miR-34 is maternally inherited in Drosophila melanogaster and Danio rerio
Upload
independentCategory
view
0download
0
Rodlet Cells and the Sensory Systemsin Zebrafish (Danio rerio)
BAHRAM SAYYAF DEZFULI,1* SILVIA CAPUANO,1 EDI SIMONI,1
MAURIZIO PREVIATI,2 AND LUISA GIARI11Department of Biology, University of Ferrara, St. Borsari, Ferrara, Italy
2Department of Morphology and Embryology, University of Ferrara, Ferrara, Italy
ABSTRACTThis preliminary work was designed to study, using routine proce-
dures for light and transmission electron microscopy, the presence ofrodlet cells (RCs) in or near the sensory systems of 12 adult specimens(4.0 6 1.2 cm, LT 6 SD) of zebrafish, Danio rerio Hamilton, 1822. Rodletcells, characterized by a distinctive cell cortex (range, 0.4–1.5 mm in thick-ness) and conspicuous inclusions named ‘‘rodlets,’’ have a round to ovoidnucleus with irregular outline. Mature RCs are 11.5 6 1.2 mm (mean 6SD) long and 7.8 6 1.1 mm (mean 6 SD) wide. These cells are morenumerous near neuromasts enclosed by an epithelial roof and/or ossifiedcanal wall. In contrast, very few RCs were noticed near superficial neuro-masts. Based on the presence of RCs around the two cranial neuromastsof each fish, a variable number from 1 to 15 rodlet cells was found (10.46 3.6, mean 6 SD). The RCs were located 1.5 mm (minimal) to 73.3 mm(maximal) from the neuromast (27.9 6 17.2, mean 6 SD). Moreover, RCswere found in olfactory epithelium and in proximity to some taste buds.Interestingly, RCs were absent in the inner ear, eye, and brain. AnatRec, 290:367–374, 2007. � 2007 Wiley-Liss, Inc.
Key words: rodlet cells; sensory systems; zebrafish; Daniorerio
Neuromasts are known as superficial mechanosensoryorgans occurring in the lateral line of fishes. They func-tion to facilitate schooling, avoid predators, capture prey,and detect vibration in the surrounding environment(Webb, 1989; Uchiyama et al., 1991; Gompel et al., 2001).The inner ear and lateral systems are closely related toeach other developmentally, functionally, and evolution-ary (Williams and Holder, 2000). They form mechano-sensory organs of vertebrates, the inner ear occurringin all vertebrates while neuromasts occurring only inaquatic anamniotes (Song et al., 1995).Two types of neuromasts have been recognized and
named depending on their position: canal and superficial(Coombs et al., 1988; Munz, 1989). In most fishes, thecanal neuromasts are enclosed within a structure suchas open grooves, bony canals (Metcalfe, 1989), or scales.In contrast, the so-called superficial or free neuromastsare not enclosed in a structure and are detectable on thesurface of the epidermis (Song et al., 1995). Structurally,neuromasts are comprised of a ring of supporting cellsencircling a central cluster of sensory hair cells.Olfactory and gustative structures are a portion of the
chemoreception organs, which play an important role in
fish behavior (Hara, 1971; Oliveira Ribeiro et al., 1995;Mandal et al., 2005). These chemical sense organs (olfac-tory epithelium and taste buds) are involved in manylife-sustaining events such as sex recognition, defenseagainst predators, parental behavior, spatial orientation,and food procurement (Døving, 1986; Hara, 1994).During this investigation, a special type of fish cell,
namely rodlet cells (RCs), was noticed adjacent to neuro-masts and within the olfactory epithelium of D. rerio.For over a century, fish histologists and pathologistshave attempted to determine the origin and functions ofthese RCs. Morphologically, they are characterized by a
*Correspondence to: Bahram Sayyaf Dezfuli, Department ofBiology, University of Ferrara, St. Borsari, 46, 44100 Ferrara,Italy. Fax: 0039-0532-291715. E-mail: [email protected]
Grant sponsor: the Italian Ministry of the University and Sci-entific Research and Technology.
Received 2 November 2006; Accepted 18 January 2007
DOI 10.1002/ar.20507Published online 7 March 2007 in Wiley InterScience (www.interscience.wiley.com).
� 2007 WILEY-LISS, INC.
