
Changes in circulating microRNAs after radiochemotherapy in head and neck cancer patients

[CANCER RESEARCH 63, 7926–7935, November 15, 2003]
Both Hepatocyte Growth Factor (HGF) and Stromal-Derived Factor-1 Regulate theMetastatic Behavior of Human Rhabdomyosarcoma Cells, But Only HGFEnhances Their Resistance to Radiochemotherapy
Kacper Jankowski,1 Magda Kucia,1 Marcin Wysoczynski,1 Ryan Reca,1 Dongling Zhao,2 Ela Trzyna,3 John Trent,1
Stephen Peiper,1 Marek Zembala,3 Janina Ratajczak,1 Peter Houghton,4 Anna Janowska-Wieczorek,2 andMariusz Z. Ratajczak1
1James Graham Brown Cancer Center, University of Louisville, Louisville, Kentucky; 2Department of Pathology and Laboratory Medicine and Department of Medicine,University of Alberta and Canadian Blood Services, Edmonton, Alberta, Canada; 3Department of Transplantology, Polish-American Children’s Hospital CMUJ, Kracow, Poland;and 4Molecular Pharmacology, St. Jude Children’s Research Hospital, Memphis, Tennessee
ABSTRACT
Rhabdomyosarcomas (RMSs) are frequently characterized by bonemarrow involvement. Recently, we reported that human RMS cells ex-press the CXC chemokine receptor-4 (CXCR4) and postulated a role forthe CXCR4 stromal-derived factor (SDF)-1 axis in the metastasis of RMScells to bone marrow. Because RMS cells also express the tyrosine kinasereceptor c-MET, the specific ligand hepatocyte growth factor (HGF) thatis secreted in bone marrow and lymph node stroma, we hypothesized thatthe c-MET-HGF axis modulates the metastatic behavior of RMS cells aswell. Supporting this concept is our observation that conditioned mediaharvested from expanded ex vivo human bone marrow fibroblasts che-moattracted RMS cells in an HGF- and SDF-1-dependent manner. Sixhuman alveolar and three embryonal RMS cell lines were examined. Wefound that although HGF, similar to SDF-1, did not affect the prolifera-tion of RMS cells, it induced in several of them: (a) locomotion; (b) stressfiber formation; (c) chemotaxis; (d) adhesion to human umbilical veinendothelial cells; (e) trans-Matrigel invasion and matrix metalloproteinasesecretion; and (f) phosphorylation of mitogen-activated protein kinasep42/44 and AKT. Moreover HGF, but not SDF-1, increased the survival ofRMS cells exposed to radio- and chemotherapy. We also found that themore aggressive alveolar RMS cells express higher levels of c-MET thanembryonal RMS cell lines and “home/seed” better into bone marrow afteri.v. injection into immunocompromised mice. Because we could not findany activating mutations in the kinase region of c-MET or any evidencefor HGF autocrine stimulation, we suggest that the increased response ofRMS cell lines depends on overexpression of functional c-MET. Weconclude that HGF regulates the metastatic behavior of c-MET-positiveRMS cells, directing them to the bone marrow and lymph nodes. Signalingfrom the c-MET receptor may also contribute to the resistance of RMScells to conventional treatment modalities.
INTRODUCTION
RMS5 is the most common soft-tissue sarcoma of adolescence andchildhood and accounts for 5% of all malignant tumors in patients
under 15 years of age (1–10). Most of these tumors originate in thehead and neck region, the urogenital tract, and the extremities. Thereare two major histological subtypes of RMS, ARMS and ERMS.Clinical evidence indicates that ARMS is more aggressive and has asignificantly worse outcome than ERMS. Almost all of the reportedcases associated with marrow involvement, including those presentingas possible acute leukemia, have been of the ARMS type (3). Geneticcharacterization of RMS has identified markers that show excellentcorrelation with histological subtype. Specifically, ARMS is charac-terized by the translocation t(2;13)(q35;q14) in 70% of cases or thevariant t(1;13)(p36;q14) in a smaller percentage of cases. Thesetranslocations disrupt the PAX3 and PAX7 genes on chromosomes 2and 1, respectively, and the FKHR gene on chromosome 13, andgenerate PAX3-FKHR and PAX7-FKHR fusion genes (7–10). Thesefusion genes encode the fusion proteins PAX3-FKHR and PAX7-FKHR, which function as novel transcription factors. As compared towild-type PAX3 and PAX7, the PAX3-FKHR and PAX7-FKHRfusion proteins demonstrate enhanced transcriptional activity and arebelieved to play a role in the survival and dysregulation of the cellcycle in alveolar RMS cells (1–10).
Why RMS cells metastasize to the bone marrow is still poorlyunderstood. It is likely that bone marrow stroma secretes chemoat-tractants for RMS cells, and that these cells are attracted into the bonemarrow microenvironment where they may find favorable conditionsfor survival and expansion. In this study we attempted to identify themolecular mechanisms that may be involved in the metastasis of RMScells to the bone marrow. We recently reported that RMS cells highlyexpress the receptor for the �-chemokine SDF-1 and that the CXCR4-SDF-1 axis regulates the metastatic behavior of RMS (11). RMS cells,however, have also been found to express the tyrosine kinase receptorc-MET (12, 13), known to mediate the multifunctional and potentiallyoncogenic activities of HGF, also known as scatter factor (14, 15),including cell motility, extracellular matrix degradation, angiogenesis,cell proliferation, and survival (16–20). Perturbation of signalingfrom the c-MET receptor because of c-MET overexpression/amplifi-cation, activating mutations, or autocrine HGF-c-MET loops has beenimplicated in the pathogenesis of various tumors (1, 20).
Because normal bone marrow stroma cells and osteoblasts highlyexpress and secrete both HGF and SDF-1 (17, 21–26), we hypothe-sized that the c-MET-HGF axis, like the CXCR4-SDF-1 axis, plays anessential role in directing RMS cells into bone marrow. We focusedon the biological responses to stimulation by exogenous HGF ofc-MET-positive ARMS and ERMS cell lines, such as phosphorylationof signaling proteins, cell proliferation, survival, adhesion, expressionof MMPs, chemotaxis, and chemoinvasion, as well as the survival ofRMS cells exposed to radio- and chemotherapy. Our findings indicatethat the HGF-c-MET axis, like the CXCR4-SDF-1 axis, regulates themetastatic behavior of RMS cells and their metastasis to the bonemarrow and could also contribute to their resistance to conventionaltreatment modalities.
Received 4/7/03; revised 8/4/03; accepted 9/9/03.Grant support: NIH Grant R01 HL61796-01 and UBN grant 3POSE10122 (to
M. Z. R.) and a Canadian Institutes of Health Research Grant (to A. J. W.).The costs of publication of this article were defrayed in part by the payment of page
charges. This article must therefore be hereby marked advertisement in accordance with18 U.S.C. Section 1734 solely to indicate this fact.
K. J. and M. K. contributed equally to this work.Requests for reprints: Mariusz Z. Ratajczak, Stem Cell Biology Program, James
Graham Brown Cancer Center, University of Louisville, 529 South Jackson Street,Louisville, KY 40202. Phone: (502) 852-1788; Fax: (502) 852-3032; E-mail: [email protected].
5 The abbreviations used are: RMS, rhabdomyosarcoma; ARMS, alveolar rhabdomy-osarcoma; ERMS, embryonal rhabdomyosarcoma; HGF, hepatocyte growth factor;SDF-1, stromal-derived factor 1; MMP, matrix metalloproteinase; MoAb, monoclonalantibody; CXCR4, CXC chemokine receptor 4; PECAM-1, platelet/endothelial cell ad-hesion molecule 1; ICAM-1, intercellular adhesion molecule-1; VCAM-1, vascular celladhesion molecule; HUVEC, human umbilical vein endothelial cell; MAPK, mitogen-activated protein kinase; STAT, signal transducers and activators of transcription; RT-PCR, reverse transcription-PCR; PI-3K, phosphatidylinositol 3-kinase; MT1-MMP, mem-brane-type 1 MMP; TIMP-2, tissue inhibitor of metalloproteinases-2; CM, conditionedmedia.
7926
Research. on December 5, 2014. © 2003 American Association for Cancercancerres.aacrjournals.org Downloaded from

MATERIALS AND METHODS
Cell Lines. Nine human RMS cell lines comprising six ARMS lines (RH1,RH2, RH4, RH28, RH30, and CW9019) and three ERMS lines (RH18, RD,and SMS-CTR), all established at St. Jude Hospital, were used. RMS cells usedfor experiments were cultured in RPMI 1640 (Sigma), supplemented with 100IU/ml penicillin, 10 �g/ml streptomycin, and 50 �g/ml neomycin (Life Tech-nologies, Inc., Grand Island, NY) in the presence of 10% heat-inactivated FCS(Life Technologies). The cells were cultured in a humidified atmosphere at 5%CO2, 37°C at an initial cell density of 2.5 � 104 cells/flask (Corning) and themedia were changed every 48 h.
FACS Analysis. The expression of c-MET on various RMS cell lines wasevaluated by FACS as described previously (11, 27). The c-MET antigen wasdetected with anti-c-MET MoAb (UPS Biotechnology, Lake Placid, NY),clone DO-24, followed by staining with secondary antibody FITC-�-murineIgG. Briefly, the cells were stained in PBS (calcium- and magnesium-free)supplemented with 5% BCS (Hyclone, Logan, Utah). After the final wash,cells were fixed in 1% paraformaldehyde, and FACS analysis was performedusing the FACscan (Becton Dickinson, San Jose, CA).
Evaluation of Adhesion Molecules. The expression of adhesion moleculeson RMS cells was evaluated by FACS. Cells were stained with specificanti-PECAM-1, ICAM-1, VCAM-1, E-selectin, and VLA-5 and VLA-4 anti-bodies detected with phycoerythrin-conjugated secondary phycoerythrin-goatanti-mouse MoAbs as described previously (28). The following antibodieswere used for this study: 4G6 (IgG2b, mouse anti-human PECAM-1), gener-ously provided by Dr. Steven Albelda; R6.5 (BIRR-1), a murine IgG2a MoAbdirected against extracellular domain two of the ICAM-1 molecule, fromBoehringer Ingelheim Pharmaceuticals Inc. (Ridgefield, CT); 4B9, an IgG1MoAb directed against human VCAM-1, from Dr. Roy Lobb, Biogen Inc.(Cambridge, MA); and ES2, (IgG1-� mouse anti-human-E-selectin MoAb), pro-vided by Dr. Rodger McEver, University of Oklahoma (Tulsa, OK). Antibodiesagainst �6�1 integrin were purchased from PharMingen (San Diego, CA).
