
THE ROLE OF INTRODUCED BIVALVE MOLLUSC SPECIES IN MARICULTURE1
Upload
independentCategory
view
1download
0
TISSUE & CELL 1979 11 (2) 337-344 P&shed by Longman Group Ltd. Printed in Great Britain
MARTIN PORVAZNIK*, JORGE LUIS RIBAS? and JOSEPH L. PARKER*
RHOMBIC PARTICLE ARRAYS IN GILL EPITHELIUM OF A MOLLUSC, APLYSlA CALIFORNICA
ABSTRACT. Gill epithelia from adult and juvenile Aplysia were examined by conven- tional thin section and freeze-fracture methods. Freeze-fracture replicas of adult gill epithelium revealed septate and gap junctions, which served as membrane markers for the epithelial cells. In these same cell membranes, non-junctional rhombic arrays of intramembranous particles were observed on prominent ridges on the membrane P fracture face of some epithelial cells. In thin sections of adult epithelium, nerve termi- nals were observed abutting the lateral plasma membranes near the basal lamina of some epithelial cells. Correlative areas of plasma membrane in freeze-fracture replicas showed a close association between rhombic particle arrays and abutting nerve termi- nals. In thin sections of juvenile Aplysia, nerve terminals abutting the epithelial cells were not recognizable, and rhombic arrays were not observed in freeze-fracture replicas. This suggested that a developmental association existed between the appearance of rhombic arrays in adult epithelia and their innervation. It is not known with certainty if, in invertebrates, rhombic arrays are an essential structural entity of all innervated cell membranes; however, in the cells thus far studied, there appears to be an associa- tive condition. In the case of the gill epithelium of Aplysia, rhombic arrays are located in the same vicinity as the abutting nerve terminals. Similar arrays of intramembranous particles have been observed in myoneural postjunctional complexes of other inverte- brates and have been interpreted to be the morphological expression of neurotransmit- ter receptors. An analogous explanation is put forth, namely that rhombic arrays may represent the structural correlates of neurotransmitter receptors and/or ionic channels in innervated membranes of invertebrates.
Introduction
RHOMBIC arrays have been described in in- vertebrates as regularly spaced, intramembra- nous particles lying at the intersections of two sets of parallel rows. These sets are oriented at an acute angle with respect to each other, resulting in the formation of a two-dimen-
* Department of Experimental Hematology, Armed Forces Radiobiology Research Institute, Bethesda, Maryland 20014.
i Department of Anatomy, School of Medicine, Uniformed Services University of the Health Sci- ences, Bethesda, Maryland 20014.
Address reprint requests to: Dr Jorge Luis Ribas Anatomy Department/USUHS 4301 Jones Bridge Road, Bethesda, MD 20014.
Received 17 July 1978. Revised 5 January 1979.
sional lattice with a rhombic-repeating unit (Quick and Johnson, 1977).
Rhombic arrays are associated with elon- gated convexities on P fracture faces; their corresponding pits, when found on concavi- ties of E faces, have lattice dimensions similar to those of particle arrays (Quick and John- son, 1977; Prescott and Brightman, 1976). As opposed to the particle arrays of inter- cellular junctions, rhombic arrays are not shared by adjacent cells or have a discernible association with internal membranes (Quick and Johnson, 1977). At their loci, neighbor- ing cells are separated by wide areas of extra- cellular space (Prescott and Brightman, 1976; Quick and Johnson, 1977). The above find- ings are taken as compelling evidence that they are not intercellular junctions, but rather
337

338
localized intramembranous specializations characteristic of certain types of cells.
Rhombic arrays are found in association with nerve and muscle cells in Dug&u (Quick and Johnson, 1977) and Aplysia (Prescott and Brightman, 1976; Ribas and Porvaznik, 1977). In earthworms, highly ordered arrays of intramembranous particles have been ob- served on the postjunctional P face of the myoneural junction; they have been inter- preted as the structural correlate of choliner- gic receptors (Rosenbluth, 1978). The fact that these structures are found in post- synaptic membranes suggests that these in- tramembranous particle arrays could be linked to mechanisms underlying chemical transmission.
