
Type III-dependent translocation of the Xanthomonas AvrBs3 protein into the plant cell
11
Molecular Microbiology (2002) 46 (1), 13–23 © 2002 Blackwell Publishing Ltd Blackwell Science, LtdOxford, UKMMIMolecular Microbiology0950-382XBlackwell Science, 200245Original Article Translocation of AvrBs3 into the plant cellB. Szurek, O. Rossier, G. Hause and U. Bonas Accepted 19 June, 2002. *For correspondence. E-mail bonas@ genetik.uni-halle.de; Tel. (+49) 345 55 26290; Fax (+49) 345 55 27277. Type III-dependent translocation of the Xanthomonas AvrBs3 protein into the plant cell Boris Szurek, 1,2 Ombeline Rossier, 1,3 Gerd Hause 4 and Ulla Bonas 1 * 1 Institut für Genetik, Martin-Luther-Universität Halle- Wittenberg, Halle, Germany. 2 INRA-URGV, Evry, France. 3 Northwestern University, Department of Microbiology and Immunology, Chicago, Illinois, USA. 4 Biozentrum der Universität, Halle, Germany. Summary Many plant pathogenic bacteria utilize a conserved type III secretion system (TTSS) to deliver effector proteins into the host tissue. Indirect evidence has suggested that at least some effector proteins are translocated from the bacterial cytoplasm into the plant cell. Using an immunocytochemical approach, we demonstrate that the type III effector AvrBs3 from Xanthomonas campestris pv. vesicatoria localizes to nuclei of infected pepper leaves. Importantly, AvrBs3 translocation was observed in situ in native tissues of susceptible and resistant plants. AvrBs3 was detected in the nucleus as soon as 4 h post infection, which was dependent on a functional TTSS and the putative translocator HrpF. N-terminal AvrBs3 dele- tion derivatives are no longer secreted by the TTSS in vitro and could not be detected inside the host cells, suggesting that the N-terminus of AvrBs3 is important for secretion. Deletion of the nuclear localization sig- nals in the AvrBs3 C-terminus, which are required for the AvrBs3-mediated induction of the hypersensitive reaction in resistant pepper plants, abolished AvrBs3 localization to the nucleus. This is the first report on direct evidence for translocation of a native type III effector protein from a plant pathogenic bacterium into the host cell. Introduction To interact with their host, many Gram-negative patho- genic bacteria have evolved a specialized type III protein secretion system (TTSS) that transports proteins across both bacterial membranes without cleavage of a classical signal peptide. It has been shown for a number of animal pathogenic bacteria that so-called effector proteins are even translocated directly into the host cell via the TTSS (Hueck, 1998). In phytopathogens such as Pseudomonas syringae , Xanthomonas subsp., Ralstonia solanacearum and Erwinia amylovora , the TTSS is encoded by hrp (h ypersensitive r eaction and p athogenicity) genes, nine of which are highly conserved in plant and animal pathogens ( hrc genes, for hrp conserved; Bogdanove et al ., 1996). Loss of hrp gene function results in a pleiotropic pheno- type: hrp mutants are unable to grow in the plant, no longer cause disease symptoms and fail to induce the hypersensitive reaction (HR) in resistant host and non- host plants (Lindgren, 1997). The HR is a rapid local, programmed cell death that is induced upon recognition of the pathogen and is concomitant with the inhibition of pathogen growth within the infected plant tissue (Klement, 1982). Whereas the core components of TTSS are highly conserved among plant and animal pathogenic bacteria, their secreted substrates appear to be extremely diverse (Cornelis and Van Gijsegem, 2000). Secreted proteins of plant pathogens include proteins essential for the function of the secretion machinery (e.g. Hrp pilus subunit; Romantschuk et al ., 2001), pathogenicity proteins, as well as triggers of the plant defence reaction, for example, harpins and avirulence (Avr) proteins (Kjemtrup et al ., 2000). Avr proteins were originally defined by their HR- inducing activity in plants that express a corresponding disease resistance ( R ) gene. In the absence of the avr or the R gene or both, no recognition takes place and dis- ease occurs (Bonas and Van den Ackerveken, 1999). However, it has become clear that the primary function of many if not all avr genes is in virulence, and that plants have probably evolved the capacity to recognize these pathogen-derived molecules. The subject of this study is the effector AvrBs3 from Xanthomonas campestris pv. vesicatoria ( Xcv ), the causal agent of bacterial spot disease on pepper and tomato. Bacteria enter the leaves via stomata or wounds and colonize the intercellular spaces of the mesophyll of sus- ceptible plants. Early disease symptoms probably corre- late with the release of water and nutrients which benefit
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Type III-dependent translocation of the Xanthomonas AvrBs3 protein into the plant cell

Molecular Microbiology (2002)
46
(1), 13–23
© 2002 Blackwell Publishing Ltd
Blackwell Science, LtdOxford, UKMMIMolecular Microbiology0950-382XBlackwell Science, 200245Original Article
Translocation of AvrBs3 into the plant cellB. Szurek, O. Rossier, G. Hause and U. Bonas
Accepted 19 June, 2002. *For correspondence. E-mail [email protected]; Tel. (+49) 345 55 26290; Fax (+49) 345 5527277.
Type III-dependent translocation of the
Xanthomonas
AvrBs3 protein into the plant cell
Boris Szurek,
1,2
Ombeline Rossier,
1,3
Gerd Hause
4
and Ulla Bonas
1
*
1
Institut für Genetik, Martin-Luther-Universität Halle-Wittenberg, Halle, Germany.
2
INRA-URGV, Evry, France.
3
Northwestern University, Department of Microbiology and Immunology, Chicago, Illinois, USA.
4
Biozentrum der Universität, Halle, Germany.
Summary
Many plant pathogenic bacteria utilize a conservedtype III secretion system (TTSS) to deliver effectorproteins into the host tissue. Indirect evidence hassuggested that at least some effector proteins aretranslocated from the bacterial cytoplasm into theplant cell. Using an immunocytochemical approach,we demonstrate that the type III effector AvrBs3 from
Xanthomonas campestris
pv.
vesicatoria
localizes tonuclei of infected pepper leaves. Importantly, AvrBs3translocation was observed
in situ
in native tissuesof susceptible and resistant plants. AvrBs3 wasdetected in the nucleus as soon as 4 h post infection,which was dependent on a functional TTSS and theputative translocator HrpF. N-terminal AvrBs3 dele-tion derivatives are no longer secreted by the TTSS
invitro
and could not be detected inside the host cells,suggesting that the N-terminus of AvrBs3 is importantfor secretion. Deletion of the nuclear localization sig-nals in the AvrBs3 C-terminus, which are required forthe AvrBs3-mediated induction of the hypersensitivereaction in resistant pepper plants, abolished AvrBs3localization to the nucleus. This is the first report ondirect evidence for translocation of a native type IIIeffector protein from a plant pathogenic bacteriuminto the host cell.
Introduction
To interact with their host, many Gram-negative patho-genic bacteria have evolved a specialized type III protein
secretion system (TTSS) that transports proteins acrossboth bacterial membranes without cleavage of a classicalsignal peptide. It has been shown for a number of animalpathogenic bacteria that so-called effector proteins areeven translocated directly into the host cell via the TTSS(Hueck, 1998). In phytopathogens such as
Pseudomonassyringae
,
Xanthomonas
subsp.,
Ralstonia solanacearum
and
Erwinia amylovora
, the TTSS is encoded by
hrp
(hypersensitive reaction and pathogenicity) genes, nine ofwhich are highly conserved in plant and animal pathogens(
hrc
genes, for
hrp
conserved; Bogdanove
et al
., 1996).Loss of
hrp
gene function results in a pleiotropic pheno-type:
hrp
mutants are unable to grow in the plant, nolonger cause disease symptoms and fail to induce thehypersensitive reaction (HR) in resistant host and non-host plants (Lindgren, 1997). The HR is a rapid local,programmed cell death that is induced upon recognitionof the pathogen and is concomitant with the inhibition ofpathogen growth within the infected plant tissue (Klement,1982). Whereas the core components of TTSS are highlyconserved among plant and animal pathogenic bacteria,their secreted substrates appear to be extremely diverse(Cornelis and Van Gijsegem, 2000). Secreted proteins ofplant pathogens include proteins essential for the functionof the secretion machinery (e.g. Hrp pilus subunit;Romantschuk
et al
., 2001), pathogenicity proteins, as wellas triggers of the plant defence reaction, for example,harpins and avirulence (Avr) proteins (Kjemtrup
et al
.,2000). Avr proteins were originally defined by their HR-inducing activity in plants that express a correspondingdisease resistance (
R
) gene. In the absence of the
avr
orthe
R
gene or both, no recognition takes place and dis-ease occurs (Bonas and Van den Ackerveken, 1999).However, it has become clear that the primary function ofmany if not all
avr
genes is in virulence, and that plantshave probably evolved the capacity to recognize thesepathogen-derived molecules.
