
Cardiovascular and respiratory developmental plasticity under oxygen depleted environment and in...
10
Cardiovascular and respiratory developmental plasticity under oxygen depleted environment and in genetically hypoxic zebrafish (Danio rerio) Nadeem Yaqoob, Thorsten Schwerte ⁎ University of Innsbruck, Institute of Zoology, Technikerstrasse 25, A-6020 Innsbruck, Austria abstract article info Article history: Received 7 December 2009 Received in revised form 25 March 2010 Accepted 28 March 2010 Available online 2 April 2010 Keywords: Hypoxia Anoxia Zebrafish Viability Cardiovascular system Hyperventilation Known vertebrate response to low oxygen concentration include change in carbohydrate metabolism, increase in nitric oxide, stimulation of red blood cell and hemoglobin production and induction of gene expression for glycolytic enzymes and hormones. Also, extreme hypoxia plays main role in pathological studies of cardiac dysfunction. The morphological and physiological developmental studies of the cardiovascular system under low oxygen are important as it is directly related to oxygen supply and consumption. Furthermore, cardiac function demands high energy during system development and thus it is most likely to be affected by hypoxia. Zebrafish (Danio rerio) can act as a model organism for oxygen demand management study as in natural environment, due to ecological disturbances, it is exposed to changes in oxygen concentrations routinely and thus would have natural ability to cope with it for survival. We have studied, in zebrafish, i) cardiovascular flexibility under extreme hypoxia (PO 2 = 20 Torr, 3 kPa) at 3–10 dpf (days post-fertilization), ii) cardiac re-animation in normoxia (PO 2 = 152 Torr, 20 kPa) after 90 min of anoxia (PO 2 = 0 Torr, 0 kPa)-induced suspended animation at 4 dpf and iii) oxygen consumption in 8 dpf von Hippel–Lindau (vhl −/− ) mutant that exhibits an artificial hypoxic response under normoxic conditions. In hypoxic fish, cardiac output, stroke volume and end-diastolic volume were elevated while intersegmental blood vessels vascularization index at 6 dpf and at 10 dpf was 22% and 11% higher respectively as compared to the normoxic fish. The heart rate in hypoxic fish was lower until 6 dpf and then showed an elevated trend. There was no significant difference in body length between the hypoxic and normoxic individuals. The observed changes may have enhanced the performance of the cardiovascular system for oxygen uptake. We also report for the first time that the post-anoxia re-animated heart rate returns to normal after 48 h. Measurement of oxygen consumption in 8 dpf hyperventilating vhl −/− mutant was, unexpectedly, significantly lower than the non-mutant fish of the same age which point towards artificial hypoxic signal from brain in these mutants. © 2010 Elsevier Inc. All rights reserved. 1. Introduction Danio rerio, commonly known as zebrafish is nowadays considered as one of the most suitable model for higher vertebrate experimental embryology and extensive genetic analysis. The transparent embryo enables to visualize and quantify the cardiac contraction, blood flow and vessel size and its patterning (Driever et al., 1994; Schwerte and Pelster, 2000; Patton and Zon, 2001; Chico et al., 2008) and thus, its choice as a model for the study of cardiovascular diseases cannot be ignored. In nature, the common environmental stress faced by fish population is hypoxia (Bagatto, 2005) that may result in changes in genetic expressions leading to modification in the physiological responses as also demonstrated by Gracey et al. (2001). Alteration of various genes functions in zebrafish under low oxygen environment (hypoxia) has been suggested in different studies (Bosworth et al., 2005; Shang et al., 2006; Marques et al., 2008). It has been reported by Padilla and Roth (2001) that when normal oxygen (normoxia) is provided, its embryo can be recovered to normal state without any deleterious effect even after 24 h exposure to zero oxygen (anoxia) conditions that resulted in suspended animation and nil heart beat. Since the cardiovascular system is most directly related to oxygen supply and consumption and itself demands high energy for system development and most likely to be affected by the low oxygen environment, studies have been carried out in the past to know about the performance pattern change in it under hypoxia. These studies were made either at moderate hypoxia with sampling time at shorter interval or at extreme hypoxia with sampling interval of longer period (Barrionuevo and Burggren, 1999; Grillitsch et al., 2005; Moore et al., 2006). Dissolved oxygen concentration of 1 mg L −1 (equivalent to 20 Torr or 3 kPa) has been denoted by Martinovic et al. (2009) as severe or extreme hypoxic for zebrafish and we have studied its embryonic and larval cardiac Comparative Biochemistry and Physiology, Part A 156 (2010) 475–484 ⁎ Corresponding author. University of Innsbruck, Institute of Zoology, Techniker- strasse 25, A-6020 Innsbruck, Austria. Tel.: + 43 512 507 6193; fax: + 43 512 507 2930. E-mail address: [email protected] (T. Schwerte). 1095-6433/$ – see front matter © 2010 Elsevier Inc. All rights reserved. doi:10.1016/j.cbpa.2010.03.033 Contents lists available at ScienceDirect Comparative Biochemistry and Physiology, Part A journal homepage: www.elsevier.com/locate/cbpa
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Cardiovascular and respiratory developmental plasticity under oxygen depleted environment and in...

Comparative Biochemistry and Physiology, Part A 156 (2010) 475–484
Contents lists available at ScienceDirect
Comparative Biochemistry and Physiology, Part A
j ourna l homepage: www.e lsev ie r.com/ locate /cbpa
Cardiovascular and respiratory developmental plasticity under oxygen depletedenvironment and in genetically hypoxic zebrafish (Danio rerio)
Nadeem Yaqoob, Thorsten Schwerte ⁎University of Innsbruck, Institute of Zoology, Technikerstrasse 25, A-6020 Innsbruck, Austria
⁎ Corresponding author. University of Innsbruck, Instrasse 25, A-6020 Innsbruck, Austria. Tel.: +43 512 507
E-mail address: [email protected] (T. Sch
1095-6433/$ – see front matter © 2010 Elsevier Inc. Aldoi:10.1016/j.cbpa.2010.03.033
a b s t r a c t
a r t i c l e i n f oArticle history:Received 7 December 2009Received in revised form 25 March 2010Accepted 28 March 2010Available online 2 April 2010
Keywords:HypoxiaAnoxiaZebrafishViabilityCardiovascular systemHyperventilation
Known vertebrate response to low oxygen concentration include change in carbohydrate metabolism,increase in nitric oxide, stimulation of red blood cell and hemoglobin production and induction of geneexpression for glycolytic enzymes and hormones. Also, extreme hypoxia plays main role in pathologicalstudies of cardiac dysfunction. The morphological and physiological developmental studies of thecardiovascular system under low oxygen are important as it is directly related to oxygen supply andconsumption. Furthermore, cardiac function demands high energy during system development and thus it ismost likely to be affected by hypoxia. Zebrafish (Danio rerio) can act as a model organism for oxygen demandmanagement study as in natural environment, due to ecological disturbances, it is exposed to changes inoxygen concentrations routinely and thus would have natural ability to cope with it for survival. We havestudied, in zebrafish, i) cardiovascular flexibility under extreme hypoxia (PO2=20 Torr, 3 kPa) at 3–10 dpf(days post-fertilization), ii) cardiac re-animation in normoxia (PO2=152 Torr, 20 kPa) after 90 min ofanoxia (PO2=0 Torr, 0 kPa)-induced suspended animation at 4 dpf and iii) oxygen consumption in 8 dpf vonHippel–Lindau (vhl−/−) mutant that exhibits an artificial hypoxic response under normoxic conditions. Inhypoxic fish, cardiac output, stroke volume and end-diastolic volume were elevated while intersegmentalblood vessels vascularization index at 6 dpf and at 10 dpf was 22% and 11% higher respectively as comparedto the normoxic fish. The heart rate in hypoxic fish was lower until 6 dpf and then showed an elevated trend.There was no significant difference in body length between the hypoxic and normoxic individuals. Theobserved changes may have enhanced the performance of the cardiovascular system for oxygen uptake. Wealso report for the first time that the post-anoxia re-animated heart rate returns to normal after 48 h.Measurement of oxygen consumption in 8 dpf hyperventilating vhl−/− mutant was, unexpectedly,significantly lower than the non-mutant fish of the same age which point towards artificial hypoxic signalfrom brain in these mutants.
stitute of Zoology, Techniker-6193; fax: +43 512 507 2930.werte).
l rights reserved.
© 2010 Elsevier Inc. All rights reserved.
