
Endurance Training Modulates Lymphocyte Function in Rats with Post-MI CHF
Upload
theconsultationcenterCategory
view
3download
0
Role of T-cell-associated lymphocytefunction-associated antigen-1 in the pathogenesisof experimental colitis
Kevin P. Pavlick1*, Dmitry V. Ostanin1*, Kathryn L. Furr1, F. Stephen Laroux1, Carla M. Brown1,Laura Gray1, Christopher G. Kevil2 and Matthew B. Grisham1
1Department of Molecular and Cellular Physiology and 2Department of Pathology, Louisiana State University Health SciencesCenter, 1501 Kings Highway, P.O. Box 33932, Shreveport, LA 71130-3932, USA
Keywords: adhesion molecules, CD11a, cytokines, inflammatory bowel disease, mesenteric lymph nodes
Abstract
The b2 integrin lymphocyte function-associated antigen-1 (LFA-1; CD11a/CD18) is important forlymphocyte trafficking and activation as well as recruitment to sites of tissue inflammation. Theobjective of this study was to assess the role of ‘T-cell-associated’ LFA-1 in the pathogenesis ofchronic colitis in vivo. Transfer of CD41CD25� T cells isolated from wild-type (wt) mice intoimmunodeficient recipients [recombinase-activating gene-1-deficient (RAG-1�/�)] produced moderateto severe colitis, whereas RAG-1�/� mice injected with CD11a-deficient (CD11a�/�; LFA-1�/�) donorT cells displayed minimal macroscopic and histological evidence of colitis. Surface expression ofL-selectin, a4, a4b7 and chemokine receptor-7 were similar for wt and CD11a�/� donor T cells.Attenuated disease in the CD11a�/� ! RAG-1�/� animals was associated with decreased numbers ofCD41 T cells in the mesenteric lymph nodes (MLNs), spleen and intestinal lamina propria (LP). Inaddition, significant reductions in Th1 cytokines were observed following ex vivo stimulation ofmononuclear cells obtained from the MLNs and colonic LP. Interestingly, mononuclear cells obtainedfrom the spleens of CD11a�/� ! RAG-1�/� exhibited enhanced pro-inflammatory cytokine productioncompared with splenocytes obtained from wt ! RAG-1�/� colitic mice. Taken together, our datasuggest that T-cell-associated CD11a (LFA-1) expression plays a dual role in the initiation of chronicgut inflammation by facilitating naive T-cell priming/activation and expansion within MLNs and byaugmenting pro-inflammatory cytokine production following secondary stimulation by antigen-presenting cells in the colonic interstitium.
Introduction
Recent experimental and clinical studies suggest that Crohn’sdisease (CD) may result from a complex interaction amonggenetic, environmental and immune factors. Indeed, a numberof different studies, using genetically engineered and immune-manipulated mice, demonstrate that chronic colitis resultsfrom a dysregulated immune response to components of thenormal gut flora (1, 2). It is well recognized that the intestinaland/or colonic interstitium of patients with active CD containssubstantial numbers of CD4+ T cells with a Th1 phenotypecharacterized by the production of IL-2, IFN-c and tumornecrosis factor-a (TNF-a). It is thought that the production ofthese T-cell-derived cytokines perpetuates or ‘drives’ chronic
gut inflammation by activating tissue macrophages to releasepro-inflammatory cytokines and mediators, including TNF-a,IL-1b, IL-12, nitric oxide and reactive oxygen species (3).Together, Th1- and macrophage-derived cytokines and medi-ators activate the microvascular endothelium within the gutto enhance expression of adhesion molecules thereby pro-moting the recruitment of potentially injurious phagocyticleukocytes such as neutrophils and monocytes. Becauseseveral different animal models of chronic colitis suggestthat CD4+ T cells play a critical role in initiating and/or per-petuating chronic gut inflammation, it is important to under-stand the mechanisms involved in leukocyte trafficking
*These authors contributed equally to this work.
Correspondence to: M. B. Grisham; E-mail: [email protected] Received 14 October 2005, accepted 29 November 2005
Transmitting editor: C. Terhorst Advance Access publication 13 January 2006
International Immunology, Vol. 18, No. 2, pp. 389–398doi:10.1093/intimm/dxh378
ª The Japanese Society for Immunology. 2006. All rights reserved.For permissions, please e-mail: [email protected]
by guest on February 23, 2016http://intim
m.oxfordjournals.org/
Dow
nloaded from

and activation in T-cell-dependent models of chronic gutinflammation.Although several different T-cell-associated adhesion mol-
ecules [e.g. CD62L, CD11a/CD18, chemokine receptor-7(CCR7), a4b7] have been identified as important moleculardeterminants for T-cell migration to and activation withinsecondary lymphoid tissue as well as recruitment to targettissue, little is known regarding their potential roles in theinitiation and perpetuation of chronic gut inflammation. Onesuch T-cell-associated adhesion molecule that is known to beimportant in T cell trafficking and activation is the b2 integrinlymphocyte function-associated antigen-1 (LFA-1; CD11a/CD18; aLb2). LFA-1 is a heterodimeric protein composed ofan alpha (CD11a, aL) and beta chain (CD18, b2), which isexpressed on T and B cells, granulocytes and macrophages(4). Multiple LFA-1 ligands have been identified that includethe intracellular adhesion molecules (ICAM-1–5) (5–9) and thejunctional adhesion molecule-1 (CD166) (10). Interaction ofLFA-1 with its counter receptor ICAM-1 has been shown topromote the recirculation of naive T cells to certain secondarylymphoid tissue such as peripheral lymph nodes (PLNs);however, its role in mediating migration to mesenteric lymphnodes (MLNs) is less well characterized. LFA-1 has also beendemonstrated to facilitate the interaction of T cells withantigen-presenting cells (APCs) to promote T cell activation,polarization and proliferation in vitro (11–14). In addition, thisT-cell-associated integrin is involved in the firm adhesion andtransendothelial cell migration of activated T cells to in-flammatory foci (15–18). Anecdotal evidence for a role forLFA-1 in intestinal inflammation comes from the observationthat CD4+ T cells obtained from the intestines of CD patientshave increased CD11a expression compared with cells fromhealthy controls (19–21).A number of intervention studies targeting different adhe-
sion molecules have been performed in inflammatory boweldisease (IBD) models with varying degrees of success. Forexample, ICAM-1 blocking strategies have been successful indown-regulating colonic inflammation in animal models oferosive, self-limiting IBD (22, 23). Additionally, anti-ICAMstrategies have been successful in attenuating inflammationobserved in human ulcerative colitis (24, 25). In other studies,delivery of anti-vascular cell adhesion molecule-1 andMAdCAM-1 mAbs demonstrated limited efficacy in attenuat-ing the severity of inflammation in both T-cell-independent and-dependent models of colitis (26–30). Furthermore, whiletransfer of T cells deficient in the b7 integrin subunit delayedthe onset of colitis in the CD45RBhigh transfer model of chroniccolitis (28), b7 deficiency did not influence the development ofspontaneous colitis in IL-2�/� mice (31). Surprisingly, few, ifany, studies have addressed the role of LFA-1 in thepathogenesis of chronic gut inflammation. This is surprisingsince efalizumab, a humanized anti-CD11a mAb, has proveneffective in both experimental and clinical studies for thetreatment of certain T-cell-dependent diseases such aspsoriasis (32–34).Therefore, the objective of this study was to assess the role
of LFA-1 in the pathogenesis of chronic gut inflammation in aT-cell-dependent model of chronic colitis. Recombinase-activating gene-1-deficient (RAG-1�/�) mice were reconsti-tuted with naive CD4+CD25� T cells obtained from either
wild-type (wt) or CD11a (LFA-1)-deficient donor animals. Wedemonstrate that transfer of CD11a-deficient T cells fail toinduce chronic colitis in RAG-1�/� recipient mice, whereastransfer of wt T cells induced severe colitis in the sameimmunodeficient recipients. Failure to induce colitis appearedto be due to defects in Tcell priming/activation within the MLNsthereby limiting the numbers of Th1 disease-producing cellsthat populate the colonic interstitium.
Methods
Animals
Wild-type (wt) and RAG-1�/� mice on a C57BL/6 backgroundwere purchased from the Jackson Laboratories (Bar Harbor,ME, USA). CD11a-deficient (LFA-1�/�) mice on a C57BL/6background were obtained from Christie M. Ballantyne (BaylorCollege of Medicine) and bred at the Louisiana StateUniversity (LSU) Health Sciences Centers animal facility.Animals were maintained on 12/12 h light/dark cycles instandardanimal cageswith filter tops under specificpathogen-free conditions. All animals were given standard laboratoryrodent chow and water ad libitum. All experimental proceduresinvolving the use of animals were reviewed and approved bythe Institutional Animal Care and Use Committee of LSU HealthSciences Center and performed according to the criteriaoutlined by the National Institute of Health.
