
A Type III Secretion System Is Required for Aeromonas hydrophila AH1 Pathogenesis
10
10.1128/IAI.72.3.1248-1256.2004. 2004, 72(3):1248. DOI: Infect. Immun. Merino, J. M. Tomas and K. Y. Leung H. B. Yu, P. S. Srinivasa Rao, H. C. Lee, S. Vilches, S. AH-1 Pathogenesis Aeromonas hydrophila A Type III Secretion System Is Required for http://iai.asm.org/content/72/3/1248 Updated information and services can be found at: These include: REFERENCES http://iai.asm.org/content/72/3/1248#ref-list-1 at: This article cites 42 articles, 28 of which can be accessed free CONTENT ALERTS more» articles cite this article), Receive: RSS Feeds, eTOCs, free email alerts (when new http://journals.asm.org/site/misc/reprints.xhtml Information about commercial reprint orders: http://journals.asm.org/site/subscriptions/ To subscribe to to another ASM Journal go to: on March 21, 2014 by guest http://iai.asm.org/ Downloaded from on March 21, 2014 by guest http://iai.asm.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
6 -
download
0
Transcript of A Type III Secretion System Is Required for Aeromonas hydrophila AH1 Pathogenesis

10.1128/IAI.72.3.1248-1256.2004.
2004, 72(3):1248. DOI:Infect. Immun. Merino, J. M. Tomas and K. Y. LeungH. B. Yu, P. S. Srinivasa Rao, H. C. Lee, S. Vilches, S.
AH-1 PathogenesisAeromonas hydrophilaA Type III Secretion System Is Required for
http://iai.asm.org/content/72/3/1248Updated information and services can be found at:
These include:
REFERENCEShttp://iai.asm.org/content/72/3/1248#ref-list-1at:
This article cites 42 articles, 28 of which can be accessed free
CONTENT ALERTS more»articles cite this article),
Receive: RSS Feeds, eTOCs, free email alerts (when new
http://journals.asm.org/site/misc/reprints.xhtmlInformation about commercial reprint orders: http://journals.asm.org/site/subscriptions/To subscribe to to another ASM Journal go to:
on March 21, 2014 by guest
http://iai.asm.org/
Dow
nloaded from
on March 21, 2014 by guest
http://iai.asm.org/
Dow
nloaded from

INFECTION AND IMMUNITY, Mar. 2004, p. 1248–1256 Vol. 72, No. 30019-9567/04/$08.00�0 DOI: 10.1128/IAI.72.3.1248–1256.2004Copyright © 2004, American Society for Microbiology. All Rights Reserved.
A Type III Secretion System Is Required for Aeromonas hydrophilaAH-1 Pathogenesis
H. B. Yu,1 P. S. Srinivasa Rao,1 H. C. Lee,1 S. Vilches,2 S. Merino,2 J. M. Tomas,2and K. Y. Leung1*
Department of Biological Sciences, Faculty of Science, National University of Singapore, Singapore 117543,Singapore,1 and Department of Microbiology, University of Barcelona, Barcelona 08071, Spain2
Received 19 September 2003/Returned for modification 12 November 2003/Accepted 27 November 2003
Aeromonas hydrophila is a gram-negative opportunistic pathogen in fish and humans. Many bacterial patho-gens of animals and plants have been shown to inject anti-host virulence determinants into the hosts via a typeIII secretion system (TTSS). Degenerate primers based on lcrD family genes that are present in every knownTTSS allowed us to locate the TTSS gene cluster in A. hydrophila AH-1. A series of genome walking steps helpedin the identification of 25 open reading frames that encode proteins homologous to those in TTSSs in otherbacteria. PCR-based analysis showed the presence of lcrD homologs (ascV) in all of the 33 strains of A. hy-drophila isolated from various sources. Insertional inactivation of two of the TTSS genes (aopB and aopD) ledto decreased cytotoxicity in carp epithelial cells, increased phagocytosis, and reduced virulence in bluegourami. These results show that a TTSS is required for A. hydrophila pathogenesis. This is the first report ofsequencing and characterization of TTSS gene clusters from A. hydrophila. The TTSS identified here may helpin developing suitable vaccines as well as in further understanding of the pathogenesis of A. hydrophila.