THE ANATOMICAL RECORD 290:367–374 (2007)

distinctive cell cortex and conspicuous inclusions, calledrodlets; hence their namesake. Since their discovery,RCs have been encountered in a wide range of tissues offreshwater and marine teleosts (Leino, 1974; Morrisonand Odense, 1978; Dezfuli et al., 1998, 2000, 2007; Bie-lek, 2005; Mazon et al., 2007). Rodlet cells have beenassociated most often with epithelia, e.g., skin (Iger andAbraham, 1997), gill (Leino, 1974; Dezfuli et al., 2003a),intestine (Paterson and Desser, 1981; Dezfuli et al.,1998, 2003b; Bielek, 2002), kidney tubule (Leino, 1996;Bielek, 2002; Kramer and Potter, 2002), endothelia(Smith et al., 1995; Koponen and Myers, 2000; Maneraet al., 2001), and mesothelia (Dezfuli et al., 2000).Differing points of view on the nature of rodlet cells
have been proposed since their first description byThelohan (1892). After 1895, several authors held theview that RCs are parasites (Laguesse, 1895; Bannister,1966; Mayberry et al., 1979; Viehberger and Bielek,1982; Richards et al., 1994). The main arguments ofthose claiming RCs are parasites include: they vary innumber from fish to fish; they often cannot be found inall individuals of the same species; they occur in differ-ent tissues of the same species; they possess a distinctcapsule; and they have sporozoite-like inclusions. For along time, authors favoring the view that RCs are para-sites believed that RCs belonged to the Apicomplexa(Bannister, 1966; Mayberry et al., 1979; Viehberger andBielek, 1982).The literature supporting the view that RCs are in
fact endogenous fish cells and not parasites is extensive.There are several points that favor the argument for theendogenous nature of these cells. First, the wide distri-bution of RCs within fish tissues and among fish speciesis not a feature of Apicomplexa, a group of parasitesnoted for having strict tissue and host specificity (Floodet al., 1975). Second, in studies involving vast numbersof individual fish, no evidence of disruption or inflamma-tion of surrounding tissue by RCs could be found(Manera and Dezfuli, 2004). Third, RCs have been foundin neonate or very young laboratory-reared fish, inembryos of viviparous teleosts, and in newly hatchedfish (Leino, 1974; Calzada et al., 1998; Kramer andPotter, 2003; Mazon et al., 2007). All of the fish in theseabove-mentioned instances were obtained under patho-gens-free controlled conditions.Catton (1951) and Bullock (1963), among others, sug-
gested that RCs resembled a type of fish granular leuko-cyte. In contrast, other researchers (Barrington, 1957;Bishop and Odense, 1966; Wilson and Westerman, 1967)concurred with Plehn’s (1906a, 1906b) original sugges-tion that RCs are of a glandular nature. In fact, ultra-structural study of these cells in several freshwater andmarine teleosts supports the latter suggestion (Leino,1974, 1996; Flood et al., 1975; Reite, 1998, 2005; Dezfuliet al., 2000, 2003a). Functionally, RCs have been consid-ered regulatory elements related to unique functionssuch as osmoregulation (Fernhead and Fabian, 1971;Mattey et al., 1979) and ion transportation (Morrisonand Odense, 1978). They have been identified as fishblood cells (Weinreb and Bilstad, 1955), secretory cells(Leino, 1974, 1996; Grunberg and Hager, 1978; Matteyet al., 1979), transport units for genetic material(Viehberger and Bielek, 1982), and nonspecific immunecells (Sulimanovic et al., 1996; Iger and Abraham, 1997;Dezfuli et al., 1998, 2000, 2002, 2003a, 2003b, 2007;
Imagawa et al., 1998; Manera et al., 2001; Mazon et al.,2007). Currently, there is only one published study inwhich the authors attributed a sensory function to theRCs (Wilson and Westerman, 1967). However, the re-sults of our previous and present study rule out this lat-ter suggestion.Our present study is the first to document the pres-
ence of RCs near the neuromasts in fish. This site wasunexpected. A hypothesis about RC function will be pre-sented because our current knowledge is far too limitedto provide a concrete explanation.
MATERIALS AND METHODS
Twelve adult specimens of zebrafish (mean LT 6 SD,4.0 6 1.2 cm) from the same parent stock were obtainedfrom a local supplier. The fish were killed by a blow tothe head, subsequently pithed and dissected.Seven whole individuals were designated for paraffin
embedding and five heads were designated for resinembedding. For light microscopy, heads and bodies werefixed separately by immersion in Bouin’s fluid for 8 hr,dehydrated, and embedded in paraffin. Tissue sections(5 mm) were stained with hematoxylin-eosin. For elec-
Fig. 1. Paraffin section of the head of a zebrafish. A neuromast(arrow) enclosed by epithelial roof. Several rodlet cells (arrowheads)occur around the neuromast and in the epithelium of the roof. Scalebar ¼ 20 mm.