Antibodies
All antibodies used in the experimental procedures werepurchased either at BD PharMingen, San Diego, CA, USA oreBioscience, San Diego, CA, USA. The following antibodiesraised against mouse antigens were used for cell isolation andflow cytometric analysis: biotin-conjugated anti-CD4 (GK1.5);FITC-conjugated anti-B220, anti-CD8a, anti-CD45RB, anti-CD44 and anti-Mac-1 (CD11b); PE-conjugated anti-CD25,anti-CD69, anti-lymphocyte Peyers patch adhesionmolecule-1(LPAM-1) (integrin a4b7 complex), anti-CD49d (integrin a4chain) and anti-CCR7; PE–Cy5-conjugated anti-CD3 and antiCD62L; allophycocyanin-conjugated anti-CD4(clone GK1.5).
T-cell transfer model of chronic colitis
Chronic colitis was induced in RAG-1�/� mice using a minormodification of a method previously described (35). Briefly,donor spleens were surgically removed from either C57BL/6wt or CD11a�/� female mice and teased into a single-cellsuspension in PBS containing 4% FBS (FACS buffer). CD4+
T cells were enriched using negative selection by first label-ing with FITC-conjugated anti-B220, anti-CD8 and anti-Mac-1and then with anti-FITCmagnetic microbeads followed by sep-aration on a type CS column using VarioMACS magneticseparator according to manufacturer’s instructions (MiltenyiBiotec, Auburn, CA, USA). Unlabeled flow-thru cells werecollected and labeled with biotin-conjugated anti-CD4(GK1.5), streptavidin and PE-conjugated anti-CD25 mAb.Cells were sorted for CD4+CD25� using FACSVantage(Becton Dickinson, San Jose, CA, USA) with >98% purity onpost-sort analysis. Male RAG-1�/� C57BL/6 mice (8- to 10-weeks old) were injected (intra-peritoneally) with 7.5 3 105
390 Role of LFA-1 in experimental colitis
by guest on February 23, 2016http://intim
m.oxfordjournals.org/
Dow
nloaded from

CD4+CD25� Tcells, from either wt or CD11a�/�, suspended in500 ll of PBS. Clinical signs of disease (e.g. loss of bodyweight and loose stool/diarrhea) were followed and recordedweekly for 8 weeks from the time of injection.
Tissue lymphocyte isolation and flow cytometric analysis
Lymphocytes were obtained from the MLNs, spleen, intestineand colon and analyzed by flow cytometry as previouslydescribed (36). Briefly, MLNs and spleens were removedaseptically and teased into a single-cell suspension using thefrosted ends of two glass slides in FACS buffer on ice. Thesuspension was then passed through a 26-gauge syringe toobtain a single-cell suspension. After pelleting, RBCs wereremoved by hypotonic lysis and the resulting leukocytes werere-suspended in FACS buffer containing anti-FcR (CD16/32)antibody at 5 3 107 cells ml�1. After incubation with anti-FcRmAb, ~1 3 106 cells were placed into individual wells ofa round bottom 96-well plate, pelleted and stained withappropriate antibodies. After the staining, cells were fixedfor 15 min on ice in freshly prepared 2% ultrapure formalde-hyde (Polysciences, Inc., Warrington, PA, USA) and analyzednext day on the FACScalibur (BD Biosciences, San Diego, CA,USA).Analysis of intestinal intra-epithelial lymphocytes (IELs) was
performed using a modification of the method describedpreviously (31). Briefly, small and large intestines wereremoved from mice flushed of luminal contents and trimmedof excess fat and connective tissue. Small and large intestineswere opened longitudinally and cut into small (0.5–1.0 cm)pieces in PBS on ice. Pieces were then incubated in pre-warmed (37�C) PBS/4% FCS/0.2 mM EDTA/10 mM D-glucoseon a rotating shaker for 20 min at 250 r.p.m. at 37�C. Afterincubation, intestinal pieces were vortexed for 3–5 s. Super-natants from individual animals were collected in separate50-ml conical tubes and kept on ice. Incubations were per-formed at least three times to insure complete removal ofepithelium. Intestinal pieces from individual animals were pro-cessedseparatelyandnevermixed.The resultantpooledsuper-natants were pelleted by centrifugation and re-suspended in30 ml of 40% Percoll (Amersham Biosciences). IELs werefurther enriched by centrifugation over a 40/70% Percollgradient for 25 min, 1000 g at room temperature. After thecentrifugation, the pellet of IELs was washed and then re-suspended in FACS buffer containing anti-FcR mAb. Viablecells were counted using 0.4% tryphan blue dye/PBS solution.Lamina propria lymphocytes (LPLs) were prepared by
digestion of finely minced intestinal pieces remaining afterIEL isolation with RPMI-1640/4% FBS and containing collage-nase type VIII (200 U ml�1) for 40 min at 250 r.p.m. in a 37�Cshaker (15). Lymphocytes were further enriched by centrifu-gation over a 40/70% Percoll gradient. The LPL pellet waswashed, re-suspended in FACS buffer containing anti-FcRmAb and counted. ~1 3 106 cells were placed in individualwells of a 96-well plate and stained. After the staining, cellswere fixed for 15 min on ice in freshly prepared 2% ultrapureformaldehyde (Polysciences, Inc.) and analyzed the next dayon the FACScalibur (BD Biosciences). Data were analyzedusing FlowJo software (Tree Star, Inc., Ashland, OR, USA;version 5.7.2 for PC). Percentages of specific subsets of
lymphocytes were compared between wt and CD11a�/�
CD4+CD25� injected RAG-1�/� animals.
Ex vivo stimulation and cytokine determinations
Mononuclear cells from spleen, MLN and colonic laminapropria (LP) of individual animals were prepared as describedabove with minor modification. Following RBC lysis, mono-nuclear leukocytes from the spleen were prepared by densitycentrifugation through Ficoll-PaqueTM PLUS (Amersham Bio-sciences). Colonic lamina propria mononuclear cells (LPMCs)were prepared as described above using a 40/70% Percolldensity gradient while a single-cell suspension of mono-nuclear cells from MLNs was used without further enrichment.All cells were washed in complete RPMI 1640 media con-taining 4% FBS, L-glutamine and 100 U ml�1 each penicillinand streptomycin and counted. A total of 53 105 mononuclearcells were plated in either control (uncoated) or CD3e-coated(BD Falcon, San Diego, CA, USA) 96-well flat bottom plates.Anti-mouse FG-purified anti-CD28 (eBioscience, 1 lg ml�1
final concentration) or medium alone (for control uncoatedplate) was added and cells were incubated for 48 h at 37�C ina humidified incubator with 10% CO2. At the end of incubation,cells were removed by centrifugation and the supernatantswere collected, frozen and stored at �70�C. Cytokine de-termination in collected supernatants was determined usingthe Inflammation and Th1/Th2 Cytometric Bead Array kits (BDBiosciences) as described by the manufacturer.
Macroscopic and histological evaluation
At 8 weeks following Tcell reconstitution, mice were euthanizedand their colons removed. Colons were cleaned of fecal ma-terial and scored for macroscopic evidence of inflammationusing a modification of the method described by Conner et al.(37). Normal colons were assigned a score of 0; mild bowelwall thickening in the absence of visible hyperemia was givena score of 1; moderate bowel wall thickening and hyperemiawas given a score of 2; severe bowel wall thickening withrigidity and marked hyperemia was assigned a score of 3 andsevere bowel thickening with rigidity, hyperemia and colonicadhesions was given a score of 4. The colons were thendivided into proximal and distal sections and each segmentwas measured and weighed for ratio comparisons as an indexof inflammation. A small piece of each section was placed in10% buffer formalin and stored overnight at 4�C. The fixedtissue was then rinsed of formalin with PBS, partially hydratedin ethanol and embedded in JB-4 medium (Polysciences, Inc.)or parafin. Samples were sectioned (5 lm) and stained withhematoxylin and eosin to evaluate standard histopathologychanges. Histopathological analysis was performed in ablinded manner and scored using a published method (35).Briefly, eight parameters were used that include (i) the degreeof inflammatory infiltrate in the LP, range 1–3; (ii) Goblet cellloss as a marker of mucin depletion, range 0–2; (iii) reactiveepithelial hyperplasia/atypia with nuclear changes, range 0–3;(iv) the number of IELs in the epithelial crypts, range 0–3;(v) abnormal crypt architecture (distortion, branching, atrophy,crypt loss), range 0–3; (vi) number of crypt abscesses, range0–2; (vii) mucosal erosion to frank ulcerations, range 0–2 and(viii) submucosal spread to transmural involvement, range 0–2.
