
Combinatorial Algorithms; Generation, Enumeration, and Search
Upload
khangminh22Category
view
0download
0
Vol. 38, No. 1APPLIED AND ENVIRONMENTAL MICROBIOLOGY, July 1979, p. 108-1130099-2240/79/07-0108/06$02.00/0
Membrane Filter Procedure for Enumeration of Aeromonashydrophila in Fresh Waters
S. R. RIPPEY AND V. J. CABELLI*Marine Field Station, Health Effects Research Laboratory-Cincinnati, U.S. Environmental Protection
Agency, West Kingston, Rhode Island 02892 and Department of Microbiology, University ofRhode Island,Kingston, Rhode Island 02881
Received for publication 11 April 1979
A membrane filter method (mA) for the enumeration ofAeromonas hydrophilain natural water samples was developed. The complex, primary medium employstrehalose as a fermentable carbohydrate and ampicillin and ethanol as selectiveinhibitors. After 20 h of incubation at 37°C, an in situ mannitol fermentation testfollowed by an in situ oxidase test is used to further differentiate A. hydrophilafrom other aquatic and terrestrial microorganisms present in freshwaters. Theprimary medium decreases background microbial growth by about two orders ofmagnitude. The recoveries on mA medium from suspensions of A. hydrophilaprepared from pure cultures and held for 24 h at 15'C exceeded 95% of therecoveries on brain-heart infusion agar spread plates. The confirmation rate forcolonies designated A. hydrophila was 98%, whereas 11% of the presumptivelynegative colonies were, in fact, A. hydrophila. Recoveries of A. hydrophila fromfresh, surface water samples exceeded recoveries by the other methods examined.
Aeromonas hydrophila is a gram-negative,oxidase-positive, facultatively anaerobic, polarlyflagellated, rod-shaped bacterium. It has mostrecently been classified as a member of the fam-ily Vibrionaceae and the genus Aeromonas (20).Numerous recommendations on speciation havebeen proposed (19, 22). Ewing et al. (10) includedonly three species within the genus, A. hydro-phila, A. salmonicida, and A. shigelloides.Their definition of A. hydrophila will be used inthe present investigation because the necessityfor further differentiation remains to be deter-mined from physiological, genetic, and ecologicalanalyses of a larger number of marine and fresh-water isolates.A. hydrophila was originally isolated and de-
scribed in 1891 by Sanarelli, who demonstratedpathogenicity of the organism for frogs. It isconsidered a resident aquatic microorganism (20,23), probably multiplying therein under appro-priate conditions. Although it has been reportedas part of the normal flora of fishes (25), it alsohas been associated with epizootic incidentsamong aquatic fauna (23).
Recent reports have increasingly associatedthe organism with human infections. A. hydro-phila has been identified as the etiological agentin a number of pathological conditions (5), in-cluding peritonitis, gastroenteritis, meningitis,and septicemia. The organism has been reportedto produce an enterotoxin (1, 14) as well as a
hemolysin and a cytotoxic protein (27). Of spe-cial interest to aquatic microbiologists are sev-eral reports of individuals with A. hydrophilawound infections associated with aquatic inju-ries (11, 18).A. hydrophila is rarely recovered from human
intestines (12, 26), although high densities of theorganism are consistently found in the influentto sewage treatment plants (J. Miescier, per-sonal communication). This implicates sewagelines as sites of multiplication for the organismsdischarged into the receiving waters. However,the densities of the organism in certain fresh-waters exceed those which could be accountedfor by direct contamination with sewage, sug-gesting possible effects of nutrient loading. Thus,there are at least three potential sources of theA. hydrophila recovered from freshwaters: fish,sewage effluents, and the aquatic environmentitself (presumably under the influence of nutri-ent inputs). Shubert (21) has attempted to iden-tify sources of the organism depending on thebiotypes present. He reports that anaerogenicbiotypes predominate in waters receiving highnutrient loads (especially "sewage waters"),whereas the aerogenic biotypes predominate inoligosaprobic waters. Unpublished data fromthis laboratory suggest that the proportions oflactose-fermenting aeromonads vary in a similarmanner; that is, significantly greater numbers oflactose-positive aeromonads occur in sewage
108

ENUMERATION OF A. HYDROPHILA 109
treatment plant effluents than in non-sewage-contaminated waters.The evaluation of the human and nonhuman
health significance of A. hydrophila in aquaticenvironments, the association of certain biotypeswith sources of pollution, and the relationship ofthe density of the organism to nutrient loadingnecessitated the development of a sensitive, fac-ile, and accurate enumeration method. Certainmedia which are selective or differential or bothhave been described for the isolation of A. hy-drophila (15, 24); however, none has been de-veloped specifically for use with membrane fil-tration procedures. The development and eval-uation of such a method is the subject of thisreport.
