
Unfolding Various Concepts of Junctional Epithelium - Thieme ...

1
Aus der Klinik und Poliklinik für Anästhesiologie und Intensivmedizin(Direktor: Univ.-Prof. Dr. med. Michael Wendt)
der Medizinischen Fakultätder Ernst – Moritz - Arndt Universität
Greifswald
Thema:Modulation des Tonus der Atemwege
durch das Epithel: Effekte von Carbocistein
Inaugural – Dissertationzur
Erlangung des akademischen Grades Eines
Doktor der Medizin (Dr. med.)
Der Medizinischen FakultätDer Ernst – Moritz - Arndt Universität
Greifswald2002
vorgelegt von:
Dragan Pavlovic04. 08. 1949, Vranje (Jugoslawien)

2
Dekan: Prof. Dr. rer. nat. Heyo K. Kroemer
1. Gutachter: Prof. S. Felix, Greifswald2. Gutachter: Prof . M. Wendt, Greifswald3. Gutachter: Dr. R. Ewert, Charite, Berlin4. Gutachter: Dr. Georges Saumon, Paris
Tag der Disputation: 27. 11 2002.

3
Modulation of the Airway Tone by the Epithelium: Effects ofCarbocisteine
Dragan Pavlovic
The Medical FacultyErnst Moritz Arndt University
Greifswald 2002

4
Short title:Tube/ring tracheal preparation

5
Summary
We improved our previous model of tracheal tube preparation and
examined the effects of oral treatment of rats with carbocisteine (CCS)
and its interaction with bronchial epithelium. The model permitted
isometric or isotonic measurements of smooth muscle contraction or
relaxation in cannulated or tracheal ring preparations, with or without
epithelium. We found that oral treatment with carbocisteine and not
preincubation of preparations in vitro, diminished sensitivity of
preparations without epithelium to carbachol (EC50, -log(M) values:
IN – luminary perfusion, -EP, controls vs. CCS: 5.8±0.06 vs. 5.5±0.09,
p<0.005; OUT - serosal perfusion, -EP, controls vs. CCS: 5.9±0.06
vs. 5.6±0.05, p<0.005), while the sensitivity to aminophylline, degree
of shortening, and the velocity of contraction of rat tracheal rings
stimulated by 10-6M carbachol was not affected. Normal sensitivity to
carbachol stimulation was re-established if preparations were
preincubated with capsaicin. We conclude that carbocisteine has small

6
inhibitory effects on the sensitivity to carbachol of the rat tracheal smooth
muscle denuded of epithelium. Described model is valuable for examining
the effects of bronchial epithelium on bronchial smooth muscle contraction.
Keywords: Smooth muscle models; tracheal epithelium; carbocisteine

7
Introduction
In this study we demonstrated the use of an improved model for testing
bronchial smooth muscle contraction in either tube (cannulated tracheal
segments) or tracheal rings preparations. Various airway tube
preparations (cannulated trachea or bronchus) have been used in the
past in an attempt to better mimic in vivo responsiveness of airways and
dissociate pure smooth muscle responses from airway epithelium
modulated smooth muscle responses (10). That interest is justified since
there is increased evidence that airway epithelium is in close relation with
underlying smooth muscle not only structurally but also functionally.
Indeed, airway epithelium, which separates inhaled gas from underlying
airway tissue, forms an important barrier between living structures and
the environment. The preservation of its normal (secretory) function is
a precondition for maintenance of an effective air - tissue barrier. In
inflammation and particularly in asthma, the mucus gel becomes thicker,
stagnant and could, together with oedema and bronchial smooth muscle

8
spasm, produce narrowing and plugging of the airways, seriously
compromising air flow and ventilation. In addition, it seams that normal
functioning of the bronchial epithelium is important for the maintenance
of normal bronchial reactivity to various bronchoconstricting or relaxing
agents. To further test the hypothesis that bronchial epithelium, either as
a diffusion barrier or by actively secreting yet not well defined mediators
(prostaglandins, leukotrienes or other mediators) could modulate
bronchial smooth muscle sensitivity to various stimulating agents (see
12, 20 and 25 for references) we used our newly adapted system of
perfused tracheal tubes.
The subject of our study was mucolytic agent carbocisteine (S-
carboxymethyl-L-cysteine) (CCS) that was found to favour production
of sialomucins at the expense of fucomucines (5, 11) which could result
in the improvement of mucociliary transport. Improving mucociliary
transport is one of the therapeutic goals in asthma therapy, where in
addition to anti-inflammatory agents (14, 26) mucolytic agents are
frequently used (2, 17). Existing controversy about the mechanisms of
action of carbocisteine (4, 15), finding that it could produce a reduction

9
(6), or an increase in sputum viscosity (24), or even to have an anti-
inflammatory action by promoting sialoglycoprotein secretion which
could have kinin-inhibitory effects (19) - further justified our aim: to
examine the role of bronchial epithelium as a specific target tissue of that
mucolytic agent and its influence on bronchial smooth muscle isometric
(cannulated) or isotonic (rings) contraction. The preliminary results that
were done in the earlier described model (20) already showed that
carbocisteine could influence sensitivity of tracheal smooth muscle of the
rat (3, 23). Particular facility of the proposed model, i.e. the possibility
to easily switch from a tube to a ring preparation, permitted us to
examine selectively effects of inner (epithelial) or outer (serosal) perfusion
with an agent which influences the bronchial smooth muscle tone.

10
Methods
Experiments were performed on tracheas taken from male Sprague -
Dawley rats weighing 390 - 420 g. All animals were housed in individual
cages and received water and food ad libitum. The group of pretreated
animals (CCSpr) received carbocisteine mixed with water (200 mg/kg/
day) for two weeks. During the third week they were sacrificed for the
experiments. They were stunned by a blow on the head and quickly
exsanguinated. The tracheas were immersed in Krebs solution (in mM:
137 NaCl, 4 KCl, 1 MgCl2 x 6H2O, 1 KH2PO4, 12 NaHCO3, 2
CaCl2, 6.5 glucose) and cleaned from surrounding tissue.
Perfusion studies. Proximal tracheal ends (10 tracheal rings long) were
used for the experiments. In one half of the preparations the epithelium
was removed (-EP) by gently rubbing with a cotton - wrapped metal
stick; in the other half of the preparations the epithelium was left intact
(+EP).
Under microscopic control two stainless steel hooks were

11
passed through the tracheal wall around two adjacent cartilagineous
rings as close as possible to the tracheal muscle insertions. The tracheal
segment was then longitudinally connected to steel tubes built in the
“carrier block” of the apparatus (in-out system) and firmly tightened with
silk thread. The apparatus used (Fig. 1A, EMKA Technologies, Paris,
France) was an improved version of the cannulated tracheal system
described previously (20). Improvements consisted of the fact that the
lower hook, which served as a “fixed point”, was attached with the silk
thread to the micrometric screw serving to adjust the hook tension on
the tracheal wall. This allowed precise adjustment of the resting tension.
The upper hook was connected to a force transducer (IT1-25, EMKA
Technologies), the latter being attached to micromanipulator which
permitted displacement of the upper hook along a strict vertical axis.
Any change in tension at the level of the tracheal muscle was registered
by the recorder (Gould AT 550) to which the amplified signal (EMKA
technologies 4 ways Amplifier) from transducer was connected. The
“carrier block” for cannulated trachea could be easily removed and
replaced by one suitable for tracheal rings mounting (Fig. 1B, ring

12
system, below). Described system accommodates for use of electrical
field stimulation, which was not used in this experiment.
The Krebs solution (at 37°C, pH 7.4, gassed with 95% O2/5%
CO2) was perfused at a constant flow rate (2ml/min) through the organ
bath (outer perfusion - OUT) and through the lumen of the tracheal
segment (inner perfusion - IN) by using peristaltic pumps (Watson
Marlow 5025, Falmouth, Cornwall, UK).
Fluid tightness of preparation. Methylene blue test. To ensure that
the hooks did not induce fluid leak through the tracheal wall, in separate
experiments (n = 4) a solution of methylene blue was perfused into the
tracheal lumen or into the bath. No cross - staining was observed. Bio-
assay. In addition to the methylene blue test a bio-assay was performed
(n = 5) in the following way. In the same organ bath in which the
cannulated tracheal segment was mounted a tracheal ring consisting of
two tracheal cartilages was suspended between the stainless steal
hooks, one served as a fix point and the other was attached to the
isometric force transducer. Following a stabilisation period of the one

13
hour the luminary (epithelial side) of the cannulated tracheal segment
was perfused with carbachol 10-3M. The tension of both preparations
(cannulated tracheal segment and the ring preparation) were recorded.
Carbachol 10-3M fully contracted the cannulated tracheal segment
whereas no change in tension was observed in the tracheal ring
preparation. This demonstrated that there was no significant leakage of
solution from the luminary side of the cannulated tracheal segment
towards outside bath solution.
Tracheal rings. To determine the effects of carbocisteine on the degree
of shortening and the velocity of contraction, tracheal ring preparations
was mounted in the customary manner using ring system of the apparatus
connected to the isotonic force transducer (Harvard Apparatus, South
Natik, MA, USA), linked to the paper recorder (GOULD AT 550,
Cleveland, OH). The tracheal muscle was stretched to its optimal length
which was established, in the preliminary experiments, to correspond to
a counter - weight of 1.0g.

14
Procedure.
Perfusion studies. In this experimental model studies were realised on
preparations with and/or without epithelium taken from animals pretreated
with carbocisteine (CCSpr) or from the control preparations (contr).
After a period of stabilisation (45-60 min.) the tracheal muscle
was stretched to its optimal length. Preliminary essays were performed
to determine optimal stretch of the muscle as previously described (22).
The length - tension relationship was not found to differ significantly
between preparations (CCSpr) and the controls (contr).
In the set of preliminary experiments cumulative concentrations
of carbachol (10-7 to 10-3M) were perfused IN (+EP or -EP) or OUT
(+EP or -EP) in (CCSpr) or from the controls (contr) (n = 8 for each).
Since the accent of the work was to test the new improved model, we
restricted control experiments to only those which were mandatory.
Accordingly, we concluded from preliminary experiments that responses
to IN and OUT stimulation in -EP preparations were identical (on the
basis of EC50
and maximal responses), and in order to further examine
observed effects in preparations -EP on (IN) stimulation with carbachol,

15
we designed another 3 sets of experiments.
First, to examine effect of preincubation, preparations taken
from untreated animals were incubated for 60 min. with carbocisteine
10-3M (CCSinc; -EP, n=8) and cumulative concentrations of carbachol
were perfused luminary the preparations (IN).
In the second set of experiments preparations CCSpr (-EP)
were preincubated for 60 minutes in indomethacin 10-6M (INDinc; n
= 8) or capsaicin 10-5M (CAPSinc, n = 6) and cumulative concentrations
of carbachol (IN) were perfused. Effects of carbocisteine on relaxant
effects of aminophylline were examined in preparations CCSpr (-EP)
which were first pre-contracted with medium concentration (10-6M) of
carbachol (OUT) and then perfused with aminophylline (IN) 10-8 to
10-3M (n =10) and compared with responses to aminophylline in
preparations (contr) (n = 6). In the third set of experiments, to examine
direct relaxant effects of carbocisteine, preparations taken from the
control animals were pre-contracted (OUT) with medium concentration
of carbachol (10-6M) and then perfused (IN) with cumulative
concentrations of carbocisteine 10-7 to 10-3M (+EP, n = 7; -EP, n =

16
7).
Tracheal rings. Degree of shortening (maximal shortening) and relative
velocity of contraction (time to 50, 80 and 100% of maximal shortening)
elicited with carbachol of preparations taken from animals pretreated
(n=11) and not pretreated (n=11) with carbocisteine (+ or - EP) were
compared. Relative degree of shortening was used because measuring
the small size change of the rat tracheal muscle required extremely
precise instrumentation not available in the laboratory. After the
equilibration period, the preparations were stimulated with carbachol
10-6M. When maximal shortening of the tracheal muscle was achieved,
they were washed with Krebs solution and allowed to relax completely.
Then, the preparations were preincubated for 60 min. in carbocisteine
(10-3M) and stimulated again with carbachol (10-6M).
Substances. The following substances were used: carbachol
(carbamylcholine chloride, Sigma), indomethacin (Sigma), aminophylline
(theophylline - ethylene diamine, Pharmacie Centrale des Hopitaux,

17
Paris, France), capsaicin (Sigma), and carbocisteine (Park-Davis,
45071 Orleans, France). The carbocisteine was diluted in 10% NaOH
and final dilutions made in Krebs solution.
Analysis of results. The data are expressed in percent of maximal
response and in absolute values (g and second) and given as means ±
s. e. means. Half - maximal concentration (EC50) values were calculated
by means of regression analysis of probit - transformed data and the
results are given as means of -log EC50 values obtained. Statistical
analysis was conducted by use of analysis of variance and the Student‘s
t-test for paired or unpaired data adjusted for multiple comparisons, as
appropriate. p<0.05 was regarded as being statistically significant.

18
Results
The animals pretreated with oral carbocisteine did not show any clinical
sign of disease or metabolic disturbances.
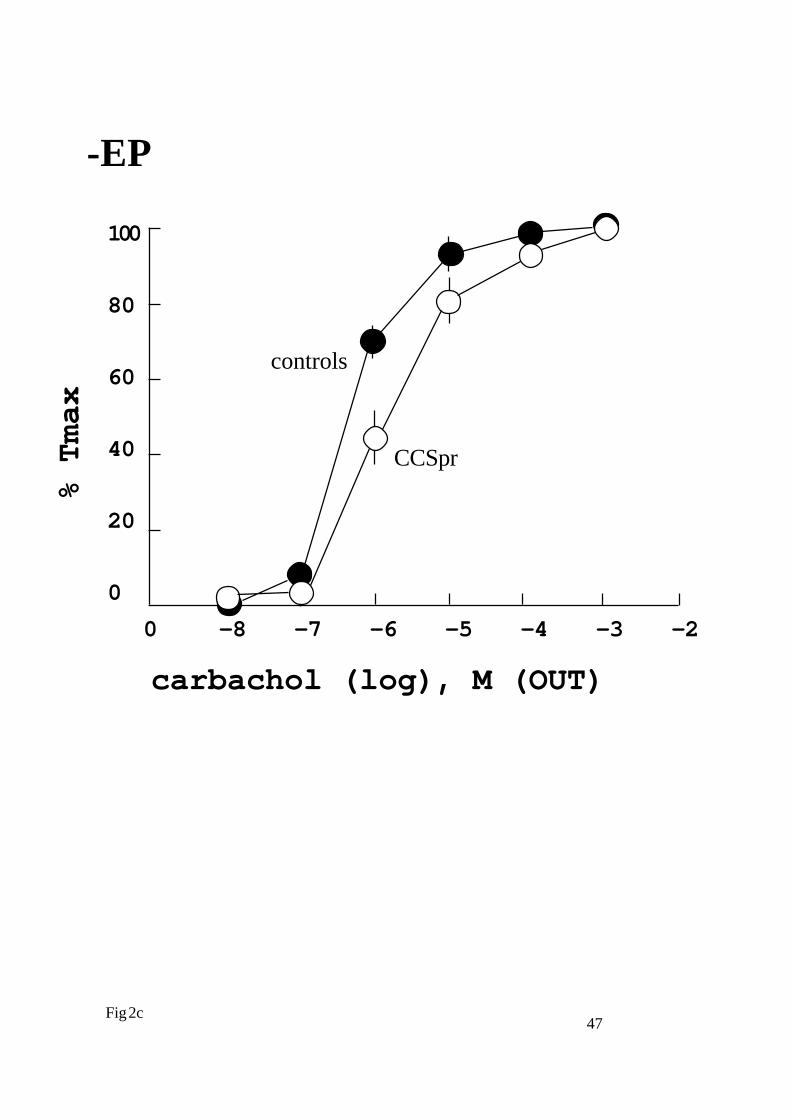
Contractility. In preparations taken from animals pretreated with oral
carbocisteine for 7 days (CCSPr) we found diminished sensitivity to
carbachol in preparations (-EP), but not in preparations (+EP) (Table
1 and 2; Fig. 2). Contrary to the oral pretreatment effect, preincubation
for 90 minutes in carbocisteine (CCSinc) did not affect sensitivity of the
preparations (-EP) to carbachol (IN) stimulation [-log EC50: 5.71 ±
0.07 (x ±s.e.m.)] (Table. 3).
Removal of the epithelium increased sensitivity to carbachol in
all preparations except in the control preparations stimulated with
carbachol (OUT) (Table 1 and 2). All preparations (+EP) were more
sensitive to carbachol (OUT) stimulation then to carbachol (IN)
stimulation. Interestingly, diminished sensitivity to carbachol (IN)
stimulation of preparations CCSpr(-EP) was absent in preparations

19
preincubated with capsaicin 10-5M (CAPSinc) [-log EC50: 5.99
±0.25 (x ±s.e.m.)] but was maintained in preparations pre-incubated
with indomethacin 10-6M (INDinc) [-log EC50: 5.59 ± 0.09 (x
±s.e.m.)] (Table 3; Fig. 3). In preparations (+EP or - EP) pre-
contracted with (10-6M) carbachol (OUT) carbocisteine alone in the
concentration range from 10-7 to 10-3M (IN) had neither contracting
nor relaxant properties. Also the maximal tension (Tmax) developed
following stimulation with carbachol did not depend on the side of
stimulation (OUT or IN), and was not affected by the removal of
epithelium, pretreatment with carbocisteine, preincubation in carbocisteine
10-3M, indomethacin 10-6M or capsaicin 10-5M (results not shown).
Sensitivity to aminophylline (IN) in the preparations (-EP) precontracted
with carbachol 10-6 (OUT) was not affected by pretreatment with
carbocisteine (-log EC50: 3.23 ±0.1 / 3.66 ±0.11; CCS / Controls; n.
s.). The degree of maximal shortening and the velocity of contraction of
the rat tracheal ring (+ and - EP) stimulated by 10-6M carbachol was
not affected either by pretreatment or preincubation with carbocisteine
(Fig. 4).