Phosphorylation of Intracellular Pathway Proteins. Western blots weredone on extracts prepared from RMS cell lines (1 � 107 cells), which werekept in RPMI 1640 containing low levels of BSA (0.5%) to render the cellsquiescent. The cells were then divided and stimulated with optimal doses ofHGF (10 ng/ml) for 1 min to 2 h at 37°C before lysing for 10 min on ice inM-Per lysing buffer (Pierce, Rockford, IL) containing protease and phospha-tase inhibitors (Sigma). Subsequently, the extracted proteins were separated oneither a 12 or 15% SDS-PAGE gel, and the fractionated proteins were trans-ferred to a nitrocellulose membrane (Schleicher & Schuell, Keene, NH) asdescribed previously (11, 29, 30). Phosphorylation of the intracellular kinases,44/42 MAPK (Thr-202/Tyr-204) and AKT, and STAT-1, -3, -5, and -6proteins was detected using commercial mouse phospho-specific MoAb (p44/42) or rabbit phospho-specific polyclonal antibodies for each of the remainder(all from New England Biolabs, Beverly, MA) with horseradish peroxidase-conjugated goat anti-mouse IgG or goat anti-rabbit IgG as a secondary anti-body (Santa Cruz Biotechnology Biotechnology, Santa Cruz Biotechnology,CA) as described (11). Equal loading in the lanes was evaluated by strippingthe blots and reprobing with appropriate MoAbs: p42/44 anti-MAPK antibodyclone #9102, anti-AKT antibody clone #9272, anti-STAT-3 #9132 (NewEngland Biolabs), anti-STAT-1 #sc-464 and anti-STAT-6 #sc-1689 (SantaCruz Biotechnology), and anti-STAT-5 #89 (Transduction Laboratories, Lex-ington, KY). The membranes were developed with an ECL reagent (Amer-sham Life Sciences, Little Chalfont, United Kingdom), dried, and subsequentlyexposed to film (HyperFilm; Amersham).
Detection of HGF by Western Blot Analysis. HGF protein was detectedin lysates derived from RMS cell lines using specific rabbit polyclonal anti-body (from R&D, Minneapolis, MN), which was detected with horseradishperoxidase-conjugated goat anti-mouse IgG or goat anti-rabbit IgG as a sec-ondary antibody (Santa Cruz Biotechnology) as described previously (11).
Isolation of mRNA and RT-PCR. For the analysis of HGF mRNA totalmRNA was isolated from RMS cells with the RNeasy Mini Kit (Qiagen Inc.,Valencia, CA), mRNA was reverse-transcribed with TaqMan Reverse Tran-scription Reagents (Applied Biosystems, Branchburg, NJ), and the PCR reac-tion was carried out with the 5�-GAG GGA CAT AAG AAA AGA AGA-3�sense primer and the 5�-GTG TGG TAT CAT GGA ACT CCA-3� antisenseprimer. The predicted size of the RT-PCR product for HGF was 383 bp.Amplified products (10 �l) were electrophoresed on a 1.5% agarose gel and
transferred to a nylon filter. The specificity of the amplified products wasfurther confirmed by Southern blotting (data not shown).
Real-Time RT-PCR. To analyze HGF and SDF-1 mRNA levels, totalmRNA was isolated from cells that were flushed out from the bone marrowcavities of nonirradiated (control) and irradiated (800 cGy) sacrificed micewith the RNeasy Mini Kit (Qiagen, Inc.) and was reverse-transcribed withTaqMan Reverse Transcription Reagents (Applied Biosystems, Foster City,CA). Detection of HGF and SDF-1 and �-actin mRNA levels was performedby real-time RT-PCR assay using an ABI PRISM 7000 Sequence DetectionSystem (ABI, Foster City, CA). A 25-�l reaction mixture contains 12.5 �l ofSYBR Green PCR Master Mix, 100 ng of cDNA template, 5�-TGC GTC CACGAG CTG TTT AC-3� forward and 5�-CCC AAG GGA GTG TCA GGTAGA G-3� reverse primers for SDF-1, and 5�-CTC ACA CCC GCT GGGAGT AC-3� forward and 5�-TCC TTG ACC TTG GAT GCA TTC-3� reverseprimers for HGF. The primers were designed with Primer Express software.The threshold cycle (Ct), i.e., the cycle number at which the amount of theamplified gene of interest reaches a fixed threshold, was determined subse-quently. Relative quantitation of HGF and SDF-1 mRNA expression wascalculated by the comparative Ct method. The relative quantitation value oftarget, normalized to an endogenous control �-actin gene and relative to acalibrator, is expressed as 2��Ct (fold), where �Ct � Ct of the target gene(HGF and SDF-1) � Ct of endogenous control gene (�-actin), and��Ct � �Ct of samples for target gene � �Ct of the calibrator for the targetgene.
Cell Proliferation and Apoptosis. Cells were plated in culture flasks at aninitial density of 104 cells/cm2 in the presence or absence of HGF (10 ng/ml).Cells were counted at 12, 24, 36, 48, and 60 h after culture initiation (in someexperiments also at 72 h). At these time points, cells were harvested from theculture flasks by trypsinization, and the number of cells was determined usinga Burker‘s hemocytometer. Apoptosis was evaluated by the Annexin-V bind-ing assay, and intracellular staining was done for activated caspase-3, asdescribed (28, 29).
Fluorescent Staining of the Actin Cytoskeleton. To visualize the actincytoskeleton, cells were cultured for 12 h on glass coverslips in RPMI 1640supplemented with 10% fetal bovine serum in the absence or presence (10ng/ml) of HGF. Subsequently, the cells were fixed in 3.7% paraformaldehyde/calcium- and magnesium-free PBS for 15 min, permeabilized by 0.1% TritonX-100 in PBS for 1 min at room temperature, and stained with TRITC-phalloidin at a concentration of 500 ng/ml for 1 h. Cells were examined usinga BX51 fluorescence microscope (Olympus America, Melville, NY) equippedin a charge-coupled device camera (Olympus America). Each staining wasrepeated three times for each cell line.
Recording Cellular Motility. Two h before starting the motility experi-ments, CW9019, RH28, RH30, RH18, and SMS-CTR cells were plated inculture dishes at a density of 2 to 2.5 � 104 cells/cm2. Cells were mock-treatedor stimulated by HGF (10 ng/ml) for 30 min, and then the tracks of individualRMS cells were recorded with an inverted Hund Wetzlar microscope usingphase-contrast optics. In brief, the images of locomoting cells were recordedwith a charge-coupled device camera, digitized, and processed. The celltrajectories were constructed from 40 subsequent cell centroid positions re-corded for 200 min at time intervals of 5 min. Fifty cell tracks were recordedunder each of the experimental conditions tested. To analyze the cellularmotility variables, the files containing the tracing data were read into theprogram Mathematica (Wolfram Research Inc., Champaign, IL; Refs. 31–33).
The following parameters characterizing cell movement were computed foreach cell: total length of cell trajectory (�m), i.e., the sum of n straight-linesegments, each corresponding to cell centroid translocation at a given timeinterval; the average speed of cell movement (calculated as length of cell pathin a given time); total length of cell displacement (�m), i.e., the distance fromthe starting point directly to the cell’s final position; average rate of celldisplacement, i.e., the distance from the starting point directly to the cell’s finalposition/time of recording; and coefficient of movement efficiency (CME),corresponding to ratio of cell displacement to cell trajectory length. The CMEequals 1 for cells moving consistently along a single straight line in a givendirection and 0 for random movement.
Transmembrane Chemotaxis. Cells were seeded in RPMI 1640 contain-ing 10% FBS into 6-well plates. After they had adhered to the dish bottom, themedium was changed, and they were made quiescent as described previously(11). Transmembrane chemotaxis across 8-�m pore polycarbonate membranes
7927
C-MET-HGF AND CXCR4-SDF-1 AXES IN RHABDOMYOSARCOMAS
Research. on December 5, 2014. © 2003 American Association for Cancercancerres.aacrjournals.org Downloaded from

covered with 50 �l of fibronectin (50 �g/ml) for 2 h at 37°C and overnight at4°C was examined. Cells were detached (with 0.5 mM EDTA), washed inRPMI 1640, resuspended in RPMI 1640 with 0.5% BSA, and seeded at adensity of 105 in 200 �l into the upper chambers of Transwell inserts (CostarTranswell). The lower chambers were filled with HGF (10 ng/ml) or 0.5%BSA RPMI 1640 (control). To determine whether migration was stimulated bythe gradient of the chemoattractant, in some experiments HGF was also addedto the upper chambers to equalize the difference in concentrations betweenchambers. After 48 h, the inserts were removed from the Transwells, cellsremaining in the upper chambers were scraped off with cotton wool, and cellsthat had transmigrated were counted either on the lower side of the membranesor on the bottom of the Transwells.
Some of the directional migration experiments to conditioned media har-vested from bone marrow-derived fibroblasts were performed on cells prein-cubated for 30 min at 37°C in the presence of 1 �M T140-truncatedpolyphemusin analogue (a gift from Dr. Nobutaka Fuji, Kyoto University,Japan) or preincubated in the presence of 1 �M c-Met inhibitor, K-252(Calbiochem-Novabiochem International). Conditioned media from bone mar-row-derived fibroblasts were derived from cells cultured for 24 h in serum-freemedium as described previously (11).
Adhesion of RMS Cells to HUVEC Cells. RMS cells were labeled withthe fluorescent dye calcein-AM and subsequently seeded (for 5 min) onto the96-well plates covered with HUVEC cells that had been pretreated with HGF(10 ng/ml). After incubation (at 37°C), the plates were washed three times, andcells that adhered to the HUVECs were dissolved using 2% SDS. Fluorescencewas measured using the Fusion Universal Microplate Analyzer (Perkin-Elmer,Boston, MA).
MMP Expression. To evaluate MMP-2 and MMP-9 activities, RMS cellswere incubated for 24 h in serum-free media in the absence (control) orpresence of HGF (10 ng/ml), and zymography was carried out as describedpreviously by us (11, 34–36). To evaluate expression of genes for MMP-2 andMMP-9, as well as MT1-MMP and TIMP-2, total RNA was extracted, and theconversion of mRNA to cDNA was carried out using AMVRT (SeigakuAmerica, Ijamsville, MD); polymerase chain reactions were performed fol-lowing the “primer dropping” method. Sequences for human MMP-2, MMP-9,MT1-MMP, and TIMP-2 were obtained from GenBank (Los Alamos, NM) andused to design primer pairs, as described by us previously (34–36).