368 DEZFULI ET AL.

tron microscopy, heads were fixed at 48C for 2 hr in 2%glutaraldehyde solution buffered with 0.1 M sodiumcacodylate (pH 7.2). The pieces were rinsed for 12 hr in0.1 M sodium cacodylate buffer containing 6% sucrose,then postfixed in 1% osmium tetroxide in the samebuffer for 2 hr before dehydrating in a graded series ofethanol. Finally, pieces were transferred to propyleneoxide and embedded in an Epon-Araldite mixture. Semi-thin sections (1.5 mm) were cut on a Reichert Om U2ultramicrotome with glass knives. Ultrathin sectionswere stained with toluidine blue, contrasted in a 50%alcohol-uranyl acetate solution and lead citrate, andexamined with a Hitachi H-800 electron microscopeoperated at 80 kV.The numbers and dimensions of rodlet and mucous
cells were registered using a computerized image analy-sis software (Lucia G 4.8; Laboratory Imaging, Praha,Czech Republic).
RESULTS
Neuromasts were observed mainly in the cranial regionof each fish (Fig. 1). At this tissue site, the vast majorityof the neuromasts were of the third type (Webb and
Shirey, 2003), namely, those enclosed by epithelial roof(Figs. 1 and 2). Moreover, superficial or free neuromasts(Fig. 3) were rarely found in the head region.The number of RCs ranged from 1 to 15 (10.4 6 3.6,
mean 6 SD) in the epithelium around mainly the thirdtype neuromast (Figs. 1, 2, and 4). The distance of RCsfrom the neuromast ranged from 1.5 to 73.3 mm (27.9 617.2, mean 6 SD). In some instances, the RC was veryclose (1.5 mm) to the peripheral-most support cell of theneuromast (Fig. 6), whereas the most distant RC (Fig. 8)was observed over 70 mm from neuromast. Often, clustersof RCs occurred within close proximity of the neuromast(Figs. 4 and 7). The number of RCs in the epithelium farfrom neuromasts (more than 200 mm) was lower andranged from 0 to 5; the mean density of RCs 6 standarddeviation was 1.1 6 1.0 per 15,000 mm2 tissue area.Almost all the RCs found around the neuromasts weremature, measuring 11.5 6 1.2 mm (mean 6 SD) long and7.8 6 1.1 mm (mean 6 SD) wide. At higher magnification(403 objective), RCs were easily differentiated fromadjacent cell types (e.g., mucous cells) based on thethickened cortex and dark spots (rodlets) within thecytoplasm (Figs. 2 and 4). In contrast, the epidermalmucous cells near the neuromast are largely ovoid,
Fig. 2. Occurrence of high number of rodlet cells (arrowheads)around the neuromast (arrow). Basal nuclei and dark spots within thecytoplasm of the rodlet cells are evident. Scale bar ¼ 20 mm.
Fig. 3. Micrograph of paraffin section shows the presence of arodlet cell (arrow) far from a superficial neuromast (open arrow). Scalebar ¼ 10 mm.
