
Intercellular and systemic spread of RNA and RNAi in plants
15
Advanced Review Intercellular and systemic spread of RNA and RNAi in plants Mohammad Nazim Uddin and Jae-Yean Kim ∗ Plants possess dynamic networks of intercellular communication that are crucial for plant development and physiology. In plants, intercellular communication involves a combination of ligand–receptor-based apoplasmic signaling, and plasmodesmata and phloem-mediated symplasmic signaling. The intercellular trafficking of macromolecules, including RNAs and proteins, has emerged as a novel mechanism of intercellular communication in plants. Various forms of regulatory RNAs move over distinct cellular boundaries through plasmodesmata and phloem. This plant-specific, non-cell-autonomous RNA trafficking network is also involved in development, nutrient homeostasis, gene silencing, pathogen defense, and many other physiological processes. However, the mechanism underlying macromolecular trafficking in plants remains poorly understood. Current progress made in RNA trafficking research and its biological relevance to plant development will be summarized. Diverse plant regulatory mechanisms of cell-to-cell and systemic long-distance transport of RNAs, including mRNAs, viral RNAs, and small RNAs, will also be discussed. © 2013 John Wiley & Sons, Ltd. How to cite this article: WIREs RNA 2013. doi: 10.1002/wrna.1160 INTRODUCTION T he non-cell-autonomous action of proteins and RNAs is a key issue in plant biology. Non- cell-autonomous proteins, such as transcription factors and RNAs that include viral RNAs, endogenous mRNAs, and small RNAs [micro-RNAs (miRNAs) and short interfering RNAs (siRNAs)], are macromolecules that can traffic between plant cells. Many research reports have demonstrated that non-cell-autonomous protein- and RNA-mediated intercellular communication participates in the establishment of developmental patterning and the integration of whole plant physiology. 1–6 In this paper, we provide an overview of cell-to-cell and systemic long-distance RNA trafficking, its regulatory mechanism, and its biological relevance, which is crucial in plant development. ∗ Correspondence to: [email protected] Division of Applied Life Science (BK21-WCU Program), Plant Molecular Biology and Biotechnology Research Center, Gyeongsang National University, Jinju 660-701, South Korea The authors have declared no conflicts of interest for this article. SYMPLASMIC COMMUNICATION PATHWAYS IN PLANTS Plasmodesmata—Symplasmic Nanochannels between Plant Cells To survive in diverse ecosystems, plants have evolved unique nanochannels of communication, which are called plasmodesmata. Each of the plasmodesmata exists as a discrete plasma membrane- lined cytoplasmic cylinder; the central region of this cylinder is covered by an appressed form of the endoplasmic reticulum (ER), which is the desmotubule that establishes endomembranous continuity between the nuclear envelopes of neighboring cells (Figure 1). The intervening space between the membranes forms a cytoplasmic sleeve, which constitutes the key conduit through which cell-to-cell movement of molecules takes place. The plasmodesmata not only allow the movement of small molecules, such as water, metabolites, and hormones, but also regulate the transport of macromolecules, such as transcription factors and various forms of RNA molecules. 7–16 The permeability of plasmodesmata is mod- ulated during development and in response to © 2013 John Wiley & Sons, Ltd.
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Intercellular and systemic spread of RNA and RNAi in plants

Advanced Review
Intercellular and systemic spreadof RNA and RNAi in plantsMohammad Nazim Uddin and Jae-Yean Kim∗
Plants possess dynamic networks of intercellular communication that are crucialfor plant development and physiology. In plants, intercellular communicationinvolves a combination of ligand–receptor-based apoplasmic signaling, andplasmodesmata and phloem-mediated symplasmic signaling. The intercellulartrafficking of macromolecules, including RNAs and proteins, has emergedas a novel mechanism of intercellular communication in plants. Variousforms of regulatory RNAs move over distinct cellular boundaries throughplasmodesmata and phloem. This plant-specific, non-cell-autonomous RNAtrafficking network is also involved in development, nutrient homeostasis,gene silencing, pathogen defense, and many other physiological processes.However, the mechanism underlying macromolecular trafficking in plants remainspoorly understood. Current progress made in RNA trafficking research andits biological relevance to plant development will be summarized. Diverseplant regulatory mechanisms of cell-to-cell and systemic long-distance transportof RNAs, including mRNAs, viral RNAs, and small RNAs, will also bediscussed. © 2013 John Wiley & Sons, Ltd.
How to cite this article:WIREs RNA 2013. doi: 10.1002/wrna.1160
INTRODUCTION
The non-cell-autonomous action of proteins andRNAs is a key issue in plant biology. Non-
cell-autonomous proteins, such as transcriptionfactors and RNAs that include viral RNAs,endogenous mRNAs, and small RNAs [micro-RNAs(miRNAs) and short interfering RNAs (siRNAs)],are macromolecules that can traffic between plantcells. Many research reports have demonstrated thatnon-cell-autonomous protein- and RNA-mediatedintercellular communication participates in theestablishment of developmental patterning and theintegration of whole plant physiology.1–6 In thispaper, we provide an overview of cell-to-cell andsystemic long-distance RNA trafficking, its regulatorymechanism, and its biological relevance, which iscrucial in plant development.
∗Correspondence to: [email protected]
Division of Applied Life Science (BK21-WCU Program), PlantMolecular Biology and Biotechnology Research Center, GyeongsangNational University, Jinju 660-701, South Korea
The authors have declared no conflicts of interest for this article.
SYMPLASMIC COMMUNICATIONPATHWAYS IN PLANTS
Plasmodesmata—Symplasmic Nanochannelsbetween Plant CellsTo survive in diverse ecosystems, plants haveevolved unique nanochannels of communication,which are called plasmodesmata. Each of theplasmodesmata exists as a discrete plasma membrane-lined cytoplasmic cylinder; the central region of thiscylinder is covered by an appressed form of theendoplasmic reticulum (ER), which is the desmotubulethat establishes endomembranous continuity betweenthe nuclear envelopes of neighboring cells (Figure 1).The intervening space between the membranes forms acytoplasmic sleeve, which constitutes the key conduitthrough which cell-to-cell movement of moleculestakes place. The plasmodesmata not only allowthe movement of small molecules, such as water,metabolites, and hormones, but also regulate thetransport of macromolecules, such as transcriptionfactors and various forms of RNA molecules.7–16
The permeability of plasmodesmata is mod-ulated during development and in response to
© 2013 John Wiley & Sons, Ltd.

Advanced Review wires.wiley.com/rna
CWCW
ERER
CWCW
CW
PD nanochannels
PDPPDP Callose
DP
PM
FIGURE 1 | Symplasmic communication channels in plant. A modelfor plasmodesmata is shown in median longitudinal (top) andtransverse (bottom) views. The abbreviations are the following: CW, cellwall; DP, docking protein; ER, endoplasmic reticulum; IPM, inner plasmamembrane leaflet; OPM, outer plasma membrane leaflet; PDP,plasmodesmal proteins; PM, plasma membrane. (Reprinted withpermission from Ref 17. Copyright 2001 Nature publishing group;Reprinted with permission from Ref 18. Copyright 2006 Elsevier Ltd)
environmental stimuli. The apertures of plasmod-esmata change from simple channels to complexbranched structures during the sink-to-source tran-sition in leaf development, accompanying withmodifications of plasmodesmal permeability.19 Alter-natively, the dynamic deposition of β-1,3-glucan (cal-lose) around plasmodesmata regulates plasmodesmalgating, and high callose accumulation at the plas-modesmata can eventually block the movement ofsignaling molecules.20
Phloem—A Unique Superhighway ofInformation in Land PlantsLand plants can transport diverse signaling molecules,such as sugars, hormones, metabolites, proteins, andRNAs, to distant organs or tissues using long-distancevascular pathways in the phloem and/or xylem.21
The phloem acts as the major signaling superhighwayby facilitating symplasmic transport of diverse non-cell-autonomous proteins via sieve tubes, which areenucleated during their maturation. Multidisciplinarystudies have provided evidence that RNA macro-molecules are selectively delivered from companioncells to sieve tubes before traveling through the vascu-lar system and spreading throughout the plant.22 Thelong-distance delivery of RNA is involved in the reg-ulation of physiological and developmental processesand stress responses, and this pathway can also becommandeered by viruses for systemic transport.17
INVASION OF EXOGENOUS RNAsINTO PLANTS
Complex RNA signaling network was observed uponplant infection by various exogenous, infectious RNAmolecules, including viroids and RNA viruses. In thefollowing section, we describe viroid and viral RNAtrafficking networks and their biological significancein plant systems.
ViroidsViroids are single-stranded, covalently closed cir-cular RNAs with genome sizes range from 250to 400 nucleotides. Viroids do not encode pro-teins; thus, to establish a systemic infection, distinctviroid sequences, structural loop/bulges, or functionalmotifs interact with the pre-existing host componentsto perform cellular functions, such as replication,pathogenesis, and trafficking between cells. Thereare approximately 30 known species of viroids, andthey are classified into two families: Avsunviroidae(type member Avocado sunblotch viroid, ASBVd)and Pospiviroidae (type member Potato spindle tuberviroid, PSTVd).
The first evidence of cell-to-cell trafficking bya viroid through plasmodesmata was provided byan experiment using microinjection.23 The authorsinjected fluorescently labeled transcripts of PSTVdinto leaf mesophyll cells (connected by plasmodes-mata) and mature guard cells (lacking connections byplasmodesmata) of tobacco plants. Viroid transcriptthat was injected into a mesophyll cell moved into theneighboring cells but not into the guard cells, whichsuggested that a plasmodesma-mediated traffickingpathway was involved. Transport of a non-mobiletranscript fused to PSTVd through the plasmodes-mata also suggested that specific motif(s) of PSTVdRNA facilitate selective, cell-to-cell trafficking.23 Amotif for the selective trafficking of this RNA fromthe bundle sheath into mesophyll was later reported.24
Viroids can also be transported over long dis-tances through the vascular system from the cell thatwas initially infected. Systemic viroid RNA spreadwas first experimentally observed in a tomato leaf.25
Although PSTVd accumulated significantly in theroots and leaves above the inoculated leaf, it didnot accumulate in the leaves below the inoculatedleaf. This result further supported the hypothesis thatsystemic RNA trafficking occurs through the phloemin plants. Similar findings were also confirmed by insitu hybridization experiments.26 To assess the impactof PSTVd on replication and systemic trafficking, agenome-wide, loss-of-function genetic analysis wasconducted. Through genome wide deletions and/or
© 2013 John Wiley & Sons, Ltd.