Role of LFA-1 in experimental colitis 391
by guest on February 23, 2016http://intim
m.oxfordjournals.org/
Dow
nloaded from
The severity of inflammation in both sections of the colon wasbased on the sum of the scores in each parameter with a max-imum score of 20.
Results
CD11a-deficient T cells fail to induce chronic colitis inimmunodeficient mice
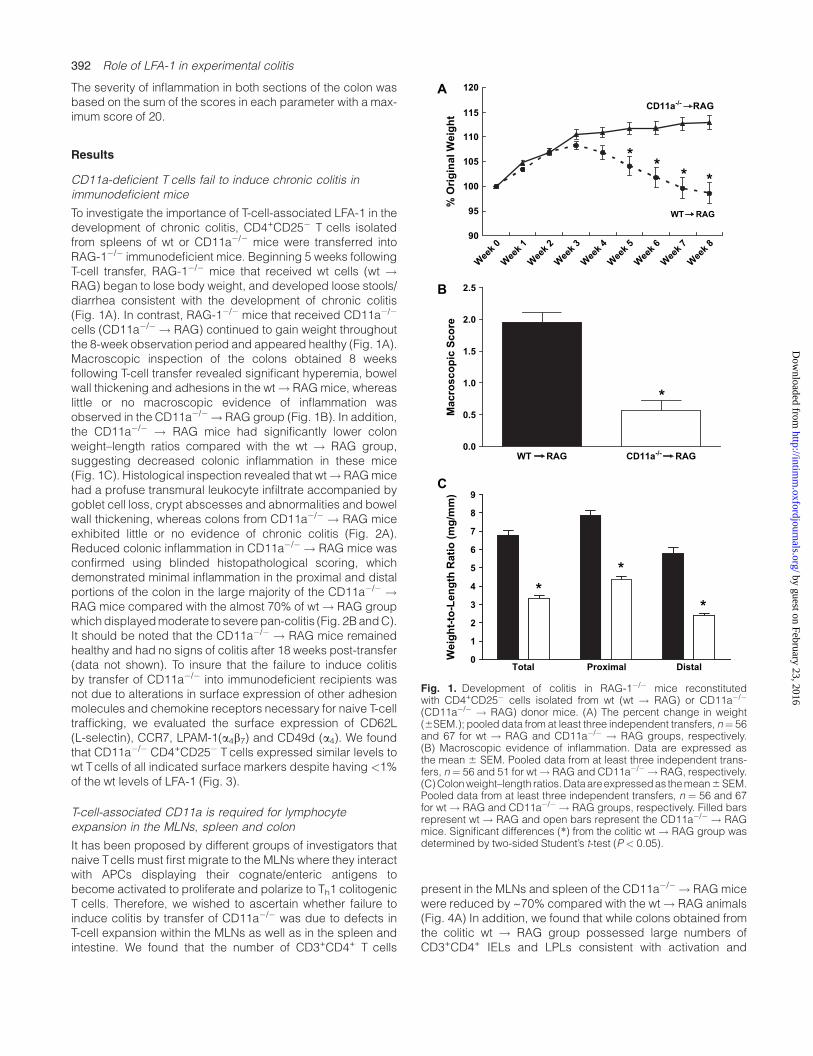
To investigate the importance of T-cell-associated LFA-1 in thedevelopment of chronic colitis, CD4+CD25� T cells isolatedfrom spleens of wt or CD11a�/� mice were transferred intoRAG-1�/� immunodeficient mice. Beginning 5 weeks followingT-cell transfer, RAG-1�/� mice that received wt cells (wt !RAG) began to lose body weight, and developed loose stools/diarrhea consistent with the development of chronic colitis(Fig. 1A). In contrast, RAG-1�/� mice that received CD11a�/�
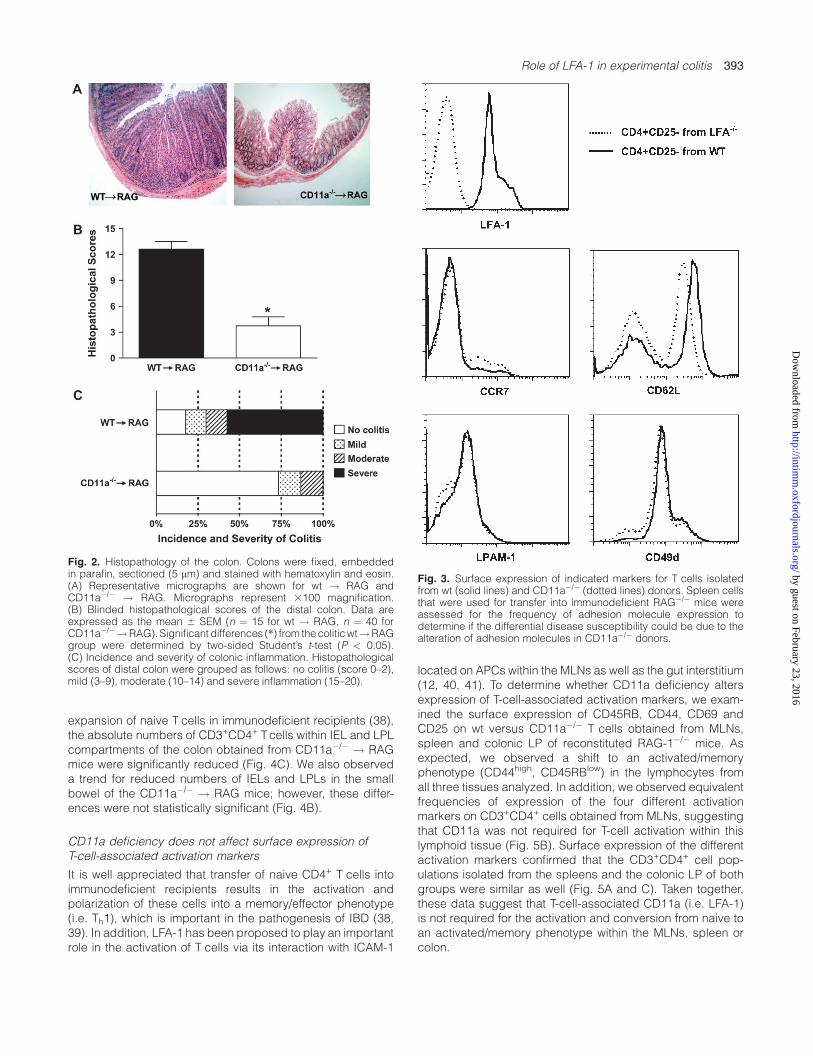
cells (CD11a�/� ! RAG) continued to gain weight throughoutthe 8-week observation period and appeared healthy (Fig. 1A).Macroscopic inspection of the colons obtained 8 weeksfollowing T-cell transfer revealed significant hyperemia, bowelwall thickening and adhesions in the wt! RAGmice, whereaslittle or no macroscopic evidence of inflammation wasobserved in the CD11a�/� ! RAG group (Fig. 1B). In addition,the CD11a�/� ! RAG mice had significantly lower colonweight–length ratios compared with the wt ! RAG group,suggesting decreased colonic inflammation in these mice(Fig. 1C). Histological inspection revealed that wt! RAGmicehad a profuse transmural leukocyte infiltrate accompanied bygoblet cell loss, crypt abscesses and abnormalities and bowelwall thickening, whereas colons from CD11a�/� ! RAG miceexhibited little or no evidence of chronic colitis (Fig. 2A).Reduced colonic inflammation in CD11a�/� ! RAG mice wasconfirmed using blinded histopathological scoring, whichdemonstrated minimal inflammation in the proximal and distalportions of the colon in the large majority of the CD11a�/� !RAG mice compared with the almost 70% of wt ! RAG groupwhichdisplayedmoderate to severepan-colitis (Fig. 2BandC).It should be noted that the CD11a�/� ! RAG mice remainedhealthy and had no signs of colitis after 18 weeks post-transfer(data not shown). To insure that the failure to induce colitisby transfer of CD11a�/� into immunodeficient recipients wasnot due to alterations in surface expression of other adhesionmolecules and chemokine receptors necessary for naive T-celltrafficking, we evaluated the surface expression of CD62L(L-selectin), CCR7, LPAM-1(a4b7) and CD49d (a4). We foundthat CD11a�/� CD4+CD25� T cells expressed similar levels towt Tcells of all indicated surface markers despite having <1%of the wt levels of LFA-1 (Fig. 3).