369RODLET CELLS AND SENSORY SYSTEMS

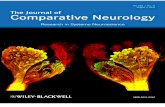
measuring 8.9 6 1.5 mm (mean 6 SD) long and 8.5 61.0 mm (mean 6 SD) wide. Moreover, the cytoplasm iscompletely filled with numerous mucus vesicles up to1.2 mm in diameter (Figs. 4 and 8) and the nucleus ispushed in the deeper zone of the cytoplasm (Fig. 8).Mucus vesicles, as seen by electron microscopy, are elec-tron-lucent or moderately lucent (Fig. 8).Ultrastructurally, the inner part of the RC plasma
membrane made contact with the cell cortex, which
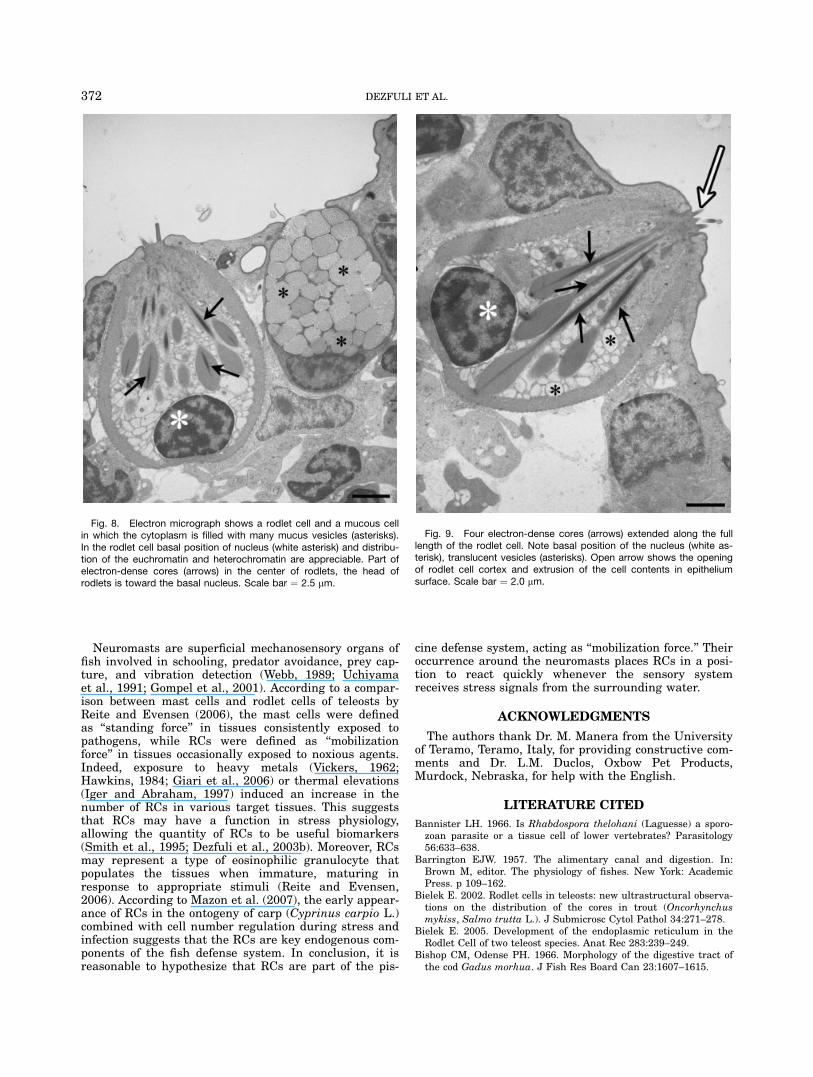
ranged in thickness from 0.4 to 0.7 mm. The cortexappeared to be composed of thin microfilaments. Nucleioccupied the basal part of the cell and were roundto ovoid with an irregular outline (Figs. 8 and 9).Euchromatin was scattered in the nucleoplasm withheterochromatin ringing the periphery (Figs. 8 and 9).Several round, elongate mitochondria were observed inthe apex of the cell. Some RCs contained translucentvesicles within the cytoplasm (Figs. 7 and 8). Club-shapedrodlets were arranged so that the globular part wasoriented toward the basal nucleus with the narrow partoriented toward an opening in the cytoplasmic borderof the RC’s apex (Figs. 7 and 9). Each rodlet possessedan electron-dense core surrounded by a fine granularsubstance. In some instances, the core extended the fulllength of the RC (Figs. 7 and 9). Often, the apical cortexof a mature RC that had reached the epithelial surfacewas interrupted by an opening with RC contents pokingthrough (Fig. 9).The olfactory organ of Danio rerio consists of several
lamellae (Fig. 5a) that insert into a midline raphe, thusforming an oval-shaped rosette. Sensory and nonsensoryregions are located separately on each lamella (Hansenand Zeiske, 1998). Based on light microscopy and theaforementioned RC morphology, lamellae were distin-guishable from the other components of zebra fish olfac-tory epithelium (Fig. 5). Olfactory epithelium of D. reriocontained from 3 to 19 RCs (9.2 6 6.0, mean 6 SD) per22,000 mm2 tissue area. These RCs were of the same sizeas those near the neuromasts. The head epithelium oftwo fish contained 2–3 RCs near the taste buds (notshown). RCs were absent in the eye, inner ear, and brainof D. rerio.
Fig. 4. Semithin section of a neuromast (arrow) and surroundingcells. Note a cluster of rodlet cells (arrowheads) and three mucouscells (open arrows). Scale bar ¼ 10 mm.
Fig. 5. a: Low magnification of an olfactory epithelium of zebrafish. Several rodlet cells (arrowheads)among the olfactory receptor cells (arrow) are evident. Open arrows show olfactory lamellae. Scale bar ¼30 mm. b: High magnification of part of a. Note the rodlet cells (arrows) and the olfactory receptor cells(open arrow). Scale bar ¼ 10 mm.