Aeromonas hydrophila, a normal inhabitant of the aquaticenvironment, is an opportunistic pathogen of a variety ofaquatic and terrestrial animals, including humans (2, 41). A.hydrophila causes motile aeromonad septicemia, which is amajor freshwater disease affecting aquaculture worldwide. Inhumans, the clinical symptoms include septicemia, wound in-fections, and gastroenteritis (21). A. hydrophila has been iden-tified as an emerging cause of diarrhea, mostly in young chil-dren (1, 8). Janda and coworkers (22) also reported that A.hydrophila was responsible for 47% of septicemia cases indifferent regions in the United States. Therefore, the levels ofA. hydrophila in drinking water and in food have been high-lighted as a potential food safety issue (15).
The pathogenesis of A. hydrophila is multifactorial. Thesefactors include O antigens, capsules (25, 47), the S layer (10),exotoxins such as hemolysins and enterotoxin (6, 19), and arepertoire of exoenzymes which digests cellular componentssuch as proteases, amylases, and lipases (24, 27). These viru-lence determinants are involved sequentially in enabling thebacteria to colonize, gain entry, establish, replicate, and causedamage in host tissues and to evade the host defense systemand spread, eventually killing the host. The mechanisms ofaction of most of these virulence factors remain unknown.
Recent studies have shown that the virulence mechanisms ofvarious pathogens are highly similar (12). One of these mech-anisms is the type III secretion system (TTSS), which playscrucial roles in host-pathogen interactions (7). The TTSS isfound in many gram-negative animal and plant pathogens. Thissystem can efficiently deliver anti-host virulence determinantsinto the host cells, directly interfering with and altering hostprocesses. Recently, a fish pathogen, Aeromonas salmonicida
subsp. salmonicida (referred to here as A. salmonicida), has beenreported to have a functional TTSS located on a large thermo-labile virulence plasmid (5, 39). Since motile aeromonads andA. salmonicida are in the same genus, it is possible that TTSSs arealso present in other Aeromonas species, including A. hydrophila.
A. hydrophila AH-1 is pathogenic to several fish, such as therainbow trout (50% lethal dose of 104.5 [J. M. Tomas, personalcommunication]) and blue gourami (LD50 of 105.6 [this study]).It belongs to the O:11 serotype, which is one of the four dom-inant serogroups (O:11, O:16, O:18, and O:34) that are asso-ciated with gastroenteritis and septicemia in clinical studies(23). In the present study, we successfully located the TTSSgene cluster in A. hydrophila AH-1 by using homology-basedanalysis. This is the first report of the cloning and character-ization of a partial TTSS gene cluster in A. hydrophila AH-1.Further inactivation of two of the TTSS genes (aopB and aopD)resulted in a delayed cytotoxic effect on carp epithelial cells,increased phagocytosis, and reduced virulence in gourami fish,showing that a TTSS is required for A. hydrophila pathogenesis.
MATERIALS AND METHODSPlasmids, bacterial strains, and growth conditions. The bacterial strains and
plasmids used in this study are listed in Table 1. A. hydrophila strains weremaintained on tryptic soy agar or in tryptic soy broth (Difco) at 25°C. Escherichiacoli strains were maintained on Luria agar or in Luria broth (Difco) at 37°C.When required, media were supplemented with ampicillin (100 �g/ml), chlor-amphenicol (5 or 25 �g/ml), gentamicin (100 �g/ml), and colistin (6.25 �g/ml).The conjugal transfer of plasmids between A. hydrophila and E. coli strains wascarried out by plate mating at 30°C for 36 h.