WIREs RNA Intercellular RNA movement in plants
base substitutions, the authors identified multipleloops/bulges in the PSTVd secondary structure thatwere essential for systemic trafficking or replicationin single cells.27 The cell-to-cell movement of viroidRNA is also dependent on cell or organ type. In sink(young) leaves, the bundle sheath-mesophyll plasmod-esmata acted as a barrier for RNA movement.24 Ininfected tobacco and tomato plants, PSTVd RNA canmove into sepals but not into other floral organs,26,28
which indicated that viroid RNA trafficking occursvia a selective transport pathway.
RNA VirusesIn contrast to viroids, RNA viruses encode one ormore viral proteins that aid in viral replication andtrafficking in host plants. Numerous studies haverevealed that viral movement proteins bind to viralRNA; this binding increases the channel size of the
plasmodesmata to facilitate the transport of the viralgenome into neighboring cells.29 Different viral move-ment proteins were shown to follow distinct pathwaysbased on the degree of structural changes of theplasmodesmata and the form of the viral ribonu-cleoprotein (vRNP) complex during cell-to-cell viralRNA trafficking.30,31 We discuss two representativeexamples below.
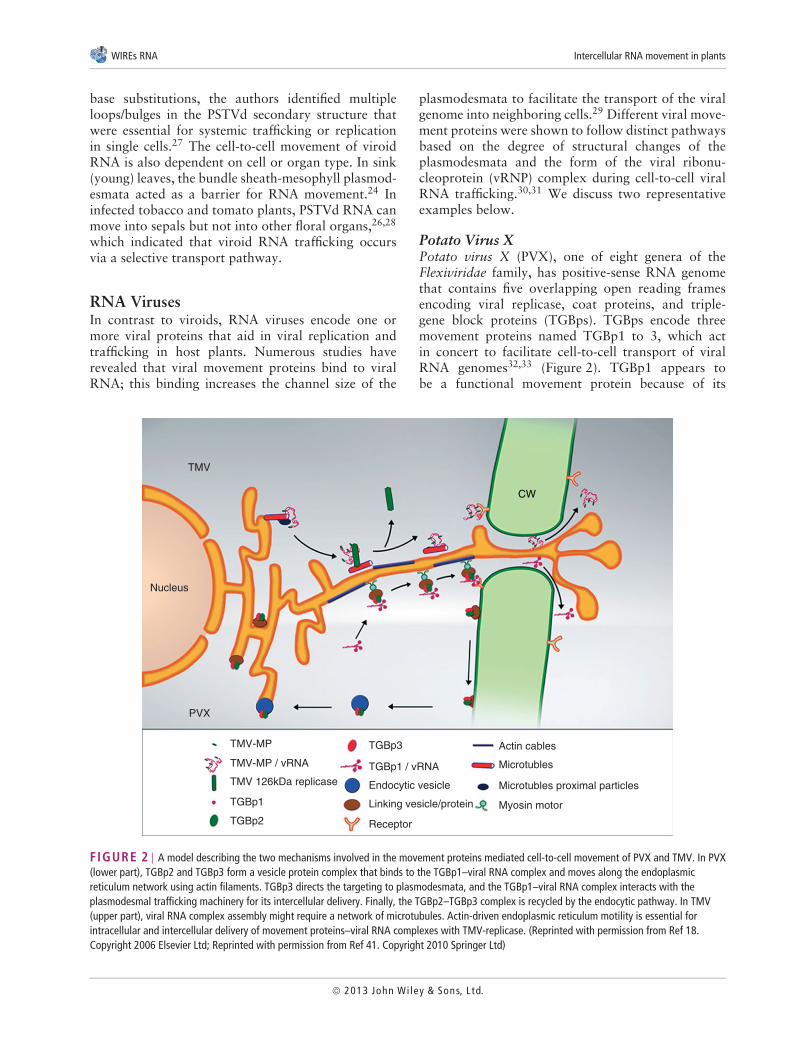
Potato Virus XPotato virus X (PVX), one of eight genera of theFlexiviridae family, has positive-sense RNA genomethat contains five overlapping open reading framesencoding viral replicase, coat proteins, and triple-gene block proteins (TGBps). TGBps encode threemovement proteins named TGBp1 to 3, which actin concert to facilitate cell-to-cell transport of viralRNA genomes32,33 (Figure 2). TGBp1 appears tobe a functional movement protein because of its
TMV
Nucleus
PVX
CW
TMV-MP
TMV-MP / vRNA
TMV 126kDa replicase
TGBp1
TGBp3 Actin cables
Microtubles
Microtubles proximal particles
Myosin motor
TGBp1 / vRNA
Endocytic vesicle
Linking vesicle/protein
ReceptorTGBp2
FIGURE 2 | A model describing the two mechanisms involved in the movement proteins mediated cell-to-cell movement of PVX and TMV. In PVX(lower part), TGBp2 and TGBp3 form a vesicle protein complex that binds to the TGBp1–viral RNA complex and moves along the endoplasmicreticulum network using actin filaments. TGBp3 directs the targeting to plasmodesmata, and the TGBp1–viral RNA complex interacts with theplasmodesmal trafficking machinery for its intercellular delivery. Finally, the TGBp2–TGBp3 complex is recycled by the endocytic pathway. In TMV(upper part), viral RNA complex assembly might require a network of microtubules. Actin-driven endoplasmic reticulum motility is essential forintracellular and intercellular delivery of movement proteins–viral RNA complexes with TMV-replicase. (Reprinted with permission from Ref 18.Copyright 2006 Elsevier Ltd; Reprinted with permission from Ref 41. Copyright 2010 Springer Ltd)
© 2013 John Wiley & Sons, Ltd.

Advanced Review wires.wiley.com/rna
capacity to bind to the 5′ untranslated region of viralRNA, increase the channel size of plasmodesmata,traffic itself via plasmodesmata, and promote viralRNA transport.34 Subcellular localization studies haveindicated that TGBp2 and TGBp3 are positioned in thevicinity of the endoplasmic reticulum.35 Interestingly,in the presence of these proteins, TGBp1 that wasfused to the green fluorescent protein (GFP) wasdirected to cell wall-embedded punctate bodies andmigrated into neighboring cells from the bombardedcell, which suggested that TGBp2 and TGBp3mediated the transport of GFP-TGBp1 through andto plasmodesmata.36 This finding is consistent witha model in which TGBp2 and TGBp3 facilitatethe delivery of the TGBp1–vRNA complex to theplasmodesmata (Figure 2). Haupt et al.37 observed theinteraction of two TGB proteins (TGBp2 and TGBp3)from Potato mop-top pomovirus (PMTV) withelements of the secretory and endocytic pathways.They also reported that TGBp2 and TGBp3 showedan early association with the endoplasmic reticulumand were colocalized in motile granules on theendoplasmic reticulum-actin network.37 Furthermore,a recent study suggested that targeting of PMTV to theplasmodesmata and intercellular PMTV movementare mediated by a membrane-anchored, tyrosine-based motif on the lumen side of the endoplasmicreticulum and the C-terminal transmembrane domainof the TGBp3 movement proteins.38 However, in thecase of Poa semi-latent virus (a hordei virus), thetargeting of TGBp3 to the plasmodesmata is mediatedby diffusion in the cytoplasm.39 Shemyakina et al.40
recently proposed that the interaction of TGBp1with microtubules is important for the formation ofvRNP-complex bodies destined for transport to theplasmodesmata and for the degradation of excessiveTGBp1.40
Tobacco Mosaic VirusTobacco mosaic virus (TMV) is a monopartite RNAvirus, a member of the Virgaviridae family, and oneof the most intensively analyzed plant viruses usedto study local and systemic viral RNA trafficking.The TMV movement protein belongs to the 30Ksuperfamily of viral movement proteins, which isrequired for the cell-to-cell spread of viral RNAthrough the manipulation of plasmodesmata18,41
(Figure 2). The TMV movement protein was thefirst viral movement protein shown to bind single-stranded RNAs to form a vRNP complex, andthe in vivo protein composition of this complexremains unknown. How the vRNP complexes aredelivered to the orifices of plasmodesmata has been
the subject of intensive investigation. An increase inthe permeability of plasmodesmata is likely correlatedwith a reduction in the accumulation of callose in theneck region of plasmodesmata. This hypothesis wasrecently supported by exciting observations of Uekiet al.42 The authors found that the TMV movementprotein interacts with a host receptor, an ankyrinrepeat containing protein at the plasmodesmata andthat this interaction decreases callose accumulationthereby increasing intercellular movement of thevirus.42 Another report also suggested that theTMV movement protein interacts with calreticulin, aprotein involved in Ca2+ sequestration and associatedwith the endoplasmic reticulum lumen, at theplasmodesmata.43 Although this interaction was notclearly proven, the overexpression of calreticulinaltered the localization of the movement protein fromthe plasmodesmata to microtubules, which resulted inreduced viral movement. Although the intercellulartrafficking of TMV movement protein primarilyoccurred on the endoplasmic reticulum membrane,endoplasmic reticulum-associated actin filaments andactin-binding proteins might also be involved in TMVtransport.44
In tobacco plants, the phosphorylation of theTMV movement protein residues Ser258, Thr261,and Ser265 activates the ability of TMV movementprotein to expand plasmodesmata and move fromcell-to-cell. Subsequent studies have revealed that theplasmodesmal-associated protein kinase 1 (PAPK1),which was identified from tobacco and Arabidopsisplants, phosphorylates viral movement proteinsand a subset of endogenous non-cell-autonomousproteins.45 The TMV movement proteins is alsophosphorylated in vitro by CK2 kinase, and a tobaccoCK2-like kinase can phosphorylate the PVX TBGp1movement protein.46 The TMV movement proteinfunctions synergistically with its replicase to controlthe translocation of TMV by lateral diffusion inthe desmotubule of tobacco plasmodesmata.47 Thedistinct mode of intercellular transport of vRNA relieson the action of movement proteins, which might co-evolve with the cell-to-cell macromolecular traffickingpathways of the viral host.