1. Introduction
Danio rerio, commonly known as zebrafish is nowadays consideredas one of the most suitable model for higher vertebrate experimentalembryology and extensive genetic analysis. The transparent embryoenables to visualize and quantify the cardiac contraction, blood flowand vessel size and its patterning (Driever et al., 1994; Schwerte andPelster, 2000; Patton and Zon, 2001; Chico et al., 2008) and thus, itschoice as a model for the study of cardiovascular diseases cannot beignored. In nature, the common environmental stress faced by fishpopulation is hypoxia (Bagatto, 2005) that may result in changes ingenetic expressions leading to modification in the physiologicalresponses as also demonstrated by Gracey et al. (2001). Alteration of
various genes functions in zebrafish under low oxygen environment(hypoxia)has been suggested indifferent studies (Bosworthet al., 2005;Shang et al., 2006; Marques et al., 2008). It has been reported by Padillaand Roth (2001) that when normal oxygen (normoxia) is provided, itsembryo can be recovered to normal state without any deleterious effecteven after 24 h exposure to zero oxygen (anoxia) conditions thatresulted in suspended animation and nil heart beat. Since thecardiovascular system is most directly related to oxygen supply andconsumption and itself demands high energy for system developmentand most likely to be affected by the low oxygen environment, studieshave been carried out in the past to know about the performancepattern change in it under hypoxia. These studies were made either atmoderate hypoxia with sampling time at shorter interval or at extremehypoxia with sampling interval of longer period (Barrionuevo andBurggren, 1999; Grillitsch et al., 2005; Moore et al., 2006). Dissolvedoxygen concentration of 1 mg L−1 (equivalent to 20 Torr or 3 kPa) hasbeen denoted by Martinovic et al. (2009) as severe or extreme hypoxicfor zebrafish and we have studied its embryonic and larval cardiac

476 N. Yaqoob, T. Schwerte / Comparative Biochemistry and Physiology, Part A 156 (2010) 475–484
performance and vasculogenesis under this environment utilizing non-invasive digital imaging techniques.
Suspended animation or the temporary suspension of life is anintriguing concept and efforts have been carried out in the past tobetter understand this behavior. In mice, it has been recorded thatsuspended animation-like state developed by 20 min pretreatmentwith H2S enhanced the capacity to survive for 6.5 h with no apparentdetrimental effects at 5% oxygen which is otherwise lethal andmortality takes place within 20 min. Furthermore, this pretreatmentwith H2S, if followed by 1 h of 5% oxygen, lead to several hourssurvival time under oxygen tension as low as 3% (Blackstone and Roth,2007). The importance of nitric oxide (NO) under extreme hypoxicconditions has been emphasized by Teodoro and O'Farrell (2003) andrecorded that it induces a reversible arrest of development, geneexpression and turnover. Some studies were targeted to define theeffect of hypoxia and anoxia on protein synthesis and it was foundthat hypoxia-tolerant cells or tissues experienced more than 90%decline in protein synthesis and a decline in membrane permeability,often the major energy-consuming processes in cells. This greatlyreduces ATP consumption, which is a crucial step for long-termhypoxia survival (Hochachka et al., 1996; Hochachka and Lutz, 2001).Interestingly, Padilla et al. (2002) have pointed out that hypoxia-inducible factors HIF-1 is important to act as mediator in response tohypoxia but not to anoxia. In Caenorhabditis elegans embryos, san-1(suspended animation-1) acts as spindle checkpoint component andits absence or a second spindle checkpoint component MDF-2 duringanoxia-induced suspended animation, fail to arrest the cell cycle andexhibit chromosome mis-seggregation and reduced viability. Thismeans that in checkpoint-deficient embryos, mitosis is still completedafter oxygen deprivation, although accompanied by massive chromo-some mis-seggregation (Nystul et al., 2003; Pandey et al., 2007).Zebrafish embryos display complete arrest of the heart beat,movement, cell cycle progression, and developmental progressionwhen exposed to anoxia (Padilla and Roth, 2001).We have carried outstudy related to post-anoxic heart beat revival in this animal. The4 dpf old zebrafish was selected for this purpose because, at thisdevelopmental stage, the animal can survive on its embryonic yolkand external feeding is not desired while the heart has been fullydeveloped with functional valves (Stainier and Fishman, 1994).Anoxia exposure of 90 min was used for our experiments as ourshort preliminary studies have supported the fact that shorter anoxiaduration failed to suspend zebrafish heart while the longer timeperiod resulted in mortality rate elevation (unpublished data).
Hyperventilation or over-breathing is the state of breathing fasterand/or deeper than necessary. In zebrafish, it is observed as behavioralresponse to hypoxia indicated by increase in frequency of buccal oropercular movements (Jonz and Nurse, 2005; Turesson et al., 2006). Inseparate studies, Vulesevic et al. (2006) differentiated this behavior inhypercapnic animal as elevated breathing amplitude rather thanbreathing frequency. Under normoxic conditions, this hyperventila-tory phenotype is displayed by zebrafish larvae that are homozygousmutant (vhl−/−) for the von Hippel–Lindau gene (van Rooijen et al.,2009) and hence termed “genetically hypoxic”. In non-mutants, thisgene encodes a tumor suppressor protein that plays a critical role inangiogenesis and oxygen homeostasis through HIF-1α. The vhlmutated phenotype is inherited in an autosomal dominant manner.The homozygous mutant (vhl−/−) embryo selected for our study wasobtained by inbreeding heterozygous (vhl+/−) adult zebrafishcontaining germline activated mutation of the ortholog vhl1,generated and generously provided to us by Ellen van Rooijen(Hubrecht Institute, Netherlands). As recorded by van Rooijen et al.(2009), these homozygous mutants (vhl−/−) display severe hyper-ventilation (at 4–5 dpf) and significantly enlarged heart (at 7 dpf) andthe animals do not survive for more than 8–11 dpf due to severeedema in the cardiac region. In addition, thesemutants show systemichypoxic response, up-regulation of hypoxia-induced genes and
polycythemia with concomitantly increased Epo/EpoR mRNA levelsand Epo signaling. As found out by Soliz et al. (2005), Epo controlsventilation at the central (brainstem) and peripheral (carotid body)levels so it was of interest to us to find out if the hyperventilationshown by vhl−/−mutant zebrafish under normoxic condition is due tothe influence of Epo over-expression or systemic increase in oxygenconsumption. For this purpose, we carried out the respirometric studiesof vhl−/− mutants and compared it with that of the non-mutants.
2. Material and methods
2.1. Cardiac functions and vascular bed formation under extremehypoxia
From the aquarium facility in Zoology department, InnsbruckUniversity, wild type (wdt) zebrafish eggs were obtained and kept insmall plastic beakers (9×6×6 cm)with two lateralmeshes and hung inan aquarium at 28 °C at 14:10 h light–dark conditions under normoxiccondition (PO2=152 Torr, 20 kPa). Nearly 100 eggswere placed in eachbeaker. For the hypoxic treatment (PO2=20 Torr, 3 kPa ), the beakerswith 24 hpf (hours post-fertilization) old eggs were hung in a locallymanufactured 30×20×20 cm glass chamber containing about 7.5 L tapwater and tightly closed with removable glass lid. The chamber wasequipped with Clark oxygen electrode (POS; Polarographic OxygenSensor) and temperature sensor that were connected to a twin-flowrespirometer control unit (Cyclobios, Innsbruck, Austria, www.cyclo-bios.at). Nitrogen was supplied to the aquarium chamber in order toreplace the oxygen and the control unit automatically switched it offwhen the desired oxygen percentage level was reached. After 5 dpf, thelarvae were fed with Zmsystems Advanced Fry Feed ZM-000 (ZM Ltd)twice a day. Images of 10 hypoxic and 5 normoxic fishwere prepared asdescribed by Kopp et al. (2007). From 3 to 10 dpf, zebrafish wereanaesthetised with 100 mg L−1, ph-7.00 adjusted MS-222 (tricainemethanosulphonate, Sigma®) for 2 min. These were then embedded inlateral position in small microscopic chambers filled with 1.5% agarose(SeaPlaque®, melting temp.≤65 °C, gelling temp. 26°–30 °C) preparedin the same concentration of anaesthetic. The images were made withan inverted microscope (Zeiss Axiovert 25, Zeiss Vienna, Austria) thatwas linked to a digital high-speed video camera (Basler A504k, BaslerAhrensburg, Germany) connected to a frame grabber card (DATACUBEMaxRevolution) installed in a common PC System (P4, 2.4 GHz). VLLToolbox and CCT+ softwares (Basler Ahrensburg, Germany) for cameraconfiguration were used and the recording of sequences of digitalpictures was managed by Video Savant 4.0 (IO Industries Ontario,Canada).