370 DEZFULI ET AL.

DISCUSSION
Very few available records on the presence of RCs inthe olfactory epithelium of fishes were published (seereview by Manera and Dezfuli, 2004). In this study, RCswere found in the olfactory epithelium of D. rerio in andnear the taste buds of only two specimens. There wereno RCs found in the inner ear, eye, or brain.A literature review emphasizes that there are numer-
ous published reports on the morphology and develop-ment of neuromasts of larval and adult zebrafish usinglight and electron microscopes (for references, see Webband Shirey, 2003). However, these studies fail to provideadequate detailed information on the type of cellssurrounding the neuromasts. This study is the first todocument the occurrence of RCs near the neuromasts ofzebra fish. The close proximity of these cells is the mostinteresting result of this study.Rodlet cells are enigmatic cells exclusive to fish. After
more than a century has passed since their discoveryand documentation in the scientific literature, it is onlyrecently that most investigators agree on the endoge-nous nature of RCs. Currently, the origin of these cellsis the most poorly studied area. Additional researchfrom the field of embryology is fundamental to advanc-ing our understanding of RC. According to Mazon et al.(2007), RCs proliferate in the kidney and then migrateto the gills. While RC function has not been conclusively
shown, a few recent experimental surveys have revealedthe importance of these cells as biomarkers for environ-mental exposure and associated effects (Dezfuli et al.,2003b, 2006; Manera and Dezfuli, 2004; Giari et al.,2006). Rodlet cells appear to contribute to a fish’s defenseagainst pathogenic organisms (Leino, 1996; Dezfuliet al., 1998, 2000, 2003a; Mazon et al., 2007), represent-ing an inflammatory cell closely related to other piscineinflammatory cells, such as mast cells (Reite, 2005; Reiteand Evensen, 2006), epithelioid cells, and mesothelialcells (Dezfuli et al., 2000).The view that RCs possess sensory function was ex-
pressed only by Wilson and Westerman (1967), althoughthe authors did not provide any morphological evidence.Based on our current and previous investigations onRCs, we believe that RCs do not have any structuralfeatures indicative of cells belonging to a fish’s sensorysystems. Rodlet cells exhibit secretory activity (Leino,1974, 1996). Moreover, they do not possess cilia. Thus,there is no connection between these cells and sensoryneurons. More importantly, while RCs can be found inall regions of the fish body, occurrence of RCs aroundthe neuromasts of D. rerio was unexpected. Thus, atpresent, we are only able to postulate the followinghypothesis of their function at the neuromast location.
Fig. 6. Electron micrograph of a rodlet cell (open arrow) close tomost peripheral support cell (asterisk) of a neuromast of D. rerio.Arrow shows the center of neuromast. Scale bar ¼ 2.9 mm.
Fig. 7. A cluster of rodlet cells. Cell cortex (open arrows) aroundeach cell is visible. In cytoplasm rodlets can be seen (arrowheads). Incytoplasm of the center cell translucent vesicles (asterisks) and theopening of the rodlet cell apex (arrow) are evident. Note irregular out-line of two nuclei (white asterisks). Scale bar ¼ 2.5 mm.
371RODLET CELLS AND SENSORY SYSTEMS
Neuromasts are superficial mechanosensory organs offish involved in schooling, predator avoidance, prey cap-ture, and vibration detection (Webb, 1989; Uchiyamaet al., 1991; Gompel et al., 2001). According to a compar-ison between mast cells and rodlet cells of teleosts byReite and Evensen (2006), the mast cells were definedas ‘‘standing force’’ in tissues consistently exposed topathogens, while RCs were defined as ‘‘mobilizationforce’’ in tissues occasionally exposed to noxious agents.Indeed, exposure to heavy metals (Vickers, 1962;Hawkins, 1984; Giari et al., 2006) or thermal elevations(Iger and Abraham, 1997) induced an increase in thenumber of RCs in various target tissues. This suggeststhat RCs may have a function in stress physiology,allowing the quantity of RCs to be useful biomarkers(Smith et al., 1995; Dezfuli et al., 2003b). Moreover, RCsmay represent a type of eosinophilic granulocyte thatpopulates the tissues when immature, maturing inresponse to appropriate stimuli (Reite and Evensen,2006). According to Mazon et al. (2007), the early appear-ance of RCs in the ontogeny of carp (Cyprinus carpio L.)combined with cell number regulation during stress andinfection suggests that the RCs are key endogenous com-ponents of the fish defense system. In conclusion, it isreasonable to hypothesize that RCs are part of the pis-
cine defense system, acting as ‘‘mobilization force.’’ Theiroccurrence around the neuromasts places RCs in a posi-tion to react quickly whenever the sensory systemreceives stress signals from the surrounding water.
