Intrinsic and extrinsic innervation of the heart in zebrafish (Danio rerio)
Upload
independentCategory
view
0download
0
Organization of the Histaminergic System in AdultZebrafish (Danio rerio) Brain: Neuron Number,Location, and Cotransmitters
Maria Sundvik and Pertti Panula*
Neuroscience Center and Institute of Biomedicine, Anatomy, 00014 University of Helsinki, Finland
ABSTRACTHistamine is an essential factor in the ascending
arousal system (AAS) during motivated behaviors. Hista-
mine and hypocretin/orexin (hcrt) are proposed to be
responsible for different aspects of arousal and wake-
fulness, histamine mainly for cognitive and motivated
behaviors. In this study we visualized the entire histami-
nergic neuron population in adult male and female
zebrafish brain and quantified the histaminergic neuron
numbers. There were 40–45 histaminergic neurons in
both male and female zebrafish brain. Further, we iden-
tified cotransmitters of histaminergic neurons in the
ventrocaudal hypothalamus, i.e., around the posterior
recess (PR) in adult zebrafish. Galanin, c-aminobutyric
acid (GABA), and thyrotropin-releasing hormone (TRH)
were colocalized with histamine in some but not all
neurons, a result that was verified by intracerebroven-
tricular injections of colchicine into adult zebrafish.
Fibers immunoreactive (ir) for galanin, GABA, TRH, or
methionine-enkephalin (mENK) were dense in the
ventrocaudal hypothalamus around the histaminergic
neurons. In histamine-ir fibers TRH and galanin immu-
noreactivities were also detected in the ventral telen-
cephalon. All these neurotransmitters are involved in
maintaining the equilibrium of the sleep–wake state.
Our results are in accordance with results from rats,
further supporting the use of zebrafish as a tool to study
molecular mechanisms underlying complex behaviors.
J. Comp. Neurol. 520:3827–3845, 2012.
VC 2012 Wiley Periodicals, Inc.
INDEXING TERMS: histamine; thyrotropin-releasing hormone; galanin; c-aminobutyric acid; methionine-enkephalin;
hypothalamus
Modulatory neurotransmitter systems are essential
for mediating complex behaviors and disturbances in
these neurotransmitter systems are often associated
with neurological disorders, although causality with
symptoms is seldom well characterized. In mammals
histamine is a modulatory neurotransmitter with diverse
functions, including higher cognitive functions, alert-
ness, and wakefulness (Haas and Panula, 2003; Haas
et al., 2008; Dere et al., 2010). It is also implicated in
sleep disorders (Kanbayashi et al., 2009; Nishino et al.,
2009). The histaminergic neurotransmitter system was
described in rats (Panula et al., 1984; Watanabe et al.,
1984) and several other species including lamprey (Bro-
din et al., 1990), bony fish (Inagaki et al., 1991; Ekstr€om
et al., 1995; Eriksson et al., 1998), and human (Panula
et al., 1990; Airaksinen et al., 1991) with histamine-
producing neurons located exclusively in the posterior
hypothalamus in the tuberomammillary nucleus in mam-
mals and widespread projections innervating most brain
areas.
The histaminergic system is the initial neurotransmitter
system that is activated in the ascending arousal system
(AAS) during motivated behavior of rats (Valdes et al.,
2005). The hypocretin/orexin (hcrt) system sends
ascending projections to all parts of the AAS in verte-
brates (Szymusiak and McGinty, 2008) and is implicated
in narcolepsy (Lin et al., 1999; Peyron et al., 2000). The
AAS in mammals consists of neurotransmitter systems
with nuclei in subcortical areas, including the histaminer-
gic tuberomammillary nucleus, cholinergic basal forebrain
Additional Supporting Information may be found in the online version ofthis article.
Grant sponsor: Academy of Finland; Grant numbers: 116177, 207352;Grant sponsors: Finnish Technology Development Fund (TEKES); SigridJuselius Foundation; Helsinki Biomedical Graduate School (M.S.); MagnusEhrnrooth Foundation University of Helsinki Funds.
*CORRESPONDENCE TO: Pertti Panula, Neuroscience Center andInstitute of Biomedicine, Anatomy, POB 63 (Haartmaninkatu 8), 00014University of Helsinki, Finland. E-mail: [email protected]
VC 2012 Wiley Periodicals, Inc.
Received March 22, 2011; Revised January 17, 2012; Accepted April 16,2012
DOI 10.1002/cne.23126
Published online April 23, 2012 in Wiley Online Library (wileyonlinelibrary.com)
The Journal of Comparative Neurology | Research in Systems Neuroscience 520:3827–3845 (2012) 3827
RESEARCH ARTICLE

and pontomesencephalic nuclei, serotonergic raphe
nuclei, noradrenergic locus coeruleus, and the lateral
hypothalamic area containing the hcrt neurons (Szymu-
siak and McGinty, 2008). In several nonmammalian spe-
cies, including zebrafish (Kaslin and Panula, 2001; Kaslin
et al., 2004; McLean and Fetcho, 2004; Mueller et al.,
2004), the connections of these main monoaminergic
systems and the cholinergic system have been mapped.
Histamine-containing fibers innervate to varying degrees
the serotonergic raphe neurons, the noradrenergic locus
coeruleus, and the tyrosine hydroxylase-positive neu-
rons in the preoptic area, posterior tuberculum, and
paraventricular organ (PVO) (Kaslin and Panula, 2001)
and hcrt neurons in the preoptic area (Kaslin et al.,
2004) in zebrafish, but none of the observed neurotrans-
mitters were found in histaminergic neurons. Other neu-
rotransmitters, such as c-aminobutyric acid (GABA)
(Sherin et al., 1998), galanin (Toppila et al., 1995; Sherin
et al., 1998; Szymusiak et al., 2007), thyrotropin-releas-
ing hormone (TRH) (Nishino et al., 1997; Gonz�alez et al.,
2009; Hara et al., 2009; Parmentier et al., 2009), and
the endogenous opioid peptide methionine-enkephalin
(mENK) (Li and van den Pol, 2008) are also involved in
equilibrating the arousal/wakefulness state. The coloc-
alization of histamine with galanin, GABA, TRH, or mENK
has been studied earlier in rodents, and the localization
has been reported to be different in mouse, rat, and
guinea pig brain (Airaksinen et al., 1992). These markers
have not been studied with regard to histamine in
zebrafish.
Zebrafish (Danio rerio) has during the last decades
emerged and established itself as an important verte-
brate model organism within biomedical research and
neurosciences (Bandmann and Burton, 2010; Panula
et al., 2010; Sager et al., 2010). Due to its small size,
large clutch size, and rapid development it is a cost-
effective model organism to keep in large quantities.
The zebrafish shares many features with mammals,
e.g., the sleep/wake cycle (Prober et al., 2006; Yoko-
gawa et al., 2007; Appelbaum et al., 2009; Rihel et al.,
2010).
In larval zebrafish, histamine is necessary for sensori-
motor gating in dark-induced flash response and normal
locomotor activity during wakefulness (Sundvik et al.,
2011). In order to further study arousal and wakefulness
in zebrafish, and the involvement of the histaminergic sys-
tem, we analyzed some of the related systems, namely,
galanin, TRH, GABA, and mENK in the area harboring his-
taminergic neurons in the ventrocaudal hypothalamus in
this model organism. We hypothesized that some of these
neurotransmitters are likely to colocalize with histamine
in neurons in zebrafish, and that they may provide affer-
ent inputs to the histaminergic neurons and therefore cor-
elease of these neurotransmitters may modulate the
response of histamine associated with different behav-
iors. Although the histaminergic neurons send wide-
spread projections throughout the brain, we decided to
focus on the coexistence of histamine in ventrocaudal
hypothalamus with galanin, TRH, mENK, or GABA, and
how fibers immunoreactive (ir) for each neurotransmitter
are distributed with regard to histaminergic neurons and
fibers in this area. We also studied if histamine-ir fibers
in the telencephalon were positive for any of these
markers. Analyzing the cotransmitters of the histaminer-
gic system and afferent inputs is a prerequisite to carry
out relevant physiological and pharmacological experi-
ments on the role of histamine in arousal and wakeful-
ness. Larval zebrafish serve as a useful tool in studies on
neural basis of behavior (Burgess and Granato, 2007a,b;
Peitsaro et al., 2007). However, for studies related to
human brain diseases, experiments on adult or aging
fish (Peitsaro et al., 2003; Anichtchik et al., 2004) are
more relevant because, e.g., neurodegenerative dis-
eases usually arise late in life. Here we describe a
method to study adult zebrafish hypothalamic anatomy
three-dimensionally using a whole-mount technique to
reveal true numbers of neurons and detailed organiza-
tional patterns. This system may be applied to analysis
of intact circuits in adult brain.
MATERIALS AND METHODS
AnimalsIn this study we used adult zebrafish (6 months to 2-
year-old females and males) of a strain that has been
maintained in the laboratory for over a decade and used
in several studies (Kaslin and Panula, 2001; Kaslin et al.,
2004; Peitsaro et al., 2007; Sallinen et al., 2009a,b,
2010). The permits for the experiments were obtained
from the Office of the Regional Government of Southern
Finland in agreement with the ethical guidelines of the Eu-
ropean convention.
Intracerebroventricular (i.c.v.) injection ofcolchicine
Adult zebrafish (n¼ 4) were anesthetized in tricaine (3-
amino benzoic acid ethyl ester, also called ethyl 3-amino-
benzoate, Sigma, St. Louis, MO) and placed on ice. Under
the dissection microscope the skull bone of the fish was
lifted with forceps and 0.3 ll of 10 mg/ml colchicine
(Merck, Whitehouse Station, NJ) solution was injected
i.c.v. with a glass capillary. After the injection fish were
moved to 28�C tank water to recover from the anesthe-
sia. The animals were killed in ice-cold water 24 hours af-
ter the injection and the brains were processed for immu-
nohistochemistry (IHC) like the uninjected ones.
Sundvik and Panula
3828 The Journal of Comparative Neurology |Research in Systems Neuroscience