20
Discussion
We used an improved in vitro model which permitted independent
perfusion of the epithelial (luminary) and serosal (outside) aspects of the
airway (20, 21, 22). Important feature of this preparation was that each
surface can be stimulated independently and two potentially different
responses distinguished. Furthermore, tracheal epithelium could be
mechanically removed and responses compared, permitting not only
comparison of the responses to luminar and outside pharmacological
stimulation, but also to better appreciate the influence of tracheal
epithelium removal on tracheal muscle contraction. The in vitro model
used this time was essentially the same as the model previously
described except (i.) the pull of the hook on the tracheal wall which could
be adjusted with precision even during the experiment (Fig 1). Before
attempting to adjust the smooth muscle resting tension, the lower hook
was pulled until minimal tension on the higher hook was perceived, and
at that position the micrometric screw was blocked. This allowed more

21
precise adjustment of the tension on the tracheal muscle and optimal
determination of the stretch. (ii.) The system (now ready to be used)
incorporates interchangeable carrier for tracheal or bronchial ring
preparation. This facilitates comparative experiments and does not
impose significant financial burden when doubling equipment and use of
both techniques is necessary.
In our experimental study we found that removal of epithelium
rendered preparations more sensitive to carbachol stimulation. On the
other hand, pretreatment with carbocisteine rendered preparations
without epithelium less sensitive to carbachol stimulation as compared
to controls (-EP). These changes, although small, were significant and
require an explanation. The results of a number of experimental studies,
including ours, have clearly demonstrated that tracheal epithelium can
modulate tracheal smooth muscle contraction (7, 12, 20, 25). However,
it seems that not only the contracting but also the relaxing effects of some
pharmacological agents are dependent on the presence of intact epithelium
(7). This may be relevant for better understanding different airway
pathologies. It has been shown that bronchial epithelium is damaged in

22
patients with severe asthma, illustrating that bronchial epithelial damage
and bronchial hyperreactivity could be linked (13).
There are studies which have demonstrated that bronchial
epithelium could be a powerful diffusion barrier (18, 20) and could
attenuate the effects of pharmacological agents applied luminary. Our
present finding of diminished sensitivity to carbachol (IN) as compared
to carbachol (OUT) in preparations with intact epithelium further
supports this hypothesis. In our earlier study we found difference of the
rate of tension development but we did not find difference in sensitivity
in response to IN and OUT stimulation (20). However previously we
used younger animals (300g) than in this study (400g). We believe that
the capacity of the epithelium to act as a diffusion barrier depends on the
tightness of intercellular junctions, epithelial cell functional properties,
and epithelial cell geometry. All of these could be species or/and age
dependent.
In the present study, diminished sensitivity of tracheal smooth
muscle denuded of epithelium was observed following pretreatment of
animals for 2 weeks with oral carbocisteine and not in tracheal preparations

23
preincubated with the agent in vitro. Our findings confirm our earlier
preliminary experimental results (3, 23) obtained in a preceding in vitro
model (20) and suggest that, following carbocisteine pretreatment, (i)
epithelial diffusion barrier increases (very low sensitivity to carbachol in
preparations +EP, IN), (ii) epithelium presence increases relative
sensitivity of preparations, and (iii) epithelium removal decreases
sensitivity of preparations; resultant effects of (i) and (ii) being visible in
preparations CCSpr, +EP, IN) and that of (iii) in preparations CCSpr,
-EP, IN or OUT.
There is evidence that blocking enkephalinase, which degrades
kinins by phosphoramidon, could increase bronchoconstriction similarly
as epithelium removal (9). Indeed, mechanical removal of the epithelium
could produce liberation of different mediators from mast cells, which
could be responsible, at least in part, for bronchial hyperresponsivnes
observed experimentally (8). It has been shown also that carbocisteine
could increase production of sialomucines (5, 11) and suggested that
sialomucines could have an anti kinin action (19), what could explain
diminished sensitivity of the preparations denuded of the epithelium to

24
carbachol stimulation.
However, capsaicin sensitive nerve terminals could secrete
various mediators (16). It is conceivable that some as yet unidentified
mediators originating from capsaicin nerve terminals could have inhibitory
effects on tracheal smooth muscle contraction. Nerve terminal destruction
by capsaicin and the disappearance of the putative inhibitory agent could
have in turn increased the sensitivity of tracheal smooth muscle to
carbachol as compared - in our experiments - to the preparations
(CCSpr) not pre-incubated with capsaicin. On the other hand,
pretreatment with carbocisteine could have promoted production of
some epithelium derived contracting factor (EpDCF). Removal of the
epithelium and removal of an excitatory agent secreted by the epithelium
could have rendered tracheal smooth muscle less sensitive to carbachol,
which would explain our finding of diminished sensitivity in CCSpr(-EP)
preparations. The prostaglandins, products of arachidonic acid
metabolism, do not seem to be involved since preincubation of the
preparations in Indomethacin did not affect the sensitivity of preparations.
The fact that carbocisteine did not have a direct and immediate effect on

25
tracheal smooth muscle reactivity suggests a complex indirect mechanism.
It is intriguing that one very recent study, that is available only in
Japanese, so we are not going to list it as a reference, suggests that
carbocisteine could modulate cAMP level - what would offer quite
comfortable explanation for its effects on bronchial smooth muscle
contractility (Ishibashi Y, Okamura T, Masumoto Y, Tachiiri T, Momo
K. Effects of carbocisteine on airway inflammation and related events
in SO2-exposed rat. Nihon Kokyuki Gakkai Zasshi, 39: 17-23, 2001).
In conclusion, we have presented an improved model of
bronchial preparation for use in cannulated preparations and, with little
adjustment, for bronchial rings, which makes the system easily accessible
to the investigators. Availability of this system could encourage further
investigation of the role of the bronchial epithelium in bronchial smooth
muscle contraction. In this improved model of cannulated isolated
trachea we have demonstrated that pretreatment of animals with
carbocisteine induces relatively small decrease of reactivity of rat
tracheal smooth muscle denuded of epithelium. As carbocisteine is
widely used as supplementary therapeutics in various pathologies

26
characterised by airway obstruction, further explanation of its mechanisms
of action would be valuable for better understanding possible beneficiary
effects.

27
References
1. Antonissen LA, Michel RW, Kroeger EA, Kepron W, Tse KS,
Stephens NL (1979) Mechanical alterations of airway smooth muscle
in a canine asthmatic model. J Appl Physiol 46: 681 - 687
2. Brown DT (1988) Carbocisteine. Drug Intell Clin Pharm 22: 603-
608
3. Cheick-Zeynneddine, Tais: Effet de la carbocisteine sur la reactivite
du muscle lisse tracheal chez le rat. Memoire pour le DEA de physiologie
et physiopathologie des appareils respiratoire et circulatoire, Annee
universutaire 1990/1991, Paris.
4. Davis SS, Deverell LC (1977) Rheological factors in mucociliary
clearance. The assessment of mucotropic agents using an in vitro model.
Mod Probl Pediatr 19: 207-217

28
5. Degand P (1973) Etude biochemique du mode d’action de la S-
carboxy-methil-cisteine. Bull Physiopathol Respir 9: 462-463,
6. Edwards GF, Steel AE, Scot JK, Jordan JW (1976) S-carboxy-
methil-cysteine in the humidification of sputum and treatment of chronic
airway disease. Chest 70: 505-513
7. Fedan JS, Hay DWP, Farmer SG, and Raeburn D (1988) Epithelial
cells: modulation of airway smooth muscle reactivity. In: Barnes PJ,
Rodger IW, Thompson NC (eds) Asthma, Basic Mechanics and
Clinical Management. Academic Press, pp. 143 - 162
8. Franconi GM, Rubinstein I, Levine EH, Ikeda S, Nadel JA (1990)
Mechanical removal of airway epithelium disrupts mast cells and
releases granules. Am J Physiol 259: L372-L377
9. Frossard N, Strretton CD, Barnes PJ (1990) Modulation of bradykinin

29
responses in airway smooth muscle by epithelial enzymes. Agents
Actions 31: 204-209,
10. Goldie RG, Lulich KM, Henry PJ, Paterson JW (1990)Evaluation
of Airways Smooth Muscle Function in Vitro. In: Agrawal DK and
Townley RG (eds.), Airway Smooth Muscle: Modulation of Receptors
and Response, CRC Press
11. Havez R Degand P, Roussel P, Randoux A (1970) Mode d’action
biochemique des derives de la cysteine sur le mucus bronchique.
Poumon Coeur 26: 81-90
12. Hay, DWP, Farmer SG, Raeburn D, Robinson VA, Fleming WW,
Fedan JS (1986) Airway epithelium modulates the reactivity of guinea-
pig respiratory smooth muscle. Eur J Pharmacol 129: 11-18
13. Laitinen LA, Heino M, Laitinen A, Kava T, Haahtela T. (1985)
Damage of the airway epithelium and bronchial reactivity in patients with

30
asthma. Am Rev Resp Dis.131: 599-606
14. Marom Z, Shelhamer JH, Alling D, Kaliner M (1984) The effect of
corticosteroids on mucous glycoprotein secretion from human airways
in vitro. Am Rev Resp Dis.129: 62-65
15. Martin R, Litt M, Marriott C (1980) The effect of mucolytic agents
on the rheologic and transport propriety of canine tracheal mucus. Am
Rev Resp Dis 121: 495-500
16. Meini S, Evangelista S, Geppetti P, Szallasi A, Blumberg P, Manzini
S (1992) Pharmacological and neurochemical evidence for the activation
of capsaicin-sensitive sensory nerves by lipoxin A4 in guinea pig
bronchus. Am Rev Resp Dis 148: 930-934
17. Molina Cl, Aiache JM, Pradelle J, Brun J, Cheminat JCl (1970)
Interet clinique des medications fluidifiantes des secretions bronchiques.
Poumon Coeur 26: 91-101

31
18. Morakawa T, Webber SE, Widdicombe JG (1991) The effect of
hydrogen peroxide on smooth muscle tone, mucus secretion and
epithelial albumin transport of the ferret trachea in vitro. Pulmonary
Pharmacol 2: 106-113
19. Muh JP, Latreille JP, Guerrin F (1968) Recherche d’inhibiteurs
naturels du bronchospasme a la bradykinine chez le cobaye. In:
Hypersecretion bronchique, Colloque international de pathologie
thoracique, p 193-202. Clichy, Poissot, Medic diffuser
20. Pavlovic D, Fournier M, Aubier M, Pariente R (1989) Epithelial vs.
serosal stimulation of tracheal muscle: role of epithelium. J Appl Physiol
67: 2522-2526
21. Pavlovic D, Brione E, De Vernejoul D, Aubier M (1993) Partial
inhibition by epithelium of tracheal smooth muscle relaxation induced by
the potassium channel activator, BRL 38227. Br J Pharmacol 110: 139-

32
144
22. Pavlovic D, Viires N, Zedda C, Fournier M, Aubier M (1998)
Effects of a high dose corticosteroids on smooth muscle function in rat
trachea. Eur Respir J 11: 575-582
23. Pavlovic, D., Cheik-Zeinedinne, T., Fournier, M., and Aubier, M.
(1994) Carbocisteine diminishes rat tracheal smooth muscle reactivity
in epithelium denuded preparations. Eur. Resp. J. Vol 7, suppl 18: 76s.
24. Puchelle E, Aug F, Polu JM. (1978) Effects of the mucoregulator
S-carboxy-methil-cisteine in patients with chronic bronchitis. Europ J
Clin Pharmacol. 14: 177-184
25. Stuart-Smith K, Vanhoutte P. (1988) Airway epithelium modulates
the responsiveness of porcine bronchial smooth muscle. J Appl Pysiol
65: 721-727

33
26. Wanner A, Zarzecki S, Hirsch J, Epstein S. (1975) Tracheal mucus
transport in experimental canine asthma. J Appl Physiol 39: 950-957

34
Table 1 EC50, -log (M), for carbachol (IN)
+EP -EP p
___________________________________________________
contr 5.40 ±0.11 5.85 ± 0.06 <0.005
CCSpr 5.19 ±0.08 5.52 ± 0.09 <0.005
p n.s. <0.005
Means (± s. e. mean) of -log EC50, -log (M) values, obtained
in preparations with (+EP) and without epithelium (-EP), taken from
controls (contr) or animals pretreated for 2 weeks with oral carbocisteine
200mg/kg/day (CCSpr), following stimulation from the epithelial side
(IN) with cumulative concentrations of carbachol (n. s. = non significant).

35
Table 2 EC50 for carbachol (OUT)
+EP -EP p
___________________________________________________
contr 6.01 ± 0.09 5.92 ± 0.06 n.s.
CCSpr 6.17 ± 0.08 5.61 ± 0.05 <0.005
p n.s. <0.005
Means (± s. e. mean) of -log EC50, -log (M) values, obtained
in preparations with (+EP) and without epithelium (-EP), taken from
controls (contr) or animals pretreated for 2 weeks with oral carbocisteine
200mg/kg/day (CCSpr), following stimulation from the serosal side
(OUT) with cumulative concentrations of carbachol (n. s. = non
significant).

36
Table 3 EC50 for carbachol (-EP, IN) following
preincubation
-EP
_____________________________________
contr 5.85 ± 0.06
CCSinc 5.71 ± 0.07
CCSpr 5.52 ± 0.09
CCSpr (INDinc) 5.59 ± 0.09
CCSpr (CAPSinc) 5.99 ± 0.25
Means (± s. e. mean) of -log EC50, -log (M) values, obtained
in preparations without epithelium (-EP) following stimulation from the
epithelial side (IN) with cumulative concentrations of carbachol.
Preparations were taken from controls (contr) and pre-incubated in
carbocisteine 10-3M (CCSinc) or from animals pretreated for 2 weeks
with oral carbocisteine 200mg/kg/day (CCSpr) and pre-incubated in

37
indomethacin 10-6M (INDinc) or capsaicin 10-5M (CAPSinc) (contr/
CCSpr p<0.005, values presented also in the table 1, CCSpr/CCSpr
(CAPSinc) p<0.03).

38
Figure legends
Figure 1
Schematic representation of the experimental apparatus. (A) The new
tracheal tube preparation with the “carrier” block. Inner, luminary (IN)
and outer (OUT) perfusion solutions are maintained at 37° C, bubbled
with 95% O2 - 5% CO2 and a constant flow rate of 2 ml/min is
maintained. Under microscopic control two stainless steel hooks were
passed through the tracheal wall around two adjacent cartilagineous
rings as close as possible to the tracheal muscle insertions. The lower
hook, which served as a “fixed point”, was attached with the silk thread
to the micrometric screw serving to adjust the hook tension on the
tracheal wall. This allowed precise adjustment of the resting tension. The
upper hook was connected to a force transducer. (B) Tracheal ring
preparation. The “carrier block” for cannulated trachea could be easily
removed and replaced by one suitable for tracheal rings mounting.

39
Figure 2
Cumulative concentration - responses curves constructed after
administration of carbachol epithelialy (IN, a and c) or serosaly (OUT,
b and d) in rat isolated trachea with (+EP, a and b) and without
epithelium (-EP, c and d). Preparations were taken from control animals
or animals pretreated for two weeks orally with carbocisteine 200mg/
kg/day (? controls, ? CCSpr). Tension is expressed as percentage of the
maximal tension (Tmax) obtained and presented as mean data ± s. e.
mean.

40
Figure 3
Cumulative concentration - responses curves constructed after
administration of carbachol epithelialy (IN) in rat isolated trachea
without epithelium (-EP). Preparations were taken from animals
pretreated for 2 weeks with carbocisteine 200mg/kg/day and not
incubated (CCSpr, ?) or incubated for 60 min. in capsaicin 10-6M
(CCSpr + capsaicin, ?). Tension is expressed as percentage of the
maximal tension (Tmax) obtained and presented as mean data ± s. e.
mean.

41
Figure 4
Relative velocity of contraction (time to 50, 80 and 100% of maximal
shortening) elicited with carbachol 10-6M in tracheal rings preparations
taken from animal pretreated with oral carbocisteine (abscissa: time in
seconds; ordinate: degree of isothonic shortening in percents of maximum
(mean ± s. e. mean); ?, CCSpr (+EP); ?, controls (+EP) ?, CCSpr (-
EP), and ?, controls (-EP); (differences not significant).

42
To transd u ce r
In n e r so lu tio n
To m icro m etric sc rew
M icro m e tricscrew
P lex ig lass h o ld erA .

43
Trach ea l r in g
To tran sd u ce r
P lex ig lass h o ld er
B .

44
100
80
60
40
20
0
0 -8 -7 -6 -5 -4 -3 -2
% T
max
carbachol (log), M (IN)
Fig 2a
+EP
controlsCCS

45
100
80
60
40
20
0
0 -8 -7 -6 -5 -4 -3 -2
% T
max
carbachol (log), M (OUT)
Fig 2b
+EP
controls
CCS

46
100
80
60
40
20
0
0 -8 -7 -6 -5 -4 -3 -2
% T
max
carbachol (log), M (IN)
Fig 2c
-EP
controls
CCS
47
100
80
60
40
20
0
0 -8 -7 -6 -5 -4 -3 -2
% T
max
carbachol (log), M (OUT)
Fig 2c
-EP
controls
CCSpr

48
100
80
60
40
20
0
0 -8 -7 -6 -5 -4 -3 -2
% T
max
carbachol (log), M (IN)
Fig 3
-EP
CCpr+ capsaicine
CCSpr

49
100
80
60
40
20
0
0 200 400 600 800 1000
% M
AX
TIME (s)
Fig 4

50
9. Appendix: Works related to the presented model
A. Own works
Pavlovic, D., M.Fournier, M.Aubier, and R.Pariente.
Modulation of the tracheal smooth muscle response to carbachol
stimulation by the epithelium in the rats.
Am.Rev.Resp.Dis. 137 (suppl):243, 1988.
Pavlovic, D., M.Fournier, M.Aubier, and R.Pariente.
Epithelial versus serosal stimulation of tracheal muscle: role of
epithelium.
J.Appl.Physiol. 67: 2522-2526, 1989. (*)
Pavlovic Dragan, Evelin Brione, Michel Fournier, et Michel Aubier.
L‘epithelium inhibe la relaxation du muscle lisse tracheal de rat
induit par un activateur des canaux potassiques (BRL 38227).
Proceedings of Journee de la Recherche (UFR Xavier Bichat), Paris,
Sept. 1990.