MATERLALS AND METHODSCultures. The A. hydrophila strains used in this
study were isolated on a complex, nonselective me-dium from water samples collected at various geo-graphical locations in the United States. This wasdone so that the test strains used in the developmentand evaluation of the medium would not have beenpreselected for antibiotic resistance.mA method. The membrane filter method (mA)
for the isolation of A. hydrophila employs growth ina selective, differential recovery medium followed bytwo in situ tests for isolate identification. The mediumis prepared by adding the following ingredients (in g/100 ml) to deionized water: tryptose, 0.5; trehalose, 0.5;yeast extract, 0.2; NaCl, 0.3; KCI, 0.2; MgSO4.7H20,0.02; FeCl366H20, 0.01; and bromothymol blue (FisherScientific Co., Fairlawn, N.J.), 0.004. The ingredientsare dissolved at room temperature, the pH is adjustedto 8.0 with 10 N NaOH, 1.5 g of agar is added, and themixture is autoclaved at 121°C for 15 min. After theaddition of 1.0 ml of ethanol, the mixture is cooled to50°C, 2 mg of ampicillin and 10 mg of sodium deoxy-cholate (Fisher Scientific Co.) are added, and themedium is dispensed into sterile petri plates (50 by 12mm; 5 ml/plate). It is recommended that the pouredplates be stored in the dark at 4°C and used within 6weeks.
After membrane filtration of water samples, thefilters (Millipore HC filters; diameter, 47 mm; poresize, 0.7 um) are placed on the mA agar plates whichare incubated in an inverted position for 20 h at 35 to37°C. Typical A. hydrophila colonies are circular,convex, yellow, and 1 to 3 mm in diameter.
Although clinical isolates of A. hydrophila havebeen reported to be resistant to both ampicillin andnovobiocin and novobiocin has been used as an inhib-itor in another isolation medium (24), the decision wasmade to use ampicillin in the primary medium. Thiscame from the examination of 113 environmentalstrains for their susceptibility to antibiotics by diskassay. These A. hydrophila strains were isolated fromsamples collected in various parts of the United States.As expected, most of the strains were resistant to bothampicillin (10-pg disk) and novobiocin (10-,ug disk).However, only 1 of the 113 strains examined (0.9%)
was sensitive to the former, whereas 13 strains (11.5%)were sensitive to the latter. It was found that ampicil-lin could be used in concentrations of up to 20 ,ug/mlwithout hindering the quantitation of A. hydrophila.
Trehalose was included in the medium as the car-bohydrate source because all strains of A. hydrophilaferment this carbohydrate (9, 19, 20). Mannitol wouldhave been a better choice than trehalose since it ismore specific for the organism. However, when man-nitol was substituted for trehalose in the primarymedium, background contamination increased, andthe color distinction between fermenters and nonfer-menters was less clear. Sodium deoxycholateheightened the yellow color of the trehalose-ferment-ing colonies, allowing a clearer distinction betweenthem and the blue-green, nonfermenting backgroundorganisms. Ethanol was included in the medium toinhibit overgrowth of the filter by Klebsiella spp.,which occasionally hindered quantification at certainsampling sites.
In situ tests. These tests were devised to augmentthe differentiation of suspected A. hydrophila coloniesfrom background organisms. Two sequential in situtests are performed as described below.
