
Torque–Speed Relationships of Na+-driven Chimeric Flagellar Motors in Escherichia coli
Upload
independentCategory
view
0download
0
Pud
LWQa
b
a
ARRAA
KFFCMSZ
1
db(2ad
cf
h0
Micron 66 (2014) 1–8
Contents lists available at ScienceDirect
Micron
j ourna l ho me page: www.elsev ier .com/ locate /micron
re-spermiogenic initiation of flagellar growth and correlativeltrastructural observations on nuage, nuclear and mitochondrialevelopmental morphology in the zebrafish Danio rerio
inli Zhanga, Ping Yanga, Yi Liua, Xunguang Biana, Shakeeb Ullaha, Qian Zhanga,ei Chena, Yuan Lea, Bing Chena, Jinxing Linb, Cheng Gaob, Jianhua Hub,
iusheng Chena,∗
Laboratory of Animal Cell Biology and Embryology, College of Veterinary Medicine, Nanjing Agricultural University, Nanjing, PR ChinaShanghai Laboratory Animal Research Centre, Shanghai Institutes for Biological Sciences, Shanghai, China
r t i c l e i n f o
rticle history:eceived 6 January 2014eceived in revised form 20 March 2014ccepted 20 March 2014vailable online 31 March 2014
eywords:ine structurelagellahromatinitochondria
permatidebrafish
a b s t r a c t
The microstructural and ultrastructural changes of germ cells during spermatogenesis of zebrafish (Daniorerio) were examined using light microscopy (LM) and transmission electron microscopy (TEM). Generallythe process of spermatogenesis in zebrafish is similar to that of other teleosts, however, here we describesome peculiar features of zebrafish spermatogenic cells which have a limited report in this species. (1)The basic events of spermiogenesis are asynchronous, location of flagellum finished in initial stage, whilechromatin condensation sharply occurred in intermediate stage and elimination of excess cytoplasmmainly taken place in final stages. (2) Surprisingly, the cilia or initial flagellae are created in sperma-tocytes, approach toward the nucleus of early stage spermatids, and then the centrioles depress intonuclear fossa and change their orientation to each other from right angle to obtuse angle about 125◦. (3)During spermatogenesis, the chromatin compaction performs in a distinctive pattern, condensed hetero-geneously from granular into chromatin clumps with central electron-lucent areas, round or long, whichdiminished to small nuclear vacuoles in spermatozoa. This finding demonstrates the origin of nuclearvacuoles in zebrafish spermatozoa for the first time. (4) Nuages are observed in both spermatogonia andspermatocytes. They are connected with the mitochondria and nuclear membrane, and are even located
in the perinuclear spaces of spermatogonia nuclei. (5) Mitochondrial morphology and distribution showsdiversity in different germ cells. The condensed mitochondria appear in pachytene spermatocytes, andmitochondria including membrane conglomerate exist in both spermatocytes and spermatids. This studywas undertaken in order to disclose specific spermatogenic cells features in zebrafish that could be helpfulfor understanding the correlative function in this model species.. Introduction
The zebrafish (Teleostei, Cyprinidae) is a model organism inevelopmental, physiological, toxicological, and genetic studiesecause it offers several advantages to researchers in these fieldsBriggs, 2002; Hill, 2005; Norton and Bally-Cuif, 2010; Traver et al.,
003). For example, the transparent embryos have been invalu-ble for developmental studies, and the short reproductive andevelopmental cycles allow for rapid and efficient data collection.∗ Corresponding author at: Department of Veterinary Medicine in Nanjing Agri-ultural University, Nanjing 210095, PR China. Tel.: +86 25 8439530;ax: +86 25 84398669.
E-mail address: [email protected] (Q. Chen).
ttp://dx.doi.org/10.1016/j.micron.2014.03.010968-4328/© 2014 Elsevier Ltd. All rights reserved.
© 2014 Elsevier Ltd. All rights reserved.
Spermatogenesis in zebrafish has been studied by relatively fewauthors (Leal et al., 2009; Rupik et al., 2011; Schulz et al., 2010).Leal et al. (2009) distinguished the types and the generation ofgerm cells, and divided spermiogenesis into three stages based onincreasing nuclear compaction, Rupik et al. (2011) presented thebasic ultrastructural characteristics of spermiogenesis of zebrafishand, in contrast to Leal et al. (2009), divided spermiogenesis intofour stages.
Although some aspects are now well described, there are noreports providing complete ultrastructural descriptions of the pro-cesses of spermatogenesis, proliferation and meiosis. There are also
some important uncertainties regarding primary events, such aswhen the initial flagellum forms, how the chromatin becomes con-centrated within spermatids and so on. These are related to the finalstructure of the spermatozoa, the dynamics of gene expression and
2 Micron 66 (2014) 1–8
tti2
sfcmcs
2
Suaa(acwsVa
BewuFti
Fig. 1. The testis of zebrafish composed of numerous milky and tortuous seminif-
FEs
L. Zhang et al. /
he mechanisms underlying loss of optimal functionality, such ashe reduced motility that occurs when zebrafish embryos developn the presence of environmental toxicants (McAllister and Kime,003).