51
Pavlovic, Dragan., Evelin Brione, Michel Fournier, and Michel Aubier:
Epithelium partialy inhibits tracheal smooth muscle relaxation
induced by potassium channel activator BRL 38227.
Br. J. Pharmacol., 110: 139-144, 1993.
Pavlovic, Dragan., Naima Viires, Christine Zedda, Michel Fournier,
and Michel Aubier:
Effects of corticosteroids on epithelial structure and smooth
muscle function of rat trachea
Am. J. Respir. Crit. Care Med., 4: A905, 1994.
Pavlovic, D., Cheik-Zeinedinne, T., Fournier, M., and Aubier, M.
Carbocisteine diminishes rat tracheal smooth muscle reactivity
in epithelium denuded preparations.
Eur. Resp. J. Vol 7,suppl 18: 76s, 1994.
Pavlovic, D., Moldovan, F., and Aubier, M.
Selective perfusion of rat tracheal preparation: no evidence for
EpDRF secretion
Eur. Resp. J. Vol 8, suppl 19: 43s, 1995
D. Pavlovic, N. Viires, F. Roux, and M. Aubier

52
Chronic Hypoxia Diminishes Sensitivity of Rat Tracheal Smooth
Muscle to Carbachol Applied Epithelialy
Am. J. Resp. Crit. Care. Med., Vol. 153, N8 4 (suppl., part 2): A840,
1996.
Ouksel, H., D. Pavlovic, N. Viires, Y. Amrani, N. Seta, and M. Aubier
Modifications in MLCK Expression Induced by Repeated
Bronchoconstriction
EMC, XXV European Muscle Congress, Montpellier, France, 14 - 17
Sept., 1996.
Ouksel, H., N. Viires, D. Pavlovic, C. Peiffer, N. Seta, Y. Amrani, and
M. Aubier
Modifications in MLCK Expression in Guinea-pig Airway Smooth
Muscle: Role of Repeated Bronchoconstrictions and Role of
Inflamation
Am. J. Resp. Crit. Care. Med. Vol 155, N8 4 (suppl, part 2): A370,
1997.
Ouksel, H., N. Viires, D. Pavlovic, C. Peiffer, C. Zedda, and M. Aubier
MLCK and SERKA Expression in Tracheal Smooth Muscle
From Sensitized Guinea-pigs

53
Eur. Resp. J. vol 10, 291s (suppl), 1997.
Ouksel, H., N. Viires, D. Pavlovic, C. Peiffer, C. Zedda, M. Pretolani,
C. RuffiÈ, and M. Aubier
Effects of Inflamation on Myosin Light Chain Konase (MLCK)
Expression in Guinea-Pig Model of Bronchial Hyperreactivity
Am. J. Crit. Care Med., Vol 157, 3 (Suppl): A519, 1998
Pavlovic, Dragan., Naima Viires, Christine Zedda, Michel Fournier,
and Michel Aubier:
Effects of a high dose corticosteroids on smooth muscle function
in rat trachea.
Eur. Respir. J., 11: 575-582, 1998
H. Ouksel, N.Viires, D. Pavlovic, C. Ruffie, C. Zedda, C. Peiffer, C.
Vizzuzaine, M. Pretolani, M. Aubier
Allergic Bronchial Hyperreactivity In The Guinea-Pig Is
Associated With An Inflammation-Dependent Increase In The
Expression Of MLCK In the Tracheal Smooth Muscle
Am. J. Crit. Care. Med. vol 161 (3) (Suppl): A840, 2000.
A. Samb, C. Lisdero, J. Callebert, D. Pavlovic, M. Pretolani, M.

54
Aubier, J. Boczkowski
Decreased Lung Expression And Activity Of Type I Nitric Oxide
Synthase (nNOS) After Ovalbumin Immunisation And Aerosol
Challenge In Guinea Pigs.
Am. J. Crit. Care. Med. vol 161 (3) (Suppl): A919, 2000.
Submitted for publication
Pavlovic, D., Cheik-Zeinedinne, T. N., Nedeljkov, V., Wendt, M., and
Aubier, M. A
New Interchangeable Tube/Ring Tracheal Preparation: Effects
of Carbocisteine.
(J. Appl. Physiol.)
Ouksel, H., N. Viires, D. Pavlovic, C. Peiffer, C. Ruffie, C. Vissuzaine,
M. Pretolani, and M. Aubier,
Quantitative Modifications of Myosin Light Chain Kinase
Associated with Airway Inflammation in a Guinea-pig Model of
Bronchial Hyperreactivity
(J Clin. Invest.)

55
B. Works of other investigators using our model
The model has been taken up by a group from Utrecht that
produced at least one PhD (Dr. Sadeghi-Hashjin G) and number
of publications. We give a short list of some of them as appear in
Medline.
Folkerts G, Kloek J, Geppetti P, Van der Linde HJ, Nijkamp FP.
Factors that determine acetylcholine responsiveness of guinea
pig tracheal tubes.
Eur J Pharmacol. 2001 May 25;420(2-3):151-7.
Sadeghi-Hashjin G, Folkerts G, Henricks PA, Van de Loo PG, Van der
Linde HJ, Dik IE, Nijkamp FP.
Induction of guinea pig airway hyperresponsiveness by
inactivation of guanylate cyclase.
Eur J Pharmacol. 1996 Apr 29;302(1-3):109-15.
Figini M, Ricciardolo FL, Javdan P, Nijkamp FP, Emanueli C, Pradelles
P, Folkerts G, Geppetti P.
Evidence that epithelium-derived relaxing factor released by

56
bradykinin in the guinea pig trachea is nitric oxide.
Am J Respir Crit Care Med. 1996 Mar;153(3):918-23.
Folkerts G, van der Linde H, Verheyen AK, Nijkamp FP.
Endogenous nitric oxide modulation of potassium-induced changes
in guinea-pig airway tone.
Br J Pharmacol. 1995 Aug;115(7):1194-8.
Folkerts G, van der Linde HJ, Nijkamp FP.
Virus-induced airway hyperresponsiveness in guinea pigs is
related to a deficiency in nitric oxide.
J Clin Invest. 1995 Jan;95(1):26-30.
Nijkamp FP, van der Linde HJ, Folkerts G.
Nitric oxide synthesis inhibitors induce airway
hyperresponsiveness in the guinea pig in vivo and in vitro. Role
of the epithelium.
Am Rev Respir Dis. 1993 Sep;148(3):727-34.

57

58
LEBENSLAUF
Dragan Pavlovic
____________________________________________________________________________________
PERSONAL
Geburt Datum und Ort: 4 August, 1949, Vranje, Jugoslawien
Staatsangehörigkeit: Französisch
Adresse:
Arbeit: Klinik und Poliklinik für Anästhesiologie und
Intensivmedozin
Ernst-Moritz-Arndt Universität, 17487
Greifswald
Friedrich-Loeffler-Strasse 23b
Tel.: (49) 3834 - 86 5800
Fax: (49) 3834 - 86 5802
e-mail: [email protected]
Hause: Walter-Rathenau-Str. 46 (2.6)17489 GreifswaldTel.: (49) 3834 - 86 2928e-mail: [email protected]
______________________________________________________________________________
DIPLOME
ArztDoktor en Medizin, Belgrad Universität, Jugoslawien (25 Juni 1976)
Approbation : 25. 10. 1977.

59
D.S.M.Diplom in Sports Medizin, The London Hospital Medical College,London, Groß Britannien. (Juni 1984)
______________________________________________________________________________
PRAKTISCHE AUSBILDUNG UND ARBEIT
1976-1977 Ärztlicher Vorbereitungsdienst (AiP), MedizinischeFakultät Belgrad Universität, Jugoslawien ( 4. 7. 1976 - 17. 7. 1977)und Sportmedizin Praxis, �Medicinar�, Belgrade, Jugoslawien
1977-197 Allgemeiner Arzt, Ambulanz Porec undKoper, Jugoslawien. (18. 7. 1977 - 20. 8 1978)
1978-1979 Assistenzarzt, Abteilung für Anästhesie undIntensivmedizin, Parazelsus Klinik, Marl, W. Deutschland. (1. 9.1978 - 31. 7. 1979)
1979-1981 Sports Medizin Praxis (Sportmedizin ÄrztlicherVorbereitungsdienst), �Medicinar� Karate Club, Belgrade,Jugoslawien. (1. 8. 1979 � 30. 7. 1981)
1981-1983 Assistenzarzt, Abteilung für Anästhesie undIntensivmedizin ,Waiblingen, W. Deutschland.(15. 8. 1981 - 31. 5. 1983)
1983-1984 Sports Medizin Fachausbildung, The London HospitalMedical College, London, Groß Britannien . (25 September 1983- 5Juni 1984)
1985 - 2000 Wissenschaftlicher Mitarbeiter, und seit 1987, Leiter,Abteilung für glatte Muskulatur, FacultÈ Xavier - Bichat, INSERMU408 (226) und Beaujon Hospital, Abteilung für Pneumologie undIntensivmedizin, Clichy, Paris, Frankreich.(6. 3. 1985 - 31.12. 1999)
2000 - ... Assistenzarzt und Wissenschaftlicher Mitarbeiter ,Klinik und Poliklinik für Anästhesiologie und Intensivmedizin, Ernst-Moritz-Arndt Universität, Greifswald, B. R. Deutschland (seit 1. 1.2000 - ... )
______________________________________________________________________________
WEITERE AUSBILDUNG
1983 Kursus in British Medical Practice auf PostgraduateMedical School, University of Exeter, Groß Britannien(April 1983 - Juni 1983).
1985-86 Kursus in Physiologie und Pharmakologie des

60
Atemweges, auf UniversitätParis V., Frankreich. (September 1985-Juni 1986)
Sommer 1989, Kursus in patch-clamp, auf Faculte des Sciences (Pr.M. Lazdunski), Nice, Frankreich, und auf Faculte de Pharmacie,Strassburg, Frankreich (Pr. Stockle).
Sommer 1991, Kursus in �Optical techniques of membrane potentialrecording and intracellular ion concentration recording�, auf Facultede Medicine , Marseille (Pr. Bongrand), Marseille, Frankreich.______________________________________________________________________________
FORCHUNG
1985 - 1999 Wissenschaftlicher Mitarbeiter, UnitÈ INSERM U408,FacultÈ Xavier - Bichat,Paris, Frankreich (6. 3. 1985-31. 12. 1999).
2000 - ... Wissenschaftlicher Mitarbeiter, Klinik und Poliklinikfür Anästhesiologie und ntensivmedizin, Ernst-Moritz-ArndtUniversität, 17487 Greifswald, B. R. Deutschland.(1. 1. 2000 -....)
______________________________________________________________________________
VERANTWORTUNGEN
1987 - 2000 Leitung: Abteilung für glatte Muskulatur, UniteINSERM U408, Faculte Xavier - Bichat, Universität Paris 7, Paris,Frankreich.
2000- ... Wissenschaftliche Mitarbeiter, Klinik und Poliklinik fürAnästhesiologie und Intensivmedozin, Ernst-Moritz-ArndtUniversität, Greifswald, B. R. Deutschland (1. 1. 2000- ...).

Epithelial vs. serosal stimulation of tracheal muscle: role of epithelium
DRAGAN PAVLOVIC, MICHEL FOURNIER, MICHEL AUBIER, AND RENE PARIENTE Service de Pneumologie et Reanimation and Institut National de la Sante et de la Recherche Mkdicale U226, Hopital Beaujon, 92110 Clichy, France
PAVLOVIC, DRAGAN, MICHEL FOURNIER, MICHEL AUBIER, AND RENE PARIENTE. Epithelial vs. serosal stimulation of tra- cheal muscle: role of epithelium. J. Appl. Physiol. 67(6): 2522- 2526,1989.-There is evidence implying an active role of airway epithelium in the modulation of bronchomotor tone. To study this phenomenon, we designed an in vitro system allowing pharmacological stimulation of either the inside or outsides of the airway lumen. Rat tracheas were excised, cannulated, and their inside and outside perfused independently with Krebs solution. Two hooks were inserted through opposite sides of the tracheal wall, the lower one was attached to a fixed point, while the upper one was connected to a force transducer. Isometric contractions of the tracheal muscle were elicited by carbachol solution perfused in single and cumulative concen- trations. In one-half of the preparations the epithelium was mechanically removed. Stimulation of the inside or outside of the trachea produced equal maximal tracheal muscle tension [l.55 t 0.14 and 1.2 * 0.09 (SE) g in and out, respectively]. The time course of tension development was longer when carbachol was administered inside the trachea: an effect that was abolished when the epithelium was removed. In addition, removal of the epithelium was found 1) to increase the maximal tension irrespective of the route of carbachol perfusion and 2) to increase the sensitivity of the preparation to carbachol stimulation.
of asthmatic patients (4, 13, l4), 2) bronchial hyperre- sponsiveness is a hallmark of asthma (8, 16), and 3) in asthmatic patients, access of constricting allergens is, most often, via airways.
The aim of the present study was therefore to design an experimental set up for in vitro studies of the airway smooth muscle responsiveness allowing an exclusive con- tact between either airway epithelium or serosa and the agent tested.
MATERIALS AND METHODS
smooth muscle; rat; carbachol
Experiments were performed on tracheas taken from male Sprague-Dawley rats (300-350 g body wt) that were stunned by a blow on the head and quickly exsanguin- ated. The tracheas were immersed in Krebs solution (in mM: 137 NaCl, 4 KCl, 1 MgC12, 1 KH2POJ, 12 NaHC03, 2 CaC12, 6.5 glucose) and cleaned from all surrounding tissue. Proximal ends (-10 rings long) were used for the experiments and distal ends were cut off. In one-half of the preparations the epithelium was removed (-EP) by gently rubbing with a cotton-wrapped metal stick; in the other half of the preparations the epithelium was left intact (+EP).
-
THERE IS EXPERIMENTAL evidence suggesting that epi- thelial damage and bronchial hyperresponsiveness are tightly linked. Indeed, numerous well-controlled studies performed in various animal species and humans have demonstrated increased airway responsiveness after damage to the epithelium caused by inhalation of citric acid (l), ozone (5, 8), or viral infection (12). In addition, it has been shown that mechanical removal of the epi- thelium in several species increases the sensitivity of tracheal strips to various bronchoconstrictor agents (2, 3, 6, 7, 9, 11). However, it should be noted that for in vitro experiments tracheal or bronchial tissue was totally immersed into physiological salt solution, with conse- quent simultaneous pharmacological stimulation of the serosal and epithelial sides of the airways. It was there- fore difficult to examine the specific role played by the epithelium in modulation of airway muscle tone.
An organ bath was constructed that permitted inde- pendent circulation of fluid within the lumen of the tracheal segment (In, epithelial side) or around the ex- terior (Out, serosal side) of the tracheal segment (Fig. 1). A modified 5-ml syringe with top and lateral openings served as the organ bath. The piston served as a support for the tubing system used to intubate and secure the tracheal segments in place. Mounting of tracheal seg- ments involved the following.
Under microscopic control two stainless steel hooks were passed through the tracheal wall around two adja- cent cartilaginous rings as close as possible to the muscle insertions. The tracheal segment was then longitudinally connected to the steel tubes built in the piston and firmly tightened with silk thread. The lower hook was attached below, serving as a fixed point. Its length was adjusted so that it did not pull down the tracheal wall.
The question as to whether airway epithelium is play- ing an active role in the modulation of underlying muscle tone is of considerable importance because 1) epithelial damage is a common pathological finding in the airways 2522 0161-7567/89 $1.50 Copyright 0 1989 the American Physiological Society
The piston was then introduced into the syringe and the upper hook connected to a force transducer (UC& Gould, Cleveland, OH), the latter being attached to the micromanipulator (Prior PO22, Prior Scientific Instru- ments, Herts, UK) that enabled the displacement of the upper hook along a strict vertical axis. Any change of tension at the level of the tracheal muscle was registered