(i) Mannitol test. The in situ mannitol test wasincluded in the procedure to differentiate A. hydro-phila from nonpigmented Chromobacterium spp. andother mannitol-negative organisms. After primary in-cubation, the trehalose-positive (yellow) colonies arescored, and the filter is transferred to the in situmannitol medium. Incubation is continued for 2 to 3h at 37'C, and only those colonies which remain yellow(i.e., those that also ferment mannitol) are scored asecond time. The in situ mannitol medium containsthe following (in g/100 ml of deionized water): tryp-tose, 0.5; mannitol, 0.5; yeast extract, 0.2; NaCl, 0.3;KCI, 0.2; MgSO4.7H20, 0.02; FeCl2.6H20, 0.01; andbromothymol blue, 0.008. The pH is adjusted to 8.0with 10 N NaOH, 1.5 g of agar is added, and themedium is autoclaved at 121°C for 15 min. After themedium has cooled to 50°C, 100 mg of sodium deoxy-cholate is added, and the medium is dispensed intopetri dishes (50 by 12 mm; 5 il/plate). The plates ofmedium can be stored for 2 months at 4°C.
(ii) Oxidase test. This test differentiates A. hydro-phila from a number of other organisms, includingmembers of the Enterobacteriaceae. After the yellowcolonies from the in situ mannitol test have beenscored, the filters are transferred to a filter pad satu-rated with phosphate-buffered saline (13) for 60 s.This partially neutralizes organic acid end productswhich can obscure the color reaction of the oxidasetest. The filters are then transferred to pads saturatedwith N,N,N',N'-tetramethyl-p-phenylenediamine di-hydrochloride (oxidase reagent); they are read within10 to 15 s, and colonies with a purple halo are consid-ered positive. Typical trehalose-positive, mannitol-positive, oxidase-positive colonies are then counted asA. hydrophila. A flow diagram for the method ispresented in Fig. 1.
Test suspensions. Test suspensions of five envi-ronmental strains were used to assess the accuracy ofthe mA method. They were prepared in the followingmanner. Brain heart infusion (BHI) broth cultures,
VOL. 38, 1979

110 RIPPEY AND CABELLI
FILTRATION
(FILTER APPLIED TO mA, INCURATED 20 -24 HR, 37' C)
GREEN OR CLEAR COLONIES (Tr*-) YELLOW COLONIES (Tre.+IGNORE MARK
IN SITU MANNITOL TEST
(TRANSFER FILTER TO MANNITOL TEST PLATE:INCURATE COLONIES AT 37'C FOR 2 - 3 HOURS)
COLONIES WHICH TURN AND YELLOW COLONIES (Tr*+, Man+(REMAIN GREEN (Man-) MARK
IGNORE
IN SITU NEUTRALIZATION PBS, 60 SEC.)
IN SITU OXIDASE TEST
(TRANSFER FILTER TO PAD SATURATED WITH OXIDASE REAGENT,EXAMINE AFTER 10 - 15 SEC.)
YELLOW COLONIES (Ox-) PURPLE COLONIES IOx+, WEAK ACID)IGNORE IGNORE
YELLOW COLONIES WITH PURPLE HALOS (Tr.+, MAN+, Ox+l.PRESUMPTIVE A. HYDROPHILA
Tre-TREHALOSE; Mon -MANNITOL; Ox-OXIDASEA. SHIGELLOIDES IS RARELY ISOLATED ON mA. IF ITS PRESENCE IS SUSPECTED,
COLONIES CAN RE PICKED TO INOSITOL BROTH.A. SHIGELLOIDES IS INOSITOL POSITIVE.
FIG. 1. Flow diagram of the mA procedure for A.hydrophila. PBS, Phosphate-buffered saline.
incubated for 18 h at 370C, were diluted in lake watersterilized by autoclaving at 1210C for 15 min. Theresulting suspensions were held at 150C for 24 h, afterwhich samples were removed. Appropriate dilutions ofthe samples were either filtered through membranesfor incubation on mA agar or spread onto BHI agarplates. All plates were inverted, incubated at 370C for24 h, and counted.