The subject of this study is the effector AvrBs3 from
Xanthomonas campestris
pv.
vesicatoria
(
Xcv
), the causalagent of bacterial spot disease on pepper and tomato.Bacteria enter the leaves via stomata or wounds andcolonize the intercellular spaces of the mesophyll of sus-ceptible plants. Early disease symptoms probably corre-late with the release of water and nutrients which benefit

14
B. Szurek, O. Rossier, G. Hause and U. Bonas
© 2002 Blackwell Publishing Ltd,
Molecular Microbiology
,
46
, 13–23
bacterial multiplication (Innes, 2001). The ability of
Xcv
tocause disease in susceptible plants and to induce the HRin resistant plants depends on the
hrp
gene cluster, whichencodes a TTSS (Bonas
et al
., 1991; Fenselau
et al
.,1992; Fenselau and Bonas, 1995; Huguet and Bonas,1997; Rossier
et al
., 2000). Type III-dependent secretion
in vitro
has recently been shown for several pathogenicityand Avr proteins from
Xcv
(Rossier
et al
., 1999; Mudgett
et al
., 2000; Rossier
et al
., 2000; Escolar
et al
., 2001).Among these proteins is AvrBs3, which triggers recogni-tion of the bacterium by pepper plants containing theresistance gene
Bs3
(Minsavage
et al
., 1990).The AvrBs3 protein belongs to a large family of con-
served proteins in species of
Xanthomonas
, which oftenplay a role in both virulence and avirulence (Swarup
et al
.,1991; 1992; Bai
et al
., 2000; Yang
et al
., 2000; Marois
et al
., 2002). An intriguing structural feature of membersof the AvrBs3 protein family is their central region, whichis composed of a variable number of nearly identical 34amino acid tandem repeats (Vivian and Arnold, 2000). The17.5 repeats of the 122 kDa AvrBs3 protein are essentialfor recognition by the
Bs3
resistance gene (Herbers
et al
.,1992) and induction of plant cell hypertrophy in suscep-tible plants (Marois
et al
., 2002). Interestingly, the C-terminal region of AvrBs3 and its homologues containstypical eukaryotic features: nuclear localization signals(NLSs) and an acidic transcription activation domain(AAD) (Yang and Gabriel, 1995; Van den Ackerveken
et al
., 1996; Zhu
et al
., 1998; Szurek
et al
., 2001). Tran-sient expression of a protein fusion between the AvrBs3C-terminus and
b
-glucuronidase (GUS) in onion epidermiscells resulted in nuclear GUS activity, indicating that theNLSs are functional (Van den Ackerveken
et al
., 1996).As AvrBs3 is secreted by the
Xcv
TTSS
in vitro
(Rossier
et al
., 1999) and the NLSs and AAD are essential for bothHR elicitation and the induction of cell hypertrophy (Vanden Ackerveken
et al
., 1996; Zhu
et al
., 1998; Szurek
et al
., 2001; Marois
et al
., 2002), AvrBs3 is likely to betranslocated into the host cell through the TTSS.
Translocation of type III effector proteins into host cellshas been demonstrated for animal pathogenic bacteria(Staskawicz
et al
., 2001), using several experimentalapproaches, for example, translational fusions to adeny-late cyclase (cya) (Sory and Cornelis, 1994), subcellularfractionation and immunofluorescence microscopy(Rosqvist
et al
., 1994; Lee
et al
., 1998). In plants, thereis currently only indirect evidence for type III effector‘injection’, the strongest being the observation that aspecific HR is induced when an
avr
gene (e.g.
avrBs3
)is expressed in resistant plant cells (Bonas and Van denAckerveken, 1999). Furthermore, the
Pseudomonassyringae
type III effector AvrRpt2 is processed duringinfection of
Arabidopsis thaliana
, and this processingwas shown to take place in the plant and to be TTSS-
dependent (Mudgett and Staskawicz, 1999). Additionalsupport for the Avr protein translocation hypothesis camefrom subcellular localization studies of
P. syringae
Avrproteins after their expression in plant cells. It wasshown that N-myristoylation motifs, which are importantfor HR-triggering activity, determine localization of the Avrprotein in the plant plasma membrane (Nimchuk
et al
.,2000; Shan
et al
., 2000). Moreover, several Avr proteinsinteract with intracellular plant proteins in yeast,
in vitro
and
in vivo
(Scofield
et al
., 1996; Tang
et al
., 1996; Leisterand Katagiri, 2000; Szurek
et al
., 2001; Mackey
et al
.,2002).
In this study, we provide direct evidence for TTSS-dependent translocation of AvrBs3 from
Xanthomonas
into infected plant cells using immunocytochemistry.Detection of AvrBs3, which is targeted to the nucleus ofpepper cells, was shown to require a functional TTSS, thepotential translocator HrpF and N-terminal sequences inAvrBs3.
Results
The N-terminus of AvrBs3 is essential for type III secretion but not for HR induction
In many type III effector proteins, signals for secretion/translocation have been mapped to the amino-terminalregion (Sory
et al
., 1995; Schesser
et al
., 1996; Mudgett
et al
., 2000; Lloyd
et al
., 2001). To address whethersequences essential for type III secretion of AvrBs3 arealso localized in the amino terminus, we deleted aminoacids 3–27 (AvrBs3
D
1) and 2–153 (AvrBs3
D
2) fromAvrBs3 respectively (Fig. 1A). The
avrBs3
gene and itsderivatives were expressed in
Xcv
strain 85* from the
lac
promoter in plasmid pDSK602, which is constitutive in
Xcv
(Murillo
et al
., 1994; Table 1). In total protein extracts, bothtruncated AvrBs3 derivatives were present although inlower amounts than the wild-type protein (Fig. 1B). Tocompensate for differences in protein stability, loading inFig. 1B (lane 4) was adjusted to obtain similar signal inten-sities (data not shown for AvrBs3
D
2). However, only thewild-type AvrBs3 protein was detected in culture superna-tants, indicating that sequences in the N-terminal regionof AvrBs3 are required for secretion (Fig. 1B, lanes 5 and8). In contrast, deletion of 83 amino acids encompassingthe NLS region in the C-terminus, which abolishes AvrBs3activity (Van den Ackerveken
et al
., 1996), did not affectAvrBs3 secretion (Fig. 1B, lane 7). As reported previously,secretion of AvrBs3 was abolished in the TTSS-mutantstrain 85*
D
hrcV
(Fig. 1B, lane 6), which lacks a conservedinner membrane protein of the core TTSS (Rossier
et al
.,1999). Infection of resistant pepper plants (ECW-30R)with strain 85* expressing either AvrBs3
D
1 or AvrBs3
D
2failed to induce the HR (Table 2). However, expression ofthe truncated
avrBs3
alleles in the plant cell using
Agro-

Translocation of AvrBs3 into the plant cell
15
© 2002 Blackwell Publishing Ltd,
Molecular Microbiology
,
46
, 13–23
bacterium tumefaciens
-mediated gene transfer resulted inthe HR induction (Table 2). This demonstrates that, onceinside the plant cell, AvrBs3 activity, i.e. recognition by
Bs3, does not require the 152 N-terminal amino acids.Taken together, these data suggest that N-terminal dele-tions of AvrBs3 abolish both its secretion and transloca-tion via the TTSS.