TRACHEAL SMOOTH MUSCLE IN RAT 2523
by the recorder (Gould AT 550) to which the transducer was connected.
The Krebs solution (at 37”C, pH 7.4, gassed with 95% OZ-5% C02) was perfused at a constant flow rate (2 ml/ min) through the syringe organ bath (outer perfusion) and through the lumen of the tracheal segment (inner perfusion) by using peristaltic pumps (Watson Marlow 5025, Falmouth, Cornwall, UK).
Fluid tightness of preparation. To ensure that hooks did not induce a fluid leak through the tracheal wall, a solution of methylene blue was perfused into the tracheal lumen and in the organ bath in separate experiments. No cross-staining was observed.
Procedure. After a period of stabilization (45-60 min), by slowly raising the transducer, the trachealis was stretched to its optimal length established in a prelimi- nary set of experiments. This consisted of a series of maximal contractions induced by 10B3 M carbachol in the outer circuit while changing incrementally tracheal muscle length, i.e., passive tension. The length-tension relationships were not found to differ significantly in preparations with and without epithelium. Appropriate amounts of carbachol (Sigma Chemical) were diluted in small volumes (100 ~1) of distilled water and added to the inner and outer Krebs solution reservoirs to obtain the required final concentrations.
In the first set of experiments (Table 1) the effects of a single concentration of carbachol (10e3 M) on tracheal muscle were examined. The agent was left in contact with the preparation until the response reached a pla- teau. The preparation was then perfused with Krebs solution and allowed to relax completely. The alternate routes of perfusion were tested in a randomly assigned order [In-(wash)-Out-(wash) or Out-(wash)-In-(wash)] for each preparation. In the second set of experiments (Table 1) lo-fold increments of concentration of car- bachol were used (10B8 to 10B3 M) in cumulative fashion. In the third set of experiments (Table 1) cumulative concentrations of carbachol were also used to produce control contractions perfusing only from the interior. This was followed by 30-min incubation with low6 M
A
TRANSDUCER
,t [
IN
FIG. 1. Schematic representation of measurement system. Inner (IN) and outer (OUT) perfusates are maintained at 37°C and bubbled with 95% 02-5% COY, and constant flow rate of 2 ml/min is maintained.
TABLE 1. Experimental protocol
Preparation n Protocol
1 C - - - - - - - - - -+ w---------------+ C - - -- - - ---- + w
+EP 18 1O-3 M lo-“M -EP 14 In (or Out) Out (or In)
2 C -e-------- + w---------------+ C - ----- -- - -+ w
+EP 14 1o-s to 1o-8 to -EP 10 lo-” M lo-" M
In (or Out) Out (or In) 3 C ------m---j w _-__ Indo __--- + C ---------- + w
+EP 8 1o-7 to 1O-6 M 1o-7 to -EP 8 1O-3 M In + Out lo-’ M
In In 4 C ------m---j w---------------+ C - -- --- - - --+ w
+EP 8 1O-7 to 1o-7 to -EP 8 lo-” M lo-' M
In + Out In + Out
+EP, epithelium intact; -EP, epithelium denuded; C, carbachol; w, wash; In, inside (epithelial side); Out, outside (serosal side); Indo, indomethacin. See text for details.
indomethacin (Indo, Sigma Chemical) and then stimu- lations with interior perfusion repeated. Finally, in the fourth set of experiments (Table l), both sides of the preparations were perfused simultaneously with at first cumulative concentrations of carbachol solutions; this was followed by one supramaximal stimulation with 10s3 M carbachol. For the latter three sets of experiments concentration-effect curves were constructed.
After completion of the experiments, the tracheal seg- ments, eight without epithelium (-EP) and four with epithelium (+EP) taken at random, were removed from the organ bath and fixed in 2.5% glutaraldehyde. Semi- thin sections from plastic-embodied blocks were then prepared, stained with toluene blue, and examined mi- croscopically for the presence of epithelium and/or for possible damage of the tracheal wall and epithelium provoked by the hooks. A quantification of the amount of epithelium present was performed by estimating the number of nuclei of epithelial cells in the whole circum- ference of the cross-section of the tracheal segment (in- tact circumference was taken to be 100%).
Analysis of results. The data are expressed in percent of maximal response and in absolute values (g and s). The results are given as means t SE. Half-maximal concentration (EC& values were calculated from regres- sion analysis of probit-transformed data, and results are given as geometric means of log EC& obtained. Statistical analysis was conducted by use of the Student’s t test for paired or unpaired samples and linear regression analy- sis, where appropriate. P < 0.05 was regarded as being statistically significant.
RESULTS
Histology. Twelve preparations, among which eight had been rubbed for epithelium removal, were selected at random for histological evaluation. In -EP prepara- tions 60-80% of the epithelium was removed without any obvious damage to the underlying submucosa or muscle layer (Fig. 2). After 3 h in the organ bath, 60- 80% of the epithelium remained intact in the control tissues (+EP).
Tissue bath studies. As shown in Fig. 3, which repre-

2524 TRACHEAL SMOOTH MUSCLE IN RAT
-
FIG. 2. Histological sections of glutaraldehyde-fixed Epon-embedded tracheal blocks (toluene blue, X640 magnification); gently rubbing of interior of tracheal segment denudes preparation of epithelial cells (control preparation is on left).
C w
! !
L......J
60s
FIG. 3. Superimposed typical tracings of rat tracheal muscle contraction after stimulation with 10-3 M carbachol (C) perfused inside(IN) and outside (OUT) lumen of an intact (+EP) preparation; note delayed rate of tension development when stimulating from interior.
sen ts typical tracings obtained in one + EP and one -EP preparation, supramaximal concentration of carbachol (10-3 M) alternately perfused in the inner and outer circuits gave superimposable maximal changes in tension. The -EP preparations were found to be more sensitive to carbachol stimulation and showed a leftward shift of concentration-effect curves with leftward displacement of EC60 (Fig. 4, Table 2). That effect was independent of the perfusion route. Moreover, removal of the epithelium was associated with a significant increase in the mean maximal tension (Tmax) developed (Fig. 5, Table 3). Although the mean Tmax developed was not influenced by the route of carbachol application in a given preparation (+EP or -EP), the time course of tension development after interior stimulation depended very much on whether epithelium was present or re-
T (91 3 WITHOUT EPITHELIUM
T(gl 3 WITH EPITHELIUM
Oo O
0
§2 o�o O 00
!:: 2 ::, 0 Ooo u 0 u
a: 0 a:: o: � u 0 0 u
5 o 0
a:: 0 0
�I I •0.83 �1 0 0
r•0.54 c:, c:,
p<0.01 p<0.05 n= 14 n■ 18
2 3 INNER CIRCUIT INNER CIRCUIT
FIG. 5. Individual values of maximal tension (Truax) recorded after stimulation with 10-:i M carbachol solution from epithelial side (in) and serosal side (out) in intact (A) and epithelium-denuded (B) preparations.
TABLE 2. Geometric means of log EC5o values
Out In
+EP{n = 14)
0.8 X 10-• 3.5 X 10-•
-EP{n = 10)
7.8 X 10-'
9.7 X 10_,
p
<0.05 <0.03
Values are expressed in M. EC.,,, half-maximal concentration; +EP, epithelium intact; -EP. epithelium denuded. Preparations were stimulated from exterior (Out) or interior (In) with cumulative concentrations of carbachol.
moved (Figs. 3 and 6, Table 4). The time required to reach 80% of Tmax was clearly influenced by the route of perfusion. Compared with outer stimulation, inner perfusion of carbachol in intact preparations showed a

TRACHEAL SMOOTH MUSCLE IN RAT 2525
WITH EPITHELIUM WITHOUT EPITHELIUM
100 A � 100 B � _,,,. 80 80
/ , , /
; , ,
j /
� 50 ,/ n, \8 50 /
I l= i tSEM ,;, "' o--o:: out -= in
r----·r-- -----,.,,,.___,__, � 50 100 200 0 50 IO0 200 300
TIME (sl TIME Is)
FIG. 4. Cumulative concentration-response curves constructed after stimulation with carbachol perfused from epithelial or serosal side of trachea with (n = 14) and without (n = 10) epithelium.
TABLE 3. Maximal tension of intact or epitheliumdenuded rat tracheal smooth muscle after stimulation from epithelial or serosal side
10-, M Carbachol Cumulative Concn of Carbachol
+EP -EPp
+EP -EPp
(n = 18) (n = 14) (n = 14) (n = 10)
Out 1.65±0.09 2.07±0.11 <0.01 l.2±0.1 2.15±0.12 <0.001In 1.57±0.07 1.99±0.11 <0.01 1.55±0.14 2.3±0.15 <0.005
Values are means± SE in g. See Table 1 footnote for definition of abbreviations.
�mo (I) z
:i: � a:
OUT
-EP
4 3 8 7
-log (CARBACHOll,M
IN
-EP
6
FIG. 6. Mean values in 18 preparations of rate of tension development recorded during supramaximal stimulation with carbachol solution either from epithelial side (e) or serosal side (0). A, with epithelium; B, without epithelium. The time required to develop 80% of maximal tension (Truax) was significantly longer (P < 0.001) in preparations with epithelium on interior stimulation compared with outer stimulation. This difference was abolished when epithelium was removed.
TABLE 4. Effects of removal of epithelium stimulated with carbachol simultaneously from epithelial and serosal sides
EC,,., Tmax, t/50% t/80% g Tmax, s Tmax, s
+EP 2.7 X 10-" 1.46±0.18 23±7.8 42±6.0 -EP 6.8 X 10-7 2.37±0.20 12±6.0 24±3.0 p <0.05 <0.01 NS NS
t/Tmax, s
240±31.7 198±47.6
NS
Values are means± SE. EC50, half-maximal carbachol concn; Tmax, maximal tension; t, time; +EP, epithelium intact; -EP, epithelium denuded.
diminished rate of tension development. Removal of the epithelium was found to abolish this effect. In the third series of experiments, 30 min of incubation in 10-6 M lndo solution did not affect contractile response of preparations to inside stimulation with carbachol solution
[EC50 (M), (n = 8): 6.8 X 10-6 vs. 6.2 X 10-0, +EP control
and +EP Indo, respectively; and 0.5 x 10-s vs. 9.8 X
10-1, -EP control and -EP Indo, respectively; differ
ences failing to show statistical significance]. Finally, simultaneous inside and outside perfusion with carbachol solution (fourth series of experiments) did not produce different responses compared with one side perfusion (Table 4). The rate of tension development was quick and comparable with the response obtained on outside stimulation only.
DISCUSSION
The main findings of this study are that removal of the epithelium 1) increases the tension developed by tracheal smooth muscle after maximal stimulation with carbachol, 2) influences the time course of the response, and 3) increases its sensitivity to carbachol stimulation.
The purpose of this study was to develop an experimental model allowing an exclusive contact between a drug and the mucosa! and/or serosal surface of the tracheal wall. In this experimental set up, however, mechanical removal of the epithelium may have altered the length-tension relationship of the tracheal smooth muscle, which in turn could have influenced our results. Therefore, preliminary studies were focused on the determination of the optimal tension. For the results to be reproducible it was found that adequate positioning of the hooks was very important. Two requirements had to be fulfilled: 1 ) hooks had to be inserted as close as possible to the muscle attachments, and 2) they had to be in the same vertical plane to avoid torque reactions when stretching the preparation. Once those precautions were taken, we found that removal of the epithelium had no appreciable effect on the basic mechanical properties of the tissue, as assessed by the shape of length-tension curve. These observations are in agreement with previous studies conducted with tracheal strips or bronchial rings from guinea pigs (9), rats (7), dogs (6), or cattle (2).
Mechanical removal of the epithelium has been shown to increase the sensitivity of airways to various bronchoconstrictor drugs in those species, but the mechanisms involved remain unclear. The possibility that epithelium removal could act as a loss of a permeability barrier, permitting access of more agonist to the underlying smooth muscle, has been proposed by Holroyde (11). The experimental model used in the present study allowed an accurate comparison between changes in tension induced by an agonist perfused alternatively from the mucosa! or serosal side of the intact preparations. The tension developed during carbachol stimulation was not affected by the route of perfusion, but the time required to reach 80% of tension developed was significantly longer when carbachol was perfused in the inner circuit compared with the outer circuit. We therefore hypothesized that + EP could act in vitro by limiting the amount of agonistreaching smooth muscle across the epithelial layer. Theobservation that epithelium removal abolishes the difference in the time course of tension development betweeninside and outside stimulation with carbachol is in agreement with this hypothesis. However, in +EP for highconcentrations of carbachol solution ( 10-3 M) and aher

2526 TRACHEAL SMOOTH MUSCLE IN RAT
sufficiently long exposure, no difference in tension de- veloped (Tmax) was observed for either route of perfu- sion. This indicates that the epithelium as a diffusion barrier is not powerful enough for these high concentra- tions of carbachol.
It was found in the present study that epithelium removal was accompanied by an increase in Tmax de- veloped, as well as an increased sensitivity of the prepa- ration to carbachol. Those effects were observed inde- pendently of the route of perfusion, suggesting that epi- thelium does not act simply as a diffusion barrier to exogenous substances. Previous studies performed on airway strips from various animal species have provided evidence that epithelium was capable of modulating the reactivity of underlying smooth muscle. For example, -EP preparations have been shown to exhibit increased sensitivity to carbachol and acetylcholine when com- pared with intact preparations; our finding is clearly in agreement with these observations.
Taken together, the results of the present study, with the use of a novel rat tracheal preparation, provide evidence that the epithelium modulates tracheal muscle response to carbachol stimulation through several mech- anisms. The epithelium acts as a diffusion barrier limit- ing access of the drug to the muscle. It may also actively influence muscle reactivity, possibly by the release of inhibitory factors. However, controversy exists regarding the existence as well as the importance of such factors. For example, by using cascade superfusion of guinea pig tracheal strips, Holroyde (11) did not find evidence for liberation of relaxing factor during stimulation with his- tamine. Conversely, Hay et al. (10) demonstrated a pro- duction of transferable relaxing factors by epithelium when challenging guinea pig tracheal preparations with antigen. An endothelium-derived factor that may cause relaxation of vascular smooth muscle has been shown to have a half-life of only a few seconds (15); a similar metabolic rate of the putative factor released from the airway epithelium could explain described experimental difficulties to demonstrate its transferability.
Finally the responses of existing neural pathways may be modified by intraepithelial amine-containing cells as well as by the action of mast cells and other inflammatory cells, if they are stimulated to release their mediators. We failed to demonstrate involvement of cyclooxygen- ase-dependent mediators, but there may be other meta- bolic pathways involved. Indeed, some intraepithelial nerve endings that are closely associated with intraepi- thelial amine-containing cells may be stimulated from the airway lumen and consequently cause an increased smooth muscle tone. The state of these neural pathways may also have been modified by mechanical removal of the epithelium in our model.
Whatever the mechanisms responsible for the in-
creased Tmax and sensitivity of the tracheal smooth muscle to carbachol in our model, our results clearly suggest that the tracheal epithelium plays an important role in the regulation of smooth muscle tone.
The new model presented may be useful in the future in determination of underlying mechanisms regulating airway smooth muscle tone.
Address for reprint requests: D. Pavlovic, Service de Pneumologie et Reanimation and INSERM U226, Hopital Beaujon, 100 Boulevard du General Leclerc, 92100 Clichy, France.
Received 12 December 1988; accepted in final form 2 August 1989.
REFERENCES
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
14.
15.
16.
ALLOTT, C. P., D. P. EVANS, AND P. W. MARCHALL. A model of irritant-induced bronchoconstriction in the spontaneously breath- ing guinea pig. Br. J. Pharmacol. 71: 165-168, 1980. BARNES, P. J., F. H. Cuss, AND J. B. PALMER. The effect of airway epithelium on smooth muscle contractility in bovine trachea. Br. J. Pharmacol. 86: 685-691, 1985. BUTLER, G. B., K. B. ADLER, J. N. EVANS, D. W. MORGAN, AND J. L. SZAREK. Modulation of rabbit airway smooth muscle respon- siveness by respiratory epithelium. Am. Reu. Respir. Dis. 135: 1099-1104,1987. DUNNILL, M. S. The pathology of asthma, with special reference to changes in bronchial mucosa. J. CZin. PathoZ. 13: 27-33, 1960. FABBRI, L. M., H. AIZAWA, S. E. ALPERT, E. H. WALTERS, P. M. 0. O’BYRNE, B. D. GOLD, J. A. NADEL, AND M. J. HOLTZMAN. Airway hyperresponsiveness and changes in cell counts in bron- choalveolar lavage after ozone exposure in dogs. Am. Reu. Respir. Dis. 129: 288-291, 1984. FLAVAHAN, N. A., L. L. AARHUS, T. J. RIMELE, AND P. M. VANHOUTTE. Respiratory epithelium inhibits bronchial smooth muscle tone. J. Appl. Physiol. 58: 834-838, 1985. FROSSARD, N., AND F. MULLER. Epithelial modulation of tracheal smooth muscle responses to antigenic stimulation. J. Appl. Physiol. 61: 1449-1456, 1986. GOLDEN, J. A., J. A. NADEL, AND H. A. BOUSHEY. Bronchial hyperirritability in healthy subjects after exposure to ozone. Am. Rev. Respir. Dis. 118: 287-294, 1978. HAY, D. W. P., S. G. FARMER, D. RAEBURN, V. A. ROBINSON, W. W. FLEMING, AND J. S. FEDAN. Airway epithelium modulates the reactivity of guinea-pig respiratory smooth muscle. Eur. J. Phar- macol. 129: 11-18, 1986. HAY, D. W. P., R. M. MUCCITELLI, D. L. HORSTEMEYER, V. A. WILSON, AND D. RAEBURN. Demonstration of the release of an epithelium-derived inhibitory factor from a novel preparation of guinea pig trachea. Eur. J. Pharmacol. 136: 247-250, 1987. HOLROYDE, M. C. The influence of epithelium on the responsive- ness of guinea pig isolated trachea. Br. J. Pharmacol. 87: 501-507, 1986. INOUE, H., S. HORIO, M. ICHINOSE, S. IDA, W. HIDA, T. TAKISH- IMA, K. OHWADA, AND M. HOMMA. Changes in bronchial reactivity to acetylcholine with type C influenza virus infection in dogs. Am. Rev. Respir. Dis. 133: 367-371, 1986. LAITINEN, L. A., M. HEINO, A. LAITINEN, T. KAVA, AND T. HAAHTELA. Damage of the airway epithelium and bronchial reac- tivity in patients with asthma. Am. Rev. Respir. Dis. 131: 599-606, 1985. NAYLOR, B. The shedding of the mucosa of the bronchial tree in asthma. Thorax 17: 69-72, 1962. VANHOUTTE, P. M. The endothelium-modulator of vascular smooth muscle tone. N. Engl. J. Med. 319: 512-513, 1988. WOOLCOCK, A. J., C. M. SALOME, AND K. YAN. The shape of the dose-response curve to histamine in asthmatic and normal subjects. Am. Rev. Respir. Dis. 130: 71-75, 1984.







