
Notch signaling can regulate endoderm formation in zebrafish
7
RESEARCH ARTICLE Notch Signaling Can Regulate Endoderm Formation in Zebrafish Yutaka Kikuchi, 1,2 * Heather Verkade, 2 Jeremy F. Reiter, 2 Cheol-Hee Kim, 3 Ajay B. Chitnis, 3 Atsushi Kuroiwa, 1 and Didier Y.R. Stainier 2 Early in vertebrate development, the processes of gastrulation lead to the formation of the three germ layers: ectoderm, mesoderm, and endoderm. The mechanisms leading to the segregation of the endoderm and mesoderm are not well understood. In mid-blastula stage zebrafish embryos, single marginal cells can give rise to both endoderm and mesoderm (reviewed by Warga and Stainier [2002] The guts of endoderm formation. In: Solnica-Krezel L, editor. Pattern formation in zebrafish. Berlin: Springer-Verlag. p 28 – 47). By the late blastula stage, however, single marginal cells generally give rise to either endoderm or mesoderm. To investigate this segregation of the blastoderm into cells with either endodermal or mesodermal fates, we analyzed the role of Notch signaling in this process. We show that deltaC, deltaD, and notch1 are expressed in the marginal domain of blastula stage embryos and that this expression is dependent on Nodal signaling. Activation of Notch signaling from an early stage leads to a reduction of endodermal cells, as assessed by sox17 and foxA2 expression. We further find that this reduction in endoderm formation by the activation of Notch signaling is preceded by a reduction in the expression of bonnie and clyde (bon) and faust/gata5, two genes necessary for endoderm formation (Reiter et al. [1999] Genes Dev 13:2983–2995; Reiter et al. [2001] Development 128:125–135; Kikuchi et al. [2001] Genes Dev 14:1279 –1289). However, activation of Notch signaling in bon mutant embryos leads to a further reduction in endodermal cells, also arguing for a bon-independent role for Notch signaling in endoderm formation. Altogether, these results suggest that Notch signaling plays a role in the formation of the endoderm, possibly in its segregation from the mesoderm. Developmental Dynamics 229: 756 –762, 2004. © 2004 Wiley-Liss, Inc. Key words: Notch; endoderm; sox17; foxA2; zebrafish Received 29 September 2003; Accepted 6 October 2003 INTRODUCTION During vertebrate development, the endodermal germ layer gives rise to the epithelial lining of the digestive tract as well as its associated organs. In zebrafish, the endoderm origi- nates from the most marginal blas- tomeres of blastula stage embryos (Warga and Nu ¨ sslein-Volhard, 1999). Fate mapping analyses show that at the mid-blastula stage single cells can give rise to both endoderm and mesoderm. By the late blastula stage, single cells generally give rise to either endoderm or mesoderm, indicating that these lineages have segregated. However, the mecha- nisms leading to the segregation of the endoderm and mesoderm are unknown. Endodermal progenitors are found no more than four cells away from the blastoderm margin inter- mingled with cells that give rise to some mesodermal derivatives such as the somites and the heart, indi- cating that endodermal and meso- 1 Division of Biological Science, Graduate School of Science, Nagoya University, Furo-cho, Chikusa-ku, Nagoya, Japan 2 Department of Biochemistry and Biophysics, Programs in Developmental Biology, Genetics and Human Genetics, University of California at San Francisco, San Francisco, California 3 Laboratory of Molecular Genetics, NICHD/NIH, Bethesda, Maryland Grant sponsors: NIH; AHA; MEXT (The Ministry of Education, Culture, Sports, Science, and Technology); Grant number: KAKENHI 13138204. Dr. Kim’s present address is Department of Biology, Chungnam National University, Daejeon 305-764, Korea. *Correspondence to: Yutaka Kikuchi, Division of Biological Science, Graduate School of Science, Nagoya University, Furo-cho, Chikusa-ku, Nagoya, 464-8602 Japan. E-mail: [email protected] DOI 10.1002/dvdy.10483 DEVELOPMENTAL DYNAMICS 229:756 –762, 2004 © 2004 Wiley-Liss, Inc.
Transcript of Notch signaling can regulate endoderm formation in zebrafish

RESEARCH ARTICLE
Notch Signaling Can Regulate EndodermFormation in ZebrafishYutaka Kikuchi,1,2* Heather Verkade,2 Jeremy F. Reiter,2 Cheol-Hee Kim,3 Ajay B. Chitnis,3 Atsushi Kuroiwa,1
and Didier Y.R. Stainier2
Early in vertebrate development, the processes of gastrulation lead to the formation of the three germ layers:ectoderm, mesoderm, and endoderm. The mechanisms leading to the segregation of the endoderm and mesodermare not well understood. In mid-blastula stage zebrafish embryos, single marginal cells can give rise to bothendoderm and mesoderm (reviewed by Warga and Stainier [2002] The guts of endoderm formation. In: Solnica-KrezelL, editor. Pattern formation in zebrafish. Berlin: Springer-Verlag. p 28–47). By the late blastula stage, however, singlemarginal cells generally give rise to either endoderm or mesoderm. To investigate this segregation of the blastoderminto cells with either endodermal or mesodermal fates, we analyzed the role of Notch signaling in this process. Weshow that deltaC, deltaD, and notch1 are expressed in the marginal domain of blastula stage embryos and that thisexpression is dependent on Nodal signaling. Activation of Notch signaling from an early stage leads to a reduction ofendodermal cells, as assessed by sox17 and foxA2 expression. We further find that this reduction in endodermformation by the activation of Notch signaling is preceded by a reduction in the expression of bonnie and clyde (bon)and faust/gata5, two genes necessary for endoderm formation (Reiter et al. [1999] Genes Dev 13:2983–2995; Reiteret al. [2001] Development 128:125–135; Kikuchi et al. [2001] Genes Dev 14:1279–1289). However, activation of Notchsignaling in bon mutant embryos leads to a further reduction in endodermal cells, also arguing for a bon-independentrole for Notch signaling in endoderm formation. Altogether, these results suggest that Notch signaling plays a role inthe formation of the endoderm, possibly in its segregation from the mesoderm. Developmental Dynamics 229:756–762, 2004. © 2004 Wiley-Liss, Inc.