Heart image analysis: for the heart video, 300 images at the rate of250 frames per second with exposure time of 3997 μs were recordedusing a 40-foldmagnification. Analyses of the images were carried outby software package Optimas 6.5 (Media Cybernetics Silversprings,USA). The time interval between three heart beats was measured andthe heart rate (bpm) was calculated in triplicate. The perimeter of theventricle image was outlined for the end-diastolic (EDV) and end-systolic volumes (ESV). Measurement analysis was carried out by “fit-to-ellipse” algorithm, which worked on calculating the center ofmass and subsequently the best fitting ellipse. The major and minoraxes were determined in this way and the data was extracted anddirectly transferred into a Microsoft Excel worksheet. Formula for aprolate spheroid, V=4/3×πab2 was applied to determine the strokevolume, where a and b represent half of the long and short axesmeasured respectively, of the ventricle. The mean stroke volumewas calculated as the difference between three ventricular EDV andESV. Cardiac output was obtained by multiplying the heart rate withstroke volume.
Vascular bed visualization and vascularization index determination:for the vascular system images, 100 images at the rate of 50 frames persecond with exposure time of 19997 μs were recorded using 10-fold

477N. Yaqoob, T. Schwerte / Comparative Biochemistry and Physiology, Part A 156 (2010) 475–484
magnification. Blood vessels network was visualized by accumulationof the shifting vectors of moving erythrocytes from a number ofsubsequent difference pictures as suggested by Schwerte and Pelster(2000). It worked on the principle that the subtraction of the twofields of a video frame can visualize any movement which occurredwithin the 20 ms time period. Directmeasurement for the velocity of amoving particle, in this case an erythrocyte, in the vascular system canbe generated by the subtraction of the length of the shifting vectors. Acomplete trace of the erythrocyte route was extracted by accumulat-ing several subsequent video frames shifting vectors. In our study,nearly 400–500 subsequent images were accumulated to obtain acomplete cast of the vascular bed of both normoxic and hypoxic fishfrom head to tail region (Fig 3B). These images were processed andjoined together by ImagePro® and CorelDraw X3® softwaresrespectively. The vascularization index was determined as describedby Pelster et al. (2003). Using Optimas 6.5 software, three equidistantlines, running parallel to the spinal cord, were drawn on the vesselbed image over the intersegmental vessels network from head to tailregion. The intersection of the vessel with the line was detected by thesoftware and displayed as a peak in the luminance spectrum. A sum ofthese peaks from all lines represented the relative index of thevascularization of that fish.
2.2. Fish length measurement
Fish total length was measured in both groups in order to check ifany retardation in the hypoxic larva body length was developed thatmay lead to differences between the two groups. Optimas 6.5software was applied to determine the fish length from the originalacquired images that were prepared to study the vascular bed. Thetransparent embryo provided the advantage to view the notochordrunning throughout the body. A line running from posterior tip of thenotochord up to the posterior margin of ear was drawn and itsmeasurement in millimeters was extracted on an excel sheet.
2.3. Anoxic viability and cardiac re-animation
From the aquarium facility in Zoology department, InnsbruckUniversity, wdt zebrafish eggs were obtained and kept in small plasticbeakers (9×6×6 cm)with two lateralmeshes andhung in anaquariumat 25 °C at 14:10 h light–dark conditions under normoxic condition. At4 dpf, 20 active and healthy embryos were selected and placed into the0.5 mL capacity, tap water filled, cylindrical, glass animal chamber withextra fine gold mesh at both ends to prevent escape of fish. It wasconnected with the twin-flow respirometer at both inflow and outflowends (Gnaiger, 1983). The animal chamber was supplied with tworeservoir bottles of 500 mL capacity independently that were also filledwith tap water. There was a manual valve to switch water supply fromeither of the bottle to the animal chamber. Continuous water flow wascarried out by a peristaltic pump joined at the outflow and adjusted at aperfusion rate of 0.533mLmin−1. The oxygen electrodes or POS, thatwere present at both the ends of animal chamber, were previouslycalibrated by zero oxygen solution containing sodium thioglycolate inthe tap water for the removal of free oxygen and its confirmation wascarried out by Redox dye Resazurinwhose blue colour diminishes underthis effect (Bright and Ellis, 1992; Padilla and Roth, 2001; Padilla et al.,2002; Hajeri et al., 2005). Normoxic reservoir was continuously purgedwith airwhile the anoxic bottlewas suppliedwith the nitrogengas froma cylinder. This whole setup was completely immersed in a water bathadjusted at 25 °C. Extreme care was taken to avoid entrapment of airbubbles inside the system. Both POS were connected to a control unitwhich automatically switched off the nitrogen supply when dissolvedoxygen concentration value reached below 0.5 % in the anoxic waterreservoir. A paper recorder was also connected to the system to getcontinuous measurement data. Prior to the experiment, anoxic bottlewas continually supplied with nitrogen for at least two and a half hours
to ensure the complete removal of oxygen from water. The animalchamber with fish inside was supplied with normoxic water for 1 h toacclimatize the fish and then the chamber was shifted to anoxic waterinflowby turning the valvemanually.Within 5 min the animal chamberwasfilledwith anoxicwater as recorded by both the inflowand outflowPOS and the fish became non-active within 10 min. The fish wasconstantly exposed to anoxia for 90 min by continuous supply fromanoxic bottle. After this exposure, fishwere removed carefully from theanimal chamber and shifted to microscopic chamber with normoxicwater at 25 °C and placed on the temperature controlled stage of aninverted microscope. The non-disintegrated fish were in a state ofsuspended animation without any heart beat and other dynamicprocesses.
Anoxic viability: to assess the viability of anoxia exposed fish, thepercentage of dead fish was noted down and their images wererecorded immediately after removing from anoxia under 10× lenspower and at each 15 min intervals up till 2 h in normoxic conditions.Partially or completely disintegrated fish were considered deadsimilar to the assays described in Mendelsohn et al. (2008). Deadembryos were removed and normoxic water was added continuously.After 2 h of study, the live fish were shifted to 9×6×6 cm plasticbeakers with two lateral meshes and hung in an aquarium at 25 °C at14:10 h light–dark conditions under normoxic condition. 24 and 48 h(5 and 6 dpf respectively) post-anoxic viability was recorded andexperiment was terminated at this point.
Cardiac re-animation: for re-animated heart beat measurement,fish were studied from 0 to 2 h and at 24 and 48 h post-anoxiaexposure under the 10× microscopic lens power. Heart rate of threerandomly selected fish (total samples=15, from five experiments)was recorded using stop watch and the statistical mean wascalculated.
Five independent experiments were carried out and the resultswere presented as their statistical mean with standard errors.
2.4. Oxygen consumption in vhl mutant
Inbreeding of vhl+/− heterozygous zebrafish (mutant typediscussed in the Introduction section) that were available in theaquarium facility in Zoology department, Innsbruck University,Austria was carried out and the eggs were shifted to petri dishesfilled with egg water and placed in incubator at 28 °C at 14:10 hlight–dark conditions under normoxic condition. The fish werenever fed during the experiment. At 6–7 dpf, the homozygousmutants (vhl−/−) were screened out under the microscope thatwere identified by hyperventilatory behavior (80 buccal move-ments per minute versus 1 per minute in non-mutants) andseverely enlarged heart. They were shifted to small plastic beakers(9×6×6 cm) with two lateral meshes and hung in an aquarium at25 °C at 14:10 h light–dark conditions under normoxic condition.At 8 dpf, respirometric study of 20 active fish was carried out intwin-flow respirometer with normoxic water supplied at aperfusion flow rate of 0.533mL min−1 to the animal chamber.This whole setup was completely immersed in a water bathadjusted at 25 °C. There was an automatic switching of the valveafter every one hour by the control unit to record the oxygenpercentage difference between fresh water (inflow) and larvautilized water (outflow) at the same POS. Oxygen reduction ratio(RO2, oxygen percentage difference divided with inflow wateroxygen percentage), water perfusion flow (V, mm3/s) and dis-solved oxygen value at specific atmospheric pressure at 25 °C ininflow water (Coxy, µmol/L) were used to determine fish oxygenconsumption (FNO2, nmol h−1) by the equation RO2×Coxy×V asdescribed by Gnaiger (1983).