In the course of a morpho-functional study of the gill and cardiovascular system in Aplysia californica, it was noticed that many of the gill epithelial cells appeared to be in- nervated. This observation prompted us to undertake a freeze-fracture study. The results of this study show that rhombic particle arrays are also present on P faces of certain gill epithelial cell membranes, and that these arrays are overwhelmingly more numerous near the sites of nerve-epithelial cell apposi- tion.
PORVAZNIK, RIBAS AND PARKER
place of water in the preparation of the para- formaldehyde and of the 0.2 M Na cacodylate stock buffer. Gills were dissected into small pieces (l-2 mm cube) and further fixed for 2 hr in the same fixative at room temperature.
Ultramicrotomy Following aldehyde fixation, the specimens were rinsed several times in buffer and sucrose or buffer and seawater overnight in these solutions at 4°C. Specimens were subse- quently osmicated for l-2 hr, dehydrated in a graded series of ethanol, and embedded in Epon or Spurr resins. Thin sections were cut with a diamond knife and stained with 05 % aqueous uranyl acetate at 40°C for lo-20 min followed by lead citrate (Venable and Cogge- shall, 1965) for l-3 min.
Freeze-fracture Following fixation with either aldehyde re- agent, the specimens were rinsed briefly in buffer and left for at least 2 hr in 30% gly- cerol. After glycerination, they were frozen in liquid Freon 22 at - 150°C and stored in liquid nitrogen until fractured and replicated with platinum and carbon in a Balzer’s 360 M apparatus at - 105°C.
Morphometry Particle sizes in freeze-fracture replicas were measured on micrographs perpendicular to the direction of platinum shadowing, from point to point on the crescent-shaped accumu- lation of platinum (Quick and Johnson, 1977).
Results
By transmission electron microscopy, the gill epithelium consisted of a simple columnar to low cuboidal epithelium containing a diversity of cell types. A detailed description
Materials and Methods
Adult (> 150 g of body weight) and juvenile (< 50 g of body weight) Aplysia californica were obtained from Pacific Bio-Marine Supply Co. (Venice, California) and kept at 15°C in a seawater aquarium. Gills were fixed in situ by immersing Aplysia in a 0.1 M Na cacodylate buffer containing 2.5 % gluta- raldehyde and 0.8 M sucrose at pH 7.6 and 15°C (Henkart, 1975) or in a modified Kar- novsky’s fixative using artificial seawater in
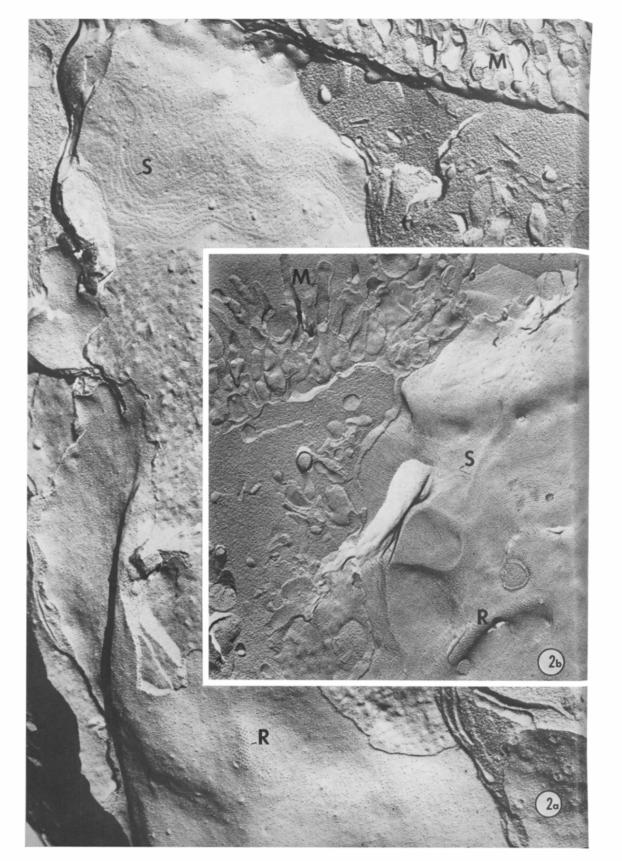
Fig. la. Thin section of an adult gill epithelium showing columnar-shaped cells with apical microvilli (M) and basally located nuclei (N). Nerve terminals (NT) are situated between the basal lamina (BL) and epithelial cells. Smooth muscle (SM); connective tissue (CT). x 14,400.