Key words: Notch; endoderm; sox17; foxA2; zebrafish
Received 29 September 2003; Accepted 6 October 2003
INTRODUCTIONDuring vertebrate development, theendodermal germ layer gives rise tothe epithelial lining of the digestivetract as well as its associated organs.In zebrafish, the endoderm origi-nates from the most marginal blas-tomeres of blastula stage embryos(Warga and Nusslein-Volhard, 1999).
Fate mapping analyses show that atthe mid-blastula stage single cellscan give rise to both endoderm andmesoderm. By the late blastulastage, single cells generally give riseto either endoderm or mesoderm,indicating that these lineages havesegregated. However, the mecha-nisms leading to the segregation of
the endoderm and mesoderm areunknown.
Endodermal progenitors arefound no more than four cells awayfrom the blastoderm margin inter-mingled with cells that give rise tosome mesodermal derivatives suchas the somites and the heart, indi-cating that endodermal and meso-
1Division of Biological Science, Graduate School of Science, Nagoya University, Furo-cho, Chikusa-ku, Nagoya, Japan2Department of Biochemistry and Biophysics, Programs in Developmental Biology, Genetics and Human Genetics, University of Californiaat San Francisco, San Francisco, California3Laboratory of Molecular Genetics, NICHD/NIH, Bethesda, MarylandGrant sponsors: NIH; AHA; MEXT (The Ministry of Education, Culture, Sports, Science, and Technology); Grant number: KAKENHI13138204.Dr. Kim’s present address is Department of Biology, Chungnam National University, Daejeon 305-764, Korea.*Correspondence to: Yutaka Kikuchi, Division of Biological Science, Graduate School of Science, Nagoya University, Furo-cho,Chikusa-ku, Nagoya, 464-8602 Japan. E-mail: [email protected]
DOI 10.1002/dvdy.10483
DEVELOPMENTAL DYNAMICS 229:756–762, 2004
© 2004 Wiley-Liss, Inc.

dermal progenitors are intermingledto some degree along the margin ofthe blastoderm.
Genetic analyses in zebrafish haveidentified three genes (bonnie andclyde [bon], faust [fau], andcasanova [cas]) that play essentialroles in endoderm formation. bonand fau mutants contain �10% and�60% of the normal number ofendodermal cells, respectively (Kiku-chi et al., 2000; Reiter et al., 2001),while cas mutants lack all endoder-mal cells (Alexander et al., 1999).bon, fau, and cas encode a Mix-type homeodomain transcriptionfactor (Kikuchi et al., 2000), the zincfinger transcription factor Gata5(Reiter et al., 1999), and a high-mo-bility group (HMG) domain transcrip-tion factor (Dickmeis et al., 2001;Kikuchi et al., 2001; Sakaguchi et al.,2001), respectively. More recently,another Mix-type homeodomaintranscription factor, Mezzo, was iden-tified to be involved in endoderm for-mation, in a manner partially redun-dant with Bon (Poulain and Lepage,2002). Gene expression analyses andmisexpression experiments in wild-type and mutant embryos have re-vealed that these four transcriptionfactors function downstream of Nodalsignaling and upstream of sox17, anHMG transcription factor gene (Alex-ander and Stainier, 1999) and foxA2,a winged helix/forkhead transcriptionfactor gene (also known as axial;Strahle et al., 1993), both of which areexpressed in endodermal cells (re-viewed in Stainier, 2002).
Numerous studies have revealedthat Notch signaling plays importantroles in cell fate specification duringdevelopment (reviewed by Artava-nis-Tsakonas et al., 1999; Mumm andKopan, 2000). A mechanism of lat-eral inhibition involving Notch signal-ing was proposed to explain the ac-quisition of different cell fates byneighboring cells. In zebrafish, lateralinhibition has been demonstrated toregulate primary neuron and oligo-dendrocyte development (Haddonet al., 1998; Appel and Eisen, 1998;Park and Appel, 2003), as well as thedecision between hypochord andnotochord cell fate (Latimer et al.,2002). bon, mezzo, and fau/gata5are expressed in all cells around thecircumference of the margin, while
cas, sox17, and foxA2 are expressedin the scattered endodermal cellsaround the marginal domain of thelate-blastula and early gastrulastage embryos. The salt-and-pepperpattern of cas, sox17, and foxA2 ex-pression suggests that endodermalcells may be selected by lateral in-hibition.
In this study, we test whetherNotch signaling regulates endodermformation in the marginal domain.Activation of the Notch signalingpathway by overexpression ofNotch intracellular domain (NICD) inzebrafish embryos leads to a reduc-tion in the number of cells expressingsox17 and foxA2. We further showthat expression of bon, which func-tions upstream of sox17 and foxA2, issignificantly reduced by the activa-tion of Notch signaling. Our datasuggest that Notch signaling may in-fluence the decision to becomeendoderm.
