
Immune response of zebrafish ( Danio rerio) against a newly isolated bacterial pathogen Aeromonas...
11
Immune response of zebrafish (Danio rerio) against a newly isolated bacterial pathogen Aeromonas hydrophila I. Rodrı ´guez, B. Novoa, A. Figueras* Instituto de Investigaciones Marinas, Consejo Superior de Investigaciones Cientı´ficas (CSIC), Eduardo Cabello 6, Vigo, Spain Received 11 February 2008; revised 7 May 2008; accepted 11 May 2008 Available online 18 May 2008 KEYWORDS Zebrafish; Aeromonas; Cytokine; Inflammation; Immune response; Flow cytometry; Extracellular products Abstract A strain of Aeromonas hydrophila associated with unusual mortalities in zebrafish (Danio rerio) culture facilities was isolated, identified and characterized. In challenge exper- iments, adult zebrafish were susceptible to infection by intraperitoneal (i.p.) injection with viable bacteria and its extracellular products (ECPs) reaching very high mortalities in a few hours. The infection, by the viable bacteria or the ECPs, caused cell death in kidney, due to the cytotoxic and haemolytic activities of the bacterial ECPs. Moreover, the infection affected the release of oxygen (ROS) and nitrogen (NO) reactive free radicals. To determine if this A. hydrophila infection induces an inflammatory response, mRNA expression levels of tumour necrosis factor-a (TNFa), interleukin-1b (IL-1b), interferon-g (IFNg) and inducible nitric oxide (iNOS) were assessed by real time PCR. The expression levels of TNFa, IL-1b and IFNg were upregulated in the kidneys of infected zebrafish with viable bacteria, heat-killed bacteria and ECPs. Expression levels of iNOS were upregulated by ECPs. Mortality rate (LD 50 ) and histo- pathology were also determined. ª 2008 Elsevier Ltd. All rights reserved. Introduction Zebrafish (Danio rerio) has been extensively used to study vertebrate development [1] and recently the interest has grown in the fields of human disease, cancer and immunol- ogy [2e10]. The zebrafish with a complete (innate and adaptative) immune system has advantages over other ver- tebrate infection models, such as mice and rats, because of its small size, relatively rapid life cycle and ease of breed- ing, transparency of early life stages and rapid growth allowing a high number of genetic screens and real-time visualization. Experimental infections using zebrafish have been conducted with different bacteria [11e17] and viruses [18e21]. Aeromonas spp. are ubiquitous aquatic ecosystems bacteria [22]. In particular, Aeromonas hydrophila has been reported as an important pathogen for humans and for lower vertebrates, including amphibians, reptiles and fish [23]. These bacteria have often been isolated from humans with diarrhoea [24e26] and in systemic illness of * Corresponding author. E-mail address: [email protected] (A. Figueras). 1050-4648/$ - see front matter ª 2008 Elsevier Ltd. All rights reserved. doi:10.1016/j.fsi.2008.05.002 available at www.sciencedirect.com journal homepage: www.elsevier.com/locate/fsi Fish & Shellfish Immunology (2008) 25, 239e249
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Immune response of zebrafish ( Danio rerio) against a newly isolated bacterial pathogen Aeromonas...

Fish & Shellfish Immunology (2008) 25, 239e249
ava i lab le at www.sc ienced i rec t . com
journa l homepage : www. e l sev i er .com/ loca te / fs i
Immune response of zebrafish (Danio rerio)against a newly isolated bacterialpathogen Aeromonas hydrophila
I. Rodrıguez, B. Novoa, A. Figueras*
Instituto de Investigaciones Marinas, Consejo Superior de Investigaciones Cientıficas (CSIC),Eduardo Cabello 6, Vigo, Spain
Received 11 February 2008; revised 7 May 2008; accepted 11 May 2008Available online 18 May 2008
KEYWORDSZebrafish;Aeromonas;Cytokine;Inflammation;Immune response;Flow cytometry;Extracellular products
* Corresponding author.E-mail address: [email protected]
1050-4648/$ - see front matter ª 200doi:10.1016/j.fsi.2008.05.002
Abstract A strain of Aeromonas hydrophila associated with unusual mortalities in zebrafish(Danio rerio) culture facilities was isolated, identified and characterized. In challenge exper-iments, adult zebrafish were susceptible to infection by intraperitoneal (i.p.) injection withviable bacteria and its extracellular products (ECPs) reaching very high mortalities in a fewhours. The infection, by the viable bacteria or the ECPs, caused cell death in kidney, due tothe cytotoxic and haemolytic activities of the bacterial ECPs. Moreover, the infection affectedthe release of oxygen (ROS) and nitrogen (NO) reactive free radicals. To determine if this A.hydrophila infection induces an inflammatory response, mRNA expression levels of tumournecrosis factor-a (TNFa), interleukin-1b (IL-1b), interferon-g (IFNg) and inducible nitric oxide(iNOS) were assessed by real time PCR. The expression levels of TNFa, IL-1b and IFNg wereupregulated in the kidneys of infected zebrafish with viable bacteria, heat-killed bacteriaand ECPs. Expression levels of iNOS were upregulated by ECPs. Mortality rate (LD50) and histo-pathology were also determined.ª 2008 Elsevier Ltd. All rights reserved.
Introduction
Zebrafish (Danio rerio) has been extensively used to studyvertebrate development [1] and recently the interest hasgrown in the fields of human disease, cancer and immunol-ogy [2e10]. The zebrafish with a complete (innate andadaptative) immune system has advantages over other ver-tebrate infection models, such as mice and rats, because of
(A. Figueras).
8 Elsevier Ltd. All rights reserved
its small size, relatively rapid life cycle and ease of breed-ing, transparency of early life stages and rapid growthallowing a high number of genetic screens and real-timevisualization. Experimental infections using zebrafish havebeen conducted with different bacteria [11e17] and viruses[18e21].
Aeromonas spp. are ubiquitous aquatic ecosystemsbacteria [22]. In particular, Aeromonas hydrophila hasbeen reported as an important pathogen for humans andfor lower vertebrates, including amphibians, reptiles andfish [23]. These bacteria have often been isolated fromhumans with diarrhoea [24e26] and in systemic illness of
.

240 I. Rodrıguez et al.
immunocompromised patients [27e29] being associatedwith high mortality rates, ranging from 24 to 68% in differ-ent studies [30e32]. This microorganism is also responsiblefor haemorrhagic septicaemia, a disease affecting a widevariety of freshwater fish species and occasionallymarine fish [33e40]. Several extracellular toxins andenzymes have been described as responsible for thevirulence of A. hydrophila. These include haemolysins,cytotoxins, enterotoxins and proteases [41,42].
In the present work we have identified and character-ized the aetiological agent associated with high mortalitiesdetected in a stock of zebrafish. Furthermore, severalimmune related functions such as the production of oxygen(ROS) and nitrogen (nitric oxide, NO) radicals or theexpression of proinflammatory cytokines were determined.
Materials and methods
Zebrafish
Care and feeding of zebrafish were conducted followingestablished protocols [1] (also see http://zfin.org/zf_info/zfbook/zfbk.html). Zebrafish wild type adults (>4 months)were anaesthetized with MS-222 (tricaine methanesulpho-nate, Argent Chemical Laboratories, USA). Euthanasia ofzebrafish was achieved by anaesthetic overdose.
Bacteria
Isolation and identification of bacteriaSeveral days after the arrival of a zebrafish stock froma supplier, mortalities were detected together withabdominal haemorrhages in moribund fish. Bacteriologicalanalyses were carried out on skin, liver and kidney usingTrypticase soy agar (TSA) plates. After incubation at roomtemperature (RT) for 24 h almost pure cultures of yellowishcolonies were recovered from most fish. Single colonieswere grown in 10 ml of Trypticase Soy Broth (TSB) for 24 hat RT and were stored at �80 �C in 15% (vol/vol) glycerol.