Chemoinvasion Assay. The ability of malignant cells to invade the recon-stituted basement membrane Matrigel is regarded as an important measure oftheir metastatic potential. Three ARMS cell lines (RH28, RH30, and CW9019)and two ERMS cell lines (RH1 and RD) were evaluated in a chemoinvasionassay as described and modified by us (11, 34–36). Briefly, cells were loadedonto the upper compartments of Boyden chambers (105 cells/chamber) andincubated for 48 h. Cells that invaded the Matrigel barrier toward media aloneor toward an HGF gradient (10 ng/ml) were counted on the undersides of filtersafter fixation and staining with crystal violet. A chemoinvasion index wascalculated as the ratio of the number of cells invading the Matrigel toward anHGF gradient to the number of cells invading toward media alone.
Sequencing. To address whether mutations activating the c-MET receptorin RMS cells could lead to the hyperreactivity of the HGF-c-MET axis inRMS, in addition to overexpression of c-MET, we sequenced c-MET DNAfrom several RMS cell lines (RH18, RH30, RH28, CW9019, SMS-CTR, andRD) and melanoma SBCL2 (negative control) and searched for potentialactivating mutations. DNA was extracted by means of the QIAamp DNA MiniKit (Qiagen). Sequencing PCR was performed on 10 ng of spectrophotometri-cally quantified DNA with the use of fluorescently labeled nucleotides andexon- specific primers as follows: (a) 5�-TCTTCCTGTTTCAGTCCCCAT-3�and 5�-AGGCCAAAGATAAAATGCTTACTG-3� for exon 15; (b) 5�-CT-CATAAAGGGTT-TGATAAATAATTATTT-3� and 5�-GTTTTATTATA-AGCTATTTATTAGGTTGCA-3� for exon 16; (c) 5�-GCTCTTCCTATCTA-AATTTGACAAAAG-3� and 5�-AGGCCTATTTTGAAGGGATG-3� forexon 17; (d) 5�-TCAGAATTCTAAGGTCAA-AATTAGAACAG-3� and 5�-TTGAACAGTGGGAAACAGATTC-3� for exon 18; (e) 5�-TCTGTA-GATATTCAGCATCATTGTAAAT-3� and 5�-AGTGATAAAACTTCAA-AAAAAGTTGG-3� for exon 19. The PCR product was run on AppliedBiosystems 310 sequencing apparatus and analyzed by means of SequencingAnalysis software (Applied Biosystems). The sequencing was performed onexons 15–19 encoding the tyrosine kinase domain.
Transplants of RMS Cells into Lethally Irradiated Mice. To evaluatethe in vivo metastatic behavior of RMS cells lines, we injected ARMS orERMS cells (2 � 106 cells/mouse) i.v. into lethally irradiated (800 cGy) mice.Cells were injected 24 h after irradiation. Then 36 h after injection of theseRMS cells, the bone marrow cavities of femora, tibiae, and humeri wereflushed to isolate cells present in the bone marrow. The presence of RMS cellsin murine bone marrow (i.e., murine-human chimerism) was evaluated by thedifferences in the level of human �-satellite DNA amplified in the extractsisolated from bone marrow-derived cells, using real-time PCR. Briefly, DNAwas isolated using the QIAamp DNA Mini Kit (Qiagen Inc.). Detection of�-satellite and �-actin DNA levels was performed by real-time PCR using anABI PRISM 7000 Sequence Detection System (ABI). A 25-�l reaction mix-ture contains 12.5 �l of SYBR Green PCR Master Mix, 50 ng of DNAtemplate, 5�-ACACTCTTTTTGCAGGATCTA-3� forward and 5�-AGCAAT-GTGAAACTCTGGGA-3� reverse primers for �-satellite; and 5�-GGA TGCAGA AGG AGA TCA CTG-3� forward and 5�-CGA TCC ACA CGG AGTACT TG-3� reverse primers for �-actin. The primers were designed withPrimer Express software. The threshold cycle (Ct), i.e., the cycle number atwhich the amount of amplified gene of interest reaches a fixed threshold, wasdetermined subsequently. Relative quantitation of �-satellite DNA was calcu-lated with the comparative Ct method described elsewhere (37). The relativequantitation value of target, normalized to a control �-actin gene and relativeto a calibrator, is expressed as 2-��Ct (fold difference), where �Ct � Ct oftarget gene (�-satellite) � Ct of control gene (�-actin), and ��Ct � �Ct ofsamples for target gene � �Ct of calibrator for the target gene.
Statistical Analysis. All results are presented as mean � SE. Statisticalanalyses of the data were performed using the nonparametric Mann-Whitneytest, with P � 0.05 considered significant.
RESULTS
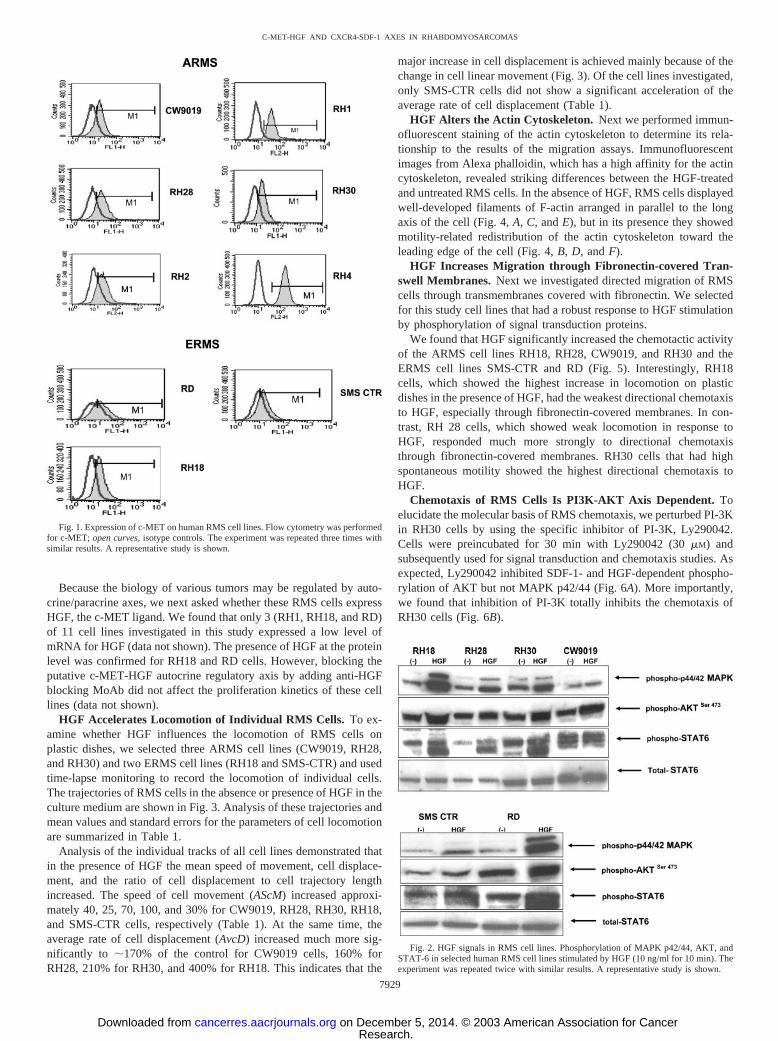
RMS Cell Lines Highly Express c-MET and CXCR4. We phe-notyped several human RMS cell lines using FACS for expression ofc-MET and CXCR4 and found that all eight ARMS cell lines tested(CW9019, RH1, RH28, RH30, RH2, and RH4) stained positively forc-MET (Fig. 1, upper panel). In three ERMS cell lines (SMS-CTR,RD, and RH18), c-MET was also expressed, although at lower levels(Fig. 1, lower panel). We observed that all eight ARMS cells linesalso stained highly positive for CXCR4 (�60% of cells; not shown),which was consistent with our previous findings of high expression ofCXCR4 in ARMS cells (11). As we reported previously (11), CXCR4is expressed at a much lower level in ERMS cells.
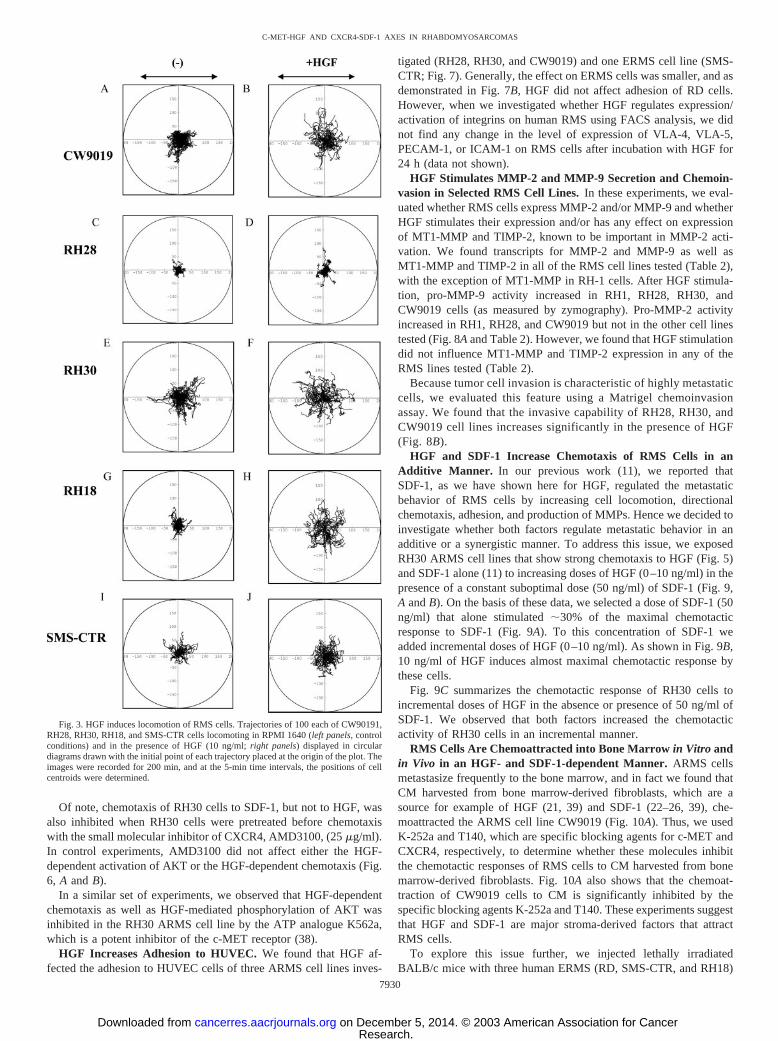
HGF Induces Phosphorylation of MAPK p42/44, AKT, andSTAT-6. Next we turned our attention to the putative role of theHGF-c-MET axis in regulating the metastatic behavior of RMS cellsand examined whether c-MET is functional in RMS cells. We deter-mined that stimulation of RMS cells by HGF induces the phospho-rylation of MAPK p42/44, serine-threonine kinase AKT, and STAT1–6 that have been reported to play an important role in cell prolif-eration, survival, and chemotaxis. We found that ARMS cell linesRH28, RH30, and CW9019 and ERMS cell lines RH18, SMS-CTR,and RD responded to HGF by phosphorylation of MAPK p42/44 orAKT or STAT-6 (Fig. 2).