T-cell-associated CD11a is required for lymphocyteexpansion in the MLNs, spleen and colon
It has been proposed by different groups of investigators thatnaive Tcells must first migrate to the MLNs where they interactwith APCs displaying their cognate/enteric antigens tobecome activated to proliferate and polarize to Th1 colitogenicT cells. Therefore, we wished to ascertain whether failure toinduce colitis by transfer of CD11a�/� was due to defects inT-cell expansion within the MLNs as well as in the spleen andintestine. We found that the number of CD3+CD4+ T cells
present in the MLNs and spleen of the CD11a�/� ! RAGmicewere reduced by ~70% compared with the wt! RAG animals(Fig. 4A) In addition, we found that while colons obtained fromthe colitic wt ! RAG group possessed large numbers ofCD3+CD4+ IELs and LPLs consistent with activation and
A
B
C
0
1
2
3
4
5
6
7
8
9
Total Proximal Distal
Weig
ht-to
-L
en
gth
R
atio
(m
g/m
m)
*
*
*
Macro
sc
op
ic S
co
re
0.0
0.5
1.0
1.5
2.0
2.5
*
WT RAG CD11a-/-
RAG%
O
rig
in
al W
eig
ht
**
**
90
95
100
105
110
115
120
Week 0
Week 1
Week 2
Week 3
Week 4
Week 5
Week 6
Week 7
Week 8
CD11a-/-
RAG
WT RAG
Fig. 1. Development of colitis in RAG-1�/� mice reconstitutedwith CD4+CD25� cells isolated from wt (wt ! RAG) or CD11a�/�
(CD11a�/� ! RAG) donor mice. (A) The percent change in weight(6SEM.); pooled data from at least three independent transfers, n ¼ 56and 67 for wt ! RAG and CD11a�/� ! RAG groups, respectively.(B) Macroscopic evidence of inflammation. Data are expressed asthe mean 6 SEM. Pooled data from at least three independent trans-fers, n ¼ 56 and 51 for wt! RAG and CD11a�/� ! RAG, respectively.(C)Colonweight–length ratios.Dataareexpressedas themean6SEM.Pooled data from at least three independent transfers, n ¼ 56 and 67for wt ! RAG and CD11a�/� ! RAG groups, respectively. Filled barsrepresent wt ! RAG and open bars represent the CD11a�/� ! RAGmice. Significant differences (*) from the colitic wt ! RAG group wasdetermined by two-sided Student’s t-test (P < 0.05).
392 Role of LFA-1 in experimental colitis
by guest on February 23, 2016http://intim
m.oxfordjournals.org/
Dow
nloaded from
expansion of naive T cells in immunodeficient recipients (38),the absolute numbers of CD3+CD4+ Tcells within IEL and LPLcompartments of the colon obtained from CD11a�/� ! RAGmice were significantly reduced (Fig. 4C). We also observeda trend for reduced numbers of IELs and LPLs in the smallbowel of the CD11a�/� ! RAG mice; however, these differ-ences were not statistically significant (Fig. 4B).
CD11a deficiency does not affect surface expression ofT-cell-associated activation markers
It is well appreciated that transfer of naive CD4+ T cells intoimmunodeficient recipients results in the activation andpolarization of these cells into a memory/effector phenotype(i.e. Th1), which is important in the pathogenesis of IBD (38,39). In addition, LFA-1 has been proposed to play an importantrole in the activation of T cells via its interaction with ICAM-1
located on APCs within the MLNs as well as the gut interstitium(12, 40, 41). To determine whether CD11a deficiency altersexpression of T-cell-associated activation markers, we exam-ined the surface expression of CD45RB, CD44, CD69 andCD25 on wt versus CD11a�/� T cells obtained from MLNs,spleen and colonic LP of reconstituted RAG-1�/� mice. Asexpected, we observed a shift to an activated/memoryphenotype (CD44high, CD45RBlow) in the lymphocytes fromall three tissues analyzed. In addition, we observed equivalentfrequencies of expression of the four different activationmarkers on CD3+CD4+ cells obtained from MLNs, suggestingthat CD11a was not required for T-cell activation within thislymphoid tissue (Fig. 5B). Surface expression of the differentactivation markers confirmed that the CD3+CD4+ cell pop-ulations isolated from the spleens and the colonic LP of bothgroups were similar as well (Fig. 5A and C). Taken together,these data suggest that T-cell-associated CD11a (i.e. LFA-1)is not required for the activation and conversion from naive toan activated/memory phenotype within the MLNs, spleen orcolon.
Fig. 2. Histopathology of the colon. Colons were fixed, embeddedin parafin, sectioned (5 lm) and stained with hematoxylin and eosin.(A) Representative micrographs are shown for wt ! RAG andCD11a�/� ! RAG. Micrographs represent 3100 magnification.(B) Blinded histopathological scores of the distal colon. Data areexpressed as the mean 6 SEM (n ¼ 15 for wt ! RAG, n ¼ 40 forCD11a�/�!RAG). Significant differences (*) from thecoliticwt!RAGgroup were determined by two-sided Student’s t-test (P < 0.05).(C) Incidence and severity of colonic inflammation. Histopathologicalscores of distal colon were grouped as follows: no colitis (score 0–2),mild (3–9), moderate (10–14) and severe inflammation (15–20).
Fig. 3. Surface expression of indicated markers for T cells isolatedfrom wt (solid lines) and CD11a�/� (dotted lines) donors. Spleen cellsthat were used for transfer into immunodeficient RAG�/� mice wereassessed for the frequency of adhesion molecule expression todetermine if the differential disease susceptibility could be due to thealteration of adhesion molecules in CD11a�/� donors.
Role of LFA-1 in experimental colitis 393
by guest on February 23, 2016http://intim
m.oxfordjournals.org/
Dow
nloaded from
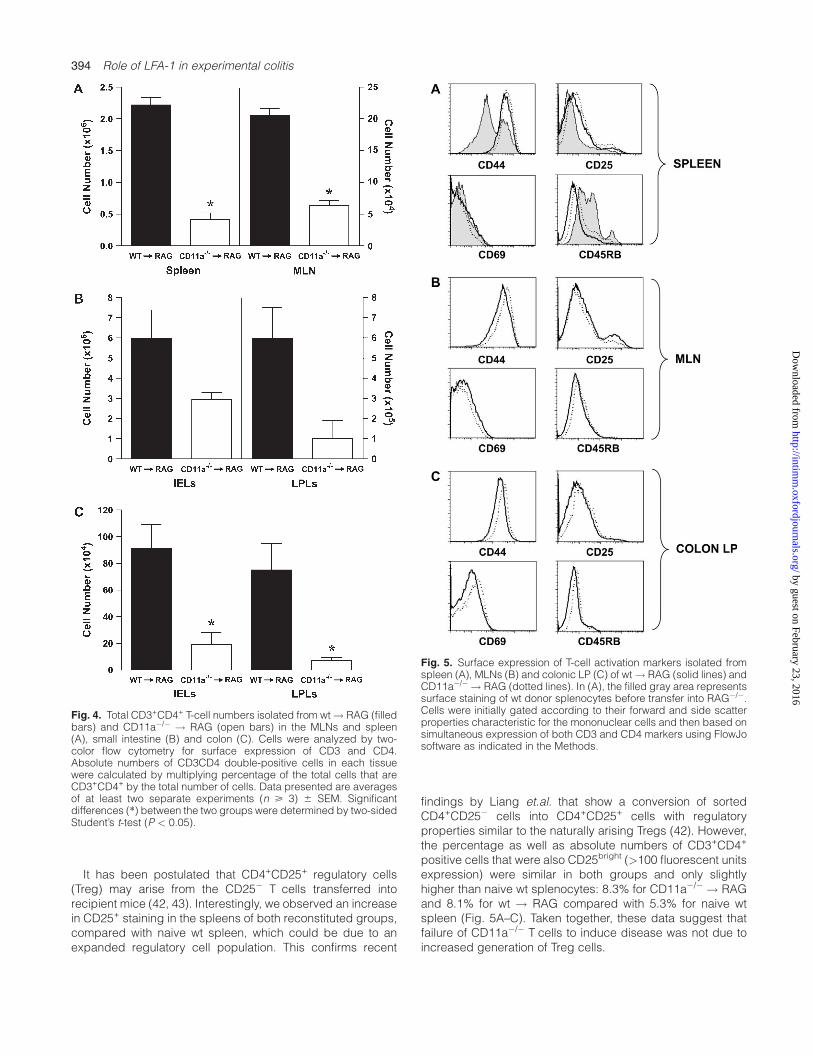
It has been postulated that CD4+CD25+ regulatory cells(Treg) may arise from the CD25� T cells transferred intorecipient mice (42, 43). Interestingly, we observed an increasein CD25+ staining in the spleens of both reconstituted groups,compared with naive wt spleen, which could be due to anexpanded regulatory cell population. This confirms recent
findings by Liang et.al. that show a conversion of sortedCD4+CD25� cells into CD4+CD25+ cells with regulatoryproperties similar to the naturally arising Tregs (42). However,the percentage as well as absolute numbers of CD3+CD4+
positive cells that were also CD25bright (>100 fluorescent unitsexpression) were similar in both groups and only slightlyhigher than naive wt splenocytes: 8.3% for CD11a�/� ! RAGand 8.1% for wt ! RAG compared with 5.3% for naive wtspleen (Fig. 5A–C). Taken together, these data suggest thatfailure of CD11a�/� T cells to induce disease was not due toincreased generation of Treg cells.
