
APOE polymorphism affects episodic memory among non demented elderly subjects
Upload
univ-paris5Category
view
5download
0
Reliving Lifelong Episodic Autobiographical MemoriesVia the Hippocampus: A Correlative Resting PET
Study in Healthy Middle-Aged Subjects
Pascale Piolino,1,2 Beatrice Desgranges,1 Valerie Hubert,1 Frederic A. Bernard,3
Vanessa Matuszewski,1 Gael Chetelat,1 Jean-Claude Baron,4 and Francis Eustache1*
ABSTRACT: We aimed at identifying the cerebral structures whosesynaptic function subserves the recollection of lifetime’s episodic auto-biographical memory (AM) via autonoetic consciousness. Twelvehealthy middle-aged subjects (mean age: 59 years 6 2.5) underwent aspecially designed cognitive test to assess the ability to relive richlydetailed episodic autobiographical memories from five time periodsusing the Remember/Know procedure. We computed an index of episo-dicity (number of Remember responses justified by the recall of specificevents and details) and an index of retrieval spontaneity, and addition-ally an index of semanticized memories (number of Know responses).The regional cerebral blood flow (rCBF) was measured in the restingstate, with H2O
15 as part of an activation PET study. The indexes werecorrelated with blood flow using volumes of interest in frontotemporalregions, including hippocampus and voxel-wise analyses in SPM. Withboth analyses, significant correlations were mainly found between theindex of episodicity and rCBF in the medial temporal lobe, includinghippocampus, across the five time periods (unlike the index of semanti-cized memories) and between the spontaneity index and rCBF in theprefrontal areas. These results highlight, in healthy subjects, the distinctrole of these two structures in AM retrieval and support the view thatthe hippocampus is needed for reexperiencing detailed episodic memo-ries no matter how old they are. VVC 2008 Wiley-Liss, Inc.
KEY WORDS: autonoetic consciousness; self; retrieval; remember/know; frontotemporal lobe
INTRODUCTION
Despite numerous neuropsychological and functional neuroimagingstudies, the neurobiological bases of long-term episodic memory remaingreatly debated (Mc Gaugh, 2000; Meeter and Murre, 2004). Accordingto a standard model (Squire, 1992; Squire and Alvarez, 1995; Teng andSquire, 1999; Murre, 2001; Squire, 2004; Bayley et al., 2005), themedial temporal lobe (MTL) structures are involved in the storage and
retrieval of declarative memory (either semantic or ep-isodic) during a limited period of a few years. Thememory trace of a consciously experienced event issparsely encoded in a distributed network, includingthe hippocampus that acts as a pointer to neocorticalregions where memories are stored. Thus, the hippo-campal memory system would serve as a temporaryindex for the retrieval of myriad aspects of the mem-ory trace, whereas the neocortex would be the perma-nent repository of long-term memory. As time passesand the memories undergo rehearsal, the linksbetween the elements of memories gradually developwithin the neocortex and the links between the MTLand neocortex vanish. An alternative model, however,the multiple trace model, considers the standard viewis correct for semantic information and semanticizedmemories, but not for episodic memories (Nadel andMoscovitch, 1997; Nadel et al., 2000; Moscovitchet al., 2005). The capacity of the MTL to recollectepisodic memories would be permanent. As timepasses, new links would be created within the MTL,so that remote episodic memories are stored evermore in multiple traces, i.e., represented by more hip-pocampal–neocortical traces. The creation of multipletraces facilitates the extraction of the common featuresof episodes and forms semantic memories that areretrieved independently of hippocampus. This theorystates that the hippocampus is needed for reexperienc-ing detailed episodic memories no matter how oldthey are. Throughout a person’s life, the hippocampuswould be involved in the recollection of the variousparts of a particular episode, especially detailed con-textual information such as time, place, emotionalcontent, and perceptual features.
Given that the timescale involvement of the MTLin episodic memory constitutes the critical divergencebetween the theories of hippocampal and/or medialtemporal lobe–neocortical interaction in memoryretention and retrieval, the studies regarding autobio-graphical memory (AM) proved very valuable in orderto test the different theories. The current theory of ep-isodic memory emphasizes autobiographical referenceand phenomenological aspects of memory (Tulving,2001, 2002). The phenomenal experience of remem-bering (i.e., reliving of perceptual, affective, and spa-
1U923, Inserm-EPHE-Universite de Caen Basse Normandie, Unite 923,Caen, France; 2CNRS FRE 2987, Laboratoire de Psychologie et Neuro-sciences Cognitives, Universite Rene Descartes, Paris 5, France; 3 InsermU 610, Hopital de la Salpetriere, Paris, France; 4Department of Neurol-ogy, University of Cambridge, Addenbrooke’s Hospital, Cambridge,United Kingdom*Correspondence to: Francis Eustache, Inserm U923, Lab. de Neuropsycho-logie, CHU Avenue de la Cote de Nacre, 14033 Caen cedex, France.E-mail: [email protected] for publication 6 December 2007DOI 10.1002/hipo.20406Published online 31 January 2008 in Wiley InterScience (www.interscience.wiley.com).
HIPPOCAMPUS 18:445–459 (2008)
VVC 2008 WILEY-LISS, INC.

tiotemporal contextual details) gives the rememberer the feel-ing that a representation is the recollection of a specific eventthat belongs to her/his personal past instead of an item ofsemantic knowledge (Brewer, 1996; Baddeley, 2001; Gardiner,2001; Tulving, 2001, 2002). More specifically, autonoetic con-sciousness—that is the feeling of reexperiencing or reliving thepast and mentally traveling back in subjective time—is centralto current conception of episodic memory (Wheeler et al.,1997; Tulving, 2002). However, AM is formed of differenttypes of representations (Tulving et al., 1988; Conway, 2001),from general knowledge about oneself (i.e., semantic compo-nent) to very specific personal events (i.e., episodic compo-nent). The semantic component is characterized by a noeticstate of consciousness in which one is capable of retrieving thegeneral facts about personal events, without reexperiencingspecific contexts. The episodic component of AM containspersonal specific events with phenomenological details situatedin time and space pertaining to one’s self and presupposesvery lengthy retention intervals. Unlike the semantic compo-nent, it is characterized by an autonoetic state of conscious-ness, which enables a conscious recollection of a personalevent, in its original encoding context, and implies a mentaltime travel. However, with the passage of time and repetitionof similar events, there is a shift in the phenomenal experienceof AM from sense of remembering to sense of just knowing,i.e., from episodic to semantic memory (Conway et al., 1997;Piolino et al., 2006). This shift is in accordance with the ideathat most parts of the very long-term memories are semanti-cized throughout time (Cermak, 1984), being a mixture ofsemantic knowledge and specific experiences (see Brainerd andReyna, 2001). Thus, given the multifaceted nature of AM, itis necessary to have stringent methods of investigation, espe-cially in order to assess the existence of truly episodic memo-ries (Moscovitch et al., 2005), indicating not only the capacityto represent a specific event and locate it in time and space,but also the capacity to recollect some specific feature of thatevent (Levine et al., 2002; Piolino et al., 2002, 2006). Thenature of AMs can be assessed with the Remember/Know pro-cedure (see Method).
Neuropsychological studies of episodic autobiographical ret-rograde amnesia have contributed to better identify the cerebralstructures involved in AM (Conway and Fthenaki, 2000;Kopelman and Kapur, 2001; Greenberg and Rubin, 2003;Moscovitch et al., 2005). Some studies have documented twomain different profiles of retrograde amnesia depending on thelocus of the lesion: either a Ribot’s temporal gradient, charac-terized by a better preservation of remote relative to recentmemories, in patients with MTL damage (Bayley et al., 2003,2005; Squire and Bayley, 2007), or a reverse temporal gradient(or reverse step-like profile), characterized by an impairment ofremote memories with preserved recent ones in patients withdamage to the anterior temporal lobe with relative sparing ofthe hippocampal complex (Graham and Hodges, 1997; Nestoret al., 2002; Piolino et al., 2003a,b). This double dissociationwould support the standard model (Meeter and Murre, 2004).However, some other studies using stringent control for the
episodic nature of AMs have found floor performances with aflat gradient in patients with damage to the MTL, discrepantwith the standard consolidation theory, but more in agreementwith the multiple trace theory instead (Fujii et al., 2000; Vis-kontas et al., 2000; Cipolotti et al., 2001; Westmacott et al.,2001; Moscovitch et al., 2005; Noulhiane et al., 2007). Someof these studies have demonstrated the absence of episodicremote AM using the Remember/Know procedure (Westmacottet al., 2001; Hirano et al., 2002; Piolino et al., 2003a; Noul-hiane et al., 2007, but see Bayley et al., 2005). The lesion stud-ies have also shown that episodic AM depends on widely dis-tributed neocortical areas, especially the frontal and lateral tem-poral. Indeed, AM is disrupted by frontal and anteriortemporal lobe lesions or a form of frontotemporal disconnec-tion (for reviews, see Kapur, 2000; Kopelman, 2000; Wheelerand McMillan, 2001). The frequent involvement of the fronto-temporal areas has also been highlighted in episodic focal retro-grade amnesia (Calabrese et al., 1996; Levine et al., 1998; Pio-lino et al., 2005). Taken together, the neuropsychological find-ings suggest that frontotemporal neocortical areas, in additionto the hippocampus (Kopelman et al., 2003; Ferbinteanu et al.,2006), play a role in AM performance.
A number of functional neuroimaging studies have exploredthe activations produced by AM tasks in healthy subjects. Theyhave highlighted the major role of the prefrontal cortex (i.e.,medial and lateral PFC) in the AM reconstructive processes,more specially ventromedial PFC, which is one of the mostubiquitous sites of activation in AM neuroimaging studies(Gilboa, 2004). Besides, they have yielded controversial resultsabout involvement of the MTL structures according to thelength of the retention interval. Some studies (e.g., Niki andLuo, 2002; Piefke et al., 2003) have provided evidence for adifferential engagement of the hippocampal region in recentand remote AM retrieval, seemingly arguing in favor of thestandard model of memory consolidation. However, most ofthe other studies showed no differential involvement of theMTL according to time interval (Conway et al., 1999; Maguireet al., 2001; Ryan et al., 2001; Addis et al., 2004; Gilboaet al., 2004; Piolino et al., 2004; Steinvorth et al., 2006; Viardet al., 2007) in contradiction with the predictions from the‘‘standard model’’ but supporting instead the multiple tracetheory. In these two sets of studies, methodological confoundmay have biased the results. Thus, in most instances, a ques-tionnaire was administered a few weeks or days prior to thescanning session in order to obtain the material to constructthe cues used during scanning to reactivate old memoriestraces. Only a few studies have tried to prevent subjects fromreencoding their memories via the hippocampus before scan-ning either by collecting cues from family members (Gilboaet al., 2004; Viard et al., 2007) or by presenting personal rele-vant cues not previously encountered by subjects (Okuda et al.,2003; Piolino et al., 2004). Moreover, in activation studies, thecontrol of the nature of memory is a problematic issue, since itcan only be made at debriefing (e.g., Graham et al., 2003;Addis et al., 2004; Gilboa et al., 2004; Piolino et al., 2004).When subjects recall memories during scanning, they inevitably
446 PIOLINO ET AL.
Hippocampus

encode them once again (Persson and Nyberg, 2000; Buckneret al., 2001). Hence, the hippocampal activation for long reten-tion intervals might reflect the processing of reencoding andnot the access per se to old memories, although some studieshave attempted to this phenomenon by including a controlcondition that involves reencoding (e.g., Conway et al., 1999;Gilboa et al., 2004). Otherwise, it is usually difficult to disen-tangle generation from hold-in-mind processes during scanning(Conway et al., 2002). Finally, the studies are generally con-ducted on young subjects in whom autobiographical memoriesare not very remote, as long retention intervals, over decades,cannot be examined.