Zebrafish were injected intraperitoneally with 10 ml ofthe bacterial suspension (106 cfu), and mortalities wererecorded. The selected bacteria were further character-ized phenotypically using the API 20E (BioMerieux, MarcyL’etoile, France) following 24 h incubation at 28 �C. Fur-thermore, enzymatic activities of the viable and heat-killedbacteria were characterized by API ZYM (BioMerieux, MarcyL’etoile, France) after incubation at 28 and 15 �C for 4 h.
The identification of the isolated bacteria was confirmedby sequencing the small subunit of 16S rRNA. The complete16S rRNA gene was amplified using primers F27 and R1492 aspreviously described [43]. PCR amplification was performedwith a PerkineElmer GeneAmp 2700 Thermocycler (AppliedBiosystems, Foster City, CA, USA). PCR reaction (25 ml)included 10 mM Tris; 50 mM KCl (pH 8.3); 2.5 mM MgCl2;400 mM of each deoxynucleotide; 1.25 U of Taq DNA poly-merase (Amplitaq; PE Applied Biosystems, Foster City,CA), 0.4 mM of each primer, and 100 ng of template DNA.Sequencing was conducted using the ABI Prism Big DyeTerminator cycle-sequencing reaction kit (version 3.1) andan ABI Prism 3700 automated sequencer (PE Applied Biosys-tems, Foster City, CA, USA) following the manufacturer’s
instructions. Sequences were examined using the BLASTsearch alignment tool comparison software (BLASTN) [44]with Aeromonas reference sequences obtained fromGenBank using ClustalX [45]. The alignment obtained wasfinally edited using the MacClade program [46] and directlytransferred to version 4.0b10 of PAUP* software [47].ModelTest software version 3.7 [48] was run together withPAUP*, as a guide to determine the best-fit maximum likeli-hood (ML) model for the edited alignment. ModelTestexamines maximum likelihood models ranging from simpleto complex. We also incorporated the best-fit model ofnucleotide evolution, calculated by ModelTest and PAUP*,into software PHYML [49], which uses a single, fast andaccurate algorithm to estimate large phylogenies bymaximum likelihood. Finally, the trees created by PHYMLwere edited using the software TreeViewX [50].
Production of the bacterial ECPsThe bacterial extracellular products (ECPs) were obtained bythe cellophane plate technique [51]. Briefly, volumes of 3 mlof overnight cultures at RT in TSB were spread onto sterilizedcellophane sheets placed on TSA plates. The plates wereincubated for 24 h and the cells washed off the cellophanewith PBS. The suspension was centrifuged at 10,000 � g for30 min at 4 �C, and the supernatants filtered through 0.22-mm membranes and stored at �80 �C until required. Theprotein concentration of the ECPs was evaluated by theBCA� (bicinchoninic acid) protein assay reagent (Pierce)according to the manufacturer’s instructions. The bacterialECPs were also characterized by API Zym.
Assay for haemolytic activityDue to the difficulty of obtaining enough zebrafish blood toconduct haemolysis assays, these were done using turbot(Scophthalmus maximus) blood. Turbot blood was obtainedfrom the tail vein, using Leibovitz medium (L-15, Gibco)supplemented with penicillin (100 IU ml�1), streptomycin(100 mg ml�1), heparin (10 units ml�1) and 0.1% FCS. Theresulting blood suspension was diluted five times with L-15supplemented with 2% FCS containing heparin. Ten-fold dilu-tions of bacteria and ECPs were made in PBS and 10 ml of eachsuspension were applied on TSA plates containing 5% (vol/vol) of turbot blood. After an incubation of 24 h at 28 �C,plates were examined for haemolytic activity.
Cytotoxicity assayCytotoxicity of bacteria and ECPs were assayed usingdifferent fish cell lines, namely epithelioma papulosumcyprini (EPC), rainbow trout gonad (RTG2), tail fin of turbot(TF), and ZF4 (cells of embryo of zebrafish). Cell mono-layers grown in 96-well plates were inoculated with 10 ml ofserial 10-fold dilutions of whole bacteria, heat-killed bacte-ria and bacterial ECPs and incubated at 20 �C (RTG2, EPC)or 25 �C (TF, ZF4). Total or partial destruction of mono-layers within a 3-day period was scored as a positivecytotoxic effect.
Pathogenicity
Experimental infectionsAdult fish were exposed to 2 � 106 cfu ml�1 of A. hydrophilain a total volume of 500 ml. Exposures were performed in

Immune response of zebrafish to Aeromonas hydrophila 241
duplicate with 10 fish per group. After immersion in thebacterial suspension for 5 h, the fish were moved to 2.5-ltanks. Control groups were not exposed to bacteria.Furthermore, additional groups of 10 fish were anaesthe-tized, prior to their immersion in water with bacteria,and a tail cut and dermis scratch was performed with a ster-ile scalpel. Control fish were treated in the same way, butwere not exposed to the pathogen. All experiments wereindependently conducted at least three times.
In order to estimate the lethal dose 50% (LD50) ofA. hydrophila and the ECPs, duplicated groups of adultzebrafish (8e10 fish per group) were injected intraperitone-ally with 10 ml of serial dilutions of A. hydrophila, from5 � 106 to 5 � 108 cfu ml�1, and with two-fold dilutions ofbacterial ECPs from 0.3 mg ml�1 to 1.5 mg ml�1. Controlgroups were injected with PBS. Mortalities were recordedfor 4 days.
Furthermore, three groups of 10 fish each were i.p.injected with 107 cfu per fish of viable bacteria, heat-inactivated bacteria (100 �C, 30 min), formalin-inactivatedbacteria (treatment with 0.5% formalin at 40% concentra-tion for 48 h), ECPs (150 mg per fish) or heat-inactivatedECPs. Washed bacteria in PBS (three times) were alsoinjected in order to determine possible contaminationwith extracellular products (ECPs).
To determine if temperature had any influence onbacteria virulence, the zebrafish A. hydrophila strain wasgrown at 15, 20 and 37 �C for 24 h, the number of cellswas adjusted to 1.5 � 109 cfu ml�1 and two groups of tenfish each were injected i.p for each bacterium grown atdifferent temperatures.
HistopathologyFish were i.p. injected with live Aeromonas, heat-inactivated bacteria (109 cfu ml�1) and ECPs (1.5 mg ml�1).Controls were injected with PBS. Four and 6 h post-injection5 fish were sampled and their organs fixed in 4% bufferedformalin and embedded in paraffin following routine proto-cols established in our laboratory. Longitudinal sectionswere cut at 5 mm and stained with haematoxilyn and eosin.
Effect of A. hydrophila on zebrafish kidney cells
Kidney cells isolation and in vitro experimental designKidneys from adult zebrafish were aseptically removed andhomogenized through a 100 mm mesh using Dulbecco’smodified Eagle’s medium:F-12 nutrient mixture (D-MEM:F12, Gibco) supplemented with penicillin (100 IUml�1), streptomycin (100 mg ml�1) and foetal bovine serum(FBS). The resulting cell suspension was dispensed into 96-well plates at a concentration of 2.5 � 106 cells ml�1.
Duplicated pools of kidney cells solution were treatedwith 0.1 ml of viable bacteria, heat-killed bacteria (109 cfuml�1) and bacterial ECPs (1.5 mg ml�1) to each well.Control cells were treated with PBS. After incubation withthe different treatments, cells were used for ROS and nitricoxide (NO) assays at different times post-infection.