DNA manipulations and Southern hybridization. Bacterial genomic DNA wasextracted as described in the manuals for the genomic DNA isolation and puri-fication kits from Promega. Plasmid DNA was extracted by using the QIAprepspin miniprep or Qiagen plasmid midi kit. Restriction endonuclease digestion wasaccomplished by standard methods (35). Southern blotting was performed withthe digoxigenin DNA labeling kit (Roche Diagnostics GmbH). Transfers of DNA tonylon membranes (GeneScreen; NEN Research Products), hybridization condi-tions, and visualization were as recommended by the manufacturer’s protocol.
PFGE. A. hydrophila AH-1 cells were embedded in low-melting-point agarose(Bio-Rad) to prepare agar plugs by using the contour-clamped homogeneous
* Corresponding author. Mailing address: Department of BiologicalSciences, Faculty of Science, National University of Singapore, ScienceDr. 4, Singapore 117543, Singapore. Phone: (65) 6874 7835. Fax: (65)6779 2486. E-mail: [email protected].
1248
on March 21, 2014 by guest
http://iai.asm.org/
Dow
nloaded from

electric field genomic DNA plug kit (Bio-Rad) according to the manufactur-er’s instructions. For PacI restriction digestion, agarose plugs were equili-brated with 500 �l of suitable restriction enzyme buffer for 30 min, exchangedwith 400 �l of fresh buffer containing 10 U of PacI restriction enzyme, andincubated overnight at 37°C. DNA fragments were separated by using aBio-Rad CHEF-DRII apparatus. Electrophoresis was carried out at 200 Vand 14°C for 22 h, with pulse times ranging from 5 to 60 s. For detecting anddetermining the sizes of large plasmids in A. hydrophila AH-1, plugs wereincubated with S1 nuclease and subjected to pulsed-field gel electrophoresis(PFGE) as described previously (3).
DNA sequencing and sequence analysis. DNA sequencing was carried out onan Applied Biosystems PRISM 3100 genetic analyzer with an ABIPRISM Big-Dye terminator cycle sequencing kit (Applied Biosystems). The sequences wereedited by using the manufacturer’s software. Sequence assembly and furtherediting were carried out with Vector NTI DNA analysis software (InforMax).BLASTN, BLASTP, and BLASTX sequence homology analyses and a proteinconserved-domain database analysis were performed by using the BLAST net-
work server of the National Center for Biotechnology Information. Terminatorprograms from the Genetics Computer Group package on a VAX4300 were usedto predict terminator sequences in the TTSS gene cluster.
Genome walking and general PCR analysis. Advantage polymerase 2 (Clon-tech) was used for genome walking and general PCR. PCRs were carried outunder the following conditions: one hold at 94°C for 2 min, followed by 32 cyclesof 94°C for 20 s, 60°C for 30 s, and 72°C for 1 min. Genome walker libraries wereconstructed by using five restriction enzymes (DraI, EcoRV, PvuII, ScaI, andStuI). The cycling parameters for genome walking were as follows: 7 cycles of 15 sat 94°C and 4 min at 72°C, followed by 32 cycles of 15 s at 94°C and 3 min at 67°C.The amplified fragments were cloned into the pGEM-T Easy vector (Promega).The recombinant DNA molecules were transformed into competent E. coliJM109 cells and sequenced.
