
Alternative sigma factor RpoN2 is required for flagellar motility and full virulence of Xanthomonas...
7
Please cite this article in press as: Tian F, et al. Alternative sigma factor RpoN2 is required for flagellar motility and full virulence of Xanthomonas oryzae pv. oryzae. Microbiol Res (2014), http://dx.doi.org/10.1016/j.micres.2014.07.002 ARTICLE IN PRESS G Model MICRES-25692; No. of Pages 7 Microbiological Research xxx (2014) xxx–xxx Contents lists available at ScienceDirect Microbiological Research j ourna l h om epage: www.elsevier.com/locate/micres Alternative sigma factor RpoN2 is required for flagellar motility and full virulence of Xanthomonas oryzae pv. oryzae Fang Tian, Chao Yu, Haiyun Li, Xiaoli Wu, Bo Li, Huamin Chen, Maosen Wu, Chenyang He ∗ State Key Laboratory for Biology of Plant Diseases and Insect Pests, Institute of Plant Protection, Chinese Academy of Agricultural Sciences, Beijing, China a r t i c l e i n f o Article history: Received 28 January 2014 Received in revised form 4 July 2014 Accepted 21 July 2014 Available online xxx Keywords: Bacterial leaf blight Flagellin Pathogenesis Sigma factor Regulatory protein a b s t r a c t Xanthomonas oryzae pv. oryzae (Xoo), the bacterial blight pathogen of rice, harbors a single polar flagellum for motility. How the flagellar system is regulated and how it is related to bacterial pathogenesis are not well understood. The genomic sequence of Xoo strain PXO99 A revealed a flagellar regulon containing over 60 contiguous genes. A gene encoding alternative sigma factor 54 ( 54 ), named as rpoN2, is located in the central region of the regulon. RT-PCR analysis demonstrated rpoN2 was transcribed in an operon with flgRR, and fleQ. Single gene deletion mutants of the rpoN2 operon were generated. The rpoN2 and fleQ mutant lost swimming motility, whereas the flgRR mutant remained motile. Quantitative RT-PCR analysis further demonstrated that expression of regulatory genes fliA and flgM, and structural genes flgG, flhB, and fliC were significantly down-regulated in the rpoN2 and fleQ mutants. These results indicated that RpoN2 and FleQ synergistically controlled flagellar motility by regulating gene expression. Interestingly, the rpoN2 mutant, but not the fleQ mutant was impaired in its virulence on rice. In addition, we showed that the flagellin gene fliC mutant, which was non-motile, was not defective in virulence. Thus, we concluded that flagellar motility might not be essential for Xoo virulence in rice, and RpoN2 probably regulated bacterial virulence through a manner independent of its role in controlling flagellar gene expression. © 2014 Elsevier GmbH. All rights reserved. 1. Introduction Xanthomonas oryzae pv. oryzae (Xoo), causes bacterial leaf blight of rice, one of the most devastating diseases of this staple crop. Due to its scientific and economic importance, Xoo has been one of the model systems to study the molecular mechanism of bacte- rial pathogenesis in plants. Xoo enters rice leaves via hydathodes or wounds, and spreads in plants through xylem (Salzberg et al., 2008). During the infection process, virulence factors such as exopolysac- charide (EPS), extracellular enzymes, adhesins, and the type III secretion system and its effectors play crucial roles (Ray et al., 2000; Das et al., 2009; White and Yang, 2009; Song and Yang, 2010). Some regulatory proteins affecting the expression of these virulence fac- tors have also been characterized (Tsuge et al., 2005; He et al., 2010; Subramoni et al., 2012; Yang et al., 2012). Like many other xanthomonads, Xoo harbors a single polar flagellum for motility. However, how the flagellar system is regulated, and whether the flagellar motility is involved in Xoo pathogenesis remain unclear. ∗ Corresponding author at: Room 207, Institute of Plant Protection, Chinese Academy of Agricultural Sciences, No. 2 Yuanmingyuan West Road, Haidian District, Beijing 100193, China. Tel.: +86 10 62894147; fax: +86 10 62894642. E-mail address: [email protected] (C. He). The function of flagella allows bacteria to move toward favorable conditions and away from harms by responding to chemical signals (Haefele and Lindow, 1987; Bourret et al., 2002; Porter et al., 2011). Flagellar motility can promote the infection of pathogenic bacte- ria by facilitating biofilm formation, colonization and attachment to host tissues (Giron et al., 2002; Ottemann and Lowenthal, 2002; Harshey, 2003; Tans-Kersten et al., 2004). It is important for the fit- ness and survival of epiphytic bacteria on leaf surfaces, and in some cases it also contributes to the virulence at post-penetration stages (Beattie and Lindow, 1995; Bahar et al., 2011). On the other hand, flagellin monomer, the building blocks for flagellar filament, has been known as a pathogen associated molecular pattern (PAMP) to induce PAMP-triggered immunity (PTI) in host (Zipfel and Felix, 2005; Franchi et al., 2006). Therefore, presence of flagella is not always an advantageous factor (Amiel et al., 2010). Assembly of flagellum is a highly organized process requiring temporal expression of dozens of genes (Macnab, 2004). Current models from several bacteria have shown that these genes are expressed in a hierarchical manner (Aldridge and Hughes, 2002; McCarter, 2006). In enteric bacteria such as Salmonella typhimurium and Escherichia coli, FlhDC are the master regulators required for transcription of class II genes including alternative sigma factor FliA ( 28 ) and some structural components (Liu and Matsumura, 1994). In single polar flagellated Vibrio cholerae and Pseudomonas http://dx.doi.org/10.1016/j.micres.2014.07.002 0944-5013/© 2014 Elsevier GmbH. All rights reserved.
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Alternative sigma factor RpoN2 is required for flagellar motility and full virulence of Xanthomonas...