Fig. lb. High magnification electron micrograph of a nerve terminal (NT) in close apposition to an epithelial cell (E). Dense-core vesicles (D) are prominent identifying features of nerve terminals. Basal lamina (BL). x 105,000.


RHOMBIC PARTICLE ARRAYS
of this epithelial lining will be published else- where. The apical border was characterized by microvilli and cilia; the basal border rested upon a prominent basal lamina. Nerve terminals, having electron-lucent dense-core vesicles or both, were found between the basal lamina and invaginations of the basal border epithelium (Fig. lb). These nerve terminals were observed in association with at least two different cell types: one charac- terized by an abundant rough endoplasmic reticulum and a prominent Golgi apparatus, and the other containing membrane-bound electron-dense bodies and prominent Golgi complexes (Figs. la, b).
The examination of freeze-fracture replicas of the gill pinules revealed a series of intra- membranous particle arrays (Figs. 2a, b). The apical border was easily identified by the presence of cilia and microvilli and by numerous particle arrangements inherent of septate and gap junctions. These apical membrane features provided a reference point for cellular orientation which was further facilitated when the epithelial cell was visual- ized in its entirety (Fig. 2a). The plasmalem- mata of the basal region contained numerous rhombic particle arrays. These arrays were found on elongated convex zones of the P fracture face; the complementary concavities were seldom observed on E fracture face images but, when found, they were character- ized by pits arranged in a lattice pattern simi- lar to those of rhombic arrays (Fig. 3b). Their striking appearance, especially on the P face, permitted their identification even at low magnification scans (Fig. 2a).
The rhombic particle arrays showed no overall orientation with respect to each other but the particle-to-particle relationship within each array was rigidly maintained, such that four adjacent intramembranous particles, when connected by straight lines, approxi-
341
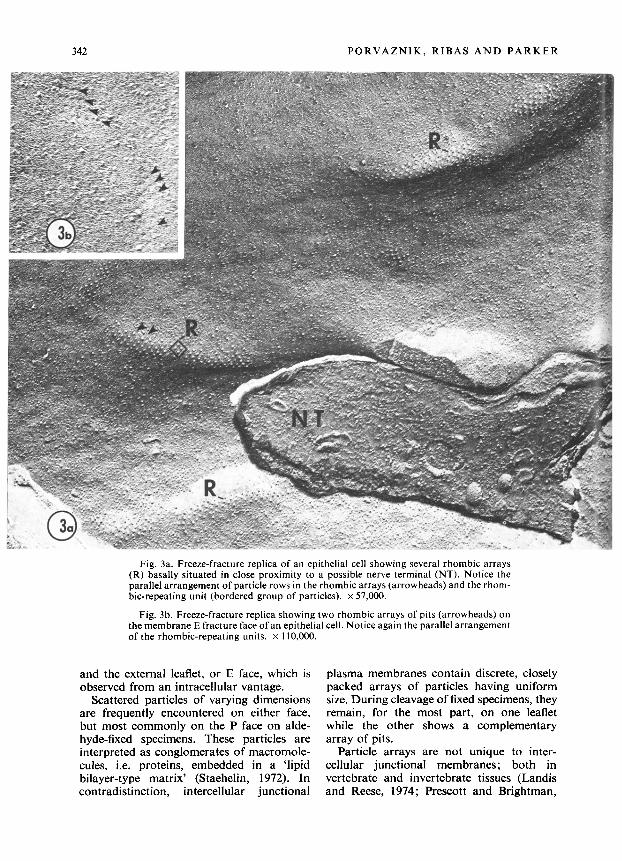
mated an equilateral parallelogram (rhom- bus), thus comprising a repeating unit (Fig. 3a).