ENDOGENOUS mRNA TRAFFICKING
Studies conducted during the past 20 years have led tothe emergence of a new paradigm in which mobilemRNAs play a pivotal role in the supra-cellularcontrol of gene regulation to coordinate plant cellulargrowth and development (Table 1), although in manycases a physiological role for the transport is notproven.
© 2013 John Wiley & Sons, Ltd.
WIREs RNA Intercellular RNA movement in plants
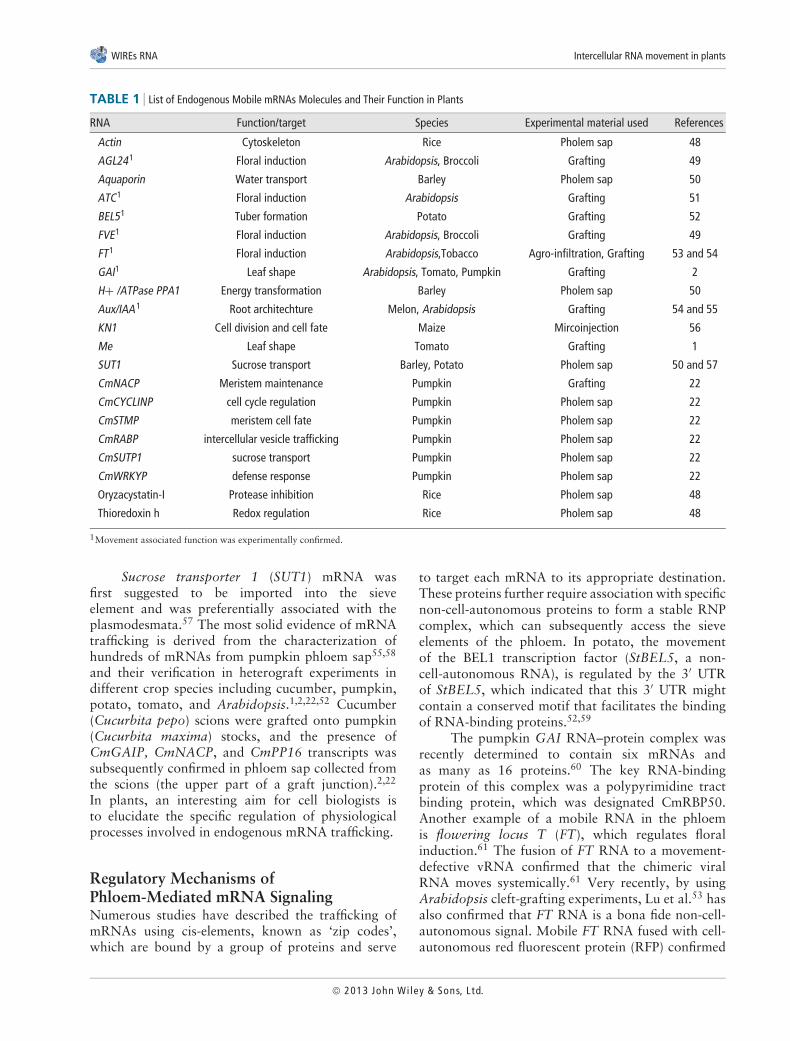
TABLE 1 List of Endogenous Mobile mRNAs Molecules and Their Function in Plants
RNA Function/target Species Experimental material used References
Actin Cytoskeleton Rice Pholem sap 48
AGL241 Floral induction Arabidopsis, Broccoli Grafting 49
Aquaporin Water transport Barley Pholem sap 50
ATC1 Floral induction Arabidopsis Grafting 51
BEL51 Tuber formation Potato Grafting 52
FVE1 Floral induction Arabidopsis, Broccoli Grafting 49
FT1 Floral induction Arabidopsis,Tobacco Agro-infiltration, Grafting 53 and 54
GAI1 Leaf shape Arabidopsis, Tomato, Pumpkin Grafting 2
H+ /ATPase PPA1 Energy transformation Barley Pholem sap 50
Aux/IAA 1 Root architechture Melon, Arabidopsis Grafting 54 and 55
KN1 Cell division and cell fate Maize Mircoinjection 56
Me Leaf shape Tomato Grafting 1
SUT1 Sucrose transport Barley, Potato Pholem sap 50 and 57
CmNACP Meristem maintenance Pumpkin Grafting 22
CmCYCLINP cell cycle regulation Pumpkin Pholem sap 22
CmSTMP meristem cell fate Pumpkin Pholem sap 22
CmRABP intercellular vesicle trafficking Pumpkin Pholem sap 22
CmSUTP1 sucrose transport Pumpkin Pholem sap 22
CmWRKYP defense response Pumpkin Pholem sap 22
Oryzacystatin-I Protease inhibition Rice Pholem sap 48
Thioredoxin h Redox regulation Rice Pholem sap 48
1Movement associated function was experimentally confirmed.
Sucrose transporter 1 (SUT1) mRNA wasfirst suggested to be imported into the sieveelement and was preferentially associated with theplasmodesmata.57 The most solid evidence of mRNAtrafficking is derived from the characterization ofhundreds of mRNAs from pumpkin phloem sap55,58
and their verification in heterograft experiments indifferent crop species including cucumber, pumpkin,potato, tomato, and Arabidopsis.1,2,22,52 Cucumber(Cucurbita pepo) scions were grafted onto pumpkin(Cucurbita maxima) stocks, and the presence ofCmGAIP, CmNACP, and CmPP16 transcripts wassubsequently confirmed in phloem sap collected fromthe scions (the upper part of a graft junction).2,22
In plants, an interesting aim for cell biologists isto elucidate the specific regulation of physiologicalprocesses involved in endogenous mRNA trafficking.
Regulatory Mechanisms ofPhloem-Mediated mRNA SignalingNumerous studies have described the trafficking ofmRNAs using cis-elements, known as ‘zip codes’,which are bound by a group of proteins and serve
to target each mRNA to its appropriate destination.These proteins further require association with specificnon-cell-autonomous proteins to form a stable RNPcomplex, which can subsequently access the sieveelements of the phloem. In potato, the movementof the BEL1 transcription factor (StBEL5, a non-cell-autonomous RNA), is regulated by the 3′ UTRof StBEL5, which indicated that this 3′ UTR mightcontain a conserved motif that facilitates the bindingof RNA-binding proteins.52,59
The pumpkin GAI RNA–protein complex wasrecently determined to contain six mRNAs andas many as 16 proteins.60 The key RNA-bindingprotein of this complex was a polypyrimidine tractbinding protein, which was designated CmRBP50.Another example of a mobile RNA in the phloemis flowering locus T (FT), which regulates floralinduction.61 The fusion of FT RNA to a movement-defective vRNA confirmed that the chimeric viralRNA moves systemically.61 Very recently, by usingArabidopsis cleft-grafting experiments, Lu et al.53 hasalso confirmed that FT RNA is a bona fide non-cell-autonomous signal. Mobile FT RNA fused with cell-autonomous red fluorescent protein (RFP) confirmed
© 2013 John Wiley & Sons, Ltd.

Advanced Review wires.wiley.com/rna
the mobility of RFP-FT RNA rather than RFP-FTprotein. Deletion analysis also revealed that 1–210nucleotides of the FT coding sequence is an RNAmobile motif which acts as a part of the systemicfloral signaling.53 It is also exciting to vascularbiologists that the presence of Auxin/Indoleacetic acid(Aux/IAA) gene family in phloem sap which encodekey transcriptional regulators in auxin signaling.Using a combination of vasculature-enriched samplingand grafting experiments, Notaghuci et al.54 identifiedArabidopsis IAA18 and IAA28 as phloem mobileRNAs which are transported from their source (invascular tissue of mature leaves) to sink (root).54
Given that numerous mobile transcripts might betrafficking through the sieve-tube system of a plant’sphloem, a single, common motif is unlikely to bepresent in all of the phloem-mobile RNAs. Therefore,a future challenge in plant vascular biology will beto understand the complex regulatory mechanismsof phloem-based RNA trafficking; research aimedat addressing this challenge will provide majorinsights into the global biological processes underlyingphysiology and development.
The Biological Function of Cell-to-CellMobile mRNAsIn higher plants, various fundamental biologicalprocesses, including developmental transitions and
patterning, are coordinated by cell-to-cell andlong-distance mRNA trafficking (Figure 3). Thisremote-control system supports precise and effectiveregulation that fine-tunes many plant developmentaland physiological programs (Figure 3). Long-distancetrafficking of pumpkin CmNACP mRNA, whichencodes a member of the NAC (NAM, ATAF1/2and CUC2) domain gene family, was involved inapical meristem development.22 In situ RT-PCRanalysis confirmed the accumulation of CmNACPmRNA in vegetative, root, and floral meristems,which is consistent with the hypothesized functionof this gene. Long-distance, phloem-based movementof several other endogenous mRNAs was validatedusing a grafting technique: CmCYCLINP is involvedin cell cycle regulation; CmGAIP is involved in theregulation of gibberellin response; CmSTMP is a cell-fate determinant in meristems; CmRABP is requiredfor intercellular vesicle trafficking; CmSUTP1 acts insieve-element sucrose transport; and CmWRKYP isinvolved in defense responses.