The present study addresses the fine structural changes in thepermatogenic cells of zebrafish, with particular focus on flagellarormation, chromatin condensation, appearance of nuclear vesi-les, distribution of nuages, and the change and arrangements ofitochondria. These results will be helpful for understanding the
orrelative functional and structural observations in this modelpecies.
. Materials and methods
Sexually mature male zebrafish were collected from thehanghai Laboratory Animal Research Center. They were rearednder controlled photoperiod (daily range: 14–10 h LD), temper-ture (daily range: 28–30 ◦C) and pH (7.0–7.8) in a recirculatingquaculture system. They were fed twice daily with dry food pelletsprotein > 38.5%). The fishes were taken at random from the tank,nd euthanized with an overdose of the anesthetic MS-222 (tri-aine methane sulfonate, 150 mg/L). (Ethics Statement: All effortsere made to minimize animal’s suffering. The slaughtering and
ampling procedures were approved by the Nanjing Agriculturaleterinary College Experimental Animal ethics committee. Thepproval ID is SYXK (SU) 2010-0005.)
Three male zebrafish, removed heads and tails, were fixed inouin’s solution overnight, embedded in paraffin wax, and thentire fishes serially sectioned (at 3 �m). The sections were stainedith hematoxylin and eosin for light microscopic observation,
sing an Olympus microscope (BX53), camera (Olympus DP73).or transmission electron microscopy (TEM), five pairs of con-ralateral testes were removed, cut into small blocks, then fixedn a mixture of 2.5% (v/v) glutaraldehyde in phosphate bufferedig. 2. The histological structure of testis in zebrafish, H&E stain. (a) Transverse sectionD, efferent ducts; I, intestine; SGA, type A spermatogonia; SGB, type B spermatogonia;permatids; T, seminiferous tubule. Scale bars: 20 �m (a) and 10 �m (b–d).
erous tubules (arrow), viewed under the dissecting microscope. Scale bar: 1 mm.
saline (PBS; 4 ◦C, pH 7.4, 0.1 M) for 24 h, post-fixed in 1% (w/v)osmium tetroxide in the same buffer for an hour, dehydrated inascending concentrations of ethyl alcohol, and infiltrated with apropylene oxide-Araldite mixture for embedding in Araldite (Zhanget al., 2007). The blocks were sectioned, and the ultrathin sec-tions (50 nm) were mounted on Formvar-coated grids, stained withuranyl acetate and lead citrate for 20 min per step. The sections
were examined using a HITACHI H-7650 transmission electronmicroscope.of the testis. (b–d) Spermatocysts containing different germ cells. Abbreviations: SS1, primary spermatocytes; SS2, secondary spermatocytes; SP, spermatozoa; ST,

L. Zhang et al. / Micron 66 (2014) 1–8 3
Fig. 3. Ultrastructure of spermatogonia in zebrafish, TEM. (a) A single type A spermatogonium with nucleus surrounded by nuages and round cristae-poor mitochondria. (b)Double type A spermatogonia in spermatocyst. (c) Two type A spermatogonia in mitosis metaphase connected by a cytoplasmic bridge and intercellular junctions. (d) Type Bspermatogonia containing fewer nuage. (e) Nuages accumulating near nucleus and appearing in the perinuclear space, and a mitochondrion attaching to nuclear membranesthrough nuage in spermatogonia A. (f) Dense material appears in the perinuclear space (double arrow). (g) The cytoplasmic bridge of spermatogonia composed of multilayerp nstruC e; Nu2
3
3
iltt(ccmtm(l(
arallel membranes (asterisk) in mitosis metaphase. (h) The cytoplasmic bridge coB, cytoplasmic bridges; Ch, chromosomes; Mi, mitochondria; N, nucleus; Na, nuag
�m (b–d), 1 �m (a), 500 nm (e, f, and h), and 200 nm (g).
. Results
.1. Histological and anatomical study of spermatogenesis
The testes of zebrafish are paired and elongated organs. Dur-ng the breeding season, the testes are about 0.5–0.6 cm inength and of white, cylindrical, and smooth appearance. Underhe dissecting microscope, seminiferous tubules were observedo be milky and tortuously distributed throughout the testisFig. 1). The germ cells are surrounded by the cytoplasmic pro-esses of Sertoli cells to form spermatocysts (Fig. 2a). Germells in spermatocysts showing basically synchronous develop-ent and various stages of cytological development (type A and
ype B spermatogonia, primary spermatocytes, secondary sper-
atocytes, spermatids) and mature spermatozoa are observedFig. 2b–d). The spermatozoa are eventually released into the tubu-ar lumen and finally pooled and collected by the efferent ductsFig. 2a).
cted by unilaminar cell membrane between type B spermatogonia. Abbreviations:, nucleolus; SC, Sertoli cell; SF, spindle fibers; IJ, intercellular junctions. Scale bars:
3.2. Ultrastructural changes of spermatogenic cells
3.2.1. SpermatogoniaType A spermatogonia, single or grouped in 2 or 4, represent
a weakly stained feature, with one or two nucleoli. A lot of elec-tron dense material appears in cytoplasm (Fig. 3a and b); this wascalled “nuage” (Billard, 1984). They are in close opposition to theedges of the nuclear membranes and connected with round andcristae-poor mitochondria (Fig. 3a and b). The nuclear membranesadjacent to nuages depress inward, and dense material appears inthe perinuclear space (Fig. 3e and f).