HGF Does Not Influence Proliferation of RMS Cell Lines. Nextwe examined eight ARMS and three ERMS cell lines to determinewhether HGF affects their proliferation. We stimulated them withHGF or not (control) in serum-free conditions or in media supple-mented with 0.5% BSA. We found that the RMS cell lines proliferatedintensively during 60 h under all types of conditions. The kinetics oftheir proliferation were similar and were not affected by the presenceof HGF in the culture, even when the cells were cultured for up to 7days (data not shown). HGF did not even influence the growth of theRH18, RH30, RD, and SMS-CTR cell lines, which had responded toHGF stimulation by phosphorylation of MAPK p42/44, as describedabove.
7928
C-MET-HGF AND CXCR4-SDF-1 AXES IN RHABDOMYOSARCOMAS
Research. on December 5, 2014. © 2003 American Association for Cancercancerres.aacrjournals.org Downloaded from
Because the biology of various tumors may be regulated by auto-crine/paracrine axes, we next asked whether these RMS cells expressHGF, the c-MET ligand. We found that only 3 (RH1, RH18, and RD)of 11 cell lines investigated in this study expressed a low level ofmRNA for HGF (data not shown). The presence of HGF at the proteinlevel was confirmed for RH18 and RD cells. However, blocking theputative c-MET-HGF autocrine regulatory axis by adding anti-HGFblocking MoAb did not affect the proliferation kinetics of these celllines (data not shown).
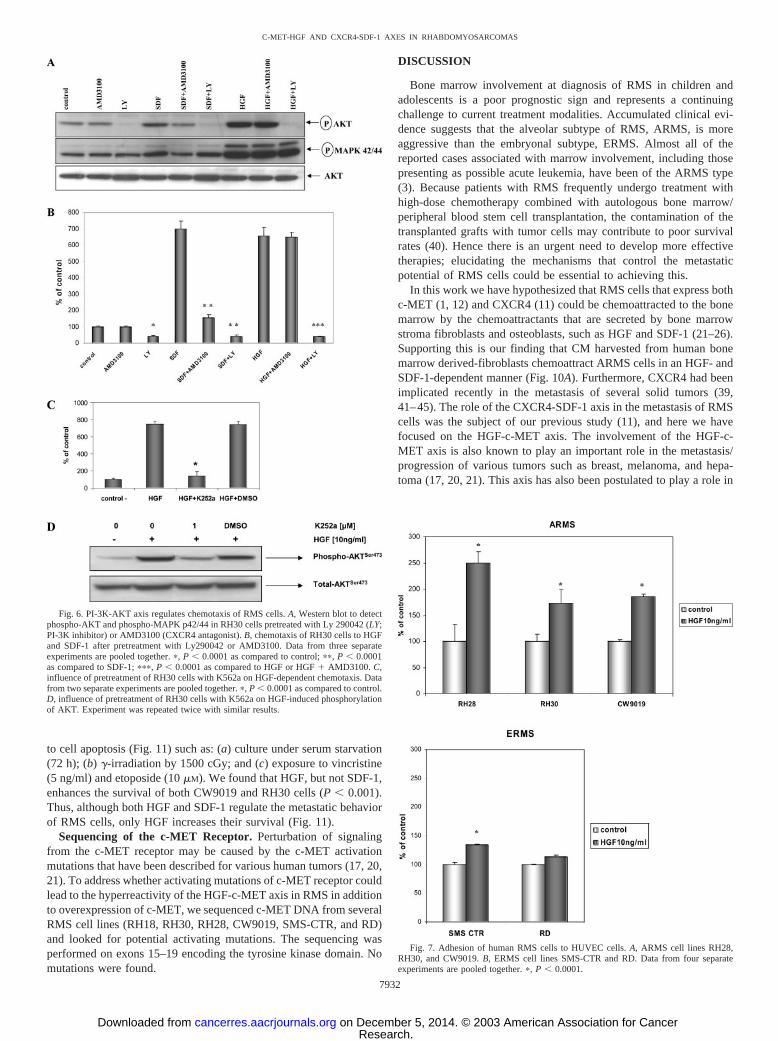
HGF Accelerates Locomotion of Individual RMS Cells. To ex-amine whether HGF influences the locomotion of RMS cells onplastic dishes, we selected three ARMS cell lines (CW9019, RH28,and RH30) and two ERMS cell lines (RH18 and SMS-CTR) and usedtime-lapse monitoring to record the locomotion of individual cells.The trajectories of RMS cells in the absence or presence of HGF in theculture medium are shown in Fig. 3. Analysis of these trajectories andmean values and standard errors for the parameters of cell locomotionare summarized in Table 1.
Analysis of the individual tracks of all cell lines demonstrated thatin the presence of HGF the mean speed of movement, cell displace-ment, and the ratio of cell displacement to cell trajectory lengthincreased. The speed of cell movement (AScM) increased approxi-mately 40, 25, 70, 100, and 30% for CW9019, RH28, RH30, RH18,and SMS-CTR cells, respectively (Table 1). At the same time, theaverage rate of cell displacement (AvcD) increased much more sig-nificantly to 170% of the control for CW9019 cells, 160% forRH28, 210% for RH30, and 400% for RH18. This indicates that the
major increase in cell displacement is achieved mainly because of thechange in cell linear movement (Fig. 3). Of the cell lines investigated,only SMS-CTR cells did not show a significant acceleration of theaverage rate of cell displacement (Table 1).
HGF Alters the Actin Cytoskeleton. Next we performed immun-ofluorescent staining of the actin cytoskeleton to determine its rela-tionship to the results of the migration assays. Immunofluorescentimages from Alexa phalloidin, which has a high affinity for the actincytoskeleton, revealed striking differences between the HGF-treatedand untreated RMS cells. In the absence of HGF, RMS cells displayedwell-developed filaments of F-actin arranged in parallel to the longaxis of the cell (Fig. 4, A, C, and E), but in its presence they showedmotility-related redistribution of the actin cytoskeleton toward theleading edge of the cell (Fig. 4, B, D, and F).
HGF Increases Migration through Fibronectin-covered Tran-swell Membranes. Next we investigated directed migration of RMScells through transmembranes covered with fibronectin. We selectedfor this study cell lines that had a robust response to HGF stimulationby phosphorylation of signal transduction proteins.
We found that HGF significantly increased the chemotactic activityof the ARMS cell lines RH18, RH28, CW9019, and RH30 and theERMS cell lines SMS-CTR and RD (Fig. 5). Interestingly, RH18cells, which showed the highest increase in locomotion on plasticdishes in the presence of HGF, had the weakest directional chemotaxisto HGF, especially through fibronectin-covered membranes. In con-trast, RH 28 cells, which showed weak locomotion in response toHGF, responded much more strongly to directional chemotaxisthrough fibronectin-covered membranes. RH30 cells that had highspontaneous motility showed the highest directional chemotaxis toHGF.
Chemotaxis of RMS Cells Is PI3K-AKT Axis Dependent. Toelucidate the molecular basis of RMS chemotaxis, we perturbed PI-3Kin RH30 cells by using the specific inhibitor of PI-3K, Ly290042.Cells were preincubated for 30 min with Ly290042 (30 �M) andsubsequently used for signal transduction and chemotaxis studies. Asexpected, Ly290042 inhibited SDF-1- and HGF-dependent phospho-rylation of AKT but not MAPK p42/44 (Fig. 6A). More importantly,we found that inhibition of PI-3K totally inhibits the chemotaxis ofRH30 cells (Fig. 6B).
Fig. 2. HGF signals in RMS cell lines. Phosphorylation of MAPK p42/44, AKT, andSTAT-6 in selected human RMS cell lines stimulated by HGF (10 ng/ml for 10 min). Theexperiment was repeated twice with similar results. A representative study is shown.
Fig. 1. Expression of c-MET on human RMS cell lines. Flow cytometry was performedfor c-MET; open curves, isotype controls. The experiment was repeated three times withsimilar results. A representative study is shown.
7929
C-MET-HGF AND CXCR4-SDF-1 AXES IN RHABDOMYOSARCOMAS
Research. on December 5, 2014. © 2003 American Association for Cancercancerres.aacrjournals.org Downloaded from
Of note, chemotaxis of RH30 cells to SDF-1, but not to HGF, wasalso inhibited when RH30 cells were pretreated before chemotaxiswith the small molecular inhibitor of CXCR4, AMD3100, (25 �g/ml).In control experiments, AMD3100 did not affect either the HGF-dependent activation of AKT or the HGF-dependent chemotaxis (Fig.6, A and B).
In a similar set of experiments, we observed that HGF-dependentchemotaxis as well as HGF-mediated phosphorylation of AKT wasinhibited in the RH30 ARMS cell line by the ATP analogue K562a,which is a potent inhibitor of the c-MET receptor (38).