ROS productionThe production of oxygen radicals was evaluated measuringthe luminol-chemoluminiscence response (CL) of zebrafish
kidney cells as the emission of relative luminescence units(RLU) produced by cell membrane stimulation with phorbolmyristrate acetate (1 mg ml�1) (PMA, Sigma-Aldrich Co.)and amplified by the addition of 5-amino-2,3-dihydro-1,4-phthalazinedione (Luminol, Sigma) prepared in dimethylsulphoxide (DMSO) at a final concentration of 10�4 M. Afterinitial exposition and 30 min, 1, 2 and 6 h of incubation withdifferent treatments 0.1 ml of Hank’s balanced salt solution(HBSS) containing luminol and PMA were added to eachwell. Controls without PMA were also included. The lumi-nescent emission was measured in a luminometer (Fluoros-kan Ascent, Labsystems) for 30 min at 5-min intervals.Triplicate wells were used in all experiments. We alsodetermined if bacteria were able to trigger the respiratoryburst activity without adding PMA as indicated above.
Nitric oxide assayThe production of NO was assayed following the methoddescribed by Tafalla and Novoa [52] based on the Griessreaction [53]. After 2, 6 and 24 h of incubation of the cellswith different treatments, 50 ml of cell supernatants wereremoved from individual wells and placed in a separate96-well plate per triplicate. One-hundred microlitres of1% sulphanilamide (Sigma) in 2.5% phosphoric acid wasadded to each well, followed by 100 ml of 0.1% N-naphthyl-ethylene-diamine (Sigma) in 2.5% phosphoric acid. Theoptical density was measured at 540 nm using a multiscanspectrophotometer (Labsystems). The molar concentrationof nitrite in the sample was determined from standardcurves previously generated, using known concentrationsof sodium nitrate.
Viability of kidney cellsThe kidney cells were obtained as indicated above. Twopools (45 fish per pool) of kidney cells were placed into 24-well plates at a concentration of 3 � 106 cells ml�1. Thekidney cells were treated with 10 ml of viable bacteria(109 cfu ml�1) or bacterial ECPs (1.5 mg ml�1). PBS wasadded to control cells. After 4, 24 and 48 h of incubationwith different treatments, the viable cell concentrationwas determined by Trypan Blue exclusion. Data areexpressed as percentage of cell viability with respect tothe initial cell number.
Flow cytometryAfter 24 h of incubation with viable or dead bacteria orECPs head kidney cells were removed from 24-well plates,as indicated above. Cells were centrifuged at 500 � g for10 min and resuspended in sterile PBS. Propidium iodide(Sigma) was added to a concentration of 1 mg ml�1 toexclude dead cells and debris. Flow cytometry analysiswas based on forward and side scatter on a FACScaliburflow cytometer (Beckton Dickinson) using previouslyreported settings and population adscriptions [54].
Gene expressionAdditional experimental infections were performed withviable bacteria, heat-killed bacteria and bacterial ECPs tostudy the expression of genes related to the inflammatoryresponse. Two pools of zebrafish (5 fish per pool) wereinjected i.p. with 10 ml of viable bacteria, heat-killedbacteria (109 cfu ml�1), bacterial ECPs (1.5 mg ml�1) or

242 I. Rodrıguez et al.
PBS and kidneys were removed after 0.5, 1, 2 and 6 h post-infection from duplicates of 5 fish of each treatment.
RNA extraction was performed using Trizol Reagent(Invitrogen). The reverse transcription was performedwith the SuperScript II Reverse Transcriptase (Invitrogen)following the manufacturer’s instructions.
Quantitative PCR assays were performed using the 7300Real Time PCR System (Applied Biosystems). cDNA amplifi-cation was performed using specific primers designed byPrimer 3 software [55]. Each primer (0.5 ml, 10 mM) wasmixed with 12.5 ml of SYBR green PCR master mix (AppliedBiosystems) in a final volume of 25 ml. The standard cyclingconditions were 95 �C for 10 min, followed by 40 cycles of95 �C 15 s and 60 �C for 1 min. The comparative CT method(2DCT method) was used to determine the expression levelof analysed genes [56]. The expression of the candidategenes was normalized using b-actin as a housekeepinggene. Fold units were calculated dividing the normalizedexpression values of infected tissues by the normalizedexpression values of the controls. Primer sequences areshown in Table 1.
Statistics
Student’s t-test was used to perform comparisons amongexperimental conditions. The results are expressed asmean � standard deviation and differences were consid-ered significant at p < 0.05.
Results
Identification and characterization of thepathogenic bacterial strain
Several bacterial colonies were isolated from a group ofdiseased zebrafish that presented haemorrhages in theabdomen and visceral cavity. After conducting severalexperimental infections with all the isolates, one of them(which was isolated from diseased fish in almost pureculture) was selected due to its virulence and pathogenic-ity. External symptoms (abdominal and visceral haemor-rhages and abdominal distension) were always observedafter experimental infections (Fig. 1B). Histologically, thefish did not show lesions (data not shown).
Table 1 Sequence of the primers
Gen Accession numberGenBank
S
b-Actin AF057040 CC
TNFa AY427649 AT
IL-1b BC098597 AA
IFNg AB158361 AA
iNOS AY324390 GG
The bacterial isolate was identified as belonging to themobile Aeromonas group (A. hydrophila/caviae and A.sobria [57] by using the API 20E system (data not shown).Furthermore, the 16S ribosomal sequence from the isolatedstrain showed a homology of 98%, with an e-value of 0, with32 sequences from the same region belonging to the GenusAeromonas. The phylogenetic analysis placed the zebrafishbacterial isolate in a cluster together with Aeromonashydrophila (Fig. 1C).
The bacteria and their extracellular products (ECPs)were characterized enzymatically using API Zym at differ-ent temperatures 15 and 28 �C (Table 2). The heat-inactivated bacteria and heat-inactivated ECPs did notpresent enzymatic activity. At 28 �C all bacteria werepositive for esterase, esterase lipase, leucine arylamidase,acid phosphatase, b-galactosidase and b-glucosidase whilethe bacteria at 15 �C were positive for esterase andesterase lipase. ECPs produced at 28 �C were positive foresterase, esterase lipase, leucine arylamidase and valinearylamidase. ECPs obtained at 15 �C had the sameenzymatic activities as ECPs at 28 �C with the exceptionof esterase lipase.
Pathogenicity
In vivo experimental infectionsAdult zebrafish were exposed to A. hydrophila zebrafishisolate by bath for 5 h. Both control and A. hydrophilaexposed fish showed no mortalities or signs of disease. Inthe immersion experiment using fish that were subjectedto a slight tail cut prior to immersion in A. hydrophila, therewas an average cumulative mortality rate of 100% after 24 hof exposure. Control fish, which had the same tail cut butwere not exposed to the pathogen, presented a 40% mortal-ity rate after 4 days (Fig. 2A).
Fish intraperitoneally injected with 5 � 105 cfu of A.hydrophila (bacterial suspension of 5 � 107 cfu ml�1)showed a mortality of 60% within 1 day post-injection(p.i.) while all fish died within 1 day p.i. when injectedwith concentrations of 5 � 108 cfu ml�1. The estimatedLD50 was 4 � 105 cfu (Fig. 2B). Since there were no differ-ences between washed and unwashed bacteria, weconcluded that potential contaminant ECPs were notinvolved in the pathogenicity of live bacterial preparations.
equence primers (50e30) F and R Size (bp)
GA GCA GGA GAT GGG AAC C 104AA CGG AAA CGC TCA TTG CCC AGG CCT TTT CTT CAG GT 303GC CCA GTC TGT CTC CTT CTAC CTG TAC CTG GCC TGT TG 227TC TCC ACC ATC TGC GAA TCTG ATT GCG CAA CAC ATG AT 189TC TTT CAG GAT TCG CAG GAGAGATGCAAGGTCAGCTTC 137GCAAAGCTCAGTGACTTCC

Figure 1 (A) Control fish. (B) Aspect of fish showing the external clinical signs of an experimental Aeromonas infection mainlycharacterized by distended visceral cavity and abdominal haemorrhages. (C) Phylogenetic tree showing the position of the zebra-fish A. hydrophila (in the box) compared with other reference strains.