Construction of defined insertion mutants. Mutants with defined insertions inaopB and aopD were constructed by using the protocol described earlier (34)with slight modifications. For better selection of insertion mutants of A. hy-drophila AH-1, we removed the kanamycin resistance cassette in pFS100 and
TABLE 1. Bacterial strains and plasmids used in this study
Strain or plasmid Genotype and/or relevant propertya Sourceb or reference
A. hydrophila strainsAer-19 O:11, virulent Human, CDHSAer-27 Virulent Human, CDHSAer-184 O:34, virulent Human, CDHSAer-186 O:16, virulent Human, CDHSAer-205 O:11, virulent Human, CDHSAer-211 O:18, virulent Human, CDHSATCC 7966 O:1, type strain Milk, ATCCAH-1 O:11, virulent, Colr Cms Fish, UMaopB mutant aopB insertion mutant from AH-1 This studyaopD mutant aopD insertion mutant from AH-1 This studyAH-3 O:34, virulent Fish, UBBa5 O:34, virulent Fish, UMJCM3968 O:6 Snake, JCMJCM3973 O:11 Unknown, JCMJCM3976 O:14 Frog, JCMJCM3978 O:16 Unknown, JCMJCM3980 O:18 Unknown, JCMJCM3981 O:19 Unknown, JCMJCM3983 O:21 Unknown, JCMJCM3984 O:22 Unknown, JCMJCM3985 O:23 Unknown, JCMJCM3996 O:34 Unknown, JCML15 O:51, avirulent Fish, BAUL31 O:91, virulent Fish, BAUL36 O:36, avirulent Fish, BAULL1 O:11, virulent Fish, UMPPD35/85 O:7, avirulent Fish, AVAPPD11/90 O:21, virulent Fish, AVAPPD64/90 O:34, avirulent Fish, AVAPPD88/90 O:16, avirulent Fish, AVAPPD45/91 Avirulent Fish, AVAPPD70/91 O:5, virulent Fish, AVAPPD122/91 O:11, virulent Fish, AVAPPD134/91 O:18, virulent Fish, AVAXs91/4/1 Virulent Fish, China
E. coli strainsJM109 Cols Amps PromegaMC1061(�pir) (�pir) thi thr-1 leu6 proA2 his-4 argE2 lacY1 galK2 ara14 xyl5 supE44, �pir 34S17-1(�pir) thi pro hsdR hsdM� recA[RP42-Tc::Mu-Km::Tn7 (Tpr Smr)Tra�] 37
PlasmidspGEM-T Easy vector Cloning vector; Apr PromegapFS100 pGP704 suicide plasmid, �pir dependent; Kmr 34pCM100 pGP704 suicide plasmid, �pir dependent; Cmr This studypCM-AOPB pCM with an internal fragment of aopB (605 bp) This studypCM-AOPD pCM with an internal fragment of aopD (565 bp) This study
a Virulent strains were defined as having a lower LD50 in blue gourami or rainbow trout (�106.5) than the avirulent strains (�107.5).b ATCC, American Type Culture Collection; AVA, Agri-Food and Veterinary Authority, Singapore; BAU, Bogor Agricultural University of Indonesia; CDHS,
California Department of Health Services; JCM, Japan Collection of Microorganisms; UB, University of Barcelona; UM, University of Montreal.
VOL. 72, 2004 TTSS IN A. HYDROPHILA 1249
on March 21, 2014 by guest
http://iai.asm.org/
Dow
nloaded from

replaced it with the chloramphenicol resistance (Cmr) cassette from pACYC184and generated the plasmid pCM100. Then, oligonucleotides aopB-F (5�-GTGGATATCTTGATCAATTGAGGAAGACGG-3�) and aopB-R (5�-GGAGATATCGGTACCAATATCAACTACCAG-3�) and oligonucleotides aopD-F (5�-GAAGATATCGATTCGAGCCTGCTGAGCAA-3�) and aopD-R (5�-GGAGATATCGCACTTCATCTTCCTTGGCATT-3�) were used to amplify internalfragments from the aopB and aopD genes, respectively. Amplified fragment wasligated to pGEM-T Easy vector (Promega) and transformed into E. coli JM109.The internal fragment was recovered by EcoRV restriction digestion (EcoRVsites in the primers are underlined) and finally ligated to EcoRV-digested anddephosphorylated pCM100 and transformed into E. coli MC1061(�pir), selectingCmr to generate plasmid pCM-AOPB and pCM-AOPD (Table 1). The recom-binant plasmid was isolated and transformed into E. coli S17-1(�pir). PlasmidspCM-AOPB and pCM-AOPD were transferred by conjugation to A. hydrophilaAH-1 (colistin resistant [Colr]) to obtain defined mutants by selecting Cmr andColr, and aopB and aopD insertion mutants, respectively, were generated. Theinsertion of plasmids on the chromosomes of these mutants was confirmed byboth PCR with appropriate primers and Southern blot analysis with pCM100suicide vectors, aopB, and aopD as probes.