M
Af
FS
a
ARRAA
KBFPSR
1
oDorwDcsDrt2xHfl
AB
h0
ARTICLE IN PRESSG ModelICRES-25692; No. of Pages 7
Microbiological Research xxx (2014) xxx–xxx
Contents lists available at ScienceDirect
Microbiological Research
j ourna l h om epage: www.elsev ier .com/ locate /micres
lternative sigma factor RpoN2 is required for flagellar motility andull virulence of Xanthomonas oryzae pv. oryzae
ang Tian, Chao Yu, Haiyun Li, Xiaoli Wu, Bo Li, Huamin Chen, Maosen Wu, Chenyang He ∗
tate Key Laboratory for Biology of Plant Diseases and Insect Pests, Institute of Plant Protection, Chinese Academy of Agricultural Sciences, Beijing, China
r t i c l e i n f o
rticle history:eceived 28 January 2014eceived in revised form 4 July 2014ccepted 21 July 2014vailable online xxx
eywords:acterial leaf blightlagellinathogenesis
a b s t r a c t
Xanthomonas oryzae pv. oryzae (Xoo), the bacterial blight pathogen of rice, harbors a single polar flagellumfor motility. How the flagellar system is regulated and how it is related to bacterial pathogenesis are notwell understood. The genomic sequence of Xoo strain PXO99A revealed a flagellar regulon containingover 60 contiguous genes. A gene encoding alternative sigma factor 54 (�54), named as rpoN2, is locatedin the central region of the regulon. RT-PCR analysis demonstrated rpoN2 was transcribed in an operonwith flgRR, and fleQ. Single gene deletion mutants of the rpoN2 operon were generated. The rpoN2 and fleQmutant lost swimming motility, whereas the flgRR mutant remained motile. Quantitative RT-PCR analysisfurther demonstrated that expression of regulatory genes fliA and flgM, and structural genes flgG, flhB, andfliC were significantly down-regulated in the rpoN2 and fleQ mutants. These results indicated that RpoN2
igma factoregulatory protein
and FleQ synergistically controlled flagellar motility by regulating gene expression. Interestingly, therpoN2 mutant, but not the fleQ mutant was impaired in its virulence on rice. In addition, we showed thatthe flagellin gene fliC mutant, which was non-motile, was not defective in virulence. Thus, we concludedthat flagellar motility might not be essential for Xoo virulence in rice, and RpoN2 probably regulatedbacterial virulence through a manner independent of its role in controlling flagellar gene expression.
© 2014 Elsevier GmbH. All rights reserved.
. Introduction
Xanthomonas oryzae pv. oryzae (Xoo), causes bacterial leaf blightf rice, one of the most devastating diseases of this staple crop.ue to its scientific and economic importance, Xoo has been onef the model systems to study the molecular mechanism of bacte-ial pathogenesis in plants. Xoo enters rice leaves via hydathodes orounds, and spreads in plants through xylem (Salzberg et al., 2008).uring the infection process, virulence factors such as exopolysac-haride (EPS), extracellular enzymes, adhesins, and the type IIIecretion system and its effectors play crucial roles (Ray et al., 2000;as et al., 2009; White and Yang, 2009; Song and Yang, 2010). Some
egulatory proteins affecting the expression of these virulence fac-ors have also been characterized (Tsuge et al., 2005; He et al.,010; Subramoni et al., 2012; Yang et al., 2012). Like many other
Please cite this article in press as: Tian F, et al. Alternative sigma facXanthomonas oryzae pv. oryzae. Microbiol Res (2014), http://dx.doi.org
anthomonads, Xoo harbors a single polar flagellum for motility.owever, how the flagellar system is regulated, and whether theagellar motility is involved in Xoo pathogenesis remain unclear.
∗ Corresponding author at: Room 207, Institute of Plant Protection, Chinesecademy of Agricultural Sciences, No. 2 Yuanmingyuan West Road, Haidian District,eijing 100193, China. Tel.: +86 10 62894147; fax: +86 10 62894642.
E-mail address: [email protected] (C. He).
ttp://dx.doi.org/10.1016/j.micres.2014.07.002944-5013/© 2014 Elsevier GmbH. All rights reserved.
The function of flagella allows bacteria to move toward favorableconditions and away from harms by responding to chemical signals(Haefele and Lindow, 1987; Bourret et al., 2002; Porter et al., 2011).Flagellar motility can promote the infection of pathogenic bacte-ria by facilitating biofilm formation, colonization and attachmentto host tissues (Giron et al., 2002; Ottemann and Lowenthal, 2002;Harshey, 2003; Tans-Kersten et al., 2004). It is important for the fit-ness and survival of epiphytic bacteria on leaf surfaces, and in somecases it also contributes to the virulence at post-penetration stages(Beattie and Lindow, 1995; Bahar et al., 2011). On the other hand,flagellin monomer, the building blocks for flagellar filament, hasbeen known as a pathogen associated molecular pattern (PAMP)to induce PAMP-triggered immunity (PTI) in host (Zipfel and Felix,2005; Franchi et al., 2006). Therefore, presence of flagella is notalways an advantageous factor (Amiel et al., 2010).
Assembly of flagellum is a highly organized process requiringtemporal expression of dozens of genes (Macnab, 2004). Currentmodels from several bacteria have shown that these genes areexpressed in a hierarchical manner (Aldridge and Hughes, 2002;McCarter, 2006). In enteric bacteria such as Salmonella typhimurium
tor RpoN2 is required for flagellar motility and full virulence of/10.1016/j.micres.2014.07.002
and Escherichia coli, FlhDC are the master regulators required fortranscription of class II genes including alternative sigma factorFliA (�28) and some structural components (Liu and Matsumura,1994). In single polar flagellated Vibrio cholerae and Pseudomonas

ARTICLE IN PRESSG ModelMICRES-25692; No. of Pages 7
2 F. Tian et al. / Microbiological Research xxx (2014) xxx–xxx
Table 1Bacterial strains and plasmids used in this study.
Strain or plasmid Description Reference or source
Bacterial strainsXoo PXO99A Wild-type strain, Philippine race 6 Lab collection (Hopkins et al., 1992)�rpoN2 rpoN2 gene deletion mutant derived from PXO99A, Gmr This work�flgRR flgRR gene deletion mutant derived from PXO99A, Gmr This work�fleQ fleQ gene deletion mutant derived from PXO99A, Gmr This work�fliC fliC gene deletion mutant derived from PXO99A, Gmr This work�rpoN2-C �rpoN2 containing plasmid pHrpoN2, Gmr Spr This work�flgRR-C �flgRR containing plasmid pHflgRR, GmrSpr This work�fleQ-C �fleQ containing plasmid pHfleQ, Gmr Spr This work�fliC-C �fliC containing plasmid pHfliC, Gmr Spr This workE. coli DH5� supE44 �lacU169 (�80lacZ�M15) hsdR17 recA1 endA1 gyrA96 thi-1 relA1 Hanahan (1983)
PlasmidspK18mobsacB Suicidal vector carrying sacB gene for mutagenesis, Kmr Schafer et al. (1994)pHM1 Broad-host-range expression vector, Spr Hopkins et al. (1992)pKrpoN2G pK18mobsacB derivative carrying a rpoN2 mutation, Kmr Gmr This workpKflgRRG pK18mobsacB derivative carrying a flgRR mutation, Kmr Gmr This workpKfleQG pK18mobsacB derivative carrying a fleQ mutation, Kmr Gmr This workpKfliCG pK18mobsacB derivative carrying a fliC mutation, Kmr Gmr This workpHrpoN2 pHM1 carrying the full-length of rpoN2 gene, Spr This work
, Spr This work Spr This workSpr This work
a�c2(Frta
gt2oflan
2
2
iT3asmbca
2
secpc
Table 2Primers used in this study.
