.. Duncan, J.A. and Marshall,
J.C. (1981) Endocrinology 109. 1360-1364.
Spona, J. (1973a) FEBS Lett. 34. 24-26.
Spona. J. (1973b) FEBS Lett. 35. 59-62.
Swennen, L., Baes, M.. Schramme, C. and Denef. C. (1985)
Neuroendocrinology 40. 78-83. Theoleyre. M.. Berault, A.. Garnier, J. and Jutisz, M. (1976)
Mol. Cell. Endocrinol. 5, 365-377.
Zolman. J.C. and Valenta, J.L. (1978) Acta Endocrinol. 89,
2322239.




