Antibody characterizationThe following primary antisera and antibodies were
used (Table 1): rabbit anti-HA19c (Panula et al., 1990)
1:10,000 and 1:50,000, rat anti-serotonin 1:250 (Milli-
pore/Chemicon, Billerica, MA, Cat. No. MAB352), mouse
anti-tyrosine hydroxylase (TH1) 1:1,000 (ImmunoStar,
Hudson, WI, Lot. No. 22941), rabbit anti-GABA (Karhunen
et al., 1993, 1H) 1:1,000, rabbit anti-TRH 1:1,000 (Antise-
rum #4319, Visser et al., 1977; Tsuruo et al., 1988a,b;
Airaksinen et al., 1992), rabbit anti-mENK 1:1,000 (Anti-
serum #36, Yang et al., 1977; Panula et al., 1984; Airaksi-
nen et al., 1992).
The antiserum against mENK was generated against a
mENK pentapeptide (from Peninsula Laboratories, San
Carlos, CA) conjugated with 1-ethyl-3,3(dimethyl-amino-
propyl) carbodiimide (EDAC, CMS Chemicals, Abingdon,
UK) to keyhole limpet hemocyanin (KLH) as described
(Yang et al., 1977; Panula et al., 1984). This antiserum
crossreacts with leucine-enkephalin 2–3%, with tetrapep-
tide Gly-Gly-Phe-Met about 1%, with tetrapeptide Tyr-Gly-
Gly-Phe less than 0.1%, and has no crossreactivity with a-
or b-endorphin in radioimmunoassay (Yang and Costa,
1979; Panula et al., 1984). The immunoreactivity in
rat brain sections is fully preabsorbed by mENK but not
with the extended mENK heptapeptide methionine-en-
kephalin-Arg-Phe or other related peptides (Panula et al.,
1984).
The TRH antiserum (Antiserum #4319) (Visser et al.,
1977; Visser and Klootwijk, 1981; Tsuruo et al., 1988a,b;
Airaksinen et al., 1992) was produced in rabbits against a
conjugate of TRH (Beckman Laboratories, Geneva, Swit-
zerland) and hemocyanin made with the bifunctional rea-
gent 1,5-difluoro-2,4-dinitrobenzene (Visser et al., 1977;
Visser and Klootwijk, 1981). It has been used extensively
in analysis of rodent, monkey, and human TRH systems
using immunocytochemistry (Tsuruo et al., 1988a–c;
Airaksinen et al., 1992; Arvidsson et al., 1992). This
immunoreactivity is fully preabsorbed by TRH peptide
(Peninsula Laboratories), but not by, e.g., histamine-
succinylated KLH (sKLH) or histamine-ovalbumin (OVA)
conjugates in sections of rat and guinea pig brain (Airaksi-
nen et al., 1992; Arvidsson et al., 1992). In human brain,
both solid-phase and liquid phase preabsorption of Anti-
serum #4319 with TRH peptide removes all staining (Fli-
ers et al., 1994). The staining was also identical with that
obtained with another highly specific antiserum (Fliers
et al., 1994).
The rabbit antiserum against histamine (rabbit anti-his-
tamine 19C, the third bleed following primary and sec-
ondary immunization) was produced against a histamine-
sKLH complex prepared with EDAC (Panula et al., 1990)
using a procedure described initially in 1984 for a related
histamine antiserum (Panula et al., 1984). In specificity
tests on nitrocellulose filters, this antiserum recognized
histamine coupled with EDAC to two different carrier pro-
teins, succinulated ovalbumin (sOVA) and bovine serum
albumin (BSA), but it did not recognize L-histidine in simi-
lar conjugates. Histamine coupled to sOVA or BSA totally
abolished the reactivity of this antiserum to histamine-
sKLH on nitrocellulose filter (Panula et al., 1990),
whereas L-histidine in similar conjugates or L-histidine-
containing peptides (e.g., LHRH) did not affect the reac-
tion. The antiserum detects histamine-immunoreactive
tuberomammillary neurons in rat, mouse, guinea pig (Air-
aksinen et al., 1992), and caudal hypothalamic neurons in
the zebrafish (Kaslin and Panula, 2001; Kaslin et al.,
2004). These neurons also express histidine decarboxyl-
ase (hdc), the sole histamine-synthesizing enzyme (Haas
and Panula, 2003). In zebrafish, the immunoreaction dis-
appears following administration of a-fluoromethylhist-
amine, suicide inhibitor of hdc (Peitsaro et al., 2003)
concomitantly with a decline in chromatographically
TABLE 1.
Primary Antibodies Used
Antigen Immunogen
Manufacturer, species antibody was raised in,
mono- vs. polyclonal, catalog or lot number Dilution used
GABA GABA- succinyl keyhole limpethemocyanin (sKLH)
Karhunen et al., 1993, rabbit polyclonal, codeGABA 1H, 7th bleed
1:1,000
Galanin Human galanin peptide-sKLH conjugate Millipore/Chemicon, rabbit polyclonal, AB5909 1:1,000 and 1:5,000Galanin Porcine galanin peptide Millipore/Chemicon, rabbit polyclonal, AB1985 1:1,000 and 1:5,000Histamine Histamine-sKLH conjugate Panula et al., 1984, 1990, rabbit polyclonal,
Antiserum #19c1:50,000
mENK mENK peptide-KLH conjugate Yang et al., 1977; Panula et al., 1984; Airaksinenet al., 1992, rabbit polyclonal, Antiserum #36
1:1,000
TH1 Mouse tyrosine hydroxylase purifiedfrom rat PC12 cells
ImmunoStar/Diasorin, mouse monoclonal, 22941 1:1,000
TRH TRH peptide Visser et al., 1977; Tsuruo et al., 1988a,b;Airaksinen et al., 1992, rabbitpolyclonal, Antiserum #4319
1:1,000
5-HT Serotonin Millipore/Chemicon, rat monoclonal, MAB352 1:250
The Journal of Comparative Neurology | Research in Systems Neuroscience 3829
Organization of zebrafish histaminergic system

measurable histamine. Translation inhibition of hdc
mRNA with morpholino oligonucleotides in larval zebra-
fish also removes the immunoreactivity (Sundvik et al.,
2011).
The antiserum against GABA (rabbit anti-GABA 1H, the
seventh bleed after primary and secondary immunization)
was prepared against GABA (Sigma) conjugated with
EDAC to succinylated KLH as described in detail (Karhu-
nen et al., 1993). In dotblot tests the antiserum detected
GABA conjugated with EDAC to sOVA, and traces of gly-
cine and glutamate similarly blotted (Karhunen et al.,
1993). In preabsorption tests, only GABA-sOVA conjugate
but not glycine or glutamate conjugate in the antibody so-
lution removed the immunoreaction from the filter. Simi-
larly, in tissue sections only GABA conjugate and not glu-
tamate or glycine conjugate abolished the reaction
obtained with the GABA antiserum.
The commercially available mouse monoclonal anti-
body against TH used here (Diasorin/Immunostar, Lot.
No. 22941) has been produced against TH isolated from
rat PC12 cells. In zebrafish it recognizes TH1 but not TH2
in double-staining procedures with cRNA probes for th1
or th2 (Chen et al., 2009), strongly suggesting that it only
detects TH1 and not TH2. This antibody detects one band
in homogenates of adult zebrafish brain (Anichtchik et al.,
2004).
The rat monoclonal serotonin (5-HT) antibody (Chemi-
con, Cat. No. MAB352) is produced against 5-HT conju-
gated to BSA with formaldehyde. The antibody shows no
crossreactivity to catecholamines, GABA, 5-hydroxytryp-
tophan, carnosine, or melatonin in fixed tissue sections.
In our pilot tests on larval zebrafish brains, it did not
stain neurons expressing th1, th2, hdc, and in adult
zebrafish brain it gave an identical staining pattern with
a previously used mouse monoclonal antibody against 5-
HT (Kaslin and Panula, 2001). This antiserum has been
previously used in zebrafish and found to give identical
staining with another rabbit antiserum against 5-HT (Lil-
lesaar et al., 2009).
The rabbit antiserum against porcine galanin (Chemi-
con, Cat. No. AB1985) has been produced against full-
length porcine galanin peptide. It has no crossreactivity
to cholecystokinin, galanin 10-29, somatostatin, sub-
stance P, growth hormone, or prolactin (Magliulo-
Cepriano et al., 1993). The rabbit antiserum against gala-
nin (Chemicon, Cat. No. AB5909) has been made against
the whole human galanin peptide conjugated to KLH. We
did the preabsorption test for both antibodies in zebrafish
tissue.
These antisera have been tested previously for specific-
ity in different mammalian species. The commercial rabbit
antiserum against human galanin (AB5909) and rabbit anti-
serum against porcine galanin (AB1985) were diluted
1:1,000–1:5,000 and tested using preabsorption tests
here. For detection of the primary antibody amplified with
the tyramide signal amplification (TSA, PerkinElmer Life
and Analytical Sciences, Boston, MA) method (the primary
antibody to be detected with TSA was in all cases rabbit
anti-histamine 1:50,000), we used a biotinylated second-
ary goat antirabbit antibody diluted 1:750 (Vectastain ABC
kit, Vector Laboratories, Burlingame, CA). Texas Red (Vec-
tor Laboratories) conjugated avidin (diluted 1:500) and
streptavidin conjugated fluorescein (Molecular Probes/
Invitrogen, Carlsbad, CA, diluted 1:50) were used to further
visualize the biotinylated antibody. To visualize the primary
antisera against serotonin, TH, galanin, GABA, TRH, mENK,
and histamine (1:10,000 in preabsorption experiment),
highly crosspurified Alexa-fluorophore conjugated goat
antirabbit antibodies (with either 488 or 561 fluorophore,
Molecular Probes) were used.
Since the antibodies of this study have been mostly
used for rodent studies, additional controls for histamine,
galanin, mENK, TRH, and GABA primary antibodies were
performed on zebrafish brain sections. The primary anti-
body and conjugate or peptide was incubated together at
4�C before IHC. The peptides and conjugates are listed in
Table 2. Histamine, serotonin, and TH antibodies have
been evaluated for zebrafish previously (Kaslin and Pan-
ula, 2001; Sallinen et al., 2009a,b). Please note that the
zebrafish has two forms of TH (Chen et al., 2009) and the
antibody used in this and previous studies detect only the
TH1 form.
TABLE 2.
Preabsorption Controls
Antigen
Preabsorption
conjugate or peptide Manufacturer, catalog number
Dilution used
(of conjugate/peptide)
GABA GABA-sOVA Karhunen et al., 1993 20 lg/mlGalanin Human galanin peptide BACHEM, H-8230 10 lMGalanin Porcine galanin peptide BACHEM, H-1365 50lMHistamine Histamine-sOVA Panula et al., 1984 2 lg/mlmENK mENK peptide Sigma-Aldrich, E-5757/ M-6638 100 lMTRH TRH peptide Peninsula Laboratories, H-4915 10 lM– Porcine dynorphin peptide Peninsula Laboratories, PEN-8697 100lM
3830 The Journal of Comparative Neurology |Research in Systems Neuroscience
Sundvik and Panula