Fecal samples. Fecal samples were collected from18 university students. A 1-g amount of feces wasdiluted in 100 ml of phosphate-buffered saline andappropriately diluted for membrane filtration. Theselectivity of the mA method against coliforms wasassessed by comparing the fermentative, oxidase-neg-ative background counts on mA medium (presumptivecoliforms) with the confirmed coliform counts as enu-merated by the mC procedure (6).Water samples. The specificity, selectivity, preci-
sion, counting range, and comparative recovery of themA method were determined by using surface watersamples collected from various geographic areas. Ingeneral, the interval between collection and assay ofthe samples did not exceed 12 h. Five replicate mem-branes on mA plates, three replicate membranes onRimler-Shotts (RS) (24) plates, and three replicateBHI agar spread plates were examined from the ap-propriate dilutions of each sample. The mA and RSplates were incubated at 370C for 20 h, after whichthey were examined as described above or as pre-scribed by the authors. The BHI agar plates wereincubated at room temperature for 48 h to estimatethe heterotrophic density of the sample.The specificity of the mA method for A. hydrophila
was determined from biochemical tests performed on20 presumptively positive and 10 presumptively neg-ative colonies from each of the 26 samples examined.
APPL. ENVIRON. MICROnIoL.
In most instances, the isolates were obtained from oneof the five replicate plates. In an effort to eliminatevisual bias, all of the colonies on a plate or a sector ofa plate were picked and streaked on nutrient agar forpurification. The cultures from the nutrient agar platesserved as the inocula for the biochemical tests. Isolateswere verified as A. hydrophila if they were (i) gramnegative, (ii) oxidase positive (4), (iii) insensitive toO/129 (17), and (iv) able to ferment mannitol and tre-halose but unable to ferment inositol (9, 20).The selectivity of the method was determined for
each of the samples examined by comparing the den-sities of the non-A. hydrophila colonies on the mAagar plates with the heterotrophic density obtained onthe BHI agar spread plates.The data on the precision of the method were
generated from the five replicate mA agar plates bythe D2 method of Eisenhart and Wilson (8).The counting range was determined as follows. In-
creasing quantities of natural water samples were as-sayed in quintuplicate. The mean A. hydrophila den-sity for each sample volume was determined. Theexpected values were obtained by extrapolating fromthe mean count nearest 30 colonies per filter by usingthe formula: Ce = C30 Ve/V30, where Ce is the expectedcount for a given sample volume (Ve) examined andC30 and V30 are the count nearest 30 colonies per filterand its corresponding volume, respectively.The recoveries of A. hydrophila on mA medium
were compared with those on the unmodified RSmedium used in conjunction with membrane filtration.This was done to increase the sample volume whichcould be examined and to permit the scoring of thetypical colonies as oxidase positive or negative withthe in situ oxidase test. The comparison was madebetween the verified recoveries by the mA procedureand the partially verified A. hydrophila estimates bythe RS method (i.e., typical, oxidase-positive, yellowcolonies). The in situ oxidase test was used with theRS method only after it was shown that the resultsobtained by the in situ method and by the method ofCowan and Steel (4) were comparable.
RESULTSThe recoveries of the five test strains of A.
hydrophila from sterile, fresh pond water sus-pensions of the organisms are presented in Table1. The values given are the average recoveriesobtained from three replicate mA filters or BHIspread plates. The mean recovery by the mAprocedure relative to that from the BHI spreadplates was no less than 85% for any of the strainstested, and the average recovery for the fivestrains was about 100%. These results, althoughhighly encouraging with regard to the accuracyof the mA procedure, were not definitive sincethey came from experiments which were basi-cally artificial in nature (i.e., stressed, pond wa-ter suspensions of pure cultures, rather thannaturally occurring populations of A. hydro-phila). Fortunately, we found several RhodeIsland ponds at which the A. hydrophila portion

VOL. 38, 1979
TABLE 1. Relative recovery ofA. hydrophila strainson nmA medium
Recovery (no. of cells/ml) on:
Straina BHI spread mA membrane % Recovery'
plate' filterbMR-16 8.2 x 10' 9.6 x 10' 116BC-7 1.4 X 102 1.5 X 102 107BL-4 8.0 X 104 6.9 x 104 86TC-14 7.9 X 103 7.3 X 103 92NC-5 3.5 X 104 3.9 x 104 111Avg 102
a Stressed at 15°C for 24 h in sterile, fresh, pondwater.