Primer Sequence (5′–3′) Restrictionenzymesite/use
GmF CTAATGCATGACGCACACCGTGGAAA NsiIGmR ATAATGCATGCGGCGTTGTGACAATTT NsiIrpoN2F GCAAGCTTATGAAGACGACCATTTCCGCC HindIIIrpoN2R ACTCTAGAGCCTTGCCGCGACAGCAATC XbaIflgRRF AGCGAATTCGCGTCGCTTGATCGAAAG EcoRIflgRRR GTCAAGCTTCCAGTCGAAGAATTTGTCGG HindIIIfleQF GCAAGCTTATGAGTGAGTCCCGCATTCT HindIIIfleQR ACTCTAGATCAGTTGGCCAGCTCGGTCTG XbaIfliCF CCGGAATTCGCAGGCGGACGGAGTTTA EcoRIfliCR CCCAAGCTTCGGGACACCAGGGTAGACA HindIIIrpoN2CF GTCAAGCTTCTTCGGAAGCGCTTCCATG HindIIIrpoN2CR AGCGAATTCTTATCCTGCCCGGGCAAGCAG EcoRIflgRRCF AGCAAGCTTAGGATAAGGACTACTATGAGC HindIIIflgRRCR GTCTCTAGATCACTCATTCCTGGCTCCTT XbaIfleQCF GCTAAGCTTGGTAAGGAGCCAGGAATGAG HindIIIfleQCR AGCGAATTCTCAGTTGGCCAGCTCGGTCTG EcoRIfliCCF GCTAAGCTTGCGGTAACGGCCAACTTGATG HindIIIfliCCR AGCGAATTCTTACTGCAGCAGGCTCAGCAC EcoRIrpoN2a CATCACCACCGGCAAATACAT RT-PCRflgRRb GAAGAGCGTGTTCACGATGCT RT-PCRflgRRa GTGGCTCAATATCGTGGGCA RT-PCRfleQb AGACATTGGCTTCGTGCAGG RT-PCRgyrBF GGCGAGCACAATGGCATT qPCRgyrBR CCATCCTTCTGCGGGATGT qPCRfliAF CCCAACCCGCAGCAGATGA qPCRfliAR CTGGCCGTGGATCTGGCAG qPCRfliCF2 GAGCGCAACCTCGGAAAACC qPCRfliCR2 GGAACCGACTTGGCCTGGG qPCRflgMF GTCAAGTTGACCGGCGAAGC qPCRflgMR GAATAGCTGCCGTTCTGC qPCRflgGF CGGCACCTTCCAGATCAAC qPCRflgGR GATGGTCAGCGACTGCGC qPCRflhBF CTATTCCGTCGTGGCCCAG qPCR
pHflgRR pHM1 carrying the full-length of flgRR genepHfleQ pHM1 carrying the full-length of fleQ gene,pHfliC pHM1 carrying the full-length of fliC gene,
eruginosa, there is a four-tiered transcriptional cascade involving54-dependent transcriptional activator FlrA/FleQ (class I), two-omponent system FlrBC/FleSR (class II) and FliA (Prouty et al.,001; Dasgupta et al., 2003). FliA is required for late flagellar geneclass III/IV) expression and its function is negatively regulated bylgM, which is an anti-sigma factor binding to FliA specifically andepress its activity (Sorenson et al., 2004). Once FlgM is secretedhrough the complete basal-body-hook structure, FliA is free toctivate the late gene expression (Karlinsey et al., 2000).
The genomic sequence of Xoo has revealed that there is a fla-ellar regulon containing over 60 contiguous genes, which encodehe complete set of components for flagellar biogenesis, (Lee et al.,005; Salzberg et al., 2008). In this study, we analyzed the functionf the master regulators RpoN2 (�54) and FleQ. We found that theagellar motility of Xoo did not contribute to its virulence directlynd RpoN2 probably regulated the virulence of Xoo through a man-er independent of its role in controlling flagellar gene expression.
. Materials and methods
.1. Bacterial strains, plasmids, primers and growth conditions
The bacterial strains and plasmids used in this study are listedn Table 1. All primers used for making constructs are listed inable 2. E. coli strains were grown in Luria-Bertain (LB) medium at7 ◦C (Hanahan, 1983). Xoo PXO99A wild type (Hopkins et al., 1992)nd the derived mutants were routinely maintained on peptoneucrose agar (PSA) plates or in liquid M210 media at 28 ◦C. A plant-imicking media XOM2 (Furutanin et al., 2003) was also used for
acterial growth assays. Antibiotics were used at the following con-entrations: kanamycin (Km) 50 �g/ml, gentamicin (Gm) 25 �g/ml,nd spectinomycin (Sp) 50 �g/ml.
.2. General DNA manipulation
General DNA manipulation was performed according totandard techniques (Sambrook and Russell, 2001). Restriction
Please cite this article in press as: Tian F, et al. Alternative sigma facXanthomonas oryzae pv. oryzae. Microbiol Res (2014), http://dx.doi.org
nzymes, T4 DNA ligase and Taq DNA polymerase were pur-hased from Takara Co. in China (Dalian, Liaoning). Oligonucleotiderimers were ordered from Invitrogen Co. in China (Beijing). Theycling conditions used for PCRs were as follows: 3 min at 94 ◦C;
flhBR CTTCCAGCGAGGGCAATTC qPCR
Restriction enzyme sites are in bold text.
35 cycles of 30 s at 94 ◦C, 30 s at 60 ◦C and 1 min at 72 ◦C; 10 min at72 ◦C.
tor RpoN2 is required for flagellar motility and full virulence of/10.1016/j.micres.2014.07.002
2.3. RT-PCR
Total RNAs were extracted with TRIzol (Qiagen) agent accordingto the manufacturer’s descriptions. 2 �g of total RNA was converted

ING ModelM
cal Res
twbjfla
2
bmpcgfipsssGcmrmGsm9pCw
flratpf
2
ddpatal
2
pwTtrS0tlaw
ARTICLEICRES-25692; No. of Pages 7
F. Tian et al. / Microbiologi
o cDNA using Superscript III reverse transcriptase (Invitrogen)ith fleQR, a specific reverse primer for fleQ. The cDNA or RNA
efore reverse transcription was used as template to amplify theunction regions of rpoN2-flgRR (598 bp), rpoN2-fleQ (987 bp), andgRR-fleQ (598 bp) with primer pairs rpoN2a/flgRRb, rpoN2a/fleQb,nd flgRRa/fleQb, respectively.
.4. Generation of PXO99A derived gene deletion mutants
The PXO99A derived gene deletion mutants were generatedy marker exchange strategy as stated below. The gene frag-ents of rpoN2, flgRR, fleQ, and fliC were amplified using the
rimers rpoN2F/R, flgRRF/R, fleQF/R, and fliCF/R respectively, andloned into suicide vector pK18mobsacB (Schafer et al., 1994). Aentamicin resistance (Gmr) gene amplified with primers GmF/Rrom pBBR1MCS-5 (Kovach et al., 1995), was then insertednto each gene fragment carried by pK18mobsacB, resulting inlasmids pKrpoN2G, pKflgRRG, pKfleQG, and pKfliCG. These con-tructs were separately electroporated into wild-type PXO99A
train, and homologous recombination was screened on PSA + 10%ucrose + Gm. Finally the single colonies that were resistant tom and high concentration sucrose, but sensitive to Km wereonsidered as mutant candidates. The rpoN2, flgRR, fleQ, and fliCutant candidates were further analyzed by PCR using primer pair
poN2F/R, flgRRF/R, fleQF/R, and fliCF/R respectively. In the rpoN2utant, an internal 483-bp fragment was replaced by the 855-bpmr gene. In the flgRR mutant, the Gmr gene was inserted into theingle PstI site of flgRR. In the fleQ mutant, an internal 489-bp frag-ent was replaced by the Gmr gene. In the fliC mutant, an internal
27-bp fragment was replaced by the Gmr gene. Therefore, the PCRroducts of the mutants are different from those of the wild type.andidate strains generating only one PCR band with correct sizeere considered as final mutants, which were used in this study.