Morphometric studies provided the fol- lowing values : width and length of each array -0.1 1 + 0.03 pm and O-60 f 0.38 pm, re- spectively; mean diameter of individual particles - 11.3 f 1.9 nm (N = 100); center- to-center distance among particIes - 25.2 4 3.4 nm (N = 65). The two sets of intersecting parallel rows were aligned at an acute angle of approximately 85”.
Although the preponderance of rhombic arrays were observed on membrane fracture faces along the lower half of the epithelial cells (Figs. 2a, 3a), in a few cuboidal cells an occasional array was found lying immediately below a septate junction (Fig. 2b). Fracture face transitions within a rhombic array re- vealing both P and E fracture faces were never observed. Nerve terminal fracture faces were rarely exposed even though other features, such as vesicular inclusions, cytoplasmic shapes and location permitted their identifi- cation in freeze-fracture replicas (Fig. 3a).
The study of thin sections from gill speci- mens of juvenile A&sia failed to reveal nerve terminals abutting on the lower half of the epithelium. Freeze-fracture replicas obtained from the same gills lacked rhombic arrays even though septate and gap junctions were regularly observed.
Discussion
It is now generally accepted that with the freeze-cleaving technique the cell membrane is fractured along the hydrophobic layer (Branton, 1966; Pinto da Silva and Branton, 1970). This results in the formation of two leaflets having complementary intramembra- nous faces: the cytoplasmic leaflet, or P face, which is viewed from the extracellular space,
Fig. 2a. Freeze-fracture replica of a cuboidal epithelial cell showing cross-fractured apical microvilli (M), a septate junction (S) on the E fracture face and a basally situated rhombic array of intramembranous particles (R) on the P fracture face. x 3 1,400.
Fig. 2b. Freeze-fracture replica showing a rhombic array (R) in an atypical location below the level of the septate junction IS). Apical microvilli (M). x 33,000.
342 PORVAZNIK, RIBAS AND PARKER
Fig. 3a. Freeze-fracture replica of an epithelial cell showing several rhombic arrays (R) basally situated in close proximity to a possible nerve terminal (NT). Notice the parallel arrangement of particle rows in the rhombic arrays (arrowheads) and the rhom- bic-repeating unit (bordered group of particles). x 57,000.
Fig. 3b. Freeze-fracture replica showing two rhombic arrays of pits (arrowheads) on themembrane E fracture face of an epithelial cell. Notice again the parallel arrangement of the rhombic-repeating units. x 110,000.
and the external leaflet, or E face, which is observed from an intracellular vantage.
Scattered particles of varying dimensions are frequently encountered on either face, but most commonly on the P face on alde- hyde-fixed specimens. These particles are interpreted as conglomerates of macromole- cules, i.e. proteins, embedded in a ‘lipid bilayer-type matrix’ (Staehelin, 1972). In contradistinction, intercellular junctional
plasma membranes contain discrete, closely packed arrays of particles having uniform size. During cleavage of fixed specimens, they remain, for the most part, on one leaflet while the other shows a complementary array of pits.
Particle arrays are not unique to inter- cellular junctional membranes; both in vertebrate and invertebrate tissues (Landis and Reese, 1974; Prescott and Brightman,

RHOMBIC PARTICLE ARRAYS
1976), ordered arrays of particles are present in non-junctional membranes, usually on the P face. One of the significant distinctions between intercellular junctional and non- junctional particle arrays is that in the former, there are two matching sets of particle arrays between adjacent plasma membranes.