Cell culture and morphological changes induced by A. hydrophila. All tissueculture reagents were obtained from Invitrogen. Epithelioma papillosum of carp(Cyprinus carpio) (EPC cells) (44) was grown in minimal essential medium withHanks salts, 10 mM HEPES (pH 7.3), 2 mM glutamine, 0.23% NaHCO3, and10% heat-inactivated fetal bovine serum (FBS) at 25°C in a 5% (vol/vol) CO2
atmosphere. Cells were grown in 75-cm2 flasks and split at least once a week bytrypsin-EDTA treatment and dilution at 1:10 in fresh medium. Studies on mor-phological changes were conducted by seeding 5 � 105 fish cells into each well ofa 24-well tissue culture plate (Falcon) and then proceeding as described previ-ously (40). EPC monolayers were infected with A. hydrophila cells suspended inphosphate-buffered saline (PBS) (137 mM NaCl, 2.7 mM KCl, 4.3 mMNa2HPO4, 1.4 mM KH2PO4 [pH 7.2]) at a multiplicity of infection (MOI)(number of bacteria per cultured cell) of 1. Changes in the cell morphology wereobserved over a period of 6 h. The fish cells were photographed under anAxiovert 25CFL phase-contrast inverted microscope (Carl Zeiss) at a magnifi-cation of �40 after 2.5 and 5.5 h of infection at 25°C.
Phagocyte isolation. Healthy blue gourami (Trichogaster trichopterus Pallas)were obtained from a commercial fish farm and maintained in well-aerated,dechlorinated water at 25 � 2°C. Phagocytes were isolated from the head kidneyof naive gourami and purified by the procedure of Secombs (36). Purified phago-
cytic cells (4 � 106 to 5 � 106 cells/well) were allowed to adhere to 24- or 48-welltissue culture plates (Falcon) in L-15 medium (Sigma) supplemented with 5%FBS. After 3 h of incubation at 25°C in a 5% CO2 atmosphere, the cells werewashed twice with Hanks balanced salt solution (HBSS) (Sigma) to remove un-attached cells. The remaining monolayer of phagocytes was infected with the wild-type and mutant strains of A. hydrophila at an MOI of 10 in all of the experiments.
Microscopic examination and phagocytosis assay. Glass coverslips wereplaced into each well of a 24-well tissue culture plate, and the wells were seededwith blue gourami phagocytes and incubated for 3 h at 25°C in a 5% CO2
atmosphere as described above. Three hours after infection, the phagocytes werewashed three times with HBSS and later stained with Giemsa stain (Merck) for30 min. After they were washed three times with PBS, the stained samples wereexamined under an Axiovert 25CFL phase-contrast inverted microscope (CarlZeiss) at a magnification of �100. For the phagocytosis assay, the phagocyteswere infected for 3 h, washed twice with HBSS, and then incubated for 1.5 h infresh L-15 medium supplemented with 5% FBS and 100 �g of gentamicin per ml.The gentamicin treatment killed extracellular bacteria but did not affect theviability of intracellular organisms. The phagocytes were then washed three timeswith HBSS to remove gentamicin and lysed with 1% (vol/vol) Triton X-100. Thiswas followed by a 1-min incubation, which released intracellular bacteria. Intra-cellular bacteria were quantified on triplicate tryptic soy agar plates. The per-centage of phagocytosis was calculated by dividing the viable bacterial count aftergentamicin treatment by the initial bacterial count. Three independent experi-ments with duplicate wells were performed.
Studies of LD50 in fish. Healthy blue gourami were obtained from a commer-cial fish farm, maintained in well-aerated dechlorinated water at 25 � 2°C, andacclimatized to the laboratory conditions for at least 15 days. The fish wereapproximately 13 g each and were about 3 months old. Three groups of ten fisheach were injected intramuscularly with 0.1 ml of PBS-washed bacterial cellsadjusted to the required concentrations. The fish were monitored for mortalityfor 7 days, and LD50s were calculated by the method of Reed and Muench (31).