For complementation of each mutant, the full length of rpoN2,gRR, fleQ, and fliC genes were amplified with primer pairspoN2CF/R, flgRRCF/R, fleQCF/R, and fliCCF/R, respectively, and lig-ted into the broad-host vector pHM1 (Hopkins et al., 1992) athe corresponding enzymes sites. The resulted constructs pHrpoN2,HflgRR, pHfleQ, and pHfliC were electroporated into each mutantor complementation studies.
.5. Motility assay
The motility of PXO99A strains was tested as previouslyescribed (Shen et al., 2001). Bacterial cells of PXO99A and theerived mutants were inoculated onto the surface of a semisolidlates containing 0.03% Bacto Peptone, 0.03% yeast extract, and 0.3%gar. Pictures were taken after bacterial growth for 4 days. Size ofhe swimming zones around the spot of inoculation indicated thebility of bacterial movement. The experiments were repeated ateast three times.
.6. Quantitative RT-PCR
The cDNA was obtained by using random hexanucleotiderimers. Specific primers for each gene were designed using soft-are (Primer 5.0). The primer names and sequences are listed in
able 2. The amplified DNA fragments were around 100 bp. Real-ime quantitative PCR was performed using the BIO-RAD iCyclereal-time PCR detection system (Bio-Rad, Hercules, CA) and iQYBR green supermix (Bio-Rad). 1 �mol of primers were used with.2 �g of cDNA template. The cycler conditions were 2 min at 95 ◦C;
Please cite this article in press as: Tian F, et al. Alternative sigma facXanthomonas oryzae pv. oryzae. Microbiol Res (2014), http://dx.doi.org
hen 40 cycles of 20 s at 95 ◦C, 30 s at 60 ◦C and 30 s at 68 ◦C; fol-owed by a dissociation curve with 80 cycles of 10 s at 55 ◦C with
0.5 ◦C increase per cycle. The relative levels of gene expressionere determined by the 2−��CT method (Livak and Schmittgen,
PRESSearch xxx (2014) xxx–xxx 3
2001) with the DNA gyrase subunit B (gyrB) gene as the internalcontrol (Tsuge et al., 2006). There were three technical repeats ineach experiment. The experiment was repeated three times.
2.7. Virulence assay
The virulence of wild-type PXO99A and mutant strains wastested on susceptible rice plant (Oryza sativa L. ssp. japonica) usingthe leaf clipping method (Ray et al., 2000). Bacterial cells weregrown in M210 overnight until saturation. They were spun downand resuspended in water at an OD600 of 1.0. 6-week old riceplants were used in this experiment. Inoculation was carried outby clipping the tips of rice leaves with scissors pretreated with thebacterial cell suspension. At least 10 leaves were used for inocula-tion of one bacterial strain. The lesion lengths were measured, andthe pictures were taken 14 days after inoculation. The experimentwas repeated three times.
3. Results
3.1. Flagellar genes of Xoo
A chromosomal locus containing over 60 contiguous flagellargenes was identified in the genome of Xoo strain PXO99A (TableS1). These genes encode proteins with various functions including:(a) structural components: hook-basal body complex (flgABCDE-FGHIJKL, and fliEF), switch (fliGMN) and filament related (fliCDS); (b)protein export apparatus (flhAB, fliOPQR and fliHIJ); (c) regulatoryfactors (rpoN2, flgRR, fleQ, fliA, flgM, flhF, and fleN); (d) glycosyl-ation island (vioA to rbfC); (e) chemotaxis related (cheAVYZ); (f)GGDEF/EAL domains (PXO 06199, PXO 06201, and PXO 06202). Theother two Xoo strains KACC and MAFF3 have the same set of flagel-lar genes except for a few encoding hypothetical proteins (Table S1).Interestingly, PXO99A is unique in that it has two identical copiesof the flagellar genes due to a large DNA duplication (Salzberg et al.,2008).
Supplementary Table S1 related to this article can befound, in the online version, at http://dx.doi.org/10.1016/j.micres.2014.07.002.
3.2. RpoN2 and FleQ were required for motility of Xoo
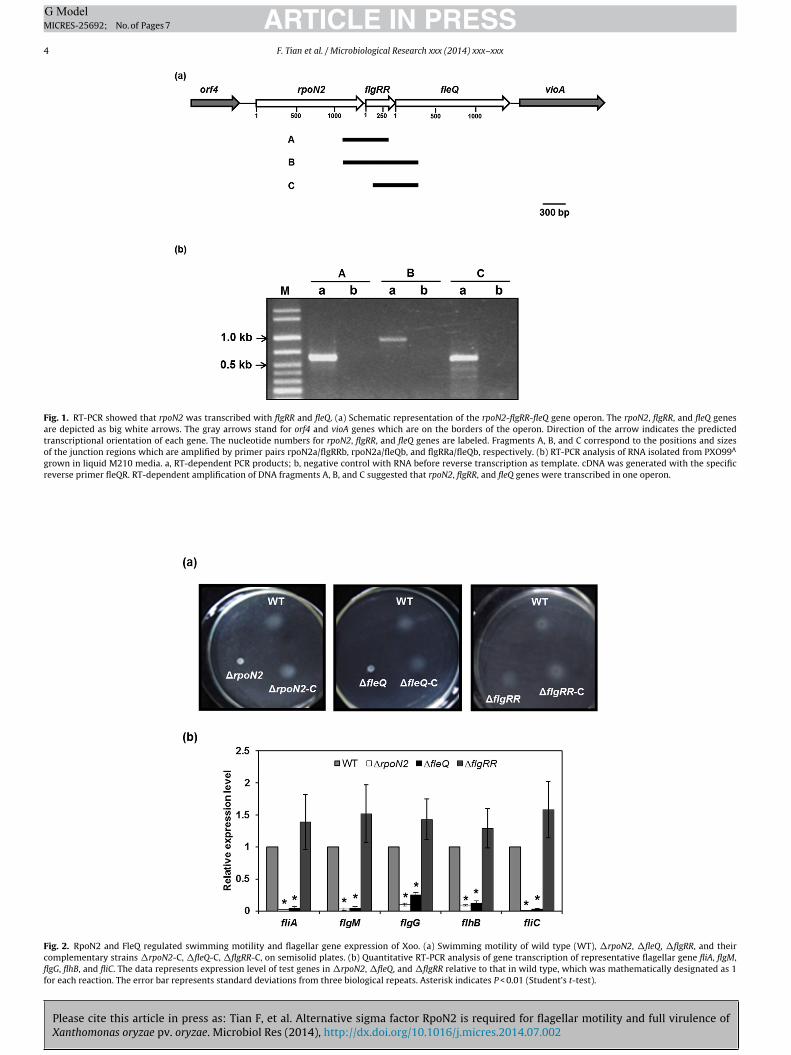
�54 encoded by rpoN gene has been show to regulate flagellargene expression and motility in several bacterial species (Proutyet al., 2001; Dasgupta et al., 2003). There are two rpoN homologs inthe genome of PXO99A, and their functions in Xoo have not beencharacterized yet. rpoN2 is located in the flagellar regulon, whilerpoN (PXO 02227) is the more ancestral copy found in most bacterialspecies. The nucleotide sequences of rpoN and rpoN2 do not sharemuch similarity, and their protein sequences share 39% identity.Downstream of rpoN2, there is a gene encoding an orphan responseregulator, named as flgRR, and a transcription activator gene fleQ(Fig. 1a). The intergenic distances of rpoN2-flgRR and flgRR-fleQare 9 bp and −3 bp, respectively, indicating they probably form anoperon. To test it experimentally, we performed RT-PCR. The totalbacterial RNAs were extracted as described in the Materials andmethods. cDNA was obtained by reverse transcription using a spe-cific primer for fleQ, then PCR was carried out to amplify the targetDNA fragments. The junctions of rpoN2-flgRR, rpoN2-fleQ, and flgRR-fleQ were obtained using the above cDNA as template, indicatingthe three genes belong to one transcriptional unit (Fig. 1b).