Apart from the activation paradigm, another powerfulapproach entails obtaining, within a short time interval (a fewdays at most), both cognitive test scores and resting-state brainmetabolism using PET, and performing correlations betweenthese two sets of data across a group of subjects. PET allowsthe study of physiological parameters such as blood flow whichis closely related to synaptic activity. It is a well-validatedmethod to map the functional neuroanatomy of a given behav-ior in neurodegenerative diseases (Desgranges et al., 1998,2002a,b; Eustache et al., 2001; Hirono et al., 2001; Lekeuet al., 2003a,b). Recently, by using measures of resting state18FDG-PET, Teipel et al. (2006) highlighted that the profile ofcortical regions that show altered metabolism was correlated tothe decline of specific cognitive functions within memory andlanguage in Alzheimer’s disease. More specially, we have usedthis approach in the field of autonoetic consciousness (Rauchset al., in press) and AM (Eustache et al., 2004). It is particu-larly useful for establishing cognitive and neurobiological mod-els of human memory, because it allows the mapping of specificbrain regions that are involved in a particular task performanceand provide complementary data to the ‘‘classic’’ activation par-adigm that unravel the cerebral networks implicated in a task(see Tulving et al., 1999; Eustache et al., 2000, for a discussionon the concepts of ‘‘how sites’’ and ‘‘what sites’’). More particu-larly, the correlative method offers an alternative approach thatfrees one from some methodological limits in AM neuroimag-ing studies, enabling the use of sophisticated cognitive assess-ments where the nature of memories can be strictly controlled,which is difficult in activation paradigms. In healthy subjects,the logic of this cognitive–metabolic approach in AM is basedon the variability concerning both cognitive performance(Levine et al., 2002; Piolino et al., 2002, 2006) and brainfunctions (Fransson, 2005; Coles et al., 2006). In this context,this approach is particularly potent in middle-aged and oldhealthy subjects who present an increasing in intersubjectvariability compared to young subjects (Eustache et al., 1995;Piefke and Fink, 2005; Coles et al., 2006). Moreover, thestudy of older subjects allows examination of the correlationbetween resting-state brain measures and thorough cognitivemeasures of AM for very long retention intervals, some lastingdecades.
The aim of the study, therefore, was to reveal the cerebralstructures whose synaptic function subserves the recollection oflifetime’s episodic AM using a correlation approach between
resting regional cerebral blood flow (rCBF) with PET and sepa-rately obtained AM indexes regarding episodicity (i.e., the rec-ollection of specific events with episodic details) and spontane-ity (i.e., the retrieval of memories without retrieval support).We assessed healthy middle-aged subjects able to remember oldmemories across five time periods covering their entire lifespanprincipally to test the two competitive theories of the role ofthe hippocampus in episodic memory retrieval. Given theircritical role in AM (Conway et al., 2002), we expected cor-relations with prefrontal and anterior temporal regions (i.e.,frontotemporal hypothesis) and MTL including hippocampus.Referring to the multiple trace theory, we mainly predicted apermanent involvement of the hippocampus regardless of thelife-time period in reliving episodic memories (i.e., episodicindex). Regarding the spontaneity of retrieval (i.e., spontaneityindex), we mainly predicted the involvement of prefrontalregions, more especially lateral and medial prefrontal areasbased on their special role in strategic retrieval search process-ing and self-referential functions.
MATERIALS AND METHODS
Subjects
We studied 12 volunteers (six males, six females) of meanage 59 yr (6 2.5; range: 55–63 yr). They were thoroughlyscreened to rule out the presence of medical, neurological orpsychiatric disorders, or dementia. In order to avoid includingmiddle-aged subjects with incipient dementia, we used theMattis Dementia Rating Scale in which all the subjectsobtained high scores on (mean 5 141.25 6 2.9). All werenative French speakers and right-handed as determined by theEdinburgh Handedness Inventory (LQ 5 95.58 6 6.07). Theywere unmedicated, had no memory complaint and had no sig-nificant abnormality on their T1 and T2-weighted magneticresonance images (magnetic resonance imaging (MRI)). Allgave written consent prior to participation and the researchprotocol was approved by the Regional Ethics Committee. Thestudy was done in line with the Declaration of Helsinki. Thisstudy was embedded in an activation PET study on new verbalepisodic learning (Bernard et al., 2007).
Experimental AM Protocol
Episodic AM was assessed according to a specially designedtask (TEMPau, Piolino et al., 2003a,b, 2005, 2006), whichtakes into account the phenomenal experience of recollection(i.e., autonoetic consciousness). The subjective phenomenalexperience can be assessed via the Remember/Know (R/K)paradigm (Tulving, 1985; Gardiner, 1988), which requires sub-jects to give either a ‘‘Remember’’ (R) response, if retrieval isaccompanied by the recollection of specific experiences presentat encoding, or a ‘‘Know’’ (K) response, if retrieval is achievedwithout access to information from the initial learning context.
RELIVING EPISODIC AUTOBIOGRAPHICAL MEMORIES VIA HIPPOCAMPUS 447
Hippocampus

The TEMPau consists of a semistructured questionnairewhich assesses the ability to recall detailed specific events situ-ated in time and space from five time periods (period 1: 0- to17-yr old, period 2: 18- to 30-yr old, period 3: more than30-yr old except for the last 5 years, period 4: last 5 yr exceptfor the last 12 months, period 5: last 12 months) covering theentire lifespan. There is no time limit but the test generallylasts 2 h.
Each time period examined, with the exception of the mostrecent one, is assessed on four topics (a meeting or an eventlinked to a person, a school and then a professional event, atrip or journey, a familial event), with instructions such as‘‘Give details of a particular event which took place duringyour family life.’’ For the most recent period, eight questionsallow a chronological study of the previous 12 months (e.g.,last summer, last weekend, yesterday). Again, subjects are giveninstructions such as ‘‘Give details of a particular event whichoccurred last week.’’ If a participant cannot spontaneously re-collect a specific event, cues are provided (for example, ‘‘on aday with a teacher or friend,’’ ‘‘in primary or secondaryschool,’’ ‘‘in the school playground,’’ or ‘‘during an exam’’),while he or she is encouraged to be specific if the memory isgeneric (e.g., do you remember a particular day?). After threecueing and/or encouragement attempts, the tester switches toanother topic.
Following the recall of each event, the subjects have to indi-cate whether or not they can consciously recollect its prioroccurrence, by making separate Remember/Know judgmentsfor the content, place, and date of the event (what, where andwhen respectively). They can also indicate if they have simplyguessed the recalled event (Mantyla, 1993). It is important tonote that the judgment categories (Conway et al., 1997;Gardiner, 2001) are explained very carefully to the participantsuntil each concept used in this test is thoroughly understood.
The general framework of the test is presented in Figure 1.First of all, the subjects are instructed to select one of these cat-egories (i.e., Remember, Know, or Guess) with reference to thefactual content (what) of the recalled event. They are theninstructed to select the appropriate categories for the place andfor date of the event. For example, the participants can make aRemember response concerning a striking meeting (i.e., what Rresponse) that took place in a particular environment (i.e.,where R response) 2 yr ago (i.e., when R response). Given thatthe kind of response is selected independently for the threecontents (what, where, and when), the participants can give forinstance, a Remember response for the factual content, a Knowresponse for the spatial content, and a Guess response for thetemporal content. Of note, in healthy subjects Know or Guessresponses mainly concern the temporal content of memories,this feature going up with the remoteness (Piolino et al.,2006).
Lastly, a procedure is performed, to check whether the sub-jects have correctly recalled a specific event (i.e., unique, situ-ated in time and space) and can justify each of their R judg-ments. Accordingly, for each R response provided whatever thekind of information (what, where and when), the subjects have
to add contextual details from the original event, if they havenot already provided them spontaneously during the memoryrecall task: specific event with phenomenal details such as theirthoughts, feelings, or perceptions for the content (what), spatialdetails and location for the place (where), and time of day ortemporal sequence for the date (when). An example of a consis-tently recollected episodic AM (i.e., R response justified interms of episodic detail) is that the participant relives the spe-cific atmosphere, his/her feelings and thoughts (justified what Rresponse), he/she recollects very specific visual details of thescene, his/her position in the scene (justified where R response),and he/she recollects the thoughts he/she has with regard to thefact that the meeting took place exactly 5 yr after his ownschool graduation and the moment of the day (justified whenR response).
Scoring
As the aim of our study was to investigate strictly episodicautobiographical memory, we focused the present study on twodifferent scores:
An index of episodicity (1) per time period, corresponding tothe total number of R responses provided taken together allkinds of information (what, where, when), justified in terms ofactual episodic details retrieved (max: 3 3 4 5 12; i.e., threejustified R responses per topic and four topics per period) and
FIGURE 1. The framework of the TEMPau and scoresrecorded. For each of the five lifetime periods, the TEMPauassesses the state of consciousness via the Remember/Know para-digm, and the ability to justify the Remember responses. Aftereach recalled event, regardless of the spontaneity level, the partici-pants estimate their state of consciousness accompanying the re-trieval, with separate assessment for the retrieval of the factual,spatial, and the temporal contents. When the participants give anR response, they must add sensory-perceptual and contextualdetails. A spontaneity index and an episodicity index are recorded(see score) to assess, respectively, (1) the total number of cues and/or encouragements to be specific that the subject required and(2) the total number of justified R responses irrespective of thecontent or according to the content.
448 PIOLINO ET AL.
Hippocampus

(2) combining all time periods, corresponding to the totalnumber of R responses provided separately according to thekind of information (what, where, when), justified in terms ofactual episodic details retrieved (max: 4).
An index of spontaneity was also recorded per time periodexamined. For each memory, the spontaneity conversely addedup the number of cues and/or encouragements to be specificthat the subject required using a four-point scale (max. 4 34 5 16). For example, the spontaneous retrieval of a specificevent was given a score of 4 points, whereas retrieval achievedafter three cues and/or prompts was given a score of 1.
In order to test the specificity of results concerning the epi-sodic index, we also recorded an index of semanticized memories(number of K responses) (1) all time period combined and (2)according to time periods (P1–P5).
Each score for the last 12 months period, which concernedeight items instead of four topics, was halved. Two independentexperts rated each memory until a consensus was reached.