Immune response of zebrafish to Aeromonas hydrophila 243
The i.p. injection of 1.5 mg ml�1 of ECPs caused a mor-tality of 80%, whereas 0.75 and 0.3 mg ml�1 caused 50 and20% mortality respectively, within 1 day p.i. No mortalitieswere recorded in any of the control fish injected with PBS(Fig. 2B).
Heat-inactivated or formalin-inactivated bacteria(107 cfu per fish) and heat-inactivated ECPs (150 mg perfish) did not produce any mortalities when injected inzebrafish (data not shown). However, a cumulativemortality of 100% was reached with whole bacteria andECPs 20 h post-injection.
When A. hydrophila was grown at 15, 20 and 37 �C for24 h and i.p. injected in adult fish, a cumulative mortalityof 100% was reached when using A. hydrophila grown at15 and 20 �C while a cumulative mortality of 70% wasreached with bacteria grown at 37 �C (Fig. 2C).
Haemolytic and cytotoxic activitiesThe haemolytic assays indicated that A. hydrophila strainand their ECPs were haemolytic on turbot blood agar plates(Table 3).
This A. hydrophila strain was also cytotoxic for severalfish cell lines, EPC, RTG2, TF and ZF4 (Table 4). HoweverECPs only had a cytotoxic effect on ZF4 cells. No changesin morphology were observed in cells that were exposedto heat-killed bacteria.
Effect of A. hydrophila on zebrafish kidney cells
Modulation of ROS and NO productionCo-incubation of zebrafish kidney cells with viableA. hydrophila triggered a fast production of oxygen radicalswithout stimulation with PMA in the first 30 min, however
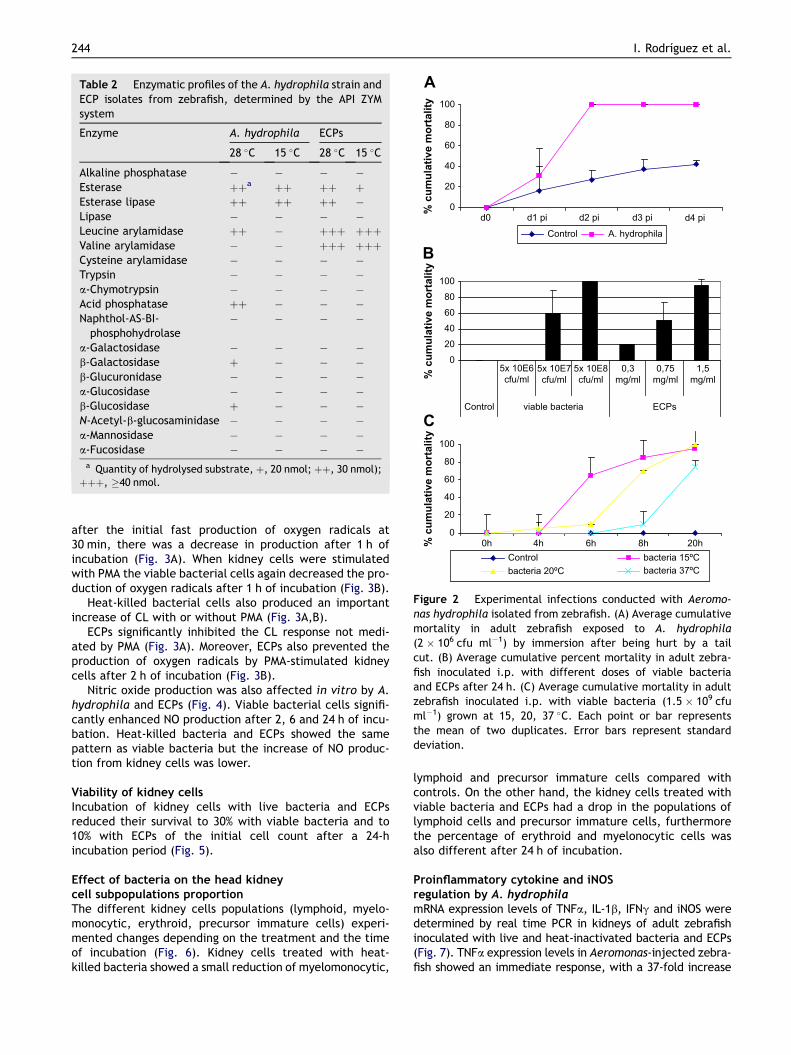
Table 2 Enzymatic profiles of the A. hydrophila strain andECP isolates from zebrafish, determined by the API ZYMsystem
Enzyme A. hydrophila ECPs
28 �C 15 �C 28 �C 15 �C
Alkaline phosphatase � � � �Esterase þþa þþ þþ þEsterase lipase þþ þþ þþ �Lipase � � � �Leucine arylamidase þþ � þþþ þþþValine arylamidase � � þþþ þþþCysteine arylamidase � � � �Trypsin � � � �a-Chymotrypsin � � � �Acid phosphatase þþ � � �Naphthol-AS-BI-
phosphohydrolase� � � �
a-Galactosidase � � � �b-Galactosidase þ � � �b-Glucuronidase � � � �a-Glucosidase � � � �b-Glucosidase þ � � �N-Acetyl-b-glucosaminidase � � � �a-Mannosidase � � � �a-Fucosidase � � � �
a Quantity of hydrolysed substrate, þ, 20 nmol; þþ, 30 nmol);þþþ, �40 nmol.
0
20
40
60
80
100
d0 d1 pi d2 pi d3 pi d4 pi
% cu
mu
lative m
ortality
Control A. hydrophila
020406080
100
5x 10E6cfu/ml
5x 10E7cfu/ml
5x 10E8cfu/ml
0,3mg/ml
0,75mg/ml
1,5mg/ml
Control ECPs
% cu
mu
lative m
ortality
0
20
40
60
80
100
0h 4h 6h 8h 20h% cu
mu
lative m
ortality
Control bacteria 15ºCbacteria 20ºC bacteria 37ºC
viable bacteria
A
C
B
Figure 2 Experimental infections conducted with Aeromo-nas hydrophila isolated from zebrafish. (A) Average cumulativemortality in adult zebrafish exposed to A. hydrophila(2 � 106 cfu ml�1) by immersion after being hurt by a tailcut. (B) Average cumulative percent mortality in adult zebra-fish inoculated i.p. with different doses of viable bacteriaand ECPs after 24 h. (C) Average cumulative mortality in adultzebrafish inoculated i.p. with viable bacteria (1.5 � 109 cfuml�1) grown at 15, 20, 37 �C. Each point or bar representsthe mean of two duplicates. Error bars represent standarddeviation.
244 I. Rodrıguez et al.
after the initial fast production of oxygen radicals at30 min, there was a decrease in production after 1 h ofincubation (Fig. 3A). When kidney cells were stimulatedwith PMA the viable bacterial cells again decreased the pro-duction of oxygen radicals after 1 h of incubation (Fig. 3B).
Heat-killed bacterial cells also produced an importantincrease of CL with or without PMA (Fig. 3A,B).
ECPs significantly inhibited the CL response not medi-ated by PMA (Fig. 3A). Moreover, ECPs also prevented theproduction of oxygen radicals by PMA-stimulated kidneycells after 2 h of incubation (Fig. 3B).
Nitric oxide production was also affected in vitro by A.hydrophila and ECPs (Fig. 4). Viable bacterial cells signifi-cantly enhanced NO production after 2, 6 and 24 h of incu-bation. Heat-killed bacteria and ECPs showed the samepattern as viable bacteria but the increase of NO produc-tion from kidney cells was lower.
Viability of kidney cellsIncubation of kidney cells with live bacteria and ECPsreduced their survival to 30% with viable bacteria and to10% with ECPs of the initial cell count after a 24-hincubation period (Fig. 5).