To investigate whether RpoN2, FlgRR, or FleQ regulates flagel-
tor RpoN2 is required for flagellar motility and full virulence of/10.1016/j.micres.2014.07.002
lar motility of Xoo, we generated PXO99A mutants defective ineach gene. Since rpoN2 is the first gene in the operon, its mutationmight have a polar effect. To check whether that had happened,we examined the transcription of fleQ in rpoN2 mutant by RT-PCR.
Please cite this article in press as: Tian F, et al. Alternative sigma factor RpoN2 is required for flagellar motility and full virulence ofXanthomonas oryzae pv. oryzae. Microbiol Res (2014), http://dx.doi.org/10.1016/j.micres.2014.07.002
ARTICLE IN PRESSG ModelMICRES-25692; No. of Pages 7
4 F. Tian et al. / Microbiological Research xxx (2014) xxx–xxx
Fig. 1. RT-PCR showed that rpoN2 was transcribed with flgRR and fleQ. (a) Schematic representation of the rpoN2-flgRR-fleQ gene operon. The rpoN2, flgRR, and fleQ genesare depicted as big white arrows. The gray arrows stand for orf4 and vioA genes which are on the borders of the operon. Direction of the arrow indicates the predictedtranscriptional orientation of each gene. The nucleotide numbers for rpoN2, flgRR, and fleQ genes are labeled. Fragments A, B, and C correspond to the positions and sizesof the junction regions which are amplified by primer pairs rpoN2a/flgRRb, rpoN2a/fleQb, and flgRRa/fleQb, respectively. (b) RT-PCR analysis of RNA isolated from PXO99A
grown in liquid M210 media. a, RT-dependent PCR products; b, negative control with RNA before reverse transcription as template. cDNA was generated with the specificreverse primer fleQR. RT-dependent amplification of DNA fragments A, B, and C suggested that rpoN2, flgRR, and fleQ genes were transcribed in one operon.
Fig. 2. RpoN2 and FleQ regulated swimming motility and flagellar gene expression of Xoo. (a) Swimming motility of wild type (WT), �rpoN2, �fleQ, �flgRR, and theircomplementary strains �rpoN2-C, �fleQ-C, �flgRR-C, on semisolid plates. (b) Quantitative RT-PCR analysis of gene transcription of representative flagellar gene fliA, flgM,flgG, flhB, and fliC. The data represents expression level of test genes in �rpoN2, �fleQ, and �flgRR relative to that in wild type, which was mathematically designated as 1for each reaction. The error bar represents standard deviations from three biological repeats. Asterisk indicates P < 0.01 (Student’s t-test).

IN PRESSG ModelM
cal Research xxx (2014) xxx–xxx 5
Taatofliflppn
3fl
rtibrpnogbplflbg
3
ncvaTlwitssmmfl
ottwmpwwmrmse(wi
Fig. 3. Virulence test of various Xoo strains. Bacterial cells of wild type (WT),�rpoN2, �fleQ, �rpoN2-C, and �fleQ-C were inoculated onto susceptible rice plants(Oryza sativa L. cv. Nipponbare) by leaf clipping method. Disease symptoms (a)and lesion lengths (b) were recorded 14 days after inoculation. The data repre-
ARTICLEICRES-25692; No. of Pages 7
F. Tian et al. / Microbiologi
he results showed that expression of fleQ in rpoN2 mutant wass good as in wild type, suggesting that mutation of rpoN2 did notlter the transcription of downstream genes (data not shown). Wehen tested the swimming motility of rpoN2, flgRR, and fleQ mutantsn semisolid plates. Compared with wild-type strain, rpoN2 andeQ mutants showed much smaller swimming zones, suggest-ng that their motility were significantly impaired (Fig. 2a). WhilegRR mutant remained motile as the wild type. The motility defecthenotypes of rpoN2 and fleQ mutants were recovered in the com-lemented stains. These results suggested that RpoN2 and FleQ, butot FlgRR are required for flagellar motility of Xoo.
.3. RpoN2 and FleQ positively regulated the expression ofagellar genes
Since rpoN2 and fleQ were co-transcribed and both wereequired for flagellar motility, they might function synergisticallyo regulate flagellar gene expression in Xoo. Therefore, we exam-ned the transcription of flagellar genes in rpoN2 and fleQ mutantsy quantitative RT-PCR, including fliA (�28), flgM (anti-�28), whichepresent regulatory factors, and flgG (basal body rod), flhB (exportathway) and fliC (flagellin), which represent structural compo-ents. As shown in Fig. 2b, transcriptions of these genes in the rpoN2r fleQ mutants dropped to only 10–30% of the wild-type level, sug-esting expressions of these flagellar genes are strictly controlledy RpoN2 and FleQ. This is consistent with the above motility defecthenotypes of the rpoN2 and fleQ mutants. As expected, expression
evels of representative flagellar genes were not repressed in thegRR mutant. These results together indicated that RpoN2 and FleQ,ut not FlgRR, positively regulated motility by controlling flagellarene expression.