Fig. 4. Total CD3+CD4+ T-cell numbers isolated from wt! RAG (filledbars) and CD11a�/� ! RAG (open bars) in the MLNs and spleen(A), small intestine (B) and colon (C). Cells were analyzed by two-color flow cytometry for surface expression of CD3 and CD4.Absolute numbers of CD3CD4 double-positive cells in each tissuewere calculated by multiplying percentage of the total cells that areCD3+CD4+ by the total number of cells. Data presented are averagesof at least two separate experiments (n > 3) 6 SEM. Significantdifferences (*) between the two groups were determined by two-sidedStudent’s t-test (P < 0.05).
Fig. 5. Surface expression of T-cell activation markers isolated fromspleen (A), MLNs (B) and colonic LP (C) of wt! RAG (solid lines) andCD11a�/� ! RAG (dotted lines). In (A), the filled gray area representssurface staining of wt donor splenocytes before transfer into RAG�/�.Cells were initially gated according to their forward and side scatterproperties characteristic for the mononuclear cells and then based onsimultaneous expression of both CD3 and CD4 markers using FlowJosoftware as indicated in the Methods.
394 Role of LFA-1 in experimental colitis
by guest on February 23, 2016http://intim
m.oxfordjournals.org/
Dow
nloaded from
Mononuclear cells obtained from CD11a�/� ! RAG micehave impaired cytokine generation
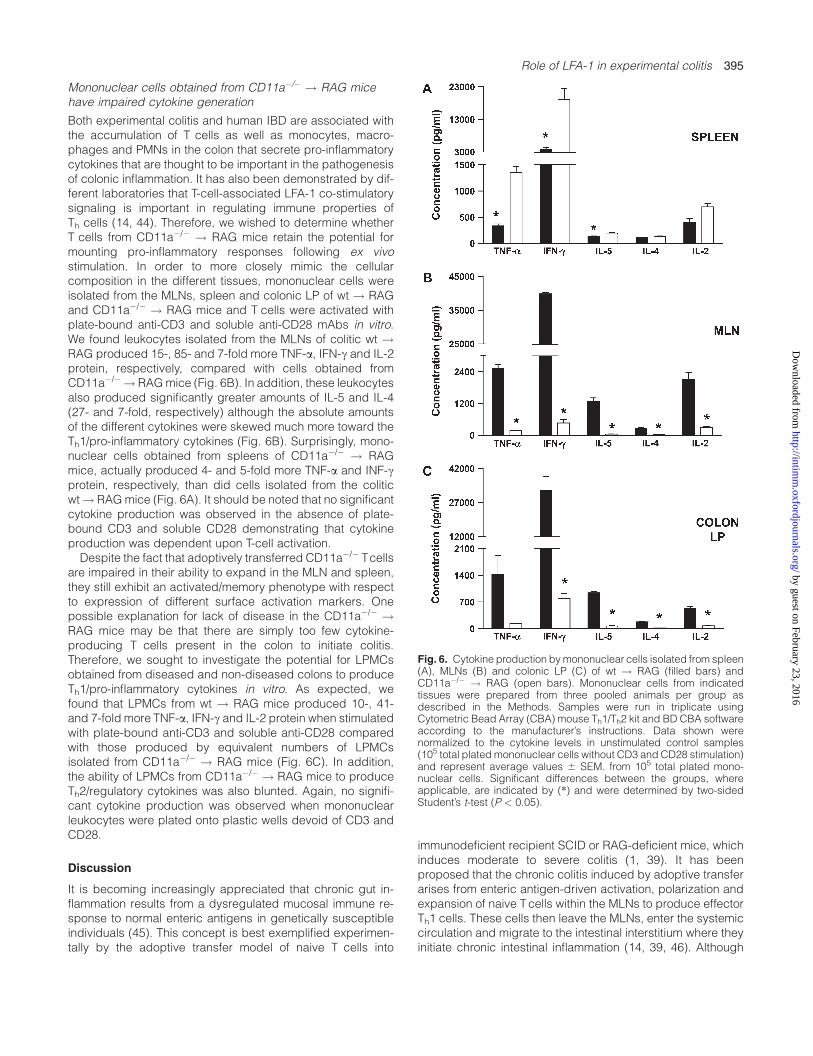
Both experimental colitis and human IBD are associated withthe accumulation of T cells as well as monocytes, macro-phages and PMNs in the colon that secrete pro-inflammatorycytokines that are thought to be important in the pathogenesisof colonic inflammation. It has also been demonstrated by dif-ferent laboratories that T-cell-associated LFA-1 co-stimulatorysignaling is important in regulating immune properties ofTh cells (14, 44). Therefore, we wished to determine whetherT cells from CD11a�/� ! RAG mice retain the potential formounting pro-inflammatory responses following ex vivostimulation. In order to more closely mimic the cellularcomposition in the different tissues, mononuclear cells wereisolated from the MLNs, spleen and colonic LP of wt ! RAGand CD11a�/� ! RAG mice and T cells were activated withplate-bound anti-CD3 and soluble anti-CD28 mAbs in vitro.We found leukocytes isolated from the MLNs of colitic wt !RAG produced 15-, 85- and 7-fold more TNF-a, IFN-c and IL-2protein, respectively, compared with cells obtained fromCD11a�/�! RAGmice (Fig. 6B). In addition, these leukocytesalso produced significantly greater amounts of IL-5 and IL-4(27- and 7-fold, respectively) although the absolute amountsof the different cytokines were skewed much more toward theTh1/pro-inflammatory cytokines (Fig. 6B). Surprisingly, mono-nuclear cells obtained from spleens of CD11a�/� ! RAGmice, actually produced 4- and 5-fold more TNF-a and INF-cprotein, respectively, than did cells isolated from the coliticwt! RAGmice (Fig. 6A). It should be noted that no significantcytokine production was observed in the absence of plate-bound CD3 and soluble CD28 demonstrating that cytokineproduction was dependent upon T-cell activation.Despite the fact that adoptively transferred CD11a�/� Tcells
are impaired in their ability to expand in the MLN and spleen,they still exhibit an activated/memory phenotype with respectto expression of different surface activation markers. Onepossible explanation for lack of disease in the CD11a�/� !RAG mice may be that there are simply too few cytokine-producing T cells present in the colon to initiate colitis.Therefore, we sought to investigate the potential for LPMCsobtained from diseased and non-diseased colons to produceTh1/pro-inflammatory cytokines in vitro. As expected, wefound that LPMCs from wt ! RAG mice produced 10-, 41-and 7-fold more TNF-a, IFN-c and IL-2 protein when stimulatedwith plate-bound anti-CD3 and soluble anti-CD28 comparedwith those produced by equivalent numbers of LPMCsisolated from CD11a�/� ! RAG mice (Fig. 6C). In addition,the ability of LPMCs from CD11a�/� ! RAG mice to produceTh2/regulatory cytokines was also blunted. Again, no signifi-cant cytokine production was observed when mononuclearleukocytes were plated onto plastic wells devoid of CD3 andCD28.