HGF Increases Adhesion to HUVEC. We found that HGF af-fected the adhesion to HUVEC cells of three ARMS cell lines inves-
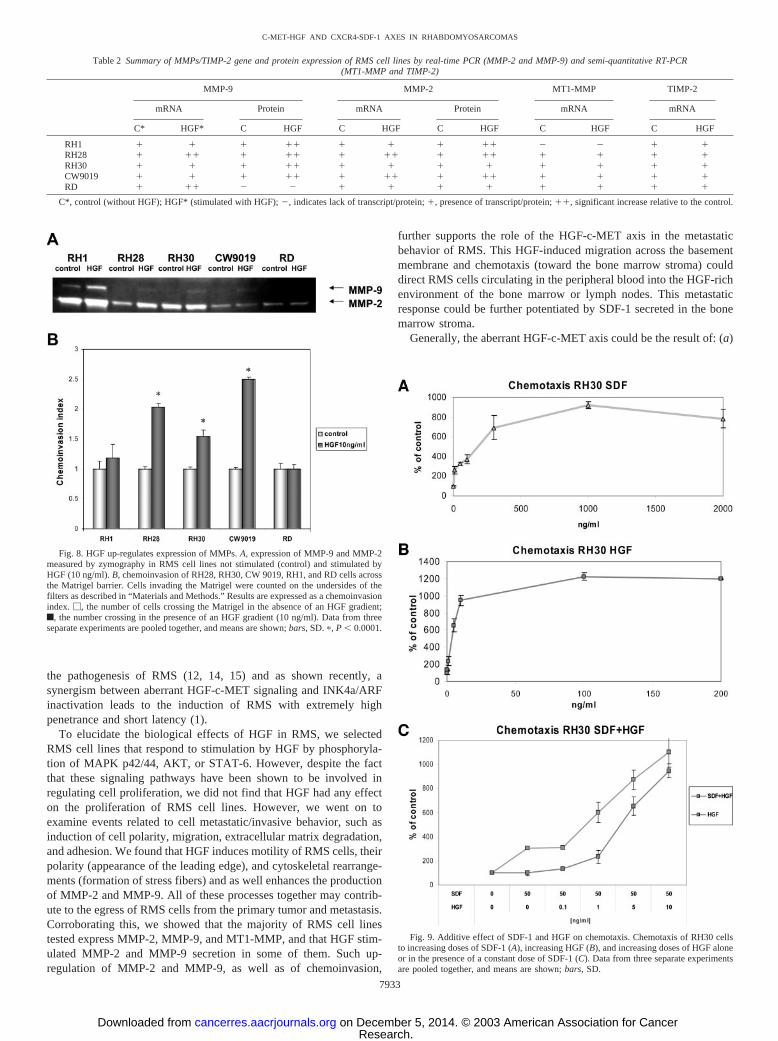
tigated (RH28, RH30, and CW9019) and one ERMS cell line (SMS-CTR; Fig. 7). Generally, the effect on ERMS cells was smaller, and asdemonstrated in Fig. 7B, HGF did not affect adhesion of RD cells.However, when we investigated whether HGF regulates expression/activation of integrins on human RMS using FACS analysis, we didnot find any change in the level of expression of VLA-4, VLA-5,PECAM-1, or ICAM-1 on RMS cells after incubation with HGF for24 h (data not shown).
HGF Stimulates MMP-2 and MMP-9 Secretion and Chemoin-vasion in Selected RMS Cell Lines. In these experiments, we eval-uated whether RMS cells express MMP-2 and/or MMP-9 and whetherHGF stimulates their expression and/or has any effect on expressionof MT1-MMP and TIMP-2, known to be important in MMP-2 acti-vation. We found transcripts for MMP-2 and MMP-9 as well asMT1-MMP and TIMP-2 in all of the RMS cell lines tested (Table 2),with the exception of MT1-MMP in RH-1 cells. After HGF stimula-tion, pro-MMP-9 activity increased in RH1, RH28, RH30, andCW9019 cells (as measured by zymography). Pro-MMP-2 activityincreased in RH1, RH28, and CW9019 but not in the other cell linestested (Fig. 8A and Table 2). However, we found that HGF stimulationdid not influence MT1-MMP and TIMP-2 expression in any of theRMS lines tested (Table 2).
Because tumor cell invasion is characteristic of highly metastaticcells, we evaluated this feature using a Matrigel chemoinvasionassay. We found that the invasive capability of RH28, RH30, andCW9019 cell lines increases significantly in the presence of HGF(Fig. 8B).
HGF and SDF-1 Increase Chemotaxis of RMS Cells in anAdditive Manner. In our previous work (11), we reported thatSDF-1, as we have shown here for HGF, regulated the metastaticbehavior of RMS cells by increasing cell locomotion, directionalchemotaxis, adhesion, and production of MMPs. Hence we decided toinvestigate whether both factors regulate metastatic behavior in anadditive or a synergistic manner. To address this issue, we exposedRH30 ARMS cell lines that show strong chemotaxis to HGF (Fig. 5)and SDF-1 alone (11) to increasing doses of HGF (0–10 ng/ml) in thepresence of a constant suboptimal dose (50 ng/ml) of SDF-1 (Fig. 9,A and B). On the basis of these data, we selected a dose of SDF-1 (50ng/ml) that alone stimulated 30% of the maximal chemotacticresponse to SDF-1 (Fig. 9A). To this concentration of SDF-1 weadded incremental doses of HGF (0–10 ng/ml). As shown in Fig. 9B,10 ng/ml of HGF induces almost maximal chemotactic response bythese cells.
Fig. 9C summarizes the chemotactic response of RH30 cells toincremental doses of HGF in the absence or presence of 50 ng/ml ofSDF-1. We observed that both factors increased the chemotacticactivity of RH30 cells in an incremental manner.
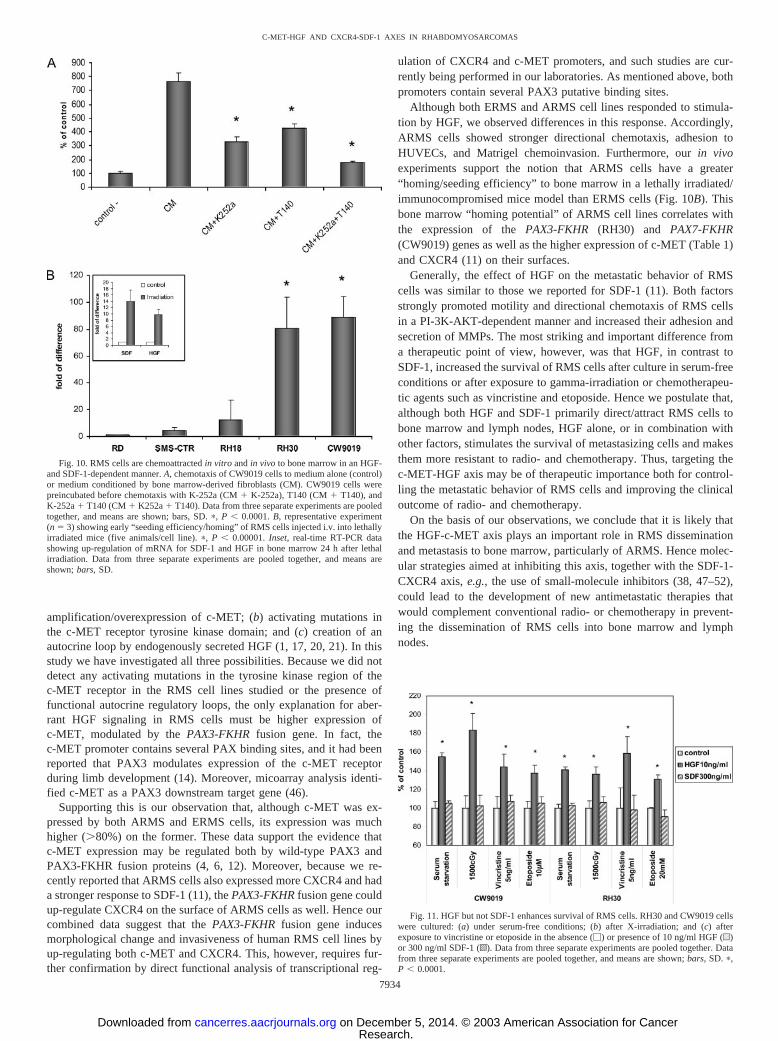
RMS Cells Are Chemoattracted into Bone Marrow in Vitro andin Vivo in an HGF- and SDF-1-dependent Manner. ARMS cellsmetastasize frequently to the bone marrow, and in fact we found thatCM harvested from bone marrow-derived fibroblasts, which are asource for example of HGF (21, 39) and SDF-1 (22–26, 39), che-moattracted the ARMS cell line CW9019 (Fig. 10A). Thus, we usedK-252a and T140, which are specific blocking agents for c-MET andCXCR4, respectively, to determine whether these molecules inhibitthe chemotactic responses of RMS cells to CM harvested from bonemarrow-derived fibroblasts. Fig. 10A also shows that the chemoat-traction of CW9019 cells to CM is significantly inhibited by thespecific blocking agents K-252a and T140. These experiments suggestthat HGF and SDF-1 are major stroma-derived factors that attractRMS cells.
To explore this issue further, we injected lethally irradiatedBALB/c mice with three human ERMS (RD, SMS-CTR, and RH18)
Fig. 3. HGF induces locomotion of RMS cells. Trajectories of 100 each of CW90191,RH28, RH30, RH18, and SMS-CTR cells locomoting in RPMI 1640 (left panels, controlconditions) and in the presence of HGF (10 ng/ml; right panels) displayed in circulardiagrams drawn with the initial point of each trajectory placed at the origin of the plot. Theimages were recorded for 200 min, and at the 5-min time intervals, the positions of cellcentroids were determined.
7930
C-MET-HGF AND CXCR4-SDF-1 AXES IN RHABDOMYOSARCOMAS
Research. on December 5, 2014. © 2003 American Association for Cancercancerres.aacrjournals.org Downloaded from

as well as the two ARMS (RH30 and CW9019) cell lines. BothARMS cells lines have the PAX3-FKHR (RH30) and PAX7-FKHR(CW9019) translocations and express more c-MET and CXCR4 re-ceptors than the PAX-FKHR-negative ERMS cell lines (Fig. 1). Thir-ty-six h after injection into lethally irradiated/immunosuppressedmice, we found that more ARMS than ERMS cells were present in thebone marrow cavities as evaluated by detection of human specific�-satellite DNA using real-time PCR. This assay suggests that bothHGF and SDF-1, which are up-regulated in irradiated bone marrowstroma (Fig. 10A, inset), strongly chemoattracted ARMS cells circu-lating in the peripheral blood in vivo.