RESULTS AND DISCUSSION
Zebrafish endodermal cells can beidentified during gastrulation by theirdistinctive morphology and expres-sion of sox17 and foxA2. During earlygastrulation, these cells are distrib-uted around the margin in a salt-and-pepper pattern (Fig. 1A-D). Theformation of the zebrafish endodermdepends upon bon, fau/gata5, andmezzo, three transcription factorgenes expressed throughout themarginal domain. That only a subsetof marginal cells expressing thesethree transcription factor genes be-come endodermal suggests thatsome other mechanism is further re-stricting the endoderm-forming ca-pacity of marginal cells. The salt-and-pepper distribution of endodermalprogenitors further suggests that a lat-eral specification mechanism mayparticipate in this restriction, prompt-ing us to investigate the role of Notchsignaling in this process.
deltaC (Smithers et al., 2000),deltaD (Haddon et al., 1998), andnotch1 (Bierkamp and Campos-Or-tega, 1993) are also expressed in thismarginal domain during both lateblastula and early gastrula stages(Fig. 1E,G,I). Because the marginalexpression of bon, fau/gata5, andcas is regulated by Nodal signaling
(reviewed by Stainier, 2002), we in-vestigated whether Nodal signalingalso regulates deltaC, deltaD, andnotch1 expression. We found thatembryos lacking both the maternaland zygotic complement of theNodal cofactor Oep (MZoep mu-tants; Gritsman et al., 1999) displayno deltaC, deltaD, and notch1 ex-pression, indicating that the expres-sion of these genes is also regulatedby Nodal signaling (Fig. 1F,H,J). Theexpression patterns of deltaC,deltaD, and notch1 during late blas-tula and early gastrula stages raisethe possibility that Notch signaling isinvolved in the formation of theendoderm and mesoderm.
Expression of the Notch intracellu-lar domain activates the Notch sig-naling pathway in a ligand-indepen-dent manner (Fortini et al., 1993;Lieber et al., 1993; Struhl et al., 1993).To investigate the role of Notch sig-naling in endoderm formation, weinjected mRNA encoding the intra-cellular domain of zebrafish Notch1(NICD; Takke et al., 1999). Injectionof 100 pg of NICD mRNA led to asubstantial decrease in the numberof sox17- and foxA2-expressingendodermal cells (Fig. 2B,E; Table 1).We also found a reduced number ofcas-expressing endodermal cellsduring early gastrulation stages andreduced endodermal tissues atpharyngula stages after NICD injec-tions (data not shown). Unfortu-nately, as specific pan-mesodermalmarkers remain to be identified inzebrafish, we have been unable toanalyze directly the role of Notchsignaling in mesoderm formation. In-stead, we generated embryos con-sisting primarily of endodermal cellsby injecting 500 pg of a constitutivelyactive form of the type-I TGF� recep-tor Taram-a, designated Taram-a*(Peyrieras et al., 1998). We found that,while 18% (39 of 214) of the embryosinjected with Taram-a* alone ex-pressed the somitic marker myoD,coinjection with 100 pg of NICD in-creased that proportion to 28% (66of 235), suggesting that activation ofthe Notch signaling pathway canconvert endodermal cells to meso-dermal cells. Altogether, these re-sults indicate that activation of theNotch signaling pathway duringearly embryonic stages can sup-
NOTCH SIGNALING AND ENDODERM FORMATION 757
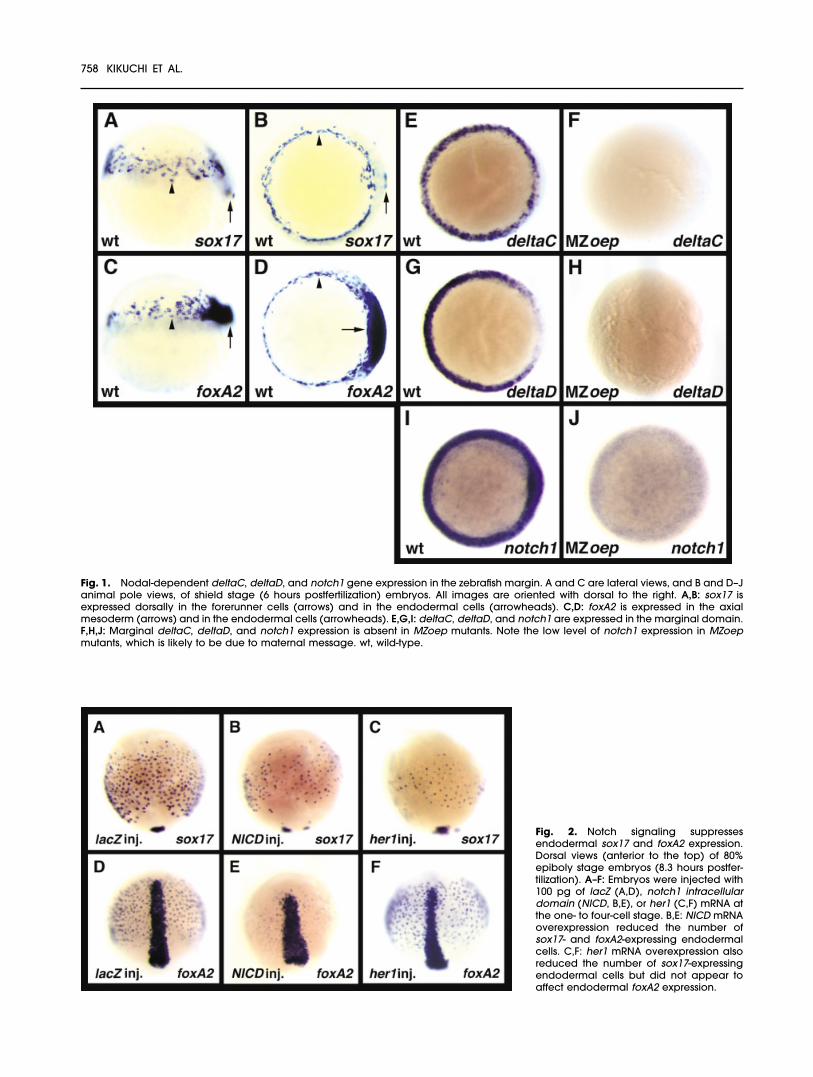
Fig. 1. Nodal-dependent deltaC, deltaD, and notch1 gene expression in the zebrafish margin. A and C are lateral views, and B and D–Janimal pole views, of shield stage (6 hours postfertilization) embryos. All images are oriented with dorsal to the right. A,B: sox17 isexpressed dorsally in the forerunner cells (arrows) and in the endodermal cells (arrowheads). C,D: foxA2 is expressed in the axialmesoderm (arrows) and in the endodermal cells (arrowheads). E,G,I: deltaC, deltaD, and notch1 are expressed in the marginal domain.F,H,J: Marginal deltaC, deltaD, and notch1 expression is absent in MZoep mutants. Note the low level of notch1 expression in MZoepmutants, which is likely to be due to maternal message. wt, wild-type.