The functional importance of a mobile tran-script affecting tomato leaf morphology was demon-strated because the PFP-LeT6 [also known as mouseears (Me)] mobile transcript was shown to producethe Me phenotype.1 The transport of GAI tran-scripts in heterografts also induced altered leafletmorphology in wild type scions, which is consistentwith the phenotypes that result from impaired GA
FIGURE 3 | A simplified model forendogenous, mRNA trafficking mechanism actingvia the phloem to regulate plant development.(a) The gibberellic acid insensitive (GAI) RNAregulates leaf morphology in Arabidopsis, tomatoand pumpkin, (b, d) NAC (a member of NACdomain gene family) RNA controls shoot and rootapical meristem development in pumpkin,(c) Movement of BEL1 transcription factor (BEL5)mRNA from leaf to stolon tip facilitates tuberformation in potato, and (e) mobile arabidopsisthaliana centroradialis homolog (ATC), andflowering locus T (FT), FVE, agamous-like24(AGL24) RNA negative and positively regulatefloral induction respectively in Arabidopsis.
Floral induction
ATC, FT, FVE, AGL24
(e)
(b)(a)
(d)(c)
Leaf development
Root apical meristem?
Shoot apical meristem?
gai
NAC
NAC
BEL5
Tuber development
© 2013 John Wiley & Sons, Ltd.

WIREs RNA Intercellular RNA movement in plants
signaling2 (Figure 3). Based on in situ hybridizationand promoter analysis, Banerjee et al.52 reported thatphloem-mobile StBEL5 mRNA induces potato tuberformation by systemically trafficking a signal from theaerial tissue to the stolon tips (Figure 3). The phloem-mobile FT, FVE and agamous-like 24 (AGL24) RNAsmight function similarly as systemic floral regulatorsin Arabidopsis.49,53 In addition, recent studies sug-gest that mobile Arabidopsis thaliana centroradialishomolog (ATC), an Arabidopsis FT homolog andAUX/IAA transcript negatively regulate flower ini-tiation and lateral root formation, respectively.51,54
Plant phloem sap contains a number of unidentifiedRNAs, which might function as long-distance sig-naling molecules that control plant development andphysiology. Future challenges in this exciting field ofresearch are to identify and characterize the molecularplayers that mediate this cell-to-cell and long-distancetransport network and to understand how these play-ers function in this unique and special delivery systemin plants.
SMALL RNAs-POTENT REGULATORSOF NON-CELL-AUTONOMOUS RNASILENCING
In general, RNA interference (RNAi; also known asRNA silencing) refers collectively to diverse RNAdegradation processes, which are triggered by adouble-stranded RNA. It requires the interactionbetween argonaute (AGO) proteins and siRNAs.62,63
In plants, small, non-protein-coding RNAs associ-ated with RNAi include miRNA and siRNAs, whichcan be post-transcriptional and transcriptional generegulators. There are also three classes of endogenoussiRNAs: trans-acting siRNAs (tasiRNAs), heterochro-matin siRNAs (hcsiRNAs) and natural antisense siR-NAs (natsiRNAs). They usually range from 21- to25-nt in length and can be classified based on theirorigin, characteristics, biogenesis, the identity of theirAGO partner and their associated-mechanism of genesilencing.
A recent, major breakthrough in plant biology isthat these small RNAs appear to function as non-cell-autonomous signaling molecules (Table 2). However,the detailed, molecular mechanisms by which thesesmall RNAs are transported in plants have been littleinvestigated.
The Complexity of RNAi TraffickingSeveral recent studies have revealed the complexity ofthe RNAi spreading network in plants and suggestedthat small RNAs, product of distinct DCL proteins, is ahallmark of RNAi signaling.12,13,41 Previous studies ofplant-specific RNA silencing and trafficking indicatethat the mobile RNAi signals involved entail twodistinct pathways: (1) plasmodesma-mediated cell-to-cell movement and (2) phloem-mediated systemicmovement. Cell-to-cell signaling further follows twosequential events: limited and extensive silencingmovements (Figure 4).70–73
TABLE 2 List of Endogenous Mobile Small RNAs Molecules and Their Function in Plants
RNA Function/target species References
miR156 Juvenile-to-adult transition Pumpkin, Oil seed rape 7 and 64
miR159 GAMYB transcription factors Pumpkin, Oil seed rape 7 and 64
miR165 HD-ZIP III, organ formation Arabidopsis 10
miR166 HD-ZIP III, organ formation Arabidopsis, Oil seed rape 10 and 64
miR167 Auxin response factors (ARF) Pumpkin, Oil seed rape 7 and 64
miR171 Scarecrow-like transcription factors Pumpkin, Oil seed rape 7 and 64
miR172 Juvenile-to-adult transition Arabidopsis, Potato 64 and 65
miR390 Adaxial and abaxial cell patterning Maize 5
miR395 Sulfur uptake Arabidopsis, Oil seed rape 64 and 66
miR398 Copper uptake Arabidopsis, Oil seed rape 64 and 66
miR399 Phosphate uptake Arabidopsis, Oil seed rape 64 and 66
IR-siRNA1 Stress response, epiallelism Arabidopsis 67
hcsiRNA1 DNA methylation, Male and female gamatogenesis Arabidopsis 13,14,67, and 68
tasiRNA Adaxial leaf polarity Arabidopsis, Maize 4 and 69
miRNA, microRNA; IR-siRNA, inverted repeat short interfering RNA; hcsiRNA, heterochromatin siRNA; tasiRNA, trans-acting siRNA.1Movement associated function was experimentally confirmed.
© 2013 John Wiley & Sons, Ltd.
Advanced Review wires.wiley.com/rna
Donor cell(a)
(b)
Recipient cell
Donor cell Recipient cell
IR transgene
dsRNA P
P
P
P P
P
P P
P
P
P
P
DCL4
AGO1
AGO1
PolIVa?
PolIVa? PolIVa? PolIVa?DCL4
DCL3? DCL3?
AGO4? AGO4?
PolIVa?
21nt siRNA
21nt siRNA
JMJ14?
JMJ14?
PolII
HEN1
HEN1
CLSY1?
RDR2?
RDR2?RDR2? RDR2?
RDR6
RDR2?
CLSY1?
Localsilencing spread
Extensivesilencing spread
?
?
?
?
?
? ?
?
? ?
?
?
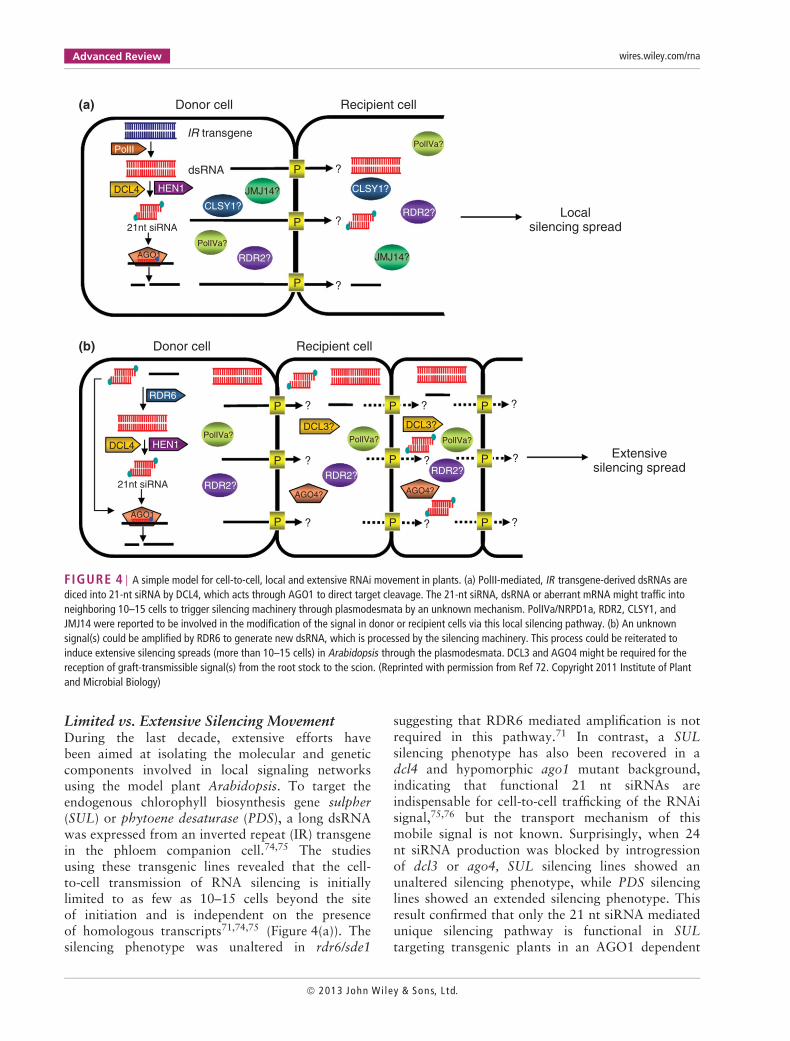
FIGURE 4 | A simple model for cell-to-cell, local and extensive RNAi movement in plants. (a) PolII-mediated, IR transgene-derived dsRNAs arediced into 21-nt siRNA by DCL4, which acts through AGO1 to direct target cleavage. The 21-nt siRNA, dsRNA or aberrant mRNA might traffic intoneighboring 10–15 cells to trigger silencing machinery through plasmodesmata by an unknown mechanism. PolIVa/NRPD1a, RDR2, CLSY1, andJMJ14 were reported to be involved in the modification of the signal in donor or recipient cells via this local silencing pathway. (b) An unknownsignal(s) could be amplified by RDR6 to generate new dsRNA, which is processed by the silencing machinery. This process could be reiterated toinduce extensive silencing spreads (more than 10–15 cells) in Arabidopsis through the plasmodesmata. DCL3 and AGO4 might be required for thereception of graft-transmissible signal(s) from the root stock to the scion. (Reprinted with permission from Ref 72. Copyright 2011 Institute of Plantand Microbial Biology)
Limited vs. Extensive Silencing MovementDuring the last decade, extensive efforts havebeen aimed at isolating the molecular and geneticcomponents involved in local signaling networksusing the model plant Arabidopsis. To target theendogenous chlorophyll biosynthesis gene sulpher(SUL) or phytoene desaturase (PDS), a long dsRNAwas expressed from an inverted repeat (IR) transgenein the phloem companion cell.74,75 The studiesusing these transgenic lines revealed that the cell-to-cell transmission of RNA silencing is initiallylimited to as few as 10–15 cells beyond the siteof initiation and is independent on the presenceof homologous transcripts71,74,75 (Figure 4(a)). Thesilencing phenotype was unaltered in rdr6/sde1
suggesting that RDR6 mediated amplification is notrequired in this pathway.71 In contrast, a SULsilencing phenotype has also been recovered in adcl4 and hypomorphic ago1 mutant background,indicating that functional 21 nt siRNAs areindispensable for cell-to-cell trafficking of the RNAisignal,75,76 but the transport mechanism of thismobile signal is not known. Surprisingly, when 24nt siRNA production was blocked by introgressionof dcl3 or ago4, SUL silencing lines showed anunaltered silencing phenotype, while PDS silencinglines showed an extended silencing phenotype. Thisresult confirmed that only the 21 nt siRNA mediatedunique silencing pathway is functional in SULtargeting transgenic plants in an AGO1 dependent
© 2013 John Wiley & Sons, Ltd.