Immunohistochemistry (IHC)The Tyramide Signal Amplification kit (TSA) was used
to enable double staining of samples with primary anti-
bodies from the same species. Brains were collected
from adult zebrafish sacrificed in ice-cold water. They
were fixed in 4% EDAC and 0.1% paraformaldehyde (PFA,
Sigma) in 0.1 M phosphate buffer pH 7.0 at 4�C. For
whole-mount IHC the brains were immediately processed,
whereas brains intended for sections and TSA procedures
were incubated in 20% sucrose in 0.1 M phosphate
buffer, pH 7.4, at 4�C, thereafter these brains were em-
bedded in embedding matrix and sectioned at 20 lm
with a Leica cryostat. Sections were stored at �20�C
until further processed. IHC was performed at room tem-
perature (RT) unless otherwise mentioned.
Whole-mount IHC of adult zebrafish brains was done in
baskets that fit the wells of a 24-well multiwell plate. Im-
mediately after fixation the brains were rinsed with phos-
phate buffered saline (PBS) 3 times and then moved to the
baskets. Brains were washed 3 � 30 minutes in PBS-Triton
X-100 0.3% (PBSTx) and incubated in preincubation buffer
(1% dimethylsulfoxide and 4% normal goat serum in PBSTx)
for 4 hours at RT or at 4�C. The incubation with the primary
antibody for histamine and 5-HT or TH1 in the same buffer
lasted for 2–4 days. After incubation with the primary anti-
bodies the samples were washed 10 minutes þ 3 � 60
minutes in PBSTx and incubated with appropriate Alexa-flu-
orophore conjugated secondary antibodies (goat antirabbit
and goat antirat or goat antimouse) diluted in the preincu-
bation buffer at 4�C. Samples were then washed for 10
minutes followed by 3 � 60 minutes in PBSTx and infil-
trated with glycerol 2 � 60 minutes in glycerol/PBS (50%)
and at 4�C in glycerol/PBS (80%). Next the samples were
mounted on slides in the space between two sets of dou-
ble 18 � 18 mm cover glasses attached to each other with
silica grease. The ventral side of the brain was mounted to-
ward the top cover glass, allowing microscopy using long
working distance objectives of the posterior hypothalamus
that harbors the histaminergic and some of the serotoner-
gic and dopaminergic neurons.
IHC with TSA was performed according to the manufac-
turer’s instructions. Samples were kept for 20 minutes at
RT and washed with PBS for 5 minutes. Endogenous per-
oxidase was blocked by incubation of the slides in 0.3%
H2O2 for 15 minutes. Samples were then rinsed in 0.1%
PBSTx for 10 minutes and incubated with blocking buffer
(0.5% blocking reagent in PBSTx) for 30 minutes. Immuno-
staining for histamine was the only procedure in which
the TSA method was used. Incubation with the histamine
antiserum was done at 4�C. The antiserum was diluted in
blocking buffer. The next day the samples were rinsed 3
times for 5 minutes in PBSTx, incubated with a biotinyl-
ated secondary antirabbit antibody in blocking buffer for
30 minutes, rinsed 3 � 5 minutes in PBSTx, incubated
with streptavidin conjugated horseradish peroxidase (SA-
HRP) diluted 1:100 in blocking buffer for 30 minutes, and
rinsed 3 � 5 minutes in PBSTx. The next step was the crit-
ical amplification step, and Biotinyl Tyramide Amplifica-
tion Reagent was diluted into Amplification diluent 1:50
and the samples were incubated in the amplification
buffer for 3–10 minutes. Samples were quickly rinsed 3
� 5 minutes in PBSTx before incubation with the fluoro-
phore-conjugated second antibody (either Texas Red or
Fluorescein) diluted in blocking buffer for 30 minutes,
rinsed 3 � 5 minutes in PBSTx. After the IHC with TSA
method, the samples were incubated with the second pri-
mary antibody solution to obtain the desired double stain-
ing. The second incubation procedure was started imme-
diately after the IHC with TSA was finished. The antisera
and antibodies used for the second primary antibody
incubation were diluted in blocking buffer and the sam-
ples were incubated at 4�C. The next day the samples
were rinsed 3 � 5 minutes in PBSTx and incubated with
secondary Alexa-fluorophore conjugated antibodies (ei-
ther 488 or 568 wavelength fluorophore, depending on
which fluorophore was initially applied to the samples
during the IHC with TSA) diluted in blocking buffer for 1–2
hours. Samples were then rinsed in PBSTx for 5 minutes
and in PBS 3 � 5 minutes and mounted in glycerol/PBS
(50% or 80%, depending on what objectives were to be
used). All stainings were done on at least three
individuals.
Microscopy and image analysisThe samples were analyzed with a Leica TCS SP2
AOBS confocal microscope (Leica Microsystems, Mann-
heim, Germany). The objectives used were the following:
Leica HC PL APO 10�/0.40 CS, HC PL APO 20�/0.70
CS, HCX PL APO 63�/1.2 W CORR CS. Excitation wave-
length for samples with streptavidin conjugated fluores-
cein or Alexa 488 fluorophore conjugated antibodies was
488 nm, and the emission was collected at 500–550 nm.
The excitation wavelength used for samples stained with
Texas Red conjugated avidin or Alexa 658 fluorophore
conjugated antibodies was 561 nm and emission was col-
lected at 600–700 nm or 590–630 nm in cases where tri-
ple-staining was applied (the narrower setting was used
for the whole-mount IHC of adult zebrafish brain) to pre-
vent band overlap.
Whole-mount IHC allowed the visualization of the intact
histaminergic neuron population in the posterior hypo-
thalamus. The brains were scanned with optimized step
size and averaging to allow 3D rendering of the data in
Imaris (Bitplane, Zurich, Switzerland). The 3D-rendered
images were exported as snapshots and movies. Quantifi-
cation of histamine neuron number was done by counting
The Journal of Comparative Neurology | Research in Systems Neuroscience 3831
Organization of zebrafish histaminergic system

the neurons from the entire stacks of images that had
been acquired with optimal settings.
From each adult zebrafish brain a single sagittal sec-
tion through the ventrocaudal hypothalamus was studied
per each staining condition, which assured that the same
neuron was not counted several times within the same
condition, and allowed us in several cases to study hista-
mine together with four different antibodies/antisera in
each individual. On the slide, the section with most hista-
mine-ir neurons was chosen, scanned, and analyzed. In
some cases this section contained only a few (2–3) hista-
minergic neurons. We were interested in whether the
neurotransmitters were found in the histamine neurons,
rather than in the fibers surrounding the histaminergic
neurons, and because of this the actual analysis was
done manually and not with a colocalization algorithm.
Single focal planes of overlay images were analyzed for
colocalization with Leica software or the open source
image analysis software Fiji (http://pacific.mpi-cbg.de/
wiki/index.php/Main_Page). The number of neurons that
coexpress two markers was counted from the whole
stacks through selected sections. Maximum intensity pro-
jection algorithm was applied to the acquired stacks with
Leica software for comparative analysis (Fig. 1). 3D-ren-
dering and an example of automated colocalization was
performed with default algorithm settings in Imaris (Bit-
plane) and exported as snapshots. Deconvolution with
Huygens Deconvolution Software (Scientific Volume
Imaging, Hilversum, The Netherlands) was applied to
some of the datasets, but there seemed to be no detecta-
ble difference between non- and deconvoluted data. All
pictures were further processed and compiled into panels
in CorelDraw (Ottawa, Canada).
Statistical analysisStudent’s t-test was applied to assess whether statisti-
cal significance could be found between two groups.
RESULTS
Antibody characterizationIn this study we performed IHC with the TSA method,
as this was the only approach that allowed us to utilize
Figure 1. Preabsorption controls for primary antibodies and antisera used in the study. In all cases the immunoreactivity was abolished
with the peptide or conjugate the antibody was raised against. More detailed information about the antibodies, peptides and conjugates is
found in Table 1. (a) anti-histamine, (b) anti-GABA, (c) anti-galanin, (d) anti-mENK, (e) anti-TRH, (f) anti-histamine and histamine-sOVA con-
jugate 2 lg/ml, (g) anti-GABA and GABA-sOVA conjugate 20 lg/ml, (h) antihuman galanin and human galanin peptide 10 lM, (i) anti-
mENK and mENK peptide 100 lM, (j) anti-TRH and TRH peptide 10 lM. All images were acquired from the ventrocaudal hypothalamus
around the posterior recess (PR), n ¼ 3 in each group. Filled arrowheads indicate positive staining and empty arrowheads indicate lack of
staining. Anterior to left. Scale bar ¼ 100 lm.
3832 The Journal of Comparative Neurology |Research in Systems Neuroscience
Sundvik and Panula

the best available antisera, all produced in the same
species, rabbit. Further, this study was done entirely on
adult zebrafish brains. Preabsorption controls have
been performed for the antibodies used in other species
at an earlier stage (Panula et al., 1990; Airaksinen et al.,
1992) and we repeated this for the zebrafish samples
(Table 1, Fig. 1). Histamine-producing neurons are
found exclusively in the nucleus of the posterior recess
in the ventrocaudal hypothalamus of zebrafish (Eriksson
et al., 1998; Kaslin and Panula, 2001). The staining with
histamine antiserum was completely abolished with
preabsorption of the antiserum with histamine-succinyl
ovalbumin (sOVA) conjugate (2 lg/ml, Fig. 1a,f). The
GABA staining was similarly abolished with GABA-sOVA
conjugate (20 lg/ml, Fig. 1b,g). It is important to note
that these two antisera have been produced against
related conjugates made with sKLH, which is an unre-
lated protein. Thus, the antisera are directed against
protein-bound histamine and GABA, respectively. Immu-
nostaining with antisera against the peptides was also
abolished when the antisera was preabsorbed with the
corresponding peptides. In this study we used two dif-
ferent galanin antisera (AB5909 and AB1985). These
antisera gave the same staining pattern, but were pre-
absorbed with different peptides. AB5909 was com-
pletely preabsorbed with 10 lM human galanin peptide
(Fig. 1c,h) and AB1985 was similarly preabsorbed with
50 lM porcine peptide (data not shown). As the staining
pattern was identical, in the following we pooled the
results acquired with the different antibodies. The anti-
sera against mENK and TRH were also completely pre-
absorbed with the following peptide concentrations
mENK (100 lM, Fig. 1d,i) and TRH (10 lM, Fig. 1e,j).
The specificity of mENK antisera was further studied by
absorption of the antisera with dynorphin peptide. In
zebrafish tissue this preabsorption did not wipe out the
mENK staining (results not shown), indicating that the
antisera detects mENK but not related peptides from
prodynorphin.
For visualizing an antigen with the TSA method two
additional controls are recommended by the manufac-
turer, a negative control and an amplification control. A
negative control, without the primary antibody (hista-
mine) but including all TSA reagents yielded no signal.
When the (histamine) antibody was included but all TSA
reagents were excluded, the amplification control, no
reaction was seen either.
Whole-mount visualization of histaminergicsystem in adult zebrafish
The nucleus containing the histaminergic neurons is
very ventrally located in the brain of zebrafish. Due to this
ventral location of the histaminergic neurons and the
small size of the brain, it was possible to quantify the total
number of histaminergic neurons in the whole-mount
brain of adult zebrafish. 3D-rendered data showed that
most of the histaminergic neurons can be found in the
most ventral area (Fig. 2a). There was no significant dif-
ference between the average neuron numbers in males
(45.7 6 5.2) and females (41.4 6 5.8, P ¼ 0.57). To ver-
ify that we were able to visualize all histamine-ir neurons
in the whole-mount adult brain, we assessed the location
of the histaminergic neurons in comparison to the seroto-
nergic and dopaminergic (TH1-ir) neurons in this region.
The data were compared to an earlier study done with the
same antibodies but on sections from adult brain (Kaslin
and Panula, 2001), and we found that all the histaminer-
gic neurons were detected in the whole-mount brain
preparation. Crosstalk between the channels and back-
ground noise were eliminated with sequential scanning
between frames, adjusting the black level of the confocal
microscope and by frame averaging (set to 2) as
described (Sallinen et al., 2009a). We observed in 3D-ren-
dered images of the intact ventrocaudal hypothalamus
that histaminergic and serotonergic neurons were local-
ized in specific areas (Fig. 2b). Histaminergic neurons are
located in the ventrocaudal hypothalamus surrounding
the posterior recess, i.e., in the edges of the caudal zone
of periventricular hypothalamus (Hc). The medial part of
Hc harbors the serotonergic neurons (Fig. 2d–l). The most
ventrolateral areas of the serotonergic Hc were inner-
vated by histaminergic processes (Fig. 2j–l). Histaminer-
gic neurons occupied both more dorsal and ventral areas
than the serotonergic neurons (Fig. 2d,g,l). Zebrafish
have two genes encoding tyrosine hydroxylase, th1 and
th2 (Chen et al., 2009). In this study we visualized the do-
paminergic neurons with an antiserum that is targeted to-
ward TH1 and that has also been used in several other
studies to detect dopaminergic neurons (Kaslin and Pan-
ula, 2001; Sallinen et al., 2009a). The distribution pattern
of the TH1-ir neurons with regard to histaminergic neu-
rons (Fig. 2c) resembled the organization of the seroto-
nergic and histaminergic neurons (Fig. 2b). TH1-ir neu-
rons are located in the same area as serotonergic
neurons, in medial Hc that did not have major histaminer-
gic input (Fig. 2m–u). In the ventrolateral parts of the Hc,
histamine-ir processes were in close contact with the
TH1-ir neurons (Fig. 2s,t) in the same manner as that
observed for the serotonergic neurons (Fig. 2j–l). Both
histaminergic and TH1-ir neurons had approximately the
same dorsal location as neurons of both systems
observed at the same tissue depth (Fig. 2m). TH2 might
colocalize with histamine as both markers are abundant
in the ventrocaudal hypothalamus, however this needs to
be studied.
The Journal of Comparative Neurology | Research in Systems Neuroscience 3833
Organization of zebrafish histaminergic system