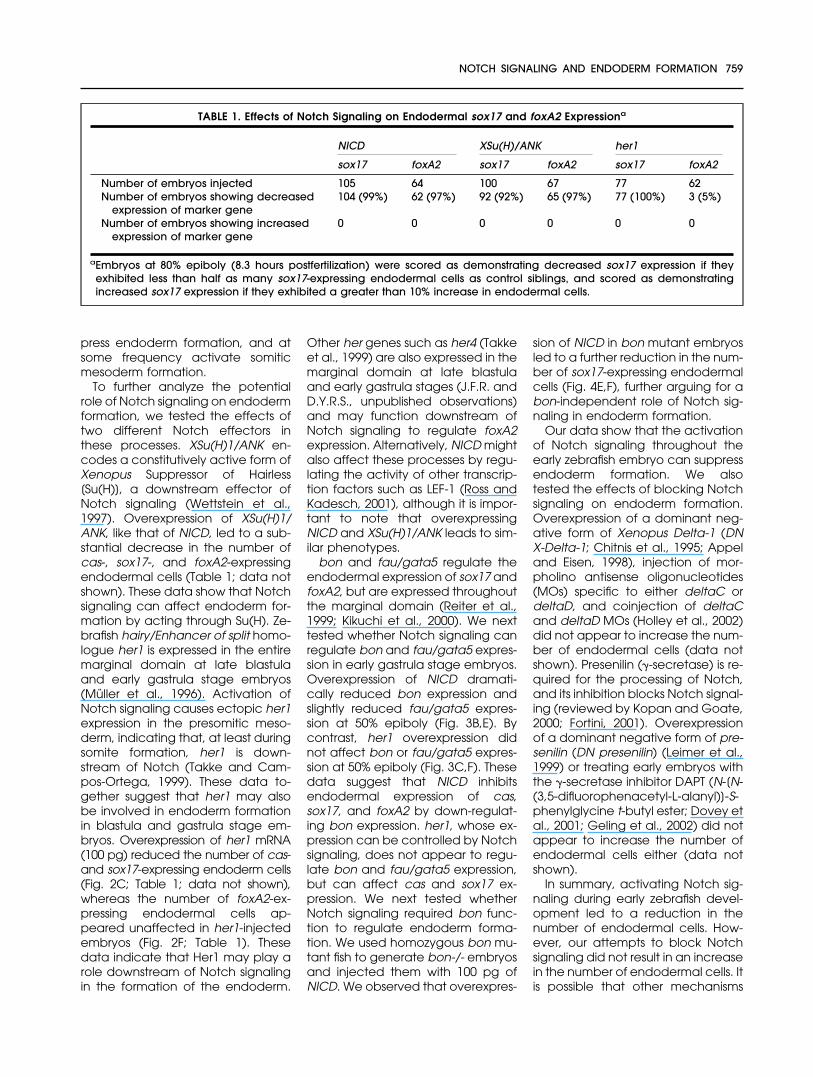
Fig. 2. Notch signaling suppressesendodermal sox17 and foxA2 expression.Dorsal views (anterior to the top) of 80%epiboly stage embryos (8.3 hours postfer-tilization). A–F: Embryos were injected with100 pg of lacZ (A,D), notch1 intracellulardomain (NICD, B,E), or her1 (C,F) mRNA atthe one- to four-cell stage. B,E: NICD mRNAoverexpression reduced the number ofsox17- and foxA2-expressing endodermalcells. C,F: her1 mRNA overexpression alsoreduced the number of sox17-expressingendodermal cells but did not appear toaffect endodermal foxA2 expression.
758 KIKUCHI ET AL.
press endoderm formation, and atsome frequency activate somiticmesoderm formation.