Background respiration or the Bacterial oxygen consumption(Dalla Via, 1983) was subtracted from fish oxygen consumption. Itwas calculated by measuring the oxygen consumption recorded in

478 N. Yaqoob, T. Schwerte / Comparative Biochemistry and Physiology, Part A 156 (2010) 475–484
the animal chamber in the absence of fish at the start and at theend of the experiment by the above mentioned procedure. Fiveindependent experiments were carried out for both the vhl−/− andnon-mutant larvae and the result is presented as their statisticalmean with standard errors.
2.5. Statistical analysis
The statistical analyses were performed with Statistica 6.0 (Stat Soft,Inc) and for the comparison of two mean values, statistical significancewas evaluated by Student's t-test. The significant difference wasdetermined at Pb0.05. The final values were expressed as the meanalong with its standard error.
Fig. 1. Cardiac activity. Comparison of (A) cardiac output (nL min−1), (B) end-diastolic volum(PO2=20 Torr, 3 kPa) zebrafish embryos. No. of samples (N): hypoxic=10, control=5, * m
3. Results
3.1. Cardiac activity
Cardiac output was higher in hypoxic fish especially at 6 dpf (57.5±8 nL min−1)while in normoxicfish this valuewas at 20.7±3.2 nL min−1
(Fig. 1A). This differencewas also recordedduring further development oflarva. Fish with low oxygen environment maintained the cardiac outputbetween67and81 nL min−1 from7 to10 dpf in contrast tonormoxicfishwhere itwaskept below37 nL min−1 for the same timeperiod. Therewassignificant difference between the two groups within 48 h (at 3 dpf) ofhypoxic exposure (hypoxic: 59.3±6.9 nL min−1, normoxic: 33.5±7.3 nL min−1). EDVwas significantly higher nearly throughout the periodof study in hypoxic group. The values were always in between 0.8 and
e (nL), (C) heart rate (bpm) and (D) stroke volume (nL) between normoxic and hypoxicarks significant differences (Pb0.05).

479N. Yaqoob, T. Schwerte / Comparative Biochemistry and Physiology, Part A 156 (2010) 475–484
1.1 nL while the normoxic larvae exhibited the values between 0.45 and0.63 nL (Fig. 1B). At 6 dpf, hypoxic EDV reached to 1.11±0.12 nL thatwasthe maximum value achieved by any fish in both the groups. The heartrate offish fromthe two treatments ranged from125 to200bpm(Fig. 1C).But, there was irregularity in the pattern in the hypoxic group and thevalue of 170.4±2.6bpm at 3 dpf was recorded that decreased to 140.8±6 bpm at 6 dpf and after that we observed a sudden increase at 7dpf(186.9±2.7 bpm) followed by a gradual decrease and it reached up to159±9.7 bpm at 10 dpf. While in the normoxic group, the heart ratemaximum value was at 4 dpf (195.2±14.2 bpm) after a slight increasefrom 166.8±6.4 bpm at 3 dpf and then a gradual decrease was observedthat reached up to 120.9±5.3 at 10 dpf. The cardiac stroke volumeexhibited significant difference between the two groups (Fig. 1D). Thegeneral trend of the hypoxic fish was to maintain the stroke volume inbetween 0.3 and 0.5 nL while in normoxic fish the values were between0.1 and 0.3 nL. It was significantly higher at 3, 6 and 10 dpf under hypoxicconditions that were 0.34±0.03, 0.4±0.06 and 0.4±0.09 nL respective-ly. The value of stroke volume in normoxic individuals at 10 dpfwas quitelower than its hypoxic counterpart (0.1±0.03 nL).
3.2. Fish length
Generally, both the groups showed nearly the same body length upto 10 dpf and no major difference was observed (Fig. 2). There wassome difference at 3 dpf and the hypoxic group showed larger bodylength (2.93±0.02 mm) than the normoxic one (2.71±0.05 mm) butthis difference was inverse at 7 dpf (hypoxic: 2.7±0.08 mm,normoxic: 3.09±0.09 mm) and 8 dpf (hypoxic: 2.96±0.01 mm,normoxic: 3.16±0.02 mm).
3.3. Blood vessels network visualization and vascularization index
Hypoxic and normoxic fish intersegmental vessels quantificationwas determined from their vascular bed images. Average vasculariza-tion index for the normoxic fish was in the range of 69 to 83 while inhypoxicfish its lowest valuewas81±2at 3 dpfwhich climbed to90±2at 6 dpf that was 22% higher than the fish that developed undernormoxic conditions. This elevated trend in vascularization index wasmaintained by hypoxic fish also during further period of developmentand at 10 dpf it was 11% higher (89.5±2.5) when compared with thenormoxicfish (80.6±2.33) (Fig. 3A). Also, at 9 and 10 dpf, the thicknessof blood vessels in hypoxic vessel was quite increased (for vessel
Fig. 2. Fish length. Comparison of length (mm) development between normoxic andhypoxic (PO2=20 Torr, 3 kPa) zebrafish embryos. No of samples (N): hypoxic=10,control=5, * marks significant differences (Pb0.05).
diameter measurement data, see Yaqoob et al, 2009) that was visible inthe whole blood vessel cast (Fig. 3B).
3.4. Anoxic viability
90 min of anoxic conditions lead to 10% mortality and the non-disintegrated fish displayed suspended animation. There was increasein mortality rate when the survived fish were re-introduced tonormoxic conditions. Within 2 h, 50% of the anoxia exposed fishdisintegrated (Fig. 4A). Further 30%mortality was recorded 24 h post-anoxia. Later, death rate decreased and only 10% more larvaedisintegrated during the next 24 h (6 dpf) (Fig. 4B). Disintegration,particularly at the head region, was observed in some larvae evenafter they regained the heart beat.
3.5. Heart re-animation
The first sign of post-anoxic re-animation appeared as very slightcardiac movement after 5 min in normoxia that was otherwise incomplete suspension and within 10 min, the heart beat reached to77 bpm. It increased to 106 and 116 bpm at 30 and 45 minrespectively that was maintained until 2 h post-anoxia (Fig. 5A).During this whole period, experimental animal had significantlylower heart beat than the controlled animals that had a heart beatnear to 180 bpm. Heart beat progression was slow in post-anoxic fishand even after 24 h, it reached near to 156±6 bpm that wassignificantly lower compared to control values at 5 dpf that was at193±3 bpm. This difference was narrowed down at 6 dpf (48 h afteranoxia) and 172 bpm was attained in anoxia exposed group. At thattime, untreated zebrafish heart rate was 186 bpm value and there wasless significant difference between the two groups (Fig. 5B).
3.6. Oxygen consumption in vhl mutant
The oxygen consumption in 8 dpf vhl−/− mutant was significantlylower with a value of 6.5±0.20 as compared to the non-mutant thatwas at 9.5±0.58 nmol h−1
fish −1.
4. Discussion
4.1. Cardiac activity
Within 48 h of hypoxia, at 3 dpf, significantly higher cardiac outputvalue was displayed and it was kept elevated up till 10 dpf thatsuggested compensation in the cardiovascular system to overcomethe demand of oxygen that was available in low quantity. Only at 4 dpfwe observed it to be same as in normoxic fish that was maybe due tothe animal's initial and short duration adjustment with the naturallydeveloped vessels and cardiac capacity. Jacob et al. (2002) studies thatwere carried out in zebrafish embryo at 10 kPa oxygen partialpressure, demonstrated nearly the same difference between thehypoxic and normoxic group at 3 and 5 dpf. They recorded highercardiac output at 4 dpf also in the hypoxic ones but in our study, bothgroups showed the same cardiac output. In agreement with theirstudies, EDV was significantly higher in the hypoxic animals from 3 to10 dpf with the exception that their hypoxic group values werebetween 0.5 and 0.8 nL in contrast to our study where it was alwaysbetween 0.8 and 1.1 nL. The difference between the two observationsmay be due to the fact that we have applied extreme hypoxicconditions rather than mild hypoxia that was used in their studies.Less availability of oxygen in the environment and, additionally,formation of new vessels may lead to heart engrossment beyond itsnatural capacity to overcome this situation that would provide forceto supply blood to remote parts of the bodywhich were not that muchvascularized under normoxic conditions.

Fig. 3. A. Vascularization index. Comparison of intersegmental vessels quantity in normoxic and hypoxic zebrafish from 3 to 10 dpf. Number of samples (N)=3–10. * markssignificant differences (Pb0.05). B. Vessel bed visualization in 9 (above) and 10 (below) dpf normoxic and hypoxic zebrafish larva made by joining ImagePro® processed imagesequences of different regions using CorelDrawX3 software. Intersegmental vessels complexity and dorsal and ventral caudal veins thickness are markedly increased in hypoxiclarva. CA = caudal artery, CV = caudal vein, Se = intersegmental vessel.