PET Study
Within an interval of few days, each subject underwent theAM assessment and a PET measurement. The rCBF was meas-ured in the resting state with H2O
15 as part of an activationPET study (Bernard et al., 2007). Each subject underwent 12consecutive scans (injections of H2O
15) during a single PETsession lasting �2 h and 30 min. The PET session was per-formed with an intravenous catheter placed in the left arm forthe administration of H2O
15. Rest was replicated four timesduring the protocol. Subjects were scanned while lying supinein a darkened and quiet room. During the Rest condition, theywere instructed to relax, to keep their eyes closed and not tofocus their mind on a precise thought, and were not subject toany stimulation. A black tent was set up all around the PETtomograph, to ensure that the subjects were in total darkness.The head was gently immobilized in a dedicated head-rest.Head position was aligned transaxially to the orbitomeatal linewith a laser beam. Measurements of regional distribution ofradioactivity were performed with an ECAT HR1 (SIEMENS)PET camera with full volume acquisition allowing the recon-struction of 63 planes (thickness: 2.4 mm; axial field-of-view:158 mm; effective resolution is about 4.2 mm in all direc-tions). Transmission scans were obtained with a 68Ga sourceprior to emission scans. The duration of each scan was 90 s.About 7 mCi of H2O
15 were administered as a slow bolus inthe left antecubital vein by means of an automated infusionpump.
PET Data Analysis
All calculations and image transformations were performedon UNIX SYSTEM workstations. First, the four rest scans dur-ing rest of each subject were realigned to each other, using theAIR 3.0 software (Woods et al., 1998). For subsequent dataanalysis, the Statistical Parametric Mapping software (SPM99,
Welcome department of Cognitive Neurology, UK) imple-mented in the MATLAB environment was used. A customizedtemplate was obtained by averaging the normalized andsmoothed structural T1-MRI of each subject. The images werenonlinearly transformed into the customized template from theMRI data and then they were smoothed using a 12-mm Gaus-sian filter. The images were scaled to an overall CBF grandmean of 50 ml/100 g/min; we therefore refer to ‘‘adjustedrCBF’’ in what follows. Anatomical localization was accordingto both the labelized atlas (‘‘toolbox aal’’) implementedin SPM99 and developed by Tzourio-Mazoyer et al. (2002)and Talairach’s Atlas (Talairach and Tournoux, 1988), usingM. Brett’s set of linear transformations (see http://www.mrc-cbu.cam.ac.uk/Imaging/mnispace.html).
First, with respect to a priori hypotheses of involvement ofthe frontotemporal neocortical structures and hippocampalregion in AM (see Introduction), we extracted the mean rCBF(corresponding to the mean adjusted rCBF from the four restscans) for each anatomical volume of interest (VOIs) selectedfrom a bilateral set of 13 VOIs covering the left and right fron-tal and temporal lobes (i.e., a total of 26 VOIs). These 13VOIs in each hemisphere were extracted from the automatedanatomical labeling atlas (aal labels, Tzourio-Mazoyer et al.,2002)—in the lateral surface of the frontal lobe (superior fron-tal dorsolateral, middle frontal, inferior frontal operculum, infe-rior triangular frontal, i.e., main Brodmann Areas: 6/8/9/44/45)—in the orbitomedial surface of the frontal lobe (superiormedial frontal, superior medial orbital frontal, superior orbitalfrontal, middle orbital frontal, inferior orbital frontal, i.e.,main BA: 10/11/47), and—in the temporal lobe (superior tem-poral pole, middle temporal pole, i.e. main BA: 21/22/38)including hippocampus and parahippocampus.
Second, we used a voxel-based analysis of the whole brainusing an exploratory approach (see later).
Statistical Analyses
We first performed an analysis of variance (ANOVA) withpost hoc tests (Bonferroni test) on the behavioral measures toassess the effect of periods on the indexes of episodicity andspontaneity. Then, we carried out forward stepwise regressionsanalyses between the performance scores obtained with theTEMPau task and the mean rCBF obtained during rest for the13 VOIs selected in the right and left sides, using a thresholdof P < 0.05. Finally, we looked for correlations between thecognitive scores and resting CBF using SPM-99. The influenceof age was controlled by setting age as a confounding variablein a single linear regression. For each score, only the correla-tions in the neurobiologically expected direction (that is, posi-tive, i.e., any decrease in performance was predicted to relate toa fall, and not an increase, in CBF) were assessed, using a sta-tistical threshold (uncorrected for multiple tests) of P < 0.005.The results were obtained in two forms: (1) projection of thesignificant voxels onto a customized template; (2) peaks withtheir MNI coordinates.
RELIVING EPISODIC AUTOBIOGRAPHICAL MEMORIES VIA HIPPOCAMPUS 449
Hippocampus
RESULTS
Behavioral Data
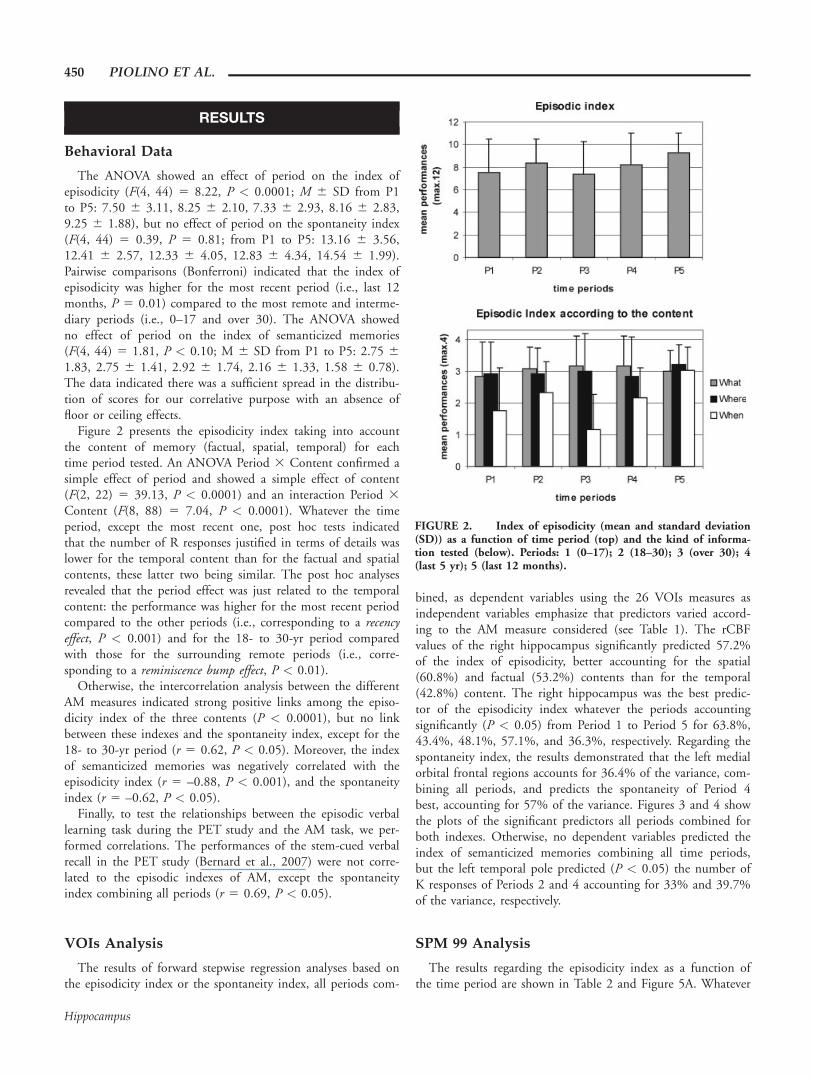
The ANOVA showed an effect of period on the index ofepisodicity (F(4, 44) 5 8.22, P < 0.0001; M 6 SD from P1to P5: 7.50 6 3.11, 8.25 6 2.10, 7.33 6 2.93, 8.16 6 2.83,9.25 6 1.88), but no effect of period on the spontaneity index(F(4, 44) 5 0.39, P 5 0.81; from P1 to P5: 13.16 6 3.56,12.41 6 2.57, 12.33 6 4.05, 12.83 6 4.34, 14.54 6 1.99).Pairwise comparisons (Bonferroni) indicated that the index ofepisodicity was higher for the most recent period (i.e., last 12months, P 5 0.01) compared to the most remote and interme-diary periods (i.e., 0–17 and over 30). The ANOVA showedno effect of period on the index of semanticized memories(F(4, 44) 5 1.81, P < 0.10; M 6 SD from P1 to P5: 2.75 61.83, 2.75 6 1.41, 2.92 6 1.74, 2.16 6 1.33, 1.58 6 0.78).The data indicated there was a sufficient spread in the distribu-tion of scores for our correlative purpose with an absence offloor or ceiling effects.
Figure 2 presents the episodicity index taking into accountthe content of memory (factual, spatial, temporal) for eachtime period tested. An ANOVA Period 3 Content confirmed asimple effect of period and showed a simple effect of content(F(2, 22) 5 39.13, P < 0.0001) and an interaction Period 3Content (F(8, 88) 5 7.04, P < 0.0001). Whatever the timeperiod, except the most recent one, post hoc tests indicatedthat the number of R responses justified in terms of details waslower for the temporal content than for the factual and spatialcontents, these latter two being similar. The post hoc analysesrevealed that the period effect was just related to the temporalcontent: the performance was higher for the most recent periodcompared to the other periods (i.e., corresponding to a recencyeffect, P < 0.001) and for the 18- to 30-yr period comparedwith those for the surrounding remote periods (i.e., corre-sponding to a reminiscence bump effect, P < 0.01).
Otherwise, the intercorrelation analysis between the differentAM measures indicated strong positive links among the episo-dicity index of the three contents (P < 0.0001), but no linkbetween these indexes and the spontaneity index, except for the18- to 30-yr period (r 5 0.62, P < 0.05). Moreover, the indexof semanticized memories was negatively correlated with theepisodicity index (r 5 –0.88, P < 0.001), and the spontaneityindex (r 5 –0.62, P < 0.05).
Finally, to test the relationships between the episodic verballearning task during the PET study and the AM task, we per-formed correlations. The performances of the stem-cued verbalrecall in the PET study (Bernard et al., 2007) were not corre-lated to the episodic indexes of AM, except the spontaneityindex combining all periods (r 5 0.69, P < 0.05).
VOIs Analysis
The results of forward stepwise regression analyses based onthe episodicity index or the spontaneity index, all periods com-
bined, as dependent variables using the 26 VOIs measures asindependent variables emphasize that predictors varied accord-ing to the AM measure considered (see Table 1). The rCBFvalues of the right hippocampus significantly predicted 57.2%of the index of episodicity, better accounting for the spatial(60.8%) and factual (53.2%) contents than for the temporal(42.8%) content. The right hippocampus was the best predic-tor of the episodicity index whatever the periods accountingsignificantly (P < 0.05) from Period 1 to Period 5 for 63.8%,43.4%, 48.1%, 57.1%, and 36.3%, respectively. Regarding thespontaneity index, the results demonstrated that the left medialorbital frontal regions accounts for 36.4% of the variance, com-bining all periods, and predicts the spontaneity of Period 4best, accounting for 57% of the variance. Figures 3 and 4 showthe plots of the significant predictors all periods combined forboth indexes. Otherwise, no dependent variables predicted theindex of semanticized memories combining all time periods,but the left temporal pole predicted (P < 0.05) the number ofK responses of Periods 2 and 4 accounting for 33% and 39.7%of the variance, respectively.