To further analyze the potentialrole of Notch signaling on endodermformation, we tested the effects oftwo different Notch effectors inthese processes. XSu(H)1/ANK en-codes a constitutively active form ofXenopus Suppressor of Hairless[Su(H)], a downstream effector ofNotch signaling (Wettstein et al.,1997). Overexpression of XSu(H)1/ANK, like that of NICD, led to a sub-stantial decrease in the number ofcas-, sox17-, and foxA2-expressingendodermal cells (Table 1; data notshown). These data show that Notchsignaling can affect endoderm for-mation by acting through Su(H). Ze-brafish hairy/Enhancer of split homo-logue her1 is expressed in the entiremarginal domain at late blastulaand early gastrula stage embryos(Muller et al., 1996). Activation ofNotch signaling causes ectopic her1expression in the presomitic meso-derm, indicating that, at least duringsomite formation, her1 is down-stream of Notch (Takke and Cam-pos-Ortega, 1999). These data to-gether suggest that her1 may alsobe involved in endoderm formationin blastula and gastrula stage em-bryos. Overexpression of her1 mRNA(100 pg) reduced the number of cas-and sox17-expressing endoderm cells(Fig. 2C; Table 1; data not shown),whereas the number of foxA2-ex-pressing endodermal cells ap-peared unaffected in her1-injectedembryos (Fig. 2F; Table 1). Thesedata indicate that Her1 may play arole downstream of Notch signalingin the formation of the endoderm.
Other her genes such as her4 (Takkeet al., 1999) are also expressed in themarginal domain at late blastulaand early gastrula stages (J.F.R. andD.Y.R.S., unpublished observations)and may function downstream ofNotch signaling to regulate foxA2expression. Alternatively, NICD mightalso affect these processes by regu-lating the activity of other transcrip-tion factors such as LEF-1 (Ross andKadesch, 2001), although it is impor-tant to note that overexpressingNICD and XSu(H)1/ANK leads to sim-ilar phenotypes.
bon and fau/gata5 regulate theendodermal expression of sox17 andfoxA2, but are expressed throughoutthe marginal domain (Reiter et al.,1999; Kikuchi et al., 2000). We nexttested whether Notch signaling canregulate bon and fau/gata5 expres-sion in early gastrula stage embryos.Overexpression of NICD dramati-cally reduced bon expression andslightly reduced fau/gata5 expres-sion at 50% epiboly (Fig. 3B,E). Bycontrast, her1 overexpression didnot affect bon or fau/gata5 expres-sion at 50% epiboly (Fig. 3C,F). Thesedata suggest that NICD inhibitsendodermal expression of cas,sox17, and foxA2 by down-regulat-ing bon expression. her1, whose ex-pression can be controlled by Notchsignaling, does not appear to regu-late bon and fau/gata5 expression,but can affect cas and sox17 ex-pression. We next tested whetherNotch signaling required bon func-tion to regulate endoderm forma-tion. We used homozygous bon mu-tant fish to generate bon-/- embryosand injected them with 100 pg ofNICD. We observed that overexpres-
sion of NICD in bon mutant embryosled to a further reduction in the num-ber of sox17-expressing endodermalcells (Fig. 4E,F), further arguing for abon-independent role of Notch sig-naling in endoderm formation.
Our data show that the activationof Notch signaling throughout theearly zebrafish embryo can suppressendoderm formation. We alsotested the effects of blocking Notchsignaling on endoderm formation.Overexpression of a dominant neg-ative form of Xenopus Delta-1 (DNX-Delta-1; Chitnis et al., 1995; Appeland Eisen, 1998), injection of mor-pholino antisense oligonucleotides(MOs) specific to either deltaC ordeltaD, and coinjection of deltaCand deltaD MOs (Holley et al., 2002)did not appear to increase the num-ber of endodermal cells (data notshown). Presenilin (�-secretase) is re-quired for the processing of Notch,and its inhibition blocks Notch signal-ing (reviewed by Kopan and Goate,2000; Fortini, 2001). Overexpressionof a dominant negative form of pre-senilin (DN presenilin) (Leimer et al.,1999) or treating early embryos withthe �-secretase inhibitor DAPT (N-[N-(3,5-difluorophenacetyl-L-alanyl)]-S-phenylglycine t-butyl ester; Dovey etal., 2001; Geling et al., 2002) did notappear to increase the number ofendodermal cells either (data notshown).
In summary, activating Notch sig-naling during early zebrafish devel-opment led to a reduction in thenumber of endodermal cells. How-ever, our attempts to block Notchsignaling did not result in an increasein the number of endodermal cells. Itis possible that other mechanisms
TABLE 1. Effects of Notch Signaling on Endodermal sox17 and foxA2 Expressiona
NICD XSu(H)/ANK her1
sox17 foxA2 sox17 foxA2 sox17 foxA2
Number of embryos injected 105 64 100 67 77 62Number of embryos showing decreased
expression of marker gene104 (99%) 62 (97%) 92 (92%) 65 (97%) 77 (100%) 3 (5%)
Number of embryos showing increasedexpression of marker gene
0 0 0 0 0 0
aEmbryos at 80% epiboly (8.3 hours postfertilization) were scored as demonstrating decreased sox17 expression if theyexhibited less than half as many sox17-expressing endodermal cells as control siblings, and scored as demonstratingincreased sox17 expression if they exhibited a greater than 10% increase in endodermal cells.