480 N. Yaqoob, T. Schwerte / Comparative Biochemistry and Physiology, Part A 156 (2010) 475–484
From 3 to 6 dpf developmental age, hypoxic bradycardia wasobserved that was in good agreement with the findings of Steele et al.(2009) who studied it at 30 and 40 Torr PO2 but, after 6 dpf, in ourstudy, we noticed a situation of increased heart rate in the samegroup. This hypoxic tachycardia has been demonstrated also by Jacobet al. (2002). The slow blood circulation in the early stages ofdevelopment (until 6 dpf) may lead to increased time for gas diffusionat gills and myocardium as also advocated by Farrell (2007). Anotherexplanation could be that the embryos under extreme hypoxiashowed tremendous individual variation and the animal develop-mental plasticity at extreme conditions is often drastic to acclimatizewith the situation as also suggested by Bagatto (2005). Stroke volume
was higher in the hypoxic animal throughout 3–10 dpf with signifi-cant difference at 3, 6 and 10 dpf. This suggested a physiologicalresponse that leads to compensate the low oxygen in the environ-ment, as it was also observed in case of cardiac output and EDV, and itmay help in enhancing the supply to the remote parts of the bodymuch more efficiently.
Cardiac output, EDV and stroke volume displayed significantdifference at 3 and 6 dpf while in the case of heart rate, at the samedevelopmental stage, the difference was not significant. It would bequite relevant to carry out future studies including genetic expressionpattern to understand this behavioral pattern changes in hypoxicindividuals for this age group.
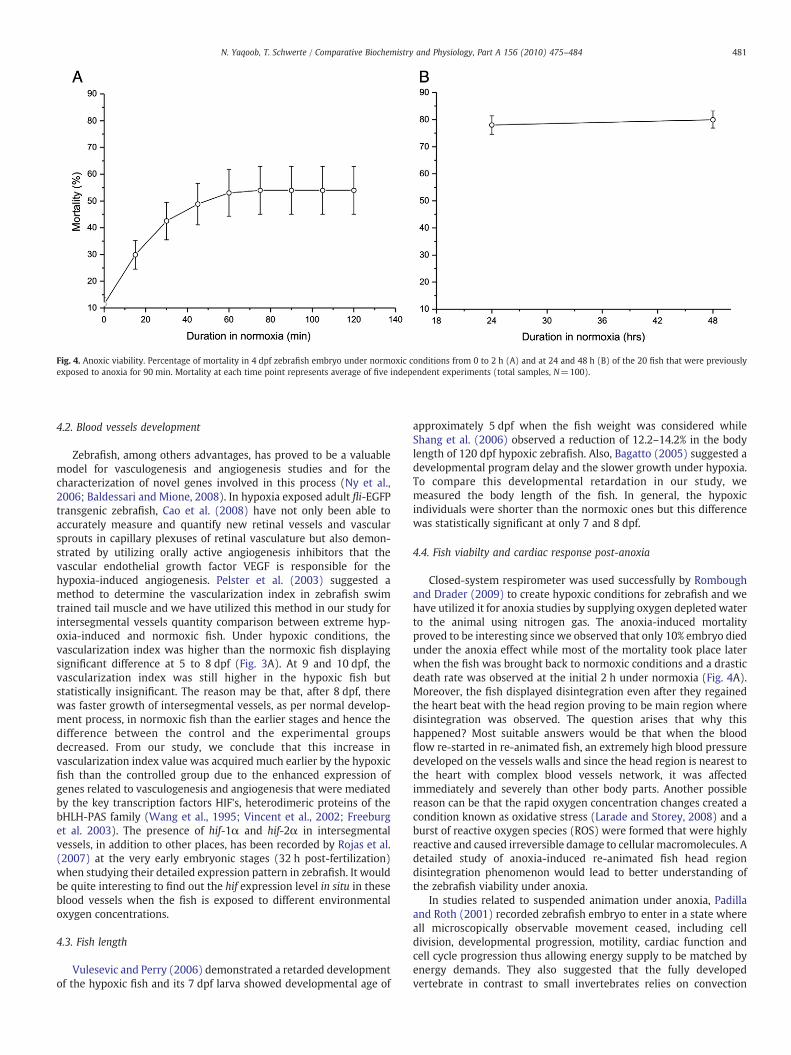
Fig. 4. Anoxic viability. Percentage of mortality in 4 dpf zebrafish embryo under normoxic conditions from 0 to 2 h (A) and at 24 and 48 h (B) of the 20 fish that were previouslyexposed to anoxia for 90 min. Mortality at each time point represents average of five independent experiments (total samples, N=100).
481N. Yaqoob, T. Schwerte / Comparative Biochemistry and Physiology, Part A 156 (2010) 475–484
4.2. Blood vessels development
Zebrafish, among others advantages, has proved to be a valuablemodel for vasculogenesis and angiogenesis studies and for thecharacterization of novel genes involved in this process (Ny et al.,2006; Baldessari and Mione, 2008). In hypoxia exposed adult fli-EGFPtransgenic zebrafish, Cao et al. (2008) have not only been able toaccurately measure and quantify new retinal vessels and vascularsprouts in capillary plexuses of retinal vasculature but also demon-strated by utilizing orally active angiogenesis inhibitors that thevascular endothelial growth factor VEGF is responsible for thehypoxia-induced angiogenesis. Pelster et al. (2003) suggested amethod to determine the vascularization index in zebrafish swimtrained tail muscle and we have utilized this method in our study forintersegmental vessels quantity comparison between extreme hyp-oxia-induced and normoxic fish. Under hypoxic conditions, thevascularization index was higher than the normoxic fish displayingsignificant difference at 5 to 8 dpf (Fig. 3A). At 9 and 10 dpf, thevascularization index was still higher in the hypoxic fish butstatistically insignificant. The reason may be that, after 8 dpf, therewas faster growth of intersegmental vessels, as per normal develop-ment process, in normoxic fish than the earlier stages and hence thedifference between the control and the experimental groupsdecreased. From our study, we conclude that this increase invascularization index value was acquired much earlier by the hypoxicfish than the controlled group due to the enhanced expression ofgenes related to vasculogenesis and angiogenesis that were mediatedby the key transcription factors HIF's, heterodimeric proteins of thebHLH-PAS family (Wang et al., 1995; Vincent et al., 2002; Freeburget al. 2003). The presence of hif-1α and hif-2α in intersegmentalvessels, in addition to other places, has been recorded by Rojas et al.(2007) at the very early embryonic stages (32 h post-fertilization)when studying their detailed expression pattern in zebrafish. It wouldbe quite interesting to find out the hif expression level in situ in theseblood vessels when the fish is exposed to different environmentaloxygen concentrations.
4.3. Fish length
Vulesevic and Perry (2006) demonstrated a retarded developmentof the hypoxic fish and its 7 dpf larva showed developmental age of
approximately 5 dpf when the fish weight was considered whileShang et al. (2006) observed a reduction of 12.2–14.2% in the bodylength of 120 dpf hypoxic zebrafish. Also, Bagatto (2005) suggested adevelopmental program delay and the slower growth under hypoxia.To compare this developmental retardation in our study, wemeasured the body length of the fish. In general, the hypoxicindividuals were shorter than the normoxic ones but this differencewas statistically significant at only 7 and 8 dpf.
4.4. Fish viabilty and cardiac response post-anoxia
Closed-system respirometer was used successfully by Romboughand Drader (2009) to create hypoxic conditions for zebrafish and wehave utilized it for anoxia studies by supplying oxygen depleted waterto the animal using nitrogen gas. The anoxia-induced mortalityproved to be interesting since we observed that only 10% embryo diedunder the anoxia effect while most of the mortality took place laterwhen the fish was brought back to normoxic conditions and a drasticdeath rate was observed at the initial 2 h under normoxia (Fig. 4A).Moreover, the fish displayed disintegration even after they regainedthe heart beat with the head region proving to be main region wheredisintegration was observed. The question arises that why thishappened? Most suitable answers would be that when the bloodflow re-started in re-animated fish, an extremely high blood pressuredeveloped on the vessels walls and since the head region is nearest tothe heart with complex blood vessels network, it was affectedimmediately and severely than other body parts. Another possiblereason can be that the rapid oxygen concentration changes created acondition known as oxidative stress (Larade and Storey, 2008) and aburst of reactive oxygen species (ROS) were formed that were highlyreactive and caused irreversible damage to cellular macromolecules. Adetailed study of anoxia-induced re-animated fish head regiondisintegration phenomenon would lead to better understanding ofthe zebrafish viability under anoxia.