Effect of bacteria on the head kidneycell subpopulations proportionThe different kidney cells populations (lymphoid, myelo-monocytic, erythroid, precursor immature cells) experi-mented changes depending on the treatment and the timeof incubation (Fig. 6). Kidney cells treated with heat-killed bacteria showed a small reduction of myelomonocytic,
lymphoid and precursor immature cells compared withcontrols. On the other hand, the kidney cells treated withviable bacteria and ECPs had a drop in the populations oflymphoid cells and precursor immature cells, furthermorethe percentage of erythroid and myelonocytic cells wasalso different after 24 h of incubation.
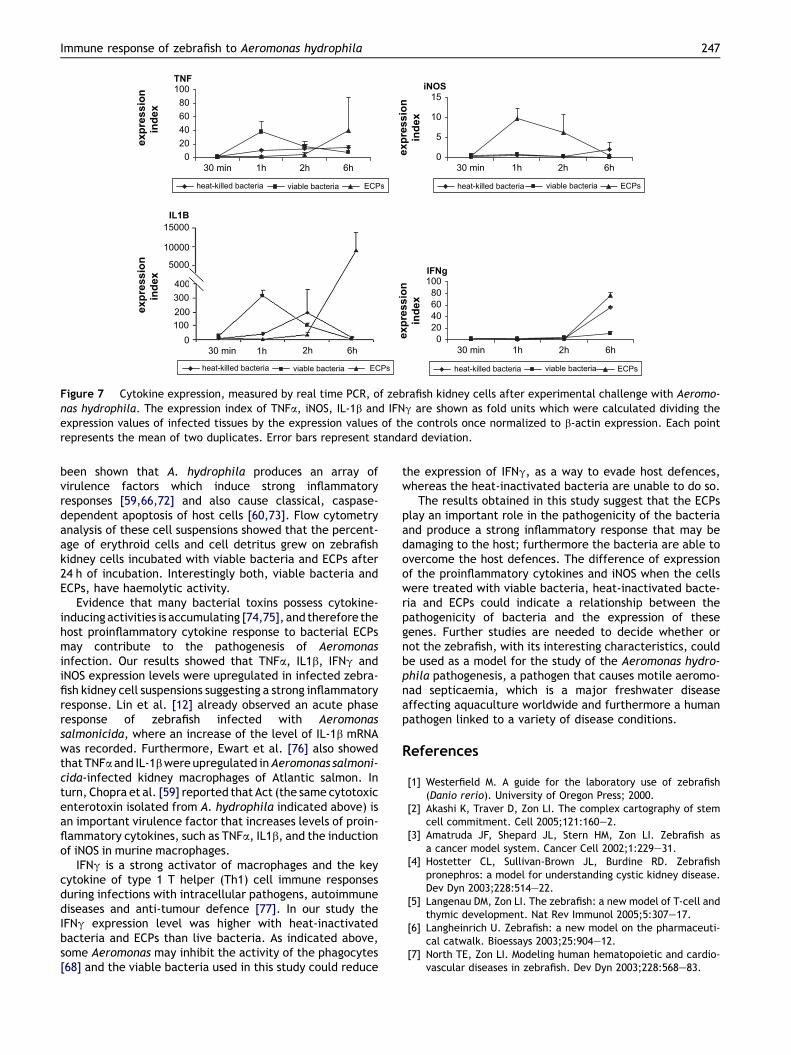
Proinflammatory cytokine and iNOSregulation by A. hydrophilamRNA expression levels of TNFa, IL-1b, IFNg and iNOS weredetermined by real time PCR in kidneys of adult zebrafishinoculated with live and heat-inactivated bacteria and ECPs(Fig. 7). TNFa expression levels in Aeromonas-injected zebra-fish showed an immediate response, with a 37-fold increase

Table 3 Cytotoxic and haemolytic activities of the strainA. hydrophila and ECPs
Cytotoxic responseon fish cell linea
Haemolysis againsterythrocytes fromb
RTG2 EPC TF ZF4 Turbot
A. hydrophila þ þ þ þ þECPs � � � þ þHeat-killed
bacteria� � � � n.d.
n.d., no data.a Toxic effect recorded after 3 days.b Haemolytic activity recorded after 24 h.
02468
10121416
PBS
viab
leba
cter
ia
heat
-ki
lled
heat
-ki
lled
heat
-ki
lled
ECPs
ECPs
PBS
viab
leba
cter
ia
viab
leba
cter
ia
PBS
ECPs
2h 6h 24h
um
n
itrito
* * *
*
* * *
* *
Figure 4 Nitric oxide, measured as nitrite concentration, re-lease of zebrafish kidney cells after in vitro infection at differ-ent times post-infection. Each bar represents the mean of twoduplicates. Error bars represent standard deviation. Statisticaldifference from PBS group (*).
Immune response of zebrafish to Aeromonas hydrophila 245
over control levels at 1 h p.i., returning at control levels at 6 hp.i. Fish injected with heat-inactivated bacteria had a slightinduction maintained throughout the different time points.ECPs induced a 40-fold increase at 6 h p.i. when comparedwith control samples. IL-1b expression levels had a 315-foldinduction at 1 h p.i. in bacteria-injected fish, they began todecline at 2 h p.i., returning to control levels at 6 h p.i. IL-1bexpression levels of heat-inactivated bacteria-injected fish
0
5
10
15
20
25
0 min 30 min 1h 2h 6h
Stim
ulatio
n In
dex
heat-killed bacteria viable bacteria ECPs
00,5
11,5
22,5
33,5
0 min 30 min 1h 2h 6h
Stim
ulatio
n In
dex
heat-killed bacteria viable bacteria ECPs
A
B
Figure 3 Effect of the Aeromonas hydrophila in vitro infec-tion on the production of oxygen radicals. (A) Triggering ofthe respiratory burst activity of head kidney cells infectedwith bacteria, heat-killed bacteria or bacterial ECPs. Stimula-tion indexes were calculated dividing the luminol amplified-chemiluminiscence response of infected cells by that ofuntreated controls. (B) Respiratory burst activity of zebrafishkidney cells mediated by PMA after in vitro infection withbacteria, heat-killed bacteria or bacterial ECPs. Stimulationindexes were calculated dividing the PMA-mediated luminolamplified-chemiluminiscence response of infected cells by thatof untreated controls. Each point represents the mean of twoduplicates. Error bars represent standard deviation.
peaked at 2 h p.i. with a 195-fold increase over the controllevels and returned to control levels at 6 h p.i. ECP-injectedfish showed a remarkable increase of 9020-fold increase at6 h p.i. The IFNg expression levels of zebrafish injected withlive bacteria showed an 11-fold increase at 6 h p.i. Whenheat-inactivated bacteria were used the expression levels at6 h p.i. had a 56-fold increase. IFNg expression levels of ECP-injected fish peaked at 6 h p.i. and was approximately 78-fold greater than control fish. iNOS was not induced in wholebacteria and heat-inactivated bacteria-infected fish through-out thesample times.HoweverECPs induceda10-fold increaseof iNOS at 1 h p.i.
Discussion
Our results indicate that the bacterial strain isolated froma group of sick zebrafish and identified as a motile Aeromo-nas strain, are responsible for the disease outbreak.
The correct identification of Aeromonas species hasbeen controversial [58]. Using the API 20E system the iso-lated bacteria was identified as a motile Aeromonas strain,three phenospecies (A. hydrophila, A. caviae and A. sobria)have been recognized within this group [57]. Since, whenusing biochemical tests, there are also difficulties in theidentification of motile Aeromonas at the species level,the 16S rRNA of the isolated strain was sequenced allowingthe identification of A. hydrophila.
0
20
40
60
80
100
4h 24h 48h
% viab
ility
Controlviable bacteriaECPs
* *
Figure 5 Effect of in vitro infection on the immune kidneycells viability and population distribution. Viability of headkidney cells incubated for 24 h with viable bacteria, heat-killedbacteria or bacterial ECPs. Each bar represents the mean oftwo duplicates. Error bars represent standard deviation. Statis-tical difference from control group (*).