NOTCH SIGNALING AND ENDODERM FORMATION 759
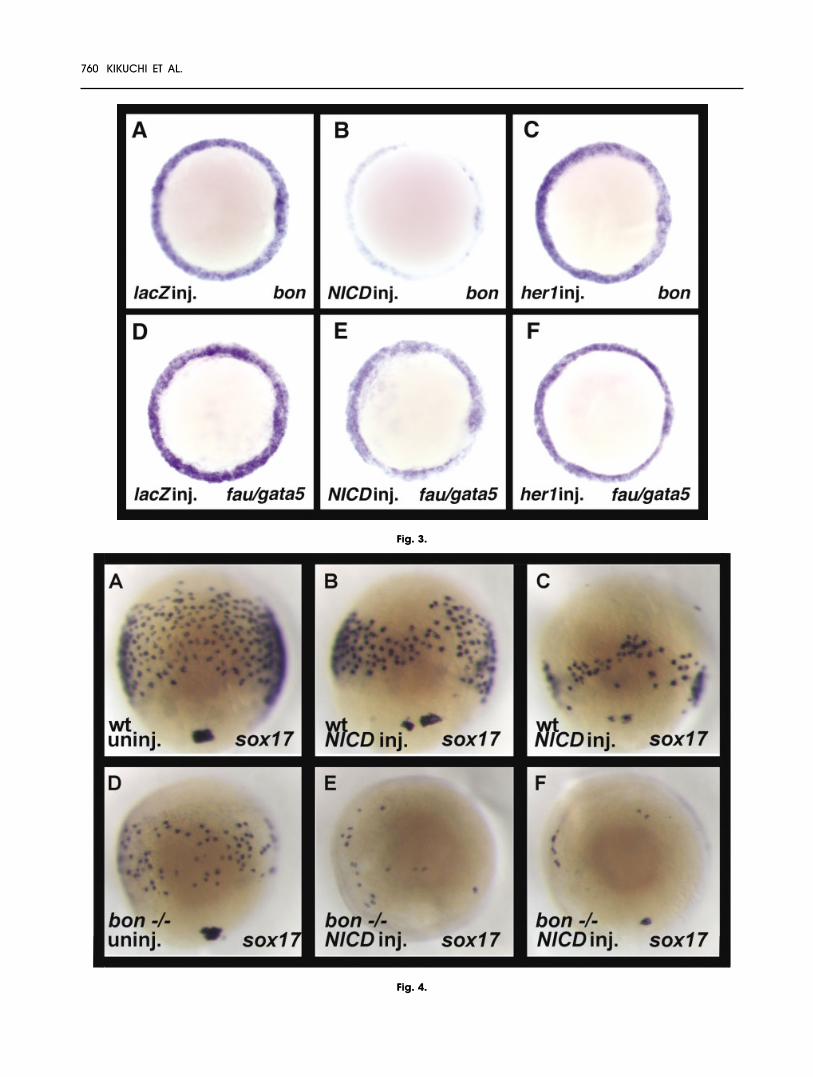
Fig. 3.
Fig. 4.
760 KIKUCHI ET AL.

are at play to regulate the numberof endodermal cells and that block-ing Notch signaling is not sufficient toovercome them. Whether Notch sig-naling is really implicated in the seg-regation of endoderm and meso-derm through a lateral inhibitionprocess remains unclear and will re-quire the development of tech-niques that allow the activation andblocking of Notch signaling in singlecells at the time when the decisionto become endoderm or mesodermis being made, as well as the abilityto monitor the fate of these singlecells and their progeny. Alterna-tively, it is possible that Notch signal-ing regulates the temporal patternof bon expression and, thereby, thenumber of endodermal cells gener-ated. bon expression initiates at themid-blastula stage and becomesundetectable by 60% epiboly. Ourdata show that premature activa-tion of Notch signaling leads to apremature reduction of bon expres-sion and a subsequent decrease inthe number of endodermal cells;thus endogenous Notch signalingmight turn off bon expression andthereby regulate endoderm forma-tion. In addition and as shown by thefurther reduction of endodermalcells in bon mutant embryos in-
jected with NICD, Notch signalingmay also regulate endoderm forma-tion in a bon-independent manner.
EXPERIMENTAL PROCEDURES
Zebrafish Strains
Adult zebrafish were maintainedand staged as described (Wester-field, 1995). Adult MZoepm134 fishwere a generous gift of Drs. Will Tal-bot (Stanford University) and MarnieHalpern (Carnegie Institution ofWashington). The bonm425 (Kikuchiet al., 2000) mutant allele was usedand bonm425 homozygous fish werecreated by injecting wild-type bonmRNA into homozygous mutant em-bryos at the one- to four-cell stageto rescue embryonic defects.
Whole-Mount In SituHybridization
Whole-mount in situ hybridizationwas performed as described (Kiku-chi et al., 1997; Alexander et al.,1998), and riboprobes prepared ac-cording to published instructions.
mRNA and MorpholinoInjections
Capped mRNA was synthesized byusing the SP6 mMessage mMachinesystem (Ambion) from the followingpreviously described templates:NICD (Takke et al., 1999), XSu(H)1/ANK (Wettstein et al., 1997), her1(Takke and Campos-Ortega, 1999),Taram-a* (Peyrieras et al., 1998), DNX-Delta-1 (Chitnis et al., 1995; Appeland Eisen, 1998), DN presenilin (Lei-mer et al., 1999). For overexpressionexperiments, mRNAs of NICD (100pg), XSu(H)1/ANK (100 pg), her1 (100pg), Taram-a* (500 pg), DN X-Delta-1(300 pg), DN presenilin (400 pg), andlacZ (100 pg) were microinjected atthe one- to four-cell stage. Injectionsof morpholino antisense oligonucle-otides for deltaC and deltaD werecarried out as described (Holley etal., 2002).
DAPT Treatments
DAPT treatments were performed asdescribed previously (Geling et al.,2002): wild-type embryos were de-chorionated at the one-cell stage
and treated with 100 �M DAPT fromthe one-cell stage until 80% epiboly(8.3 hours postfertilization).