There are also important differences be- tween vertebrate and invertebrate non- junctional particle arrays. In the majority of vertebrate studies, it appears that the size of the particles is smaller and the distance be- tween particles is shorter than in inverte- brates. Moreover, the particle arrays in the latter seem to be restricted to innervated cell membranes, i.e. nerve and smooth muscle, while in vertebrates they have been observed in many non-innervated cells as well (for review, see Quick and Johnson, 1977), in- cluding cultured hepatoma cells (Porvaznik et al., 1976).
Non-junctional particle arrays have been described in rat astrocytes as ‘assemblies’ (Landis and Reese, 1974), in Aplysia as ‘grills’ (Prescott and Brightman, 1976) and as ‘rhombic arrays’ in Du,gesia (Quick and Johnson, 1977). Since ‘grills’ and ‘rhombic arrays’ are structurally similar, we have adopted the latter term in describing the non- junctional particle arrays present in gill epithelium.
The nature and function of non-junctional particle arrays remain conjectural. In verte- brates, it has been proposed that they func- tion in the transport or permeability of certain substances (Landis and Reese, 1974) or represent receptor sites (Rash and Ellisman, 1974). In vertebrates, rhombic arrays are thought to represent sites of attachment of the sarcolemma to extracellular or intra- cellular elements (Prescott and Brightman, 1976), or to alter the electrical properties of their membranes (Quick and Johnson, 1977).
Non-junctional particle arrays have also been studied extensively in the longitudinal musculature of the earthworm body wall; they have been interpreted as cholinergic receptors extending at least as far as the hydrophobic layer of the sarcolemma (Rosen- bluth, 1978). In the abdominal musculature of the crayfish, inhibitory and excitatory synapses are known to exist. Characteristic postjunctional particle arrays are present for each synaptic type, presumably reflecting
343
chemical differences in neurotransmitter receptors and/or ionic channels (Franzini- Armstrong, 1976). These ordered arrays are not structurally identical to rhombic arrays but are similar in their confinement to inner- vated membranes.
The differences observed among some post- junctional intramembranous arrays may reflect real structural variations or be the result of the techniques used in the prepara- tions of specimens (Rosenbluth, 1978 ; Franzini-Armstrong, 1976). The former may be a more consistent interpretation since purposeful methodological variations pro- duced no discernible changes in the present or previous studies (Prescott and Brightman, 1976).
In this report we have observed rhombic particle arrays in epithelial cell membranes identical to those described by others (Quick and Johnson, 1977) in nerve and muscle. We have found that the epithelial rhombic arrays were generally restricted to sites of innerva- tion. Furthermore, when thin sections of the gill epithelium from juvenile Apfysiu ap- peared to lack nerve terminals, the corre- sponding freeze-fracture images also failed to reveal the presence of rhombic arrays. This suggests that innervation in gill epi- thelium, and therefore the presence of rhom- bic arrays, may be related to the develop- mental stage of the animal.
Rhombic arrays are a feature in innervated cells of some invertebrates. If they are struc- tural entities mediating functional events concerning stimulus-response coupling mechanisms, i.e. secretion or contraction, what is their precise role(s) ? At the present, a conclusive answer is hampered by the dearth of evidence concerning the neurotransmit- ter(s) involved or other anatomical data which would make possible a correlation between rhombic arrays seen in freeze- fracture replicas with some structural fea- ture(s) of the postjunctional membrane as revealed by thin sections. In the earthworm, the correlation has been made between the arrangement and location of intramembra- nous particles and the structural details of en face thin sections of postjunctional mem- brane (Rosenbluth, 1978). In this system, the evidence that these particle arrays are recep- tors is indirect but this seems a likely expla- nation. Similar suggestions have been made in the postsynaptic membrane of Torpedo

344
electroplax (Orci et al., 1974) and crayfish abdominal musculature (Franzini-Arm- strong, 1976).