During mitotic metaphase, type A spermatogonia have the char-acteristics of a typical metaphase cell, displaying chromosomescombining with spindle fibers and arranging on the metaphase
plate (Fig. 3c). A small number of nuages are diffused into thecytoplasm. Spermatogonia are connected with each other by inter-cellular junctions and cytoplasmic bridges which are strengthenedby multilayer membranes (Fig. 3c and g).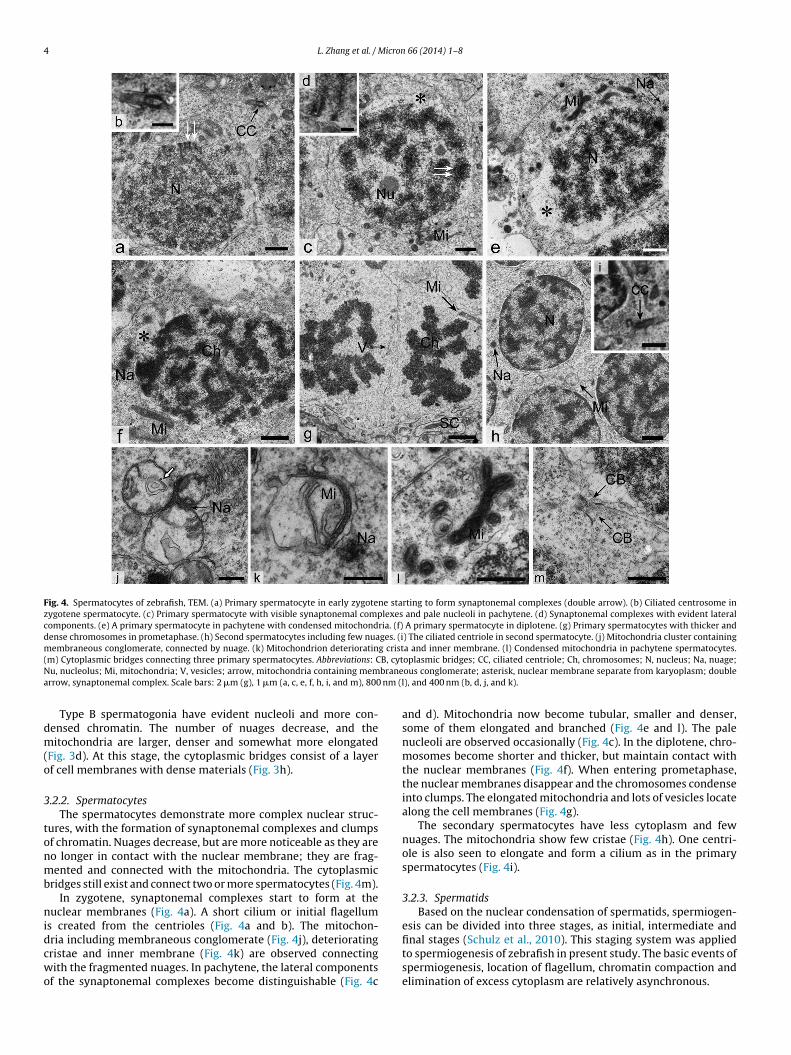
4 L. Zhang et al. / Micron 66 (2014) 1–8
Fig. 4. Spermatocytes of zebrafish, TEM. (a) Primary spermatocyte in early zygotene starting to form synaptonemal complexes (double arrow). (b) Ciliated centrosome inzygotene spermatocyte. (c) Primary spermatocyte with visible synaptonemal complexes and pale nucleoli in pachytene. (d) Synaptonemal complexes with evident lateralcomponents. (e) A primary spermatocyte in pachytene with condensed mitochondria. (f) A primary spermatocyte in diplotene. (g) Primary spermatocytes with thicker anddense chromosomes in prometaphase. (h) Second spermatocytes including few nuages. (i) The ciliated centriole in second spermatocyte. (j) Mitochondria cluster containingmembraneous conglomerate, connected by nuage. (k) Mitochondrion deteriorating crista and inner membrane. (l) Condensed mitochondria in pachytene spermatocytes.( B, cytN branea nm (l
dm(o
3
tonmb
nidcwo
m) Cytoplasmic bridges connecting three primary spermatocytes. Abbreviations: Cu, nucleolus; Mi, mitochondria; V, vesicles; arrow, mitochondria containing memrrow, synaptonemal complex. Scale bars: 2 �m (g), 1 �m (a, c, e, f, h, i, and m), 800
Type B spermatogonia have evident nucleoli and more con-ensed chromatin. The number of nuages decrease, and theitochondria are larger, denser and somewhat more elongated
Fig. 3d). At this stage, the cytoplasmic bridges consist of a layerf cell membranes with dense materials (Fig. 3h).