ACKNOWLEDGMENTSWe thank Alex Navarro and StevenWaldron for invaluable technical as-sistance; Le Trinh for bon homozy-gous fish; Jose Campos-Ortega, Sha-ron Amacher, and Julian Lewis forDNA templates; and Jonathan Alex-ander, Yee-Ming Chan, Xin Sun,Mike Verzi, and Pia Aanstad for dis-cussions and comments on themanuscript. Y.K. was funded by Post-doctoral Fellowships from the Amer-ican Heart Association and the Hu-man Frontier Science Program(HFSP). H.V. was funded by a Postdoc-toral Fellowship from the HFSP. J.F.R.was a member of the Medical Scien-tist Training Program of the NIH and anAchievement Rewards for CollegeScientists scholarship recipient. C.-H.Kim was supported by a VascularSystem Research Center grant fromKorea Science and EngineeringFoundation. This work was supportedin part by grants to D.Y.R.S. from theAmerican Heart Association, thePackard Foundation and the Lifeand Health Insurance Medical Re-search Fund, and by a Grant-in-Aidfor Scientific Research to Y.K. fromThe Ministry of Education, Culture,Sports, Science, and Technology(MEXT).
REFERENCES
Alexander J, Stainier DY. 1999. A molec-ular pathway leading to endoderm for-mation in zebrafish. Curr Biol 9:1147–1157.
Alexander J, Stainier DY, Yelon D. 1998.Screening mosaic F1 females for muta-tions affecting zebrafish heart induc-tion and patterning. Dev Genet 22:288–299.
Alexander J, Rothenberg M, Henry GL,Stainier DY. 1999. casanova plays anearly and essential role in endodermformation in zebrafish. Dev Biol 215:343–357.
Appel B, Eisen JS. 1998. Regulation ofneuronal specification in the zebrafishspinal cord by Delta function. Develop-ment 125:371–380.
Artavanis-Tsakonas S, Rand MD, Lake RJ.1999. Notch signaling: cell fate controland signal integration in development.Science 284:770–776.
Bierkamp C, Campos-Ortega JA. 1993. Azebrafish homologue of the Drosophilaneurogenic gene Notch and its pat-
Fig. 3. Notch signaling inhibits bon expres-sion. A–F: Embryos were injected with 100pg of lacZ (A,D), NICD (B,E), or her1 (C,F)mRNA at the one- to four-cell stage, andexamined for bon or fau/gata5 expressionat 50% epiboly (5.3 hours postfertilization).A–F: Animal pole views, dorsal to the right.B,E: NICD overexpression dramatically re-duced bon expression and slightly re-duced fau/gata5 expression. C,F: UnlikeNICD, her1 overexpression did not affectbon or fau/gata5 expression.Fig. 4. Notch signaling also functions in-dependently of bon function in endodermformation. Dorsal views (anterior to the top)of 80% epiboly stage embryos (8.3 hourspostfertilization). Embryos are wild-type (wt,A,B,C) or from an incross of bon-/- fish(D,E,F). Embryos were left uninjected (A,D),or injected with 100 pg of NICD mRNA at theone- to four-cell stage (B,C,E,F). A,D: Theloss of bon function reduces the numberof sox17 expressing endodermal cells.B,C: NICD overexpression also reduces thenumber of sox17 expressing endodermalcells. E,F: NICD overexpression in bon mu-tants further reduces the number of sox17expressing endodermal cells.
NOTCH SIGNALING AND ENDODERM FORMATION 761

tern of transcription during early em-bryogenesis. Mech Dev 43:87–100.
Chitnis A, Henrique D, Lewis J, Ish-Horow-icz D, Kintner C. 1995. Primary neuro-genesis in Xenopus embryos regulatedby a homologue of the Drosophila neu-rogenic gene Delta. Nature 375:761–766.
Dickmeis T, Mourrain P, Saint-Etienne L,Fischer N, Aanstad P, Clark M, Strahle U,Rosa F. 2001. A crucial component ofthe endoderm formation pathway,CASANOVA, is encoded by a novelsox-related gene. Genes Dev 15:1487–1492.
Dovey HF, John V, Anderson JP, Chen LZ,de Saint Andrieu P, Fang LY, FreedmanSB, Folmer B, Goldbach E, HolsztynskaEJ, Hu KL, Johnson-Wood KL, KennedySL, Kholodenko D, Knops JE, Latimer LH,Lee M, Liao Z, Lieberburg IM, Motter RN,Mutter LC, Nietz J, Quinn KP, Sacchi KL,Seubert PA, Shopp GM, Thorsett ED,Tung JS, Wu J, Yang S, Yin CT, SchenkDB, May PC, Altstiel LD, Bender MH,Boggs LN, Britton TC, Clemens JC, CzilliDL, Dieckman-McGinty DK, Droste JJ,Fuson KS, Gitter BD, Hyslop PA, John-stone EM, Li WY, Little SP, Mabry TE,Miller FD, Audia JE. 2001. Functional�-secretase inhibitors reduce �-amy-loid peptide levels in brain. J Neuro-chem 76:173–181.
Fortini ME. 2001. Notch and presenilin: aproteolytic mechanism emerges. CurrOpin Cell Biol 13:627–634.
Fortini ME, Rebay I, Caron LA, Artavanis-Tsakonas S. 1993. An activated Notchreceptor blocks cell-fate commitmentin the developing Drosophila eye. Na-ture 365:555–557.
Geling A, Steiner H, Willem M, Bally-Cuif L,Haass C. 2002. A �-secretase inhibitorblocks Notch signaling in vivo andcauses a severe neurogenic pheno-type in zebrafish. EMBO Rep 3:688–694.
Gritsman K, Zhang J, Cheng S, HeckscherE, Talbot WS, Schier AF. 1999. The EGF-CFC protein one-eyed pinhead is es-sential for nodal signaling. Cell 97:121–132.