ACKNOWLEDGMENTS
The authors thank Dr. M. Manera from the Universityof Teramo, Teramo, Italy, for providing constructive com-ments and Dr. L.M. Duclos, Oxbow Pet Products,Murdock, Nebraska, for help with the English.
LITERATURE CITED
Bannister LH. 1966. Is Rhabdospora thelohani (Laguesse) a sporo-zoan parasite or a tissue cell of lower vertebrates? Parasitology56:633–638.
Barrington EJW. 1957. The alimentary canal and digestion. In:Brown M, editor. The physiology of fishes. New York: AcademicPress. p 109–162.
Bielek E. 2002. Rodlet cells in teleosts: new ultrastructural observa-tions on the distribution of the cores in trout (Oncorhynchusmykiss, Salmo trutta L.). J Submicrosc Cytol Pathol 34:271–278.
Bielek E. 2005. Development of the endoplasmic reticulum in theRodlet Cell of two teleost species. Anat Rec 283:239–249.
Bishop CM, Odense PH. 1966. Morphology of the digestive tract ofthe cod Gadus morhua. J Fish Res Board Can 23:1607–1615.
Fig. 8. Electron micrograph shows a rodlet cell and a mucous cellin which the cytoplasm is filled with many mucus vesicles (asterisks).In the rodlet cell basal position of nucleus (white asterisk) and distribu-tion of the euchromatin and heterochromatin are appreciable. Part ofelectron-dense cores (arrows) in the center of rodlets, the head ofrodlets is toward the basal nucleus. Scale bar ¼ 2.5 mm.
Fig. 9. Four electron-dense cores (arrows) extended along the fulllength of the rodlet cell. Note basal position of the nucleus (white as-terisk), translucent vesicles (asterisks). Open arrow shows the openingof rodlet cell cortex and extrusion of the cell contents in epitheliumsurface. Scale bar ¼ 2.0 mm.
372 DEZFULI ET AL.

Bullock WL. 1963. Intestinal histology of some salmonid fishes withparticular reference to the histopathology of acanthocephalaninfections. J Morphol 112:23–35.
Calzada A, Medina A, Gonzales De Canales ML. 1998. Fine struc-ture of the intestine development in cultured sea bream larvae.J Fish Biol 53:340–365.
Catton WT. 1951. Blood cell formation in certain teleost fishes.Blood 6:39–60.
Coombs S, Janssen J, Webb J. 1988. Diversity of lateral line sys-tems: evolutionary and functional considerations. In: Atema J,Fay RR, Popper AN, Tavolga WN, editors. Sensory biology ofaquatic animals. New York: Springer. p 553–593.
Dezfuli BS, Capuano S, Manera M. 1998. A description of rodletcells from the alimentary canal of Anguilla anguilla andtheir relationship with parasitic helminths. J Fish Biol 53:1084–1095.
Dezfuli BS, Simoni E, Rossi R, Manera M. 2000. Rodlet cells andother inflammatory cells of Phoxinus phoxinus infected withRaphidascaris acus (Nematoda). Dis Aquat Org 43:61–69.
Dezfuli BS, Giari L, Konecny R, Jaeger P, Manera M. 2003a. Immu-nohistochemistry, ultrastructure and pathology of gills of Abramisbrama (L.) from lake Mondsee (Austria) due to Ergasilus sieboldi(Copepoda). Dis Aquat Org 53:257–262.
Dezfuli BS, Giari L, Simoni E, Palazzi D, Manera M. 2003b. Altera-tion of rodlet cells in chub caused by the herbicide Stam1 M-4(Propanil). J Fish Biol 63:232–239.
Dezfuli BS, Simoni E, Giari L, Manera M, 2006. Effects of experi-mental terbuthylazine exposure on the cells of Dicentrarchus lab-rax (L.). Chemosphere 64:1684–1694.
Dezfuli BS, Giari L, Shinn AP. 2007. The role of rodlet cells in theinflammatory response in Diplostomum infected Phoxinus phoxi-nus brains. Fish Shellfish Immunol (in press).
Døving KB. 1986. Functional properties of the fish olfactory system.In: Autrum H, Ottoson D, Perl ER, Schmidt RF, Shimazu H, WillsWD, editors. Progress in sensory physiology. Berlin: Springer.p 39–104.