It is not unreasonable to entertain the possibility that rhombic arrays may also play a similar role in invertebrate epithelial cells since they are present in innervated membranes at approximately the same anato- mical location where nerves terminate. How- ever, it must be emphasized that even though a causality appears to exist between nerve terminals and rhombic arrays, the relatively wide extracellular space between nerves and epithelial cells makes it difficult to obtain a favourable fracture plane through both membranes. Consequently, there is no abso- lute certainty that in epithelial cells, rhombic
PORVAZNIK, RIBAS AND PARKER
arrays are only restricted to postjunctional membranes. However, the present evidence suggest that rhombic arrays are a feature of innervated invertebrate membranes, and therefore they may be the morphological expressions of neurotransmitter receptors and/or ionic channels.
Acknowledgements
We wish to thank Drs Milton Brightman and Thomas Reese for the use of their freeze- fracture facilities; Dr David 0. Carpenter for critical review of the manuscript, Mrs Gwendolyn Harley for her excellent technical assistance and Miss Pamela Compart for her skillful preparation of the manuscript.
References
BRANTON, D. 1966. Fracture faces of frozen membranes. Proc. nnfn Acad. Sci. (Wash.), 55, 1048-1056. FRANZINI-ARMSTRONG, C. 1976. Freeze-fracture of excitatory and inhibitory synapses in crayfish neuro-
muscular junction. J. Microsc. Biol. Cell., 25, 217-222. HENKART, M. 1975. Light-induced changes in the structure of pigmented granules in Aplysia neurons. Science,
N. Y., 188, 155-I 57. LANDIS, D. M. D. and REESE, T. S. 1974. Arrays of particles in freeze-fractured astrocytic membranes. J. Cell
Biol., 60, 3 16-320. ORCI, L., PERRELET, A. and DUNANT, Y. 1974. A peculiar substructure in the postsynaptic membrane of
Torpedo electroplax. Proc. natn Acad. Sri. (Wash.), 71, 307-3 IO. PINTO DA SILVA, P. and BRANTON, D. 1970. Membrane-splitting in freeze-etching: covalently bound ferritin
as a membrane marker. J. Cell Biol., 45, 598-605. PDRVAZNIK, M., JOHNSON, R. G. and SHERIDAN, J. D. 1976. Intercellular junctions and other cell surface
differentiations of H4-III3 hepatoma cells in vitro. J. Ultrastruct. Res., 55, 343-359. PRESCOTT, L. and BRIGHTMAN, M. W. 1976. The sarcolemma of Aplysia smooth muscle in freeze-fracture
preparations. Tissue & Cell, 8, 241-258. QUICK, D. C. and JOHNSON, R. G. 1977. Gap junctions and rhombic particle arrays in Planaria. J. Ultrastru~~t.
Res., 60, 348-361. RASH, J. E. and ELLISMAN, M. H. 1974. Studies of excitable membranes. I. Macromolecular specializations
of the neuromuscular junction and the non-junctional sarcolemma. J. Cell Biol., 63, 567-586. RIBAS, J. L. and PORVAZNIK, 1977. Unpublished observations. ROSENBLUTH, J. 1978. Particle arrays in earthworm post-junctional membranes. J. Cell Rio/., 76, 76-86. STAEHELIN, L. A. 1972. Three types of gap junctions interconnecting intestinal epithelial cells visualized by
freeze-etching. Proc. natn. Acnd. Sri. (Wash.), 69, 13 18-l 32 I. VENABLE, J. H. and COGGESHALL, R. 1965. A simplified lead citrate for use in electron microscopy. J. Cc4
Biol., 25, 407.
Copyright © 2022 FDOKUMEN










![Animal husbandry and mollusc gathering [in the Hellenistic town of New Halos]](https://static.fdokumen.com/doc/165x107/632b357dba70062a77056249/animal-husbandry-and-mollusc-gathering-in-the-hellenistic-town-of-new-halos.jpg)