.2.2. SpermatocytesThe spermatocytes demonstrate more complex nuclear struc-
ures, with the formation of synaptonemal complexes and clumpsf chromatin. Nuages decrease, but are more noticeable as they areo longer in contact with the nuclear membrane; they are frag-ented and connected with the mitochondria. The cytoplasmic
ridges still exist and connect two or more spermatocytes (Fig. 4m).In zygotene, synaptonemal complexes start to form at the
uclear membranes (Fig. 4a). A short cilium or initial flagellums created from the centrioles (Fig. 4a and b). The mitochon-
ria including membraneous conglomerate (Fig. 4j), deterioratingristae and inner membrane (Fig. 4k) are observed connectingith the fragmented nuages. In pachytene, the lateral componentsf the synaptonemal complexes become distinguishable (Fig. 4c
oplasmic bridges; CC, ciliated centriole; Ch, chromosomes; N, nucleus; Na, nuage;ous conglomerate; asterisk, nuclear membrane separate from karyoplasm; double), and 400 nm (b, d, j, and k).
and d). Mitochondria now become tubular, smaller and denser,some of them elongated and branched (Fig. 4e and l). The palenucleoli are observed occasionally (Fig. 4c). In the diplotene, chro-mosomes become shorter and thicker, but maintain contact withthe nuclear membranes (Fig. 4f). When entering prometaphase,the nuclear membranes disappear and the chromosomes condenseinto clumps. The elongated mitochondria and lots of vesicles locatealong the cell membranes (Fig. 4g).
The secondary spermatocytes have less cytoplasm and fewnuages. The mitochondria show few cristae (Fig. 4h). One centri-ole is also seen to elongate and form a cilium as in the primaryspermatocytes (Fig. 4i).
3.2.3. SpermatidsBased on the nuclear condensation of spermatids, spermiogen-
esis can be divided into three stages, as initial, intermediate and
final stages (Schulz et al., 2010). This staging system was appliedto spermiogenesis of zebrafish in present study. The basic events ofspermiogenesis, location of flagellum, chromatin compaction andelimination of excess cytoplasm are relatively asynchronous.
L. Zhang et al. / Micron 66 (2014) 1–8 5
Fig. 5. Spermatids in initial stage of zebrafish, TEM. (a) The chromatin of spermatids concentrating asymmetrically and forming granular chromatin. (b) The centriolesapproaching toward lateral of nuclei where formed shallow nuclear fossa. (c) Initial segment of flagellum, formation of cytoplasmic canal and pristine cytoplasmic sheath. (d)The distal centriole partially wrapped by nucleus membrane. (e) The centrioles arranging at obtuse-angulate to each other, about 125 deg. (f) The posterior part of proximalc nteriot in; Cym centri
ts(nnt1mflcTcmpj
entriole (cross section) incompletely surrounded by nucleus membrane. (g) The ahe initial segment of flagellum. Abbreviations: CB, cytoplasmic bridge; Ch, chromat
itochondria; N, nucleus; NM, nuclear membrane; NF, nuclear fossa; PC, proximal
In the initial stage, the most notable changes are location ofhe flagellum and formation of the nuclear fossa. The chromo-omes begin concentrating asymmetrically and becoming granularFig. 5a). The centrosome approaches toward the lateral of ovalucleus, where nuclear membrane invaginates and forms a shallowuclear fossa (Fig. 5b–g). Two centrioles change their orienta-ion to each other from perpendicular to obtuse-angulate, about25 deg (Fig. 5e). Meanwhile, the cytoplasmic canal and cytoplas-ic sheath emerges in this stage. The cell membrane around the
agellum demonstrates inward depression, forming cytoplasmicanal, which separates the flagellum from the cytoplasm (Fig. 5c).he thin cytoplasm that surrounds the cytoplasmic canal forms a
ytoplasmic sheath around the cytoplasm canal. The mitochondriaove toward the initial segment of the flagellum (Fig. 5h). The cyto-lasmic bridges are more obvious, but the number of intercellularunctions appear to be reduced (Fig. 5b).
r part of proximal centriole inserted into nuclear fossa. (h) Mitochondria aroundC, cytoplasmic canal; CS, cytoplasmic sheath; DC, distal centriole; F, flagellum; Mi,ole. Scale bars: 1 �m (a, b, and h), 500 nm (f), and 300 nm (c–e and g).