Immunodetection of AvrBs3 in pepper leaf tissue after transient expression
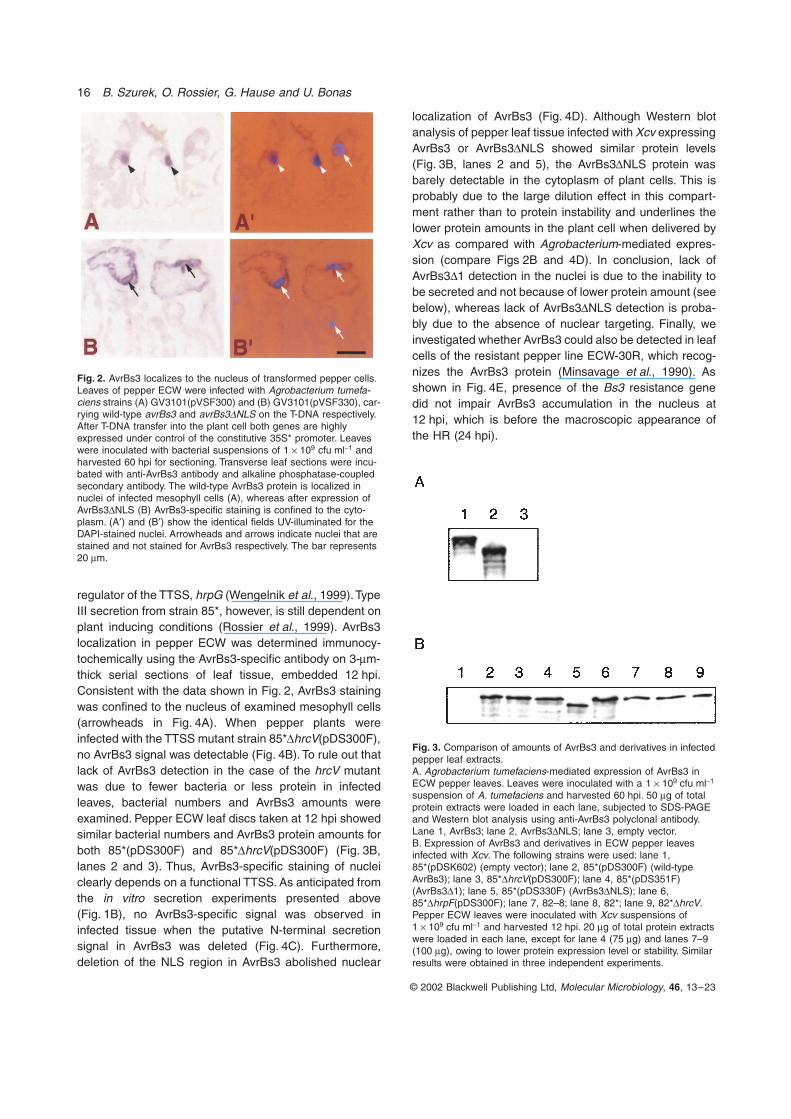
Next, we tested whether AvrBs3 could be detected inplant cells using immunocytochemistry. As proteinamounts delivered by Xcv are believed to be low, we firstestablished immunocytochemistry protocols for peppercells expressing avrBs3 via Agrobacterium-mediatedgene transfer under control of the constitutive cauliflowermosaic virus 35S* promoter. This promoter contains abacterial transcription terminator resulting in undetectableexpression in bacteria (Van den Ackerveken et al., 1996).The subcellular localization of AvrBs3 was determined inserial transverse sections of susceptible pepper (ECW)leaf tissue expressing avrBs3, using an AvrBs3-specificpolyclonal antibody and alkaline phosphatase-coupledsecondary antibody (see Experimental procedures).DNA-specific staining with DAPI revealed that the AvrBs3protein was detectable in the nucleus of some but not allmesophyll cells (Fig. 2A and A¢). This is probably due tothe fact that not all cells are transformed by Agrobacte-rium. Thus, the untransformed cells act as negative con-trol for the immunostaining. No significant signals wereobserved in leaf sections of tissue infected with Agrobac-terium carrying the empty vector (data not shown). Asexpected, deletion of the NLS region in the C-terminus ofAvrBs3 (avrBs3DNLS) abolished its targeting to thenucleus and led to specific staining in the cytoplasmof infected pepper cells (Fig. 2B and B¢). Similar datawere obtained using fluorescein-isothiocyanate (FITC)-conjugated secondary antibody (data not shown).
Importantly, immunoblotting analysis of pepper leaf tis-sue expressing avrBs3 or avrBs3DNLS after A. tumefa-ciens-mediated gene delivery showed similar proteinlevels indicating that the NLS deletion does not affectprotein stability (Fig. 3A, lanes 1 and 2). No specific signalwas detectable in total protein extracts prepared fromleaves infiltrated with A. tumefaciens carrying the emptyvector (Fig. 3A, lane 3). Altogether, these results demon-strate that avrBs3 expression in the plant results in tar-geting of the protein to the nucleus. Moreover, deletionof the NLS region, which previously was shown toabolish AvrBs3 recognition in resistant plants (Van denAckerveken et al., 1996; Table 2), prevents nuclear local-ization of AvrBs3.
In situ immunolocalization of AvrBs3 in infected leaf tissue
We then examined whether AvrBs3 could be detectedinside the plant cell after delivery by Xcv. To maximizeAvrBs3 transfer by the TTSS, AvrBs3 was expressedunder control of the lac promoter in Xcv strain85*(pDS300F). Strain 85* carries the hrpG* mutation thatleads to the expression of a constitutively active master
Fig. 1. The N-terminal region of AvrBs3 is needed for type III secretion.A. AvrBs3 wild-type and deletion constructs used in this study. Struc-tural features of the 122 kDa AvrBs3 protein are depicted. The central region consists of 17.5 tandem, nearly identical 34-amino-acid repeats. Two functional nuclear localization signals (NLSs) and an acidic activation domain (AAD) are in the C-terminal region (see also Table 1).B. Immunoblotting analysis of total protein extracts (lanes 1–4) and culture supernatants (lanes 5–8) of the following strains: 85*(pDS300F) (wild-type AvrBs3, lanes 1 and 5), 85*DhrcV(pDS300F) (TTSS mutant, lanes 2 and 6), 85*(pDS330F) (AvrBs3DNLS, lanes 3 and 7) and 85*(pDS351F) (AvrBs3D1, lanes 4 and 8). 3 ml aliquots of total protein extracts and 6 ml aliquots of supernatants were loaded. To compensate for AvrBs3D1 protein instability, 7 and 14 ml were loaded in lanes 4 and 8 respectively. Proteins were separated by SDS-PAGE (8% polyacrylamide) and transferred to nitrocellulose. The blot was reacted with an AvrBs3-specific polyclonal antibody, and reprobed with a specific antibody against the intracellular protein HrcN to ensure that no bacterial lysis had occurred.
16 B. Szurek, O. Rossier, G. Hause and U. Bonas
© 2002 Blackwell Publishing Ltd, Molecular Microbiology, 46, 13–23
Fig. 2. AvrBs3 localizes to the nucleus of transformed pepper cells. Leaves of pepper ECW were infected with Agrobacterium tumefa-ciens strains (A) GV3101(pVSF300) and (B) GV3101(pVSF330), car-rying wild-type avrBs3 and avrBs3DNLS on the T-DNA respectively. After T-DNA transfer into the plant cell both genes are highly expressed under control of the constitutive 35S* promoter. Leaves were inoculated with bacterial suspensions of 1 ¥ 109 cfu ml-1 and harvested 60 hpi for sectioning. Transverse leaf sections were incu-bated with anti-AvrBs3 antibody and alkaline phosphatase-coupled secondary antibody. The wild-type AvrBs3 protein is localized in nuclei of infected mesophyll cells (A), whereas after expression of AvrBs3DNLS (B) AvrBs3-specific staining is confined to the cyto-plasm. (A¢) and (B¢) show the identical fields UV-illuminated for the DAPI-stained nuclei. Arrowheads and arrows indicate nuclei that are stained and not stained for AvrBs3 respectively. The bar represents 20 mm.
Fig. 3. Comparison of amounts of AvrBs3 and derivatives in infected pepper leaf extracts.A. Agrobacterium tumefaciens-mediated expression of AvrBs3 in ECW pepper leaves. Leaves were inoculated with a 1 ¥ 109 cfu ml-1 suspension of A. tumefaciens and harvested 60 hpi. 50 mg of total protein extracts were loaded in each lane, subjected to SDS-PAGE and Western blot analysis using anti-AvrBs3 polyclonal antibody. Lane 1, AvrBs3; lane 2, AvrBs3DNLS; lane 3, empty vector.B. Expression of AvrBs3 and derivatives in ECW pepper leaves infected with Xcv. The following strains were used: lane 1, 85*(pDSK602) (empty vector); lane 2, 85*(pDS300F) (wild-type AvrBs3); lane 3, 85*DhrcV(pDS300F); lane 4, 85*(pDS351F) (AvrBs3D1); lane 5, 85*(pDS330F) (AvrBs3DNLS); lane 6, 85*DhrpF(pDS300F); lane 7, 82–8; lane 8, 82*; lane 9, 82*DhrcV. Pepper ECW leaves were inoculated with Xcv suspensions of 1 ¥ 109 cfu ml-1 and harvested 12 hpi. 20 mg of total protein extracts were loaded in each lane, except for lane 4 (75 mg) and lanes 7–9 (100 mg), owing to lower protein expression level or stability. Similar results were obtained in three independent experiments.