Eur Respir J 1998; 11: 575–582DOI: 10.1183/09031936.98.11030575Printed in UK - all rights reserved
Copyright ©ERS Journals Ltd 1998European Respiratory Journal
ISSN 0903 - 1936
Effects of corticosteroids on epithelial structure and smoothmuscle function of rat trachea
D. Pavlovic, N. Viires, C. Zedda, M. Fournier, M. Aubier
aa
Several studies have shown that the most prominent fea-ture of bronchial asthma is an inflammatory process thataffects the airway mucosa. In this connection, corticoster-oids, which are widely used in the treatment of asthma fortheir anti-inflammatory properties, have proven their effect-iveness on airway hyperreactivity as well as providing symp-tomatic relief [1]. However, in order to control the mucosalinflammation of the airways, prolonged treatment withcorticosteroids is often required [2]. Long-term corticos-teroid administration has been shown to induce various sideeffects. Apart from the well-known glucocorticoid inducedmyopathy [3–5], long-term corticosteroid treatment hasbeen shown to induce pronounced atrophy of the gut anduterine smooth muscle [6]. Furthermore, corticosteroidscould affect connective tissue, inducing skin atrophy andmarked microvascular changes [7, 8].
Although patients with severe or unstable asthma oftenreceive high doses of corticosteroids, the effects of thesecompounds on normal bronchial tissue and airway smoothmuscle contractility are still poorly evaluated. Further-more, corticosteroids may also affect the function of air-way epithelium. Since there is experimental evidence invarious preparations of tracheal smooth muscle that theairway epithelium modulates contraction of the underly-ing smooth muscle in respect to various stimulating agents
[9], we also evaluated the effect of corticosteroids on thetracheal epithelium. This was performed by comparingepithelial and serosal application of a pharmacological ag-ent in preparations with and without epithelium. The pur-pose of this study was, therefore, to examine the effects ofhigh doses of corticosteroids on rat tracheal epitheliumand tracheal smooth muscle function.
Materials and methods
Animals
Male Sprague-Dawley rats (body weight 290–310 g) werestudied. Rats were separated into three groups and housedin individual cages. On day 0 steroid-treated animalsreceived hind limb intramuscular injections of Kenacort(triamcinolone acetonid; Squibb Laboratories, Neuilly-sur-Seine, France) 1.2 mg·kg-1·day-1. Since triamcinolone isequipotential to prednisolone, this dose would correspondto a low (small animal) anti-inflammatory dose of pred-nisolone [10, 11]. The animals were allowed food andwater ad libitum. Pair-fed animals (PF) (the control group)were given water ad libitum and the same amount of foodas the rats had received in the steroid-treated group. To
Effects of corticosteroids on epithelial structure and smooth muscle function of rat trachea.D. Pavlovic, N. Viires, C. Zedda, M. Fournier, M. Aubier. ERS Journals Ltd 1998.ABSTRACT: Corticosteroids, efficient drugs for the treatment of severe asthma, mayhave numerous side effects. We investigated the effects of 7 days of treatment with tri-amcinolone (1.2 mg·kg-1·day-1) on the epithelial structure, tracheal smooth musclecross-sectional area and contractility in the rat.
The corticosteroid-injected rats were compared to pair-fed, and pair-weighed animals.Histological studies were performed on transverse sections of glutaraldehyde-fixedtracheal blocks embedded in plastic. In the preparations taken from corticosteroid-injected, pair-fed and pair-weighed animals, pharmacological stimulation with single(10-3 M) or cumulative (10-8–10-3 M) concentrations of carbachol (in corticosteroid-injected and pair-fed animals), either inside (In) or outside (Out) of the tracheallumen, was performed and contractions of the tracheal smooth muscle were rec-orded.
We found that triamcinolone administration: 1) reduced the number of epithelialcells and the tracheal smooth muscle cross-sectional area; 2) induced a decrease inmaximal tension (Tmax (g); Out: 2.42±0.17, 1.03±0.1 in pair-fed and corticosteroid-injected, respectively; In: 2.55±0.16, 1.1±0.16, respectively) without affecting the sen-sitivity of the tracheal smooth muscle; and 3) reduced the time required to reach 50%Tmax in carbachol (In) preparations.
We conclude that the observed changes resulted from atrophy of tracheal smoothmuscle induced by undernutrition and atrophy of tracheal smooth muscle and tra-cheal epithelium induced by corticosteroid treatment.Eur Respir J 1998; 11: 575–582.
Unite INSERM 408, Faculté Xavier -Bichat, Paris, France.
Correspondence: M. AubierService de PneumologieHôpital Bichat46 rue Henri Huchard75018 ParisFranceFax: 33 1 40258818
Keywords: Carbacholcorticosteroidsratsmooth muscletrachea
Received: February 16 1996Accepted after revision July 30 1997

576 D. PAVLOVIC ET AL.
verify the effects of corticosteroid treatment on the epithe-lium and maximal tension (Tmax) and the rate of tensiondevelopment (time to Tmax) a third group of animals wasintroduced. Animals in this pair-weighed (PW) group rec-eived restricted amounts of food in order to achieve, bythe seventh day, the same body weight as the steroid-treated animals. All groups of animals received dailyintraperitoneal injections of 0.1 mL Bactrim (sulphameth-oxasole 400 mg and trimethoprim 80 mg per 5 mL; RocheLaboratories, Neuilly-sur-Seine, France), to prevent Pneu-mocystis carinii infection. On the seventh day of treatmentthe animals were sacrificed for the experiments.
Study design
The steroid-treated rats were compared to PF and PWanimals. Histological studies were performed on transversesections of glutaraldehyde-fixed, plastic-embedded trach-eal blocks. Pharmacological stimulation with single (10-3
M) or cumulative (10-8–10-3 M) concentrations of carba-chol (in steroid-treated and PF animals), either inside (In)or outside (Out) the tracheal lumen, were performed in thepreparations of steroid-treated, PF and PW animals, andcontractions of the tracheal smooth muscle were recorded.
Methods
The method used to prepare tracheal smooth musclehas been described previously [10, 11]. Briefly, tracheaswere taken from rats after they had been stunned by ablow on the head and quickly exsanguinated. The tracheascut from the cricoid cartilage to the bifurcation of twomain bronchi, were immersed in Krebs solution (mM:NaCl 137, KCl4, MgCl2 11, KH2PO4 1, NaHCO3 12, CaCl22, Glucose 6.5) and cleaned from all surrounding tissue.The upper 10–12 cartilaginous rings of each trachea wereused for the study of muscle contractility. From threeentirely separate groups of animals (PF, PW, and steroid-treated), tracheas were dissected to be used for subsequenthistological examination.
Histological examination. Tracheal segments (superior twotracheal rings) were fixed in 2.5% glutaraldehyde (TAAB,Aldermaston, UK) in 0.14 cacodylate buffer pH 7.3 at 4°Cfor 2 h. Tissue blocks were dehydrated in an alcoholseries, cleared in propylene oxide and embedded in Epon(Fluka Chemica-biochemica, Buchs, Switzerland). Onemicrometre semithin transverse sections were cut on aLKB ultrotome III (LKB, Broma, Sweden), stained withtoluidine blue at pH 4 and mounted. The preparationsfrom the three groups of animals (steroid-treated, n=8, PFn=4, and PW n=5) that had undergone the above-des-cribed corticosteroid or dietary protocols were examinedmicroscopically. Two methods were used to estimate changesof the epithelium and smooth muscle: microscopy withplanimetry and computer-assisted image analysis.
Computer-assisted image analysis. The preparations wereexamined by a light microscope (BH2; Olympus, Tokyo,Japan; SCOP S. A. Olympus, Ringis, France) linked toa video camera (COHU, San Diego, CA, USA; Clara Vis-ion, Orsey, France) and to a computer equipped with an
image analysis program (Perfect image 2.01; Clara Vis-ion, Orsey, France). Observations were performed as fol-lows.
Epithelium. Four fields, equidistant from each other, oneoverlying the middle of the muscle region, and the otherthree spaced at about 45° around the tracheal ring, wererandomly selected from each section by the examiner,who did not know which group of preparations were beinganalysed. In each field the number of epithelial cells (Nc)was counted; epithelial cell height (Hc) and the corre-sponding basement membrane length (Lm) were traced onthe screen and the distances computed to the lengths. Theratio Nc/Lm was calculated for each field and one meanvalue is reported for each section.
Smooth muscle surface. The limit of the total area of thetracheal smooth muscle for the given section was drawnfrom the computer screen and the smooth muscle cross-sectional area (SM) was automatically computed. The sec-tion of the basement membrane that corresponded to thesmooth muscle was also traced and computed to the length(Lm). The ratio SM/Lm was calculated for each section.
Tracheal smooth muscle (TSM) contractility. Figure 1shows a schematic representation of the experimental set-up, which represented an improved modification of the
Hook to transducer
In
In
Out
Tracheal segmentHook
Hook
Fig. 1. – Schematic representation of the experimental apparatus. Fluidfreely circulates with the lumen of the tracheal segment (In: epithelialside) or around the exterior (Out: serosal side) of the tracheal segment.These inner and outer perfusion solutions are maintained at 37°C, bub-bled with 95% O2 and 5% CO2, and a constant flow rate of 2 mL·min-1 ismaintained.

EFFECTS OF CORTICOSTEROIDS ON RAT TRACHEA 577
apparatus described previously [11]. Under microscopiccontrol, two stainless steel hooks were passed through thetracheal wall around two adjacent cartilaginous ringsclose to the points of muscle insertion. The tracheal seg-ment was then longitudinally connected to steel tubes andtightened firmly with silk thread. The lower hook wasattached below and served as a fixed point. The intubatedtrachea was then put into the organ bath and the upperhook connected to a force transducer (UC2; Gould Inc,Cleveland, OH, USA) which could be moved with a micro-manipulator (Prior Scientific Instruments Ltd, Herts, UK)along a vertical axis. Changes in tension were registeredon a paper recorder (AT 550; Gould Inc.) connected to thetransducer. The organ bath used for the experiments per-mitted independent circulation of fluid within the lumen ofthe tracheal segment (In: epithelial side) or around theexterior (Out: serosal side). The Krebs solution (37°C, pH= 7.4, gassed with 95% O2 and 5% CO2) was perfused at aconstant flow rate (2 mL·min-1). A peristaltic pump (WatsonMarlow 5025, Falmouth, Cornwall, UK) was used to main-tain outer perfusion. Tracheal epithelium was removedmechanically from one group of tracheas taken from corti-costeroid-treated animals (-Ep group) by gently rubbingthe interior of the tracheal preparation with a moistenedcotton-wrapped metal stick, as described previously [11].To verify successful epithelium removal at the end of theexperiment each steroid-treated -EP preparation was exam-ined histologicaly, as described previously [11].
For tests of tracheal smooth muscle contractility, thetracheas were mounted on the apparatus (as described ab-ove) and, after a period of stabilisation (45–60 min), theywere transversally stretched to an optimal muscle length,established in a preliminary set of experiments.
Length-tension relationship. Preliminary assays were per-formed to determine optimal stretch of the muscle. Theseexperiments could not be performed on the cannulated tra-cheal segments since pulling the hooks apart would, byincreasing the distance between hooks, stretch not onlythe two hooked cartilaginous rings but also the adjacent tra-cheal rings and corresponding muscle fibres which would,in turn, contribute to the maximal contraction registered(fig. 2a). The contribution of the muscle fibres adjacent tothose between the two hooks was found, in a separate set ofexperiments, performed with canulated preparations withthe muscle between hooks sectioned, but the adjacentmuscle left intact (fig. 2b), to be 5–8% of the maximaltension obtained at optimal length. To obtain contractionsof the muscle just between the two cartilaginous ringswithout the contribution of adjacent muscle fibres, mount-ed cannulated tracheal preparations were sectioned trans-versely as close as possible to the insertions of the hooks(fig. 2c). In a preliminary set of experiments, we foundthat transverse sectioning of unstretched dog trachea (tra-chea taken out of thorax, n=3, results not shown) did notby itself change the distance between cartilaginous ends,meaning that muscle resting length did not change. Weassumed that rat trachea would behave in the same man-ner. This permitted us to perform experiments on trachealrings consisting of only two tracheal cartilages. Experi-ments for determining the optimal length-tension rela-tionship consisted of maximal induced contractions with
10-6 M carbachol at a given muscle length (i.e. given pas-sive tension). Following the relaxation a new contractionwas elicited with the same agent, but at different musclelengths.
Experimental protocol. Appropriate amounts of carbachol(Sigma Chimie S.a.r.l., St Quentin Fallavier, France) werediluted in small volumes (100 µL) of distilled water andadded to the inner or outer Krebs solution reservoirs toobtain the required final concentrations. In the first setof experiments the effects of a single supramaximal con-centration of carbachol (10-3 M) on tracheal muscle Tmaxand rate of tension development (time to Tmax) were ex-amined. The agent was left in contact with the preparationuntil the response reached a plateau. The preparation wasthen perfused with Krebs solution and allowed to relax
a)
b)
c)
HookIntact
tracheal segment
Longitudinal section
Transverse section
Fig. 2. – Positions of the hooks on the tracheal wall showing distensionof the trachea when the hooks are pulled away in a) intact trachea, b)when longitudinally sectioning the tracheal smooth muscle between thehooks, and c) when transversally sectioning the tracheal smooth musclefor determination of the length-tension relationship.
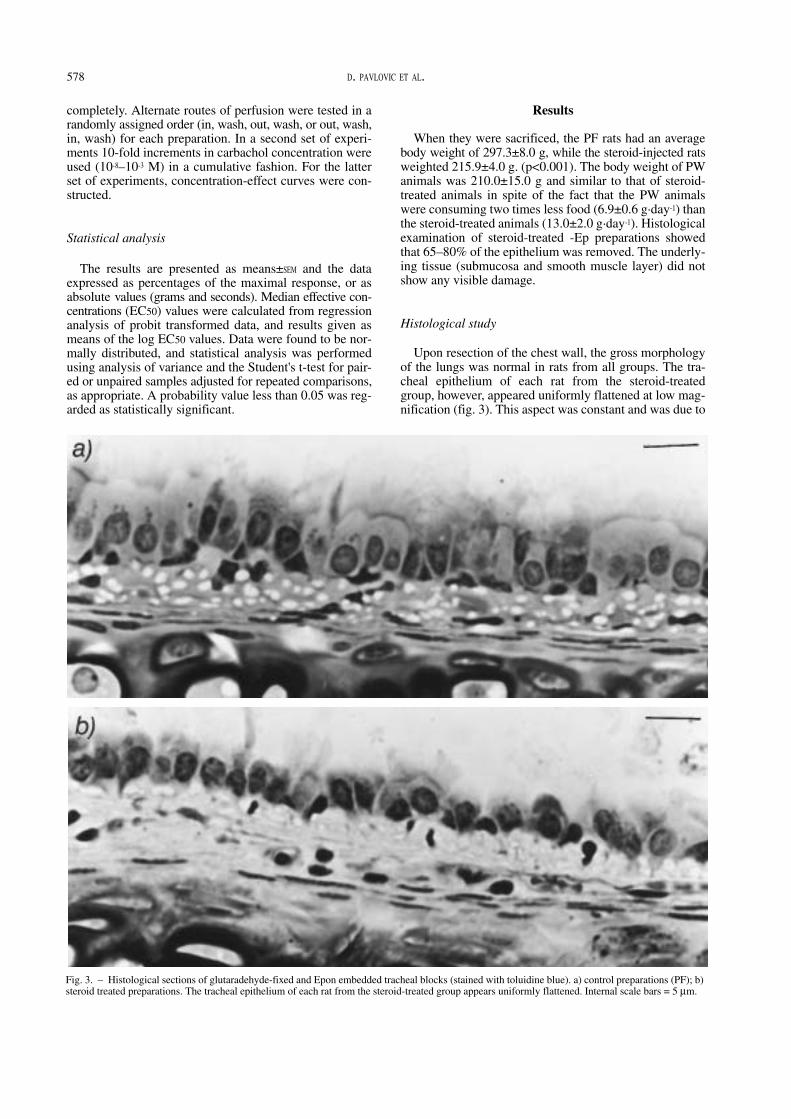
578 D. PAVLOVIC ET AL.
completely. Alternate routes of perfusion were tested in arandomly assigned order (in, wash, out, wash, or out, wash,in, wash) for each preparation. In a second set of experi-ments 10-fold increments in carbachol concentration wereused (10-8–10-3 M) in a cumulative fashion. For the latterset of experiments, concentration-effect curves were con-structed.
Statistical analysis
The results are presented as means±SEM and the dataexpressed as percentages of the maximal response, or asabsolute values (grams and seconds). Median effective con-centrations (EC50) values were calculated from regressionanalysis of probit transformed data, and results given asmeans of the log EC50 values. Data were found to be nor-mally distributed, and statistical analysis was performedusing analysis of variance and the Student's t-test for pair-ed or unpaired samples adjusted for repeated comparisons,as appropriate. A probability value less than 0.05 was reg-arded as statistically significant.
Results
When they were sacrificed, the PF rats had an averagebody weight of 297.3±8.0 g, while the steroid-injected ratsweighted 215.9±4.0 g. (p<0.001). The body weight of PWanimals was 210.0±15.0 g and similar to that of steroid-treated animals in spite of the fact that the PW animalswere consuming two times less food (6.9±0.6 g·day-1) thanthe steroid-treated animals (13.0±2.0 g·day-1). Histologicalexamination of steroid-treated -Ep preparations showedthat 65–80% of the epithelium was removed. The underly-ing tissue (submucosa and smooth muscle layer) did notshow any visible damage.
Histological study
Upon resection of the chest wall, the gross morphologyof the lungs was normal in rats from all groups. The tra-cheal epithelium of each rat from the steroid-treatedgroup, however, appeared uniformly flattened at low mag-nification (fig. 3). This aspect was constant and was due to
Fig. 3. – Histological sections of glutaradehyde-fixed and Epon embedded tracheal blocks (stained with toluidine blue). a) control preparations (PF); b)steroid treated preparations. The tracheal epithelium of each rat from the steroid-treated group appears uniformly flattened. Internal scale bars = 5 µm.