Main events in the intermediate stage include chromatin con-centration and elimination of excessive nuclear membrane intocytoplasm. The chromatin in the nucleus become concentratedand compacted into chromosome clumps sharply, but with largeelectron-lucent areas in their central regions, round or tubularopening outward. (Fig. 6a–e). Denser mitochondria gather alongthe nuclear and cell membranes in linear arrangements, adjacentto the initial segment of the flagellum (Fig. 6b and c). The redundantnuclear membranes are discharged into the cytoplasm throughvesicles (Fig. 6b and e). With the progress of chromatin concen-tration, the size of electron-lucent areas in the nucleus diminish.At the end of this stage, the nucleus shows uniform electron-dense
chromatin except for 2–3 round large nuclear vacuoles (Fig. 7a andb). There is also a great deal of cytoplasm at this stage.During the final stage, spermatids exclude redundant cytoplasmand become separated from each other. Spermatids are separated

6 L. Zhang et al. / Micron 66 (2014) 1–8
Fig. 6. Spermatids in intermediate stage of zebrafish, TEM. (a) Spermatids showing nonuniform concentration of chromatin with electron-lucent areas in central, round ortubular opening outward. (b) Spermatids with mitochondrial linear gathering along the nuclear and cell membrane and a laterally located flagellum. (c) Cross section throughthe initial segment of flagellum, note flagellum lying in cytoplasmic canal and mitochondria arranging along the nuclear membrane. (d) Mitochondria close to nucleus. (e)Spermatid eliminating nuclear membrane in the form of vesicles. Abbreviations: CB, cytoplasmic bridges; Ch, chromatin; CS, cytoplasmic sheath; CyC, cytoplasmic canal;E ining
v
bwnbfcm
4
ummpepts
sicgtaSt
L, electron-lucent area; F, flagellum; Mi, mitochondria; arrow, mitochondria contaesicles; asterisk, intercellular gap. Scale bars: 2 �m (a and b) and 1 �m (c–e).
y the fracture of cytoplasmic bridges and intercellular junctions,hile excess cytoplasm is gradually removed (Fig. 7c). Inside theucleus the condensation of chromatin makes the nuclear vacuolesecome smaller (Fig. 7c and d). Finally, spermatogenesis finishes,orming mature spermatozoon with a round non-acrosomic headontaining small nuclear vacuoles, a middle piece with giantitochondria and a laterally located flagellum (Fig. 7c and e).
. Discussion
The process of spermatogenesis is very conserved in vertebrates,ndergoing spermatogonia proliferation and differentiation, sper-atocyte meiosis and maturation of spermatids, however, there isore diversity in anamniotes (fish, amphibia), for forming the mor-
hology of the species-specific spermatozoon to adapt to aqueousnvironments (Pudney, 1995). In this study, we found the generalrocess of spermatogenesis in zebrafish is similar to other teleosts,hat is consistent with previous reports (Leal et al., 2009), however,ome peculiar features previously unknown were found.
Unexpectedly the cilia or initial flagellae growing from centro-omes were observed in primary spermatocytes of the zebrafishn present study. In most teleosts, the flagellum has widely beenonsidered to form at or soon after the beginning of spermio-enesis. However, the initial flagellum has only been detected in
eleost spermatogonia and spermatocytes in a few species, suchs Oryzias latipes (Grier, 1976), Poecilia reticulate (Billard, 1984),orubim lima (Quagio-Grassiotto and Carvalho, 1999). Formation ofhe initial flagellum before the beginning spermiogenesis has alsomembraneous conglomerate; N, nucleus; PC, proximal centriole; SC, Sertoli cell; V,
been reported in invertebrates as well, such as Dermatobia homi-nis (Quagio-Grassiotto and De Lello, 1996) and Helcton pellucidus(Sousa and Oliveira, 1994). The formation of initial flagella beforespermiogenesis may represent a real difference between species orit may perhaps be an essential feature of germ cells.
During spermiogenesis, the mode of chromatin condensationvaries in teleosts, probably determined by the form of nuclearproteins present (Saperas et al., 1993). The chromatin of sper-matids becomes homogeneously compacted from fine granulesinto coarser granules or clusters in characidae (Veríssimo-Silveiraet al., 2006), golden gray mullet Liza aurata risso (Brusl, 1981),Piaractus mesopotamicus (De Cruz-Landim et al., 2003), blueheadThalassoma bifasciatum (Koulish et al., 2002) and cichlid fish(Fishelson, 2003), while in some teleosts, the chromatin com-paction is not homogeneous. Some authors have reported thatchromatin becomes concentrated from the periphery toward thecenter, forming thick filaments, as arranged in Hoplias malabari-cus (Quagio-Grassiotto et al., 2001) and Coreius heterodon (Zheng,1997). In the spermatids of zebrafish, chromatin condensationwas heterogeneous from granular into chromosome clumps withelectron-lucent areas in central, which later formed homogeneouscompact head of spermatozoa with small nuclear vacuoles.