Discussion
It is becoming increasingly appreciated that chronic gut in-flammation results from a dysregulated mucosal immune re-sponse to normal enteric antigens in genetically susceptibleindividuals (45). This concept is best exemplified experimen-tally by the adoptive transfer model of naive T cells into
immunodeficient recipient SCID or RAG-deficient mice, whichinduces moderate to severe colitis (1, 39). It has beenproposed that the chronic colitis induced by adoptive transferarises from enteric antigen-driven activation, polarization andexpansion of naive Tcells within the MLNs to produce effectorTh1 cells. These cells then leave the MLNs, enter the systemiccirculation and migrate to the intestinal interstitium where theyinitiate chronic intestinal inflammation (14, 39, 46). Although
Fig. 6. Cytokine production bymononuclear cells isolated from spleen(A), MLNs (B) and colonic LP (C) of wt ! RAG (filled bars) andCD11a�/� ! RAG (open bars). Mononuclear cells from indicatedtissues were prepared from three pooled animals per group asdescribed in the Methods. Samples were run in triplicate usingCytometric Bead Array (CBA) mouse Th1/Th2 kit and BD CBA softwareaccording to the manufacturer’s instructions. Data shown werenormalized to the cytokine levels in unstimulated control samples(105 total plated mononuclear cells without CD3 and CD28 stimulation)and represent average values 6 SEM. from 105 total plated mono-nuclear cells. Significant differences between the groups, whereapplicable, are indicated by (*) and were determined by two-sidedStudent’s t-test (P < 0.05).
Role of LFA-1 in experimental colitis 395
by guest on February 23, 2016http://intim
m.oxfordjournals.org/
Dow
nloaded from

recognition and interaction of the MHC Class II-bound antigenby the TCR is critical for the initial immune response within theMLN, a variety of different adhesion molecules localized onT cells are also required for co-stimulatory signals as well ascell–cell interactions. One adhesion molecule thought to playa critical role in T-cell biology yet has received little attention inimmune-based models of IBD is the b2 integrin LFA-1 (47). Ithas been shown that LFA-1 plays an important role in severalaspects of cell-mediated immunity including migration (re-circulation) to and activation/expansion within MLNs as well asthe recruitment of activated/memory T cells to inflamed tissue(48). Data obtained in the current study demonstrate thattransfer of CD11a�/� CD4+CD25� Tcells into immunodeficientRAG-1�/� mice fails to induce chronic colitis, whereas transferof wt T cells induced moderate to severe gut inflammation.Failure to induce colonic inflammation correlatedwith a paucityof CD3+CD4+ T cells within the MLNs, colonic interstitium andspleen. These data are consistent with the proposal that LFA-1is critical for migration and/or priming within the MLN and thesubsequent recruitment of these T cells into the colonicinterstitium to initiate disease. Recent studies have shownthat expression of different adhesion molecules can influencethe inhibitory nature of anti-LFA treatment using in vitroadhesion assays, supporting an accessory role for LFA-1 intrafficking of naive Tcells to some secondary lymphoid tissues(49). It is well documented for example that LFA-1 is importantfor the migration (i.e. recirculation) of naive T cells from theblood into PLNs (40, 50, 49). However, these studies alsodemonstrated that LFA-1 had only ‘a limited role’ in thehoming of lymphocytes to MLNs and Peyer’s patches and noeffect on homing to the spleen. This accessory role of LFA-1 issupported by the findings that the a4 integrins (a4b1 and a4b7)are important for the migration of lymphocytes into certainsecondary lymphoid tissue (e.g. MLNs) and can compensatefor the loss of LFA-1 (51, 52). These data coupled to the factthat CD62L, CCR7, a4 and a4b7 expression on CD11a�/� werevirtually identical to those observed on wt CD4+CD25� donorTcells suggest that CD11a�/� Tcells were able to migrate intoMLNs via T-cell-associated a4 integrins, CCR7 and possiblyCD62L (i.e. L-selectin). Consequently, we would propose thatthe paucity of lymphocytes in the MLNs and colonic tissue ofCD11a�/� ! RAGmice versus wt! RAGmice may be due todefects in the initial T-cell priming/activation and consequentexpansion within the MLNs.The extent of the primary response of naive CD4 T cells,
which involves activation, expansion and polarization, dictatesif a proper immune response is mounted to a specific antigen.Activation of T cells requires two signals; the first signal isantigen dependent, involving the interaction of the antigen–MHC on an APC and the TCR, while the second signal isantigen independent, involving multiple T-cell surface mol-ecules and their ligands on APCs. In particular, LFA-1/ICAMinteractions are important for optimal T-cell activation bymediating and stabilizing the T-cell-APC contact referred to asthe immunologic synapse (53). LFA-1 on T cells is thought tolower the threshold level of activation and promote proliferationof T cells. The generalized lack of T-cell expansion in multipletissues, including the spleen, would appear to support theidea that LFA-1 is more important for priming and expansionthan for trafficking to the MLNs (Fig. 4). These in vivo data
agree well with other investigators who have demonstratedthat LFA-1�/� lymphocytes have major defects in alloantigen(MLR)- or Con A-stimulated proliferation in vitro (15, 54). Inaddition to priming within the MLNs, our data would suggestthat T-cell-associated LFA-1 is important for Th1 cytokineproduction as T cells obtained from MLNs and colonic LP ofCD11a�/� ! RAG produced much smaller amounts of IFN-cand/or TNF-a following ex vivo stimulation with plate-boundCD3 and soluble CD28 compared with the colitic wt ! RAGanimals (Fig. 6). It could be argued that the cytokine profilesobserved in our ex vivo studies were due to non-specificactivation of the different cell types (Tcells, macrophages anddendritic cells) found in our mononuclear cell preparations ofMLN, spleen and colon. However, as noted previously, nocytokine generation could be observed by cells from anytissue in the absence of CD3 and CD28, suggesting that T-cellactivation was required for cytokine synthesis. Although it ispossible that certain T-cell-derived cytokines (e.g. IFN-c)could influence the production of macrophage and/or den-dritic cell cytokines during the incubation period, the factremains that Th1 cytokines were depressed to a much greaterextent in cells derived from the CD11a�/� ! RAG mice thantheir colitic counterparts. These data are interesting for tworeasons. First, although we observed large and significantreductions in IFN-c and/or TNF-a production by cells obtainedfrom CD11a�/� ! RAG MLNs and colon, T-cell activationmarker expression was consistent with activated/antigen-experienced cells and virtually identical to those obtainedfrom the colitic wt! RAG animals, suggesting that expressionof activation markers may be dissociated from cell proliferationand cytokine expression. A second interesting observationmade in these studies is that colonic LPMCs isolated fromCD11a�/� ! RAG animals appeared to be deactivated i.e.they appeared to be refractory in their ability to produce bothTh1 and Th2 cytokines following TCR activation. These datasuggest the novel finding that LFA-1 is required not only forpriming within the MLNs but is also required for ‘secondary’activation of T cells that have gained access to the gutinterstitium.One possible mechanism to account for reduced cytokine
production by MLNs and LPMCs isolated from CD11a�/� !RAG mice is that lack of LFA-1 on T cells somehow skews thepolarization of T cells to a Th2 and/or regulatory phenotype.Indeed, it is known that blocking the interaction of LFA-1 withICAM-1 expressed on APCs inhibits Th1 polarization andcytokine production (12, 44, 55). In fact, blocking of bothICAM-1 and ICAM-2 in the interaction of naive T cells withAPCs induces a >100-fold increase in the production ofTh2 cytokines, IL-4 and IL-5, which will down-regulate Th1-dependent inflammation (44). Our data would suggest thatthis is not the case as we found that although the production ofTh1 cytokines from MLNs and LPMCs of CD11a�/� ! RAGmice were significantly reduced compared with their coliticcounterparts, the production of Th2/regulatory cytokines wasalmost undetectable. In addition, we observed that thenumbers of CD4+CD25+ T cells isolated from the MLNs,spleen and gut were very small and were not different fromthose isolated from tissues of the colitic wt ! RAG mice.Another unexpected finding in the current study was thatsplenic mononuclear cells obtained from CD11a�/� ! RAG
396 Role of LFA-1 in experimental colitis
by guest on February 23, 2016http://intim
m.oxfordjournals.org/
Dow
nloaded from

mice produced significantly larger amounts of IFN-c and TNF-acompared with cells obtained from wt ! RAG mice. Themechanisms responsible for this rather surprising finding werenot delineated in the current study; however, it may be thata larger percentage of the mononuclear cells in the spleenpreparation were IFN-c- and TNF-a-producing T cells and/orNK cells. This exaggerated cytokine production profile raisesthe interesting possibility that extra-intestinal tissue inflamma-tion (i.e. lung, liver, spleen) may be present in the CD11a�/� !RAG mice. Although possible, the animals appeared healthyand gained weight throughout the 8- to 10-week experiment.In addition to recirculation and activation, LFA-1 is known to
be important for the firm adhesion of activated lymphocytes tothe post-capillary microvascular endothelium (56–58). Dinget al. (59) showed that T-cell adhesion and migration acrossstimulated endothelium (e.g. TNF-a, INF-c and MIP-1a) is me-diated by LFA-1 and not VLA-4. However, LFA-1-independentpathways exist and it has also been shown that humanlymphocytes can migrate across stromal cell-derived factor-1stimulated human umbilical vein endothelial cells monolayersindependent of both VLA-4 and LFA-1 (56, 59, 60). Therefore,it is reasonable to propose that specific T-cell populationsutilize several adhesion pathways to migrate from theperipheral circulation into the tissue after they have adheredto the endothelium. Additional adhesion molecules may beable to facilitate LFA-1-deficient T-cell migration across theendothelium and thus explaining the small but significantnumbers of Tcells in the colon of CD11a�/� ! RAG mice. Theobservation of decreased numbers of T cells in the colons ofthese mice is consistent with previous findings using a T-cell-dependent delayed type sensitivity model, which showed thatinfiltration of LFA-1�/� lymphocytes injected into wt mice wasreduced by 50% in ears sensitized with oxazolone (50). Sincethe isolation of T cells from the colons of CD11a�/� ! RAGmice was performed at one specific point in time, it is possiblethat these cells were recruited to the colonic interstitium innumbers similar to those observed in colitic wt! RAGmice atan earlier time but were rendered either anergic or unable tobe retained within the interstitium. Data obtained in the currentstudy demonstrate that LPMCs isolated from the colons ofCD11a�/� ! RAG mice may exist in a ‘deactivated’ orrefractory state, suggesting that LFA-1 is critical for secondaryactivation within the tissue. Current studies are underway toaddress these potential mechanisms.In summary, we have shown that T-cell-associated LFA-1 is
necessary for the induction of chronic colonic inflammationin a T-cell-dependent mouse model of Crohn’s colitis. Ourfindings indicate that T-cell-associated LFA-1 is critical for thepriming/activation, expansion and polarization of naive T cellswithin the MLNs. In addition, our data suggest that LFA-1 isalso important for secondary activation of T cells gainingaccess to the colonic interstitium. We propose that LFA-1 mayrepresent an important target for immune therapy for patientswith IBD.