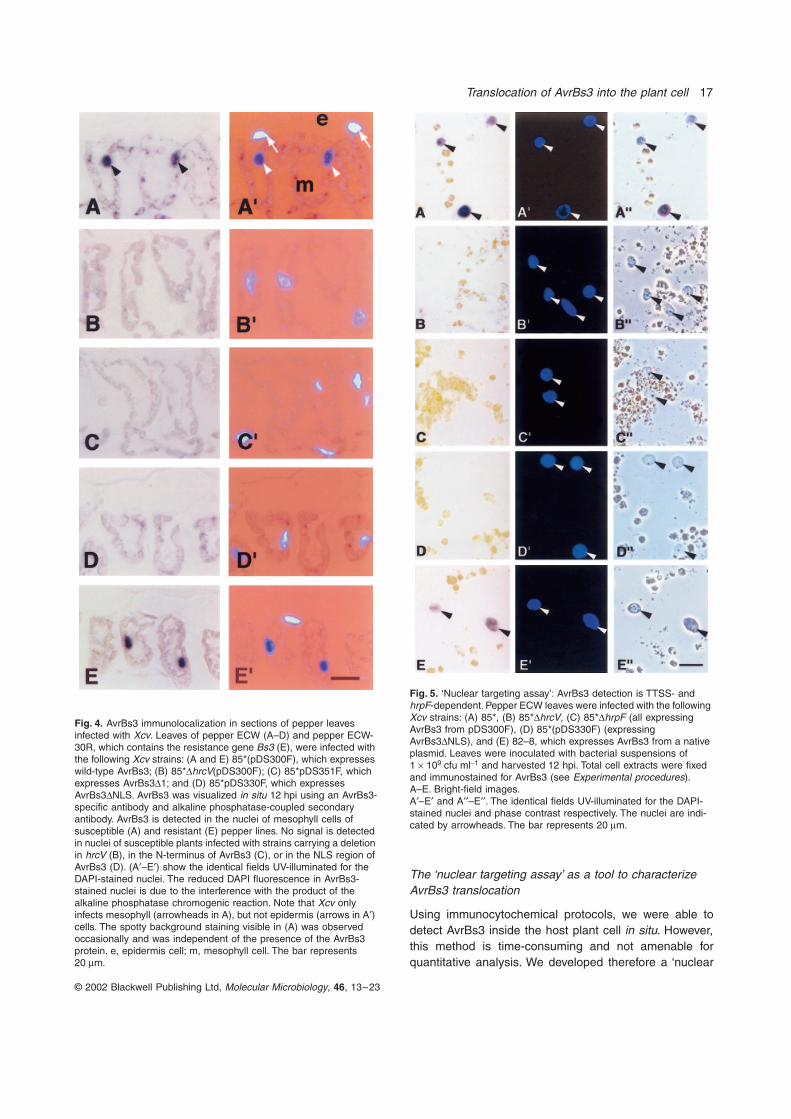
regulator of the TTSS, hrpG (Wengelnik et al., 1999). TypeIII secretion from strain 85*, however, is still dependent onplant inducing conditions (Rossier et al., 1999). AvrBs3localization in pepper ECW was determined immunocy-tochemically using the AvrBs3-specific antibody on 3-mm-thick serial sections of leaf tissue, embedded 12 hpi.Consistent with the data shown in Fig. 2, AvrBs3 stainingwas confined to the nucleus of examined mesophyll cells(arrowheads in Fig. 4A). When pepper plants wereinfected with the TTSS mutant strain 85*DhrcV(pDS300F),no AvrBs3 signal was detectable (Fig. 4B). To rule out thatlack of AvrBs3 detection in the case of the hrcV mutantwas due to fewer bacteria or less protein in infectedleaves, bacterial numbers and AvrBs3 amounts wereexamined. Pepper ECW leaf discs taken at 12 hpi showedsimilar bacterial numbers and AvrBs3 protein amounts forboth 85*(pDS300F) and 85*DhrcV(pDS300F) (Fig. 3B,lanes 2 and 3). Thus, AvrBs3-specific staining of nucleiclearly depends on a functional TTSS. As anticipated fromthe in vitro secretion experiments presented above(Fig. 1B), no AvrBs3-specific signal was observed ininfected tissue when the putative N-terminal secretionsignal in AvrBs3 was deleted (Fig. 4C). Furthermore,deletion of the NLS region in AvrBs3 abolished nuclear
localization of AvrBs3 (Fig. 4D). Although Western blotanalysis of pepper leaf tissue infected with Xcv expressingAvrBs3 or AvrBs3DNLS showed similar protein levels(Fig. 3B, lanes 2 and 5), the AvrBs3DNLS protein wasbarely detectable in the cytoplasm of plant cells. This isprobably due to the large dilution effect in this compart-ment rather than to protein instability and underlines thelower protein amounts in the plant cell when delivered byXcv as compared with Agrobacterium-mediated expres-sion (compare Figs 2B and 4D). In conclusion, lack ofAvrBs3D1 detection in the nuclei is due to the inability tobe secreted and not because of lower protein amount (seebelow), whereas lack of AvrBs3DNLS detection is proba-bly due to the absence of nuclear targeting. Finally, weinvestigated whether AvrBs3 could also be detected in leafcells of the resistant pepper line ECW-30R, which recog-nizes the AvrBs3 protein (Minsavage et al., 1990). Asshown in Fig. 4E, presence of the Bs3 resistance genedid not impair AvrBs3 accumulation in the nucleus at12 hpi, which is before the macroscopic appearance ofthe HR (24 hpi).
Translocation of AvrBs3 into the plant cell 17
© 2002 Blackwell Publishing Ltd, Molecular Microbiology, 46, 13–23
Fig. 5. ‘Nuclear targeting assay’: AvrBs3 detection is TTSS- and hrpF-dependent. Pepper ECW leaves were infected with the following Xcv strains: (A) 85*, (B) 85*DhrcV, (C) 85*DhrpF (all expressing AvrBs3 from pDS300F), (D) 85*(pDS330F) (expressing AvrBs3DNLS), and (E) 82–8, which expresses AvrBs3 from a native plasmid. Leaves were inoculated with bacterial suspensions of 1 ¥ 109 cfu ml-1 and harvested 12 hpi. Total cell extracts were fixed and immunostained for AvrBs3 (see Experimental procedures).A–E. Bright-field images.A¢–E¢ and A¢¢–E¢¢. The identical fields UV-illuminated for the DAPI-stained nuclei and phase contrast respectively. The nuclei are indi-cated by arrowheads. The bar represents 20 mm.
Fig. 4. AvrBs3 immunolocalization in sections of pepper leaves infected with Xcv. Leaves of pepper ECW (A–D) and pepper ECW-30R, which contains the resistance gene Bs3 (E), were infected with the following Xcv strains: (A and E) 85*(pDS300F), which expresses wild-type AvrBs3; (B) 85*DhrcV(pDS300F); (C) 85*pDS351F, which expresses AvrBs3D1; and (D) 85*pDS330F, which expresses AvrBs3DNLS. AvrBs3 was visualized in situ 12 hpi using an AvrBs3-specific antibody and alkaline phosphatase-coupled secondary antibody. AvrBs3 is detected in the nuclei of mesophyll cells of susceptible (A) and resistant (E) pepper lines. No signal is detected in nuclei of susceptible plants infected with strains carrying a deletion in hrcV (B), in the N-terminus of AvrBs3 (C), or in the NLS region of AvrBs3 (D). (A¢–E¢) show the identical fields UV-illuminated for the DAPI-stained nuclei. The reduced DAPI fluorescence in AvrBs3-stained nuclei is due to the interference with the product of the alkaline phosphatase chromogenic reaction. Note that Xcv only infects mesophyll (arrowheads in A), but not epidermis (arrows in A¢) cells. The spotty background staining visible in (A) was observed occasionally and was independent of the presence of the AvrBs3 protein. e, epidermis cell; m, mesophyll cell. The bar represents 20 mm.