SPM 99 Analysis
The results regarding the episodicity index as a function ofthe time period are shown in Table 2 and Figure 5A. Whatever
FIGURE 2. Index of episodicity (mean and standard deviation(SD)) as a function of time period (top) and the kind of informa-tion tested (below). Periods: 1 (0–17); 2 (18–30); 3 (over 30); 4(last 5 yr); 5 (last 12 months).
450 PIOLINO ET AL.
Hippocampus
the time period, the positive correlation (Z values) concernedthe hippocampus, either bilaterally (Periods 1 and 2) or unilat-erally—right-sided (Periods 3 and 4) or —left-sided (Period 5).The correlations also concerned a network including the para-hippocampus, precuneus, lingual gyri, and thalamus, regardlessof time periods. Moreover, for some Periods (1, 3, and 4), thecorrelations involved some temporal and prefrontal neocorticalregions.
As regards the factual, spatial, or temporal content, the cor-relations concerned principally the right hippocampus/parahip-pocampus as well as right middle temporal gyrus. The numberof voxels and Z score were highest for the spatial content (Ta-ble 3, Fig. 5B).
Concerning the spontaneity index (Table 4, Fig. 6), themain correlations (Z scores and cluster size) concerned the pre-frontal cortex bilaterally, more specially the orbitomedial surface(ventromedial part of BA10; medial parts of BA11 and 47),except for Period 2 which involved the left hippocampus.
To deal with the issue of the specificity of the link betweenperfusion of hippocampus and the episodic index, we looked atthe correlations with the index of semanticized memories as afunction of the time period and combining all periods. Therewas no significant correlation, neither for only one period norfor all combined period (data not shown). We also looked atthe correlations between the baseline PET scans and the verbalmemory performances (stem-cued recall) at retrieval in thePET study (Bernard et al., 2007). At the same threshold as forthe AM data (P < 0.005, k > 30), there was no correlation inthe MTL (data not shown).
DISCUSSION
The main aim of the present study was to examine inhealthy subjects the underlying cerebral regions of correlationbetween episodic AM retrieval and resting rCBF.
TABLE 1.
Summary of Forward Stepwise Regression Analysis Using the Selected 26 VOIs Which Were Entered Into and Kept in
Predictive Model for Episodic Index or Spontaneity Index Combined on All Periods
Step Dependent variable Predictive independent variable R2 F(P) df
1 Episodic index R hippocampus 0.572 13.34** 1,11
1 What-EI R hippocampus 0.532 11.37** 1,11
1 Where-EI R hippocampus 0.608 15.52** 1,11
1 When-EI R hippocampus 0.464 8.67* 1,11
1 Spontaneity index L medial orbital frontal 0.364 5.72* 1,11
Caption for the independent variables entered: right and left superior frontal dorsolateral, middle frontal, inferior frontal operculum, inferior triangular frontal,superior medial frontal, superior medial orbital frontal, superior orbital frontal, medial orbital frontal, inferior orbital frontal, hippocampus, parahippocampus,superior temporal pole, middle temporal pole. The R2 indicates the total proportion of variance mediated by significant predictors. The F ratios show the signifi-cance of the total R2.*P < 0.05; **P < 0.01.
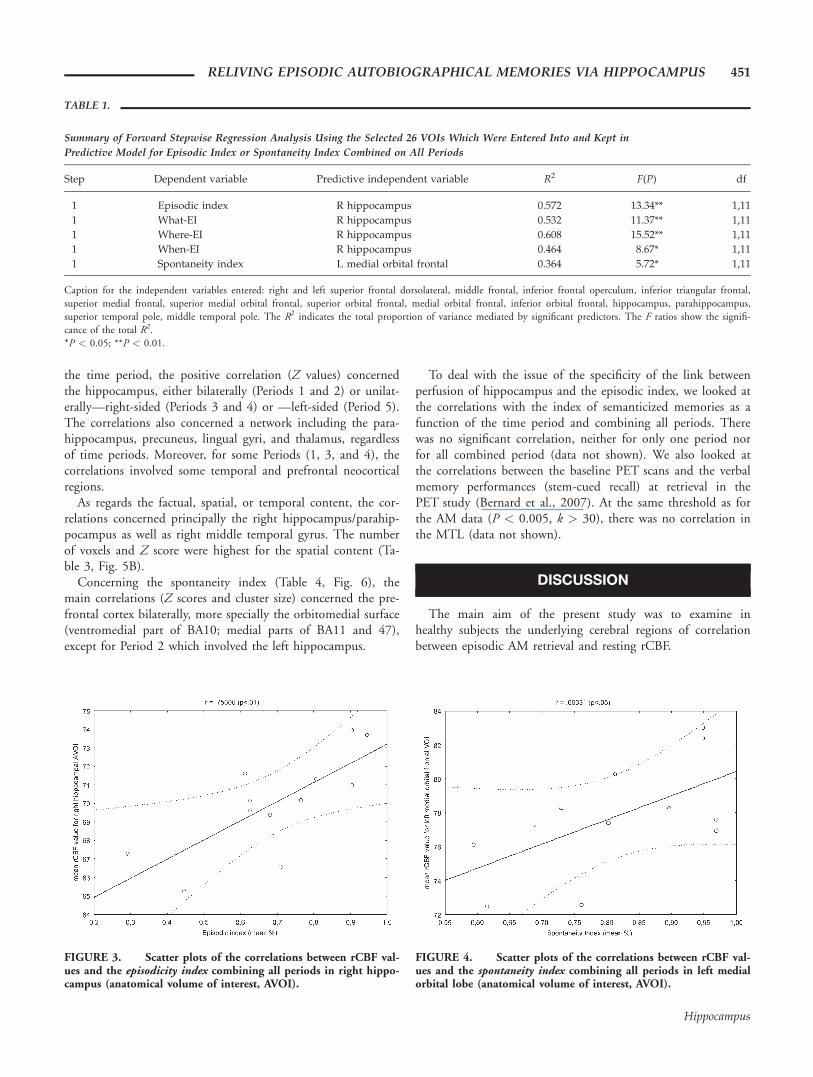
FIGURE 3. Scatter plots of the correlations between rCBF val-ues and the episodicity index combining all periods in right hippo-campus (anatomical volume of interest, AVOI).
FIGURE 4. Scatter plots of the correlations between rCBF val-ues and the spontaneity index combining all periods in left medialorbital lobe (anatomical volume of interest, AVOI).
RELIVING EPISODIC AUTOBIOGRAPHICAL MEMORIES VIA HIPPOCAMPUS 451
Hippocampus

As regards the behavioral data, the study showed that themajority of memories gave rise to a sense of rememberingwhich was justified in terms of specificity and details and theywere rather spontaneously retrieved regardless of the remote-ness. The temporal distribution of the episodicity index morespecially was characterized by a recency effect and a reminiscencebump effect. The presence of both effects in healthy middle-aged subjects corroborates the literature on AM (Rubin et al.,1998; Levine et al., 2002; Piolino et al., 2002) and confirmsthe consistency of these effects using strict criteria of assessmentbased on the remember/know paradigm (see also Piolino et al.,2006). More specially, the reminiscence bump phenomenonconcerns the rise of vivid memories from adolescence andyoung adulthood compared to surrounding remote periodswhich represent a strong landmark for the current self (Conwayand Pleydell-Pearce, 2000; Conway et al., 2004) and, thereafter,for maintaining a sense of identity and continuity in the pres-ent. The episodicity index applied in this study was particularlystringent as it was based on the subjective feeling of mentaltraveling back in time (i.e., R responses) checked for the abilityto relive specific events with details of encoding episodes (i.e.,phenomenal details such as their thoughts, feelings or percep-tions, and spatio-temporal details), indicating that the subjecttruly reexperienced the original event. The justified R responsesprovided for the temporal content were clearly less numerousthan for the factual and the spatial contents, a pattern already
observed in previous studies on healthy subjects regardless oftheir age (Piolino et al., 2006).
The stepwise regression analyses indicated that the rCBF val-ues of the right hippocampus solely predicted the index of epi-sodicity regardless of the age of memories, while that of the leftmedial orbital frontal gyrus solely predicted the index of spon-taneity. The results using SPM encompassing the whole brainconfirm these striking relationships between episodic AM andrCBF in the medial temporal regions including hippocampus,and between the spontaneity of access to specific memories andrCBF in frontal regions. Interestingly, the relationship betweenthe episodicity index and the hippocampus was regardless ofthe content of information, but more importantly for the spa-tial content. These findings are discussed in the framework ofAM and neuroimaging data.
Cerebral Sites of Correlation for Episodic IndexAccording to Time Period and Content
The involvement of hippocampus in autonoetic memories isin keeping with studies carried out in healthy subjects using ep-isodic learning tasks that suggested that MTL structures, nota-bly the hippocampus, may take part to this phenomenon (Ran-ganath et al., 2003). Most importantly, the link with the perfu-sion of the hippocampus whatever the time period arguesagainst the standard theory of consolidation (Squire and
TABLE 2.
Main Peaks of Significant (P < 0.005, Uncorrected, SPM Maps Thresholded at T > 3.17, Z > 2.58, Minimal Cluster Size k > 30 voxels)
Correlations Between Episodic Index as Function of Periods and rCBF
Periods
No. of
voxels Regions BA x y z Z
1 959 R. hippocampus/parahippocampus/lingual/ thalamus 27/30/35 18 232 2 3.68
77 R insula 48 34 30 6 3.33
86 L rectus/orbital frontal G 11 212 30 220 3.31
465 R superior polar temporal G/insula 48 50 24 2 3.26
77 L precuneus/B paracentral 5 0 240 64 3.23
175 L superior polar temporal G/inferior orbital frontal G 38 246 14 28 3.18
75 R middle temporal G 22 56 226 210 3.17
308 L hippocampus/ thalamus 27 214 228 4 3.12
40 R rolandic operculum 48 48 226 18 2.80
2 75 L hippocampus 37 224 234 22 3.80
220 R hippocampus/parahippocampus/lingual thalamus 27/30 14 234 210 2.91
3 60 R. superior orbital frontal G 11 214 30 220 3.99
489 R hippocampus/parahippocampus/lingual/thalamus 27 18 232 2 3.50
121 R middle/superior temporal G 20 54 226 210 3.14
4 897 R hippocampus/parahippocampus/lingual/
thalamus/superior frontal G
27/30 14 222 216 4.35
80 R rolandic operculum 48 52 214 20 2.94
45 L superior polar temporal G 38 252 10 214 2.91
5 38 R precuneus/cuneus/calcarine 23 10 264 24 3.17
32 L hippocampus 20 232 220 216 2.90
The clusters are listed in decreasing order of peak Z score. BA, Brodmann’s area; x, y, z, coordinates of peaks in Talairach’s system; L, left; R, right; G, gyrus.Periods: 1 (0–17); 2 (18–30); 3 (over 30); 4 (last 5 years); 5 (last 12 months).