Acknowledgements
This work was supported by DK64023, the Arthritis Center ofExcellence at LSU Health Sciences Center and the YamanouchiUSA Foundation.
Abbreviations
APC antigen-presenting cellCCR7 chemokine receptor-7CD Crohn’s diseaseIBD inflammatory bowel diseaseICAM intracellular adhesion moleculeIEL intra-epithelial lymphocyteLFA-1 lymphocyte function-associated antigen-1LP lamina propriaLPAM-1 lymphocyte Peyers patch adhesion molecule-1LPL lamina propria lymphocyteLPMC lamina propria mononuclear cellLSU Louisiana State UniversityMLN mesenteric lymph nodePLN peripheral lymph nodeRAG-1�/� recombinase-activating gene-1 deficientTNF-a tumor necrosis factor-awt wild type
References
1 Powrie, F. andLeach,M.W.1995.Geneticandspontaneousmodelsof inflammatory bowel disease in rodents: evidence for abnormal-ities in mucosal immune regulation. Ther. Immunol. 2:115.
2 Groux, H. and Powrie, F. 1999. Regulatory Tcells and inflammatorybowel disease. Immunol. Today 20:442.
3 Laroux, F. S. and Grisham, M. B. 2001. Immunological basis ofinflammatory bowel disease: role of the microcirculation. Microcir-culation 8:283.
4 Springer, T. A., Dustin, M. L., Kishimoto, T. K. and Marlin, S. D.1987. The lymphocyte function-associated LFA-1, CD2, and LFA-3molecules: cell adhesion receptors of the immune system. Annu.Rev. Immunol. 5:223.
5 Butini, L., de Fougerolles, A. R., Vaccarezza, M. et al. 1994.Intercellular adhesion molecules (ICAM)-1 ICAM-2 and ICAM-3function as counter-receptors for lymphocyte function-associatedmolecule 1 in human immunodeficiency virus-mediated syncytiaformation. Eur. J. Immunol. 24:2191.
6 de Fougerolles, A. R. and Springer, T. A. 1992. Intercellularadhesion molecule 3, a third adhesion counter-receptor forlymphocyte function-associated molecule 1 on resting lympho-cytes. J. Exp. Med. 175:185.
7 Klickstein, L. B., York, M. R., Fougerolles, A. R. and Springer, T. A.1996. Localization of the binding site on intercellular adhesionmolecule-3 (ICAM-3) for lymphocyte function-associated antigen 1(LFA-1). J. Biol. Chem. 271:23920.
8 Bailly, P., Tontti, E., Hermand, P., Cartron, J. P. and Gahmberg,C. G. 1995. The red cell LW blood group protein is an intercellularadhesionmolecule which binds to CD11/CD18 leukocyte integrins.Eur. J. Immunol. 25:3316.
9 Yoshihara, Y., Oka, S., Nemoto, Y. et al. 1994. An ICAM-relatedneuronal glycoprotein, telencephalin, with brain segment-specificexpression. Neuron 12:541.
10 Ostermann,G.,Weber, K. S., Zernecke,A., Schroder, A. andWeber,C. 2002. JAM-1 is a ligand of the beta(2) integrin LFA-1 involved intransendothelial migration of leukocytes. Nat. Immunol. 3:151.
11 Abraham, C., Griffith, J. and Miller, J. 1999. The dependence forleukocyte function-associated antigen-1/ICAM-1 interactions inTcell activation cannot be overcome by expression of high densityTCR ligand. J. Immunol. 162:4399.
12 Perez, O. D., Mitchell, D., Jager, G. C. et al. 2003. Leukocytefunctional antigen 1 lowers T cell activation thresholds and sig-naling through cytohesin-1 and Jun-activating binding protein 1.Nat. Immunol. 4:1083.
13 Ragazzo, J. L., Ozaki, M. E., Karlsson, L., Peterson, P. A. andWebb,S.R. 2001.Costimulation via lymphocyte function-associatedantigen 1 in the absence of CD28 ligation promotes anergy of naiveCD4+ Tcells. Proc. Natl Acad. Sci.USA 98:241.
14 Bachmann, M. F., McKall-Faienza, K., Schmits, R. et al. 1997.Distinct roles for LFA-1 and CD28 during activation of naive Tcells:adhesion versus costimulation. Immunity 7:549.
Role of LFA-1 in experimental colitis 397
by guest on February 23, 2016http://intim
m.oxfordjournals.org/
Dow
nloaded from

15 Schmits, R., Kundig, T. M., Baker, D. M. et al. 1996. LFA-1-deficientmice show normal CTL responses to virus but fail to rejectimmunogenic tumor. J. Exp. Med. 183:1415.
16 Springer, T. A. 1994. Traffic signals for lymphocyte recirculationand leukocyte emigration: the multistep paradigm. Cell 76:301.
17 Sligh, J. E., Jr, Ballantyne, C. M., Rich, S. S. et al. 1993. In-flammatory and immune responses are impaired in mice deficientin intercellular adhesion molecule 1. Proc. Natl Acad. Sci. USA90:8529.
18 Lehmann, J. C., Jablonski-Westrich, D., Haubold, U., Gutierrez-Ramos, J. C., Springer, T. and Hamann, A. 2003. Overlapping andselective roles of endothelial intercellular adhesion molecule-1(ICAM-1) and ICAM-2 in lymphocyte trafficking. J. Immunol.171:2588.
19 Bernstein, C. N., Sargent, M. and Rector, E. 2002. Alteration inexpression of beta 2 integrins on lamina propria lymphocytes inulcerative colitis and Crohn’s disease. Clin. Immunol. 104:67.
20 Kirman, I. and Nielsen, O. H. 1996. LFA-1 subunit expression inulcerative colitis patients. Dig. Dis. Sci. 41:670.
21 Vainer, B., Nielsen, O. H. and Horn, T. 2000. Comparative studies ofthe colonic in situ expression of intercellular adhesion molecules(ICAM-1, -2, and -3), beta2 integrins (LFA-1, Mac-1, and p150,95),and PECAM-1 in ulcerative colitis and Crohn’s disease. Am. J.Surg. Pathol. 24:1115.
22 Bendjelloul, F., Maly, P., Mandys, V. et al. 2000. Intercellular adhesionmolecule-1 (ICAM-1) deficiency protects mice against severe formsof experimentally induced colitis. Clin. Exp. Immunol. 119:57.
23 Bennett, C. F., Kornbrust, D., Henry, S. et al. 1997. An ICAM-1antisense oligonucleotide prevents and reverses dextran sulfatesodium-induced colitis in mice. J. Pharmacol. Exp. Ther. 280:988.