0 200 400 600 800 1000FSC-H
R1 58,60%
R2 13,46%
R3 9,34%R4 7,38%
200 400 600 800 10000FSC-H
R1 79,20%
R2 14,30%
R3 0,10%R4 0%
0 200 400 600 800 1000FSC-H
R1 81%
R2 10,50%
R3 0,60%R4 0,08%
0 200 400 600 800 1000
FSC-H
R1 79,50%
R2 7,38%
R3 3,52%R4 1,18%
SS
C-H
104
SS
C-H
SS
C-H
SS
C-H
100
101
102
103
104
100
101
102
103
104
100
101
102
103
104
100
101
102
103
A
C
B
D
Figure 6 Flow cytometry of zebrafish kidney cell populations analysed by size (forward scatter; FSC) and granularity (side scat-ter; SSC). Kidney cells were untreated (A) or treated with viable (B) or heat-killed bacteria (C) and ECPs (D) for 24 h. Erythroid (R1),myeloid (R2), lymphoid (R3) or precursors (R4) populations are indicated in each panel.
246 I. Rodrıguez et al.
A. hydrophila and other motile Aeromonas can causefatal septicaemia in fish but also seems to be more harmfulfor humans than was previously believed, mainly due to: (i)its ability to produce an important number of virulencefactors such as enterotoxins, proteases, lipases, etc; (ii)its resistance to many common antibiotics such as penicillinand ampicillin; and (iii) the significant effects that thebacteria produce on the host including cytotoxicity andmassive inflammation [59e61].
We were able to reproduce disease symptoms, similar tothose obtained by Pullium et al. [38] and mortalities in fishafter experimental infection by i.p. injection or woundedimmersed fish. According to the degree of virulencedescribed by Mittal et al. [62], our isolated strain wasincluded in the virulent category (LD50, 104e105 cfu). Thesefish showed clinical symptoms such as haemorrhages andabdominal swelling. However histological lesions were notobserved because the peracute form of disease may killfish before any changes are evident [63].
A. hydrophila virulence is frequently associated with theproduction of several extracellular toxins and enzymes[41,64]. The extracellular products of our A. hydrophilastrain are able to produce similar effects to those of viablebacteria, sharing most of the enzymatic activities andhaemolytic and cytotoxic activity. The lethal dose rangedfrom 12 to 60 mg of protein per gram of fish; this rangewas comparable to those reported previously for Aeromo-nas species by Santos et al. [65].
The lack of mortalities in the experimental infectionwith heat- or formalin-inactivated bacteria together with
the absence of bacterial growth in the histopathologicalanalysis of zebrafish tissues suggests that the quick mortal-ity response found is the effect of potent heat-labilevirulence factors or toxins (ECPs) that may play animportant role in this bacterial pathogenicity.
Bacterial infection was able to induce the release ofROS. These results agree with the detection of respiratoryburst related genes on an SSH library conducted on A.salmonicida and Staphylococcus aureus-infected zebrafish[12]. Furthermore, Ribardo et al. [66] showed that a cyto-toxic enterotoxin (Act) isolated from A. hydrophilaincreased the ROS production in murine macrophages. How-ever, this response lasts a very short time. The resistance ofA. hydrophila against oxygen radicals has already beenreported [67]. These authors describe that some Aeromo-nas bacteria possess an Mn-superoxide dismutase (Mn-SOD)and catalase that protects them from the external anionsuperoxide and hydrogen peroxide generated by macro-phages. Furthermore, Jin and Houston [68] found thatsome Aeromonas had toxins that may inhibit the phago-cytes’ activity. The inhibition of the respiratory burst activ-ity was previously shown in fish when kidney macrophageswere incubated with Vibrio pelagius and its ECPs [69].
NO production was enhanced by viable bacteria, heat-inactivated bacteria and ECPs. Campos-Perez et al. [70] haveshown in rainbow trout that an increase of NO may havea role against bacterial infections, however NO can alsobe potentially dangerous for the host cells and tissues [71].
Kidney cell suspension viability was reduced to 30% whenincubated with viable bacteria and to 10% with ECPs. It has
TNF
020406080
100
30 min 1h 2h 6h
exp
ressio
n
in
dex
exp
ressio
n
in
dex
heat-killed bacteria viable bacteria ECPs
iNOS
0
5
10
15
30 min 1h 2h 6h
exp
ressio
n
in
dex
heat-killed bacteria viable bacteria ECPs
IFNg
020406080
100
30 min 1h 2h 6h
exp
ressio
n
in
dex
heat-killed bacteria viable bacteria ECPsheat-killed bacteria viable bacteria ECPs
15000
10000
5000
400300
1000
30 min 1h 2h 6h
IL1B
200
Figure 7 Cytokine expression, measured by real time PCR, of zebrafish kidney cells after experimental challenge with Aeromo-nas hydrophila. The expression index of TNFa, iNOS, IL-1b and IFNg are shown as fold units which were calculated dividing theexpression values of infected tissues by the expression values of the controls once normalized to b-actin expression. Each pointrepresents the mean of two duplicates. Error bars represent standard deviation.
Immune response of zebrafish to Aeromonas hydrophila 247
been shown that A. hydrophila produces an array ofvirulence factors which induce strong inflammatoryresponses [59,66,72] and also cause classical, caspase-dependent apoptosis of host cells [60,73]. Flow cytometryanalysis of these cell suspensions showed that the percent-age of erythroid cells and cell detritus grew on zebrafishkidney cells incubated with viable bacteria and ECPs after24 h of incubation. Interestingly both, viable bacteria andECPs, have haemolytic activity.
Evidence that many bacterial toxins possess cytokine-inducing activities is accumulating [74,75], and therefore thehost proinflammatory cytokine response to bacterial ECPsmay contribute to the pathogenesis of Aeromonasinfection. Our results showed that TNFa, IL1b, IFNg andiNOS expression levels were upregulated in infected zebra-fish kidney cell suspensions suggesting a strong inflammatoryresponse. Lin et al. [12] already observed an acute phaseresponse of zebrafish infected with Aeromonassalmonicida, where an increase of the level of IL-1b mRNAwas recorded. Furthermore, Ewart et al. [76] also showedthat TNFa and IL-1b were upregulated in Aeromonas salmoni-cida-infected kidney macrophages of Atlantic salmon. Inturn, Chopra et al. [59] reported that Act (the same cytotoxicenterotoxin isolated from A. hydrophila indicated above) isan important virulence factor that increases levels of proin-flammatory cytokines, such as TNFa, IL1b, and the inductionof iNOS in murine macrophages.
IFNg is a strong activator of macrophages and the keycytokine of type 1 T helper (Th1) cell immune responsesduring infections with intracellular pathogens, autoimmunediseases and anti-tumour defence [77]. In our study theIFNg expression level was higher with heat-inactivatedbacteria and ECPs than live bacteria. As indicated above,some Aeromonas may inhibit the activity of the phagocytes[68] and the viable bacteria used in this study could reduce
the expression of IFNg, as a way to evade host defences,whereas the heat-inactivated bacteria are unable to do so.
The results obtained in this study suggest that the ECPsplay an important role in the pathogenicity of the bacteriaand produce a strong inflammatory response that may bedamaging to the host; furthermore the bacteria are able toovercome the host defences. The difference of expressionof the proinflammatory cytokines and iNOS when the cellswere treated with viable bacteria, heat-inactivated bacte-ria and ECPs could indicate a relationship between thepathogenicity of bacteria and the expression of thesegenes. Further studies are needed to decide whether ornot the zebrafish, with its interesting characteristics, couldbe used as a model for the study of the Aeromonas hydro-phila pathogenesis, a pathogen that causes motile aeromo-nad septicaemia, which is a major freshwater diseaseaffecting aquaculture worldwide and furthermore a humanpathogen linked to a variety of disease conditions.