452 PIOLINO ET AL.
Hippocampus

Alvarez, 1995), but supports the multiple trace theory instead(Nadel and Moscovitch, 1997; Fujii et al., 2000; Moscovitchet al., 2005). With respect to the latter theory, the findingssubstantiate that the hippocampus is involved in the mecha-nism that reactivates all the neocortical regions where the mul-tifaceted components of episodic memories are represented.Interestingly, using a similar correlative approach in a group ofpatients with mild-to-moderate Alzheimer’s disease, we foundan involvement of the right hippocampus restricted to the mostrecent time period studied (i.e., previous 5 yr) (Eustache et al.,2004). The findings were compatible with either the standardconsolidation theory or the multitrace theory (see introduction)as we showed that preserved remote memories were personalsemantic knowledge instead of episodic memories. Our presentresults substantiate the MTT as we found that strictly episodic
memories were dependent on the hippocampus whatever theirage unlike semanticized ones (i.e., associated to K responses).
The regression analyses stressed the role of the right hippo-campus whatever the period considered. Nevertheless, the cor-relations using SPM concerned the hippocampus either bilater-ally (i.e., the two most remote periods), or unilaterally (i.e.,right-sided for the over 30- and last 5-yr periods and left-sidedfor the last 12 months period). Yet, the issue of hippocampallaterality is still debated especially in AM. Gilboa et al. (2005),unlike Kopelman et al. (2003), found in patients with brainlesions a significant correlation between remote AM and theamount of remaining tissue in bilateral MTL, stronger on theright than the left, (see also, Noulhiane et al., 2007). Most ofthe neuroimaging studies on AM which showed a constantinvolvement of the hippocampus regardless of the age of mem-
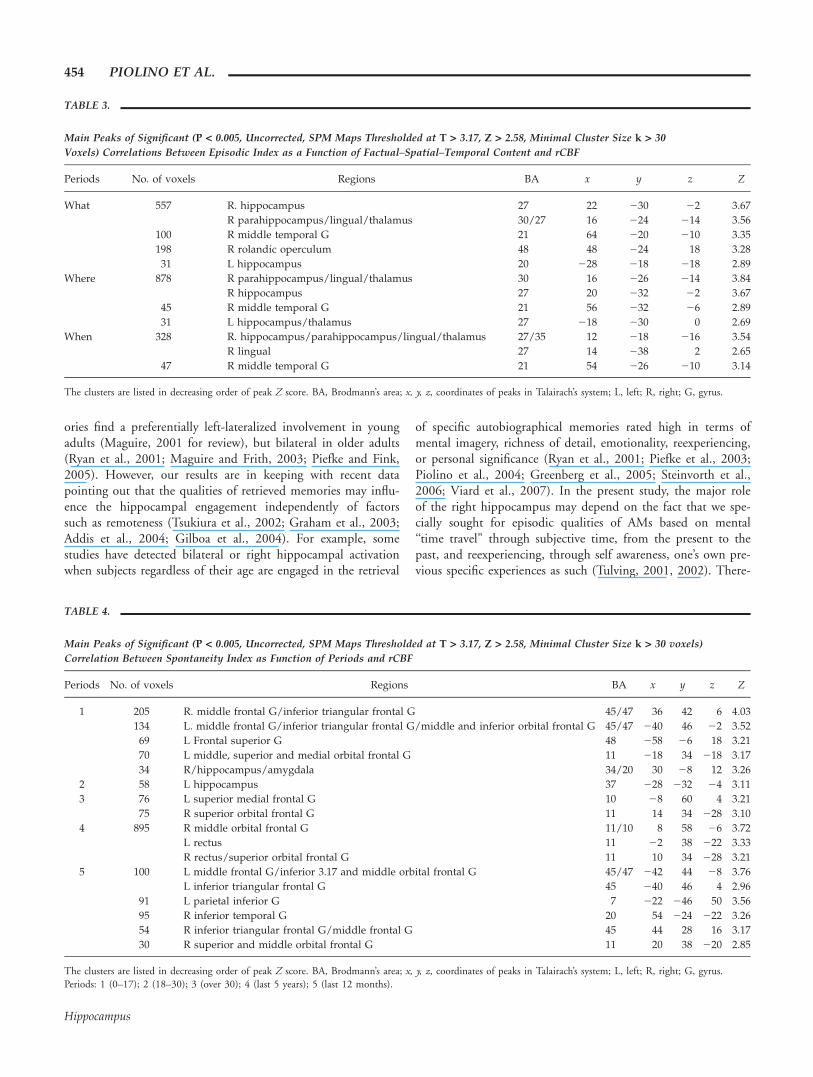
FIGURE 5. Results from SPM99 analysis: positive correlations(P < 0.005, uncorrected; SPM maps thresholded at Z > 2.58)between episodicity index and rCBF. For anatomical orientation,the significant correlations are shown as colored voxels superim-posed onto a customized template based on the whole group ofsubject. The right hemisphere corresponds to the right side of the
figure. Results are shown (A) according to time periods (P1–P5)and (B) according to contents (what, where, and when). The crossindicates the locus of the main correlation (Z values). Periods: 1(0–17); 2 (18–30); 3 (over 30); 4 (last 5 yr); 5 (last 12 months).[Color figure can be viewed in the online issue, which is availableat www.interscience.wiley.com.]
RELIVING EPISODIC AUTOBIOGRAPHICAL MEMORIES VIA HIPPOCAMPUS 453
Hippocampus
ories find a preferentially left-lateralized involvement in youngadults (Maguire, 2001 for review), but bilateral in older adults(Ryan et al., 2001; Maguire and Frith, 2003; Piefke and Fink,2005). However, our results are in keeping with recent datapointing out that the qualities of retrieved memories may influ-ence the hippocampal engagement independently of factorssuch as remoteness (Tsukiura et al., 2002; Graham et al., 2003;Addis et al., 2004; Gilboa et al., 2004). For example, somestudies have detected bilateral or right hippocampal activationwhen subjects regardless of their age are engaged in the retrieval
of specific autobiographical memories rated high in terms ofmental imagery, richness of detail, emotionality, reexperiencing,or personal significance (Ryan et al., 2001; Piefke et al., 2003;Piolino et al., 2004; Greenberg et al., 2005; Steinvorth et al.,2006; Viard et al., 2007). In the present study, the major roleof the right hippocampus may depend on the fact that we spe-cially sought for episodic qualities of AMs based on mental‘‘time travel" through subjective time, from the present to thepast, and reexperiencing, through self awareness, one’s own pre-vious specific experiences as such (Tulving, 2001, 2002). There-
TABLE 3.
Main Peaks of Significant (P < 0.005, Uncorrected, SPM Maps Thresholded at T > 3.17, Z > 2.58, Minimal Cluster Size k > 30
Voxels) Correlations Between Episodic Index as a Function of Factual–Spatial–Temporal Content and rCBF
Periods No. of voxels Regions BA x y z Z
What 557 R. hippocampus 27 22 230 22 3.67
R parahippocampus/lingual/thalamus 30/27 16 224 214 3.56
100 R middle temporal G 21 64 220 210 3.35
198 R rolandic operculum 48 48 224 18 3.28
31 L hippocampus 20 228 218 218 2.89
Where 878 R parahippocampus/lingual/thalamus 30 16 226 214 3.84
R hippocampus 27 20 232 22 3.67
45 R middle temporal G 21 56 232 26 2.89
31 L hippocampus/thalamus 27 218 230 0 2.69
When 328 R. hippocampus/parahippocampus/lingual/thalamus 27/35 12 218 216 3.54
R lingual 27 14 238 2 2.65
47 R middle temporal G 21 54 226 210 3.14
The clusters are listed in decreasing order of peak Z score. BA, Brodmann’s area; x, y, z, coordinates of peaks in Talairach’s system; L, left; R, right; G, gyrus.
TABLE 4.
Main Peaks of Significant (P < 0.005, Uncorrected, SPM Maps Thresholded at T > 3.17, Z > 2.58, Minimal Cluster Size k > 30 voxels)
Correlation Between Spontaneity Index as Function of Periods and rCBF
Periods No. of voxels Regions BA x y z Z
1 205 R. middle frontal G/inferior triangular frontal G 45/47 36 42 6 4.03
134 L. middle frontal G/inferior triangular frontal G/middle and inferior orbital frontal G 45/47 240 46 22 3.52
69 L Frontal superior G 48 258 26 18 3.21
70 L middle, superior and medial orbital frontal G 11 218 34 218 3.17
34 R/hippocampus/amygdala 34/20 30 28 12 3.26
2 58 L hippocampus 37 228 232 24 3.11
3 76 L superior medial frontal G 10 28 60 4 3.21
75 R superior orbital frontal G 11 14 34 228 3.10
4 895 R middle orbital frontal G 11/10 8 58 26 3.72
L rectus 11 22 38 222 3.33
R rectus/superior orbital frontal G 11 10 34 228 3.21
5 100 L middle frontal G/inferior 3.17 and middle orbital frontal G 45/47 242 44 28 3.76
L inferior triangular frontal G 45 240 46 4 2.96
91 L parietal inferior G 7 222 246 50 3.56
95 R inferior temporal G 20 54 224 222 3.26
54 R inferior triangular frontal G/middle frontal G 45 44 28 16 3.17
30 R superior and middle orbital frontal G 11 20 38 220 2.85
The clusters are listed in decreasing order of peak Z score. BA, Brodmann’s area; x, y, z, coordinates of peaks in Talairach’s system; L, left; R, right; G, gyrus.Periods: 1 (0–17); 2 (18–30); 3 (over 30); 4 (last 5 years); 5 (last 12 months).
454 PIOLINO ET AL.
Hippocampus
fore, our result may suggest that the right hippocampus con-tributes to the successful retrieval of episodic memories greatlydependent upon the richness of reexperiencing phenomenaland spatiotemporal details.
More specifically, our findings pointed out the major linkbetween perfusion in right hippocampus and spatial episodicrepresentations (O’Keefe and Nadel, 1978; Hartley et al.,2000; see Burgess et al., 2002) unlike schematic representationof spatial context as discussed by Moscovitch et al. (2005).Otherwise, while we might have expected the frontal lobe to beinvolved in temporal memory in reference with Nadel andMoscovitch (1997) and Kopelman et al. (1997), the pattern ofcorrelations also involved the hippocampus in keeping with itscritical role in memory for sequences of events (Fortin et al.,2002).