The ‘nuclear targeting assay’ as a tool to characterize AvrBs3 translocation
Using immunocytochemical protocols, we were able todetect AvrBs3 inside the host plant cell in situ. However,this method is time-consuming and not amenable forquantitative analysis. We developed therefore a ‘nuclear

18 B. Szurek, O. Rossier, G. Hause and U. Bonas
© 2002 Blackwell Publishing Ltd, Molecular Microbiology, 46, 13–23
Table 2. Type III secretion and recognition of AvrBs3 derivatives.
AvrBs3 construct
Expression in XcvExpression in planta
via Agrobacterium tumefaciens
in vitro secretion Plant phenotype on pepper Bs3a Plant phenotype on pepper Bs3a
AvrBs3 + HR HRAvrBs3D1 – WS HRAvrBs3D2 – WS HRAvrBs3DNLS + WS No HR
a. HR, hypersensitive reaction (defence); WS, watersoaking (disease symptoms).Identical results were obtained in four independent experiments.
Table 1. Xanthomonas strains and plasmids used in this study.
Designation Relevant characteristics Reference/source
Xcv strains85* Derivative of 85–10 carrying hrpG*; constitutive expression of hrp genes Wengelnik et al. (1999)85*DhrcV hrcV deletion mutant of 85* Rossier et al. (2000)85*DhrpF hrpF deletion mutant of 85* Büttner et al. (2002)82–8 Pepper race 1; wild-type strain expressing AvrBs3 from its own promoter Minsavage et al. (1990)82* Derivative of 82–8 carrying hrpG* Wengelnik et al. (1999)82*DhrcV hrcV deletion mutant of 82* Rossier et al. (1999)
PlasmidspDS300F pDSK602 expressing AvrBs3 from the lac promoter Van den Ackerveken et al. (1996)pDS351F pDSK602 expressing AvrBs3 deleted in 25 N-terminal aa This studypDS356F pDSK602 expressing AvrBs3 deleted in 152 N-terminal aa This studypDS330F pDSK602 expressing AvrBs3 deleted in 83 C-terminal aa including the NLS region Van den Ackerveken et al. (1996)
targeting assay’, taking advantage of AvrBs3 localizationto the plant nucleus. First, susceptible pepper leaves(ECW) were infiltrated with Agrobacterium strains carryingdifferent constructs. Nuclei and other organelles prepared60 hpi were fixed on microscopy glass slides (see Exper-imental procedures). Using the AvrBs3-specific antibodyand alkaline phosphatase-coupled secondary antibody,followed by DAPI staining, the AvrBs3 protein wasdetected in plant nuclei after Agrobacterium-mediatedexpression of avrBs3 (data not shown). In control experi-ments using Agrobacterium containing the empty vectoror avrBs3DNLS on the T-DNA, no staining was observedin plant cell nuclei. As described above the N-terminus ofAvrBs3 is essential for type III secretion but not for AvrBs3recognition in planta (see Fig. 1 and Table 2). As expectedfrom the biological activity assays (Table 2), Agrobacte-rium-mediated expression of the N-terminal deletionmutant AvrBs3D1 in the plant led to strong staining innuclei (data not shown), indicating that the N-terminus ofAvrBs3 is dispensable for nuclear targeting. In conclusion,the establishment of the ‘nuclear targeting assay’ usingAgrobacterium was successful and used to furtheranalyse the translocation of AvrBs3 into the plant cellfollowing natural infection with Xanthomonas. Nucleiand other organelles, prepared 12 hpi from susceptiblepepper leaves (ECW) infected with Xanthomonas strain85*(pDS300F), were fixed on microscopy glass slides.
Using the AvrBs3-specific antibody, the AvrBs3 proteinwas detected in plant nuclei (Fig. 5A), which were identi-fied by DAPI staining (Fig. 5A¢) and phase contrastmicroscopy (Fig. 5A¢¢). In control experiments using theTTSS mutant strain DhrcV, AvrBs3 was not detectable inplant cell extracts (Fig. 5B). As anticipated from the datapresented above, no AvrBs3 signal was observed whenthe putative N-terminal secretion signal (data not shown)or the NLS region (Fig. 5D) was deleted in AvrBs3. In anattempt to compensate for the apparently lower amountof AvrBs3D1 compared with the wild-type protein(Fig. 3B), leaves were inoculated with lower density(1.5 ¥ 108 cfu ml-1) of the wild-type strain 85*(pDS300F).Although in this case the relative amounts of AvrBs3and AvrBs3D1 per leaf disk were similar (data notshown), AvrBs3-stained nuclei were only observed with85*(pDS300F). In addition to the TTSS mutant DhrcV, wetested Xcv strains carrying a deletion in hrpF. hrpFmutants still secrete proteins via the TTSS in vitro but nolonger cause disease or induce the HR in the plant, sug-gesting that hrpF encodes the ‘translocator’ (Rossieret al., 2000; Büttner et al., 2002). As shown in Fig. 5C, nostained nuclei were obtained from pepper leaves infectedwith the mutant strain 85*DhrpF(pDS300F). Takentogether with the results presented above (Fig. 4), detec-tion of AvrBs3 in plant nuclei depends on both secretionfrom the bacteria and translocation into the plant cytosol.

Translocation of AvrBs3 into the plant cell 19
© 2002 Blackwell Publishing Ltd, Molecular Microbiology, 46, 13–23
To study the time-course of AvrBs3 translocation,nuclei from pepper ECW leaves infected with strain85*(pDS300F) were examined in 2 h-intervals from 0 to12 hpi. The AvrBs3 protein was detected in plant nucleias early as 4 hpi (not shown). The number of stainednuclei increased over time: 8% of the nuclei (80 out of 998analysed) at 4 hpi and 63% of the nuclei (526 out of 835)at 12 hpi stained positive for AvrBs3.
Visualization of AvrBs3 delivered by a naturally occurring Xcv strain
The experiments described above were performed usingXcv strains harbouring a plasmid in which avrBs3 is underthe control of the lac promoter. To address whetherAvrBs3 is also detectable in plant nuclei after pepperinfection with a strain expressing wild-type levels ofAvrBs3, we used Xcv strain 82*, which expresses avrBs3under the control of its own promoter (Table 1). The‘nuclear targeting assay’ developed above was performedwith leaf tissue of pepper ECW infected with Xcv strain82*. Nuclei exhibited significant AvrBs3 staining 12 hpi butnot at 10 hpi (data not shown). In contrast, no AvrBs3signal was detected in nuclei isolated from leaves infectedwith the isogenic TTSS mutant 82*DhrcV (data notshown). All the strains analysed so far in this studyexpress components of the TTSS constitutively. When weused the naturally occurring strain 82–8, AvrBs3 wasclearly detected in the nuclei at 12 hpi (Fig. 5E and E¢),although the protein expression level was lower than withXcv 85*(pDS300F) (Fig. 3B, lanes 2 and 7). In conclusion,translocation of AvrBs3 expressed in the wild-type straincould be clearly demonstrated.
Discussion
Using immunocytochemical detection protocols thetype III effector AvrBs3 was demonstrated to be deliveredby the plant pathogen Xcv into host plant cells in situ.Significantly, TTSS-dependent AvrBs3 translocation wasobserved in the native tissues of plants infected with Xan-thomonas, whereas translocation studies in systemsinvolving animal pathogenic bacteria used cultured HeLacells or macrophages, but not the intact host organism(Rosqvist et al., 1994; Collazo et al., 1995; Sory et al.,1995; Wood et al., 1996). Moreover, we provide experi-mental evidence for AvrBs3 targeting to the plant nucleus.Nuclear localization of AvrBs3 confirmed our previousdata showing that a fusion between the AvrBs3 C-terminalregion and b-glucuronidase resulted in nuclear targetingof the reporter protein in onion epidermis cells (Van denAckerveken et al., 1996).
We show here that translocation was not only depen-dent on a functional TTSS, but also on the HrpF protein,
which is itself secreted but not required for type III-dependent secretion in vitro. As hrpF mutants are unableto grow in planta and do not provoke any plant reaction,it has been suggested that the HrpF protein plays a roleafter secretion, i.e. in translocation (Rossier et al., 2000).The data presented here corroborate this model. WhetherHrpF interacts with the host plasma membrane is notknown. However, it has recently been shown that HrpFhas lipid-binding activity and forms a pore in lipid bilayers,supporting the function of HrpF as a translocon protein(Büttner et al., 2002).