EFFECTS OF CORTICOSTEROIDS ON RAT TRACHEA 579
a change in the epithelial structure, which appeared mon-ostratified. No cell lysis, local oedema or epithelial shed-ding was observed and the usual cell subtypes, such asciliated or goblet cells, were present. The basement mem-brane appeared normal and inflammatory cells occasion-ally infiltrated the lamina propria to the same extent as inPF animals. Other structures of the tracheal wall had nopeculiar features.
Computer assisted image analysis (table 1) showed thatfor the comparable length of basement membrane (Lm) tra-cheal sections from one steroid-treated group (n=8) showeda diminution in Nc and Hc as compared to the PF group(n=4) (Nc/Lm ratio=790.93±20.1 versus 984.05± 29.3;p<0.001) and smaller (although nonsignificant) total sur-face area of the tracheal SM, and a significantly smaller
Tmax
%
100
50
0
a)
●
●
●
●
●
●●
●
●
Tmax
%
0
0 1 2 3 4 5 6∆L 140 µm·division-1
b)
100
50●
●
●
●
●
●
●
●
●
●
Fig. 4. – Length-tension relationship of the rat tracheal smooth musclebetween two cartilaginous rings taken from transversally semisectionedtracheas in a) controls and b) corticosteroid-treated animals. ýL: lengthchanges achieved by pulling apart the hooks inserted close to the muscleinsertions in the cartilaginous rings; Tmax: maximal tension followingstimulation with carbachol (106 M). ❍: active tension; ●: passive ten-sion. Values are presented as mean±SEM.
Table 1. – Histology of rat trachea
Epithelium MuscleLm Nc Hc Nc/Lm SM LM SM/LM
PF(n=4)PW(n=5)ST(n=8)
PF/PWPF/STPW/ST
0.0396±0.00273
0.0389±0.00533
0.0385±0.00046
NS
NS
NS
38.87±2.598736.31±1.918930.53±0.67
NS
<0.005<0.05
0.00575±0.00009
0.00512±0.00033
0.00393±0.00026
NS
<0.001<0.05
984.05±29.3931.14±27.1790.93±20.1
NS
<0.001<0.005
0.1825±0.022
0.0931±0.022
0.054±0.046
NS
NS
NS
0.261±0.01385
0.201±0.01385
0.1323±0.00951
<0.05<0.001<0.005
0.69±0.05
0.46±0.03
0.40±0.02
<0.01<0.001
NS
Values are presented as mean±SEM. Tracheal cross-sections were taken from the controls (pair-fed(PF) and pair-weighted (PW)) and steroid-treated (ST) animals. Lm: epithelium length (length ofthe basement membrane); Nc: number of epithelium cells; Hc: height of the epithelium; SM:total smooth muscle area; LM: basement membrane length in the region of the smooth muscle.
Tmax
g,
In
4
3
2
1
0
a)
▼▼▼▼▼▼▼▼▼▼▼▼▼▼▼▼▼▼
▼
▼
▼
▼
▼
▼▼
▼▼▼
▼
▼
Tmax
g,
In
4
3
2
1
00 1 2 3 4
Tmax g, Out
b)
●
●●●●
●
●
●
Fig. 5. – Individual values of maximal tension (Tmax) recorded follow-ing stimulation with (10-3 M) carbachol solution from epithelial (In) andserosal (Out) sides, in a) pair-fed (▼), and pair-weighted ( ) prepara-tions, and b) taken from triamcinolone pretreated rats, with epitheliumintact (●) and without epithelium (❍) preparations. Symbols show meanvalues with error bars showing SEM.
ý

580 D. PAVLOVIC ET AL.
ratio of muscle surface/length of the basement membranewhich corresponded to the muscle length (SM/LM =0.4±0.02 versus 0.69±0.05; p<0.001). The values obtained forthe PW group (n=5) were between those of steroid-treatedand PF group.
Tracheal smooth muscle contractility
We found that the length-tension relationship did notdiffer significantly between control (PF) and steroid-treat-ed groups (fig. 4). As shown in figure 5 and in table 2,supramaximal concentrations of carbachol (10-3 M) alter-nately perfused In and Out, gave superimposable maximalchanges in tension for a given preparation. However, thepreparations taken from steroid-treated (n=8), steroid-treat-ed -Ep (n=6), PW (n=8) and PW -Ep (n=5) groups of ani-mals were found to generate considerably less Tmax oncarbachol stimulation than did the PF (n=11) and PF -Ep(n=5) groups (p<0.005), maximal tension developed as aconsequence of the preparations of the PW and PW -Epgroups being slightly superior to those of the steroid-treat-ed and steroid-treated -Ep groups (p<0.05). Conversely, treat-ment with corticosteroids did not seem to influence thesensitivity of the preparations to cumulative carbachol stim-ulation (second set of experiments). Indeed, mean EC50values calculated from dose-response curves obtained inthe steroid-treated group (n=8) did not differ from those of
the PF group (n=8), PW group (n=8), or steroid-treated-Ep group (n=6), and were not subject to significant changeupon inner or outer carbachol stimulation (table 3). Never-theless, we found that preparations without epithelium, PF-Ep (n=5) and PW -Ep (n=6), were more sensitive to car-bachol stimulation (Out) as compared to the correspond-ing control preparations with epithelium (p<0.05 and 0.001,
Table 2. – Maximal tension (Tmax) in rat tracheal prep-arations
Tmax gOut In
PFPF (-Ep)PWPW (-Ep)STST (-Ep)
2.42±0.172.37±0.221.76±0.191.66±0.251.03±0.101.06±0.13
2.55±0.162.21±0.171.68±0.241.72±0.271.1±0.161.01±0.14
Tmax following supramaximal stimulation with carbachol (10-3
M) from the serosal (Out) or epithelial (In) side of rat trachea incontrol preparations taken from pair-fed (PF) or pair-weighted(PW) animals or from corticosteroid-treated (ST) animals, withintact epithelium or without epithelium (-Ep). Values are pres-ented as mean±SEM.
Table 3. – Log median effective concentrations (EC50) ofcarbachol in pair-fed (PF) or pair-weighed (PW) controland steroid-treated rats
Out In
PFPF (-Ep)PWPW (-Ep)Steroid-treatedSteroid-treated (-Ep)
5.89±0.076.15±0.115.91±0.036.12±0.055.77±0.065.80±0.08
5.65±0.175.89±0.25.73±0.075.92±0.095.43±0.195.78±0.02
Values are geometric means (±SEM) of log EC50 values obtainedin preparations taken from controls (PF and PW), or triamci-nolone pretreated rats with intact epithelium or without epithe-lium (-Ep) following stimulation from the epithelial (In) orserosal side (Out) with cumulative concentrations of carbachol10-8–10-3 M solution (all differences of comparable responses arestatistically nonsignificant except from Out: PF, -Ep and PW, -Ep versus controls, p<0.05 and <0.001, respectively.
100
80
50
0Tm
ax %
a) ●
●
●●
●
●
100
80
50
0
Tmax
%
b)
●
●
●
●●
●
100
80
50
0
Tmax
%
c)
0 50 100 300 500 700Time s
●
●●
●●
●
Fig. 6. – The rate of tension development (time to Tmax) (mean±SEM)recorded during supramaximal stimulation with carbachol solution (10-3
M) either from the epithelial side (In; ❍) or serosal side (Out; ●), withepithelium (+Ep; ) and without epithelium (-Ep; ). The time toreach 50% (and 80% in PF) of Tmax was longer during inner (In) thanduring outer perfusion (Out) with 10-3 M carbachol in control prepara-tions (panel a and b, PF and PW, +Ep versus -Ep, In versus Out,p<0.001) but not in the preparations taken from corticosteroid treatedanimals (panel c, steroid-treated and steroid-treated -Ep). Tmax: maxi-mum tension.

EFFECTS OF CORTICOSTEROIDS ON RAT TRACHEA 581
respectively). Maximal response of the steroid-treated pre-parations (with and without epithelium) to stimulation withcumulative concentrations of carbachol was again, as inthe first set of experiments, inferior to the maximal res-ponse of PF and PW preparations with or without epithe-lium (results not shown).
The time course of tension development (time to Tmax)during carbachol (10-3 M) stimulation was different in thesteroid-treated group as compared with PF and PW ani-mals (fig. 6). Indeed, in the PF and PW groups, the timerequired to reach 50 and 80% of Tmax was clearly influ-enced by the route of perfusion: the time to reach 50%(and 80% in PF) of Tmax was significantly longer (p<0.001)during inner than during outer perfusion with carbachol(fig. 6a and b). On the other hand, in the steroid-treatedand steroid-treated -Ep group and in control preparationsPF -Ep or PW -Ep (results not shown), no difference in timeto Tmax was observed in response to inner as compared toouter perfusion with carbachol (fig. 6c).
Discussion
The results of this study demonstrate that corticosteroidadministration reduces rat tracheal muscle cross-sectionalarea, as well as the number of tracheal epithelial cells, anddecreases the maximal tension elicited by carbachol stim-ulation of rat tracheal smooth muscle.
The steroid-treated animals lost considerable weight dur-ing the study period. This could only partly be explainedby a reduced food intake, since PF animals did not loseweight, and PW animals (which ate much less than thesteroid-treated group) showed the same weight loss. Thedose of triamcinolone used corresponded to an anti-inflam-matory animal dose. Triamcinolone is equipotent to pred-nisolone [10] and the recommended anti-inflammatorydose of prednisolone for small animals is 1–5 mg·kg-1
[11]. However, our finding of substantial weight loss 1.2mg·kg-1·day-1 in triamcinolone-treated animals suggests thatthe dose used was probably high. The excessive weightloss observed in steroid-treated animals could be due toimpaired food utilization and/or increased catabolism.There is considerable evidence demonstrating that corti-costeroid treatment produces skeletal muscle atrophy anddecreases force generated by the muscle. The fluorinatedcorticosteroids (triamcinolone, betamethasone and dexa-methasone) appear most potent in this regard [12]. Histo-logical studies have shown an atrophy of type 2 musclefibres (fast glycolytic fibres) with little change in type 1fibres (slow fibres). The atrophy was attributed primarilyto decreased protein synthesis [5, 13, 14] and/or increasedprotein degradation [15, 16]. Tracheal smooth musclecould have also been affected in a similar manner by cor-ticosteroid treatment [6]. Recent findings that dexametha-sone can inhibit mitogen-induced proliferation of humancultured airway smooth muscle indicate that trachealsmooth muscle atrophy could be expected following treat-ment with corticosteroids [17].
Our findings strongly suggest that this was the case. In-deed, the histological study (planimetry of tracheal cross-sections) showed that tracheal smooth muscle mass wasdiminished in the corticosteroid-treated animals. This smoothmuscle atrophy may, at least in part, be responsible for thedecrease in maximal tension developed during carbachol
stimulation in corticosteroid-treated animals, as comparedto the controls.
Nevertheless, other structures such as cartilage, mucosaand connective tissue could also have been affected bycorticosteroids. Indeed, it has been shown that corticoster-oid treatment decreases connective tissue synthesis [18,19]. This latter effect accompanied by smooth muscle at-rophy could have produced changes in the length-tensionrelationship of the tracheal smooth muscle, explaining thediminished maximal tension that we observed in corti-costeroid-treated animals. Other investigators who used thesame experimental model to examine only the sensitivityof the preparation to various agents did not routinely per-form experiments to determine length-tension relationships,probably assuming that it was relatively unimportantwhen examining sensitivity [20–22]. However, if the rest-ing pretension was determined arbitrarily, maximal tensionreported in some of those experiments [20, 21] would beunreliable, since it is strongly influenced by the smoothmuscle length. A quite elaborate procedure of determiningthe length-tension relationship was important in our ex-periments since we examined, in addition to the sensitiv-ity, the maximal tension developed. In these experimentswe found that tracheal smooth muscle preparations fromboth PF and steroid-treated animals, had the same length-tension relationship.
Concomitantly to tracheal smooth muscle atrophy, adirect depressant effect of corticosteroids on tracheal mus-cle contractility cannot be ruled out. Our finding of adiminished smooth muscle cross-sectional area and Tmaxin PW animals indicates that undernutrition was partlyresponsible for a loss of force of tracheal smooth muscle.However, the fact that Tmax was lower in the steroid-treated group, as compared to the PW animals implies thatcorticosteroid treatment induces tracheal smooth muscleweakness by some other additional mechanism. Indeed,corticosteroids may have affected cell membrane excita-bility or smooth muscle contractility. There is evidence,for example, that in striated muscle, treatment with corti-costeroids decreases calcium uptake by the sarcoplasmaticreticulum [23] which, in turn, could result in decreasedforce generation. We suggest, although with some reserve,that this mechanism may explain the decrease in maxi-mal tension observed in preparations taken from cortico-steroid-treated animals. On the other hand it has beenrepeatedly demonstrated that mechanical removal of theepithelium renders bronchial smooth muscle more sensi-tive to various agents. An epithelium-derived inhibitoryfactor (EpDRF) has been proposed to explain these effects.In steroid-treated -Ep preparations we did not observe anychange in the sensitivity to carbachol stimulation. How-ever, if corticosteroid treatment were to depress produc-tion of EpDRF, treated preparations (steroid-treated orsteroid-treated -Ep) would have been more sensitive to thecarbachol stimulation and EC50 would have increased, butthis was not the case. Questions that remain open are:whether inhibitory effects of putative EpDRF are too weakand masked by overly strong corticosteroid depressionof tracheal smooth muscle contractility; is the experiment-al model used sufficiently sensitive; and, finally, whetherexistence of such an inhibitory agent is a valuable hypo-thesis.
Using the same experimental model, we have previous-ly shown that the time to reach peak tension (the rate of

582 D. PAVLOVIC ET AL.
tension development) of tracheal smooth muscle duringinterior stimulation with carbachol was longer when theepithelium was intact than in epithelium-denuded prepara-tions [11]. This phenomenon was abolished in corticoster-oid-treated animals. The reduced time to peak tensionobserved during inside stimulation in the corticosteroidgroup with intact epithelium indicates a dysfunction of thetracheal epithelium, suggesting that its properties as a dif-fusion barrier were greatly reduced. This is supported bythe results of histological examination of whole trachealcross sections. Indeed, the decrease in the number of thetracheal epithelial cells observed may be considered as acorticosteroid-induced atrophy. These structural changesmay have been accompanied by important functional im-pairment of the tracheal epithelium in corticosteroid-treated animals. Observed changes in the rate of tensiondevelopment in corticosteroid-treated animals which was,as also shown previously [11], similar to the one obtainedin preparations of PF and PW denuded of epithelium,strongly supports this hypothesis.
In conclusion, we have demonstrated that corticosteroidtreatment in rats produces significant decrease in the res-ponsiveness of tracheal smooth muscle to carbachol stim-ulation, which could be explained, at least partially, byan atrophy of tracheal smooth muscle. Furthermore, weobserved pronounced atrophy of the tracheal epithelium,which was accompanied by its significant dysfunction. Thesefindings may be important for better understanding of themechanism of action, as well as the possible side effects,of corticosteroids in patients with asthma.
References
1. Peake MD, Cayton RM, Howard P. Triamcinolone incorticosteroid resistant asthma. Br J Dis Chest 1979; 73:39–45.
2. Rebuck AS, Read J. Assessment and management ofsevere asthma. Am J Med 1971; 51: 788–798.
3. Bunim JJ, Ziff M, McEven C. Evaluation of prolongedcortisone therapy in rheumatoid arthritis: 4 year study.Am J Med 1955; 18: 27–40.
4. Duvenci J, Chodosh S, Segal MS. Dexamethasone ther-apy in bronchial asthma. Ann Allergy 1959; 17: 695–700.
5. Goldberg AL, Tischler M, DeMartino G, Griffith G. Nor-mal regulation of protein degradation and synthesis inskeletal muscle. Fed Proc 1980; 39: 31–36.
6. Kelly FJ, Goldspink DF. The differing responses of fourmuscle types to dexamethasone treatment in the rat. Bio-chem J 1982; 208: 147–151.
7. MacAllen M. Long term side effects of corticosteroids.Respiration 1970; 27 (Suppl.): 250–259.
8. Smyllie HC, Connolly CK. Incidence of serious compli-cations of corticosteroid therapy in respiratory disease. Aretrospective survey of patients in the Brompton hospital.Thorax 1968; 23: 571–581.
9. Pavlovic D, Fournier M, Aubier M, Pariente R. Epithelialversus saerosal stimulation of tracheal muscle: role ofepithelium. J Appl Physiol 1989; 67: 2522–2526.
10. Rogers HJ, Spector RG, Trounce JR. A Textbook of Clin-ical Pharmacology. London, Hodder and Stoughton, 1981;p.596.
11. Zaslow IM. Veterinary Trauma and Critical Care. Phila-delphia, Lea and Fabiger, 1984; p. 329.
12. Askari AP, Vignos J, Moskowitz RW. Steroid myopathyin connective tissue diseases. Am J Med 1976; 61: 485–492.
13. Rannels SR, Rannels DE, Pett AE, Jefferson LS. Gluco-corticoid effect on peptide-chain initiation in skeletal mus-cle and heart. Am J Physiol 1978; 235: E 134–139.
14. Rannels SR, Jefferson LS. Effects of glucocorticoids onmuscle protein turnover in perfused rat hemicorpus. Am JPhysiol 1980; 238: E 564–572.
15. Clark AF, Vignos PJ. Experimental corticosteroid myopa-thy: effect on myofibrilar ATPase activity and proteindegradation. Muscle Nerve 1979; 2: 265–273.
16. Mayer M, Rosen F. Interaction of glucocorticoids andandrogens with skeletal muscle. Metabolism 1977; 26:937–962.
17. Fernandes D, Tomlinson PR, Harris T, Stewart AG. Anti-inflammatory steroids inhibit mitogen-induced prolifera-tion of human cultured airway smooth muscle. Am J RespirCrit Care Med 1995; 4: A47.
18. Keeley FW, Johnson DJ. Age differences in the effect ofhydrocortisone on the synthesis of insoluble elastin inaortic tissue of growing chicks. Conn Tissue Res 1987;16: 259–268.
19. Monthorpe R. Glucocorticosteroid effect on vascular con-nective tissue. Acta Endocrin 1983; 259 (Suppl. ): 1–40.
20. Folkerts GH, Van der Linde J, Nijkamp FP. Virus-inducedairway hyperresponsivness in guinea-pigs is related to adeficiency in nitric oxide. J Clin Invest 1995; 95: 26–30.
21. Nijkamp FP, Van der Linde HJ, Folkerts G. Nitric oxidesynthesis inhibitors induce airway hyperresponsiveness inthe guinea-pig in vivo and in vitro. Am Rev Respir Dis1993; 148: 727–734.
22. Small RC, Good DM, Dixon JS, Kennedy I. The effectsof epithelium removal on the actions of cholinomimeticdrugs in opened segments and perfused tubular prepara-tions of guinea-pig trachea. Br J Pharmacol 1990; 100:516–522.
23. Shoji S, Takagi A, Sugita H, Toyokura Y. Disfunction ofsarcoplasmatic reticulum in rabbit and human steroidmyopathy. Exp Neurol 1979; 51: 304–309.