Our SPM findings also showed that the episodic index waslinked to the CBF of some other limbic structures (e.g., para-hippocampus) and several other structures consistent with AMneuroimaging studies (Maguire, 2001; Conway et al., 2002;Cabeza et al., 2004). In fact, regardless of the period, the corre-lations concerned regions involved in visual and spatial memory(i.e., parahippocampus and lingual/precuneus/cuneus/calcarine),highly implicated in autobiographical retrieval processing(Greenberg and Rubin, 2003). Interestingly, the parahippocam-pus was involved regardless of the remoteness (except the mostrecent period) consistent with the MTT model that suggeststhat various MTL regions including hippocampus may perma-nently contribute to the different aspects of episodic AM (Mos-covitch et al., 2005). Moreover, for most of the periods, thecorrelations also involved some PFC and temporal neocortical
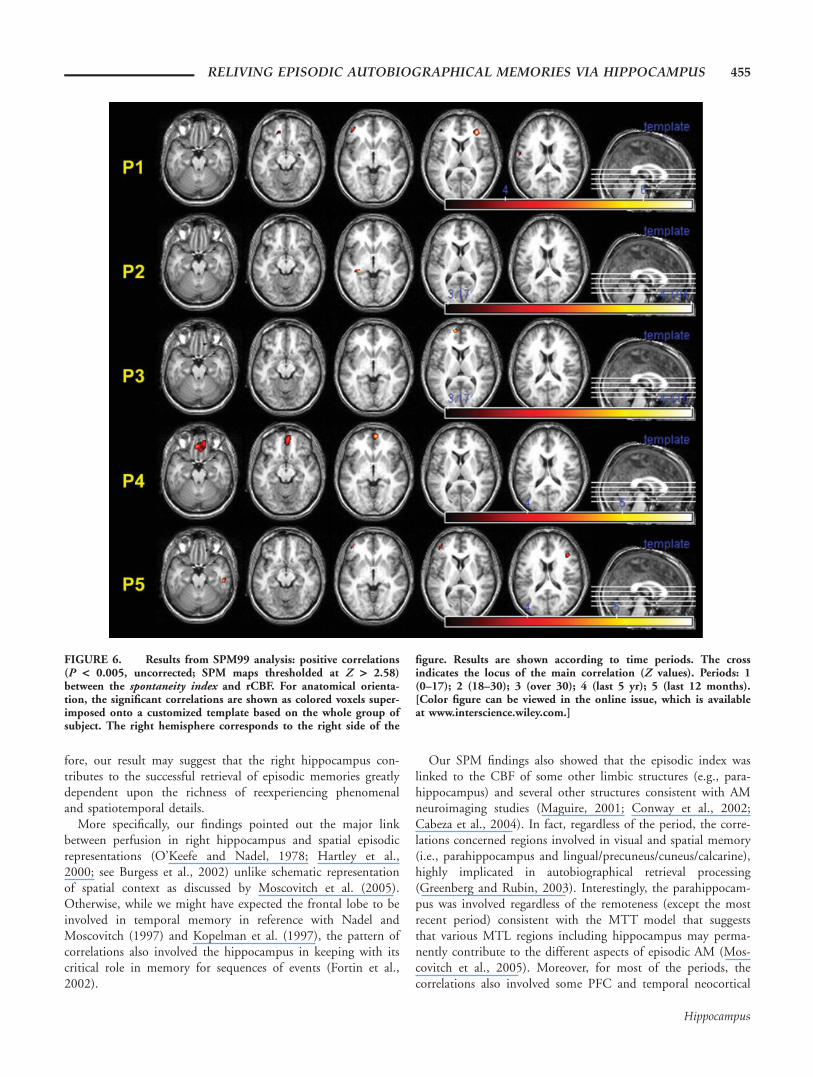
FIGURE 6. Results from SPM99 analysis: positive correlations(P < 0.005, uncorrected; SPM maps thresholded at Z > 2.58)between the spontaneity index and rCBF. For anatomical orienta-tion, the significant correlations are shown as colored voxels super-imposed onto a customized template based on the whole group ofsubject. The right hemisphere corresponds to the right side of the
figure. Results are shown according to time periods. The crossindicates the locus of the main correlation (Z values). Periods: 1(0–17); 2 (18–30); 3 (over 30); 4 (last 5 yr); 5 (last 12 months).[Color figure can be viewed in the online issue, which is availableat www.interscience.wiley.com.]
RELIVING EPISODIC AUTOBIOGRAPHICAL MEMORIES VIA HIPPOCAMPUS 455
Hippocampus

areas including the polar area (BA 38) in keeping with ourfrontotemporal hypothesis. The regions in PFC are concernedby self-referential (orbitomedial) and strategic retrieval processes(dorsolateral) while those in the temporal neocortex are entailedin self-conceptual knowledge which takes part in AM (Finket al., 1996; Conway et al., 2002). Of note, we found that Kresponses were predicted by the left temporal pole. The mostrecent and 18- to 30-yr-old periods were linked to a less exten-sive set of regions. Of note, for both periods the intersubjectvariability was relatively low compared to the other periods,which might be a plausible explanation for a less extensive setof correlations.
Cerebral Sites of Correlation for SpontaneityIndex According to Time Period
Concerning the spontaneity index, we expected the specialinvolvement of prefrontal regions. Both the stepwise regressionsand SPM analyses pinpointed the role of the left orbitomedialPFC cortex. Nevertheless using SPM, the pattern of correla-tions also concerned the lateral PFC (BA 45) and right orbito-medial PFC. The correlations with orbitomedial PFC are inkeeping with Gilboa’s review (2004) of the neuroimaging litera-ture which suggests that some subregions in left prefrontal cor-tex, particularly parts of the frontal pole and ventromedialaspects, may be activated preferentially during retrieval of auto-biographical as compared to other types of episodic memories.Interestingly, our study extends the previous data by emphasiz-ing that it may particularly depend on the spontaneity of re-trieval, that is, on the number of external cues and/or encour-agements to be specific that the subject required during genera-tive retrieval. The less spontaneous is a memory, thus the lessself-generated, the poorer perfusion in left orbitomedial PFC.Taken as a whole, the findings regarding the PFC are also inagreement with neuropsychological studies (Conway and Fthe-naki, 2000; Kopelman and Kapur, 2001). For example, themechanisms of AM retrieval were correlated to executive/frontaltasks in a group of traumatic brain injury patients (Piolinoet al., 2007a) or patients with frontotemporal dementia(Matuszewski et al., 2006) tested upon anterograde and retro-grade periods using the present AM task. We recently demon-strated that AM retrieval deficits in frontotemporal dementiawere mainly correlated with the dysfunction of metabolism inleft orbitofrontal lobe, regardless of the time period (Piolinoet al., 2007b). These data are consistent with the role of PFCas a working-with-memory structure that is involved in strate-gic aspects of retrieval such as establishing a retrieval modeand goals, initiating and guiding search, and monitoring andverifying the memories that have been retrieved (Moscovitchand Winocur, 2002; Badre and Wagner, 2004; Umeda et al.,2005).
According to Conway’s framework (Conway and Pleydell-Pearce, 2000; Conway, 2001; Conway et al., 2004), the recol-lection of episodic AMs that ensures a sense of rememberingrequires a complex strategic retrieval process depending on theprefrontal cortex, which allows access to sensory/perceptual
event-specific knowledge from personal semantic knowledge.The left PFC would be involved during the AM generativeprocess when triggering off episodic AM. In line with thismodel, the spontaneity of retrieval would depend on thecapacity of the subjects to self-generate (without external sup-port) a retrieval model in relation to self-goals representation.We suggest that the major role of the left orbitomedial PFC inspontaneity reflects the critical role of self-referential representa-tion (Craik, 1999; Frith and Frith, 1999; Northoff and Berm-pohl, 2004; Northoff et al., 2006) in AM generative process.Of note, we tested very few memories by period and thus, themost accessible ones, which could account in the present studyfor the relatively minor role of the lateral surface of the frontallobe involved in effortful strategic retrieval search processing(Petrides, 2000). However, intriguingly the retrieval spontaneityregarding the 18- to 30-yr-old period (reminiscence bump)which is a landmark for self-identity did not yield correlationin the orbitomedial prefrontal cortex, but in the hippocampusinstead. The access to those episodic memories probably mighthave been more dependent on direct retrieval processes (i.e.,ecphoric processes) and thus, relatively unconstrained by frontalregions.
In sum, our paradigm gives us the opportunity to single outtwo distinct aspects of AM retrieval by recording on the onehand the generative processes (i.e., spontaneity index) and onthe other hand the recollection based on autonoetic conscious-ness with reliving of episodic details (i.e., episodicity index).The complete set of data may be consistent with Conway’smodel. In effect, when the retrieval of autobiographical memo-ries is deliberate and intentional, as is the case in our AM test,the model predicts that the construction of consciously-recol-lected episodic memories involves frontal/executive processesrelated to current goals (i.e., the working self ), and a sense ofre-experiencing which emerges by linking all the componentsof a specific AM experience. Our study highlights that the orbi-tofrontal cortex would play a role in the working self whereasthe hippocampus would play a role in successful re-experienc-ing no matter how old the memories are. Finally, there is evi-dence that the ‘‘default network’’ engaged during passive taskstates or rest resembles to the brain network engaged when sub-jects are instructed to initiate self-referential mental processessuch as autobiographical reminiscing (Gusnard et al., 2001;Gusnard and Raichle, 2001; Raichle, 2001; Fransson, 2005;Northoff et al., 2006). This network prominently includesfrontal regions along the midline, a network of lateral andmedial parietal regions, and MTL structures (Buckner and Vin-cent, 2007). Thus, our results substantiate that it seems partic-ularly powerful to correlate AM measures and resting state inhealthy subjects to highlight some regions in this network thatare necessary to a specific task performance.
In conclusion, the findings reported in the present studyemphasize in healthy middle-aged subjects the permanent roleof the hippocampus in reexperiencing lifetime episodic detailsof AM relevant to MTT, and the role of the prefrontal cortexas a self working-with-memory structure that initiates andguides self-generated retrieval to trigger off memories.
456 PIOLINO ET AL.
Hippocampus

Acknowledgments
The authors thank for their assistance the cyclotron staff(GIP Cyceron of Caen) and the subjects who participated inthe protocol.
REFERENCES
Addis DR, Moscovitch M, Crawley AP, McAndrews MP. 2004. Recol-lective qualities modulate hippocampal activation during autobio-graphical memory retrieval. Hippocampus 14:752–762.
Baddeley AD. 2001. The concept of episodic memory. Philos Trans RSoc Lond B Biol Sci 356:1545–1550.
Badre D, Wagner AD. 2004. Selection, integration, and conflict moni-toring: Assessing the nature and generality of prefrontal cognitivecontrol mechanisms. Neuron 41:473–487.
Bayley PJ, Hopkins RO, Squire LR. 2003. Successful recollection ofremote autobiographical memories by amnesic patients with medialtemporal lobe lesions. Neuron 38:135–144.
Bayley PJ, Gold JJ, Hopkins RO, Squire LR. 2005. The neuroanat-omy of remote memory. Neuron 2:799–810.
Bernard FA, Desgranges, Eustache F, Baron JC. 2007. Neural corre-lates of age-related verbal episodic memory decline: A PET studywith combined subtraction/correlation analysis. Neurobiol aging28:1568–1576.
Brainerd CJ, Reyna VF. 2001. Fuzzy-trace theory: Dual processes inmemory, reasoning, and cognitive neuroscience. Adv Child DevBehav 28:41–100.
Brewer W. 1996. What is recollective memory? In: Rubin DC, editor.Remembering Our Past: Studies in Autobiographical Memory.Cambridge, England: Cambridge University Press. pp 19–66.
Buckner RL, Vincent JL. 2007. Unrest at rest: Default activity andspontaneous network correlations. Neuroimage 37:1091–1096.
Buckner RL, Wheeler ME, Sheridan MA. 2001. Encoding processesduring retrieval tasks. J Cogn Neurosci 13:406–415.
Burgess N, Maguire EA, O’Keefe J. 2002. The human hippocampusand spatial and episodic memory. Neuron 35:625–641.