References
[1] Westerfield M. A guide for the laboratory use of zebrafish(Danio rerio). University of Oregon Press; 2000.
[2] Akashi K, Traver D, Zon LI. The complex cartography of stemcell commitment. Cell 2005;121:160e2.
[3] Amatruda JF, Shepard JL, Stern HM, Zon LI. Zebrafish asa cancer model system. Cancer Cell 2002;1:229e31.
[4] Hostetter CL, Sullivan-Brown JL, Burdine RD. Zebrafishpronephros: a model for understanding cystic kidney disease.Dev Dyn 2003;228:514e22.
[5] Langenau DM, Zon LI. The zebrafish: a new model of T-cell andthymic development. Nat Rev Immunol 2005;5:307e17.
[6] Langheinrich U. Zebrafish: a new model on the pharmaceuti-cal catwalk. Bioessays 2003;25:904e12.
[7] North TE, Zon LI. Modeling human hematopoietic and cardio-vascular diseases in zebrafish. Dev Dyn 2003;228:568e83.

248 I. Rodrıguez et al.
[8] Stern HM, Zon LI. Cancer genetics and drug discovery in thezebrafish. Nat Rev Cancer 2003;3:533e9.
[9] Trede NS, Langenau DM, Traver D, Look AT, Zon LI. The use ofzebrafish to understand immunity. Immunity 2004;20:367e79.
[10] Weber GJ, Choe SE, Dooley KA, Paffett-Lugassy NN, Zhou Y,Zon LI. Mutant-specific gene programs in the zebrafish. Blood2005;106:521e30.
[11] Davis JM, Clay H, Lewis JL, Ghori N, Herbomel P,Ramakrishnan L. Real-time visualization of mycobacteriume
macrophage interactions leading to initiation of granulomaformation in zebrafish embryos. Immunity 2002;17:693e702.
[12] Lin B, Chen S, Cao Z, Lin Y, Mo D, Zhang H, et al. Acute phaseresponse in zebrafish upon Aeromonas salmonicida and Staph-ylococcus aureus infection: Striking similarities and obviousdifferences with mammals. Mol Immunol 2007;44:295e301.
[13] Neely MN, Pfeifer JD, Caparon M. Streptococcusezebrafishmodel of bacterial pathogenesis. Infect Immun 2002;70:3904e14.
[14] O’Toole R, Von Hofsten J, Rosqvist R, Olsson PE, Wolf-Watz H.Visualisation of zebrafish infection by GFP-labelled Vibrioanguillarum. Microb Pathog 2004;37:41e6.
[15] Pressley ME, Phelan 3rd PE, Witten PE, Mellon MT, Kim CH.Pathogenesis and inflammatory response to Edwardsiellatarda infection in the zebrafish. Dev Comp Immunol 2005;29:501e13.
[16] van der Sar AM, Appelmelk BJ, Vandenbroucke-Grauls CM,Bitter W. A star with stripes: zebrafish as an infection model.Trends Microbiol 2004;12:451e7.
[17] van der Sar AM, Musters RJ, van Eeden FJ, Appelmelk BJ,Vandenbroucke-Grauls CM, Bitter W. Zebrafish embryos asa model host for the real time analysis of Salmonellatyphimurium infections. Cell Microbiol 2003;5:601e11.
[18] LaPatra SE, Barone L, Jones GR, Zon LI. Effects of infectioushematopoietic necrosis virus and infectious pancreaticnecrosis virus infection on hematopoietic precursors of thezebrafish. Blood Cells Mol Dis 2000;26:445e52.
[19] Novoa B, Romero A, Mulero V, Rodriguez I, Fernandez I,Figueras A. Zebrafish (Danio rerio) as a model for the studyof vaccination against viral haemorrhagic septicemia virus(VHSV). Vaccine 2006;24:5806e16.
[20] Phelan PE, Pressley ME, Witten PE, Mellon MT, Blake S,Kim CH. Characterization of snakehead rhabdovirus infectionin zebrafish (Danio rerio). J Virol 2005;79:1842e52.
[21] Sanders GE, Batts WN, Winton JR. Susceptibility of zebrafish(Danio rerio) to a model pathogen, spring viremia of carpvirus. Comp Med 2003;53:514e21.
[22] Monfort P, Baleux B. Dynamics of Aeromonas hydrophila,Aeromonas sobria, and Aeromonas caviae in a sewage treat-ment pond. Appl Environ Microbiol 1990;56:1999e2006.
[23] Janda JM, Abbott SL. Evolving concepts regarding the genusAeromonas: an expanding Panorama of species, diseasepresentations, and unanswered questions. Clin Infect Dis1998;27:332e44.
[24] Begue RE, Castellares G, Hayashi KE, Ruiz R, Meza R,English CK, et al. Diarrheal disease in Peru after theintroduction of cholera. Am J Trop Med Hyg 1994;51:585e9.
[25] Kuhn I, Albert MJ, Ansaruzzaman M, Bhuiyan NA, Alabi SA,Islam MS, et al. Characterization of Aeromonas spp. isolatedfrom humans with diarrhea, from healthy controls, and fromsurface water in Bangladesh. J Clin Microbiol 1997;35:369e73.
[26] Ogunsanya TI, Rotimi VO, Adenuga A. A study of theaetiological agents of childhood diarrhoea in Lagos, Nigeria.J Med Microbiol 1994;40:10e4.
[27] Funada H, Matsuda T. Aeromonas bacteremia in patients withhematologic diseases. Intern Med 1997;36:171e4.
[28] Gonzalez-Barca E, Ardanuy C, Carratala J, Sanchez A, Fernandez-Sevilla A, Granena A. Fatal myofascial necrosis due to imipenem-resistantAeromonashydrophila. ScandJ InfectDis 1997;29:91e2.
[29] Takano Y, Asao Y, Kohri Y, Oida K, Imanaka T. Fulminantpneumonia and sepsis due to Aeromonas hydrophila in analcohol abuser. Intern Med 1996;35:410e2.
[30] Clark NM, Chenoweth CE. Aeromonas infection of thehepatobiliary system: report of 15 cases and review of theliterature. Clin Infect Dis 2003;37:506e13.
[31] Ko WC, Chuang YC. Aeromonas bacteremia: review of 59episodes. Clin Infect Dis 1995;20:1298e304.
[32] Lau SM, Peng MY, Chang FY. Outcomes of Aeromonas bacter-emia in patients with different types of underlying disease.J Microbiol Immunol Infect 2000;33:241e7.
[33] Candan A, Kucuker MA, Karatas S. Motile aeromonad septice-mia in Salmo salar cultured in the black sea in Turkey. Bull EurAssoc Fish Pathol 1995;15:195e6.
[34] Gratzek JB. An overview of the diseases of ornamental fishes.Proceedings of the 4th KalKan Symposium 1980:25e39.
[35] Kozinska A, Figueras MJ, Chacon MR, Soler L. Phenotypic char-acteristics and pathogenicity of Aeromonas genomospeciesisolated from common carp (Cyprinus carpio L.). J Appl Micro-biol 2002;93:1034e41.
[36] Ogara WO, Mbuthia PG, Kaburia HFA, Sorum H, Kagunya DK,Nduthu DI, et al. Motile aeromonads associated with rainbowtrout (Oncorhynchus mykiss) mortality in Kenya. Bull Eur AssocFish Pathol 1998;18:7e9.
[37] Paniagua C, Rivero O, Anguita J, Naharro G. Pathogenicityfactors and virulence for rainbow trout (Salmo gairdneri) ofmotile Aeromonas spp. isolated from a river. J Clin Microbiol1990;28:350e5.
[38] Pullium JK, Dillehay DL, Webb S. High mortality in zebrafish(Danio rerio). Contemp Top Lab Anim Sci 1999;38:80e3.