.4. RpoN2 contributed to the virulence of Xoo in rice
The relation between flagellar motility and Xoo virulence hasot been tested previously. rpoN2 and fleQ mutants were signifi-antly impaired in their flagellar motility. Were they also affected inirulence? We then tested the virulence of rpoN2 and fleQ mutantsnd their complemented strains on rice by leaf clipping method.he disease symptoms were recorded by photography and theesion lengths were measured 14 days post inoculation. Compared
ith the wild type, the fleQ mutant did not show much differencen its ability to cause disease on rice leaves (Fig. 3). Interestingly,he rpoN2 mutant caused much less severe disease symptoms andhorter lesions (Fig. 3), which were recovered in the complementedtrain. Since FleQ, which has a similar regulatory role on flagellarotility, did not contribute to virulence, we speculated that RpoN2ight regulate Xoo virulence through a manner independent of
agellar motility.To exclude the possibility that the reduced virulence phenotype
f the rpoN2 mutant was due its growth defect, we performed bac-erial growth assays to compare the growth rate between WT andhe rpoN2 mutant. As shown in Fig. 4a, no significant differenceas observed between them when growing in the nutrient-richedium M210. A nutrient-scarce medium XOM2 has been used
reviously to induce the hrp system of Xoo (Furutanin et al., 2003),hich was considered to mimic the growth conditions in planta. Weondered whether they grow differently in this plant-mimickingedium. However, no growth was detected for either WT or the
poN2 mutant, probably due to there was little carbon source in theedium. Therefore, we added 0.5% sucrose in XOM2 medium to
upport bacterial growth. Interestingly, the rpoN2 mutant showed
Please cite this article in press as: Tian F, et al. Alternative sigma facXanthomonas oryzae pv. oryzae. Microbiol Res (2014), http://dx.doi.org
nhanced growth rate than WT for an undermined mechanismFig. 4b). Overall, these results suggested that the rpoN2 mutantas not defective in growth, which further supported that RpoN2
s a novel virulence determinant in Xoo.
sent average and standard deviation of measured lesion lengths from 10 leaves.The experiment was repeated three times with similar results. Asterisk indicatesP < 0.05 (Student’s t-test).
3.5. A fliC mutant remained virulent as the wild type
In the above experiments, we showed both rpoN2 and fleQmutants were defective in flagellar motility, but only rpoN2 mutantwas impaired in virulence, implying that flagellar motility mightnot contribute to virulence directly. To further clarify it, we gen-erated a mutant knocking out the flagellin gene fliC. As expected,the PXO99A fliC mutant was not motile (Fig. 5a). The motility wasrestored in the complemented strain expressing fliC gene on a plas-mid in trans. Then the virulence of fliC mutant was tested on riceleaves. The disease phenotypes and lesion lengths were recorded14 days post inoculation (Fig. 5b and c). Compared with that of wildtype, the disease symptoms of the fliC mutant did not change much.The average lesion length was slightly increased (statistically sig-nificant), while that of the complemented strain was restored towild-type level. This result supported the statement that motilitywas not required for the virulence of Xoo in rice leaves.
4. Discussion
Alternative sigma factor RpoN has been implicated as a mul-tifunctional regulator in controlling various bacterial phenotypes
tor RpoN2 is required for flagellar motility and full virulence of/10.1016/j.micres.2014.07.002
such as nitrogen assimilation, motility, stress resistance, and vir-ulence factor production (Reitzer and Schneider, 2001; Dong andMekalanos, 2012; Ancona et al., 2014). Different from the othersigma factors, RpoN requires an additional transcriptional factor to

ARTICLE ING ModelMICRES-25692; No. of Pages 7
6 F. Tian et al. / Microbiological Re
Fig. 4. Growth curves of wild type (WT) and �rpoN2 in rich media M210 (a)and plant-mimicking media XOM2 plus 0.5% sucrose (b). Bacterial growth wasdetermined by measuring OD600 at 12-h intervals with cultures starting from con-centration of OD600 at 0.1. Each experiment was performed with three biologicalrd
iFtDflwXsInflac
that the fliC mutant caused increased lesion length on rice leaves
F�i
eplicates and each repeat was assayed in triplicate. Error bars indicate standardeviation.
nitiate transcription (Reitzer and Schneider, 2001). For example,leQ has been shown as an RpoN-dependent transcriptional factoro regulate bacterial motility in several species (Prouty et al., 2001;asgupta et al., 2003). Here, we demonstrated that the rpoN2 andeQ were co-transcribed in the same operon (Fig. 1), and they bothere required for flagellar motility and flagellar gene expression ofoo (Fig. 2). In P. aeruginosa, the alternative sigma factor fliA washown to be constitutively expressed, thus was grouped as class
(Dasgupta et al., 2003). We found that expression of fliA and itsegative regulator flgM were significantly reduced in rpoN2 and
Please cite this article in press as: Tian F, et al. Alternative sigma facXanthomonas oryzae pv. oryzae. Microbiol Res (2014), http://dx.doi.org
eQ mutants, suggesting fliA and flgM were under control of RpoN2nd FleQ, and may belong to class II in the flagellar gene expressionascade of Xoo. This results is consistent with the previous report
ig. 5. Motility and virulence phenotypes of �fliC. (a) �fliC lost motility, which was restfliC compared with wild type. (c) The average disease lesion length caused by �fliC wa
ndicates P < 0.05 (Student’s t-test).
PRESSsearch xxx (2014) xxx–xxx
of X. campestris pv. campestris, in which a three-level hierarchy offlagellar gene expression model was proposed (Yang et al., 2009).
Although most bacteria carry only one copy of rpoN gene,there are some species that have two or more copies. Rhodobac-ter sphaeroides is a unique example in which four different rpoNgenes were identified (Poggio et al., 2002). The function of each rpoNgene was not interchangeable, suggesting there is specialization ofrpoN genes during evolution (Poggio et al., 2002; Domenzain et al.,2012). Two copies of rpoN genes exist in the genome of Xoo. Muta-tion of rpoN2 has resulted in loss of motility and reduced virulence(Figs. 2 and 3), suggesting the two rpoN genes may not be func-tionally redundant. In addition, the rpoN2 mutant demonstratedenhanced growth rate than the wild type in the plant-mimickingmedium (Fig. 4b). These results implied that rpoN2 might playmultiple roles in Xoo. By identifying the regulon of rpoN2 gene atgenome-wide level will help to reveal its regulatory mechanisms.
Whether flagellar motility contributes to the pathogenesis ofplant bacterial pathogens varies in different species. For Ralsto-nia solanacearum, which causes bacterial wilt disease on manyplants, flagellar motility seemed to be an important virulence factorduring the early infection stage (Tans-Kersten et al., 2001, 2004).Flagellum was also required for biofilm formation and pathogeni-city of X. axonopodis pv. citri, the causative agent of citrus canker(Malamud et al., 2011, 2013). However, it was also noticed thatmost bacterial cells were non-motile once inside of the host xylem(Tans-Kersten et al., 2004). Here we showed that the fleQ and fliCmutants of Xoo were not impaired in causing leaf blight disease onrice (Figs. 3 and 5), suggesting that flagellar motility might not beessential for the virulence of Xoo. Recently, the published genomesequence of X. fuscans subsp. fuscans strain 4834-R revealed thatit harbors an incomplete flagellar gene cluster and is non-motile(Darrasse et al., 2013), which further implicated that flagellar motil-ity might not be essential for the fitness of plant pathogens. If so,the question will be what drives the bacterial cells to move insidethe host tissues. More work in the future will be needed to answerthis question.