Cabeza R, Prince SE, Daselaar SM, Greenberg D, Budde M, DolcosF, LaBar KS, Rubin DC. 2004. Brain activity during episodicretrieval of autobiographical and laboratory events: An fMRIstudy using a novel photo paradigm. J Cogn Neurosci 169:1583–1594.
Calabrese P, Markowitsch HJ, Durwen HF, Widlitzek H, Haupts M,Holinka B, Gehlen W. 1996. Right temporofrontal cortex as criti-cal locus for the ecphory of old episodic memories. J Neurol Neu-rosurg Psychiatry 61:304–310.
Cermak LS. 1984. The episodic-semantic distinction in amnesia. In:Squire LR, Butters N, editors. Neuropsychology of Memory. New-York: Guilford Press. pp 55–62.
Cipolotti L, Shallice T, Chan D, Fox N, Scahill R, Harrison G, Ste-vens J, Rudge P. 2001. Long-term retrograde amnesia: The crucialrole of the hippocampus. Neuropsychologia 39:151–172.
Coles JP, Fryer TD, Bradley PG, Nortje J, Smielewski P, Rice K, ClarkJC, Pickard JD, Menon DK. 2006. Intersubject variability andreproducibility of 15O PET studies. J Cereb Blood Flow Metab26:48–57.
Conway MA. 2001. Sensory-perceptual episodic memory and its con-text: Autobiographical memory. Philos Trans R Soc Lond B BiolSci 356:1375–1384.
Conway MA, Fthenaki A. 2000. Disruption and loss of autobiographi-cal memory. In: Cermak LS, editor. Handbook of Neuropsychol-ogy: Memory and Its Disorders, Vol. 2. Amsterdam: Elsevier. pp257–288.
Conway MA, Pleydell-Pearce CW. 2000. The construction of autobio-graphical memories in the self-memory system. Psychol Rev 107:261–288.
Conway MA, Gardiner JM, Perfect TJ, Anderson SJ, Cohen GM.1997. Changes in memory awareness during learning: The acquisi-tion of knowledge by psychology undergraduates. J Exp Psychol:General 126:393–413.
Conway MA, Turk DJ, Miller SL, Logan J, Nebes RD, Meltzer CC,Becker JT. 1999. A positron emission tomography (PET) study ofautobiographical memory retrieval. Memory 7:679–702.
Conway MA, Pleydell-Pearce CW, Whitecross SE, Sharpe H. 2002.Brain imaging autobiographical memory. Psychol Learn Motiv: AdvRes Theory 41:229–263.
Conway MA, Singer JA, Tagini A. 2004. The self and autobiographi-cal memory: Correspondence and coherence. Soc Cogn 22:491–529.
Craik IM. 1999. In search of the self. A PET study. Psychol Sci10:26–34.
Desgranges B, Baron JC, de la Sayette V, Petit-Taboue MC, Benali K,Landeau B, Lechevalier B, Eustache F. 1998. The neural substratesof memory systems impairment in Alzheimer’s disease. A PETstudy of resting brain glucose utilization. Brain 121:611–631.
Desgranges B, Baron JC, Lalevee C, Giffard B, Viader F, de la SayetteV, Eustache F. 2002a. The neural substrates of episodic memoryimpairment in Alzheimer’s disease as revealed by FDG-PET: Rela-tionship to degree of deterioration. Brain 125:1116–1124.
Desgranges B, Baron JC, Giffard B, Chetelat G, Lalevee C, Viader F,de la Sayette V, Eustache F. 2002b. The neural basis of intrusionsin free recall and cued recall: A PET study in Alzheimer’s disease.Neuroimage 17:1658–1664.
Eustache F, Rioux P, Desgranges B, Marchal G, Petit-Taboue MC,Dary M, Lechevalier B, Baron JC. 1995. Healthy aging, memorysubsystems and regional cerebral oxygen consumption. Neuropsy-chologia 33:867–887.
Eustache F, Desgranges B, Aupee AM, Guillery B, Baron JC. 2000.Functional neuroanatomy of amnesia: Positron emission tomogra-phy studies. Micros Res Technol 51:94–100.
Eustache F, Desgranges B, Giffard B, de la Sayette V, Baron JC. 2001.Entorhinal cortex disruption causes memory deficit in early Alzhei-mer’s disease as shown by PET. Neuroreport 12:683–685.
Eustache F, Piolino P, Giffard B, Viader F, de la Sayette V, Baron JC,Desgranges B. 2004. ‘In the course of time’: A PET study of thecerebral substrates of autobiographical amnesia in Alzheimer’s dis-ease. Brain 127:1549–1560.
Ferbinteanu J, Kennedy PJ, Shapiro ML. 2006. Episodic memory—From Brain to mind. Hippocampus 16:691–703.
Fink GR, Markowitsch HJ, Reinkemeier M, Bruckbauer T, Kessler J,Heiss WD. 1996. Cerebral representation of one’s own past: Neu-ral networks involved in autobiographical memory. J Neurosci 16:4275–4282.
Fortin NJ, Agster KL, Eichenbaum H. 2002. Critical role of the hip-pocampus in memory for sequences of events. Nature Neurosci5:458–462.
Fransson P. 2005. Spontaneous low-frequency BOLD signal fluctua-tions: An fMRI investigation of the resting-state default mode ofbrain function hypothesis. Hum Brain Mapp 26:15–29.
Frith CD, Frith U. 1999. Interacting minds—a biological basis. Sci-ence 286:1692–1695.
Fujii T, Moscovitch M, Nadel L. 2000. Memory consolidation, retro-grade amnesia, and the temporal lobe. In: Boller F, Grafman J edi-tors. The Handbook of Neuropsychology, Vol. 4. Amsterdam:Elsevier. pp 223–250.
Gardiner JM. 1988. Functional aspects of recollective experience.Mem Cogn 16:309–313.
Gardiner JM. 2001. Episodic memory and autonoetic consciousness:A first-person approach. Philos Trans R Soc Lond B Biol Sci 356:1351–1361.
RELIVING EPISODIC AUTOBIOGRAPHICAL MEMORIES VIA HIPPOCAMPUS 457
Hippocampus

Gilboa A. 2004. Autobiographical and episodic memory-one and thesame? Evidence from prefrontal activation in neuroimaging studies.Neuropsychologia 42:1336–1349.
Gilboa A, Winocur G, Grady CL, Hevenor SJ, Moscovitch M. 2004.Remembering our past: Functional neuroanatomy of recollection ofrecent and very remote personal events. Cereb Cortex 14:1214–1225.
Gilboa A, Ramirez J, Kohler S, Westmacott R, Black SE, MoscovitchM. 2005. Retrieval of autobiographical memory in Alzheimer’s dis-ease: Relation to volumes of medial temporal lobe and other struc-tures. Hippocampus 15:535–550.
Graham KS, Hodges JR. 1997. Differentiating the roles of the hippo-campal complex and the neocortex in long-term memory storage:Evidence from the study of semantic dementia and Alzheimer’s dis-ease. Neuropsychology 11:77–89.
Graham KS, Lee AC, Brett M, Patterson K. 2003. The neural basis ofautobiographical and semantic memory: New evidence from threePET studies. Cogn Affect Behav Neurosci 3:234–254.
Greenberg DL, Rubin DC. 2003. The neuropsychology of autobio-graphical memory. Cortex 39:687–728.
Greenberg DL, Rice HJ, Cooper JJ, Cabeza R, Rubin DC, Labar KS.2005. Co-activation of the amygdala, hippocampus and inferiorfrontal gyrus during autobiographical memory retrieval. Neuropsy-chologia 43:659–674.
Gusnard DA, Raichle ME. 2001. Searching for a baseline: Functionalimaging and the resting human brain. Nat Rev Neurosci 2:685–694.
Gusnard DA, Akbudak E, Shulman GL, Raichle ME. 2001. Medial pre-frontal cortex and self-referential mental activity: Relation to a defaultmode of brain function. Proc Natl Acad Sci USA 98:4259–4264.
Hartley T, Burgess N, Lever C, Cacucci F, O’Keefe J. 2000. Modelingplace fields in terms of the cortical inputs to the hippocampus.Hippocampus 10:369–379.
Hirano M, Noguchi K, Hosokawa T, Takayama T. 2002. I cannotremember, but I know my past events: Remembering and knowingin a patient with amnesic syndrome. J Clin Exp Neuropsychol 24:548–555.
Hirono N, Mori E, Ishii K, Imamura T, Tanimukai S, Kazui H,Hashimoto M, Takatsuki Y, Kitagaki H, Sasaki M. 2001. Neuronalsubstrates for semantic memory: A positron emission tomographystudy in Alzheimer’s disease. Dement Geriatr Cogn Disord 12:15–21.
Kapur N. 2000. Focal retrograde amnesia and the attribution of cau-sality: An exceptionally benign commentary. Cogn Neuropsychol17:623–637.
Kopelman MD. 2000. The neuropsychology of remote memory. In:Cermak L, editor. Handbook of Neuropsychology. Amsterdam:Elsevier. pp 251–280.
Kopelman MD, Kapur N. 2001. The loss of episodic memories in ret-rograde amnesia: Single-case and group studies. Philos Trans R SocLond B Biol Sci 356:1409–1421.
Kopelman MD, Stanhope N, Kingsley D. 1997. Temporal and spatialcontext memory in patients with focal frontal, temporal lobe, anddiencephalic lesions. Neuropsychologia 35:1533–1545.
Kopelman MD, Lasserson D, Kingsley DR, Bello F, Rush C, StanhopeN, Stevens TG, Goodman G, Buckman JR, Heilpern G, KendallBE, Colchester AC. 2003. Retrograde amnesia and the volume ofcritical brain structures. Hippocampus 13:879–891.
Lekeu F, Van der Linden M, Chicherio C, Collette F, Degueldre C,Franck G, Moonen G, Salmon E. 2003a. Brain correlates of per-formance in a free/cued recall task with semantic encoding in Alz-heimer disease. Alzheimer Dis Assoc Disord 17:35–45.
Lekeu F, Van der Linden M, Degueldre C, Lemaire C, Luxen A, FranckG, Moonen G, Salmon E. 2003b. Effects of Alzheimer’s disease onthe recognition of novel versus familiar words: Neuropsychologicaland clinico-metabolic data. Neuropsychology 17:143–154.
Levine B, Black SE, Cabeza R, Sinden M, McIntosh AR, Toth JP,Tulving E, Stuss DT. 1998. Episodic memory and the self in a caseof isolated retrograde amnesia. Brain 121:1951–1973.
Levine B, Svoboda E, Hay JF, Winocur G, Moscovitch M. 2002.Aging and autobiographical memory: Dissociating episodic fromsemantic memory. Psychol Aging 17:677–689.
Maguire EA. 2001. Neuroimaging studies of autobiographical eventmemory. Philos Trans R Soc Lond B Biol Sci 356:1441–1451.
Maguire EA, Frith CD. 2003. Aging affects the engagement of thehippocampus during autobiographical memory retrieval. Brain126:1511–1523.
Maguire EA, Henson RN, Mummery CJ, Frith CD. 2001. Activity inprefrontal cortex, not hippocampus, varies parametrically with theincreasing remoteness of memories. Neuroreport 12:441–444.
Mantyla T. 1993. Knowing but not remembering: Adult age differen-ces in recollective experience. Mem Aging 21:379–388.