In studies related to suspended animation under anoxia, Padillaand Roth (2001) recorded zebrafish embryo to enter in a state whereall microscopically observable movement ceased, including celldivision, developmental progression, motility, cardiac function andcell cycle progression thus allowing energy supply to be matched byenergy demands. They also suggested that the fully developedvertebrate in contrast to small invertebrates relies on convection

Fig. 5. Heart rate in re-animated zebrafish embryo. Comparison of heart rate (bpm) between 4 dpf zebrafish control zebrafish embryo and 90 min anoxia exposed embryo of thesame age that were re-immersed under normoxic conditions for 0–2 h (A) and at 24 and 48 h (B). At each data point, fish were examined under microscope and heart rate in 3 viablefish was recorded. Five independent experiments were carried out and average value (total samples, N=15) presented. * marks significant difference (Pb0.05).
482 N. Yaqoob, T. Schwerte / Comparative Biochemistry and Physiology, Part A 156 (2010) 475–484
instead of diffusion and therefore may be unable to adjust rapidlyenough to survive anoxia. But, one must not overlook the fact that thehypoxia and anoxia difference is related to organism morphology.Sollid et al. (2005), in crucian carp Carassius carassius, observed 7.5-fold increase of respiratory surface area under hypoxia but thisresponse was not triggered in anoxia. Also, in response to 7 days ofanoxia, there was a reduced incidence of S-phase cells (from 12.2 to5.0%) in gills, decreased number of S-phase cells in intestine (from 8.1to 1.8%) and there was no change in the fraction of S-phase cells(nearly1%) in liver revealing a considerable rate of DNA synthesis.Involvement of chemical factors in suspended animation has alsobeen observed by different authors. Nitric oxide (NO) inducedsuspended animation has been recorded to promote survival duringhypoxia. It promotes vaso-dilation and its levels are thought to rise atsites that are hypoxic due to increased respiratory demands or poorperfusion. It is a putative hypoxia signal and induce a reversible arrestof development, gene expression and turnover (Fritsche et al., 2000;Teodoro and O'Farrell, 2003). In case of carbon monoxide (CO), it hasbeen found out that its induction protects against hypoxic damagewhen in suspended animation. Carbon monoxide binds to sites whereoxygen binds in the body and it has been found that C. elegansembryos can be put into a state of suspended animation using carbonmonoxide and its embryo was protected from damage underintermediate concentrations of oxygen (0.01 and 0.1 kPa, normallylethal) when CO was induced. In addition, CO also rescued thehypoxia-sensitive mutant hif-l(ia04) from lethality in hypoxia (Nystuland Roth, 2004). Zebrafish embryo has been recorded to surviveunder the absence of oxygen. When exposed to anoxia for a longerperiod, it shows a state of suspended animation but re-animates whenoxygen is supplied again (referred earlier).We have selected the heartrate for our study in re-animated zebrafish embryo as it providesdirect estimation of suspension or re-animation activity. The anoxiaexposed fish regained the normal heart rate after 48 h when broughtto normoxia again. Before that it was significantly lower than theembryos in control condition.
Many attempts were carried out by us to determine the oxygenconsumption in the re-animated embryo but it was not found feasible byusing the twin-flow respirometer that was available to us as there weremany factors involved that lead toerroneous results. Therewere strikinglydifferent results in independent experiment. Themain problemswere thedifference in fish viability in individual experiment and delay in responseof the POSwhen normoxicwaterwas re-supplied to the animal chamber.
But, the oxygen consumption in re-animated embryo would be aninteresting studyand itwouldhelp inunderstandingvariousphysiologicalbehaviors related to survival under anoxia andwesuggest that someothermethod should be devised to carry out this study.
4.5. Comparison of genetically hypoxic vhl mutant with environmentallyinfluenced hypoxic non-mutant
We were interested in correlating the observed cardiovascularbehavior under hypoxia with that of the genetically modifiedzebrafish. Some studies related to mutants for this purpose havebeen carried out by different authors to search for genes related tocardiac morphogenesis and heart rhythm (Kopp et al., 2005). Wepropose that the valentine (vtn), heart of glass (heg) or the santa (san)genes related to large distended heart pointed out by Stainier et al.(1996) and Chen et al. (1996) were maybe up-regulated in ourhypoxic individuals since we have observed quite large EDV in themin comparison with the control animals. van Rooijen et al. (2009)observed general systemic hypoxic response in zebrafish lines thatcarried inactivating germline mutations in vhl gene involving up-regulated hypoxia-induced genes, severe hyperventilation and car-dio-physiological response. Its 8 dpf mutant displayed cardiacdilation, cardiomegaly, stretched cardiomyocytes and also increasedblood volume. They also recorded elevated heart rate, cardiac outputand stroke volume in vhl mutants. These behaviors of vhl mutants arestrikingly similar with the hypoxic stressed non-mutant individualsthat we studied but there were some differences between them incardiac performance. At 4 dpf, 3.2-fold elevated cardiac output wasrecorded in vhl mutant while the hypoxic fish in our study did notshow any significant difference with the normoxic fish. Similarly, at10 dpf, the vhl mutant's cardiac output was 15-fold increased while itwas only 7-fold increased in the hypoxic fish at the same age. Theheart rate in vhl mutant was elevated at 7 dpf that was in agreementwith the heart rate observed by us in hypoxic individuals but, inaddition, we recorded an increased cardiac output also (Fig 1A). Thisincrease was displayed by the vhl mutants with a delay of one day (at8 dpf). So, we conclude that our data only partly support thestatement that vhl mutant zebrafish display hypoxic response. Theincrease in Epo production under hypoxic conditions has beenconfirmed previously (Yaqoob et al., 2009). The increased in buccalmovement frequency at 8 dpf observed by van Rooijen et al. (2009)and other cardiovascular physiological behaviors lead us to measure

483N. Yaqoob, T. Schwerte / Comparative Biochemistry and Physiology, Part A 156 (2010) 475–484
the oxygen consumption of these individuals as this data was lackingand interestingly, we found that the vhl−/− mutant displayedsignificantly low oxygen consumption than the non-mutants. Asalso pointed out by Soliz et al. (2005), Epo controls ventilation at thecentral (brainstem) and peripheral (carotid body) levels so, weconclude that the hyperventilation in vhl mutants was maybe due toover-expression of Epo and it would be quite relevant to find out if thenon-mutant zebrafish display vhl mutant ventilatory behavior underthe effect of experimentally increased Epo levels. A possible techniquewould be to inject zEpo (zebrafish erythropoietin) mRNA intozebrafish embryos at the one-cell stage as demonstrated by Chuet al. (2007) who used it to rescue the animal frommorpholino effect.This study is currently in progress.
5. Conclusion
Quite extensive research is going on to know about the cardiacpatterning (Reifers et al., 2000), morphogenesis (Rohr et al., 2006;Rohr et al., 2008), regeneration (Lien et al., 2006) and genetic control(Huang et al., 2003; Beis et al., 2005). We have selected zebrafish forstudies relating to the physiological aspect of this important organunder hypoxic, anoxic and normoxic environment. For the heartimaging and vessel bed view we have used inverted microscopeconnected with high-speed video imaging camera and with the helpof computer software we have been able to determine heartperformance and vessels development non-invasively. Also we havesuccessfully determined the oxygen consumption in hyperventilating,genetically hypoxic (vhl−/−) larva with the help of twin-flowrespirometer.
In zebrafish, it has been suggested that the cardiac activity is notdesired until 12 dpf and the oxygen supply to tissues for aerobicmetabolism is mostly accomplished by the diffusion through skin(Pelster and Burggren, 1996; Jacob et al., 2002; Rombough, 2002). Butit can be argued that under extremely low oxygen condition, theremust be a need of an additional help to make the best use of limitedresources for survival. This task, may be, taken up by the heart since itis responsible for supply of oxygen to the remotest and the deepesttissues of the body and the increased cardiac output, stroke volumeand EDV in our studied hypoxic individuals provided its evidence. Wecan correlate the increased cardiac activity with the findings ofRombough and Drader (2009) who also observed in zebrafish larvaeimportant role of hemoglobin in enhancing oxygen uptake duringextreme hypoxia. Our study clearly demonstrates that there wassignificant effect of extreme hypoxia on the cardiovascular perfor-mance and development. It is in contrast with the finding of Territoand Altimiras (1998) but in good agreement with Jacob et al. (2002).