Haddon C, Smithers L, Schneider-Mau-noury S, Coche T, Henrique D, Lewis J.1998. Multiple delta genes and lateralinhibition in zebrafish primary neuro-genesis. Development 125:359–370.
Holley SA, Julich D, Rauch GJ, Geisler R,Nusslein-Volhard C. 2002. her1 and thenotch pathway function within the os-cillator mechanism that regulates ze-brafish somitogenesis. Development129:1175–1183.
Kikuchi Y, Segawa H, Tokumoto M, Tsub-okawa T, Hotta Y, Uyemura K, Oka-moto H. 1997. Ocular and cerebellardefects in zebrafish induced by overex-pression of the LIM domains of the Is-let-3 LIM/homeodomain protein. Neu-ron 18:369–382.
Kikuchi Y, Trinh LA, Reiter JF, Alexander J,Yelon D, Stainier DY. 2000. The zebrafishbonnie and clyde gene encodes a Mixfamily homeodomain protein that reg-ulates the generation of endodermalprecursors. Genes Dev 14:1279–1289.
Kikuchi Y, Agathon A, Alexander J, ThisseC, Waldron S, Yelon D, Thisse B, StainierDY. 2001. casanova encodes a novelSox-related protein necessary and suf-ficient for early endoderm formation inzebrafish. Genes Dev 15:1493–1505.
Kopan R, Goate A. 2000. A common en-zyme connects notch signaling andAlzheimer’sdisease.GenesDev14:2799–2806.
Latimer AJ, Dong X, Markov Y, Appel B.2002. Delta-Notch signaling induceshypochord development in zebrafish.Development 129:2555–2563.
Leimer U, Lun K, Romig H, Walter J, Grun-berg J, Brand M, Haass C. 1999. Ze-brafish (Danio rerio) presenilin pro-motes aberrant amyloid �-peptideproduction and requires a critical as-partate residue for its function in amy-loidogenesis. Biochemistry 38:13602–13609.
Lieber T, Kidd S, Alcamo E, Corbin V,Young MW. 1993. Antineurogenic phe-notypes induced by truncated Notchproteins indicate a role in signal trans-duction and may point to a novelfunction for Notch in nuclei. Genes Dev7:1949–1965.
Muller M, v. Weizsacker E, Campos-Or-tega JA. 1996. Expression domains of azebrafish homologue of the Drosophilapair-rule gene hairy correspond to pri-mordia of alternating somites. Devel-opment 122:2071–2078.
Mumm JS, Kopan R. 2000. Notch signal-ing: from the outside in. Dev Biol 228:151–165.
Park HC, Appel B. 2003. Delta-Notch sig-naling regulates oligodendrocytespecification. Development 130:3747–3755.
Peyrieras N, Strahle U, Rosa F. 1998. Con-version of zebrafish blastomeres to anendodermal fate by TGF-�-related sig-nalling. Curr Biol 8:783–786.
Poulain M, Lepage T. 20002. Mezzo, apaired-like homeobox protein is an im-mediate target of Nodal signaling and
regulates endoderm specification inzebrafish. Development 129:4901–4914.
Reiter JF, Alexander J, Rodaway A, YelonD, Patient R, Holder N, Stainier DY. 1999.Gata5 is required for the developmentof the heart and endoderm in ze-brafish. Genes Dev 13:2983–2995.
Reiter JF, Kikuchi Y, Stainier DY. 2001. Mul-tiple roles for Gata5 in zebrafishendoderm formation. Development128:125–135.
Ross DA, Kadesch T. 2001. The notch in-tracellular domain can function as acoactivator for LEF-1. Mol Cell Biol 21:7537–7544.
Sakaguchi T, Kuroiwa A, Takeda H. 2001.A novel sox gene, 226D7, acts down-stream of Nodal signaling to specifyendoderm precursors in zebrafish.Mech Dev 107:25–38.
Smithers L, Haddon C, Jiang Y, Lewis J.2000. Sequence and embryonic ex-pression of deltaC in the zebrafish.Mech Dev 90:119–123.
Stainier DY. 2002. A glimpse into the mo-lecular entrails of endoderm formation.Genes Dev 16:893–907.
Strahle U, Blader P, Henrique D, InghamPW. 1993. Axial, a zebrafish gene ex-pressed along the developing bodyaxis, shows altered expression in cy-clops mutant embryos. Genes Dev 7:1436–1446.
Struhl G, Fitzgerald K, Greenwald I. 1993.Intrinsic activity of the Lin-12 and Notchintracellular domains in vivo. Cell 74:331–345.
Takke C, Campos-Ortega JA. 1999. her1,a zebrafish pair-rule like gene, actsdownstream of notch signalling to con-trol somite development. Develop-ment 126:3005–3014.
Takke C, Dornseifer P, v. Weizsacker E,Campos-Ortega JA. 1999. her4, a ze-brafish homologue of the Drosophilaneurogenic gene E(spl), is a target ofNOTCH signalling. Development 126:1811–1821.
Warga RM, Nusslein-Volhard C. 1999. Or-igin and development of the zebrafishendoderm. Development 126:827–838.
Westerfield M. 1995. The zebrafish book: aguide for the laboratory use of ze-brafish. Eugene, OR: University of Ore-gon Press.
Wettstein DA, Turner DL, Kintner C. 1997.The Xenopus homolog of DrosophilaSuppressor of Hairless mediates Notchsignaling during primary neurogenesis.Development 124:693–702.
762 KIKUCHI ET AL.




