Fernhead EA, Fabian BL. 1971. The ultrastructure of the gill ofMonodactylus argenteus (an euryhaline teleost fish) with particu-lar reference to morphological changes associated with changes insalinity. South Afr Assoc Marine Biol Res Oceanogr Res InstInvest Rep 26:1–39.
Flood MT, Nigrelli RF, Gennaro JF Jr. 1975. Some aspects of theultrastructure of the ‘‘Stabchendrusenzellen,’’ a peculiar cell asso-ciated with the endothelium of the bulbus arteriosus and withother fish tissue. J Fish Biol 7:129–138.
Giari L, Manera M, Simoni E, Dezfuli BS. 2006. Cellular alterationsin different organs of European sea bass Dicentrarchus labrax(L.) exposed to cadmium. Chemosphere (in press).
Gompel N, Dambly-Chaudiere C, Ghisen A. 2001. Neuronal differ-ences prefigure somatotopy in the zebrafish lateral line. Develop-ment 128:387–393.
Grunberg GW, Hager G. 1978. Zur Ultrastruktur der ‘‘Stabchendru-senzellen’’ (rodlet cells, pear-shaped cells) im bulbus Arteriosusdes Karpfen, Cyprinus carpio L. (Pices: Cyprinidae). Anat Anz134:277–290.
Hansen A, Zeiske E. 1998. The peripheral olfactory organ of thezebrafish, Danio rerio: an ultrastructural study. Chem Senses23:39–48.
Hara TJ. 1971. Chemoreception. In: Hoar WS, Randall DJ, editors.Fish physiology: sensory system and electric organs. New York:Academic Press. p 79–114.
Hara TJ. 1994. Olfaction and gestation in fish: an overview. ActaPhysiol Scand 152:207–217.
Hawkins WE. 1984. Ultrastructure of rodlet cells: response to cad-mium damage in the kidney of the spot Leiostomus xanthurusLacepede. Gulf Res Rep 7:365–372.
Iger Y, Abraham M. 1997. Rodlet cells in the epidermis of fishexposed to stressors. Tissue Cell 29:431–438.
Imagawa T, Kltagawa H, Uehara M. 1998. An association betweenrodlet cells and the vascular endothelial cells in the head kidneyof carp, Cyprinus carpio L.: ultrastructural observation. J FishDis 21:153–157.
Koponen K, Myers MS. 2000. Seasonal changes in intra and inter-organ occurrence of rodlet cells in freshwater bream. J Fish Biol56:250–263.
Kramer CR, Potter H. 2002. Ultrstructural observations on rodletcell development in the head kidney of the southern platyfish,Xiphophorus maculatus. Can J Zool 80:1422–1436.
Kramer CR, Potter H. 2003. Rodlet cells in the posterior intestineof embryos and neonates of two poecilid species. J Fish Biol 62:1211–1216.
Laguesse E. 1895. Les ‘‘Stabchendrusenzellen’’ (M. Plehn) sont dessporozoares parasites. Anat Anz 28:414–416.
Leino RL. 1974. Ultrastructure of immature, developing, and secre-tory rodlet cells in fish. Cell Tissue Res 155:367–381.
Leino RL. 1996. Reaction of rodlet cells to a myxosporean infectionin kidney of the bluegill, Lepomis macrochirus. Can J Zool 74:217–225.
Mandal DK, Roy D, Ghosh L. 2005. Structural organization of theolfactory epithelium of a spotted snakehead fish, Channa puncta-tus. Acta Ich Pisc 35:45–50.
Manera M, Simoni E, Dezfuli BS. 2001. Effect of dexamethasoneadministration on occurrence and ultrastructure of rodlet cells inCarassius auratus L. with particular reference to bulbus arterio-sus. J Fish Biol 59:1239–1248.
Manera M, Dezfuli BS. 2004. Rodlet cells in teleosts: a new insightinto their nature and functions. J Fish Biol 65:597–619.
Mattey DL, Morgan M, Wright DE. 1979. Distribution and develop-ment of rodlet cells in the gills and pseudobranch of the bass,Dicentrarchus labrax (L). J Fish Biol 15:363–370.
Mayberry LF, Marchiondo AA, Ubelaker JE, Kazic D. 1979. Rhab-dospora thelohani Laguesse, 1895 (Apicomplexa): new host andgeographic records with taxonomic consideration. J Protozool 26:168–178.