b Average of three replicate plates.'Recovery on mA medium/recovery on BHI agar
spread plates x 100.
of the recoverable bacterial population was solarge during the summer that the densities couldbe estimated by using the mA basal mediumwithout the inhibitors. This medium quantita-tively recovered A. hydrophila relative to BHIagar spread plates when fresh pond water sus-pensions of the organism were used (data notshown). Table 2 shows that, at four of five sitestested, the relative recoveries (mA comparedwith mA without inhibitors) of natural popula-tions of A. hydrophila exceeded 90% and thatthe average recovery at all five sites was 91%.The selectivity of the mA method when nat-
ural water samples were used was variable on asite-to-site basis. A 3-log reduction of the back-ground organisms was the target. The averagereduction was only 2.15 logs. When fecal sampleswere assayed, the reduction of coliforms wasapproximately 2.75 logs. This was observed froma comparison of the number of oxidase-negative,fermentative organisms on mA medium with thenumber on mC medium. A. hydrophila was notrecovered from any of the fecal samples.The specificity of the mA method was deter-
mined from the examination of more than 700isolates (Table 3). The overall confirmation ratefor colonies designated A. hydrophila was 98%,whereas 11% of the presumptively negative col-onies were, in fact, A. hydrophila.The precision of the mA method for each of
the sites was determined from D2 values forassay variability calculated from the followingequation: D2 = [NZXi2-(:Xi)2]/Xi, where :Xiwas the summation of the plate counts Xi,X2.... Xn and N (the number of replicate platesper sample) was five. These data (not shown)indicated that the values were randomly distrib-uted and that the data could therefore be sub-jected to further statistical analyses.The counting range was determined as de-
ENUMERATION OF A. HYDROPHILA 111
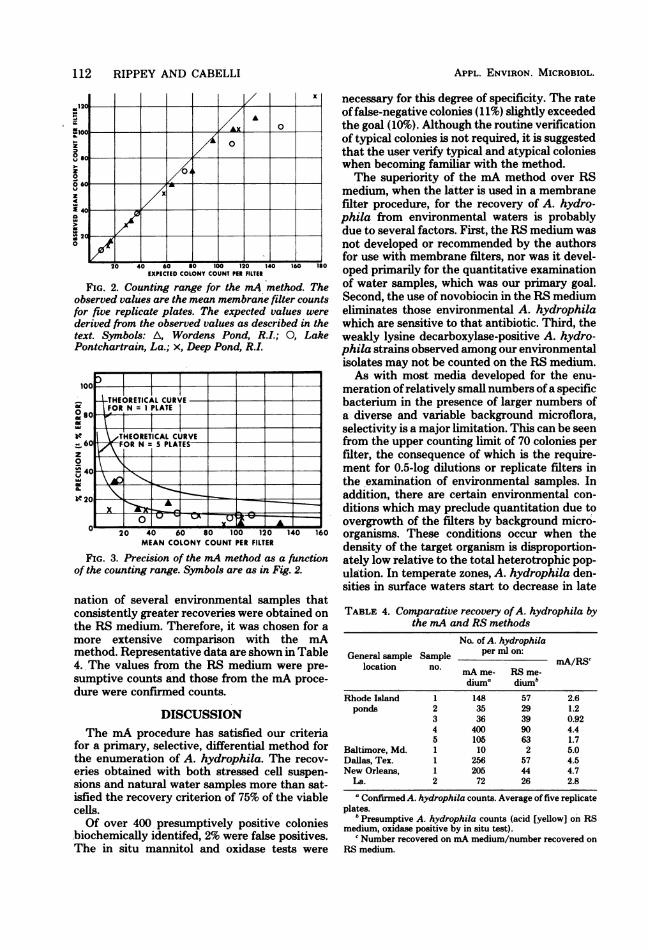
scribed above. The observed and expected val-ues for the recovery ofA. hydrophila from fresh-water samples are shown in Fig. 2. The upperlimit to the counting range was approximately70 A. hydrophila colonies per filter. A furtheranalysis of these data allowed the counting rangeof the mA method for natural water samples tobe evaluated as a function of its precision. Figure3, which shows the precision of the method,indicates that the variability within samples ap-proximates that which is expected from randomsampling error alone.