Figure 2. Visualization of the intact histaminergic system in a whole-mount sample of adult zebrafish brain. a: 3D-rendered histamine system
in the adult brain. b: The anatomical relationships between histaminergic (green) and serotonergic (magenta) neurons in the ventrocaudal
hypothalamus (n ¼ 4). A snapshot image of a 3D-rendered image. c: The anatomical relationships between histaminergic (green) and dopami-
nergic (magenta, TH) neurons in the ventrocaudal hypothalamus (n ¼ 8). A snapshot image of a 3D-rendered image. The appearance of
colocalization is due to the high density of TH-ir neurons. d–l: Overlay images of histamine (green) and serotonin (magenta) throughout the
ventrocaudal hypothalamus. m–u: Overlay images of histamine (green) and TH-ir (magenta) throughout the ventrocaudal hypothalamus. Hc,
caudal zone of periventricular hypothalamus; PR, posterior recess. Anterior toward top of page in all images. Scale bars ¼ 50 lm.
3834 The Journal of Comparative Neurology |Research in Systems Neuroscience
Sundvik and Panula

Innervation of histaminergic neurons bygalanin-, TRH-, mENK-, or GABA-ir fibers
The innervation patterns of fibers immunoreactive for
one of four different transmitters, galanin, TRH, mENK, or
GABA, with regard to histaminergic neurons were eval-
uated using confocal microscopy, as described in the pre-
vious section. In zebrafish, a dense histaminergic fiber
network surrounds the histaminergic somata (Eriksson
et al., 1998; Kaslin and Panula, 2001). We found that in
adult zebrafish a dense galanin-ir fiber network was pres-
ent around the histaminergic neurons (Fig. 3a–c) in the
ventrocaudal hypothalamus. The pattern of TRH-ir fibers
resembled that of galanin, but the TRH-ir fiber network
around the histaminergic neurons was denser (Fig. 3d–f).
Similar fiber distribution pattern was observed with
mENK-ir fibers, as these are found close to histaminergic
neurons (Fig. 3g–i). GABA-ir fiber terminals were also
observed in close contact with the histaminergic neurons
(Fig. 3j–l).
Coexisting transmitters in histaminergicneurons
To assess the colocalization of other transmitters with
histamine we counted the total number of histamine-ir
neurons and analyzed the coexistence of histamine and
each of the four transmitters studied at each time. The
data were acquired by analyzing stacks of images
acquired with �0.2–1.0-lm intervals from 20-lm thick
samples. An example of a neuron positive for both hista-
mine and galanin is shown in Figure 4a–j. 3D-rendering of
the data illustrated the same colocalization as observed
when single focal plane images were analyzed (Fig. 4a vs.
4b–j). Analysis with standard parameters for colocaliza-
tion algorithm in Imaris revealed the same pattern (data
not shown). The software calculates the number of voxels
that contain two colors. Because we were interested in
whether or not the neurotransmitters are present in hista-
minergic neuronal cell bodies rather than in surrounding
fibers or terminals, we analyzed the data manually
through whole stacks, as shown in Figure 4b–j.
The number of histaminergic neurons that were posi-
tive for histamine and a second marker were quite few
and the majority of the neurons displayed only histamine
immunoreactivity (compiled in Table 3). We found that
galanin was colocalized with histamine (Movie 1a; 63�magnification and 5.41� zoom, and 1b; only 63� magni-
fication in Supporting Information), as we detected 25
galanin-ir and histamine-ir positive neurons within a total
of 79 histamine-ir neurons in 11 individuals (Fig. 4k–m).
Approximately 30% of the histaminergic neurons
observed were therefore positive for both histamine and
galanin. This result rendered galanin the most commonly
coexisting neuropeptide in histaminergic neurons in our
study. TRH was also colocalized in the histaminergic neu-
rons (Fig. 4n–p and Movie 2 in Supporting Information),
but only in a very few cases. We detected only two dou-
ble-stained neurons for TRH-ir and histamine-ir of 28 his-
tamine-ir neurons observed in total, i.e., 7% in six individu-
als. mENK was not found in the histaminergic neurons as
not a single double-stained neuron for both mENK and
histamine was found within the total of 51 histamine-ir
observed neurons (Fig. 4q–s and Movie 3 in Supporting
Information) in eight individuals. Finally, we found that
GABA was colocalized with histamine in a few cases
(Fig. 4t–v and Movie 4 in Supporting Information). We
detected eight GABA-ir and histamine-ir positive neurons
of a total of 54 histamine-ir neurons from 11 individuals.
This corresponds to about 14% of the histaminergic
neurons.
To increase the detection of neuropeptides within neu-
ronal somata, axonal transport can be disrupted by col-
chicine (to perturb microtubule integrity) treatment
before the animals are sacrificed, as previously demon-
strated (Johansson and H€okfelt, 1980). This treatment,
however, may activate genes that are not normally
expressed in cells and this may lead to false-positive
results (R�ethelyi et al., 1991). Despite this, as there were
quite few histaminergic neurons that were positive for
another neurotransmitter, we did i.c.v. injections of col-
chicine into adult zebrafish. The result corroborated the
earlier finding that galanin, GABA, and TRH are cotrans-
mitters of histaminergic neurons. The data are presented
as Supporting Movies 5–8.
Distribution of galanin-, TRH-, mENK-, andGABA-ir with regard to histamine-ir intelencephalon
The region with the highest density of afferent hista-
mine-ir fibers is the dorsal and to a lesser extent the ven-
tral telencephalon in zebrafish. We studied the distribu-
tion of all five neurotransmitters in telencephalon, and
only detected double labeling in the ventral part. We
found that galanin-ir fibers are sparse in the dorsal telen-
cephalon (Fig. 5a–c) and that the fibers mainly innervate
the ventral telencephalon. The same pattern of immuno-
reactive fiber distribution was observed for TRH (Fig. 5d–
f) and mENK (Fig. 5g–i). In a few cases the histamine-ir
fibers also showed galanin-ir (insert in Fig. 5c and Movie
9 in Supporting Information) and TRH-ir (inset in Fig. 5f
and Movie 10 in Supporting Information), whereas no his-
tamine-ir fibers were positive for mENK (insert in Fig. 5i
and Movie 11 in Supporting Information). mENK-ir neu-
rons were found in magnocellular preoptic nucleus (PM),
which was not innervated by histamine-ir fibers (Fig. 5j–l).
The Journal of Comparative Neurology | Research in Systems Neuroscience 3835
Organization of zebrafish histaminergic system
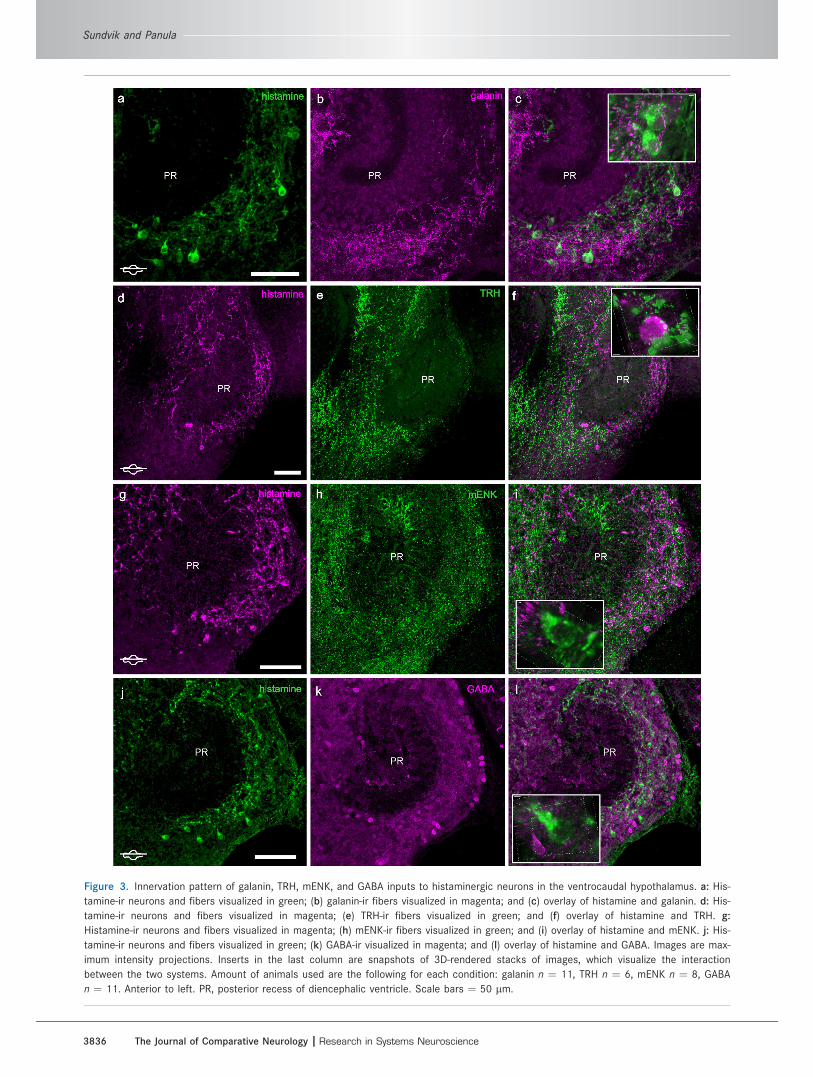
Figure 3. Innervation pattern of galanin, TRH, mENK, and GABA inputs to histaminergic neurons in the ventrocaudal hypothalamus. a: His-
tamine-ir neurons and fibers visualized in green; (b) galanin-ir fibers visualized in magenta; and (c) overlay of histamine and galanin. d: His-
tamine-ir neurons and fibers visualized in magenta; (e) TRH-ir fibers visualized in green; and (f) overlay of histamine and TRH. g:
Histamine-ir neurons and fibers visualized in magenta; (h) mENK-ir fibers visualized in green; and (i) overlay of histamine and mENK. j: His-
tamine-ir neurons and fibers visualized in green; (k) GABA-ir visualized in magenta; and (l) overlay of histamine and GABA. Images are max-
imum intensity projections. Inserts in the last column are snapshots of 3D-rendered stacks of images, which visualize the interaction
between the two systems. Amount of animals used are the following for each condition: galanin n ¼ 11, TRH n ¼ 6, mENK n ¼ 8, GABA
n ¼ 11. Anterior to left. PR, posterior recess of diencephalic ventricle. Scale bars ¼ 50 lm.
3836 The Journal of Comparative Neurology |Research in Systems Neuroscience
Sundvik and Panula

Figure 4. Quantification of colocalization and localization of galanin, TRH, and GABA but not mENK in histaminergic neurons around the
posterior recess. a: 3D rendering of histamine and galanin-ir in a single neuron in zebrafish. b–j: A series of single focal plane images
taken from a 20-lm thick stack shows the localization of histamine and galanin in a single cell in the ventrocaudal hypothalamus. Hista-
mine-ir is visualized in magenta and galanin-ir in green. k–l: Histamine-ir (magenta) and galanin-ir (green). m: Overlay of histamine and gal-
anin, showing the colocalization of galanin with histamine in a neuron. n–o: Histamine-ir (magenta) and TRH-ir (green). p: Overlay of
histamine and TRH, showing the colocalization of TRH and histamine. q–r: Histamine-ir (magenta) and mENK-ir (green). s: Overlay of hista-
mine and mENK, showing the lack of colocalization of mENK and histamine in neurons. t–u: Histamine-ir (magenta) and GABA-ir (green). v:
Overlay of histamine and GABA, showing the colocalization of GABA and histamine in a neuron. Single focal plane images are presented
to exclude false detection of colocalization. Only (a) contains information from a stack of images. Arrows indicate the approximate area
that was assessed for colocalization. Animals used: galanin n ¼ 11, TRH n ¼ 6, mENK n ¼ 8, GABA n ¼ 11. Anterior to left. Scale bars
¼ 5 lm in a; 20 lm in all other panels.