Post-anoxia, coma situationwas observed in zebrafish as it enteredinto non-reactive, but nondormant, state. It failed to respond normallyto touch or light stimulus and did not take voluntary actions but thebasic requirement of the body to keep it alive including heart beat,yolk consumption, ventilation and gut movement were there. The fishseemed to be entered into a “stand-by”mode to conserve energy. Onecan hypothesize that it may allow anoxia affected body's internalmechanisms to heal more efficiently. This risk avoiding strategy, innature, has also been observed in Arctosa fulvolineata, the salt-marshspider to survive water inundation in its habitat that leads to hypoxicenvironment (Petillon et al., 2009). We also observed cardiac re-animation and blood flow revival post-anoxia in zebrafish and thuswe can conclude that, among other advantages, it can act as apromising vertebrate model for coma and reperfusion studies.
Acknowledgments
This work was supported by the University of Innsbruck andHigher Education Commission of Pakistan (HEC).
References
Bagatto, B., 2005. Ontogeny of cardiovascular control in zebrafish (Danio rerio): effectsof developmental environment. Comp. Biochem. Physiol. 141, 391–400.
Baldessari, D., Mione, M., 2008. How to create the vascular tree? (Latest) help from thezebrafish. Pharmacol. Ther. 118, 206–230.
Barrionuevo, W.R., Burggren, W.W., 1999. O2 consumption and heart rate in developingzebrafish (Danio rerio): influence of temperature and ambient O2. Am. J. Physiol.276, R505–R513.
Beis, D., Bartman, T., Jin, S.W., Scott, I.C., D'Amico, L.A., Ober, E.A., Verkade, H., Frantsve, J.,Field, H.A., Wehman, A., Baier, H., Tallafuss, A., Bally-Cuif, L., Chen, J.N., Stainier, D.Y.,Jungblut, B., 2005. Genetic and cellular analyses of zebrafish atrioventricular cushionand valve development. Development 132, 4193–4204.
Blackstone, E., Roth, M.B., 2007. Suspended animation-like state protects mice fromlethal hypoxia. Shock 27, 370–372.
Bosworth, C.A., Chou, C.W., Cole, R.B., Rees, B.B., 2005. Protein expression patterns inzebrafish skeletal muscle: initial characterization and the effects of hypoxicexposure. Proteomics 5, 1362–1371.
Bright, C.M., Ellis, D., 1992. Intracellular pH changes induced by hypoxia and anoxia inisolated sheep heart Purkinje fibres. Exp. Physiol. 77, 165–175.
Cao, R., Jensen, L.D., Soll, I., Hauptmann, G., Cao, Y., 2008. Hypoxia-induced retinalangiogenesis in zebrafish as a model to study retinopathy. PLoS ONE 3 (7), e2748.
Chen, J.N., Haffter, P., Odenthal, J., Vogelsang, E., Brand, M., van Eeden, F.J., Furutani-Seiki, M.,Granato, M., Hammerschmidt, M., Heisenberg, C.P., Jiang, Y.J., Kane, D.A., Kelsh, R.N.,Mullins, M.C., Nusslein-Volhard, C., 1996. Mutations affecting the cardiovascular systemand other internal organs in zebrafish. Development 123, 293–302.
Chico, T.J., Ingham, P.W., Crossman, D.C., 2008. Modeling cardiovascular disease in thezebrafish. Trends Cardiovasc. Med. 18, 150–155.
Chu, C.Y., Cheng, C.H., Chen, G.D., Chen, Y.C., Hung, C.C., Huang, K.Y., Huang, C.J., 2007.The zebrafish erythropoietin: functional identification and biochemical character-ization. FEBS Lett. 581, 4265–4271.
Dalla Via, G.J., 1983. Bacterial growth and antibiotics in animal respirometry. In:Gnaiger, E., Forstner, H. (Eds.), Polarographic Oxygen Sensors, Aquatic andPhysiological Applications. Springer Verlag, Berlin, pp. 202–218.
Driever, W., Stemple, D., Schier, A., Solnica-Krezel, L., 1994. Zebrafish: genetic tools forstudying vertebrate development. Trends Genet. 10, 152–159.
Farrell, A.P., 2007. Tribute to P. L. Lutz: a message from the heart—why hypoxicbradycardia in fishes? J. Exp. Biol. 210, 1715–1725.
Freeburg, P.B., Robert, B., St, John P.L., Abrahamson, D.R., 2003. Podocyte expression ofhypoxia-inducible factor (HIF)-1 and HIF-2 during glomerular development. J. Am.Soc. Nephrol. 14, 927–938.
Fritsche, R., Schwerte, T., Pelster, B., 2000. Nitric oxide and vascular reactivity indeveloping zebrafish, Danio rerio. Am. J. Physiol. Regul. Integr. Comp. Physiol. 279,R2200–R2207.
Gnaiger, E., 1983. The twin-flow respirometer and simultaneous calorimetry. In:Gnaiger, E., Forstner, H. (Eds.), Polarographic Oxygen Sensors, Aquatic andPhysiological Applications. Springer Verlag, Berlin, pp. 134–166.
Gracey, A.Y., Troll, J.V., Somero, G.N., 2001. Hypoxia-induced gene expression profilingin the euryoxic fish Gillichthys mirabilis. Proc. Natl Acad. Sci. U. S. A. 98, 1993–1998.
Grillitsch, S., Medgyesy, N., Schwerte, T., Pelster, B., 2005. The influence ofenvironmental Po2 on hemoglobin oxygen saturation in developing zebrafishDanio rerio. J. Exp. Biol. 208, 309–316.
Hajeri, V.A., Trejo, J., Padilla, P.A., 2005. Characterization of sub-nuclear changes inCaenorhabditis elegans embryos exposed to brief, intermediate and long-termanoxia to analyze anoxia-induced cell cycle arrest. BMC Cell Biol. 6, 47.
Hochachka, P.W., Lutz, P.L., 2001. Mechanism, origin, and evolution of anoxia tolerancein animals. Comp. Biochem. Physiol. B 130, 435–459.
Hochachka, P.W., Buck, L.T., Doll, C.J., Land, S.C., 1996. Unifying theory of hypoxiatolerance: molecular/metabolic defense and rescue mechanisms for survivingoxygen lack. Proc. Natl Acad. Sci. U. S. A. 93, 9493–9498.
Huang, C.J., Tu, C.T., Hsiao, C.D., Hsieh, F.J., Tsai, H.J., 2003. Germ-line transmission of amyocardium-specific GFP transgene reveals critical regulatory elements in thecardiac myosin light chain 2 promoter of zebrafish. Dev. Dyn. 228, 30–40.
Jacob, E., Drexel, M., Schwerte, T., Pelster, B., 2002. Influence of hypoxia and ofhypoxemia on the development of cardiac activity in zebrafish larvae. Am. J.Physiol. Regul. Integr. Comp. Physiol. 283, 911–917.
Jonz, M.G., Nurse, C.A., 2005. Development of oxygen sensing in the gills of zebrafish. J.Exp. Biol. 208, 1537–1549.
Kopp, R., Schwerte, T., Pelster, B., 2005. Cardiac performance in the zebrafishbreakdance mutant. J. Exp. Biol. 208, 2123–2134.
Kopp, R., Pelster, B., Schwerte, T., 2007. How does blood cell concentration modulatecardiovascular parameters in developing zebrafish (Danio rerio)? Comp. Biochem.Physiol. 146, 400–407.
Larade, K., Storey, K.B., 2008. Identification of a granulin-like transcript expressedduring anoxic exposure and translated during aerobic recovery in a marinegastropod. Gene 410, 37–43.
Lien, C.L., Schebesta, M., Makino, S., Weber, G.J., Keating, M.T., 2006. Gene expressionanalysis of zebrafish heart regeneration. PLoS Biol. 4, e260.
Marques, I.J., Leito, J.T., Spaink, H.P., Testerink, J., Jaspers, R.T., Witte, F., van den Berg, S.,Bagowski, C.P., 2008. Transcriptome analysis of the response to chronic constanthypoxia in zebrafish hearts. J. Comp. Physiol. B 178, 77–92.
Martinovic, D., Villeneuve, D.L., Kahl, M.D., Blake, L.S., Brodin, J.D., Ankley, G.T., 2009.Hypoxia alters gene expression in the gonads of zebrafish (Danio rerio). Aquat.Toxicol. 95, 258–272.