Preliminary to the full scale evaluation of mA,two other enumeration media, McCoy-Pilchermedium (15) and RS medium (24), were com-pared for their recovery of A. hydrophila onmembrane filters. Although the two latter mediawere not developed for use with membrane fil-tration techniques, they were so applied in thepresent study in order to maintain uniformityand allow for the examination of large volumesof water. It was demonstrated from the exami-
TABLE 2. Recovery of naturally occurring A.hydrophila populations on mA medium
Recovery (no. of cells/Rhode Island ml) on:a
sites ~~~~~~%Recovery'sites mA basal mA me-
mediumb dium'
Wordens Pond 467 460 99Tucker Pond 303 200 66Barber Pond 30 28 94100 Acre Pond 53 53 100Wood River 47 45 97Avg 91
a Includes fermentative, oxidase-positive organisms.b mA without ampicillin, sodium deoxycholate, and
ethanol.' Average of three replicate plates.d Recovery on mA medium/recovery on mA basal
medium x 100.
TABLE 3. Verification of colonies isolated by themA procedure
Typical coloniesb Atypical colonies'
Sample locationa No. ex- No. con- No: ex- No. con-firmed a firmedamined positive ammed negative
Rhode Island 293 290 187 160Baltimore, Md. 16 16 9 9Dallas, Tex. 38 38 26 26New Orleans, La. 59 57 28 25Georgia 30 30 20 20% Confirmed 98 89
a Includes all freshwater samples.bTypical A. hydrophila colonies are described in the text.cAll colonies other than those noted in footnote b.
112 RIPPEY AND CABELLI
,R12
0 1 0 20 4 0 _1|
lo
0
06
4
M
0/0
20 40 60Uso 100 120 140 160 Igo
EXPECTED COLONY COUNT PER FILTER
FIG. 2. Counting range for the mA method. Theobserved values are the mean membrane filter countsfor five replicate plates. The expected values werederived from the observed values as described in thetext. Symbols: A, Wordens Pond, R.I.; 0, LakePontchartrain, La.; x, Deep Pond, R.I.
Ir
be
z0
U,
ST
-z0 4U DU du IUU lBU I4U IOUMEAN COLONY COUNT PER FILTER
FIG. 3. Precision of the mA method as a functionof the counting range. Symbols are as in Fig. 2.
nation of several environmental samples thatconsistently greater recoveries were obtained onthe RS medium. Therefore, it was chosen for a
more extensive comparison with the mAmethod. Representative data are shown in Table4. The values from the RS medium were pre-
sumptive counts and those from the mA proce-
dure were confirmed counts.
DISCUSSIONThe mA procedure has satisfied our criteria
for a primary, selective, differential method forthe enumeration of A. hydrophila. The recov-
eries obtained with both stressed cell suspen-
sions and natural water samples more than sat-isfied the recovery criterion of 75% of the viablecells.Of over 400 presumptively positive colonies
.biochemically identifed, 2% were false positives.The in situ mannitol and oxidase tests were
APPL. ENVIRON. MICROBIOL.