Statistical analysis. All data were expressed as means � standard errors of themeans. Data were analyzed by a one-way analysis of variance and a Duncanmultiple-range test (SAS software; SAS Institute). P values of �0.05 were con-sidered significant.
Nucleotide sequence accession number. The nucleotide sequence data for theA. hydrophila AH-1 TTSS gene cluster have been deposited in GenBank underaccession no. AY394563.
TABLE 2. Putative A. hydrophila AH-1 proteins and their homologs in other bacteria
A. hydrophila proteinHomolog (I/S)a in:
Putative functionA. salmonicida P. luminescens Yersinia spp. P. aeruginosa
AscU LscU (75/84) YacU (69/81) PscU (68/81) Regulation of secretionAscT LscT (54/63) YscT (51/62) UnknownAscS LscS (82/90) YscS (77/89) PscS (78/90) UnknownAscR LscR (78/87) YscR (77/84) PscR (79/87) UnknownAscQ LscQ (47/58) YscQ (44/60) PscQ (47/59) UnknownAscP LscP (36/53) YscP (39/57) PscP (34/51) Regulation of secretionAscO LscO (40/47) YscO (37/53) PscO (42/53) Regulation of secretionAscN AscN (NDb) LscN (90/90) YscN (87/90) PscN (83/90) ATP synthaseAopN AopN (86/91) LopN (66/78) YopN (51/66) PopN (59/74) Regulation of translocationAcr1 Acr1 (90/94) LssA (71/86) TyeA (54/74) Pcr1 (57/75) Translocation apparatusAcr2 Acr2 (81/89) LssN (63/78) SycN (60/75) Pcr2 (46/62) ChaperoneAscX AscX (80/89) LssB (59/73) YscX (50/70) Pcr3 (46/61) Type III secretion apparatusAscY AscY (58/62) LssC (46/52) YscY (37/43) Pcr4 (46/54) Type III secretion apparatusAscV AscV (86/88) LssD (78/84) LcrD (76/82) PcrD (74/82) Type III secretion apparatusAcrR AcrR (60/70) LssR (51/64) LcrR (58/74) PcrR (47/61) UnknownAcrG AcrG (43/56) LssG (43/56) LcrG (47/63) PcrG (46/55) Regulation of low-calcium responseAcrV LssV (42/59) LcrV (41/63) PcrV (32/48) Protective antigen, anti-host factorAcrH LssH (58/69) LcrH (61/75) PcrH (61/73) ChaperoneAopB LopB (36/50) YopB (32/48) PopB (36/51) Translocation apparatusAopD LopD (49/66) YopD (45/61) PopD (47/64) Translocation apparatusHscY LscY (67/80) ExsC (56/80) UnknownAscW LscW (38/58) YscW (35/55) ExsB (33/52) Regulation of secretionAscA LscA (75/82) VirF (62/76) ExsA (64/77) Transcriptional activatorAscZ LscZ (41/54) ExsD (33/51) UnknownAscB LscB (54/67) YscB (45/64) PscB (44/60) Unknown
a I, percent identity; S, percent similarity.b ND, not determined due to incomplete sequence.
1250 YU ET AL. INFECT. IMMUN.
on March 21, 2014 by guest
http://iai.asm.org/
Dow
nloaded from

RESULTS AND DISCUSSIONSequencing and genetic organization of a TTSS gene cluster
in AH-1. TTSSs are present in many gram-negative bacteria(13, 20). Various components of the type III secretion genesare highly conserved. lcrD homologs are one of those compo-
nents that are present in all known TTSSs, and sequence sim-ilarities between individual members of the LcrD family varybetween 36 and 66% (20). Multiple-sequence alignment ofproteins of the LcrD family from A. salmonicida (5), Pseudo-monas aeruginosa (46), and Yersinia enterocolitica (accession
FIG. 1. Genetic organizations of TTSSs in A. hydrophila and other bacteria. Arrows indicate the proposed directions of transcription.