The nuclear vacuoles are non-membranous cavities in thenuclei of spermatids or spermatozoa, with a variable size and
number (Czaker, 1985; Fawcett, 1958). They have been demon-strated in various mammalian spermatozoa, such as, bovine(Saacke and Almquist, 1964), guinea pig (Ohtomo, 1981) and mice(Czaker, 1985). They could also be observed in some teleosts
L. Zhang et al. / Micron 66 (2014) 1–8 7
Fig. 7. Spermatids in intermediate stage, final stage and spermatozoa of zebrafish, TEM. (a) Later spermatids in intermediate stage, containing lot cytoplasm and compactchromatin with large round nuclear vacuoles. (b) A later spermatid in intermediate stage, with a large round nuclear vacuole and pristine cytoplasmic sheath. (c) Spermatidsin final stage separated from each other within spermatocyst (upper half of the schematic), and spermatozoa in lumen of seminiferous tubule (lower half). (d) A spermatidi . (e) Mn ctronS bars:
llsnHwnfsn2it
tcmctMimim
n final stage, with a laterally located flagellum and cytoplasm around the nucleusuclear vacuoles. Abbreviations: CS, cytoplasmic sheath; DC, distal centriole; EL, eleertoli cell; SP, spermatozoa; ST, spermatids; T, lumen of seminiferous tubule. Scale
ike, P. mesopotamicus (De Cruz-Landim et al., 2003) and gold-ined seabream Rhabdosargus sarda (Liu and Yang, 2004). In humanpermatozoa, nuclear vacuoles were considered to probably origi-ate from defects of the chromatin condensation (Fawcett, 1958).owever, in zebrafish, this seems to be a normal structure. Theyere generally found in spermatids and spermatozoa. And theuclear vacuoles of zebrafish spermatozoa were confirmed coming
rom heterogeneous concentrated chromatin during spermiogene-is. This is probably a result of zebrafish sperm genome packaged inucleosomes and histone variants while not protamine (Wu et al.,011). Chen (2000) suggested that the nuclear vacuoles play a role
n reducing mechanical stress during vigorous movements of fer-ilization.
Morphology and distribution of mitochondria is variable, andhey can fuse and separate to adapt to variable functional state ofells (Bereiter-Hahn and Vöth, 1994; Westermann, 2012). In nor-al rats germ cells, at least three types of mitochondria, usual
ristae type (‘orthodox-type’), intermediate type and condensedype demonstrating a high oxidative capacity are detected (De
artino et al., 1979). The condensed mitochondria also appeared
n the pachytene spermatocytes of the zebrafish. In addition, manyitochondria including membrane conglomerate were observedn both spermatocytes and spermatids. Similar morphological
itochondria have been detected in germ cells of the mouse and
ature spermatozoon containing few cytoplasmic in cytoplasmic sheath and small-lucent area; F, flagellum; Mi, mitochondria; N, nucleus; NV, nuclear vacuoles; SC,
2 �m (a and c), 1 �m (b and d), and 500 nm (e).
was presumed following by release of mitochondrial molecules(Reunov, 2006). This type of mitochondria as well as the mitochon-drion deteriorating cristae and inner membrane may be eliminatedthrough residual body, for zebrafish mature spermatozoa reservingonly a limited number of mitochondria. During spermiogene-sis, mitochondria of spermatids can fuse and finally form giantmitochondria within the midpiece of spermatozoa in Salminus(Veríssimo-Silveira et al., 2006). In spermatids of zebrafish, themitochondria became linearly arranged along the nucleus and cellmembrane, gathering and forming giant mitochondrion at onset ofthe flagellum.
Nuages, a kind of electron-dense materials in the cytoplasm,somewhat like chromatin, specially exist in the embryonic andgerm cells (Braat et al., 1999; Reunov, 2006). They were reportedas only existing in the spermatogonia of zebrafish (Rupik et al.,2011). However, in the present study, nuages were found bothin spermatogonia and spermatocytes. These have been presumedto arise from both the nucleus and mitochondria in sea urchin(Reunov et al., 2000). Both interpretations may be true because,as shown in this study, in zebrafish, the nuages were both con-
nected with mitochondria and found in the perinuclear space. Theorigin of nuage of zebrafish seems to be closely related to nuclei andmitochondria from ultrastructural findings. They were reported tobe appeared in spermatocytes and spermatids of Wistar albino rats,
8 Micron
a2mnt
pIcsGettmtGzpowaz
ohdcmtmc
A
Ursa3
R
B
B
B
B
C
C
C
D
D
F
L. Zhang et al. /
nd were inferred to be a mRNA degradation site (Haraguchi et al.,005). In this study, a lot of nuages emerged at the edge of sper-atogonia nuclei, moving toward the cytoplasm and reduced the
umber in the spermatocytes, means maintaining its function tillhe meiosis stage of spermatocytes in zebrafish.
The cytoplasmic bridges between germ cells result from incom-lete cytokinesis during mitosis and meiosis (Fawcett et al., 1959).
n zebrafish, the cytoplasmic bridges in general germ cells areonstructed by unilaminar cell membranes, as though showingtrengthening during the mitotic division, as in S. lima (Quagio-rassiotto and Carvalho, 1999) and H. malabaricus (Cunha Negrãot al., 2002). These intercellular structures represent a way forhe spermatids to share organelles (Ventelä et al., 2003), pro-ein (Morales et al., 1998) and some genetic products such as
RNA (Morales et al., 2002). They are also presumably relatedo the synchronization of germ cell development (Zamboni andondos, 1968). In this study, more than two germ cells ofebrafish were frequently observed to be connected by a cyto-lasmic bridge (Figs. 4m and 5b). It indicated that the groupsf germ cells were not linear arranged and had side branches,hich may lead to more convenient and rapid communication
mong germ cells, to adapt to the short spermatogenic cycle ofebrafish.