Clinical and Experimental Pharmacology and Physiology (2008) 35, 663–669 doi: 10.1111/j.1440-1681.2007.04857.x
Blackwell Publishing AsiaOriginal Articless-Carboxymethylcysteine and airwaysD Pavlovic et al.s-CARBOXYMETHYLCYSTEINE INHIBITS CARBACHOL-INDUCED CONSTRICTION OF EPITHELIUM-DENUDED RAT AND HUMAN
AIRWAY PREPARATIONS
Dragan Pavlovic,* Helge Frieling,*§ Taras Usichenko,* Vladimir Nedeljkov,† Thais Nafissi,‡ Christian Lehmann,* Michel Aubier‡ and Michael Wendt*
*Department of Anaesthesiology and Intensive Care Medicine, Ernst-Moritz-Arndt University, Greifswald, Germany, †Institute for Pathophysiology, Medical Faculty, Belgrade University, Belgrade, Serbia and
‡INSERM U408, Faculte Xavier-Bichat, Universite Paris 7, Paris, France
SUMMARY
1. The effects of S-carboxymethyl-L-cysteine (S-CMC), eitheradministered orally to rats or incubated with tissue preparationsfrom rats and humans, on isometric contractions of trachealsmooth muscle were investigated in the present study using animproved in vitro model of tracheal tube or ring preparations.The involvement of the tracheal epithelium in the observedeffects was also investigated.
2. The experimental model permitted selective perfusion ofthe airway tube, luminal-IN or serosal–OUT, and measurementof airway smooth muscle contraction or relaxation in preparationswith (+) or without (–) epithelium (Ep), excluding direct effectsof airway mucus.
3. We found that oral pretreatment of rats with S-CMC(mixed with water; 200 mg/kg per day for 2 weeks), but not shortpre-incubation of preparations in vitro (10–3 mol/L S-CMC for1 h), diminished the sensitivity of –Ep preparations to carbacholcompared with controls (EC50 (–log10 mol/L) values: 5.5 ± 0.1 vs5.8 ± 0.1, respectively, for IN perfusion (P < 0.005); 5.6 ± 0.1 vs5.9 ± 0.1, respectively, for OUT perfusion (P < 0.005)), whereasthe sensitivity of preparations to aminophylline was not affected.Normal sensitivity to carbachol stimulation was re-establishedif preparations were pre-incubated with capsaicin.
4. It was also found that longer pre-incubation (4 h) of ring-preparations of human bronchus with S-CMC (10–5 mol/L) invitro resulted in a diminished response to carbachol stimulation.
5. In conclusion, S-CMC had small inhibitory effects on thesensitivity of rat and human airway smooth muscle to carbachol,particularly in endothelium-denuded preparations. Whether theepithelium was responding to S-CMC by producing somecontracting factor(s) requires further investigation.
Key words: epithelium, rat trachea, s-carboxymethylcysteine,tracheal ring, tracheal tube.
INTRODUCTION
The airway epithelium, which separates inhaled gas from theunderlying airway tissue, forms an important barrier between livingstructures and the environment. Preservation of the normal functionof the bronchial epithelium is a precondition for maintenance of aneffective air–tissue barrier. That barrier may be disturbed in airwayinflammation and asthma. One objective of our earlier and thepresent study has been to test the hypothesis that the bronchialepithelium, either as a diffusion barrier or by actively secreting yetnot well defined mediators (prostaglandins, leukotrienes or othermediators), can modulate bronchial smooth muscle sensitivity tovarious stimulating agents.1–3 Experimentally, we use a system ofperfused tracheal tubes because a tube airway preparation allowsselective pharmacological stimulation of the epithelial or serosalside of the airway in vitro. In the present study, the model describedearlier2,4,5 was improved so that it allowed investigation of contractionsof the entire airway tube first and then as a ring preparation.
We centred our study on the mucolytic agent s-carboxymethylcysteine(s-carboxymethyl-l-cysteine; S-CMC; carbocysteine), used acrossEurope. Improving mucociliary transport is one of the therapeuticapproaches in asthma therapy, in which, in addition to anti-inflammatory agents,6,7 mucolytic agents are occasionally used.8,9
Indeed, beneficial effects of S-CMC for chronic obstructive bronchitishave been demonstrated in a large multicentre controlled trail.10
Nevertheless, the mechanism of action of S-CMC remains unclear.It was found that S-CMC could enhance the production of sialomucinsat the expense of fucomucins,11,12 which may result in an improvementin mucociliary transport. Several other findings have implicatedeither reduced or increased sputum viscosity13–16 or even an anti-inflammatory property17–19 that may be related to increasedsialoglycoprotein secretion as a mechanism of action of S-CMC.
Nevertheless, asthmatic subjects often have airway hyperrespon-siveness that is accompanied by secretion of thick mucus and variousdegrees of inflammation. Recent evidence indicating that S-CMCmay indirectly favour smooth muscle relaxation18,20,21 furtherjustified the aim of the present study, which was to test the effectsof S-CMC on airway smooth muscle (ASM) responsiveness. Moreprecisely, we wanted to examine the effect of S-CMC on bronchialepithelium function and/or tracheal smooth muscle in healthy animalsand its effect on the isometric contraction of bronchial smoothmuscle in cannulated preparations or ring preparations. Thesecond objective of the study was to construct and test a slightly
Correspondence: Dragan Pavlovic, Klinik und Poliklinik für Anästhesieund Intensivmedizin, Ernst-Moritz-Arndt Universität, Friedrich-Loeffler Str.23B, 17487 Greifswald, Germany. Email: [email protected]
§Present address: University Hospital Erlangen, Department of Psychiatryand Psychotherapy, Erlangen, Germany.
Received 16 January 2007; revision 20 November 2007; accepted 21November 2007.
© 2008 The Authors Journal compilation © 2008 Blackwell Publishing Asia Pty Ltd

664 D Pavlovic et al.
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Asia Pty Ltd
modified in vitro apparatus that can be used with an airway tubeor, alternatively, an airway ring preparation.
METHODS
Animal preparation
Experiments were performed on tracheas taken from male Sprague-Dawleyrats, weighing 390–420 g. Animal treatment and experimental procedureswere in accordance with the recommendations of INSERM and with the localInstructions for Animal Care of Greifswald University. All animals werehoused in individual cages and received water and food ad libitum. Thepretreated group of animals (S-CMCpr) received S-CMC mixed with water(200 mg/kg per day) for 2 weeks. During the 3rd week, rats were killed forthe experiments by stunning and quick exsanguination. The tracheas wereimmersed in modified Krebs’–Henseleit solution (KH; composition (inmmol/L): NaCl 113; KCl 4.8; MgCl2·6H2O 1.3; KH2PO4 1.2; NaHCO3 25;CaCl2 2.5; glucose 5.7) and cleaned from surrounding tissue.
Perfusion studies
Proximal tracheal ends (10 tracheal rings long) were used for the experiments.In half the preparations the epithelium was removed (–Ep) by gently rubbing
the luminal side with a cotton-wrapped metal stick; in the other half, theepithelium was left intact (+Ep).
Under magnification (¥2–4), two stainless-steel hooks were passedthrough the tracheal wall around two adjacent cartilaginous rings as closeas possible to the tracheal muscle insertion. The tracheal segment was thenconnected longitudinally to steel tubes built in the ‘carrier block’ of theapparatus (in–out system) and firmly tightened with silk thread. The apparatusused (Fig. 1a–c; EMKA Technologies, Paris, France) was an improvedversion of the cannulated tracheal system described previously.2 Theimprovement4 consisted of the fact that the lower hook, which served as afixed point, was attached via the silk thread to the micrometric screw servingto adjust hook tension on the tracheal wall. This allowed precise adjustmentof the resting tension. The upper hook was connected to a force transducer(IT1-25; EMKA Technologies), the latter being attached to a micromanipulatorthat permitted displacement of the upper hook along a strict vertical axis.Any change in tension at the level of the tracheal muscle was registered bythe recorder (AT 550; Gould Instrument Systems, Valley View, OH, USA),to which the amplified signal (EMKA Technologies four-way amplifier) fromthe transducer was connected. The ‘carrier block’ for cannulated tracheacould be removed easily and replaced by one suitable for mounting trachealrings (Fig. 1b; ring system and human airway, see below).
The KH solution (37!C, pH 7.4, gassed with 95% O2/5% CO2) was perfusedat a constant flow rate (2 mL/min) through the organ bath (outer perfusion–OUT)and through the lumen of the tracheal segment (inner perfusion–IN) usingperistaltic pumps (Watson Marlow 5025, Falmouth, Cornwall, UK).
Fig. 1 (a) Schematic representationof the experimental apparatus. (a) Thenew tracheal tube preparation with the‘carrier’ block. Inner luminally (IN)and outer (OUT) perfusion solutionsare maintained at 37!C and bubbledwith 95% O2–5% CO2 and a constantflow rate of 2 mL/min is maintained.Under microscopic control, twostainless-steel hooks were passedthrough the tracheal wall around twoadjacent cartilagineous rings as closeas possible to the tracheal muscleinsertion. The lower hook, whichserved as a ‘fixed point’, was attachedvia silk thread to the micrometricscrew serving to adjust hook tensionon the tracheal wall. This allowedprecise adjustment of resting tension.The upper hook was connected to aforce transducer. (b) Tracheal ringpreparation. The ‘carrier block’ forcannulated trachea could be easilyremoved and replaced by an identicalone which the lower part has beenadapted for the mounting of trachealrings. (c) A photograph of the newtracheal tube preparation. (d) Trachealtube with the hooks that are placed asclose as possible to the end of thetracheal rings and close to the trachealmuscle.

S-Carboxymethylcysteine and airways 665
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Asia Pty Ltd
Procedure
Perfusion studies
In this experimental model, studies were conducted using preparationswith and/or without epithelium taken from animals pretreated with S-CMC(S-CMCpr) or from control animals preparations.
After a period of stabilization (45–60 min), the tracheal muscle wasstretched to its optimal length, corresponding to approximately 1.5 g passivepretension. Preliminary assays were performed to determine the optimalstretch of the muscle, as described previously.4 The length–tension relationshipdid not differ significantly between S-CMCpr and control preparations.
In the initial set of experiments, cumulative concentrations of carbachol(10–7 to 10–3 mol/L) were perfused IN (+Ep or –Ep) or OUT (+Ep or –Ep)in S-CMCpr or controls preparations (n = 8 for each). We concluded fromthese experiments that responses to IN and OUT stimulation in –Ep preparationswere identical (on the basis of EC50 values and maximal responses) and, tofurther examine the effects of S-CMC in –Ep preparations on IN stimulationwith carbachol, we designed another four sets of experiments.
First, to examine the effect of pre-incubation, preparations taken fromuntreated animals were incubated with 10–3 mol/L S-CMC (S-CMCinc; –Ep;n = 8) for 60 min and cumulative concentrations of carbachol were perfusedluminally (IN).
In the second set of experiments, S-CMCpr (–Ep) preparations werepre-incubated with 10–6 miol/L indomethacin (INDinc; both sides; n = 8) or10–5 mol/L capsaicin (CAPSinc; both sides; n = 6) for 60 min and prior toperfusion of cumulative concentrations of carbachol IN. The effects of S-CMC on the relaxant effects of aminophylline were examined in S-CMCpr(–Ep) preparations that were first precontracted with a medium concentration(10–6 mol/L) of carbachol OUT and then perfused IN with 10–8 to 10–3 mol/Laminophylline (n = 10) and compared with control responses to aminophylline(n = 6). In the third set of experiments, to examine direct relaxant effects ofS-CMC, preparations taken from control animals were precontracted OUTwith an EC50 concentration of carbachol (10–6 mol/L) and then perfused INwith cumulative concentrations of S-CMC, ranging from 10–7 to 10–3 mol/L(+Ep, n = 7; –Ep, n = 7).
Human airway
Preparations of human bronchi (3–4 mm in diameter, n = 4 in each group)were obtained during thorax surgery (partial lung resection due to lung cancer),stored for approximately 1 h in cold (+4!C) physiological salt solution (PSS;NaCl 0.9%) and then dissected into 2–3 mm ring preparations and incubatedwith 10–5 mol/L S-CMC for 4 h. Paired controls were left in cold PSS duringthat time. In either one of the paired preparations, the epithelium wasremoved before the preparations were mounted for in vitro experiments.Owing to scarcity of the specimens, cannulation was not possible andtherefore the experiments were performed only on bronchial rings. In thesepreparations only dose–response curves to carbachol were obtained.
Substances
The following substances were used: carbachol (carbamylcholine chloride,Sigma Chimie, St Quentin Fallavier, France), indomethacin (Sigma Chimie),aminophylline (theophylline–ethylenediamine; Pharmacie Centrale desHopitaux, Paris, France), capsaicin (Sigma Chimie) and S-CMC (Park-Davis, Orleans, France). The S-CMC was diluted in 10% NaOH, whereasindomethacin was diluted in methanol; final dilutions of both stock solutionswere made in KH solution. Final solutions contained less then 0.01%methanol and NaOH; all controls were exposed to the same concentrationsof solvent as the treatment groups.
Statistical analyses
Data are expressed as a percentage of the maximal response and in absolutevalues (g or s) and given as the mean±SEM. Half-maximal concentration
(EC50) values were calculated by means of non-linear regression using theHill–Langmuir equation implemented in GraphPad Prism (GraphPadSoftware, San Diego, CA, USA) and the results given as the mean of –log10
EC50 values obtained. Statistical analysis was perfomed using analysis ofvariance and Student’s t-test for paired or unpaired data adjusted for multiplecomparisons (Bonferroni), as appropriate. P < 0.05 was regarded as beingstatistically significant. The False Discovery Rate (FDR) procedure wasalso used for multiple comparisons.22,23 All statistical analyses wereperformed using the software package Graph Pad Prism 4 for Windows(Graph Pad Software), except for the FDR, for which a short routine waswritten in Microsoft Excel (Microsoft Office 2000; Microsoft, Redmond,VA, USA).
RESULTS
Animals pretreated with oral S-CMC did not show any clinical signsof disease or metabolic disturbances. They showed normal cageactivity, no disturbances in the sleep–wake pattern, normalspontaneous food and water intake, weight gain ordinary stoolconsistency and normal eyes and no piloerection.
Perfusion studies
Effects of oral pretreatment and pre-incubation with S-CMC and capsaicin and indomethacin
In preparations taken from S-CMCpr amimals, we found diminishedsensitivity to carbachol in –Ep, but not +Ep, preparations(Tables 1,2; Fig. 2). Interestingly, the diminished sensitivity followingepithelial application (inside, IN) of carbachol in –Ep preparationsfrom S-CMCpr was absent in preparations pre-incubated with10–5 mol/L capsaicin (CAPSinc; Table 3; Fig. 3), but was maintainedin preparations pre-incubated with 10–6 mol/L indomethacin
Table 1 EC50 values for carbachol (IN)
+Ep –Ep P
Control 5.40 ± 0.11 5.85 ± 0.06 < 0.005S-CMCpr 5.19 ± 0.08 5.52 ± 0.09 < 0.005P NS < 0.005
Data are the mean±SEM of EC50 values (–log10; mol/L), obtained inpreparations with (+Ep) and without (–Ep) epithelium, taken from controlsor animals pretreated for 2 weeks with with s-carboxymethyl-l-cysteine (S-CMC; 200 mg/kg per day, p.o.; S-CMCpr), following stimulation from theepithelial side (IN) with cumulative concentrations of carbachol (F = 10.45).
Table 2 EC50 values for carbachol (OUT)
+Ep –Ep P
Control 6.01 ± 0.09 5.92 ± 0.06 NSS-CMCpr 6.17 ± 0.08 5.61 ± 0.05 < 0.005P NS < 0.005
Data are the mean±SEM of EC50 values (–log10; mol/L), obtained inpreparations with (+Ep) and without (–Ep) epithelium, taken from controlsor animals pretreated for 2 weeks with with s-carboxymethyl-l-cysteine (S-CMC; 200 mg/kg per day, p.o.; S-CMCpr), following stimulation from theserosal side (OUT) with cumulative concentrations of carbachol (F = 11.14).

666 D Pavlovic et al.
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Asia Pty Ltd
(INDinc; Table 3). In control experiments, we observed that pre-incubation of ring preparations with capsaicin did not affect theirsensitivity (EC50) or the maximal force developed (Tmax) followingcarbachol stimulation.
In contrast with the effects of oral pretreatment, pre-incubationof tissues with S-CMC (S-CMCinc) for 60 min did not affect thesensitivity of –Ep preparations to carbachol IN stimulation (Table 3).However, all +Ep preparations were more sensitive to carbacholOUT stimulation then to carbachol IN stimulation. Removal of theepithelium increased sensitivity to carbachol in all preparationsexcept for control preparations stimulated with carbachol (OUT;Tables 1,2).
Direct effect of S-CMC, Tmax and aminophylline
In +Ep or –Ep preparations precontracted with carbachol (10–6 mol/L)OUT, S-CMC alone in the concentration range 10–7 to 10–3 mol/LIN had neither contracting nor relaxant effects. In addition, the Tmax
developed following stimulation with carbachol did not depend onthe side of stimulation (OUT or IN) and was not affected by removalof the epithelium, pretreatment with S-CMC or pre-incubation
Fig. 2 Cumulative concentration–responses curves constructed after administration of carbachol (a,c) epithelialy (IN) or (b,d) serosaly (OUT) in rat isolatedtrachea with (a,b) and without (c,d) epithelium. Preparations were taken from control animals or animals pretreated for 2 weeks with s-carboxymethyl-l-cysteine (S-CMC; 200 mg/kg per day, p.o.). (!), control; ("), S-CMC pretreated. Tension is expressed as a percentage of the maximal tension (Tmax) obtainedand presented as the mean±SEM.
Table 3 EC50 values for carbachol following pre-incubation withindomethacin or capsaicin
–Ep (IN)
Control 5.85 ± 0.06*S-CMCinc 5.71 ± 0.07S-CMCpr 5.52 ± 0.09S-CMCpr (INDinc) 5.59 ± 0.09†‡
S-CMCpr (CAPSinc) 5.99 ± 0.15*†
Data are the mean±SEM of EC50 values (–log10; mol/L), obtained inpreparations without epithelium (–Ep) following stimulation from theepithelial side (IN) with cumulative concentrations of carbachol. Preparationswere taken from controls and pre-incubated in s-carboxymethyl-l-cysteine(S-CMC; 10–3 mol/L; S-CMCinc) or from animals pretreated for 2 weekswith oral S-CMC (200 mg/kg per day, p.o.; S-CMCpr) and pre-incubatedwith 10–6 mol/L indomethacin (INDinc) or 10–5 mol/L capsaicin (CAPSinc).*P < 0.03 compared with S-CMCpr (F = 5.444; see also Table 2); †P < 0.05compared with S-CMCinc (FDR procedure); ‡P < 0.05 compared withS-CMCpr (FDR procedure).