In Arabidopsis, receptor kinase FLS2 mediated PTI induced bybacterial flagellin through directly interacting with flg22, the pep-tide representing the elicitor-active epitope of flagellin (Zipfel et al.,2004; Chinchilla et al., 2006). OsFLS2, the rice ortholog of Arabidop-sis FLS2, demonstrated similar activity by complementing the fls2mutant of Arabidopsis (Takai et al., 2008). However, it was not con-firmed that OsFLS2 could recognize the flagellin from rice bacterialpathogens such as Acidovorax avenae (Takai et al., 2008). Whetheror not flagellin from Xoo triggers PTI in rice is not clear. We observed
tor RpoN2 is required for flagellar motility and full virulence of/10.1016/j.micres.2014.07.002
than the wild type (Fig. 5), which might be due to reduced defenseresponses triggered by the mutant during infection. Compara-tive transcriptome analysis revealed some rice genes expressed
ored in �fliC-C. (b) No significant difference observed on the disease symptoms ofs slightly bigger than that of the wild type, which was restored in �fliC-C. Asterisk

ING ModelM
cal Res
d(w
A
wgRC
R
A
A
A
B
B
B
C
D
D
D
D
D
F
F
G
H
H
H
H
H
K
K
ARTICLEICRES-25692; No. of Pages 7
F. Tian et al. / Microbiologi
ifferentially in responding to wild type PXO99A and the fliC mutantunpublished data). Nevertheless, how bacterial flagellin interactith rice cells remains to be an intriguing question.
cknowledgements
We thank Dr. Ching-Hong Yang for helpful discussion. This workas supported by grants from the National Basic Research Pro-
ram of China (2011CB100700) and the National High Technologyesearch and Development Program of China (2012AA101504) to.H.
eferences
ldridge P, Hughes KT. Regulation of flagellar assembly. Curr Opin Microbiol2002;5(2):160–5.
miel E, Lovewell RR, O’Toole GA, Hogan DA, Berwin B. Pseudomonas aeruginosaevasion of phagocytosis is mediated by loss of swimming motility and is inde-pendent of flagellum expression. Infect Immun 2010;78(7):2937–45.
ncona V, Li W, Zhao Y. Alternative sigma factor RpoN and its modulation pro-tein YhbH are indispensable for Erwinia amylovora virulence. Mol Plant Pathol2014;15(1):58–66.
ahar O, Levi N, Burdman S. The cucurbit pathogenic bacterium Acidovorax cit-rulli requires a polar flagellum for full virulence before and after host-tissuepenetration. Mol Plant-Microbe Interact 2011;24(9):1040–50.
eattie GA, Lindow SE. The secret life of foliar bacterial pathogens on leaves. AnnuRev Phytopathol 1995;33:145–72.
ourret RB, Charon NW, Stock AM, West AH. Bright lights, abundantoperons—fluorescence and genomic technologies advance studies of bac-terial locomotion and signal transduction: review of the BLAST meeting,Cuernavaca, Mexico, 14 to 19 January 2001. J Bacteriol 2002;184(1):1–17.
hinchilla D, Bauer Z, Regenass M, Boller T, Felix G. The Arabidopsis receptor kinaseFLS2 binds flg22 and determines the specificity of flagellin perception. Plant Cell2006;18(2):465–76.
arrasse A, Carrere S, Barbe V, Boureau T, Arrieta-Ortiz ML, Bonneau S, et al. Genomesequence of Xanthomonas fuscans subsp. fuscans strain 4834-R reveals thatflagellar motility is not a general feature of xanthomonads. BMC Genomics2013;14(1):761.
as A, Rangaraj N, Sonti RV. Multiple adhesin-like functions of Xanthomonas oryzaepv. oryzae are involved in promoting leaf attachment, entry, and virulence onrice. Mol Plant-Microbe Interact 2009;22(1):73–85.
asgupta N, Wolfgang MC, Goodman AL, Arora SK, Jyot J, Lory S, et al. A four-tieredtranscriptional regulatory circuit controls flagellar biogenesis in Pseudomonasaeruginosa. Mol Microbiol 2003;50(3):809–24.
omenzain C, Camarena L, Osorio A, Dreyfus G, Poggio S. Evolutionary origin of theRhodobacter sphaeroides specialized RpoN sigma factors. FEMS Microbiol Lett2012;327(2):93–102.
ong TG, Mekalanos JJ. Characterization of the RpoN regulon reveals differentialregulation of T6SS and new flagellar operons in Vibrio cholerae O37 strain V52.Nucleic Acids Res 2012;40(16):7766–75.
ranchi L, Amer A, Body-Malapel M, Kanneganti TD, Ozoren N, Jagirdar R, et al.Cytosolic flagellin requires Ipaf for activation of caspase-1 and interleukin 1betain salmonella-infected macrophages. Nat Immunol 2006;7(6):576–82.
urutanin A, Tsuge S, Oku T, Tsuno K, Inoue Y, Ochiai H, et al. Hpa1 secretion viatype III secretion system in Xanthomonas oryzae pv. oryzae. J Gen Plant Pathol2003;69:271–5.
iron JA, Torres AG, Freer E, Kaper JB. The flagella of enteropathogenic Escherichia colimediate adherence to epithelial cells. Mol Microbiol 2002;44(2):361–79.
aefele DM, Lindow SE. Flagellar motility confers epiphytic fitness advantages uponPseudomonas syringae. Appl Environ Microbiol 1987;53(10):2528–33.
anahan D. Studies on transformation of Escherichia coli with plasmids. J Mol Biol1983;166(4):557–80.
arshey RM. Bacterial motility on a surface: many ways to a common goal. AnnuRev Microbiol 2003;57:249–73.
e YW, Wu J, Cha JS, Zhang LH. Rice bacterial blight pathogen Xanthomonas oryzaepv. oryzae produces multiple DSF-family signals in regulation of virulence factorproduction. BMC Microbiol 2010;10:187.
opkins CM, White FF, Choi SH, Guo A, Leach JE. Identification of a family of avir-ulence genes from Xanthomonas oryzae pv. oryzae. Mol Plant-Microbe Interact1992;5(6):451–9.
arlinsey JE, Tanaka S, Bettenworth V, Yamaguchi S, Boos W, Aizawa SI, et al.Completion of the hook-basal body complex of the Salmonella typhimurium
Please cite this article in press as: Tian F, et al. Alternative sigma facXanthomonas oryzae pv. oryzae. Microbiol Res (2014), http://dx.doi.org
flagellum is coupled to FlgM secretion and fliC transcription. Mol Microbiol2000;37(5):1220–31.
ovach ME, Elzer PH, Hill DS, Robertson GT, Farris MA, Roop RM 2nd, et al. Fournew derivatives of the broad-host-range cloning vector pBBR1MCS, carryingdifferent antibiotic-resistance cassettes. Gene 1995;166(1):175–6.