In conclusion, this study presents a morphological descriptionf zebrafish spermatogenesis and describes some observations thatave not previously been noted. These new findings, especially theistribution of nuages, formation of flagellum, the special mode ofhromosome condensation in spermatid and the arrangement ofitochondria supplement the existing information, and will con-
ribute to further comparisons with other species, providing normalorphological evidence for the pathology and reproductive toxi-
ology studies of this model species.
cknowledgments
We are grateful to William V. Holt (University of Sheffield,K) for his help and critical comments on the manuscript. The
esearch was supported by Science and Technology Commis-ion of Shanghai Municipality (Grant Number: 11140902300)nd the National Science Foundation of China (Grant Number:1272521).
eferences
ereiter-Hahn, J., Vöth, M., 1994. Dynamics of mitochondria in living cells: shapechanges, dislocations, fusion, and fission of mitochondria. Microsc. Res. Tech.27, 198–219.
raat, A.K., Zandbergen, T., Van De Water, S., Goos, H.J.T., Zivkovic, D., 1999. Charac-terization of zebrafish primordial germ cells: morphology and early distributionof vasa RNA. Dev. Dyn. 216, 153–167.
riggs, J.P., 2002. The zebrafish: a new model organism for integrative physiology.Am. J. Physiol. Regul. Integr. Comp. Physiol. 282, R3–R9.
rusl, S., 1981. Ultrastructure of spermiogenesis in Liza aurata Risso, 1810 (Teleostei,Mugilidae). Cell Tissue Res. 217, 415–424.
hen, D., 2000. Impregnation Biology – Impregnation Mechanism and ProcreationEngineering, first ed. Science Press, Beijing, China.
unha Negrão, J.N., Carvalho, E.D., Foresti, F., Quagio-Grassiotto, I., 2002. Sper-matogonia and spermatocyte ultrastructure in Hoplias malabaricus (Teleostei,Characiformes: Erythrinidae). J. Zool. 257, 287–293.
zaker, R., 1985. On the origin of nuclear vacuoles in spermatozoa a fine structuraland cytochemical study in mice. Andrologia 17, 547–557.
e Cruz-Landim, C., Camargo Abdalla, F., De Cruz-Höfling, M.A., 2003. Morphologicalstudy of the spermatogenesis in the teleost Piaractus mesopotamicus. Biocell 27,319–328.
e Martino, C., Floridi, A., Marcante, M., Malorni, W., Barcellona, P.S., Bel-
locci, M., Silvestrini, B., 1979. Morphological, histochemical and biochemicalstudies on germ cell mitochondria of normal rats. Cell Tissue Res. 196,1–22.awcett, D.W., 1958. The structure of the mammalian spermatozoon. Int. Rev. Cytol.7, 195–234.
66 (2014) 1–8
Fawcett, D.W., Ito, S., Slautterback, D., 1959. The occurrence of Intercellular bridgesin groups of cells exhibiting synchronous differentiation. J. Biophys. Biochem.Cytol. 5, 453–460.
Fishelson, L., 2003. Comparison of testes structure, spermatogenesis, and sperma-tocytogenesis in young, aging, and hybrid cichlid fish (Cichlidae, Teleostei). J.Morphol. 256, 285–300.
Grier, H.J., 1976. Sperm development in the teleost Oryzias latipes. Cell Tissue Res.168, 419–431.
Haraguchi, C.M., Mabuchi, T., Hirata, S., Shoda, T., Hoshi, K., Akasaki, K., Yokota, S.,2005. Chromatoid bodies: aggresome-like characteristics and degradation sitesfor organelles of spermiogenic cells. J. Histochem. Cytochem. 53, 455–465.
Hill, A.J., 2005. Zebrafish as a model vertebrate for investigating chemical toxicity.Toxicol. Sci. 86, 6–19.
Koulish, S., Kramer, C.R., Grier, H.J., 2002. Organization of the male gonad in a pro-togynous fish, Thalassoma bifasciatum (Teleostei: Labridae). J. Morphol. 254,292–311.
Leal, M.C., Cardoso, E.R., Nobrega, R.H., Batlouni, S.R., Bogerd, J., Franca, L.R., Schulz,R.W., 2009. Histological and stereological evaluation of zebrafish (Danio rerio)spermatogenesis with an emphasis on spermatogonial generations. Biol. Reprod.81, 177–187.
Liu, X.Z., Yang, W.X., 2004. The fine structure of the spermatozoon of teleost Rhab-dosargus sarda (Forskai). Donghai Mar. Sci. 22, 43–48.
McAllister, B.G., Kime, D.E., 2003. Early life exposure to environmental levels of thearomatase inhibitor tributyltin causes masculinisation and irreversible spermdamage in zebrafish (Danio rerio). Aquat. Toxicol. 65, 309–316.
Morales, C.R., Lefrancois, S., Chennathukuzhi, V., El-Alfy, M., Wu, X.Q., Yang, J.X.,Gerton, G.L., Hecht, N.B., 2002. A TB-RBP and Ter ATPase complex accompaniesspecific mRNAs from nuclei through the nuclear pores and into intercellularbridges in mouse male germ cells. Dev. Biol. 246, 480–494.