24 van Assche, G. and Rutgeerts, P. 2002. Antiadhesion moleculetherapy in inflammatory bowel disease. Inflamm. Bowel Dis. 8:291
25 van Deventer, S. J., Tami, J. A. and Wedel, M. K. 2004. Arandomised, controlled, double blind, escalating dose study ofalicaforsen enema in active ulcerative colitis. Gut 53:1646.
26 Hesterberg, P. E., Winsor-Hines, D., Briskin, M. J. et al. 1996. Rapidresolution of chronic colitis in the cotton-top tamarin with anantibody to a gut-homing integrin alpha 4 beta 7. Gastroenterology111:1373.
27 Podolsky, D. K., Lobb, R., King, N. et al. 1993. Attenuation of colitisin the cotton-top tamarin by anti-alpha 4 integrin monoclonalantibody. J. Clin. Invest. 92:372.
28 Picarella, D., Hurlbut, P., Rottman, J., Shi, X., Butcher, E. andRingler, D. J. 1997. Monoclonal antibodies specific for beta 7integrin and mucosal addressin cell adhesion molecule-1(MAdCAM-1) reduce inflammation in the colon of scid mice recon-stituted with CD45RBhigh CD4+ T cells. J. Immunol. 158:2099.
29 Burns, R. C., Rivera-Nieves, J., Moskaluk, C. A., Matsumoto, S.,Cominelli, F. and Ley, K. 2001. Antibody blockade of ICAM-1 andVCAM-1 ameliorates inflammation in the SAMP-1/Yit adoptive trans-fer model of Crohn’s disease in mice. Gastroenterology 121:1428.
30 Soriano, A., Salas, A., Salas, A. et al. 2000. VCAM-1, but notICAM-1orMAdCAM-1, immunoblockadeamelioratesDSS-inducedcolitis in mice. Lab. Invest. 80:1541.
31 Sydora, B. C., Wagner, N., Lohler, J. et al. 2002. beta7 Integrinexpression is not required for the localization of T cells to theintestine and colitis pathogenesis. Clin. Exp. Immunol. 129:35.
32 Dedrick, R. L., Walicke, P. and Garovoy, M. 2002. Anti-adhesionantibodies—efalizumab, a humanized anti-CD11a monoclonalantibody. Transpl. Immunol. 9:181.
33 Gottlieb, A. B., Krueger, J. G., Wittkowski, K., Dedrick, R., Walicke,P. A. and Garovoy, M. 2002. Psoriasis as a model for T-cell-mediated disease—immunobiologic and clinical effects of treat-ment with multiple doses of efalizumab, an anti-CD11a antibody.Arch. Dermatol. 138:591.
34 Lebwohl, M., Tyring, S. K., Hamilton, T. K. et al. 2003. A novel tar-geted T-cell modulator, efalizumab, for plaque psoriasis. N. Engl.J. Med. 349:2004.
35 Laroux, F. S., Norris, H. H., Houghton, J. et al. 2004. Regulation ofchronic colitis in athymic nu/nu (nude) mice. Int. Immunol. 16:77.
36 Ostanin, D. V., Pavlick, K. P., Bharwani, S. et al. 2006. T-cell-inducedinflammation of the small and large intestine in immuno-deficientmice. Am. J .Physiol. Gastrointest. Liver Physiol., 289:G109.
37 Conner, E. M., Brand, S., Davis, J. M. et al. 1997. Proteasomeinhibition attenuates nitric oxide synthase expression, VCAM-1transcription and the development of chronic colitis. J. Pharmacol.Exp. Ther. 282:1615.
38 Aranda, R., Sydora, B. C., McAllister, P. L. et al. 1997. Analysis ofintestinal lymphocytes in mouse colitis mediated by transfer ofCD4+, CD45RBhigh T cells to SCID recipients. J. Immunol.158:3464.
39 Powrie, F., Mauze, S. and Coffman, R. L. 1997. CD4+ T-cells in theregulation of inflammatory responses in the intestine. Res.Immunol. 148:576.
40 Warnock, R. A., Askari, S., Butcher, E. C. and von Andrian, U. H.1998. Molecular mechanisms of lymphocyte homing to peripherallymph nodes. J. Exp. Med. 187:205.
41 Shimizu, Y. 2003. LFA-1: more than just Tcell velcro. Nat. Immunol.4:1052.
42 Liang, S., Alard, P., Zhao, Y., Parnell, S., Clark, S. L. and Kosiewicz,M. M. 2005. Conversion of CD4+. J. Exp. Med. 201:127.
43 Curotto de Lafaille, M. A., Lino, A. C., Kutchukhidze, N. andLafaille, J. J. 2004. J. Immunol. 173:7259.
44 Salomon,B.andBluestone,J.A.1998. LFA-1 interactionwith ICAM-1and ICAM-2 regulates Th2 cytokine production. J. Immunol. 161:5138.
45 Singh, B., Read, S., Asseman, C. et al. 2001. Control of intestinalinflammation by regulatory T cells. Immunol. Rev. 182:190.
46 Powrie, F. 1995. T cells in inflammatory bowel disease: protectiveand pathogenic roles. Immunity 3:171.
47 Anderson, D. C. and Springer, T. A. 1987. Leukocyte adhesiondeficiency: an inherited defect in the Mac-1, LFA-1, and p150,95glycoproteins. Annu. Rev. Med. 38:175.
48 Pribila, J. T., Quale, A. C., Mueller, K. L. and Shimizu, Y. 2004.Integrins and Tcell-mediated immunity.Annu. Rev. Immunol. 22:157.
49 Hamann, A., Jablonski-Westrich, D., Duijvestijn, A. et al. 1988.Evidence for an accessory role of LFA-1 in lymphocyte-highendothelium interaction during homing. J. Immunol. 140:693.
50 Andrew, D. P., Spellberg, J. P., Takimoto, H., Schmits, R., Mak, T. W.and Zukowski, M. M. 1998. Transendothelial migration andtrafficking of leukocytes in LFA-1-deficient mice. Eur. J. Immunol.28:1959.
51 Berlin-Rufenach, C., Otto, F., Mathies, M. et al. 1999. Lymphocytemigration in lymphocyte function-associated antigen (LFA)-1-deficient mice. J. Exp. Med. 189:1467.
52 Issekutz, T. B. 1991. Inhibition of in vivo lymphocyte migration toinflammation and homing to lymphoid tissues by the TA-2 mono-clonal antibody. A likely role for VLA-4 in vivo. J. Immunol. 147:4178.
53 Dustin, M. L. and Shaw, A. S. 1999. Costimulation: building animmunological synapse. Science 283:649.
54 Shier, P., Otulakowski, G., Ngo, K. et al. 1996. Impaired immuneresponses toward alloantigens and tumor cells but normal thymicselection in mice deficient in the beta2 integrin leukocyte function-associated antigen-1. J. Immunol. 157:5375.
55 Smits, H. H., de Jong, E. C., Schuitemaker, J. H. et al. 2002.Intercellular adhesion molecule-1/LFA-1 ligation favors human Th1development. J. Immunol. 168:1710.
56 Kavanaugh, A. F., Lightfoot, E., Lipsky, P. E. and Oppenheimer-Marks, N. 1991. Role of CD11/CD18 in adhesion and trans-endothelial migration of T cells. Analysis utilizing CD18-deficientT cell clones. J. Immunol. 146:4149.
57 Oppenheimer-Marks, N., Davis, L. S., Bogue, D. T., Ramberg, J.and Lipsky, P. E. 1991.Differential utilization of ICAM-1 andVCAM-1during the adhesion and transendothelial migration of humanT lymphocytes. J. Immunol. 147:2913.
58 Van Epps, D. E., Potter, J., Vachula, M., Smith, C. W. and Anderson,D. C. 1989. Suppression of human lymphocyte chemotaxis andtransendothelial migration by anti-LFA-1 antibody. J. Immunol.143:3207.
59 Ding, Z., Xiong, K. and Issekutz, T. B. 2001. Chemokines stimulatehuman T lymphocyte transendothelial migration to utilize VLA-4 inaddition to LFA-1. J. Leukoc. Biol. 69:458.
60 Ding, Z. M., Babensee, J. E., Simon, S. I. et al. 1999. Relativecontribution of LFA-1 and Mac-1 to neutrophil adhesion andmigration. J. Immunol. 163:5029.
398 Role of LFA-1 in experimental colitis
by guest on February 23, 2016http://intim
m.oxfordjournals.org/
Dow
nloaded from
Copyright © 2022 FDOKUMEN




