HGF but not SDF-1 Increases Survival of RMS Cells. Activa-tion of the tyrosine kinase receptor c-MET, in contrast to activation ofG-protein-coupled CXCR4, influences cell survival in several celllines (17, 19, 21). Thus, to find potential differences between HGFand SDF-1, we evaluated the effects of both factors on the survival ofRMS cells. Two ARMS cell lines, RH30 and CW9019, were exposedto HGF or SDF-1 in three experimental models of cell stress that lead
Fig. 4. Stress fiber formation in human RMS cells. Upper panel, RH18 cells; middlepanel, CW9019 cells; lower panel, RH30 cells. Left half of the figure, cells not exposedto HGF; right half of the figure, cells exposed to HGF (10 ng/ml). Representative cellswere selected.
Fig. 5. Chemotaxis of RMS cells across Transwell membranes covered with fibronec-tin. A, ARMS cell lines RH28, CW9019, and RH30. B, ERMS cell lines SMS-CTR, RD,and RH18. �, chemotaxis to control medium (no HGF in lower chamber); f, chemotaxisto HGF (10 ng/ml). Data from four separate experiments are pooled together. �,P � 0.0001.
Table 1 Parameters characterizing the movement of CW9019, RH28, and RH30 ARMS and RH18 and SMS-CTR ERMS cells in control medium (�) and in the presence ofHGF (HGF)
Cell line TLcT AScM TLcD AvcD CME
CW919(�) 106.78 � 1.91 0.53 � 0.01 32.46 � 2.00 0.16 � 0.01 0.31 � 0.02HGF 146.93 � 4.46a 0.73 � 0.02a 54.47 � 3.36 0.27 � 0.02 0.32 � 0.02
RH28(�) 91.40 � 2.45 0.46 � 0.01 14.09 � 1.34 0.07 � 0.01 0.15 � 0.01HGF 114.65 � 2.74a 0.57 � 0.01a 22.44 � 2.32a 0.11 � 0.02a 0.20 � 0.02a
RH30(�) 110.53 � 6.22 0.55 � 0.03 37.23 � 2.61 0.19 � 0.01 0.37 � 0.02HGF 184.61 � 4.94a 0.92 � 0.02a 79.75 � 5.24 0.40 � 0.03 0.44 � 0.03
RH18(�) 78.27 � 1.93 0.39 � 0.01 13.60 � 1.16 0.07 � 0.01 0.17 � 0.01HGF 157.66 � 5.22a 0.79 � 0.03a 68.82 � 5.31a 0.34 � 0.03a 0.44 � 0.03a
SMS-CTR(�) 110.54 � 6.12 0.55 � 0.03 37.70 � 2.56 0.19 � 0.01 0.37 � 0.02HGF 144.34 � 3.96a 0.72 � 0.02a 42.72 � 2.92 0.21 � 0.01 0.29 � 0.02
Note: Values are given as the mean � SE. TLcT, Total length of cell trajectory (�m); AscM, Average speed of cell movement (�m/min.) � the average speed of cell; TLcD, Totallength of cell displacement (�m); AvcD, Average velocity of cell displacement (�m/min) � the average velocity of cell displacement is defined as length of final cell displacement/timeof recording; CME, Coefficient of movement efficiency � the ratio of cell displacement to cell trajectory length. CME would equal 1 for cells moving persistently along one straightline in one direction and 0 for random movement.
a Statistically significant at P � 0.05.
7931
C-MET-HGF AND CXCR4-SDF-1 AXES IN RHABDOMYOSARCOMAS
Research. on December 5, 2014. © 2003 American Association for Cancercancerres.aacrjournals.org Downloaded from
to cell apoptosis (Fig. 11) such as: (a) culture under serum starvation(72 h); (b) �-irradiation by 1500 cGy; and (c) exposure to vincristine(5 ng/ml) and etoposide (10 �M). We found that HGF, but not SDF-1,enhances the survival of both CW9019 and RH30 cells (P � 0.001).Thus, although both HGF and SDF-1 regulate the metastatic behaviorof RMS cells, only HGF increases their survival (Fig. 11).
Sequencing of the c-MET Receptor. Perturbation of signalingfrom the c-MET receptor may be caused by the c-MET activationmutations that have been described for various human tumors (17, 20,21). To address whether activating mutations of c-MET receptor couldlead to the hyperreactivity of the HGF-c-MET axis in RMS in additionto overexpression of c-MET, we sequenced c-MET DNA from severalRMS cell lines (RH18, RH30, RH28, CW9019, SMS-CTR, and RD)and looked for potential activating mutations. The sequencing wasperformed on exons 15–19 encoding the tyrosine kinase domain. Nomutations were found.
DISCUSSION
Bone marrow involvement at diagnosis of RMS in children andadolescents is a poor prognostic sign and represents a continuingchallenge to current treatment modalities. Accumulated clinical evi-dence suggests that the alveolar subtype of RMS, ARMS, is moreaggressive than the embryonal subtype, ERMS. Almost all of thereported cases associated with marrow involvement, including thosepresenting as possible acute leukemia, have been of the ARMS type(3). Because patients with RMS frequently undergo treatment withhigh-dose chemotherapy combined with autologous bone marrow/peripheral blood stem cell transplantation, the contamination of thetransplanted grafts with tumor cells may contribute to poor survivalrates (40). Hence there is an urgent need to develop more effectivetherapies; elucidating the mechanisms that control the metastaticpotential of RMS cells could be essential to achieving this.
In this work we have hypothesized that RMS cells that express bothc-MET (1, 12) and CXCR4 (11) could be chemoattracted to the bonemarrow by the chemoattractants that are secreted by bone marrowstroma fibroblasts and osteoblasts, such as HGF and SDF-1 (21–26).Supporting this is our finding that CM harvested from human bonemarrow derived-fibroblasts chemoattract ARMS cells in an HGF- andSDF-1-dependent manner (Fig. 10A). Furthermore, CXCR4 had beenimplicated recently in the metastasis of several solid tumors (39,41–45). The role of the CXCR4-SDF-1 axis in the metastasis of RMScells was the subject of our previous study (11), and here we havefocused on the HGF-c-MET axis. The involvement of the HGF-c-MET axis is also known to play an important role in the metastasis/progression of various tumors such as breast, melanoma, and hepa-toma (17, 20, 21). This axis has also been postulated to play a role in
Fig. 6. PI-3K-AKT axis regulates chemotaxis of RMS cells. A, Western blot to detectphospho-AKT and phospho-MAPK p42/44 in RH30 cells pretreated with Ly 290042 (LY;PI-3K inhibitor) or AMD3100 (CXCR4 antagonist). B, chemotaxis of RH30 cells to HGFand SDF-1 after pretreatment with Ly290042 or AMD3100. Data from three separateexperiments are pooled together. �, P � 0.0001 as compared to control; ��, P � 0.0001as compared to SDF-1; ���, P � 0.0001 as compared to HGF or HGF AMD3100. C,influence of pretreatment of RH30 cells with K562a on HGF-dependent chemotaxis. Datafrom two separate experiments are pooled together. �, P � 0.0001 as compared to control.D, influence of pretreatment of RH30 cells with K562a on HGF-induced phosphorylationof AKT. Experiment was repeated twice with similar results.
Fig. 7. Adhesion of human RMS cells to HUVEC cells. A, ARMS cell lines RH28,RH30, and CW9019. B, ERMS cell lines SMS-CTR and RD. Data from four separateexperiments are pooled together. �, P � 0.0001.
7932
C-MET-HGF AND CXCR4-SDF-1 AXES IN RHABDOMYOSARCOMAS
Research. on December 5, 2014. © 2003 American Association for Cancercancerres.aacrjournals.org Downloaded from
the pathogenesis of RMS (12, 14, 15) and as shown recently, asynergism between aberrant HGF-c-MET signaling and INK4a/ARFinactivation leads to the induction of RMS with extremely highpenetrance and short latency (1).
To elucidate the biological effects of HGF in RMS, we selectedRMS cell lines that respond to stimulation by HGF by phosphoryla-tion of MAPK p42/44, AKT, or STAT-6. However, despite the factthat these signaling pathways have been shown to be involved inregulating cell proliferation, we did not find that HGF had any effecton the proliferation of RMS cell lines. However, we went on toexamine events related to cell metastatic/invasive behavior, such asinduction of cell polarity, migration, extracellular matrix degradation,and adhesion. We found that HGF induces motility of RMS cells, theirpolarity (appearance of the leading edge), and cytoskeletal rearrange-ments (formation of stress fibers) and as well enhances the productionof MMP-2 and MMP-9. All of these processes together may contrib-ute to the egress of RMS cells from the primary tumor and metastasis.Corroborating this, we showed that the majority of RMS cell linestested express MMP-2, MMP-9, and MT1-MMP, and that HGF stim-ulated MMP-2 and MMP-9 secretion in some of them. Such up-regulation of MMP-2 and MMP-9, as well as of chemoinvasion,
further supports the role of the HGF-c-MET axis in the metastaticbehavior of RMS. This HGF-induced migration across the basementmembrane and chemotaxis (toward the bone marrow stroma) coulddirect RMS cells circulating in the peripheral blood into the HGF-richenvironment of the bone marrow or lymph nodes. This metastaticresponse could be further potentiated by SDF-1 secreted in the bonemarrow stroma.
Generally, the aberrant HGF-c-MET axis could be the result of: (a)
Fig. 8. HGF up-regulates expression of MMPs. A, expression of MMP-9 and MMP-2measured by zymography in RMS cell lines not stimulated (control) and stimulated byHGF (10 ng/ml). B, chemoinvasion of RH28, RH30, CW 9019, RH1, and RD cells acrossthe Matrigel barrier. Cells invading the Matrigel were counted on the undersides of thefilters as described in “Materials and Methods.” Results are expressed as a chemoinvasionindex. �, the number of cells crossing the Matrigel in the absence of an HGF gradient;f, the number crossing in the presence of an HGF gradient (10 ng/ml). Data from threeseparate experiments are pooled together, and means are shown; bars, SD. �, P � 0.0001.
Fig. 9. Additive effect of SDF-1 and HGF on chemotaxis. Chemotaxis of RH30 cellsto increasing doses of SDF-1 (A), increasing HGF (B), and increasing doses of HGF aloneor in the presence of a constant dose of SDF-1 (C). Data from three separate experimentsare pooled together, and means are shown; bars, SD.