Despite extensive efforts to demonstrate the trans-location of effector proteins into the plant cell by the TTSSof a plant pathogenic bacterium, direct evidence waslacking until now. During the course of the preparationof this manuscript, type III-dependent translocation ofthe Xcv protein AvrBs2 was shown using cya fusions(Casper-Lindley et al., 2002). In our work, AvrBs3 accu-mulation in the nucleus was key to facilitate detection byimmunocytochemistry. In the ‘nuclear targeting assay’,AvrBs3-staining was only observed in nuclei after freezingthe samples, probably because this facilitates the pene-tration of antibodies into the nuclei. Moreover, the useof alkaline phosphatase-coupled secondary antibodyappeared to be important, because AvrBs3 was undetect-able in nuclei of Xcv-infected leaves using FITC conju-gates here and in previous studies (Van den Ackervekenet al., 1996). Localization of AvrBs3 by immunofluores-cence was only successful when the protein was overex-pressed in the plant cell using Agrobacterium-mediatedgene transfer (data not shown). Although bacteria couldbe identified by DAPI staining or phase contrast using the‘nuclear targeting assay’ (data not shown), they were notstained for AvrBs3. Possible explanations are the struc-tural differences between the bacterial and nuclear enve-lope and differential permeability for antibodies aftertreatment with Triton X-100.
In this study, we have established the ‘nuclear targetingassay’ that will enable us to analyse the translocationsignal of AvrBs3. Type III translocation signals have beenmost extensively studied in Yersinia Yops (Yersinia outerproteins) (Cornelis, 2000). For several Yops, a minimalregion of 15–17 N-terminal amino acids was found to beessential for secretion in vitro, whereas additional 50–70amino acids were required for translocation into theeukaryotic cell (Plano et al., 2001). Interestingly, a puta-tive translocation motif consisting of a proline residuesurrounded by basic amino acids is conserved in the N-terminus of all avirulence proteins characterized so far inXcv (Escolar et al., 2001). An additional potential applica-tion of the ‘nuclear targeting assay’ is the use of thismethod to demonstrate the translocation of other type IIIeffectors fused to the AvrBs3 NLSs into the plant cell.
Visualization of AvrBs3 in the plant nucleus was depen-

20 B. Szurek, O. Rossier, G. Hause and U. Bonas
© 2002 Blackwell Publishing Ltd, Molecular Microbiology, 46, 13–23
dent on the NLSs in the C-terminus, which is consistentwith our earlier observation that the NLSs are requiredfor AvrBs3 activity (Van den Ackerveken et al., 1996).Nuclear localization of AvrBs3 is probably due to its inter-action with pepper importin a, a component of the nuclearimport machinery, which was recently isolated from pep-per by a yeast two-hybrid screen and which also interactswith AvrBs3 in vitro (Szurek et al., 2001). One of the keyquestions for future research is the biochemical functionof AvrBs3 and the nature of the cellular target(s) of AvrBs3in the plant. Intriguingly, AvrBs3 and its homologues con-tain an acidic transcription activation domain in the C-terminus, reinforcing the presumed function of membersof this effector protein family in the nucleus (Zhu et al.,1998; Yang et al., 2000; Szurek et al., 2001). We thereforebelieve that AvrBs3 modulates transcription and thusinduces changes in the plant metabolism, which contrib-ute to disease development. The observation that AvrBs3induces hypertrophy of mesophyll cells in susceptibleplants (Marois et al., 2002) fits with this idea. Althoughthere is now direct evidence for effector translocation froma plant pathogen into the host cell, it is still a mystery howthe TTSS transfers proteins across the plant cell wall andplasma membrane. The finding of TTSS-specific pili thatmight serve as conduit (Brown et al., 2001; Jin and He,2001; Jin et al., 2001) is a step towards a better under-standing of this crucial process.
Experimental procedures
Bacterial strains, growth conditions and plasmids
Bacterial strains used in this study were Escherichia colistrain DH5a (Bethesda Research Laboratories), Agrobacte-rium tumefaciens strain GV3101 (Van Larebeke et al., 1974),and different Xanthomonas campestris pv. vesicatoria (Xcv)strains (see Table 1). E. coli cells were cultivated at 37∞C inLuria–Bertani medium, Xcv at 30∞C in NYG broth (Danielset al., 1984), and A. tumefaciens at 30∞C in YEB medium(Van den Ackerveken et al., 1996). Plasmids were introducedinto E. coli by electroporation, and into Xcv and A. tumefa-ciens by conjugation, using pRK2013 as a helper plasmid intriparental mating (Figurski and Helinski, 1979).
Construction of plasmids
The N-terminal deletions in AvrBs3 were constructed as fol-lows: for avrBs3D1, a linker (obtained with oligonucleotides5¢-GATCGTCAGGCCTATGCA-3¢ and 5¢-TAGGCCTGAC-3¢)was ligated into pUS300F digested with BamHI–PstI (Vanden Ackerveken et al., 1996), yielding pUS351F. pUS351Fencodes AvrBs3D1 in which amino acid residues 3–27 arereplaced by a short peptide (RQAS). AvrBs3D2 lacks aminoacids 2–153: a 2886 basepair (bp) fragment, obtained byBstYI digestion of pUS300F, was ligated to the 3308 bpBamHI fragment from pUS300F, resulting in pUS356F. The
avrBs3 derivatives were cloned into the expression vectorpDSK602 (Murillo et al., 1994), resulting in pDS351F andpDS356F. For Agrobacterium-mediated gene transfer,avrBs3 derivatives were cloned into pBI1.4t (Mindrinos et al.,1994).
Plant material and plant inoculations
Pepper (Capsicum annuum) plants of cultivar Early CalWonder (ECW) and the near-isogenic line ECW-30R, whichcontains the resistance gene Bs3 (Minsavage et al., 1990),were grown in the greenhouse. Xcv and A. tumefaciens sus-pensions of 5 ¥ 108 colony forming units (cfu) ml-1 in 10 mMMgCl2 were infiltrated into the intercellular space of leavesusing a needle-less syringe as described (Bonas et al., 1989;Van den Ackerveken et al., 1996). To rule out that lack ofhypersensitive reaction (HR) was due to lower proteinamounts in the case of bacteria expressing AvrBs3D1 orAvrBs3D2 (Table 2), leaves of pepper ECW-30R were infil-trated with a higher inoculum (1 ¥ 109 cfu ml-1). No HR wasobserved while bacteria expressing wild-type AvrBs3 wereable to induce the HR when inoculated at 1 ¥ 108 cfu ml-1.Macroscopic reactions were scored 2–5 days after inocula-tion. For immunocytochemistry, the inoculum concentrationwas approximately 109 cfu ml-1. Bacterial growth in plantawas determined as described (Bonas et al., 1991).
Protein secretion experiments and immunoblot analysis
In vitro secretion experiments were performed as described(Rossier et al., 1999). Total protein extracts and culturesupernatants were concentrated 10- and 100-fold respec-tively. Western blots were reacted with affinity-purifiedAvrBs3-specific polyclonal antibody (diluted 1:15 000)(Knoop et al., 1991) and with anti-rabbit horseradish peroxi-dase-conjugated secondary antibody, which was detectedby enhanced chemiluminescence (Amersham PharmaciaBiotech). The membranes were reprobed with a specific anti-body against the intracellular protein HrcN to ensure that nobacterial lysis had occurred (Rossier et al., 1999). Proteinextracts from infected leaves were prepared by grinding twoleaf discs (0.8 cm in diameter) from different leaves in liquidnitrogen in 80 ml of protein extraction buffer (20 mM Tris-HCl,pH 7.5, 150 mM NaCl, 1 mM EDTA, 1% Triton X-100, 0.1%SDS) to which 20 ml of 5¥ Laemmli buffer was added. Proteinsamples were separated by SDS-PAGE and subjected toimmunoblot analysis.