The scattered GABAergic neurons in the dorsal telen-
cephalon were found to be contacted by histamine-ir
fibers (Fig. 5m–o) and a higher magnification illustrates
this more clearly (insert in Fig. 5o).
DISCUSSION
In this study we show that nerve fibers immunoreactive
either for GABA, galanin, or TRH innervate the histaminer-
gic neurons and these neurotransmitters are to a low
degree colocalized with histamine in zebrafish. mENK and
histamine do not colocalize but mENK-ir fibers are dense
around the histaminergic neurons. The results are in ac-
cordance with what has been found in rat (Kohler et al.,
1986; Airaksinen et al., 1992), although the number of
neurons and fibers in which two markers coexist is low in
zebrafish. Thus, this study does not indicate functional
importance of the coexistence. Previous studies, includ-
ing those from our group (Airaksinen et al., 1992; Kukko-
Lukjanov and Panula, 2003), have strived to identify the
cotransmitters of histaminergic neurons. In this case the
TSA approach has not been used, but rather different
markers for histaminergic neurons such as vesicular
monoamine transporter (Kukko-Lukjanov and Panula,
2003) was used for double staining of cultured rat hista-
minergic neurons, or the mirror sectioning method was
applied to rat brain (Airaksinen et al., 1992) to define
whether histamine is present with some other neuro-
transmitters or not. In the present study it is likely that
the counts obtained underestimate the number of neu-
rons that produce several transmitters. We detected
quite elaborate neuropeptide fiber networks, whereas we
did not observe many neuropeptide-positive cell somata.
Results from the colchicine injection further verified the
coexistence data. Another approach would have been to
detect the mRNA of neuropeptides by in situ hybridization
(ISH). However, successful histamine staining with the
antiserum against histamine requires specific fixation
with the bifunctional fixative EDAC, which is not compati-
ble with ISH. We therefore decided to perform IHC of the
peptides for identification instead of ISH. Published
results on mRNA expression of the studied transmitters
in zebrafish confirm that GABAergic neurons are found in
ventrocaudal hypothalamus (Gonz�alez-Nu~nez et al.,
2005). The expression trh mRNA has so far only been
detected in preoptic region (Lohr et al., 2009) in zebra-
fish, whereas TRH-ir neurons have been detected in pos-
terior hypothalamus (Diaz et al., 2002). For mENK there
no available published data on mRNA expression in zebra-
fish. The galanin gene is expressed in four paired groups
of neurons in developing zebrafish brain (Podlasz et al.
2012), and one of these groups located in caudal periven-
tricular hypothalamus overlaps with the histamine cell
group. All studied neurotransmitters are likely to be im-
portant for the function of histaminergic neurons, as they
are colocalized and/or innervate the histaminergic neu-
rons. A discrepancy between the neurotransmitter pheno-
type of the histaminergic neurons has been reported
between rat, mouse, and guinea pig (Airaksinen et al.,
1992). In rat, histamine is colocalized with GABA, galanin,
and TRH but not with mENK (Airaksinen et al., 1992). Fur-
thermore, in mouse histamine was colocalized with GABA
and galanin, whereas in guinea pig histamine was colocal-
ized with GABA and mENK (Airaksinen et al., 1992). In
human tuberomammillary nucleus histamine coexists
with GABA but not with galanin (Trottier et al., 2002). In
this study we found that the zebrafish histaminergic neu-
rons have a similar neurotransmitter phenotype as the rat
histaminergic neurons. Therefore, the similarities of the
rat and zebrafish systems encourage the use of zebrafish
as a simple tool and model organism to study the molecu-
lar and cellular mechanisms of complex behaviors.
Intact histaminergic system in adultzebrafish brain
In this study we visualized the histaminergic neurons in
whole-mount adult zebrafish brain and quantified the
exact neuron numbers in both males and females. No
significant differences were observed between the sexes.
The anatomical distributions of the serotonergic and do-
paminergic neurotransmitter systems were in accordance
with what was previously reported on sections from adult
zebrafish brain (Kaslin and Panula, 2001). We merely
used this as a map to be certain that the entire histamine
neuron population was detected. Histamine-ir processes
extended to the ventrolateral Hc area, which expressed
densely packed serotonergic and TH1-ir neurons. Hista-
minergic neurons occupied both more dorsal and ventral
areas than the serotonergic neurons. A similar distribu-
tion pattern was observed for the histaminergic neurons
with regard to TH1-ir neurons, although some of the TH1-
ir neurons were also observed as dorsally as the histami-
nergic neurons. Taken together, the core of Hc surround-
ing PR was devoid of histaminergic neuron somata, and
as shown earlier, the histaminergic neurons were located
on the edges of Hc (Kaslin and Panula, 2001). We were
TABLE 3.
Quantified Colocalization in Wildtype Adult Zebrafish
Brain
Total number of
histaminergic cells
examined for double label
Number of histaminergic
cells that colabel with
the particular marker
Galanin 54 25TRH 26 2mENK 51 0GABA 46 8
Sundvik and Panula
3838 The Journal of Comparative Neurology |Research in Systems Neuroscience

Figure 5. Analysis of galanin-, TRH-, GABA-, and mENK-ir fibers and cells in the telencephalon. a,d,g,m: Histamine-ir fibers in the telen-
cephalon innervate both the dorsal (D) and the ventral (V) telencephalon. b: Galanin-ir fibers are found in low density throughout the telen-
cephalon. c: Overlay of histamine- and galanin-ir in telencephalon. A small fraction of the fibers were double positive for both galanin and
histamine (indicated in white and with arrows). e: TRH-ir fibers are mainly detected in V. f: Overlay of histamine- and TRH-ir. A small por-
tion of the fibers were positive for TRH and histamine (indicated in white and with arrows). h: mENK-ir fiber distribution is similar to TRH-ir
with the highest intensity of fibers observed in V. i: Overlay of histamine- and mENK-ir. No fibers were found to be positive for both hista-
mine and mENK. j: No histamine-ir was detected in the magnocellular preoptic nucleus (PM) whereas (k) mENK-ir cells were observed in
PM. l: Overlay of histamine- and mENK-ir. n: GABA-ir cells in D. o: Overlay of histamine-ir and GABA-ir in D. Histamine-ir fibers made close
contact with GABA-ir neurons, but no colocalization was observed in this area. All images are maximum projection images. Arrows indicate
site of coexistence. D, dorsal telencephalon (pallium); PM, magnocellular preoptic nucleus; V, ventral telencephalon (subpallium). Anterior
to left. Scale bars ¼ 50 lm in a–o; 10 lm in inlays.

able in the whole-mount preparation to visualize the
intact histaminergic, dopaminergic, and serotonergic neu-
ron clusters. This approach can prove to be useful for
analysis of phenotypes related to, e.g., disease mecha-
nisms, because many human disease gene mutant fish
will soon be available (Bandmann and Burton, 2010).
General organization of galanin, GABA,mENK, and TRH systems in vertebrates
In rats, the galanin system is widely distributed in the
brain, including the hypothalamus (Skofitsch and Jacobo-
witz, 1985; Melander et al., 1986, 1988) and involved in
many physiological functions from energy metabolism,
sleep regulation, to learning and memory (Mitsukawa
et al., 2008). Galanin signals through a total of three G-
protein-coupled receptors (GPCRs), GALR1, GALR2, and
GALR3, which are all distributed in different brain areas
including the hypothalamus (Branchek et al., 2000). There
is so far one descriptive study regarding the general orga-
nization of the galaninergic system in zebrafish (Castro
et al., 2006) reporting that a small number of galaninergic
neurons are found in the dorsal periventricular hypothala-
mus. We found that galanin-ir fibers surround the histami-
nergic neurons in the ventrocaudal hypothalamus, which
is in accordance with studies on rats (Skofitsch and Jaco-
bowitz, 1985; Melander et al., 1988). We also found
scarce histamine-ir fibers in the ventral telencephalon
that were positive for galanin. Double-positive fibers lend
further support to the concept that part of the histaminer-
gic neurons in zebrafish also use galanin as a transmitter.
The galanin-like peptide in rats is expressed in histami-
nergic, catecholaminergic, and GABAergic neurons (Mel-
ander et al., 1986). In cultured histaminergic tuberomam-
millary neurons from fetal rat, galanin can also be
detected (Kukko-Lukjanov and Panula, 2003). Distribu-
tion of galanin immunoreactivity, the development of the
system, and sexual dimorphism has been studied earlier
in brown trout (Salmo trutta fario) (Rodrıguez et al., 2003)
and goldfish (Carassius auratus) (Prasada Rao et al.,
1996). The ventral telencephalon and the hypothalamus
are areas that are highly galanin-ir in these species and in
zebrafish. A similar galanin-ir distribution pattern has also
been reported for Siberian sturgeon (Acipenser baeri)
(Adrıo et al., 2005).
TRH neurons are found in the hypothalamus and me-
dulla oblongata of rats (Johansson and H€okfelt, 1980) and
these neurons send widespread projections to the entire
brain (H€okfelt et al., 1975). In zebrafish the TRH system
has been characterized, with TRH-positive neurons and
fibers found throughout the brain in telencephalon, dien-
cephalon, mesencephalon, and rhombencephalon (Diaz
et al., 2002), and the overall structure of the systems in
the two species is quite similar. TRH mediates its function
via activation of GPCRs TRHR1 and TRHR2 (Gershengorn
and Osman, 1996; Itadani et al., 1998; O’Dowd et al.,
2000). TRH is classically associated with hypothalamic–
pituitary–adrenal axis-associated endocrine function, but
as TRH and TRHRs are widespread in the brain, it is likely
that TRH mediates many physiological functions of which
not all are yet known. TRH-ir in two salmon species (Matz
and Takahashi, 1994; Diaz et al., 2001) show a similar
distribution pattern as observed for zebrafish. In these
three studied cases TRH-ir fibers can be observed at high
density in the same anatomical structures: the ventral tel-
encephalon and the ventrocaudal hypothalamus. In the
ventral telencephalon we observed histamine-ir fibers
that were also TRH-ir-positive. This finding further sup-
ports our colocalization result that indicates that some
histaminergic neurons also include TRH.
Endogenous opioid peptides enkephalins (methionine-
and leucine-enkephalin) are formed from a common pro-
enkephalin gene (Noda et al., 1982). mENK is an inhibi-
tory neuropeptide and growth factor peptide (Zagon
et al., 2002), implicated in nociception (Westlund, 2009)
and a ligand for the delta opioid receptor (Zagon et al.,
2000). The rat brain has about 30 different clusters of en-
kephalin-positive neurons and therefore the fibers also in-
nervate most areas of the brain (Johansson and H€okfelt,
1980). The zebrafish proenkephalin has 40% similarity to
the mammalian gene and is mainly expressed in the brain
(Gonz�alez-Nu~nez et al., 2003), as is the zebrafish d opioid
receptor (Barrallo et al., 1998). In rainbow trout (Onco-
rhynchus mykiss) mENK-ir neurons are found in the ven-
tral telencephalon, the ventromedial thalamus, the mes-
encephalic tegmentum, and the cerebellum (Vecino et al.,
1992). We did not find any histaminergic neurons in the
ventrocaudal hypothalamus that were mENK-positive.
mENK-ir cell bodies are found in magnocellular preoptic
nucleus, which is in accordance with what has been
reported for dogfish (Scyliorhinus canicula) (Vallarino
et al., 1994). No histamine-ir input was observed in this
area, suggesting that histamine has no effect on mENK
neurons, whereas the opposite is more likely. The distri-
bution of mENK-ir fibers in the ventral telencephalon of
zebrafish resembles the pattern found previously in rain-
bow trout (Vecino et al., 1992) and West African lungfish
(Protopterus annectens) (Vallarino et al., 1998). We did
not observe any histamine-ir fibers positive for mENK-ir.
When compared to galanin, TRH, and mENK, the
GABAergic neurotransmitter system has been studied
extensively in zebrafish. Zebrafish have two forms of glu-
tamate decarboxylase (GAD) that convert glutamate to
GABA (Martin et al., 1998). These two isoforms are
expressed throughout the larval brain (Martin et al.,
1998). In the adult brain GABAergic neurons have gener-
ally an inhibitory role through GABA-A receptors. GABA-ir
3840 The Journal of Comparative Neurology |Research in Systems Neuroscience
Sundvik and Panula