WIREs RNA Intercellular RNA movement in plants
manner. The explanation of the expansion of PDSsilencing phenotype is that mutation of DCL3 andAGO4 releases the self-silencing mechanism, andincreased accumulation of the PDS dsRNA generatesthe enhanced silencing phenotype.75
In order to identify mobile signal(s), an elegantstudy was carried out on SUL silencing line byDunoyer et al.12 Through a series of experimentsusing cell-specific rescue of DCL4 function and cell-specific inhibition of RNAi movement by a viralsuppressor, P19, they concluded that exogenous andendogenous siRNAs, as opposed to their precursormolecules, act as mobile silencing signals betweenplant cells. Using a bombardment experiment, theyfurther confirmed that double stranded siRNAs trafficcell-to-cell12 (Figure 4(a)). However, an alternativepossibility that has not been excluded is that ssRNAsmove from cell to cell.
The mobility of miRNAs has also beendemonstrated using artificial miRNA (amiRNA).Using a system based on the silencing of the CH42gene, Felippes and colleagues77 have tracked themobility of silencing signals that were initiated inphloem companion cells by amiRNA and tasiRNA.By genetic analysis, the authors concluded that atasiRNA has a greater range of activity in the non-cell-autonomous pathway than that of a miRNA againstCH42, which indicated that biogenesis can define thenon-autonomous effects of small RNAs. In agreementwith this hypothesis, they also demonstrated that thesilencing signals initiated by different small RNAsdiffered in their genetic requirements.77
Curiously, extensive RNAi movement wasobserved when the trigger (silencing inducer)and target genes were both exogenous, whereasendogenous gene silencing was limited to local sectors(Figure 4b). When an AtSUC2 promoter driving aninverted repeat construct corresponding to the 5′ part(GF) of GFP was introduced into a GFP transgenicline (GFP142), silencing of the GFP occurred fromthe companion cell to the whole leaf lamina.71 Twoenzymes namely RNA-dependent RNA polymerase(RDR6) and silencing defective 3 (SDE3, a putativeRNA helicase-like protein) were reported to functionin the spread of extensive RNA silencing, sinceGFP silencing was restricted to around the vein(∼10–15 cells) in rdr6 and sde3 mutants.71 In thissystem, the accumulation of 21-nt and 24-nt siRNAcorresponding to ‘GF’ was similar to the wild typein rdr6 and sde3, whereas the accumulation of ‘P’-specific (3′ part of GFP), 21-nt secondary siRNAs washigher in wild type than in mutant plants. These resultsindicate a pivotal role of secondary siRNA in extensiveRNA silencing spread.63 Recently, the root-to-shoot
movement of silencing that was extensive and long-distance was observed in Arabidopsis; the silencingwas spread from cell-to-cell through the cells of thecentral stele by a template-dependent, reiterated, andshort-distance spread.78 By grafting experiments usingnon-silenced GFP shoots and silenced GFP roots,Brosnan et al.79 previously showed that rootstockswith a dcl1 mutation or dcl2/dcl3/dcl4 mutationshave the capacity to transmit a silencing signal intothe shoot, which indicated that distinct DCLs arenot involved in the systemic signaling network. Theauthors suggested that long RNAs are the mobile RNAsignal(s)79 (Figure 4(b)). However, the functionalredundancy of different DCLs in RNAi80 might alsoprovide the complexity of a non-cell-autonomousRNAi signaling network in plants.
Systemic Transmission of RNAi SignalsIn plants, unlike the cell-to-cell movement of an RNAisignal, systemic RNAi trafficking occurs over thecourse of days. A mobile signal that is initiated ina few cells of a source leaf (mature leaf) can beactively transmitted over long distances through thephloem to induce silencing in sink organs (youngleaf, inflorescence, and root). The genetic dissectionof phloem-based RNAi movement was first done intobacco plants using grafting experiments.81 Thesegrafting experiments provided clear evidence that highamounts of target transcripts were required for thereception of the silencing signal over long distances.Various forms of nucleic acids, including small RNAsand small RNA precursors, were considered as mobilesignaling molecules because they could be monitoredin silenced phloem.12,13,67 Analyses of phloem sapalso confirmed the existence of small RNAs in thephloem, which further supported the involvement ofsmall RNAs as part of the mobile silencing signal.7
However, the exact molecular form of a mobile RNAsignal in phloem has not been identified.
More recently, conclusive grafting experimentsusing a dcl2/dcl3/dcl4 mutant as either sourceor recipient tissue revealed that the transgene-derived siRNAs and a substantial proportion of theendogenous siRNAs (21–24-nt), had moved across thegraft junction.13 Similarly, using grafting approaches,Dunoyer et al.67 showed that all the size classes ofsiRNAs derived from the endogenous IR were notcell-autonomous and could be transmitted through thegraft union over long distances. In this case, the 24-ntsiRNAs were able to trigger both RNAi and RNA-d ependent DNA methylation of the endogenous IRtarget at a distance. These data established a geneticoverlap for the biogenesis, stability and activity ofsmall RNAs generated from both exogenous and
© 2013 John Wiley & Sons, Ltd.
Advanced Review wires.wiley.com/rna
PHO2
siRNA
siRNA
miR399
SSS
SRS(3)
miR399
LRS
Sho
otS
tem
Roo
t
Xyl
em (
1)
Pi
Pi
Phloem
(4)
Phloem
(2)
IPS1/At4
PHO2
IPS1/At4
Pi uptake and transport
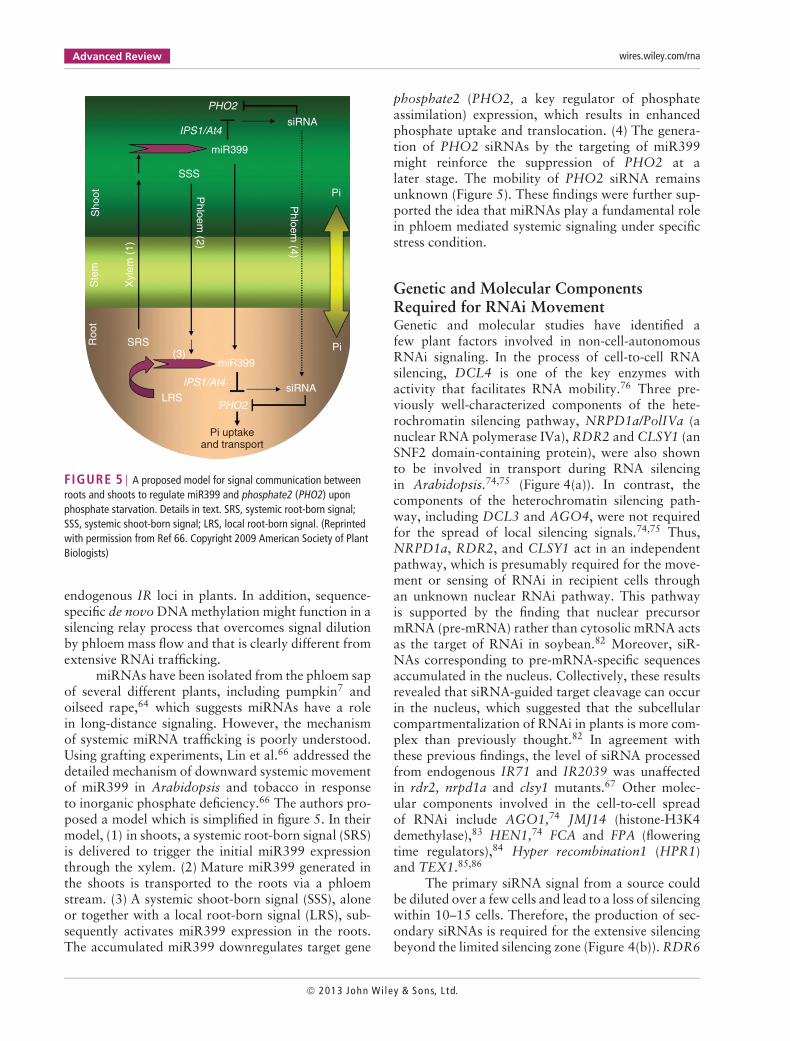
FIGURE 5 | A proposed model for signal communication betweenroots and shoots to regulate miR399 and phosphate2 (PHO2) uponphosphate starvation. Details in text. SRS, systemic root-born signal;SSS, systemic shoot-born signal; LRS, local root-born signal. (Reprintedwith permission from Ref 66. Copyright 2009 American Society of PlantBiologists)
endogenous IR loci in plants. In addition, sequence-specific de novo DNA methylation might function in asilencing relay process that overcomes signal dilutionby phloem mass flow and that is clearly different fromextensive RNAi trafficking.