S-Carboxymethylcysteine and airways 667
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Asia Pty Ltd
with 10–3 mol/L S-CMC, 10–6 mol/L indomethacin or 10–5 mol/Lcapsaicin (data not shown). Sensitivity to aminophylline IN in –Eppreparations precontracted with carbachol 10–6 mol/L OUT was notaffected by pretreatment with S-CMC (–log10 EC50: 3.2 ± 0.1 vs3.66 ± 0.11 for S-CMCpr vs controls, respectively; NS).
Human bronchial rings
The carbachol concentration–response curves for human bronchialrings pre-incubated for 4 h with S-CMC, without epithelium, were lesssensitive to carbachol stimulation (Fig. 4, Table 4), thus indicatingthat removal of the epithelium increased sensitivity to carbacholstimulation only in preparations not pre-incubated with S-CMC,whereas pre-incubation with S-CMC seemed to diminish sensitivityto carbachol.
DISCUSSION
In the present study, we found that removal of the epitheliumrendered preparations more sensitive to carbachol stimulation and,alternatively, that pretreatment with S-CMC rendered preparationswithout epithelium less sensitive to carbachol stimulation comparedwith controls (–Ep). These changes, although small, were significantand require an explanation. In addition, we were able to demonstratesimilar small effects in human bronchus denuded of epithelium (i.e.preparations incubated with S-CMC were less sensitive to carbachol).
The results of various experimental studies, including our own,clearly demonstrate that tracheal epithelium can modulate trachealsmooth muscle contraction.1–3,24,29 In most of these studies, theinhibitory effects of the bronchial epithelium were found to be small(except in dogs). However, it has been notoriously difficult to identifya single underlying pathophysiological mechanism. In addition, itseems that not only the contracting, but also the relaxing effects ofsome pharmacological agents are dependent on the presence of an
intact epithelium.25 This may be relevant for a better understandingof different airway pathologies. It has been shown that the bronchialepithelium is damaged in patients with severe asthma, indicating thatbronchial epithelial damage and airway hyperresponsiveness couldbe linked.26 Several studies have demonstrated that the bronchialepithelium constitutes a powerful diffusion barrier2,27 and can attenuatethe effects of pharmacological agents applied luminally. The findingsof the present study, although demonstrating small effects, supportthis hypothesis. It appears that the epithelium in rats mediates lessinhibition compared with guinea-pigs. In guinea-pigs, we have foundbigger differences following serosal compared with epithelialperfusion (D Pavlovic et al., unpubl. obs., 1999). However, it hasbeen quite difficult to demonstrate airway hyperresponsivenessin vitro in airways already hyperresponsive in vivo. The significanceof small in vitro changes that we observed may (but do not have to)correspond to more important in vivo changes, which we would liketo verify in further experiments.
Incubation with S-CMC appears to influence human ringpreparations but not rat preparations. However, in the present study,rat preparations were incubated for 60 min whereas human ring
Fig. 3 Cumulative concentration–responses curves constructed afteradministration of carbachol epithelialy (IN) in rat isolated trachea withoutepithelium. Preparations were taken from animals pretreated for 2 weeks withs-carboxymethyl-l-cysteine (S-CMC; 200 mg/kg per day, p.o.) and either notincubated (!) or incubated (") for 60 min with 10–6 mol/L capsaicin.Tension is expressed as a percentage of the maximal tension (Tmax) obtainedand presented as the mean±SEM.
Fig. 4 Cumulative concentration–responses curves constructed after theadministration of carbachol in human isolated bronchial rings. Half thebronchial rings were stored at 4!C, whereas the other half were incubatedfor 4 h in s-carboxymethyl-l-cysteine (S-CMC; 10–5 mol/L). Then, in pairedpreparations, the epithelium was either removed or left intact (–Ep and +Ep,respectively). Tension is expressed as a percentage of the maximal tension(Tmax) obtained and presented as the mean±SEM. (!), no S-CMC, –Ep;("), no S-CMC, +Ep; (#), S-CMC, –Ep; ($), S-CMC, +Ep.
Table 4 EC50 values for human bronchus
+Ep –Ep P
Control 7.2 7.7 NSS-CMC 6.9 6.8 NSP NS < 0.05
Data are the mean±SEM of EC50 values (–log10; mol/L), obtained in humanpreparations with (+Ep) and without (–Ep) epithelium and either incubatedfor 4 h in the presence of s-carboxymethyl-l-cysteine (S-CMC; 10–5 mol/L;F = 8.54) or not (control).

668 D Pavlovic et al.
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Asia Pty Ltd
preparations were incubated for 4 h and this may explain thedifferences observed.
In a recent animal study, it was shown that in SO2-exposed ratscarbocysteine diminished fucose, sialic acid and protein content, aswell as the number of inflammatory cells, and reduced free radicalsand elastase activity in bronchoalveolar lavage fluid.20 In addition,an increase in cAMP in tracheal tissue was observed20 and patch-clamp techniques revealed increased activity and density of cAMP-dependent Cl– channel.21 These findings could, in principle, explainour observations. However, numerous other mechanisms may alsobe involved.
There is evidence that blocking enkephalinase (which degradeskinins) by phosphoramidon increases bronchoconstriction similarto removal of the epithelium.28 Indeed, mechanical removal of theepithelium could promote the release of different mediators frommast cells, which could be responsible, at least in part, for thebronchial hyperresponsiveness observed experimentally.29 It has alsobeen shown that S-CMC increases the production of sialomucins11,12
and suggested that sialomucins could have an antikinin action.17 Theanti-inflammatory activity of S-CMC was confirmed in an animalstudy where it reduced neutrophil infiltration provoked by intratrachealinjection of interleukin (IL)-1.18 The same study demonstrated thatS-CMC diminished smoke-induced bronchial hyperresponsivenessin guinea-pigs. Very recently, it was demonstrated that treatmentwith S-CMC effectively reduces airway hyperreactivity and airwayinflammation at different phases of the response to secondaryallergen challenge in mice, implicating the possible importance ofthe timing of S-CMC administration.19
Indeed, capsaicin-sensitive nerve terminals secrete various,although only partially identified, mediators.30–33 It is conceivablethat some as yet unidentified mediator/s originating from capsaicin-sensitive nerve terminals could have inhibitory effects on trachealsmooth muscle contraction. In addition, mechanical removal of theepithelium could have contributed to the release of such mediator/s.29
Nerve terminal destruction by capsaicin and the disappearance ofthe putative inhibitory agent could, in turn, have increased thesensitivity of tracheal smooth muscle to carbachol compared (in thepresent experiments) with S-CMCpr preparations not pre-incubatedwith capsaicin. It is tempting to hypothesize that S-CMS binds nitricoxide (NO) released from sensory nerves in epithelium-denudedtissues and thereby suppresses the responsiveness to carbachol.34 Asa result of treatment with capsaicin, the NO stores may be depleted andS-CMS will not inhibit carbachol-induced contractions any further.
Other hypotheses, such as an anti-oxidant action for S-CMC,similar to one recently described for carbocysteine lysine saltmonohydrate (SCMC-Lys),35 should be examined. Last, but notleast, one simple feed-back mechanism could be proposed that couldexplain our finding of diminished sensitivity in –Ep preparationsfrom S-CMCpr: removal of the epithelium and removal of one ormore excitatory agent(s) secreted by the epithelium renders trachealsmooth muscle less sensitive to carbachol. The prostaglandins,products of arachidonic acid metabolism, do not seem to be involvedbecause pre-incubation of preparations with indomethacin did notaffect the sensitivity of the preparations. The therapeutic effects ofS-CMC may be variable and it has been proposed that variation inthe efficacy of S-CMC and pharmacogenomics may be underlyingfactors in this variation.36–38 Certainly, further insights into themechanisms of action of S-CMC are needed to understand its effects,especially in human tissues.
In the present study, we used a slightly improved in vitro modelthat permitted independent perfusion of the epithelial (luminal) andserosal (outside) layers of the airway.2,4,5,24,39–41 However, an alternativemodel is also available.42 This system incorporates interchangeablecarriers for tracheal or bronchial ring preparations that facilitatecomparative experiments and is not a significant financial burdenwhen the use of both techniques is necessary.
The principle finding of the present study is that pretreatment ofanimals, or longer incubation of human bronchus, with S-CMCinduces relatively small decrease of reactivity of rat or human airwaysmooth muscle denuded of epithelium. Whether S-CMC hasbeneficial effects in hyperresponsive airways with injured epitheliumhas to be examined. Because S-CMC is widely used throughoutEurope as a supplementary therapeutic in various pathologiescharacterized by airway obstruction and hyperresponsiveness,further explanation of the mechanisms of action underlying itsbeneficial effects, probably in a model of sensitized, allergic animals,would be valuable.
ACKNOWLEDGEMENTS
This study was funded by the research budget of INSERM U408,Paris, and EMA University, Greifswald.
REFERENCES
1. Hay DW, Farmer SG, Raeburn D et al. Airway epithelium modulatesthe reactivity of guinea-pig respiratory smooth muscle. Eur. J. Pharmacol.1986; 129: 11–18.
2. Pavlovic D, Fournier M, Aubier M, Pariente R. Epithelial vs. serosalstimulation of tracheal muscle: Role of epithelium. J. Appl. Physiol.1989; 67: 2522–6.
3. Stuart-Smith K, Vanhoutte PM. Airway epithelium modulates theresponsiveness of porcine bronchial smooth muscle. J. Appl. Physiol.1988; 65: 721–7.
4. Pavlovic D, Viires N, Zedda C, Fournier M, Aubier M. Effects ofcorticosteroids on epithelial structure and smooth muscle function ofrat trachea. Eur. Respir. J. 1998; 11: 575–82.
5. Pavlovic D, Brione E, De Vernejoul D, Aubier M. Partial inhibition byepithelium of tracheal smooth muscle relaxation induced by thepotassium channel activator, BRL 38227. Br. J. Pharmacol. 1993; 110:139–44.
6. Marom Z, Shelhamer J, Alling D, Kaliner M. The effects of corticos-teroids on mucous glycoprotein secretion from human airways in vitro.Am. Rev. Respir. Dis. 1984; 129: 62–5.
7. Wanner A, Zarzecki S, Hirsch J, Epstein S. Tracheal mucous transportin experimental canine asthma. J. Appl. Physiol. 1975; 39: 950–7.
8. Brown DT. Carbocysteine. Drug Intell. Clin. Pharm. 1988; 22: 603–8.9. Molina C, Aiache JM, Pradelle J, Brun J, Cheminat JC. Interet clinique
des medications fluidifiantes des secretions bronchiques. Poumon.Coeur 1970; 26: 91–102.
10. Allegra L, Cordaro CI, Grassi C. Prevention of acute exacerbations ofchronic obstructive bronchitis with carbocysteine lysine salt monohydrate:A multicenter, double-blind, placebo-controlled trial. Respiration 1996;63: 174–80.
11. Degand P. Etude biochemique du mode d’action de la s-carboxy-methyl-cysteine. Bull. Physiophathol. Respir. 1973; 9: 462–3.
12. Havez R, Degand P, Roussel P, Randoux A. Mode d’action biochemiquedes derives de la cysteine sur le mucus bronchique. Poumon. Coeur1970; 26: 81–90.
13. Davis SS, Deverell LC. Rheological factors in mucociliary clearance.The assessment of mucotropic agents using an in vitro model. Mod.Probl. Paediatr. 1976; 19: 207–17.

S-Carboxymethylcysteine and airways 669
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Asia Pty Ltd
14. Martin R, Litt M, Marriott C. The effect of mucolytic agents on therheologic and transport properties of canine tracheal mucus. Am. Rev.Respir. Dis. 1980; 121: 495–500.
15. Edwards GF, Steel AE, Scott JK, Jordan JW. s-Carboxymethylcysteinein the fluidification of sputum and treatment of chronic airway obstruction.Chest 1976; 70: 506–13.
16. Puchelle E, Aug F, Polu JM. Effect of the mucoregulator s-carboxy-methyl-cysteine in patients with chronic bronchitis. Eur. J. Clin.Pharmacol. 1978; 14: 177–84.
17. Muh J, Latreille J, Guerrin F. Recherche d’inhibiteurs naturels dubronchospasme a la bradykinine chez le cobaye. In: ColloaqueInternational de Pathologie Thoracique. Poissot, Medic diffuser,Clichy. 1968; 193–202.
18. Asti C, Melillo G, Caselli GF et al. Effectiveness of carbocysteinelysine salt monohydrate on models of airway inflammation andhyperresponsiveness. Pharmacol. Res. 1995; 31: 387–92.
19. Takeda K, Miyahara N, Kodama T et al. s-Carboxymethylcysteinenormalises airway responsiveness in sensitised and challenged mice.Eur. Respir. J. 2005; 26: 577–85.
20. Ishibashi Y, Okamura T, Masumoto Y, Tachiiri T, Momo K. Effectsof carbocisteine on airway inflammation and related events in SO2-exposed rats. Nihon Kokyuki Gakkai Zasshi 2001; 39: 17–23.
21. Meyer G, Doppierio S, Daffonchio L, Cremaschi D. s-Carbocysteine-lysine salt monohydrate and cAMP cause non-additive activation of thecystic fibrosis transmembrane regulator channel in human respiratoryepithelium. FEBS Lett. 1997; 404: 11–14.
22. Hochberg Y, Benjamini Y. Controlling of the false discovery rate: Apractical and powerful approach to multiple testing. J. R. Soc. Ser. B1995; 57: 289–300.
23. Curran-Everett D. Multiple comparisons: Philosophies and illus-trations. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000; 279: R1–8.
24. Folkerts G, Nijkamp FP. Airway epithelium: More than just a barrier!Trends Pharmacol. Sci. 1998; 19: 334–41.
25. Fedan J, Hay D, Farmer S, Raeburn D. Epithelial cells: Modulation ofairway smooth muscle reactivity. In: Barnes PJ, Rodger I, Thompson N(eds). Asthma, Basic Mechanics and Clinical Management. AcademicPress, London. 1988; 143–62.
26. Laitinen LA, Heino M, Laitinen A, Kava T, Haahtela T. Damage ofthe airway epithelium and bronchial reactivity in patients with asthma.Am. Rev. Respir. Dis. 1985; 131: 599–606.
27. Morikawa T, Webber SE, Widdicombe JG. The effect of hydrogenperoxide on smooth muscle tone, mucus secretion and epithelial albumintransport of the ferret trachea in vitro. Pulmon. Pharmacol. 1991; 4:106–13.
28. Frossard N, Stretton CD, Barnes PJ. Modulation of bradykinin responsesin airway smooth muscle by epithelial enzymes. Agents Actions 1990;31: 204–9.
29. Franconi GM, Rubinstein I, Levine EH, Ikeda S, Nadel JA. Mechanicalremoval of airway epithelium disrupts mast cells and releases granules.Am. J. Physiol. 1990; 259: L372–7.
30. Meini S, Evangelista S, Geppetti P et al. Pharmacologic and neuro-chemical evidence for the activation of capsaicin-sensitive sensorynerves by lipoxin A4 in guinea pig bronchus. Am. Rev. Respir. Dis.1992; 146: 930–4.
31. Kawao N, Ikeda H, Kitano T et al. Modulation of capsaicin-evokedvisceral pain and referred hyperalgesia by protease-activated receptors1 and 2. J. Pharmacol. Sci. 2004; 94: 277–85.
32. Ono K, Saito A, Goto K, Kasuya Y, Shigenobu K. Elucidation withthe protease alpha-chymotrypsin of the inhibitory modulating actionof endogenous neuropeptide Y over sympathetic neurotransmission inrat vas deferens. J. Pharmacol. Sci. 2003; 91: 113–24.
33. Paulino N, Dantas AP, Bankova V et al. Bulgarian propolis inducesanalgesic and anti-inflammatory effects in mice and inhibits in vitrocontraction of airway smooth muscle. J. Pharmacol. Sci. 2003; 93: 307–13.
34. Kloek J, van Ark I, Bloksma N et al. Glutathione and other low-molecular-weight thiols relax guinea pig trachea ex vivo: Interactions with nitricoxide? Am. J. Physiol. Lung Cell Mol. Physiol. 2002; 283: L403–8.
35. Brandolini L, Allegretti M, Berdini V et al. Carbocysteine lysine saltmonohydrate (SCMC-LYS) is a selective scavenger of reactive oxygenintermediates (ROIs). Eur. Cytokine Netw. 2003; 14: 20–6.
36. Steventon GB. A methodological and metabolite identification study ofthe metabolism of s-carboxymethyl-l-cysteine in man. Chromatographia1998; 48: 561–8.
37. Steventon GB, Waring R, Sturman S, Heafield M, Williams A. Themetabolism of s-carboxymethyl-l-cysteine in man: A neurodegenerativedisease study. Med. Sci. J. 1999; 27: 119–20.
38. Steventon GB. Diurnal variation in the metabolism of s-carboxymethyl-l-cysteine in humans. Drug Metab. Dispos. 1999; 27: 1092–7.
39. Folkerts G, van der Linde HJ, Nijkamp FP. Virus-induced airwayhyperresponsiveness in guinea pigs is related to a deficiency in nitricoxide. J. Clin. Invest. 1995; 95: 26–30.
40. Folkerts GLH, Verheyen AK, Nijkamp FP. Endogenous nitric oxidemodulation of potassium-induced changes in guinea-pig airway tone.Br. J. Pharmacol. 1995; 115: 1194–8.
41. Folkerts G, Kloek J, Geppetti P, van der Linde HJ, Nijkamp FP. Factorsthat determine acetylcholine responsiveness of guinea pig trachealtubes. Eur. J. Pharmacol. 2001; 420: 151–7.
42. Munakata M, Huang I, Mitzner W, Menkes H. Protective role ofepithelium in the guinea pig airway. J. Appl. Physiol. 1989; 66: 1547–52.
Copyright © 2022 FDOKUMEN




