neurons are found in several brain areas such as the ol-
factory bulb, telencephalon, optic tectum, and hypothala-
mus (Kim et al., 2004) and surround the Hc in larval
zebrafish brain (Mueller et al., 2006). We also found
GABAergic neurons throughout the brain and we identi-
fied dense GABAergic fiber projection terminals at hista-
minergic neurons. In the dorsal telencephalon, histamine-
ir fibers were correspondingly in close contact with the
GABAergic neurons. In lamprey (Mel�endez-Ferro et al.,
2002; Robertson et al., 2007), lungfish (Trabucchi et al.,
2008), and zebrafish (Mueller et al., 2006; Mueller and
Guo, 2009) the development and distribution of the
GABAergic neurons and system is similar and seems to
be conserved with the system development observed in
mice and rats.
Role of the interconnected systems onphysiological function, with focus on arousal
Neurons communicate by releasing neurotransmitters
through two different mechanisms: 1) by local release in
classical chemical synapses and 2) by 3D diffusion
according to the model called ‘‘volume transmission’’
through releasing neurotransmitters into the extracellular
space, where they diffuse toward the target cells and act
on the appropriate receptors (Agnati et al., 1995). Hista-
mine is released in the latter fashion and modulates the
basic level of brain activity in response to changes in the
environment (Freeman, 2005). As galanin, GABA, and
TRH all were expressed in the histaminergic neurons and
fibers containing these markers innervated the ventro-
caudal hypothalamus, corelease of these neurotrans-
mitters may modulate the response of histamine associ-
ated with, e.g., arousal.
Previous studies have shown that these neurotrans-
mitters not only modulate the histaminergic system, but
also the hcrt system. Galanin has been reported to have
an inhibitory effect on the release of histamine from rat
hypothalamus (Arrang et al., 1991) and an excitatory
effect on hcrt neurons in rat lateral hypothalamus
(Kageyama et al., 2006). Sleep-deprived rats express
increased galanin levels (Toppila et al., 1995). TRH has an
excitatory effect on both histaminergic (Parmentier et al.,
2009) and hcrt neurons (Gonz�alez et al., 2009), but the
effect of a TRH analog on waking behavior in mice is com-
pletely abolished in histidine decarboxylase knockout
mice lacking histamine (Parmentier et al., 2009). Further-
more, the symptoms of narcolepsy in a canine model are
alleviated after treatment with TRH analogs (Nishino
et al., 1997). These results suggest that the arousal-pro-
moting effect of TRH is mainly mediated through the his-
taminergic neurotransmitter system. mENK inhibits the
firing rate of hcrt neurons (Li and van den Pol, 2008) and
thus has a sleep-promoting effect. As mENK was widely
distributed and the fibers containing mENK innervated
densely histaminergic neurons in zebrafish, it is likely that
mENK significantly affects the activity of histaminergic
neurons. This has, however, not yet been reported. The
main inhibitory neurotransmitter GABA and galanin are
colocalized in histaminergic neurons in rat (Kohler et al.,
1986; Eriksson et al., 2001; Kukko-Lukjanov and Panula,
2003). Histaminergic neurons in mammals are regulated
by GABAergic and galanin-containing neurons from the
preoptic area (Sherin et al., 1998). It has been proposed
that together these two inhibitory neurotransmitter sys-
tems form the sleep-promoting part of the sleep/wake
cycle (McGinty and Szymusiak, 2003). Thereby, GABA
and galanin target and silence one of the initial compo-
nents of the AAS.
A study by Anaclet et al. (2009) utilized knockout
mouse models and found that histamine and hcrt are re-
sponsible for different aspects of arousal and wakeful-
ness. Histamine is suggested to be responsible for the
cognitive arousal/wakefulness, whereas hcrt is responsi-
ble for the locomotor-associated arousal (Anaclet et al.,
2009). The precise roles of histamine and hcrt in
arousal/wakefulness and their interaction are still
unclear. In zebrafish, the main focus has been on how the
hcrt system regulates arousal (Prober et al., 2006; Yoko-
gawa et al., 2007; Appelbaum et al., 2009, 2010),
whereas the role of histamine has not been extensively
studied. To date, few studies have assessed the role of
histaminergic ligands on activity (Peitsaro et al., 2007)
and sleep/wake regulation in zebrafish (Rihel et al.,
2010). Treatment of zebrafish larvae with the histamine
receptor 1 (hrh1) antagonist resulted in a behavioral phe-
notype exhibiting reduced locomotor activity (Peitsaro
et al., 2007). Similarly, treatment of zebrafish larvae with
hrh1 antagonist resulted in increased rest when the
sleep/wake cycle was observed in a large pharmacologi-
cal screen (Rihel et al., 2010).
We have furthermore shown that translation inhibition
of hdc with morpholino oligonucleotides (MO) decreases
brain histamine levels by about 70% in larval zebrafish
and leads to a significant decline in locomotor activity
during wakefulness (Sundvik et al., 2011). This procedure
also abolishes a dark-induced fast behavioral locomotor
response, i.e., dark-induced flash response, characteris-
tic of the larvae, suggesting involvement of histamine in
sensorimotor gating. Surprisingly, hdc MO treatment also
significantly downregulates orexin/hcrt mRNA expression
(Sundvik et al., 2011). This physiological significance of
the histaminergic system renders the structural and
organizational data presented here useful for further func-
tional studies. The present study further supports the use
of zebrafish as a tool organism to study the role of the
The Journal of Comparative Neurology | Research in Systems Neuroscience 3841
Organization of zebrafish histaminergic system

histaminergic system in wakefulness, as the comparative
analysis of the neurotransmitter phenotype of the hista-
minergic system shows no major differences to the rat
histaminergic system.
CONCLUSION
In this study we visualized the intact histaminergic neu-
ron population in an adult zebrafish brain and show that
about 40–45 histaminergic neurons are found in both
male and female brain. In zebrafish galanin, GABA, and
TRH were colocalized with histamine in a few neurons in
the ventrocaudal hypothalamus. We also found that fibers
containing galanin-, TRH-, or mENK-ir innervated the his-
taminergic neurons. Taken together, we show that the
histaminergic neurons in adult zebrafish are similar to the
histamine neurons in rats, as they express the same
neurotransmitters, although this is observed to varying
degrees. A similar comparative analysis of the cotransmit-
ters of histaminergic neurons and the locations of the sys-
tems with regard to each other in the target area of hista-
minergic fibers in telencephalon is not available for any
other teleost species. These results further support the
use of zebrafish in studies on regulation of wakefulness.
ACKNOWLEDGMENT
We thank Henri Koivula, BSc, and Susanna Norrbacka,
BSc, for excellent fish care.
LITERATURE CITEDAdrıo F, Rodrıguez MA, Rodrıguez-Moldes I. 2005. Distribution
of galanin-like immunoreactivity in the brain of the Siberiansturgeon (Acipenser baeri). J Comp Neurol 487:54–74.
Agnati LF, Zoli M, Str€omberg I, Fuxe K. 1995. Intercellularcommunication in the brain: wiring versus volume trans-mission. Neuroscience 69:711–726.
Airaksinen MS, Paetau A, Palj€arvi L, Reinikainen K, RiekkinenP, Suomalainen R, Panula P. 1991. Histamine neurons inhuman hypothalamus: anatomy in normal and Alzheimerdiseased brains. Neuroscience 44:465–481.
Airaksinen MS, Alanen S, Szabat E, Visser TJ, Panula P. 1992.Multiple neurotransmitters in the tuberomammillary nu-cleus: comparison of rat, mouse, and guinea pig. J CompNeurol 323:103–116.
Anaclet C, Parmentier R, Ouk K, Guidon G, Buda C, Sastre JP,Akaoka H, Sergeeva OA, Yanagisawa M, Ohtsu H, FrancoP, Haas HL, Lin JS. 2009. Orexin/hypocretin and hista-mine: distinct roles in the control of wakefulness demon-strated using knock-out mouse models. J Neurosci 29:14423–14438.
Anichtchik OV, Kaslin J, Peitsaro N, Scheinin M, Panula P.2004. Neurochemical and behavioural changes in zebrafishDanio rerio after systemic administration of 6-hydroxydopa-mine and 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine.J Neurochem 88:443–453.
Appelbaum L, Wang GX, Maro GS, Mori R, Tovin A, Marin W,Yokogawa T, Kawakami K, Smith SJ, Gothilf Y, Mignot E,Mourrain P. 2009. Sleep-wake regulation and hypocretin-melatonin interaction in zebrafish. Proc Natl Acad Sci U SA 106:21942–21947.
Appelbaum L, Wang G, Yokogawa T, Skariah GM, Smith SJ,Mourrain P, Mignot E. 2010. Circadian and homeostaticregulation of structural synaptic plasticity in hypocretinneurons. Neuron 68:87–98.
Arrang JM, Gulat-Marnay C, Defontaine N, Schwartz JC. 1991.Regulation of histamine release in rat hypothalamus andhippocampus by presynaptic galanin receptors. Peptides12:1113–1117.
Arvidsson U, Ulfhake B, Cullheim S, Shupliakov O, Brodin E,Franck J, Bennett GW, Fone KC, Visser TJ, H€okfelt T. 1992.Thyrotropin-releasing hormone (TRH)-like immunoreactivityin the grey monkey (Macaca fascicularis) spinal cord andmedulla oblongata with special emphasis on the bulbospi-nal tract. J Comp Neurol 322:293–310.
Bandmann O, Burton EA. 2010. Genetic zebrafish models ofneurodegenerative diseases. Neurobiol Dis 40:58–65.
Barrallo A, Gonz�alez-Sarmiento R, Porteros A, Garcia-IsidoroM, Rodriguez RE. 1998. Cloning, molecular characteriza-tion, and distribution of a gene homologous to delta opioidreceptor from zebrafish (Danio rerio). Biochem Biophys ResCommun 245:544–548.
Branchek TA, Smith KE, Gerald C, Walker MW. 2000. Galaninreceptor subtypes. Trends Pharmacol Sci 21:109–117.
Brodin L, H€okfelt T, Grillner S, Panula P. 1990. Distribution ofhistaminergic neurons in the brain of the lamprey Lampetrafluviatilis as revealed by histamine-immunohistochemistry.J Comp Neurol 292:435–442.
Burgess HA, Granato M. 2007a. Modulation of locomotor ac-tivity in larval zebrafish during light adaptation. J Exp Biol210:2526–2539.
Burgess HA, Granato M. 2007b. Sensorimotor gating in larvalzebrafish. J Neurosci 27:4984–4994.
Castro A, Becerra M, Manso MJ, Anad�on R. 2006. Calretininimmunoreactivity in the brain of the zebrafish, Danio rerio:distribution and comparison with some neuropeptides andneurotransmitter-synthesizing enzymes. I. Olfactory organand forebrain. J Comp Neurol 494:435–59.
Chen YC, Priyadarshini M, Panula P. 2009. Complementarydevelopmental expression of the two tyrosine hydroxylasetranscripts in zebrafish. Histochem Cell Biol 132:375–381.
Dere E, Zlomuzica A, De Souza Silva MA, Ruocco LA, SadileAG, Huston JP. 2010. Neuronal histamine and the interplayof memory, reinforcement and emotions. Behav Brain Res215:209–220.
Diaz ML, Becerra M, Manso MJ, Anad�on R. 2001. Develop-ment of thyrotropin-releasing hormone immunoreactivity inthe brain of the brown trout Salmo trutta fario. J CompNeurol 429:299–320.
Diaz ML, Becerra M, Manso MJ, Anad�on R. 2002. Distributionof thyrotropin-releasing hormone (TRH) immunoreactivity inthe brain of the zebrafish (Danio rerio). J Comp Neurol450:45–60.
Ekstr€om P, Holmqvist BI, Panula P. 1995. Histamine-immuno-reactive neurons in the brain of the teleost Gasterosteusaculeatus L. Correlation with hypothalamic tyrosine hydrox-ylase- and serotonin-immunoreactive neurons. J Chem Neu-roanat 8:75–85.
Eriksson KS, Peitsaro N, Karlstedt K, Kaslin J, Panula P. 1998.Development of the histaminergic neurons and expressionof histidine decarboxylase mRNA in the zebrafish brain inthe absence of all peripheral histaminergic systems. Eur JNeurosci 10:3799–3812.
Eriksson KS, Sergeeva O, Brown RE, Haas HL. 2001. Orexin/hypocretin excites the histaminergic neurons of the tubero-mammillary nucleus. J Neurosci 21:9273–9279.
Fliers E, Noppen NW, Wiersinga WM, Visser TJ, Swaab DF.1994. Distribution of thyrotropin-releasing hormone (TRH)-
3842 The Journal of Comparative Neurology |Research in Systems Neuroscience
Sundvik and Panula