miRNAs have been isolated from the phloem sapof several different plants, including pumpkin7 andoilseed rape,64 which suggests miRNAs have a rolein long-distance signaling. However, the mechanismof systemic miRNA trafficking is poorly understood.Using grafting experiments, Lin et al.66 addressed thedetailed mechanism of downward systemic movementof miR399 in Arabidopsis and tobacco in responseto inorganic phosphate deficiency.66 The authors pro-posed a model which is simplified in figure 5. In theirmodel, (1) in shoots, a systemic root-born signal (SRS)is delivered to trigger the initial miR399 expressionthrough the xylem. (2) Mature miR399 generated inthe shoots is transported to the roots via a phloemstream. (3) A systemic shoot-born signal (SSS), aloneor together with a local root-born signal (LRS), sub-sequently activates miR399 expression in the roots.The accumulated miR399 downregulates target gene
phosphate2 (PHO2, a key regulator of phosphateassimilation) expression, which results in enhancedphosphate uptake and translocation. (4) The genera-tion of PHO2 siRNAs by the targeting of miR399might reinforce the suppression of PHO2 at alater stage. The mobility of PHO2 siRNA remainsunknown (Figure 5). These findings were further sup-ported the idea that miRNAs play a fundamental rolein phloem mediated systemic signaling under specificstress condition.
Genetic and Molecular ComponentsRequired for RNAi MovementGenetic and molecular studies have identified afew plant factors involved in non-cell-autonomousRNAi signaling. In the process of cell-to-cell RNAsilencing, DCL4 is one of the key enzymes withactivity that facilitates RNA mobility.76 Three pre-viously well-characterized components of the hete-rochromatin silencing pathway, NRPD1a/PolIVa (anuclear RNA polymerase IVa), RDR2 and CLSY1 (anSNF2 domain-containing protein), were also shownto be involved in transport during RNA silencingin Arabidopsis.74,75 (Figure 4(a)). In contrast, thecomponents of the heterochromatin silencing path-way, including DCL3 and AGO4, were not requiredfor the spread of local silencing signals.74,75 Thus,NRPD1a, RDR2, and CLSY1 act in an independentpathway, which is presumably required for the move-ment or sensing of RNAi in recipient cells throughan unknown nuclear RNAi pathway. This pathwayis supported by the finding that nuclear precursormRNA (pre-mRNA) rather than cytosolic mRNA actsas the target of RNAi in soybean.82 Moreover, siR-NAs corresponding to pre-mRNA-specific sequencesaccumulated in the nucleus. Collectively, these resultsrevealed that siRNA-guided target cleavage can occurin the nucleus, which suggested that the subcellularcompartmentalization of RNAi in plants is more com-plex than previously thought.82 In agreement withthese previous findings, the level of siRNA processedfrom endogenous IR71 and IR2039 was unaffectedin rdr2, nrpd1a and clsy1 mutants.67 Other molec-ular components involved in the cell-to-cell spreadof RNAi include AGO1,74 JMJ14 (histone-H3K4demethylase),83 HEN1,74 FCA and FPA (floweringtime regulators),84 Hyper recombination1 (HPR1)and TEX1.85,86
The primary siRNA signal from a source couldbe diluted over a few cells and lead to a loss of silencingwithin 10–15 cells. Therefore, the production of sec-ondary siRNAs is required for the extensive silencingbeyond the limited silencing zone (Figure 4(b)). RDR6
© 2013 John Wiley & Sons, Ltd.

WIREs RNA Intercellular RNA movement in plants
and SDE3 are the key factors amplifying secondarymobile signals.71 Using grafting approaches, Brosnanand colleagues79 showed that in addition to RDR6,the PolIVa-RDR2-DCL3-AGO4 chromatin silencingpathway is involved in the reception of extensive root-to-shoot silencing. Genetic analysis also indicated thatTEX1 processes or transports long RNA molecules foruse as substrates in secondary siRNA production.86
In the Arabidopsis ovule, AGO9, RDR6 and SGS3control female gamete formation by restricting thespecification of gametophytic precursors in a dosage-dependent, non-cell-autonomous manner.14
Most proteins involved in the spread of systemicsilencing remain unknown. Using biochemicalanalyses, Yoo et al.7 first identified phloem smallRNA binding protein 1 (PSRP1), which binds to 25-ntsmall RNAs with high affinity, from pumpkin phloemsap. No clear homologs have been characterizedin Arabidopsis or tobacco, and genetic evidencesupporting the function of this protein is lacking.
Biological Significance of RNAi Spreading inPlantsRNA silencing and its non-cell-autonomous naturemight have a significant role in the coordination ofa vast array of complex plant biological processes,including growth, development, disease resistance,stress response, and the maintenance of nutrient sig-naling. However, research describing the biologicalsignificance of the spread of silencing is still in itsinfancy. In tobacco, a mobile RNAi signal is involvedin viral defense.87,88 During infection, the signal is pri-marily generated in an infected cell, which is translo-cated into growing tissues with or ahead of the system-ically mobile vRNA. The signal triggers an immediatesilencing response against the virus and slows downthe systemic spread of the virus in these tissues.
Endogenous 24-nt siRNAs have the potentialto direct DNA methylation over long distances inrecipient tissues because their epigenetic effects canbe transmitted through grafts.13,67 IR-derived, 24-ntsiRNAs might also have implications in the adaptationof plants to various environmental conditions andin epiallelism.67 Transposon-derived, mobile siRNAsignals also control epigenetic reprogramming inthe male gametophyte in Arabidopsis pollen.68 Inaddition, Olmedo-Monfil et al.14 proposed that themovement of transposon-derived 24-nt hcsiRNAsilencing out of somatic companion cells is necessaryfor the control of gametophyte specification.
Many miRNAs are relatively immobile andcell autonomous, whereas some miRNAs functionas intercellular signals that orchestrate plant tissue
specification and differentiation. For example, in theArabidopsis root, miRNA165/166 moves from theendodermis to the vascular cylinder to regulate classIII homeodomain-leucine zipper transcription factorsacting in xylem patterning in the stele periphery.10
The expression of miR395, miR399, and miR398are known to be induced in the phloem undersulfate-, phosphate-, and copper-deprived conditions,respectively.66 In particular, during the onset ofphosphate deficiency, Arabidopsis mature miR399 iscrucial for enhancing inorganic phosphate uptake andtranslocation66 (Figure 5).
In maize, laser micro-dissection experimentsrevealed an extensive accumulation of miR390precursors (miR390a and miR390b) within theepidermal layer of the incipient leaf, whereas maturemiR390 also accumulates in the epidermal andsub-epidermal layers, which suggested that miR390moves.6 During leaf development, the movement ofmature miR390 through only a few cells regulatedadaxial and abaxial cell patterning via the tasiRNApathway.6
Like miRNAs, RNAi mediated by mobiletasiRNAs also plays diverse roles in the developmentalprocesses of flowering plants. In Arabidopsis, tasiRNAregulation results in vegetative phase change defectsthat are similar to those seen in ago7, rdr6,and sgs3 mutants. In Arabidopsis, TAS3-driventasiRNAs behave non-cell-autonomously to controlauxin response factor (ARF) activities. They alsocontribute to the specification of adaxial leaf polarityby controlling the expression of their targets, theabaxial determinants ARF3 and ARF4.4,69 However,ago7, rdr6, and sgs3 single mutants do not exhibitobvious leaf polarity defects most likely because ofthe existence of functional overlap between the TAS3tasiRNA and Asymmetric leaves (AS) pathways inArabidopsis.69 In contrast, in other plant species,such as maize or rice, mutations in the homologsof Arabidopsis DCL4, SGS3, and RDR6 havemuch stronger developmental phenotypes than thoseexhibited by the Arabidopsis mutants.5,89
CONCLUSION
Over the past decade, many efforts, includingcombinations of genetic, biochemical, molecular andstructural approaches, in the field of intercellular RNAsignaling have provided a molecular underpinning ofthe regulatory mechanisms governing local signalsbetween cells or systemic signals at the whole-plantlevel. Interesting questions include the molecularbasis for the determination of plasmodesma-mediatedcell-to-cell and vascular-mediated long-distance RNA
© 2013 John Wiley & Sons, Ltd.

Advanced Review wires.wiley.com/rna
trafficking, and how the molecular components ofplasmodesmata mediate their gate-keeping function.The current lack of information may be caused bythe indispensable role of macromolecular movement,which provides the RNAs necessary for plant growthand development: this would lead to difficulties in the
isolation of mutants, which could be lethal or havereduced viability. Therefore, an outstanding challengeis to develop innovative and worthwhile approachesto get deeper insights about the mechanisms and bio-logical significance of non-cell-autonomous RNA andRNAi pathways in integrative plant biology.
REFERENCES1. Kim M, Canio W, Kessler S, Sinha N. Develop-
mental changes due to long distance movement of ahomeobox fusion transcript in tomato. Science 2001,293:287–289.
2. Haywood V, Yu TS, Huang NC, Lucas WJ. Phloemlong distance trafficking of GIBBERELLIC ACID-INSENSITIVE RNA regulates leaf development. PlantJ 2005, 42:49–68.
3. Gallagher KL, Benfey PN. Not just another hole inthe wall: understanding intercellular protein trafficking.Genes Dev 2005, 19:189–195.