Mazon AF, Huising MO, Taverne-Thiele AJ, Bastiaans J, Verburg-vanKemenade BML. 2007. The first appearance of rodlet cells in carp(Cyprinus carpio L.) ontogeny and their possible roles during stressand parasite infection. Fish Shellfish Immunol 22:27–37.
Metcalfe W. 1989. Organization and development of the zebrafishposterior lateral line. In: Munz H, editor. The mechanosensorylateral line. New York: Springer. p 147.
Morrison CM, Odense PH. 1978. Distribution and morphology ofthe rodlet cell in fish. J Fish Res Board Can 35:101–116.
Munz H. 1989. Functional organization of the lateral line periphery.In: Coombs S, Gorner P, Munz H, editors. The mechanosensorylateral line. New York: Springer. p 285–298.
Oliveira Ribeiro CA, Fernandes LM, Carvalho CS, Cardoso RI, Tur-catti NM. 1995. Acute effects of mercuric chloride on the olfactoryepithelium of Trichomycterus brasiliensis. Ecotox Environ Safe 31:104–109.
Paterson WB, Desser SS. 1981. Rhabdospora thelohani Laguesse,1906 is not a member of the Apicomplexa. J Parasitol 67:741–744.
Plehn M. 1906a. Uber eigentmuliche Drusenzellen im Gefu und Sys-tem und in aderen Organen bei Fischen. Anat Anz 28:192–203.
PlehnM. 1906b. Drusenzellen oder parasiten? Anat Anz 29:152–157.Reite OB. 1998. Mast cell eosinophilic granule cells of teleostean
fish: a review focusing on staining properties and functionalresponses. Fish Shellfish Immunol 8:489–513.
Reite OB. 2005. The rodlet cells of teleostean fish: their potentialrole in host defence in relation to the role of mast cells/eosino-philic granule cells. Fish Shellfish Immunol 19:253–267.
Reite OB, Evensen Ø. 2006. Inflammatory cells of teleostean fish: areview focusing on mast cells/eosinophilic granule cells and rodletcells. Fish Shellfish Immunol 20:192–208.
Richards DT, Hoole D, Arme C, Lewis JW, Ewens E. 1994. Phagocy-tosis of rodlet cells (Rhabdospora thelohani Laguesse, 1895) bycarp (Cyprinus carpio L.) macrophages and neutrophils. Helmin-thologia 31:29–33.
Smith SA, Caceci T, Marei HE-S, El-Habback HA. 1995. Observa-tions on rodlet cells found in the vascular system and extravascu-lar space of angelfish. J Fish Biol 46:241–254.
Song J, Young Yan H., Popper AN. 1995. Damage and recovery ofhair in fish canal (but not superficial) neuromasts after gentami-cin exposure. Hearing Res 91:63–71.
373RODLET CELLS AND SENSORY SYSTEMS

Sulimanovic D, Curic S, Zeba L, Berc A. 1996. The possible role ofrodlet cells in the immune system of carp (Cyprinus carpio L.).Vet Arhiv 66:103–109.
Thelohan P. 1892. Sur des sporozoaires indetermines parasites despoissons. J Anat Physiol Paris 28:163–171.
Uchiyama M, Iwasaki S, Murakami T. 1991. Surface and subsurfacestructures of neuromasts in tadpoles of the crab eating frog, Ranacarnivora. J Morphol 207:157–163.
Vickers T. 1962. A study of the intestinal epithelium of the goldfishCarassius auratus: its normal structure, the dynamics of cellreplacement, and the changes induced by salts of cobalt and man-ganese. Q J Micros Sci 103:93–110.
Viehberger G, Bielek E. 1982. Rodlet-cells: gland cell or protozoon?Experientia 38:1216–1218.
Webb J. 1989. Neuromast morphology and lateral line trunk canalontogeny in two species of cichlids: an SEM study. J Morphol202:53–60.
Webb JF, Shirey JE. 2003. Postembryonic development of the cra-nial lateral line canals and neuromasts in zebrafish. Dev Dynam228:370–385.
Weinreb EL, Bilstad NM. 1955. Histology of the digestive tract andadjacent structures of the rainbow trout, Salmo gairdneri irideus.Copeia 3:194–204.
Williams JA, Holder N. 2000. Cell turnover of zebrafish larvae.Hearing Res 143:171–181.
Wilson JAF, Westerman RA. 1967. The fine structure of the olfac-tory mucosa and nerve in the teleost Carassius auratus L. Z Zell-forsch Mikrosk Anat 83:196–206.
374 DEZFULI ET AL.
Copyright © 2022 FDOKUMEN