Immunostaining of leaf sections
Leaf segments were fixed for 3 h with 3% paraformaldehydein phosphate-buffered saline (PBS), washed, dehydrated andembedded in polyethylene glycol (Van Lammeren et al.,1985). Then, 3 mm serial sections were mounted on glassslides, incubated with 0.1 M NH4Cl, washed with PBS, andincubated in PBS-3% BSA and anti-AvrBs3 antibody(diluted1:3000). Antibody binding was visualized by sheepanti-rabbit IgG conjugated to alkaline phosphatase (Roche)according to the manufacturer's instructions. To identify

Translocation of AvrBs3 into the plant cell 21
© 2002 Blackwell Publishing Ltd, Molecular Microbiology, 46, 13–23
nuclei the samples were incubated with 0.1 mg ml-1 of 4¢,6-diamidino-2-phenylindole (DAPI) and mounted in Citifluor-glycerol (Citifluor). Fluorescence images were obtained usinga Zeiss (Jena) Axioplan 2 microscope equipped with a FujiDig HC-300Z digital camera (Fujifilm) using filter blocks forDAPI. Experiments were repeated three times with similarresults. Approximately 100 sections were examined perassay for each tested strain.
‘Nuclear targeting assay’
Leaf discs (1 cm in diameter) were chopped on glass slidesin 4% formaldehyde in PBS, pH 7, 0.05% Triton X-100. Sam-ples were frozen in liquid nitrogen and treated as describedabove for leaf sections. In addition to fluorescence images,we used phase-contrast, enabling the visualization of bothnuclei and chloroplasts. Experiments were repeated at leastthree times with similar results.
Acknowledgements
We are grateful to R. Kahmann, E. Marois and R. Koebnikfor critically reading the manuscript, and to A. Fischer fortechnical advice. This work was supported by a grant fromthe Deutsche Forschungsgemeinschaft (SFB 363) to U.B.
References
Bai, J., Choi, S.H., Ponciano, G., Leung, H., and Leach, J.E.(2000) Xanthomonas oryzae pv. oryzae avirulence genescontribute differently and specifically to pathogen aggres-siveness. Mol Plant-Microbe Interact 13: 1322–1329.
Bogdanove, A., Beer, S.V., Bonas, U., Boucher, C.A.,Collmer, A., Coplin, D.L., et al. (1996) Unified nomencla-ture for broadly conserved hrp genes of phytopathogenicbacteria. Mol Microbiol 20: 681–683.
Bonas, U., and Van den Ackerveken, G. (1999) Gene-for-gene interactions: bacterial avirulence proteins specifyplant disease resistance. Curr Opin Microbiol 2: 94–98.
Bonas, U., Stall, R.E., and Staskawicz, B. (1989) Genetic andstructural characterization of the avirulence gene avrBs3from Xanthomonas campestris pv. vesicatoria. Mol GenGenet 218: 127–136.
Bonas, U., Schulte, R., Fenselau, S., Minsavage, G.V.,Staskawicz, B.J., and Stall, R.E. (1991) Isolation of a gene-cluster from Xanthomonas campestris pv. vesicatoria thatdetermines pathogenicity and the hypersensitive responseon pepper and tomato. Mol Plant-Microbe Interact 4: 81–88.
Brown, I.R., Mansfield, J.W., Taira, S., Roine, E., andRomantschuk, M. (2001) Immunocytochemical localizationof HrpA and HrpZ supports a role for the Hrp pilus in thetransfer of effector proteins from Pseudomonas syringaepv. tomato across the host plant cell wall. Mol Plant-Microbe Interact 14: 394–404.
Büttner, D., Nennstiel, D., Klüsener, B., and Bonas, U. (2002)Functional analysis of HrpF, a putative type III transloconprotein from Xanthomonas campestris pv. vesicatoria. JBacteriol 184: 2389–2398.
Casper-Lindley, C., Dahlbeck, D., Clark, E.T., andStaskawicz, B.J. (2002) Direct biochemical evidence fortype III secretion-dependent translocation of the AvrBs2effector protein into plant cells. Proc Natl Acad Sci USA99: 8336–8341.
Collazo, C.M., Zieler, M.K., and Galán, J.E. (1995) Functionalanalysis of the Salmonella typhimurium invasion genes invland invJ and identification of a target of the protein secre-tion apparatus encoded in the inv locus. Mol Microbiol 15:25–38.
Cornelis, G.R. (2000) Molecular and cell biology aspects ofplague. Proc Natl Acad Sci USA 97: 8778–8783.
Cornelis, G.R., and Van Gijsegem, F. (2000) Assembly andfunction of type III secretory systems. Annu Rev Microbiol54: 735–774.
Daniels, M.J., Barber, C.E., Turner, P.C., Sawczyc, M.K.,Byrde, R.J.W., and Fielding, A.H. (1984) Cloning of genesinvolved in pathogenicity of Xanthomonas campestris pv.campestris using the broad host range cosmid pLAFR1.EMBO J 3: 3323–3328.
Escolar, L., Van den Ackerveken, G., Rossier, O., Pieplow,S., and Bonas, U. (2001) Type III secretion and in plantarecognition of the Xanthomonas avirulence proteinsAvrBs1 and AvrBsT. Mol Plant Pathol 2: 287–296.
Fenselau, S., and Bonas, U. (1995) Sequence and expres-sion analysis of the hrpB pathogenicity operon of Xan-thomonas campestris pv. vesicatoria which encodes eightproteins with similarity to components of the Hrp, Ysc, Spa,and Fli secretion systems. Mol Plant-Microbe Interact 8:845–854.
Fenselau, S., Balbo, I., and Bonas, U. (1992) Determinantsof pathogenicity in Xanthomonas campestris pv. vesicato-ria are related to proteins involved in secretion in bacterialpathogens of animals. Mol Plant-Microbe Interact 5: 390–396.
Figurski, D.H., and Helinski, D.R. (1979) Replication of anorigin-containing derivative of plasmid RK2 dependent ona plasmid function provided in trans. Proc Natl Acad SciUSA 76: 1648–1652.
Herbers, K., Conrads-Strauch, J., and Bonas, U. (1992)Race-specificity of plant resistance to bacterial spot dis-ease determined by repetitive motifs in a bacterial aviru-lence protein. Nature 356: 172–174.
Hueck, C.J. (1998) Type III protein secretion systems inbacterial pathogens of animals and plants. Microbiol MolBiol Rev 62: 379–433.
Huguet, E., and Bonas, U. (1997) hrpF of Xanthomonascampestris pv. vesicatoria encodes an 87-kDa protein withhomology to NolX of Rhizobium fredii. Mol Plant-MicrobeInteract 10: 488–498.
Innes, R.W. (2001) Targeting the targets of type III effectorproteins secreted by phytopathogenic bacteria. Mol PlantPathol 2: 109–115.
Jin, Q., and He, S.Y. (2001) Role of the Hrp pilus in type IIIprotein secretion in Pseudomonas syringae. Science 294:2556–2558.
Jin, Q., Hu, W., Brown, I., McGhee, G., Hart, P., Jones, A.L.,and He, S.Y. (2001) Visualization of secreted Hrp and Avrproteins along the Hrp pilus during type III secretion inErwinia amylovora and Pseudomonas syringae. Mol Micro-biol 40: 1129–1139.

22 B. Szurek, O. Rossier, G. Hause and U. Bonas
© 2002 Blackwell Publishing Ltd, Molecular Microbiology, 46, 13–23
Kjemtrup, S., Nimchuk, Z., and Dangl, J.L. (2000) Effectorproteins of phytopathogenic bacteria: bifunctional signalsin virulence and host recognition. Curr Opin Microbiol 3:73–78.
Klement, Z. (1982) Hypersensitivity. In PhytopathogenicProkaryotes, Vol. 2. Mount, M.S., and Lacy, G.H. (eds).New York: Academic Press, pp. 149–177.
Knoop, V., Staskawicz, B., and Bonas, U. (1991) Expressionof the avirulence gene avrBs3 from Xanthomonas campes-tris pv. vesicatoria is not under the control of hrp genesand is independent of plant factors. J Bacteriol 173: 7142–7150.
Lee, V.T., Anderson, D.M., and Schneewind, O. (1998)Targeting of Yersinia Yop proteins into the cytosol of HeLacells: one-step translocation of YopE across bacterial andeukaryotic membranes is dependent on SycE chaperone.Mol Microbiol 28: 593–601.
Leister, R.T., and Katagiri, F. (2000) A resistance geneproduct of the nucleotide binding site: leucine rich repeatsclass can form a complex with bacterial avirulence proteinsin vivo. Plant J 22: 345–354.
Lindgren, P.B. (1997) The role of hrp genes during plant–bacterial interactions. Annu Rev Phytopathol 35: 129–152.
Lloyd, S.A., Norman, M., Rosqvist, R., and Wolf-Watz, H.(2001) Yersinia YopE is targeted for type III secretion by N-terminal, not mRNA, signals. Mol Microbiol 39: 520–532.