Mendelsohn, B.A., Kassebaum, B.L., Gitlin, J.D., 2008. The zebrafish embryo as a dynamicmodel of anoxia tolerance. Dev. Dyn. 237, 1780–1788.

484 N. Yaqoob, T. Schwerte / Comparative Biochemistry and Physiology, Part A 156 (2010) 475–484
Moore, F.B., Hosey, M., Bagatto, B., 2006. Cardiovascular system in larval zebrafishresponds to developmental hypoxia in a family specific manner. Frontiers Zool. 3, 4.
Ny, A., Autiero, M., Carmeliet, P., 2006. Zebrafish and Xenopus tadpoles: small animalmodels to study angiogenesis and lymphangiogenesis. Exp. Cell Res. 312, 684–693.
Nystul, T.G., Roth, M.B., 2004. Carbonmonoxide-induced suspended animation protectsagainst hypoxic damage in Caenorhabditis elegans. Proc. Natl Acad. Sci. U. S. A. 101,9133–9136.
Nystul, T.G., Goldmark, J.P., Padilla, P.A., Roth, M.B., 2003. Suspended animation in C.elegans requires the spindle checkpoint. Science 302, 1038–1041.
Padilla, P.A., Roth, M.B., 2001. Oxygen deprivation causes suspended animation in thezebrafish embryo. Proc. Natl Acad. Sci. U. S. A. 98, 7331–7335.
Padilla, P.A., Nystul, T.G., Zager, R.A., Johnson, A.C., Roth, M.B., 2002. Dephosphorylationof cell cycle-regulated proteins correlates with anoxia-induced suspendedanimation in Caenorhabditis elegans. Mol. Biol. Cell 13, 1473–1483.
Pandey, R., Heeger, S., Lehner, C.F., 2007. Rapid effects of acute anoxia on spindle kinetochoreinteractions activate the mitotic spindle checkpoint. J. Cell Sci. 120, 2807–2818.
Patton, E.E., Zon, L.I., 2001. The art and design of genetic screens: zebrafish. Nat. Rev.Genet. 2, 956–966.
Pelster, B., Burggren, W.W., 1996. Disruption of hemoglobin oxygen transport does notimpact oxygen-dependent physiological processes in developing embryos of zebrafish (Danio rerio). Circ. Res. 79, 358–362.
Pelster, B., Sanger, A.M., Siegele, M., Schwerte, T., 2003. Influence of swim training oncardiac activity, tissue capillarization, andmitochondrial density in muscle tissue ofzebrafish larvae. Am. J. Physiol. Regul. Integr. Comp. Physiol. 285, R339–R347.
Petillon, J., Montaigne, W., Renault, D., 2009. Hypoxic coma as a strategy to surviveinundation in a salt-marsh inhabiting spider. Biol. Lett. 5, 442–445.
Reifers, F., Walsh, E.C., Leger, S., Stainier, D.Y., Brand, M., 2000. Induction anddifferentiation of the zebrafish heart requires fibroblast growth factor 8 (fgf8/acerebellar). Development 127, 225–235.
Rohr, S., Bit-Avragim, N., Abdelilah-Seyfried, S., 2006. Heart and soul/PRKCi and nagieoko/Mpp 5 regulate myocardial coherence and remodeling during cardiacmorphogenesis. Development 133, 107–115.
Rohr, S., Otten, C., Abdelilah-Seyfried, S., 2008. Asymmetric involution of themyocardialfield drives heart tube formation in zebrafish. Circ. Res. 102, e12–e19.
Rojas, D.A., Perez-Munizaga, D.A., Centanin, L., Antonelli, M., Wappner, P., Allende, M.L.,Reyes, A.E., 2007. Cloning of hif-1alpha and hif-2alpha and mRNA expressionpattern during development in zebrafish. Gene Expr. Patterns 7, 339–345.
Rombough, P., 2002. Gills are needed for ionoregulation before they are needed for O2
uptake in developing zebrafish, Danio rerio. J. Exp. Biol. 205, 1787–1794.Rombough, P., Drader, H., 2009. Hemoglobin enhances oxygen uptake in larval zebrafish
(Danio rerio) but only under conditions of extreme hypoxia. J. Exp. Biol. 212, 778–784.Schwerte, T., Pelster, B., 2000. Digital motion analysis as a tool for analysing the shape
and performance of the circulatory system in transparent animals. J. Exp. Biol. 203,1659–1669.
Shang, E.H.H., Yu, R.M.K., Wu, R.S.S., 2006. Hypoxia affects sex differentiation anddevelopment, leading to a male-dominated population in zebrafish (Danio rerio).Environ. Sci. Technol. 40, 3118–3122.
Soliz, J., Joseph, V., Soulage, C., Becskei, C., Vogel, J., Pequignot, J.M., Ogunshola, O.,Gassmann, M., 2005. Erythropoietin regulates hypoxic ventilation in mice byinteracting with brainstem and carotid bodies. J. Physiol. 568, 559–571.
Sollid, J., Kjernsli, A., De Angelis, P.M., Rohr, A.K., Nilsson, G.E., 2005. Cell proliferationand gill morphology in anoxic crucian carp. Am. J. Physiol. Regul. Integr. Comp.Physiol. 289, R1196–R1201.
Stainier, D.Y.R., Fishman, M.C., 1994. The zebrafish as a model system to studycardiovascular development. Trends Cardiovasc. Med. 4, 207–212.
Stainier, D.Y., Fouquet, B., Chen, J.N., Warren, K.S., Weinstein, B.M., Meiler, S.E.,Mohideen, M.A., Neuhauss, S.C., Solnica-Krezel, L., Schier, A.F., Zwartkruis, F.,Stemple, D.L., Malicki, J., Driever, W., Fishman, M.C., 1996. Mutations affecting theformation and function of the cardiovascular system in the zebrafish embryo.Development 123, 285–292.
Steele, S.L., Lo, K.H., Li, V.W., Cheng, S.H., Ekker, M., Perry, S.F., 2009. Loss of M2
muscarinic receptor function inhibits development of hypoxic bradycardia andalters cardiac beta-adrenergic sensitivity in larval zebrafish (Danio rerio). Am. J.Physiol. Regul. Integr. Comp. Physiol. 297, R412–R420.
Teodoro, R.O., O'Farrell, P.H., 2003. Nitric oxide-induced suspended animationpromotes survival during hypoxia. EMBO J. 22, 580–587.
Territo, P.R., Altimiras, J., 1998. The ontogeny of cardio-respiratory function underchronically altered gas compositions in Xenopus laevis. Respir. Physiol. 111,311–323.
Turesson, J., Schwerte, T., Sundin, L., 2006. Late onset of NMDA receptor-mediatedventilatory control during early development in zebrafish (Danio rerio). Comp.Biochem. Physiol. 143, 332–339.
van Rooijen, E., Voest, E.E., Logister, I., Korving, J., Schwerte, T., Schulte-Merker, S., Giles,R.H., van Eeden, F.J., 2009. Zebrafish mutants in the von Hippel–Lindau tumorsuppressor display a hypoxic response and recapitulate key aspects of Chuvashpolycythemia. Blood 113, 6449–6460.
Vincent, K.A., Feron, O., Kelly, R.A., 2002. Harnessing the response to tissue hypoxia:HIF-1 alpha and therapeutic angiogenesis. Trends Cardiovasc. Med. 12, 362–367.
Vulesevic, B., Perry, S.F., 2006. Developmental plasticity of ventilatory control inzebrafish, Danio rerio. Respir. Physiol. Neurobiol. 154, 396–405.
Vulesevic, B., McNeill, B., Perry, S.F., 2006. Chemoreceptor plasticity and respiratoryacclimation in the zebrafish Danio rerio. J. Exp. Biol. 209, 1261–1273.
Wang, G.L., Jiang, B.H., Rue, E.A., Semenza, G.L., 1995. Hypoxia-inducible factor 1 is abasic-helix–loop-helix–PAS heterodimer regulated by cellular O2 tension. Proc.Natl Acad. Sci. U. S. A. 92, 5510–5514.
Yaqoob, N., Holotta, M., Prem, C., Kopp, R., Schwerte, T., 2009. Ontogenetic developmentof erythropoiesis can be studied non-invasively in GATA-1:DsRed transgeniczebrafish. Comp. Biochem. Physiol. 154, 270–278.




