VOL. 72, 2004 TTSS IN A. HYDROPHILA 1251
on March 21, 2014 by guest
http://iai.asm.org/
Dow
nloaded from

no. AF102990) helped in locating two regions which are highlyconserved at both the amino acid and nucleotide levels. A pairof degenerate primers (ascV-F [5�-GTAARCAGATGAGTATCGATGG-3�] and ascV-R [5�-GAGACSCGGGTGACGATAAT-3�]) were designed, and the truncated lcrD-homologousgene (renamed ascV in this study; 331 bp in size) was success-fully amplified in A. hydrophila AH-1. The 331-bp fragmentshowed 92 and 99% identity to the ascV gene of A. salmonicidaat the nucleotide and amino acid levels, respectively. Subse-quently, the TTSS gene cluster was partially sequenced by aseries of genome walking steps. This led to the identification of25 open reading frames (ORFs) whose products showed highhomology with the TTSS proteins in A. salmonicida (5), Photo-rhabdus luminescens (42), P. aeruginosa (46), and Yersinia spe-cies (4, 26) (Table 2). Most of the putative TTSS proteins inA. hydrophila were designated according to the Yersinia no-menclature, which was also used to designate the ORFs of theA. salmonicida TTSS (Table 2). The genetic organization ofthis cluster is highly similar to that of P. aeruginosa (Fig. 1). Itis also similar to those of A. salmonicida, P. luminescens, andYersinia species but differs from the gene order in P. lumines-cens and Yersinia species, where the homologous ascU-aopDregions are inverted.
Further sequence analysis of this TTSS gene cluster identi-fied five putative promoter regions (Fig. 1), three of whichpossess ExsA binding sites (TxAAAAxA) that are similar tothose in A. salmonicida (5) and P. aeruginosa (18). In P. aerugi-nosa, these consensus sequences are bound by ExsA, a tran-scriptional activator of exoenzyme S regulon (18). These threepromoters are located upstream of aopN, acrG, and hscY. Wealso have identified AscA, which shows 77% similarity to ExsA(Table 2) and which may regulate the transcription of theseoperons in the AH-1 TTSS cluster. Further work is required toconfirm this hypothesis. The other two putative promoters are
upstream of ascN and ascZ, without the characteristic ExsAconsensus sequences being identified. Terminator analysisshowed the presence of three possible transcription termina-tors. They are downstream of ascU (G, 3.30 kcal/mol), aopD(G, 18.60 kcal/mol), and ascA (G, 6.40 kcal/mol) (Fig.1). Based on the analysis described above, we speculate thatthere may be five operons in the AH-1 TTSS gene cluster.They are ascN to ascU, aopN to aopD, acrG to aopD, hscY toascA, and ascZ and beyond.
The TTSS is located on the AH-1 chromosome. TTSS geneclusters can be either on a plasmid or on the bacterial chro-mosome (30). PFGE of bacterial DNA of A. hydrophila AH-1was carried out to investigate whether it contained any plas-mids. The PFGE results showed that the DNA cut with S1nuclease was brighter than the uncut DNA (Fig. 2A). Doddand Pemberton (9) also reported that the treatment of A.hydrophila JMP636 genomic DNA with S1 nuclease produceda highly intense band compared to untreated DNA, indicatingthe digestion of circular DNA by S1 nuclease. PFGE of an S1nuclease-treated A. hydrophila AH-1 genomic DNA plug showedonly one band as an uncut genomic DNA, indicating that theremay not be a large plasmid in AH-1. We further prepared agenomic DNA plug digested with PacI and carried out South-ern blot analysis with the ascV probe. The results revealed thatthe ascV probe hybridized with S1 nuclease-cut genomic DNA,uncut genomic DNA, and a 200-kb PacI-digested fragment(Figs. 2B and D). A 20-kb plasmid was detected by using aplasmid purification kit which was capable of purifying plas-mids of up to 150 kb. This small plasmid was not detected inPFGE, which could be due to the low copy number of thisplasmid. ascV probe did not hybridize with this 20-kb plasmid(data not shown). These results indicate that the TTSS genecluster is located on the chromosome of A. hydrophila AH-1,which is similar to the case for other pathogens such as P.aeruginosa (46) and P. luminescens (42). A. hydrophila AH-1 isunlike A. salmonicida, in which the TTSS is located on a largethermolabile plasmid of approximately 140 kb (39).