Morales, C.R., Wu, X.Q., Hecht, N.B., 1998. The DNA/RNA-binding protein, TB-RBP,moves from the nucleus to the cytoplasm and through intercellular bridges inmale germ cells. Dev. Biol. 201, 113–123.
Norton, W., Bally-Cuif, L., 2010. Adult zebrafish as a model organism for behaviouralgenetics. BMC Neurosci. 11, 90.
Ohtomo, K., 1981. Ultrastructural changes in the spermatid nucleolus duringspermiogenesis in the guinea pig. Gamete Res. 4, 387–394.
Pudney, J., 1995. Spermatogenesis in nonmammalian vertebrates. Microsc. Res. Tech.32, 459–497.
Quagio-Grassiotto, I., Carvalho, E.D., 1999. The ultrastructure of Sorubim lima(Teleostei, Siluriformes, Pimelodidae) spermatogenesis: premeiotic and meioticperiods. Tissue Cell 31, 561–577.
Quagio-Grassiotto, I., De Lello, E., 1996. Centriole behaviour during meiosis of malegerm cells of Dermatobia hominis (Diptera: Cuterebridae). Cytobios 341, 73–80.
Quagio-Grassiotto, I., Negrao, J.N.C., Carvalho, E.D., Foresti, F., 2001. Ultrastructure ofspermatogenic cells and spermatozoa in Hoplias alabaricus (Teleostei, Characi-formes, Erythrinidae). J. Fish Biol. 59, 1494–1502.
Reunov, A., 2006. Structures related to the germ plasm in mouse. Zygote 14, 231–238.Reunov, A., Isaeva, V., Au, D., Wu, R., 2000. Nuage constituents arising from mitochon-
dria: is it possible? Dev. Growth Differ. 42, 139–143.Billard, R., 1984. Ultrastructural changes in the spermatogonia and spermatocytes
of Poecilia reticulata during spermatogenesis. Cell Tissue Res. 237, 219–226.Rupik, W., Huszno, J., Klag, J., 2011. Cellular organisation of the mature testes and
stages of spermiogenesis in Danio rerio (Cyprinidae; Teleostei) – structural andultrastructural studies. Micron 42, 833–839.
Saacke, R., Almquist, J., 1964. Ultrastructure of bovine spermatozoa. I. The head ofnormal, ejaculated sperm. Am. J. Anat. 115, 143–161.
Saperas, N., Ribes, E., Buesa, C., García-Hegart, F., Chiva, M., 1993. Differences in chro-matin condensation during spermiogenesis in two species of fish with distinctprotamines. J. Exp. Zool. 265, 185–194.
Schulz, R.W., de Franc a, L.R., Lareyre, J.-J., LeGac, F., Chiarini-Garcia, H., Nobrega, R.H.,Miura, T., 2010. Spermatogenesis in fish. Gen. Comp. Endocrinol. 165, 390–411.
Sousa, M., Oliveira, E., 1994. An ultrastructural study of spermatogenesis in Helcionpellucidus (Gastropoda, Prosobranchia). Invertebr. Reprod. Dev. 2, 119–125.
Traver, D., Herbomel, P., Patton, E.E., Murphey, R.D., Yoder, J.A., Litman, G.W., Catic,A., Amemiya, C.T., Zon, L.I., Trede, N.S., 2003. The zebrafish as a model organismto study development of the immune system. Adv. Immunol. 81, 254–330.
Ventelä, S., Toppari, J., Parvinen, M., 2003. Intercellular organelle traffic throughcytoplasmic bridges in early spermatids of the rat: mechanisms of haploid geneproduct sharing. Mol. Biol. Cell 14, 2768–2780.
Veríssimo-Silveira, R., Gusmão-Pompiani, P., Vicentini, C., Quagio-Grassiotto, I.,2006. Spermiogenesis and spermatozoa ultrastructure in Salminus and Brycon,two primitive genera in Characidae (Teleostei: Ostariophysi: Characiformes).Acta Zool. (Stockholm) 87, 305–313.
Westermann, B., 2012. Bioenergetic role of mitochondrial fusion and fission.Biochim. Biophys. Acta 1817, 1833–1838.
Wu, S.-F., Zhang, H., Cairns, B.R., 2011. Genes for embryo development are packagedin blocks of multivalent chromatin in zebrafish sperm. Genome Res. 21, 578–589.
Zamboni, L., Gondos, B., 1968. Intercellular bridges and synchronization of germ cell
differentiation during oogenesis in the rabbit. J. Cell Biol. 36, 276–282.Zhang, L., Han, X.K., Li, M.Y., Bao, H.J., Chen, Q.S., 2007. Spermiogenesis in soft-shelledturtle, Pelodiscus sinensis. Anat. Rec. 290, 1213–1222.
Zheng, S.M., 1997. Ultrastructural study on the spermatogenesis of Coreius heterodon.J. Hydroecol., 22–24 (in Chinese).
Copyright © 2022 FDOKUMEN




