necessary for this degree of specificity. The rateoffalse-negative colonies (11%) slightly exceededthe goal (10%). Although the routine verificationof typical colonies is not required, it is suggestedthat the user verify typical and atypical colonieswhen becoming familiar with the method.The superiority of the mA method over RS
medium, when the latter is used in a membranefilter procedure, for the recovery of A. hydro-phila from environmental waters is probablydue to several factors. First, the RS medium was
not developed or recommended by the authorsfor use with membrane filters, nor was it devel-oped primarily for the quantitative examinationof water samples, which was our primary goal.Second, the use of novobiocin in the RS mediumeliminates those environmental A. hydrophilawhich are sensitive to that antibiotic. Third, theweakly lysine decarboxylase-positive A. hydro-phila strains observed among our environmentalisolates may not be counted on the RS medium.As with most media developed for the enu-
meration of relatively small numbers of a specificbacterium in the presence of larger numbers ofa diverse and variable background microflora,selectivity is a major limitation. This can be seenfrom the upper counting limit of 70 colonies perfilter, the consequence of which is the require-ment for 0.5-log dilutions or replicate filters inthe examination of environmental samples. Inaddition, there are certain environmental con-
ditions which may preclude quantitation due toovergrowth of the filters by background micro-organisms. These conditions occur when thedensity of the target organism is disproportion-ately low relative to the total heterotrophic pop-
ulation. In temperate zones, A. hydrophila den-sities in surface waters start to decrease in late
TABLE 4. Comparative recovery ofA. hydrophila bythe mA and RS methods
No. of A. hydrophilaGeneral sample Sample per ml on: c
locationno.~~mA/RS~
mA me- RS me-diuma diumb
Rhode Island 1 148 57 2.6ponds 2 35 29 1.2
3 36 39 0.924 400 90 4.45 105 63 1.7
Baltimore, Md. 1 10 2 5.0Dallas, Tex. 1 256 57 4.5New Orleans, 1 205 44 4.7
La. 2 72 26 2.8
a Confirmed A. hydrophila counts. Average of five replicateplates.
'Presumptive A. hydrophila counts (acid [yellow] on RSmedium, oxidase positive by in situ test).
'Number recovered on mA medium/number recovered onRS medium.
THEORETICAL CURVE - _FOR N = 1 PLATE
so
THEORETICAL CURVE60 FOR N = PLATES
20

ENUMERATION OF A. HYDROPHILA 113
fall, are extremely low in the winter, and are notappreciable again until late spring (2). This maybe due to the organism's sensitivity to low tem-peratures. A similar situation occurs during theremainder of the year in highly oligotrophicbodies of water where A. hydrophila densitiesare very low relative to other heterotrophic mi-croflora. Both these situations require the ex-amination of large sample volumes, and thisincreases the probability of a decrease in theupper counting limit if not overgrowth by thebackground microflora. A third situation inwhich quantitation appears to be a problem is inwaters receiving carbohydrate-rich industrial ef-fluents, such as those of pulp mills and textilefinishing plants. At least two groups of orga-nisms, Klebsiella spp. (7) and Pseudomonasspp. (3) are present in very high densities in sucheffluents, and the mA method inhibitors, in theconcentrations used, are relatively ineffectiveagainst such high densities of these organisms.The selectivity of the mA medium against the
coliforms in feces was such that this mediumshould prove useful in the clinical examinationof human stools for A. hydrophila.
LITERATURE CITED
1. Annapurna, E., and S. C. Sanyal. 1977. Enterotoxicityof Aeromonas hydrophila. J. Med. Microbiol. 10:317-323.
2. Cabenli, V. J., F. T. Brezenaki, A. P. Dufour, and M.A. Levin. 1974. Microbiological methods for monitoringmarine waters for possible health effects. Proceedingsof seminar on methodology for monitoring the marineenvironment. U.S. Environmental Protection AgencyPublication EPA-600/4-74-004. U.S. EnvironmentalProtection Agency, Washington, D.C.
3. Cabelli, V. J., H. Kennedy, and M. A. Levin. 1976.Pseudomonas aeruginosa-fecal coliform relationshipsin estuarine and fresh recreational waters. J. WaterPoll. Control Fed. 48:367-376.
4. Cowan, S. T., and K. J. Steel. 1970. Cowan & Steel'smanual for the identification of medical bacteria. Cam-bridge University Press, Cambridge.
5. Davis, W. A., J. G. Kane, and V. F. Garagusi. 1978.Human Aeromonas infections: a review of the literatureand a case report of endocarditis. Medicine 57:267-277.
6. Dufour, A. P., and V. J. Cabelli. 1975. Membrane filterprocedure for enumerating the component genera of thecoliform group in seawater. Appl. Microbiol. 29:826-833.
7. Dufour, A. P., and V. J. Cabelli. 1976. Characteristicsof Klebsiella from textile finishing plant effluents. J.Water Poll. Control Fed. 48:872-879.