4. Chitwood DH, Nogueira FT, Howell MD, MontgomeryTA, Carrington JC, Timmermans MC. Pattern for-mation via small RNA mobility. Genes Dev 2009,23:549–554.
5. Nogueira FT, Madi S, Chitwood DH, Juarez MT, Tim-mermans MC. Two small regulatory RNAs establishopposing fates of a developmental axis. Genes Dev2007, 21:750–755.
6. Nogueira FT, Chitwood DH, Madi S, Ohtsu K, Schn-able PS, Scanlon MJ, Timmermans MC. Regulation ofsmall RNA accumulation in the maize shoot apex. PLoSGenet 2009, 5:e1000320.
7. Yoo BC, Kragler F, Varkonyi-Gasic E, Haywood V,Archer-Evans S, Lee YM, Lough TJ, Lucas WJ. A sys-temic small RNA signaling system in plants. Plant Cell2004, 16:1979–2000.
8. Kurata T, Ishida T, Kawabata-Awai C, Noguchi M,Hattori S, Sano R, Nagasaka R, Tominaga R, Koshino-Kimura Y, Kato T, et al. Cell-to-cell movement of theCAPRICE protein in Arabidopsis root epidermal celldifferentiation. Development 2005, 132:5387–5398.
9. Kim JY. Regulation of short-distance transport of RNAand protein. Curr Opin Plant Biol 2005, 8:45–52.
10. Carlsbecker A, Lee JY, Roberts CJ, Dettmer J, Lehes-ranta S, Zhou J, Lindgren O, Moreno-Risueno MA,Vaten A, Thitamadee S, et al. Cell signalling bymicroRNA165/6 directs gene dose dependent root cellfate. Nature 2010, 465:316–321.
11. Ding B, Wang Y. Viroids: small probes for exploringthe vast universe of RNA trafficking in plants. J IntegrPlant Biol 2010, 52:28–39.
12. Dunoyer P, Schott G, Himber C, Meyer D, TakedaA, Carrington JC, Voinnet O. Small RNA duplexes
function as mobile silencing signals between plant cells.Science 2010, 328:912–916.
13. Molnar A, Melnyk CW, Bassett A, Hardcastle TJ, DunnR, Baulcombe DC. Small silencing RNAs in plants aremobile and direct epigenetic modification in recipientcells. Science 2010, 328:872–875.
14. Olmedo-Monfil V, Duran-Figueroa N, Arteaga-Vazquez M, Demesa-Arevalo E, Autran D, GrimanelliD, Slotkin RK, Martienssen RA, Vielle-Calzada JP.Control of female gamete formation by a small RNApathway in Arabidopsis. Nature 2010, 464:628–632.
15. Brosnan CA, Voinnet O. Cell-to-cell and long-distancesiRNA movement in plants: mechanisms and biologicalimplications. Curr Opin Plant Biol 2011, 14:580–587.
16. Rim Y, Huang L, Chu H, Han X, Cho WK, Jeon CO,Kim HJ, Hong JC, Lucas WJ, Kim JY. Analysis of Ara-bidopsis transcription factor families revealed extensivecapacity for cell-to-cell movement as well as discretetrafficking patterns. Mol Cells 2011, 32:519–526.
17. Lucas WJ, Yoo B-C, Kragler F. RNA as a long-distanceinformation macromolecule in plants. Nat Rev Mol CellBiol 2001, 2:849–857.
18. Lucas WJ. Plant viral movement proteins: Agents forcell-to-cell trafficking of viral genomes. jtlVirology2006, 344:169–184.
19. Oparka KJ, Roberts AG, Boevink P, Santa Cruz S,Roberts I, Pradel KS, Imlau A, Kotlizky G, Sauer N, EpelB. Sample, but not branched, plasmodesmata allow thenonspecific trafficking of proteins in developing tobaccoleaves. Cell 1999, 97:743–754.
20. Rinne PLH, van der Schoot C. Plasmodesmata atthe crossroads between development dormancy, anddefense. Botany 2003, 81:1182–1197.
21. Chen XY, Kim JY. Transport of macromoleculesthrough plasmodesmata and the phloem. Physiol Plant2006, 126:560–571.
22. Ruiz-Medrano R, Xoconostle-Caızares B, Lucas WJ.Phloem long-distance transport of CmNACP mRNA:implications for supracellular regulation in plants.Development 1999, 126:4405–4419.
23. Ding B, Kwon MO, Hammond R, Owens R. Cell-to-cellmovement of potato spindle tuber viroid. Plant J 1997,12:931–936.
24. Qi Y, Pelissier T, Itaya A, Hunt E, Wassenegger M,Ding B. Direct role of a viroid RNA motif in mediating
© 2013 John Wiley & Sons, Ltd.

WIREs RNA Intercellular RNA movement in plants
directional RNA trafficking across a specific cellularboundary. Plant Cell 2004, 16:1741–1752.
25. Palukaitis P. Potato spindle tuber viroid: investigation ofthe long-distance, intra-plant transport route. Virology1987, 158:239–241.
26. Zhu Y, Green L, Woo YM, Owens R, Ding B. Cellularbasis of potato spindle tuber viroid systemic movement.Virology 2001, 279:69–77.
27. Zhong X, Archual AJ, Amin AA, Ding B. A genomicmap of viroid RNA motifs critical for replication andsystemic trafficking. Plant Cell 2008, 20:35–47.
28. Zhu Y, Qi Y, Xun Y, Owens R, Ding B. Movementof potato spindle tuber viroid reveals regulatory pointsof phloem-mediated RNA traffic. Plant Physiol 2002,130:138–146.
29. Roberts IM, Boevink P, Roberts AG, Sauer N, ReichelC, Oparka KJ. Dynamic changes in the frequencyand architecture of plasmodesmata during the sink-source transition in tobacco leaves. Protoplasma 2001,218:31–44.
30. Benitez-Alfonso Y, Faulkner C, Ritzenthaler C, MauleAJ. Plasmodesmata: gateways to local and systemicvirus infection. Mol Plant–Microbe Interact 2010,23:1403–1412.
31. Niehl A, Heinlein M. Cellular pathways for viraltransport through plasmodesmata. Protoplasma 2011,248:75–99.
32. Fedorkin ON, Solovyev AG, Yelina NE, Jr, ZamyatninAA, Zinovkin RA, Makinen K, Schiemann J, MorozovSY. Cell-to-cell movement of Potato virus X involvesdistinct functions of the coat protein. J Gen Virol 2001,82:449–458.
33. Verchot-Lubicz J. A new cell-to-cell transport modelfor potexviruses. Mol Plant Microb Interact 2005,18:283–290.
34. Lough TJ, Lee RH, Emerson SJ, Forster RL, LucasWJ. Functional analysis of the 5′ untranslated region ofpotexvirus RNA reveals a role in viral replication andcell-to-cell movement. Virology 2006, 351:455–465.
35. Morozov SY, Solovyev AG. Triple gene block: modu-lar design of a multifunctional machine for plant virusmovement. J Gen Virol 2003, 84:1351–1366.
36. Zamyatnin AA, Jr. Solovyev AG, Savenkov EI, Ger-mundsson A, Sandgren M, Valkonen JP, Morozov SY.Transient coexpression of individual genes encoded bythe triple gene block of potato mop-top virus revealsrequirements for TGBp1 trafficking. Mol Plant MicrobInteract 2004, 17:921–930.
37. Haupt S, Cowan GH, Ziegler A, Roberts AG, OparkaKJ, Torrance L. Two plant-viral movement proteinstraffic in endocytic recycling pathway. Plant Cell 2005,17:164–181.
38. Tilsner J, Cowan GH, Roberts AG, Chapman SN,Ziegler A, Savenkov E, Torrance L. Plasmodesmaltargeting and intercellular movement of potato mop-top pomovirus is mediated by a membrane anchored
tyrosine-based motif on the lumenal side of the endo-plasmic reticulum and the C-terminal transmembranedomain in the TGB3 movement protein. Virology 2010,402:41–51.
39. Schepetilnikov MV, Solovyev AG, Gorshkova EN,Schiemann J, Prokhnevsky AI, Dolja VV, MorozovSY. Intracellular targeting of a hordeiviral membrane-spanning movement protein: sequence requirements andinvolvement of an unconventional mechanism. J Virol2008, 82:1284–1293.
40. Shemyakina EA, Solovyev AG, Leonova OG, PopenkoVI, Schiemann J, Morozov SY. The role of microtubuleassociation in plasmodesmal targeting of potato mop-top virus movement protein TGBp1. Open Virol J 2011,5:1–11.
41. Hyun TK, Uddin MN, Rim Y, Kim J-Y. Cell-to-celltrafficking of RNA and RNA silencing through plas-modesmata. Protoplasma 2011, 248:101–116.
42. Ueki S, Spektor R, Natale DM, Citovsky V. ANK, ahost cytoplasmic receptor for the tobacco mosaic viruscell to cell movement protein, facilitates intercellulartransport through plasmodesmata. PLoS Path 2010,6:e1001201.
43. Chen M-H, Tian G-W, Gafni Y, Citovsky V. Effects ofcalreticulin on viral cell-to-cell movement. Plant Physiol2005, 138:1866–1876.
44. Hofmann C, Niehl A, Sambade A, Steinmetz A, Hein-lein M. Inhibition of tobacco mosaic virus movementby expression of an actin-binding protein. Plant Physiol2009, 149:1810–1823.
45. Lee JY, Taoka KI, Yoo BC, Ben-Nissan G, Lim DJ,Lucas WJ. Plasmodesmal-associated protein kinase intobacco and Arabidopsis recognizes a subset of non-cell-autonomous proteins. Plant Cell 2005, 17:2817–2831.
46. Modena NA, Zelada AM, Conte F, Mentaberry A.Phosphorylation of the TGBp1 movement protein ofPotato virus X by a Nicotiana tabacum CK2-like activ-ity. Virus Res 2008, 137:16–23.