containing cells and fibers in the human hypothalamus.J Comp Neurol 350:311–323.
Freeman WJ. 2005. Ndn, volume transmission, and self-organi-zation in brain dynamics. J Integr Neurosci 4:407–421.
Gershengorn MC, Osman R. 1996. Molecular and cellular biol-ogy of thyrotropin-releasing hormone receptors. PhysiolRev 76:175–191.
Gonz�alez-Nu~nez V, Gonz�alez Sarmiento R, Rodrıguez RE.2003. Characterization of zebrafish proenkephalin revealsnovel opioid sequences. Brain Res Mol Brain Res 114:31–39.
Gonz�alez-Nu~nez V, Arsequell G, Szemenyei E, Toth G, ValenciaG, Rodrıguez RE. 2005. Binding profile of the endogenousnovel heptapeptide Met-enkephalin-Gly-tyr in zebrafish andrat brain. J Pharmacol Exp Ther 314:862–867.
Gonz�alez JA, Horjales-Araujo E, Fugger L, Broberger C, Burda-kov D. 2009. Stimulation of orexin/hypocretin neurones bythyrotropin-releasing hormone. J Physiol 587:1179–1186.
Haas H, Panula P. 2003. The role of histamine and the tubero-mamillary nucleus in the nervous system. Nat Rev Neuro-sci 4:121–130.
Haas HL, Sergeeva OA, Selbach O. 2008. Histamine in thenervous system. Physiol Rev 88:1183–1241.
Hara J, Gerashchenko D, Wisor JP, Sakurai T, Xie X, Kilduff TS.2009. Thyrotropin-releasing hormone increases behavioralarousal through modulation of hypocretin/orexin neurons.J Neurosci 29:3705–3714.
H€okfelt T, Fuxe K, Johansson O, Jeffcoate S, White N. 1975.Distribution of thyrotropin-releasing hormone (TRH) in thecentral nervous system as revealed with immunohisto-chemistry. Eur J Pharmacol 34:389–392.
Inagaki N, Panula P, Yamatodani A, Wada H. 1991. Organiza-tion of the histaminergic system in the brain of the teleost,Trachurus trachurus. J Comp Neurol 310:94–102.
Itadani H, Nakamura T, Itoh J, Iwaasa H, Kanatani A, Borkow-ski J, Ihara M, Ohta M. 1998. Cloning and characterizationof a new subtype of thyrotropin-releasing hormone recep-tors. Biochem Biophys Res Commun 250:68–71.
Johansson O, H€okfelt T. 1980. Thyrotropin releasing hormone,somatostatin, and enkephalin: distribution studies usingimmunohistochemical techniques. J Histochem Cytochem28:364–366.
Kageyama H, Kita T, Toshinai K, Guan JL, Date Y, Takenoya F,Kato S, Matsumoto H, Ohtaki T, Nakazato M, Shioda S.2006. Galanin-like peptide promotes feeding behaviour viaactivation of orexinergic neurones in the rat lateral hypo-thalamus. J Neuroendocrinol 18:33–41.
Kanbayashi T, Kodama T, Kondo H, Satoh S, Inoue Y, Chiba S,Shimizu T, Nishino S. 2009. CSF histamine contents in nar-colepsy, idiopathic hypersomnia and obstructive sleepapnea syndrome. Sleep 32:181–187.
Karhunen T, Airaksinen MS, Tuomisto L, Panula P. 1993. Neu-rotransmitters in the nervous system of Macoma balthica(Bivalvia). J Comp Neurol 334:477–488.
Kaslin J, Panula P. 2001. Comparative anatomy of the histami-nergic and other aminergic systems in zebrafish (Daniorerio). J Comp Neurol 440:342–377.
Kaslin J, Nystedt JM, Ostergard M, Peitsaro N, Panula P.2004. The orexin/hypocretin system in zebrafish is con-nected to the aminergic and cholinergic systems. J Neuro-sci 24:2678–2689.
Kim YJ, Nam RH, Yoo YM, Lee CJ. 2004. Identification andfunctional evidence of GABAergic neurons in parts of thebrain of adult zebrafish (Danio rerio). Neurosci Lett 355:29–32.
Kohler C, Ericson H, Watanabe T, Polak J, Palay SL, Palay V,Chan-Palay V. 1986. Galanin immunoreactivity in hypo-thalamic neurons: further evidence for multiple chemical
messengers in the tuberomammillary nucleus. J CompNeurol 250:58–64.
Kukko-Lukjanov TK, Panula P. 2003. Subcellular distribution ofhistamine, GABA and galanin in tuberomamillary neuronsin vitro. J Chem Neuroanat 25:279–292.
Li Y, van den Pol AN. 2008. Mu-opioid receptor-mediateddepression of the hypothalamic hypocretin/orexin arousalsystem. J Neurosci 28:2814–2819.
Lillesaar C, Stigloher C, Tannhauser B, Wullimann MF, Bally-Cuif L. 2009. Axonal projections originating from rapheserotonergic neurons in the developing and adult zebrafish,Danio rerio, using transgenics to visualize raphe-specificpet1 expression. J Comp Neurol 512:158–182.
Lin L, Faraco J, Li R, Kadotani H, Rogers W, Lin X, Qiu X, deJong PJ, Nishino S, Mignot E. 1999. The sleep disorder ca-nine narcolepsy is caused by a mutation in the hypocretin(orexin) receptor 2 gene. Cell 98:365–376.
Lohr H, Ryu S, Driever W. 2009. Zebrafish diencephalic A11-related dopaminergic neurons share a conserved transcrip-tional network with neuroendocrine cell lineages. Develop-ment 136:1007–1017.
Magliulo-Cepriano L, Schreibman MP, Blum V. 1993. The dis-tribution of immunoreactive FMRF-amide, neurotensin, andgalanin in the brain and pituitary gland of three species ofXiphophorus from birth to sexual maturity. Gen CompEndocrinol 92:269–280.
Martin SC, Heinrich G, Sandell JH. 1998. Sequence andexpression of glutamic acid decarboxylase isoforms in thedeveloping zebrafish. J Comp Neurol 396:253–266.
Matz SP, Takahashi TT. 1994. Immunohistochemical localiza-tion of thyrotropin-releasing hormone in the brain of chi-nook salmon (Oncorhynchus tshawytscha). J Comp Neurol345:214–223.
McGinty D, Szymusiak R. 2003. Hypothalamic regulation ofsleep and arousal. Front Biosci 8:1074–1083.
McLean DL, Fetcho JR. 2004. Ontogeny and innervation pat-terns of dopaminergic, noradrenergic, and serotonergicneurons in larval zebrafish. J Comp Neurol 480:38–56.
Melander T, H€okfelt T, Rokaeus A, Cuello AC, Oertel WH, Ver-hofstad A, Goldstein M. 1986. Coexistence of galanin-likeimmunoreactivity with catecholamines, 5-hydroxytrypta-mine, GABA and neuropeptides in the rat CNS. J Neurosci6:3640–3654.
Melander T, Kohler C, Nilsson S, H€okfelt T, Brodin E, Theo-dorsson E, Bartfai T. 1988. Autoradiographic quantitationand anatomical mapping of 125I-galanin binding sites inthe rat central nervous system. J Chem Neuroanat 1:213–233.
Mel�endez-Ferro M, Perez-Costas E, Villar-Cheda B, Abalo XM,Rodriguez-Munoz R, Rodicio MC, Anad�on R. 2002. Ontog-eny of gamma-aminobutyric acid-immunoreactive neuronalpopulations in the forebrain and midbrain of the sea lam-prey. J Comp Neurol 446:360–376.
Mitsukawa K, Lu X, Bartfai T. 2008. Galanin, galanin receptorsand drug targets. Cell Mol Life Sci 65:1796–1805.
Mueller T, Guo S. 2009. The distribution of GAD67-mRNA inthe adult zebrafish (teleost) forebrain reveals a prosomericpattern and suggests previously unidentified homologies totetrapods. J Comp Neurol 516:553–568.
Mueller T, Vernier P, Wullimann MF. 2004. The adult centralnervous cholinergic system of a neurogenetic model ani-mal, the zebrafish Danio rerio. Brain Res 1011:156–169.
Mueller T, Vernier P, Wullimann MF. 2006. A phylotypic stagein vertebrate brain development: GABA cell patterns inzebrafish compared with mouse. J Comp Neurol 494:620–634.
Nishino S, Arrigoni J, Shelton J, Kanbayashi T, Dement WC,Mignot E. 1997. Effects of thyrotropin-releasing hormone
The Journal of Comparative Neurology | Research in Systems Neuroscience 3843
Organization of zebrafish histaminergic system