8. Eisenhart, C., and P. W. Wilson. 1943. Statistical meth-ods and control in bacteriology. Bacteriol. Rev. 7:57-137.
9. Ewing, W. H., and R. Hugh. 1974. Aeromonas, p. 230-238, 262, and 263. In E. H. Lennette, E. H. Spaulding,and J. P. Truant (ed.), Manual of clinical microbiology,2nd ed. American Society for Microbiology, Washing-ton, D.C.
10. Ewing, W. H., R. Hugh, and J. S. Johnson. 1961.Studies on the Aeromonas group. Center for DiseaseControl, Atlanta.
11. Hanson, P. G., J. Standridge, F. Jarrett, and D. G.Maki. 1977. Fresh water wound infection due to Aero-monas hydrophila. J. Am. Med. Assoc. 238:1053-1054.
12. Lautrop, H. 1961. Aeromonas hydrophila isolated fromhuman faeces and its possible pathological significance.Acta. Pathol. Microbiol. Scand. 144(Suppl.):299-301.
13. Levin, M. A., and V. J. Cabelli. 1977. Membrane filtertechnique for enumeration of Pseudomonas aerugi-nosa. Appl. Microbiol. 24:864-870.
14. Lungh, A., M. Popoff, and T. Wadstrom. 1977. Aero-monas hydrophila in acute diarrheal diseases: detectionof enterotoxin and biotyping of strains. J. Clin. Micro-biol. 6:96-100.
15. McCoy, R. H., and K. S. Pilcher. 1974. Peptone beefextract glycogen agar, a selective and differential Aero-monots medium. J. Fish. Res. Board Can. 31:1553-1555.
16. McCracken, A. W., and R. Barkley. 1972. Isolation ofAeromonas species from clinical sources. J. Clin. Pa-thol. 25:970-975.
17. Merkel, J. R. 1972. Influence of salts on the vibriostaticaction of 2,4-diamino-6,7-diisopropyl pteridine. Arch.Microbiol. 81:379-382.
18. Phillips, J. A., H. E. Bernhardt, and S. G. Rosenthal.1974. Aeromonas hydrophila infections. Pediatrics 53:110-112.
19. Popoff, M., and M. Veron. 1976. A taxonomic study ofthe Aeromonas hydrophila-Aeromonas punctatagroup. J. Gen. Microbiol. 94:11-22.
20. Schubert, R. H. W. 1974. Aeromonas, p. 345-348. In R.E. Buchanan and N. E. Gibbons (ed.), Bergey's manualof determinative bacteriology, 8th ed. The Williams &Wilkins Co., Baltimore.
21. Schubert, R. H. W. 1975. The relation of aerogenic toanaerogenic aeromonads of the "hydrophila-punctata-group" in river water depending on the load of waste.Zentralbl. Bakteriol. Parasitenkd. Infektionskr. Hyg.Abt. 1 Orig. Reihe B. 160:237-245.
22. Sedlak, J., and M. Slajsova. 1962. On the taxonomy ofthe genus Aeromonas. Prirodovedecka Faculta 438:508-510.
23. Shotts, E. B., Jr., J. L Gaines, L Martin, and A. K.Prestwood. 1972. Aeromonas-induced deaths amongfish and reptiles in an eutrophic inland lake. J. Am. Vet.Med. Assoc. 161:603-607.
24. Shotts, E. B., Jr., and R. Rimmler. 1973. Medium forthe isolation of Aeromonas hydrophila. Appl. Micro-biol. 26:550-553.
25. Trust, T. J., and R. A. H. Sparrow. 1974. The bacterialflora in the alimentary tract of fresh water salmonidfishes. Can. J. Microbiol. 20:1219-1228.
26. Von Graevenitz, A., and L Zinterhofer. 1970. Thedetection ofAeromonas hydrophila in stool specimens.Health Lab. Sci. 7:124-127.
27. Wadstrom, T., A. Ljungh, and B. Wretlind. 1976.Enterotoxin, haemolysin, and cytotoxic protein in Aero-monas hydrophila from human infections. Acta Pathol.Microbiol. Scand. Sect. B 84:112-114.
VOL. 38, 1979
Copyright © 2022 FDOKUMEN




