Matuszewski V, Piolino P, de la Sayette V, Lalevee C, Pelerin A,Dupuy B, Viader F, Eustache F, Desgranges B. 2006. Retrievalmechanisms for autobiographical memories: Insights from the fron-tal variant of frontotemporal dementia. Neuropsychologia 44:2386–2397.
Meeter M, Murre JM. 2004. Consolidation of long-term memory:Evidence and alternatives. Psychol Bull 130:843–857.
Mc Gaugh JL. 2000. Memory—A century of consolidation. Science287:248–251.
Moscovitch M, Winocur G. 2002. The frontal cortex and workingwith memory. In: Stuss DT, Knight RT, editors. The FrontalLobes. Oxford: Oxford University Press. pp 188–209.
Moscovitch M, Rosenbaum RS, Gilboa A, Addis DR, Westmacott R,Grady C, McAndrews MP, Levine B, Black S, Winocur G, NadelL. 2005. Functional neuroanatomy of remote episodic, semanticand spatial memory: A unified account based on multiple tracetheory. J Anat 207:35–66.
Murre JMJ, Graham KS, Hodges JR. 2001. Semantic dementia: Rele-vance to connectionist models of long-term memory. Brain 124:647–675.
Nadel L, Moscovitch M. 1997. Memory consolidation, retrograde am-nesia and the hippocampal complex. Curr Opin Neurobiol 7:217–227.
Nadel L, Samsonovich A, Ryan L, Moscovitch M. 2000. Multipletrace theory of human memory: Computational, neuroimaging,and neuropsychological results. Hippocampus 10:352–368.
Nestor PJ, Graham KS, Bozeat S, Simons JS, Hodges JR. 2002. Memoryconsolidation and the hippocampus: Further evidence from studies ofautobiographical memory in semantic dementia and frontal variantof frontotemporal dementia. Neuropsychologia 40:633–654.
Niki K, Luo J. 2002. An fMRI study on the time-limited role of themedial temporal lobe in long-term topographical autobiographicmemory. J Cogn Neurosci 14:500–507.
Northoff G, Bermpohl F. 2004. Cortical midline structures and theself. Trends Cogn Sci 8:102–107.
Northoff G, Heinzel A, de Greck M, Bermpohl F, Dobrowolny H,Panksepp J. 2006. Self-referential processing in our brain—a meta-analysis of imaging studies on the self. Neuroimage 1:440–457.
Noulhiane M, Piolino P, Hasboun D, Baulac M, Samson S. 2007. Ep-isodic autobiographical memory in epileptic patients with medialtemporal lobe resection: Neuropsychological Findings and Relationto Volumes of Medial Temporal Lobe. Brain 130:3184–3199.
O’Keefe J, Nadel L. 1978. The Hippocampus as a Cognitive Map.Oxford: Oxford University Press.
Okuda J, Fujii T, Ohtake H, Tsukiura T, Tanji K, Suzuki K, Kawa-shima R, Fukuda H, Itoh M, Yamadori A. 2003. Thinking of thefuture and past: The roles of the frontal pole and the medial tem-poral lobes. Neuroimage 19:1369–1380.
Persson J, Nyberg L. 2000. Conjunction analysis of cortical activationscommon to encoding and retrieval. Microsc Res Technol 51:39–44.
Petrides M. 2000. Frontal lobe and memory. In: Boller F, Grafman J,editors. Handbook of Neuropsychology, 2nd ed. Amsterdam: Elsev-ier. Vol. 2, pp 67–84.
458 PIOLINO ET AL.
Hippocampus

Piefke M, Fink GR. 2005. Recollections of one’s own past: The effectsof aging and gender on the neural mechanisms of episodic autobio-graphical memory. Anat Embryol 210:497–512.
Piefke M, Weiss PH, Zilles K, Markowitsch HJ, Fink GR. 2003. Dif-ferential remoteness and emotional tone modulate the neural corre-lates of autobiographical memory. Brain 126:650–668.
Piolino P, Desgranges B, Benali K, Eustache F. 2002. Episodic andsemantic remote autobiographical memory in ageing. Memory10:239–257.
Piolino P, Desgranges B, Belliard S, Matuszewski V, Lalevee C, De laSayette V, Eustache F. 2003a. Autobiographical memory and auton-oetic consciousness: Triple dissociation in neurodegenerative dis-eases. Brain 126:2203–2219.
Piolino P, Belliard S, Desgranges B, Perron M, Eustache F. 2003b.Autobiograhical memory and autonoetic consciousness in a case ofsemantic dementia. Cogn Neuropsychol 20:619–639.
Piolino P, Giffard-Quillon G, Desgranges B, Chetelat G, Baron J-C,Eustache F. 2004. Re-experiencing old memories via hippocampus:A PET study of autobiographical memory. Neuroimage 22:1371–1383.
Piolino P, Hannequin D, Desgranges B, Girard C, Beaunieux H, Gif-fard B, Lebreton K, Eustache F. 2005. Right ventral frontal hypo-metabolism and abnormal sense of self in a case of disproportion-ate retrograde amnesia. Cogn Neuropsychol 22:1005–1034.
Piolino P, Desgranges B, Clarys D, Guillery-Girard B, Isingrini M,Eustache F. 2006. Autobiographical memory and sense of remem-bering in aging. Psychol Aging 21:510–525.
Piolino P, Desgranges B, Manning L, North P, Jokic F, Eustache F.2007a. Autobiographical memory, the sense of recollection and exec-utive functions after severe closed head injury. Cortex 43:176–195.
Piolino P, Chetelat G, Matuszewski V, Landeau B, De La Sayette, Eus-tache F, Desgranges B. 2007b. In search of autobiographical mem-ories: A PET study in frontotemporal dementia. Neuropsychologia45:2730–2743.
Raichle ME. 2001. Cognitive neuroscience. Bold insights. Nature412:128–130.
Rauchs G, Piolino P, Mezenge F, Landeau B, Lalevee C, Pelerin A,Viader F, de la Sayette V, Eustache F, Desgranges D. 2007. Auto-noetic consciousness in Alzheimer’s disease: Neuropsychological andPET findings using an episodic learning and recognition task. Neu-robiol Aging 28:1410–1420.
Ranganath C, Yonelinas AP, Cohen MX, Dy CJ, Tom SM, D’EspositoM. 2003. Dissociable correlated of recollection and familiaritywithin the medial temporal lobes. Neuropsychologia 42:2–13.
Rubin DC, Rahhal TA, Poon LW. 1998. Things learned in earlyadulthood are remembered best. Mem Cogn 26:3–19.
Ryan L, Nadel L, Keil K, Putnam K, Schnyer D, Trouard T, Mosco-vitch M. 2001. Hippocampal complex and retrieval of recent andvery remote autobiographical memories: Evidence from functionalmagnetic resonance imaging in neurologically intact people. Hip-pocampus 11:707–714.
Squire LR. 1992. Memory and the hippocampus: A synthesis fromfindings with rats, monkeys and humans. Psychol Rev 99:195–231.
Squire LR. 2004. Memory systems of the brain: A brief history andcurrent perspective. Neurobiol Learn Mem 82:171–177.
Squire LR, Alvarez P. 1995. Retrograde amnesia and memory consoli-dation: A neurobiological perspective. Curr Opin Neurobiol 5:169–177.
Squire LR, Bayley PJ. 2007. The neuroscience of remote memory.Curr Opin Neurobiol 17:185–196.
Steinvorth S, Corkin S, Halgren E. 2006. Ecphory of autobiographicalmemories: An fMRI study of recent and remote memory retrieval.Neuroimage 30:285–298.
Talairach J, Tournoux P. 1988. Co-planar Stereotaxic Atlas of theHuman Brain. New York: Thieme Medical Publishers.
Teipel SJ, Willoch F, Ishii K, Burger K, Drzezga A, Engel R, Barten-stein P, Moller HJ, Schwaiger M, Hampela H. 2006. Resting stateglucose utilization and the CERAD cognitive battery in patientswith Alzheimer’s disease. Neurobiol Aging 27:681–690.
Teng E, Squire LR. 1999. Memory for places learned long ago isintact after hippocampal damage. Nature 400:675–677.
Tulving E. 1985. Memory and consciousness. Can Psychol 25:1–12.
Tulving E. 2001. Episodic and common sense: How far apart? PhilosTrans R Soc Lond B Biol Sci 356:1505–1515.
Tulving E. 2002. Episodic memory: From mind to brain. Ann RevPsychol 53:1–25.
Tulving E, Schacter DL, McLachlan DR., Moscovitch M. 1988. Pri-ming of semantic autobiographical knowledge: A case study of ret-rograde amnesia. Brain Cogn 8:3–20.
Tulving E, Habib R, Nyberg L, Lepage M, McIntosh AR. 1999. Posi-tron emission tomography correlations in and beyond medial tem-poral lobes. Hippocampus 9:71–82.
Tzourio-Mazoyer N, Landeau B, Papathanassiou D, Crivello F, EtardO, Delcroix N, Mazoyer B, Joliot M. 2002. Automated anatomicallabeling of activations in SPM using a macroscopic anatomical par-cellation of the MNI MRI single-subject brain. Neuroimage15:273–289.
Tsukiura T, Fujii T, Okuda J, Ohtake H, Kawashima R, Itoh M,Fukuda H, Yamadori A. 2002. Time-dependent contribution ofthe hippocampal complex when remembering the past: A PETstudy. Neuroreport 13:2319–2323.
Umeda S, Akine Y, Kato M, Muramatsu T, Mimura M, Kandatsu S,Tanada S, Obata T, Ikehira H, Suhara T. 2005. Functional networkin the prefrontal cortex during episodic memory retrieval. Neuro-image 26:932–940.
Viard A, Piolino P, Desgranges B, Lebreton K, Chetelat G, LandeauB, Young A, De La Sayette V, Eustache F. Autobiographical memo-ries and sense of recollection over the entire lifetime of elderly sub-jects: an fMRI study. Cereb Cortex 17:2453–2467.
Viskontas IV, McAndrews MP, Moscovtich M. 2000. Remote episodicmemory deficits in patients with unilateral temporal lobe epilepsyand excisions. J Neurosci 20:5853–5857.
Westmacott R, Leach L, Freedman M, Moscovitch M. 2001. Differentpatterns of autobiographical memory loss in semantic dementiaand medial temporal lobe amnesia: A challenge to consolidationtheory. Neurocase 7:37–55.
Wheeler MA, McMillan CT. 2001. Focal retrograde amnesia and theepisodic-semantic distinction. Cogn Affect Behav Neurosc 1:22–36.
Wheeler MA, Stuss DT, Tulving E. 1997. Toward a theory of episodicmemory: The frontal lobes and autonoetic consciousness. PsycholBull 121:331–354.
Woods RP, Grafton ST, Holmes CJ, Cherry SR, Mazziotta JC. 1998.Automated image registration. I. General methods and intrasubject,intramodality validation. J Comput Assist Tomogr 22:139–152.
RELIVING EPISODIC AUTOBIOGRAPHICAL MEMORIES VIA HIPPOCAMPUS 459
Hippocampus
Copyright © 2022 FDOKUMEN




