Table 2 Summary of MMPs/TIMP-2 gene and protein expression of RMS cell lines by real-time PCR (MMP-2 and MMP-9) and semi-quantitative RT-PCR(MT1-MMP and TIMP-2)
MMP-9 MMP-2 MT1-MMP TIMP-2
mRNA Protein mRNA Protein mRNA mRNA
C* HGF* C HGF C HGF C HGF C HGF C HGF
RH1 � � RH28 RH30 CW9019 RD � �
C*, control (without HGF); HGF* (stimulated with HGF); �, indicates lack of transcript/protein; , presence of transcript/protein; , significant increase relative to the control.
7933
C-MET-HGF AND CXCR4-SDF-1 AXES IN RHABDOMYOSARCOMAS
Research. on December 5, 2014. © 2003 American Association for Cancercancerres.aacrjournals.org Downloaded from
amplification/overexpression of c-MET; (b) activating mutations inthe c-MET receptor tyrosine kinase domain; and (c) creation of anautocrine loop by endogenously secreted HGF (1, 17, 20, 21). In thisstudy we have investigated all three possibilities. Because we did notdetect any activating mutations in the tyrosine kinase region of thec-MET receptor in the RMS cell lines studied or the presence offunctional autocrine regulatory loops, the only explanation for aber-rant HGF signaling in RMS cells must be higher expression ofc-MET, modulated by the PAX3-FKHR fusion gene. In fact, thec-MET promoter contains several PAX binding sites, and it had beenreported that PAX3 modulates expression of the c-MET receptorduring limb development (14). Moreover, micoarray analysis identi-fied c-MET as a PAX3 downstream target gene (46).
Supporting this is our observation that, although c-MET was ex-pressed by both ARMS and ERMS cells, its expression was muchhigher (�80%) on the former. These data support the evidence thatc-MET expression may be regulated both by wild-type PAX3 andPAX3-FKHR fusion proteins (4, 6, 12). Moreover, because we re-cently reported that ARMS cells also expressed more CXCR4 and hada stronger response to SDF-1 (11), the PAX3-FKHR fusion gene couldup-regulate CXCR4 on the surface of ARMS cells as well. Hence ourcombined data suggest that the PAX3-FKHR fusion gene inducesmorphological change and invasiveness of human RMS cell lines byup-regulating both c-MET and CXCR4. This, however, requires fur-ther confirmation by direct functional analysis of transcriptional reg-
ulation of CXCR4 and c-MET promoters, and such studies are cur-rently being performed in our laboratories. As mentioned above, bothpromoters contain several PAX3 putative binding sites.
Although both ERMS and ARMS cell lines responded to stimula-tion by HGF, we observed differences in this response. Accordingly,ARMS cells showed stronger directional chemotaxis, adhesion toHUVECs, and Matrigel chemoinvasion. Furthermore, our in vivoexperiments support the notion that ARMS cells have a greater“homing/seeding efficiency” to bone marrow in a lethally irradiated/immunocompromised mice model than ERMS cells (Fig. 10B). Thisbone marrow “homing potential” of ARMS cell lines correlates withthe expression of the PAX3-FKHR (RH30) and PAX7-FKHR(CW9019) genes as well as the higher expression of c-MET (Table 1)and CXCR4 (11) on their surfaces.
Generally, the effect of HGF on the metastatic behavior of RMScells was similar to those we reported for SDF-1 (11). Both factorsstrongly promoted motility and directional chemotaxis of RMS cellsin a PI-3K-AKT-dependent manner and increased their adhesion andsecretion of MMPs. The most striking and important difference froma therapeutic point of view, however, was that HGF, in contrast toSDF-1, increased the survival of RMS cells after culture in serum-freeconditions or after exposure to gamma-irradiation or chemotherapeu-tic agents such as vincristine and etoposide. Hence we postulate that,although both HGF and SDF-1 primarily direct/attract RMS cells tobone marrow and lymph nodes, HGF alone, or in combination withother factors, stimulates the survival of metastasizing cells and makesthem more resistant to radio- and chemotherapy. Thus, targeting thec-MET-HGF axis may be of therapeutic importance both for control-ling the metastatic behavior of RMS cells and improving the clinicaloutcome of radio- and chemotherapy.
On the basis of our observations, we conclude that it is likely thatthe HGF-c-MET axis plays an important role in RMS disseminationand metastasis to bone marrow, particularly of ARMS. Hence molec-ular strategies aimed at inhibiting this axis, together with the SDF-1-CXCR4 axis, e.g., the use of small-molecule inhibitors (38, 47–52),could lead to the development of new antimetastatic therapies thatwould complement conventional radio- or chemotherapy in prevent-ing the dissemination of RMS cells into bone marrow and lymphnodes.
Fig. 11. HGF but not SDF-1 enhances survival of RMS cells. RH30 and CW9019 cellswere cultured: (a) under serum-free conditions; (b) after X-irradiation; and (c) afterexposure to vincristine or etoposide in the absence (�) or presence of 10 ng/ml HGF (u)or 300 ng/ml SDF-1 (o). Data from three separate experiments are pooled together. Datafrom three separate experiments are pooled together, and means are shown; bars, SD. �,P � 0.0001.
Fig. 10. RMS cells are chemoattracted in vitro and in vivo to bone marrow in an HGF-and SDF-1-dependent manner. A, chemotaxis of CW9019 cells to medium alone (control)or medium conditioned by bone marrow-derived fibroblasts (CM). CW9019 cells werepreincubated before chemotaxis with K-252a (CM K-252a), T140 (CM T140), andK-252a T140 (CM K252a T140). Data from three separate experiments are pooledtogether, and means are shown; bars, SD. �, P � 0.0001. B, representative experiment(n � 3) showing early “seeding efficiency/homing” of RMS cells injected i.v. into lethallyirradiated mice (five animals/cell line). �, P � 0.00001. Inset, real-time RT-PCR datashowing up-regulation of mRNA for SDF-1 and HGF in bone marrow 24 h after lethalirradiation. Data from three separate experiments are pooled together, and means areshown; bars, SD.
7934
C-MET-HGF AND CXCR4-SDF-1 AXES IN RHABDOMYOSARCOMAS
Research. on December 5, 2014. © 2003 American Association for Cancercancerres.aacrjournals.org Downloaded from

REFERENCES
1. Sharp, R., Recio, J. A., Jhappan, C., Otsuka, T., Liu, S., Yu, Y., Liu, W., Anver, M.,Navid, F., Helman, L. J. DePinho, R. A., and Merlino, G. Synergism betweenINK4a/ARF inactivation and aberrant HGF signaling in rhabdomyosarcomagenesis.Nat. Med., 8: 1276–1280, 2002.
2. Ruymann, F. B., Newton, W. A., Ragab, A. H., Donaldson, M. H., and Foulkes, M.Bone marrow metastases at diagnosis in children and adolescents with rhabdomyo-sarcoma. Cancer (Phila.), 53: 368–373, 1984.
3. Sandberg, A. A., Stone, J. F., Czarnecki, L., and Cohen, J. D. Hematologic masquer-ade of rhabdomyosarcoma. Am. J. Hematol., 68: 51–57, 2001.
4. Barr, F. G., Galili, N., Holick, J., Biegel, J. A., Rovera, G., and Emanuel, B. S.Rearrangement of the PAX3 paired box gene in the paediatric solid tumour alveolarrhabdomyosarcoma. Nat. Genet., 3: 113–117, 1993.
5. Hazelton, B. J., Houghton, J. A., Parham, D. M., et al. Characterization of cell linesderived from xenografts of childhood rhabdomyosarcoma. Cancer Res., 47: 4501–4507, 1987.
6. Davis, R. J., and Barr, F. G. Fusion genes resulting from alternative chromosomaltranslocations are overexpressed by gene-specific mechanisms in alveolar rhabdomy-osarcoma. Proc. Natl. Acad. Sci. USA, 94: 8047–8051, 1997.
7. Davis, R. J., D’Cruz, C. M., Lovell, M. A., Biegel, J. A., and Barr, F. G. Fusion ofPAX7 to FKHR by the variant t(1;13)(p36;q14) translocation in alveolar rhabdomy-osarcoma. Cancer Res., 54: 2869–2872, 1994.
8. Collins, M. H., Zhao, H., Womer, R. B., and Barr, F. G. Proliferative and apoptoticdifferences between alveolar rhabdomyosarcoma subtypes: a comparative study oftumors containing PAX3-FKHR gene fusions. Med. Pediatr. Oncol., 37: 83–89,2000.
9. Bennicelli, J. L., Advani, S., Schafer, B. W., and Barr, F. G. PAX3 and PAX7 exhibitconserved cis-acting transcription repression domains and utilize a common gain offunction mechanism in alveolar rhabdomyosarcoma. Oncogene, 18: 4348–4356,1999.
10. Kelly, K. M., Womer, R. B., and Barr, F. 3-FKHR and PAX7-FKHR fusions inrhabdomyosarcoma. J. Pediatr. Hematol. Oncol., 20: 517–518, 1998.
11. Libura, J., Drukala, J., Majka, M., et al. CXCR4-SDF-1 signaling is active inrhabdomyosarcoma cells and regulates locomotion, chemotaxis, and adhesion. Blood,100: 2597–2606, 2002.
12. Ginsberg, J. P., Davis, R. J., Bennicelli, J. L., Nauta, L. E., and Barr, F. G.Up-regulation of MET but not neural cell adhesion molecule expression by thePAX3-FKHR protein in alveolar rhabdomyosarcoma. Cancer Res., 58: 3542–3546,1998.
13. Anderson, J., Ramsay, A., Gould, S., Pritchard-Jones K. PAX3-FKHR inducesmorphological change and enhances cellular proliferation and invasion in rhabdomy-osarcoma. Am. J. Pathol., 159: 1089–1096, 2001.
14. Epstein, J. A., Shapiro, D. N., Cheng, J., Lam, P. Y. P., and Maas, R. L. Pax3modulates expression of the c-Met receptor during limb muscle development. Proc.Natl. Acad. Sci. USA, 93: 4213–4218, 1996.
15. Ferracini, R., Olivero, M. DiRenzo, M. F., Martano, M., DeGiovanni, C., Nanni, P.,Basso, G., Scotlandi, K., Lollini, P. L., and Comoglio, P. M. Retrogenic expressionof the MET proto-oncogene correlates with the invasive phenotype of human rhab-domyosarcoma. Oncogene, 12: 1697–1705, 1996.
16. Corti, S., Salani, S., Del Bo, R., et al. Chemotactic factors enhance myogenic cellmigration across an endothelial monolayer. Exp. Cell Res., 268: 36–44, 2001.