and its analogs on daytime sleepiness and cataplexy in ca-nine narcolepsy. J Neurosci 17:6401–6408.
Nishino S, Sakurai E, Nevsimalova S, Yoshida Y, Watanabe T,Yanai K, Mignot E. 2009. Decreased CSF histamine in nar-colepsy with and without low CSF hypocretin-1 in compari-son to healthy controls. Sleep 32:175–180.
Noda M, Teranishi Y, Takahashi H, Toyosato M, Notake M,Nakanishi S, Numa S. 1982. Isolation and structural orga-nization of the human preproenkephalin gene. Nature 297:431–434.
O’Dowd BF, Lee DK, Huang W, Nguyen T, Cheng R, Liu Y,Wang B, Gershengorn MC, George SR. 2000. TRH-R2exhibits similar binding and acute signaling but distinctregulation and anatomic distribution compared with TRH-R1. Mol Endocrinol 14:183–193.
Panula P, Yang HY, Costa E. 1984. Histamine-containing neu-rons in the rat hypothalamus. Proc Natl Acad Sci U S A81:2572–2576.
Panula P, Airaksinen MS, Pirvola U, Kotilainen E. 1990. A his-tamine-containing neuronal system in human brain. Neuro-science 34:127–132.
Panula P, Chen YC, Priyadarshini M, Kudo S, Semenova S,Sundvik M, Sallinen V. 2010. The comparative neuroanat-omy and neurochemistry of zebrafish CNS systems of rele-vance to human neuropsychiatric diseases. Neurobiol Dis40:46–57.
Parmentier R, Kolbaev S, Klyuch BP, Vandael D, Lin JS, Sel-bach O, Haas HL, Sergeeva OA. 2009. Excitation of hista-minergic tuberomamillary neurons by thyrotropin-releasinghormone. J Neurosci 29:4471–4483.
Peitsaro N, Kaslin J, Anichtchik OV, Panula P. 2003. Modula-tion of the histaminergic system and behaviour by alpha-fluoromethylhistidine in zebrafish. J Neurochem 86:432–441.
Peitsaro N, Sundvik M, Anichtchik OV, Kaslin J, Panula P.2007. Identification of zebrafish histamine H1, H2 and H3receptors and effects of histaminergic ligands on behavior.Biochem Pharmacol 73:1205–1214.
Peyron C, Faraco J, Rogers W, Ripley B, Overeem S, CharnayY, Nevsimalova S, Aldrich M, Reynolds D, Albin R, Li R,Hungs M, Pedrazzoli M, Padigaru M, Kucherlapati M, Fan J,Maki R, Lammers GJ, Bouras C, Kucherlapati R, Nishino S,Mignot E. 2000. A mutation in a case of early onset narco-lepsy and a generalized absence of hypocretin peptides inhuman narcoleptic brains. Nat Med 6:991–997.
Podlasz P, Sallinen V, Chen Y-C, Kudo H, Fedorowska N, Pan-ula P. 2012. Galanin gene expression and effects of itsknock-down on the developoment of the nervous systemin larval zebrafish. J Comp Neurol 520:3846–3862.
Prasada Rao PD, Murthy CK, Cook H, Peter RE. 1996. Sexualdimorphism of galanin-like immunoreactivity in the brainand pituitary of goldfish, Carassius auratus. J Chem Neuro-anat 10:119–135.
Prober DA, Rihel J, Onah AA, Sung RJ, Schier AF. 2006. Hypo-cretin/orexin overexpression induces an insomnia-like phe-notype in zebrafish. J Neurosci 26:13400–13410.
R�ethelyi M, Mohapatra NK, Metz CB, Petrusz P, Lund PK.1991. Colchicine enhances mRNAs encoding the precursorof calcitonin gene-related peptide in brainstem motoneur-ons. Neuroscience 42:531–539.
Rihel J, Prober DA, Arvanites A, Lam K, Zimmerman S, Jang S,Haggarty SJ, Kokel D, Rubin LL, Peterson RT, Schier AF.2010. Zebrafish behavioral profiling links drugs to biologi-cal targets and rest/wake regulation. Science 327:348–351.
Robertson B, Auclair F, Menard A, Grillner S, Dubuc R. 2007.GABA distribution in lamprey is phylogenetically conserved.J Comp Neurol 503:47–63.
Rodrıguez MA, Anad�on R, Rodrıguez-Moldes I. 2003. Develop-ment of galanin-like immunoreactivity in the brain of thebrown trout (Salmo trutta fario), with some observations onsexual dimorphism. J Comp Neurol 465:263–285.
Sager JJ, Bai Q, Burton EA. 2010. Transgenic zebrafish modelsof neurodegenerative diseases. Brain Struct Funct 214:285–302.
Sallinen V, Torkko V, Sundvik M, Reenila I, Khrustalyov D,Kaslin J, Panula P. 2009a. MPTP and MPPþ target specificaminergic cell populations in larval zebrafish. J Neurochem108:719–731.
Sallinen V, Sundvik M, Reenila I, Peitsaro N, Khrustalyov D,Anichtchik O, Toleikyte G, Kaslin J, Panula P. 2009b.Hyperserotonergic phenotype after monoamine oxidase in-hibition in larval zebrafish. J Neurochem 109:403–415.
Sallinen V, Kolehmainen J, Priyadarshini M, Toleikyte G, ChenYC, Panula P. 2010. Dopaminergic cell damage and vulner-ability to MPTP in Pink1 knockdown zebrafish. NeurobiolDis 40:93–101.
Sherin JE, Elmquist JK, Torrealba F, Saper CB. 1998. Innerva-tion of histaminergic tuberomammillary neurons byGABAergic and galaninergic neurons in the ventrolateralpreoptic nucleus of the rat. J Neurosci 18:4705–4721.
Skofitsch G, Jacobowitz DM. 1985. Immunohistochemicalmapping of galanin-like neurons in the rat central nervoussystem. Peptides 6:509–546.
Sundvik M, Kudo H, Toivonen P, Rozov S, Chen YC, Panula P.2011. The histaminergic system regulates wakefulness andorexin/hypocretin neuron development via histamine re-ceptor H1 in zebrafish. FASEB J 25:1–10.
Szymusiak R, McGinty D. 2008. Hypothalamic regulation ofsleep and arousal. Ann N Y Acad Sci 1129:275–286.
Szymusiak R, Gvilia I, McGinty D. 2007. Hypothalamic controlof sleep. Sleep Med 8:291–301.
Toppila J, Stenberg D, Alanko L, Asikainen M, Urban JH, TurekFW, Porkka-Heiskanen T. 1995. REM sleep deprivationinduces galanin gene expression in the rat brain. NeurosciLett 183:171–174.
Trabucchi M, Trudeau VL, Drouin G, Tostivint H, Ihrmann I,Vallarino M, Vaudry H. 2008. Molecular characterizationand comparative localization of the mRNAs encoding twoglutamic acid decarboxylases (GAD65 and GAD67) in thebrain of the African lungfish, Protopterus annectens.J Comp Neurol 506:979–988.
Trottier S, Chotard C, Traiffort E, Unmehopa U, Fisser B,Swaab DF, Schwartz JC. 2002. Co-localization of histaminewith GABA but not with galanin in the human tuberomamil-lary nucleus. Brain Res 939:52–64.
Tsuruo Y, H€okfelt T, Visser TJ, Kimmel JR, Brown JC, Verhof-stadt A, Walsh J. 1988a. TRH-like immunoreactivity in en-docrine cells and neurons in the gastro-intestinal tract ofthe rat and guinea pig. Cell Tissue Res 253:347–356.
Tsuruo Y, Ceccatelli S, Villar MJ, H€okfelt T, Visser TJ, TereniusL, Goldstein M, Brown JC, Buchan A, Walsh J. 1988b.Coexistence of TRH with other neuroactive substances inthe rat central nervous system. J Chem Neuroanat 1:235–253.
Tsuruo Y, H€okfelt T, Visser TJ. 1988c. Thyrotropin-releasinghormone (TRH)-immunoreactive neuron populations in therat olfactory bulb. Brain Res 447:183–187.
Valdes JL, Farias P, Ocampo-Garces A, Cortes N, Seron-FerreM, Torrealba F. 2005. Arousal and differential Fos expres-sion in histaminergic neurons of the ascending arousal sys-tem during a feeding-related motivated behaviour. Eur JNeurosci 21:1931–1942.
Vallarino M, Bucharles C, Facchinetti F, Vaudry H. 1994.Immunocytochemical evidence for the presence of Met-en-kephalin and Leu-enkephalin in distinct neurons in the
3844 The Journal of Comparative Neurology |Research in Systems Neuroscience
Sundvik and Panula

brain of the elasmobranch fish Scyliorhinus canicula.J Comp Neurol 347:585–597.
Vallarino M, Thoumas JL, Masini MA, Trabucchi M, Chartrel N,Vaudry H. 1998. Immunocytochemical localization of enke-phalins in the brain of the African lungfish, Protopterusannectens, provides evidence for differential distribution ofMet-enkephalin and Leu-enkephalin. J Comp Neurol 396:275–287.
Vecino E, Pinuela C, Arevalo R, Lara J, Alonso JR, Aijon J.1992. Distribution of enkephalin-like immunoreactivity inthe central nervous system of the rainbow trout: an immu-nocytochemical study. J Anat 180:435–453.
Visser TJ, Klootwijk W. 1981. Approaches to a markedlyincreased sensitivity of the radioimmunoassay for thyrotro-pin-releasing hormone by derivatization. Biochim BiophysActa 673:454–466.
Visser TJ, Klootwijk W, Docter R, Hennemann G. 1977. A newradioimmunoassay of thyrotropin-releasing hormone. FEBSLett 83:37–40.
Watanabe T, Taguchi Y, Shiosaka S, Tanaka J, Kubota H, Ter-ano Y, Tohyama M, Wada H. 1984. Distribution of the his-taminergic neuron system in the central nervous system of
rats; a fluorescent immunohistochemical analysis with his-tidine decarboxylase as a marker. Brain Res 295:13–25.
Westlund KN. 2009. Gene therapy for pancreatitis pain. GeneTher 16:483–492.
Yang HY, Costa E. 1979. Estimating ‘‘in vivo’’ the turnoverrate of brain enkephalins. Adv Biochem Psychopharmacol20:131–142.
Yang HY, Hong JS, Costa E. 1977. Regional distribution ofLEU and MET enkephalin in rat brain. Neuropharmacology16:303–307.
Yokogawa T, Marin W, Faraco J, Pezeron G, Appelbaum L,Zhang J, Rosa F, Mourrain P, Mignot E. 2007. Characteriza-tion of sleep in zebrafish and insomnia in hypocretin re-ceptor mutants. PLoS Biol 5:e277.
Zagon IS, Verderame MF, Allen SS, McLaughlin PJ. 2000.Cloning, sequencing, chromosomal location, and functionof cDNAs encoding an opioid growth factor receptor(OGFr) in humans. Brain Res 856:75–83.
Zagon IS, Verderame MF, McLaughlin PJ. 2002. The biology ofthe opioid growth factor receptor (OGFr). Brain Res BrainRes Rev 38:351–376.
The Journal of Comparative Neurology | Research in Systems Neuroscience 3845
Organization of zebrafish histaminergic system
Copyright © 2022 FDOKUMEN