PRESSearch xxx (2014) xxx–xxx 7
Lee BM, Park YJ, Park DS, Kang HW, Kim JG, Song ES, et al. The genome sequence ofXanthomonas oryzae pathovar oryzae KACC10331, the bacterial blight pathogenof rice. Nucleic Acids Res 2005;33(2):577–86.
Liu X, Matsumura P. The FlhD/FlhC complex, a transcriptional activator of theEscherichia coli flagellar class II operons. J Bacteriol 1994;176(23):7345–51.
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(−Delta Delta C(T)) method. Methods2001;25(4):402–8.
Macnab RM. Type III flagellar protein export and flagellar assembly. Biochim BiophysActa 2004;1694(1–3):207–17.
Malamud F, Homem RA, Conforte VP, Yaryura PM, Castagnaro AP, Marano MR, et al.Identification and characterization of biofilm formation-defective mutants ofXanthomonas citri subsp. citri. Microbiology 2013;159(9):1911–9.
Malamud F, Torres PS, Roeschlin R, Rigano LA, Enrique R, Bonomi HR, et al. TheXanthomonas axonopodis pv. citri flagellum is required for mature biofilm andcanker development. Microbiology 2011;157(3):819–29.
McCarter LL. Regulation of flagella. Curr Opin Microbiol 2006;9(2):180–6.Ottemann KM, Lowenthal AC. Helicobacter pylori uses motility for initial coloniza-
tion and to attain robust infection. Infect Immun 2002;70(4):1984–90.Poggio S, Osorio A, Dreyfus G, Camarena L. The four different sigma(54) factors
of Rhodobacter sphaeroides are not functionally interchangeable. Mol Microbiol2002;46(1):75–85.
Porter SL, Wadhams GH, Armitage JP. Signal processing in complex chemotaxispathways. Nat Rev 2011;9(3):153–65.
Prouty MG, Correa NE, Klose KE. The novel sigma54- and sigma28-dependentflagellar gene transcription hierarchy of Vibrio cholerae. Mol Microbiol2001;39(6):1595–609.
Ray SK, Rajeshwari R, Sonti RV. Mutants of Xanthomonas oryzae pv. oryzae defi-cient in general secretory pathway are virulence deficient and unable to secretexylanase. Mol Plant-Microbe Interact 2000;13(4):394–401.
Reitzer L, Schneider BL. Metabolic context and possible physiological themesof sigma(54)-dependent genes in Escherichia coli. Microbiol Mol Biol Rev2001;65(3):422–44.
Salzberg SL, Sommer DD, Schatz MC, Phillippy AM, Rabinowicz PD, Tsuge S, et al.Genome sequence and rapid evolution of the rice pathogen Xanthomonas oryzaepv. oryzae PXO99A. BMC Genomics 2008;9:204.
Sambrook J, Russell DW. Molecular cloning: a laboratory manual. Cold Spring Harbor,NY: Cold Spring Harbor Laboratory Press; 2001.
Schafer A, Tauch A, Jager W, Kalinowski J, Thierbach G, Puhler A. Small mobilizablemulti-purpose cloning vectors derived from the Escherichia coli plasmids pK18and pK19: selection of defined deletions in the chromosome of Corynebacteriumglutamicum. Gene 1994;145(1):69–73.
Shen Y, Chern M, Silva FG, Ronald P. Isolation of a Xanthomonas oryzae pv. oryzae fla-gellar operon region and molecular characterization of flhF. Mol Plant-MicrobeInteract 2001;14(2):204–13.
Song C, Yang B. Mutagenesis of 18 type III effectors reveals virulence function ofXopZ(PXO99) in Xanthomonas oryzae pv. oryzae. Mol Plant-Microbe Interact2010;23(7):893–902.
Sorenson MK, Ray SS, Darst SA. Crystal structure of the flagellar sigma/anti-sigmacomplex sigma(28)/FlgM reveals an intact sigma factor in an inactive conforma-tion. Mol Cell 2004;14(1):127–38.
Subramoni S, Pandey A, Vishnu Priya MR, Patel HK, Sonti RV. The ColRS system ofXanthomonas oryzae pv. oryzae is required for virulence and growth in iron-limiting conditions. Mol Plant Pathol 2012;13(7):690–703.
Takai R, Isogai A, Takayama S, Che FS. Analysis of flagellin perception mediatedby flg22 receptor OsFLS2 in rice. Mol Plant-Microbe Interact 2008;21(12):1635–42.
Tans-Kersten J, Huang H, Allen C. Ralstonia solanacearum needs motility for invasivevirulence on tomato. J Bacteriol 2001;183(12):3597–605.
Tans-Kersten J, Brown D, Allen C. Swimming motility, a virulence trait of Ralstoniasolanacearum, is regulated by FlhDC and the plant host environment. Mol Plant-Microbe Interact 2004;17(6):686–95.
Tsuge S, Terashima S, Furutani A, Ochiai H, Oku T, Tsuno K, et al. Effects on promoteractivity of base substitutions in the cis-acting regulatory element of HrpXo reg-ulons in Xanthomonas oryzae pv. oryzae. J Bacteriol 2005;187(7):2308–14.
Tsuge S, Nakayama T, Terashima S, Ochiai H, Furutani A, Oku T, et al. Gene involvedin transcriptional activation of the hrp regulatory gene hrpG in Xanthomonasoryzae pv. oryzae. J Bacteriol 2006;188(11):4158–62.
White F, Yang B. Host and pathogen factors controlling the rice–Xanthomonas oryzaeinteraction. Plant Physiol 2009;150(4):1677–86.
Yang F, Tian F, Sun L, Chen H, Wu M, Yang C-H, et al. A novel two-component systemPdeK/PdeR regulates c-di-GMP turnover and virulence of Xanthomonas oryzaepv. oryzae. Mol Plant-Microbe Interact 2012;25(10):1361–9.
Yang TC, Leu YW, Chang-Chien HC, Hu RM. Flagellar biogenesis of Xanthomonascampestris requires the alternative sigma factors RpoN2 and FliA and is tempo-rally regulated by FlhA, FlhB, and FlgM. J Bacteriol 2009;191(7):2266–75.
tor RpoN2 is required for flagellar motility and full virulence of/10.1016/j.micres.2014.07.002
Zipfel C, Felix G. Plants and animals: a different taste for microbes? Curr Opin PlantBiol 2005;8(4):353–60.
Zipfel C, Robatzek S, Navarro L, Oakeley EJ, Jones JD, Felix G, et al. Bacte-rial disease resistance in Arabidopsis through flagellin perception. Nature2004;428(6984):764–7.




