17. Danilkovitch-Miagkova, A., and Zbar, B. Dysregulation of Met receptor tyrosinekinase activity in invasive tumors. J. Clin. Invest., 109: 863–867, 2002.
18. Comoglio, P. M., and Trusolino, L. Invasive growth: from development to metastasis.J. Clin. Invest., 109: 857–862, 2002.
19. Goshima, Y., Ito, T., Sasaki, Y., Nakamura, F. Semaphorins as signals for cellrepulsion and invasion. J. Clin. Invest., 109: 993–998, 2002.
20. Mualik, G., Shrikhande, A., Kijima, T., et al. Role of hepatocyte growth factorreceptor, c-Met, in oncogenesis and potential therapeutic inhibition. Cytokine GrowthFactor Rev., 13: 41–59, 2002.
21. Tamagnone, L., and Comoglio, P. M. Control of invasive growth by hepatocytegrowth factor (HGF) and related scatter factors. Cytokine Growth Factor Rev., 8:129–142, 1997.
22. Zou, Y., Kottmann, A. H., Kuroda, M., Taniuchi, I., and Littman, D. R. Function ofthe chemokine receptor CXCR4 in haematopoiesis and in cerebellar development.Nature (Lond.), 393: 595–599, 1998.
23. Aiuti, A., Webb, I. J., Bleul, C., Springer, T., and Gutierrez-Ramos, J. C. Thechemokine SDF-1 is a chemoattractant for human CD34 hematopoietic progenitorcells and provides a new mechanism to explain the mobilization of CD34 progen-itors to peripheral blood. J. Exp. Med., 185: 111–120, 1997.
24. Mohle, R., Bautz, F., Rafii, S., Moore, M. A., Brugger, W., and Kanz, L. Thechemokine receptor CXCR4 is expressed on CD34 hematopoietic progenitors andleukemic cells and mediates transendothelial migration induced by stromal cell-derived factor-1. Blood, 91: 4523–4530, 1998.
25. Peled, A., Grabovsky, V., Habler, L., et al. The chemokine SDF-1 stimulatesintegrin-mediated arrest of CD34 cells on vascular endothelium under shear flow.J. Clin. Invest., 104: 1199–1211, 1997.
26. Wang, J-F., Park, I-W., and Groopman, J. E. Stromal cell-derived factor-1� stimu-lates tyrosine phosphorylation of multiple focal adhesion proteins and induces mi-
gration of hematopoietic progenitor cells: roles of phosphoinositide-3 kinase andprotein kinase C. Blood, 95: 2505–2513, 2000.
27. Ratajczak, M. Z., Marlicz, W., Ratajczak, J., Wasik, M., Machalinski, B., Carter, A.,and Gewirtz, A. M. Effect of hepatocyte growth factor on early human haemopoieticcell development. Br. J. Haematol., 99: 229–236, 1997.
28. Kijowski, J., Baj, M., Majka, M., et al. The SDF-1-CXCR4 axis stimulates VEGFsecretion and activates integrins but does not affect proliferation and survival inlymphohematopoietic cells. Stem Cells, 19: 453–466, 2001.
29. Majka, M., Janowska-Wieczorek, A., Ratajczak, J., et al. Stromal derived factor-1 andthrombopoietin regulate distinct aspects of human megakaryopoiesis. Blood, 96:4142–4151, 2000.
30. Majka, M., Ratajczak, J., Lee, B., et al. The role of HIV related chemokine receptorsand chemokines in human erythropoiesis in vitro. Stem Cells, 18: 128–138, 2000.
31. Majka, M., Janowska-Wieczorek, A., Ratajczak, J., et al. Numerous growth factors,cytokines and chemokines are secreted by human CD34 cells, myeloblasts, eryth-roblasts and megakaryoblasts and regulate normal hematopoiesis in an autocrine/paracrine manner. Blood, 97: 3075–3085, 2001.
32. Madeja, Z., Szymkiewicz, I., Zaczek, A., Sroka, J., Miekus, K., and Korohoda, W.Contact-activated migration of melanoma B16 and sarcoma XC cells. Biochem. CellBiol., 79: 1–16, 2001.
33. Korohoda, W., and Madeja, Z. Contact of sarcoma cells with aligned fibroblastsaccelerates their displacement: computer-assisted analysis of tumour cell locomotionin co-culture. Biochem. Cell Biol., 75: 263–276, 1997.
34. Janowska-Wieczorek, A., Marquez, L. A., Matsuzaki, A., et al. Expression of matrixmetalloproteinases (MMP-2 and -9) and tissue inhibitors of metalloproteinases(TIMP-1 and -2) in acute myelogenous leukaemia blasts: comparison with normalbone marrow cells. Br. J. Haematol., 105: 402–411, 1999.
35. Janiak, M., Hashmi, H. R., and Janowska-Wieczorek, A. Use of the Matrigel-basedassay to measure the invasiveness of leukemic cells. Exp. Hematol., 22: 559–565,1994.
36. Janowska-Wieczorek, A., Marquez, L. A., Nabholtz, J-M., et al. Growth factors andcytokines upregulate gelatinase expression in bone marrow CD34 cells and theirtransmigration through reconstituted basement membrane. Blood, 93: 3379–3390,1999.
37. Livak, K. J., and Schmittgen, T. D. Analysis of relative gene expression data usingreal-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods, 25:402–408, 2001.
38. Morotti, A., Mila, S., Accornero, P., Tagliabue, E., and Ponzetto, C. K.252a inhibitsthe oncogenic properties of Met, the HGF receptor. Oncogene, 21: 4885–4893, 2002.
39. Taichman, R. S., Cooper, C., Keller, E. T., Pienta, K. J., Taichman, N. S., andMcCauley, L. K. Use of the stromal cell-derived factor-1/CXCR4 pathway in prostatecancer metastasis to bone. Cancer Res., 62: 1832–1837, 2002.
40. Weigel, B. J., Breitfeld, P. P., Hawkins, D., Crist, W. M., and Baker, K. S. Role ofhigh-dose chemotherapy with hematopoietic stem cell rescue in the treatment ofmetastatic or recurrent rhabdomyosarcoma. J. Pediatr. Hematol. Oncol., 23: 272–276,2001.
41. Mueller, A., Homey, B., Soto, H., et al. Involvement of chemokine receptors in breastcancer metastasis. Nature (Lond.), 410: 50–56, 2000.
42. Geminder, H., Sagi-Assif, O., Goldberg, L., et al. A possible role for CXCR4 and itsligand, the CXC chemokine stromal cell-derived factor-1, in the development of bonemarrow metastases in neuroblastoma. J. Immunol., 167: 4747–4757, 2001.
43. Bachelder, R. E., Wendt, M. A., and Mercurio, A. M. Vascular endothelial growthfactor promotes breast carcinoma invasion in an autocrine manner by regulating thechemokine receptor CXCR4. Cancer Res., 62: 7203–7206, 2002.
44. Scotton, C. J., Wilson, J., Scott, K., Stamp, G., Wilbanks, G. D., Fricker, S., Bridger,G., and Balkwill, R. Multiple action of the chemokine CXCL12 on epithelial tumorcells in human ovarian cancer. Cancer Res., 62: 5930–5938, 2002.
45. Kijima, T., Maulik, G., Ma, P. C., Tibaldi, E. V., Turner, R. E., Rollins, B., Sattler,M., Johnson, B. E., and Salgia, R. Regulation of cellular proliferation, cytoskeletalfunction and signal transduction through CXCR4 and c-kit in small cell lung cancercells. Cancer Res., 62: 6304–6311, 2002.
46. Mayanil, C. S. K., George, D., Freilich, L., Miljan, E. J., Mania-Farnell, B., McLone,D. G., and Bremer, E. G. Microarray analysis detects novel Pax3 downstream targetgenes. J. Biol. Chem., 276: 49299–49309, 2001.
47. Murphy, P. M. Chemokines and the molecular basis of cancer metastasis. N. Engl.J. Med., 345: 833–835, 2001.
48. Zhang, W. W., Navenot, J. M., Haribabu, B., et al. A point mutation that confersconstitutive activity to CXCR4 reveals T140 is an inverse agonist and AMD3100 andALX40–4C are weak partial agonists. J. Biol. Chem., 277: 24515–24521, 2002.
49. Tamamura, H., Xu, Y., Hattori, T., et al. A low-molecular-weight inhibitor against thechemokine receptor CXCR4: a strong anti-HIV peptide T140. Biochem. Biophys.Res. Commun., 253: 877–882, 1998.
50. Hatse, S., Princen, K., Bridger, G., De Clercq, E., and Schols, D. Chemokine receptorinhibition by AMD3100 is strictly confined to CXCR4. FEBS Lett., 527: 255–262,2002.
51. Webb, C. P., Hose, C. D., Koochekpour, S., et al. The geldanamycins are potentinhibitors of the hepatocyte growth factor/scatter factor Met-urokinase plasminogenactivator plasmin proteolytic network. Cancer Res., 60: 342–349, 2000.
52. Bardelli, A., Longati, P., Williams, T. A., et al. A peptide representing the carboxyl-terminal tail of the Met receptor inhibits kinase activity and invasive growth. J. Biol.Chem., 274: 29274–29281, 1999.
7935
C-MET-HGF AND CXCR4-SDF-1 AXES IN RHABDOMYOSARCOMAS
Research. on December 5, 2014. © 2003 American Association for Cancercancerres.aacrjournals.org Downloaded from

2003;63:7926-7935. Cancer Res Kacper Jankowski, Magda Kucia, Marcin Wysoczynski, et al. Resistance to RadiochemotherapyRhabdomyosarcoma Cells, But Only HGF Enhances TheirFactor-1 Regulate the Metastatic Behavior of Human Both Hepatocyte Growth Factor (HGF) and Stromal-Derived
Updated version
http://cancerres.aacrjournals.org/content/63/22/7926
Access the most recent version of this article at:
Cited Articles
http://cancerres.aacrjournals.org/content/63/22/7926.full.html#ref-list-1
This article cites by 49 articles, 21 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/63/22/7926.full.html#related-urls
This article has been cited by 24 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
SubscriptionsReprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
Research. on December 5, 2014. © 2003 American Association for Cancercancerres.aacrjournals.org Downloaded from
Copyright © 2022 FDOKUMEN


















![t] x| f} + of] hgf -cf=j](https://static.fdokumen.com/doc/165x107/631922cab41f9c8c6e099861/t-x-f-of-hgf-cfj.jpg)