[39] Toranzo AE, Baya AM, Romalde JL, Hetrick FM. Association ofAeromonas sobria with mortalities of adult gizzard shad,Dorosoma cepedianum Lesueur. J Fish Dis 1989;12:439e48.
[40] Wang C, Silva JL. Prevalence and characteristics of Aeromonasspecies isolated from processed channel catfish. J Food Prot1999;62:30e4.
[41] Allan BJ, Stevenson RM. Extracellular virulence factors ofAeromonas hydrophila in fish infections. Can J Microbiol1981;27:1114e22.
[42] Ljungh A, Wretlind B, Mollby R. Separation and characteriza-tion of enterotoxin and two haemolysins from Aeromonashydrophila. Acta Pathol Microbiol Scand B 1981;89:387e97.
[43] Edwards U, Rogall T, Blocker H, Emde M, Bottger EC. Isolationand direct complete nucleotide determination of entiregenes. Characterization of a gene coding for 16S ribosomalRNA. Nucleic Acids Res 1989;17:7843e53.
[44] Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic localalignment search tool. J Mol Biol 1990;215:403e10.
[45] Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F,Higgins DG. The CLUSTAL_X windows interface: flexiblestrategies for multiple sequence alignment aided by qualityanalysis tools. Nucleic Acids Res 1997;25:4876e82.
[46] Maddison WP, Maddison DR. Interactive analysis of phylogenyand character evolution using the computer programMacClade. Folia Primatol (Basel) 1989;53:190e202.
[47] Swofford DL. PAUP*. Phylogenetic analysis using parsimony(* and other methods). Version 4.0b4a. Sunderland, MA:Sinauer Associates; 2000.
[48] Posada D, Crandall KA. MODELTEST: testing the model of DNAsubstitution. Bioinformatics 1998;14:817e8.
[49] Guindon S, Gascuel O. A simple, fast, and accurate algorithmto estimate large phylogenies by maximum likelihood. SystBiol 2003;52:696e704.
[50] Page RD. TreeView: an application to display phylogenetictrees on personal computers. Comput Appl Biosci 1996;12:357e8.
[51] Liu PV. Survey of hemolysin production among species ofpseudomonads. J Bacteriol 1957;74:718e27.

Immune response of zebrafish to Aeromonas hydrophila 249
[52] Tafalla C, Novoa B. Requirements for nitric oxide productionby turbot (Scophthalmus maximus) head kidney macrophages.Dev Comp Immunol 2000;24:623e31.
[53] Green LC, Wagner DA, Glogowski J, Skipper PL,Wishnok JS, Tannenbaum SR. Analysis of nitrate, nitrite,and [15N]nitrate in biological fluids. Anal Biochem 1982;126:131e8.
[54] Traver D, Paw BH, Poss KD, Penberthy WT, Lin S, Zon LI.Transplantation and in vivo imaging of multilineage engraft-ment in zebrafish bloodless mutants. Nat Immunol 2003;4:1238e46.
[55] Rozen S, Skaletsky H. Primer3 on the WWW for general usersand for biologist programmers. Methods Mol Biol 2000;132:365e86.
[56] Livak KJ, Schmittgen TD. Analysis of relative gene expressiondata using real-time quantitative PCR and the 2(-Delta DeltaC(T)) method. Methods 2001;25:402e8.
[57] Popoff M. Genus III Aeromonas. In: Holt JG, editor. Bergey’smanual of determinative bacteriology. Baltimore: Williams &Wilkins; 1984. p. 545e8.
[58] Cipriano RC. Aeromonas hydrophyla and motile aeromonadsepticemias of fish. Revision of Fish Disease Leaflet 2001;68.
[59] Chopra AK, Xu X, Ribardo D, Gonzalez M, Kuhl K, Peterson JW,et al. The cytotoxic enterotoxin of Aeromonas hydrophilainduces proinflammatory cytokine production and activatesarachidonic acid metabolism in macrophages. Infect Immun2000;68:2808e18.
[60] Galindo CL, Gutierrez Jr C, Chopra AK. Potential involvementof galectin-3 and SNAP23 in Aeromonas hydrophila cytotoxicenterotoxin-induced host cell apoptosis. Microb Pathog 2006;40:56e68.
[61] Saavedra MJ, Guedes-Novais S, Alves A, Rema P, Tacao M,Correia A, et al. Resistance to beta-lactam antibiotics inAeromonas hydrophila isolated from rainbow trout (Onco-rhynchus mykiss). Int Microbiol 2004;7:207e11.
[62] Mittal KR, Lalonde G, Leblanc D, Olivier G, Lallier R.Aeromonas hydrophila in rainbow trout: relation betweenvirulence and surface characteristics. Can J Microbiol 1980;26:1501e3.
[63] Walsh AH. Biology and disease of fish. Orlando, FL: AcademicPress; 1984.
[64] Ljungh A, Wadstrom T. Aeromonas toxins. Pharmacol Ther1981;15:339e54.
[65] Santos Y, Toranzo AE, Barja JL, Nieto TP, Villa TG. Virulenceproperties and enterotoxin production of Aeromonas strainsisolated from fish. Infect Immun 1988;56:3285e93.
[66] Ribardo DA, Kuhl KR, Boldogh I, Peterson JW, Houston CW,Chopra AK. Early cell signaling by the cytotoxic enterotoxinof Aeromonas hydrophila in macrophages. Microb Pathog2002;32:149e63.
[67] Leclere V, Bechet M, Blondeau R. Functional significance ofa periplasmic Mn-superoxide dismutase from Aeromonashydrophila. J Appl Microbiol 2004;96:828e33.
[68] Jin GF, Houston CW. Effect of Aeromonas hydrophila entero-toxins on function of mouse phagocytes. Dig Dis Sci 1992;37:1697e703.
[69] Villamil L, Figueras A, Aranguren R, Novoa B. Non-specific im-mune response of turbot, Scophthalmus maximus (L.), exper-imentally infected with a pathogenic Vibrio pelagius. J FishDis 2003;26:321e9.
[70] Campos-Perez JJ, Ellis AE, Secombes CJ. Toxicity of nitric ox-ide and peroxynitrite to bacterial pathogens of fish. Dis AquatOrgan 2000;43:109e15.
[71] Braun JS, Novak R, Gao G, Murray PJ, Shenep JL. Pneumolysin,a protein toxin of Streptococcus pneumoniae, induces nitricoxide production from macrophages. Infect Immun 1999;67:3750e6.
[72] Galindo CL, Sha J, Ribardo DA, Fadl AA, Pillai L, Chopra AK.Identification of Aeromonas hydrophila cytotoxic entero-toxin-induced genes in macrophages using microarrays. JBiol Chem 2003;278:40198e212.
[73] Galindo CL, Fadl AA, Sha J, Gutierrez Jr C, Popov VL,Boldogh I, et al. Aeromonas hydrophila cytotoxic enterotoxinactivates mitogen-activated protein kinases and induces apo-ptosis in murine macrophages and human intestinal epithelialcells. J Biol Chem 2004;279:37597e612.
[74] Henderson B, Poole S, Wilson M. Bacterial modulins: a novel classof virulence factorswhich cause host tissue pathology by inducingcytokine synthesis. Microbiol Rev 1996;60:316e41.
[75] Henderson B, Wilson M, Wren B. Are bacterial exotoxins cyto-kine network regulators? Trends Microbiol 1997;5:454e8.
[76] Vanya Ewart K, Williams J, Richards RC, Gallant JW, Melville K,Douglas SE. The early response of Atlantic salmon (Salmo salar)macrophagesexposed invitro toAeromonassalmonicidaculturedin broth and in fish. Dev Comp Immunol 2008;32:380e90.
[77] Bogdan C, Mattner J, Schleicher U. The role of type I inter-ferons in non-viral infections. Immunol Rev 2004;202:33e48.




