47. Guenoune-Gelbart D, Elbaum M, Sagi G, Levy A,Epel BL. Tobacco mosaic virus (TMV) replicase andmovement protein function synergistically in facilitatingTMV spread by lateral diffusion in the plasmodes-mal desmotubule of Nicotiana benthamiana. Mol PlantMicrobe Interact 2008, 21:335–345.
48. Sasaki T, Chino M, Hayashi H, Fujiwara T. Detectionof several mRNA species in rice phloem sap. Plant CellPhysiol 1998, 39:895–897.
49. Yang H-W, Yu T-S. Arabidopsis floral regulators FVEand AGL24 are phloem-mobile RNAs. Bot Stud 2010,51:17–26.
50. Doering-Saad C, Newbury HJ, Bale JS, Pritchard J. Useof aphid stylectomy and RT-PCR for the detection oftransporter mRNAs in sieve elements. J Exp Bot 2002,53:631–637.
51. Huang N-C, Jane W-N, Chen J, Yu T-S. Arabidopsisthaliana CENTRORADIALIS homologue (ATC) acts
© 2013 John Wiley & Sons, Ltd.

Advanced Review wires.wiley.com/rna
systemically to inhibit floral initiation in Arabidopsis.Plant J 2012, 72:175–184.
52. Banerjee AK, Chatterjee M, Yu Y, Suh SG, Miller WA,Hannapel DJ. Dynamics of a mobile RNA of potatoinvolved in a long distance signaling pathway. PlantCell 2006, 18:3443–3457.
53. Lu K-J, Huang N-C, Liu Y-S, Lu C-A, Yu T-S.Long-distance movement of Arabidopsis FLOWER-ING LOCUS T RNA participates in systemic floralregulation. RNA Biol 2012, 9:653–662.
54. Notaghuci M, Wolf S, Lucas WJ. Phloem-mobileAux/IAA transcripts target to the root tip andmodify root architecture. J Integr Plant Biol 2012,54:760–772.
55. Omid A, Keilin T, Glass A, Leshkowitz D, Wolf S.Characterization of phloem-sap transcription profile inmelon plants. J Exp Bot 2007, 58:3645–3656.
56. Kim JY, Rim Y, Wang J, Jackson D. A novel cell-to-cell trafficking assay indicates that the KNOXhomeodomain is necessary and sufficient for intercel-lular protein and mRNA trafficking. Genes Dev 2005,19:788–793.
57. Kuhn C, Franceschi VR, Schulz A, Lemoine R, FrommerWB. Macromolecular trafficking indicated by localiza-tion and turnover of sucrose transporters in enucleatesieve elements. Science 1997, 275:1298–1300.
58. Kehr J, Buhtz A. Long distance transport and move-ment of RNA through the phloem. J Exp Bot 2008,59:85–92.
59. Banerjee AK, Lin T, Hannapel DJ. Untranslated regionsof mobile transcript mediate RNA metabolism. PlantPhysiol 2009, 151:1831–1843.
60. Ham BK, Brandom JL, Xoconostle-Cazares B, Ring-gold V, Lough TL, Lucas WJ. A polypyrimidine tractbinding protein, pumpkin RBP50, forms the basis of aphloem-mobile ribonucleoprotein complex. Plant Cell2009, 21:197–215.
61. Li C, Zhang K, Zeng X, Jackson S, Zhou Y, HongY. A cis element within Flowering Locus T mRNAdetermines its mobility and facilitates trafficking ofheterologous viral RNA. J Virol 2009, 83:3540–3548.
62. Baulcombe D. RNA silencing in plants. Nature 2004,431:356–363.
63. Heo I, Kim VN. Regulating the regulators: posttrans-lational modifications of RNA silencing factors. Cell2009, 139:28–31.
64. Buhtz A, Springer F, Chappell L, Baulcombe DC, KehrJ. Identification and characterization of small RNAsfrom the phloem of Brassica napus. Plant J 2008,53:739–749.
65. Martin A, Adam H, Diaz-Mendoza M, Zur-czak M, Gonzalez-Schain ND, Suarez-Lopez P.Graft-transmissible induction of potato tuberizationby the micro-RNA miR172. Development 2009,136:2873–2881.
66. Lin SI, Chiang SF, Lin WY, Chen JW, Tseng CY, WuPC, Chiou TJ. Regulatory network of microRNA399and PHO2 by systemic signaling. Plant Physiol 2008,147:732–746.
67. Dunoyer P, Brosnan CA, Schott G, Wang Y, Jay F,Alioua A, Himber C, Voinnet O. An endogenous,systemic RNAi pathway in plants. EMBO J 2010,29:1699–1712.
68. Slotkin RK, Vaughn M, Borges F, Tanurdzic M, BeckerJD, Feijo JA, Martienssen RA. Epigenetic reprogram-ming and small RNA silencing of transposable elementsin pollen. Cell 2009, 136:461–472.
69. Garcia D, Collier SA, Byrne ME, Martienssen RA. Spec-ification of leaf polarity in Arabidopsis via the trans-acting siRNA pathway. Curr Biol 2006, 16:933–938.
70. Chitwood DH, Timmermans MCP. Small RNAs are onthe move. Nature 2010, 467–:415–467.
71. Himber C, Dunoyer P, Moissiard G, Ritzenthaler C,Voinnet O. Transitivity-dependent and -independentcell-to-cell movement of RNA silencing. EMBO J 2003,22:4523–4533.
72. Uddin MN, Kim JY. Non-cell-autonomous RNA silenc-ing spread in plants. Bot Stud 2011, 52:129–136.
73. Kalantidis K, Schumacher HT, Alexiadis T, Helm JM.RNA silencing movement in plants. Biol Cell 2008,100:13–26.
74. Dunoyer P, Himber C, Ruiz-Ferrer V, Alioua A,Voinnet O. Intra- and intercellular RNA interferencein Arabidopsis thaliana requires components of themicroRNA and heterochromatic silencing pathways.Nat Genet 2007, 39:848–856.
75. Smith LM, Pontes O, Searle I, Yelina N, Yousafzai FK,Herr AJ, Pikaard CS, Baulcombe DC. An SNF2 proteinassociated with nuclear RNA silencing and the spreadof a silencing signal between cells in Arabidopsis. PlantCell 2007, 19:1507–1521.
76. Dunoyer P, Himber C, Voinnet O. DICER-LIKE 4is required for RNA interference and produces the21-nucleotide small interfering RNA component ofthe plant cell-tocell silencing signal. Nat Genet 2005,37:1356–1360.
77. Felippes FF, Ott F, Weigel D. Comparative analysisof non-autonomous effects of tasiRNAs and miR-NAs in Arabidopsis thaliana. Nucleic Acids Res 2011,39:2880–2889.
78. Liang D, White RG, Waterhouse PM. Gene silencingin Arabidopsis spreads from the root to the shoot,through a gating barrier, by template-dependent, non-vascular, cell-to-cell movement. Plant Physiol 2012,159:984–1000.
79. Brosnan CA, Mitter N, Christie M, Smith NA,Waterhouse PM, Carroll BJ. Nuclear gene silencingdirects reception of long-distance mRNA silencingin Arabidopsis. Proc Natl Acad Sci U S A 2007,104:14741–14746.
© 2013 John Wiley & Sons, Ltd.

WIREs RNA Intercellular RNA movement in plants
80. Deleris A, Bartolome JG, Bao J, Kasschau KD, Carring-ton JC, Voinnet O. Hierarchical action and inhibitionof plant Dicer-Like proteins in antiviral defense. Science2006, 313:68–71.
81. Palauqui J-C, Elmayan T, Pollien J-M, Vaucheret H.Systemic acquired silencing: transgene-specific post-transcriptional silencing is transmitted by grafting fromsilenced stocks to non-silenced scions. EMBO J 1997,16:4738–4745.
82. Hoffer P, Ivashuta S, Pontes O, Vitins A, Pikaard C,Mroczka A, Wagner N, Voelker T. Posttranscriptionalgene silencing in nuclei. Proc Natl Acad Sci USA 2011,108:409–414.
83. Searle IR, Pontes O, Melnyk CW, Smith LM, BaulcombeDC. JMJ14, a JmjC domain protein, is required for RNAsilencing and cell-to-cell movement of an RNA silencingsignal in Arabidopsis. Genes Dev 2010, 24:986–991.
84. Baurle I, Smith LMA, Baulcombe DC, Dean C.Widespread role for the flowering time regulators FCAand FPA in siRNA-directed chromatin silencing. Science2007, 318:109–112.
85. Jauvion V, Elmayan T, Vaucheret H. The conservedRNA trafficking proteins HPR1 and TEX1 are involvedin the production of endogenous and exogenous smallinterfering RNA in Arabidopsis. Plant Cell 2010,22:2697–2709.
86. Yelina NE, Smith LM, Jones AME, Patel K, KellyKA, Baulcombe DC. Putative Arabidopsis THO/TREXmRNA export complex is involved in transgene andendogenous siRNA biosynthesis. Proc Natl Acad Sci US A 2010, 107:13948–13953.
87. Ratcliff F, Harrison BD, Baulcombe DC. A similar-ity between viral defense and gene silencing in plants.Science 1997, 276:1558–1560.
88. Schwach F, Vaistij FE, Jones L, Baulcombe DC. AnRNA dependent RNA-polymerase prevents meristeminvasion by Potato virus X and is required for the activ-ity but not the production of a systemic silencing signal.Plant Physiol 2005, 138:1842–1852.
89. Liu B, Chen Z, Song X, Liu C, Cui X, Zhao X, Fang J,Xu W, Zhang H, Want X, et al. Oryza sativa dicer-like4reveals a key role for small interfering RNA silencing inplant development. Plant Cell 2007, 19:2705–2718.
© 2013 John Wiley & Sons, Ltd.




