Mackey, D., Holt, B.F., Wiig, A., and Dangl, J. (2002) RIN4interacts with Pseudomonas syringae type III effectormolecules and is required for RPM1-mediated resistancein Arabidopsis. Cell 108: 743–754.
Marois, E., van den Ackerveken, G., and Bonas, U. (2002)The Xanthomonas type III effector protein AvrBs3 modu-lates plant gene expression and induces cell hypertrophyin the susceptible host. Mol Plant-Microbe Interact 15:637–646.
Mindrinos, M., Katagiri, F., Yu, G.L., and Ausubel, F.M.(1994) The A. thaliana disease resistance gene RPS2encodes a protein containing a nucleotide-binding site andleucine-rich repeats. Cell 78: 1089–1099.
Minsavage, G.V., Dahlbeck, D., Whalen, M.C., Kearny, B.,Bonas, U., Staskawicz, B.J., and Stall, R.E. (1990) Gene-for-gene relationships specifying disease resistance inXanthomonas campestris pv. vesicatoria – pepper interac-tions. Mol Plant-Microbe Interact 3: 41–47.
Mudgett, M.B., and Staskawicz, B.J. (1999) Characterizationof the Pseudomonas syringae pv. tomato AvrRpt2 protein:demonstration of secretion and processing during bacterialpathogenesis. Mol Microbiol 32: 927–941.
Mudgett, M.B., Chesnokova, O., Dahlbeck, D., Clark, E.T.,Rossier, O., Bonas, U., and Staskawicz, B.J. (2000)Molecular signals required for type III secretion and trans-location of the Xanthomonas campestris AvrBs2 protein topepper plants. Proc Natl Acad Sci USA 97: 13324–13329.
Murillo, J., Shen, H., Gerhold, D., Sharma, A., Cooksey, D.A.,and Keen, N.T. (1994) Characterization of pPT23B, theplasmid involved in syringolide production by Pseudomo-nas syringae pv. tomato PT23. Plasmid 31: 275–287.
Nimchuk, Z., Marois, E., Kjemtrup, S., Leister, R.T., Katagiri,F., and Dangl, J.L. (2000) Eukaryotic fatty acylation drivesplasma membrane targeting and enhances function of sev-
eral type III effector proteins from Pseudomonas syringae.Cell 101: 353–363.
Plano, G.V., Day, J.B., and Ferracci, F. (2001) Type III export:new uses for an old pathway. Mol Microbiol 40: 284–293.
Romantschuk, M., Roine, E., and Taira, S. (2001) Hrp pilus:reaching through the plant cell wall. Eur J Plant Pathol 107:153–160.
Rosqvist, R., Magnusson, K.E., and Wolf-Watz, H. (1994)Target cell contact triggers expression and polarized trans-fer of Yersinia YopE cytotoxin into mammalian cells. EMBOJ 1390: 964–972.
Rossier, O., Wengelnik, K., Hahn, K., and Bonas, U. (1999)The Xanthomonas Hrp type III system secretes proteinsfrom plant and mammalian pathogens. Proc Natl Acad SciUSA 96: 9368–9373.
Rossier, O., Van den Ackerveken, G., and Bonas, U. (2000)HrpB2 and HrpF from Xanthomonas are type III-secretedproteins and essential for pathogenicity and recognition bythe host plant. Mol Microbiol 38: 828–838.
Schesser, K., Frithz-Lindsten, E., and Wolf-Watz, H. (1996)Delineation and mutational analysis of the Yersiniapseudotuberculosis YopE domains which mediate translo-cation across bacterial and eukaryotic cellular membranes.J Bacteriol 178: 7227–7233.
Scofield, S.R., Tobias, C.M., Rathjen, J.P., Chang, J.H.,Lavelle, D.T., Michelmore, R.W., and Staskawicz, B.J.(1996) Molecular basis of gene-for-gene specificity in bac-terial speck disease of tomato. Science 274: 2063–2065.
Shan, L., Thara, V.K., Martin, G.B., Zhou, J.-M., and Tang,X. (2000) The Pseudomonas AvrPto protein is differentiallyrecognized by tomato and tobacco and is localized to theplant plasma membrane. Plant Cell 12: 2323–2337.
Sory, M.P., and Cornelis, G.R. (1994) Translocation of ahybrid YopE-adenylate cyclase from Yersinia enterocoliticainto HeLa cells. Mol Microbiol 14: 583–594.
Sory, M.P., Boland, A., Lambermont, I., and Cornelis, G.R.(1995) Identification of the YopE and YopH domainsrequired for secretion and internalization into the cytosol ofmacrophages, using the cyaA gene fusion approach. ProcNatl Acad Sci USA 92: 11998–12002.
Staskawicz, B.J., Mudgett, M.B., Dangl, J.L., and Galan, J.E.(2001) Common and contrasting themes of plant and ani-mal diseases. Science 292: 2285–2280.
Swarup, S., De Feyter, R., Brlansky, R.H., and Gabriel, D.W.(1991) A pathogenicity locus from Xanthomonas citrienables strains from several pathovars of Xanthomonascampestris to elicit canker-like lesions on citrus. Phytopa-thology 81: 802–809.
Swarup, S., Yang, Y., Kingsley, M.T., and Gabriel, D.W.(1992) A Xanthomonas citri pathogenicity gene, pthA,pleiotropically encodes gratuitous avirulence on nonhosts.Mol Plant-Microbe Interact 5: 204–213.
Szurek, B., Marois, E., Bonas, U., and Van den Ackerveken,G. (2001) Eukaryotic features of the Xanthomonas type IIIeffector AvrBs3: protein domains involved in transcriptionalactivation and the interaction with nuclear import receptorsfrom pepper. Plant J 26: 523–534.
Tang, X., Frederick, R.D., Zhou, J., Halterman, D.A., Jia, Y.,and Martin, G.B. (1996) Initiation of plant disease resis-tance by physical interaction of AvrPto and Pto kinase.Science 274: 2060–2063.

Translocation of AvrBs3 into the plant cell 23
© 2002 Blackwell Publishing Ltd, Molecular Microbiology, 46, 13–23
Van den Ackerveken, G., Marois, E., and Bonas, U. (1996)Recognition of the bacterial avirulence protein AvrBs3occurs inside the host plant cell. Cell 87: 1307–1316.
Van Lammeren, A.A.M., Keijzer, C.J., Willemse, M.T.M., andKieft, H. (1985) Structure and function of the microtubularcytoskeleton during pollen development in Gasteria verru-cosa (Mill.) H. Duval. Planta 165: 1–11.
Van Larebeke, N., Engler, G., Holsters, M., Van den Elsacker,S., Zaenen, I., Schilperoort, R.A., and Schell, J. (1974)Large plasmid in Agrobacterium tumefaciens essential forcrown gall-inducing ability. Nature 252: 169–170.
Vivian, A., and Arnold, D.L. (2000) Bacterial effector genesand their role in host–pathogen interactions. J Plant Pathol82: 163–178.
Wengelnik, K., Rossier, O., and Bonas, U. (1999) Mutationsin the regulatory gene hrpG of Xanthomonas campestrispv. vesicatoria result in constitutive expression of all hrpgenes. J Bacteriol 181: 6828–6831.
Wood, M.W., Rosqvist, R., Mullan, P.B., Edwards, M.H., andGalyov, E.E. (1996) SopE, a secreted protein of Salmo-nella dublin, is translocated into the target eukaryotic cellvia a sip-dependent mechanism and promotes bacterialentry. Mol Microbiol 22: 327–338.
Yang, Y., and Gabriel, D.W. (1995) Xanthomonas avirulence/pathogenicity gene family encodes functional plantnuclear targeting signals. Mol Plant-Microbe Interact 8:627–631.
Yang, B., Zhu, W., Johnson, L.B., and White, F.F. (2000) Thevirulence factor AvrXa7 of Xanthomonas oryzae pv. oryzaeis a type III secretion pathway-dependent nuclear-localizeddouble-stranded DNA-binding protein. Proc Natl Acad SciUSA 97: 9807–9812.
Zhu, W.G., Yang, B., Chittoor, J.M., Johnson, L.B., andWhite, F.F. (1998) AvrXa10 contains an acidic transcrip-tional activation domain in the functionally conserved Cterminus. Mol Plant-Microbe Interact 11: 824–832.




