Distribution of TTSSs in A. hydrophila. In several gram-negative pathogens, TTSSs are encoded by pathogenicity is-lands, which are present in virulent strains but absent from
FIG. 2. Location of the ascV gene as determined by PFGE andSouthern blot analysis. (A) PFGE of S1 nuclease-treated (lane 1) andintact (lane 2) genomic plugs. (B) Southern blot analysis with ascV asa probe against the gel in panel A. (C and D) Gel showing a PacI-digested genomic DNA plug from AH-1 (C) and Southern blot withascV as a probe against the gel in panel C.
FIG. 3. aopB and aopD mutants show a decrease in the pathogen-esis of blue gourami infections. The number of fish alive after beinginjected with 106 CFU of A. hydrophila is plotted against the numberof days that it took for the fish to die. The fish were monitored for 7days. WT, wild-type AH-1.
1252 YU ET AL. INFECT. IMMUN.
on March 21, 2014 by guest
http://iai.asm.org/
Dow
nloaded from
avirulent strains (7, 43). Hence, the distribution of TTSS geneclusters among 33 A. hydrophila strains isolated from differentsources, such as fish and humans, from various geographicallocations was surveyed (Table 1). These isolates representeddiverse serotypes with various levels of virulence. The samepair of degenerate primers (ascV-F and ascV-R) was used todetect the ascV gene in all 33 strains. PCR results showed thata 331-bp DNA fragment was present in all of the A. hydrophilastrains. FlhA, an essential component of the flagellar exportapparatus, has also shown some similarity to the LcrD familyprotein in the TTSS (20). Hence, DNA sequencing was per-formed to confirm that these 33 PCR products were highlysimilar to ascV but not to flhA. All of the sequences showedhigh identity to ascV of A. hydrophila AH-1, with amino acidand nucleotide identities ranging from 83 to 100% and from 73to 99%, respectively. Our results indicate, therefore, that apartial or an entire TTSS gene cluster may be present in all 33A. hydrophila strains that we examined.
Similarly, the TTSS has also been shown to be ubiquitouslypresent in both clinical and environmental isolates of P. aerugi-nosa (11). Hence, the presence of TTSS genes may not corre-late with virulence for some of the bacteria. On the other hand,the TTSSs in nonpathogenic strains of A. hydrophila may not
be functional. Pierson and Falkow (29) have reported the pres-ence of nonfunctional invA-homologous genes in nonpatho-genic Y. enterocolitica.
Construction of mutants and LD50 studies. To ascertain thatthe TTSS in A. hydrophila AH-1 was functional, we carried outinsertional mutagenesis of aopB and aopD. They are the ho-mologs of yopB and yopD, respectively, in Y. enterocolitica,which may form the translocation apparatus for functional typeIII secretion (20). Disruptions in yopB and/or yopD reducedvirulence in mice (16, 17). In A. hydrophila AH-1, both aopBand aopD mutants showed growth rates in tryptic soy broththat were similar to that of the wild type (data not shown). TheLD50 of AH-1 by intramuscular injection into blue gouramiwas estimated to be 105.6, while those of the aopB and aopDmutants were 106.6 and 106.4, respectively, which was about 1log unit higher than that of the wild type. When blue gouramiwere inoculated with the same dosage (106), most of the fishinjected with aopB and aopD mutants recovered within 4 daysand displayed no marked skin ulceration at the injection site.However, 8 out of 10 fish injected with the wild type diedwithin 3 days (Fig. 3). To confirm the stability of the insertionalaopB and aopD mutant genes, bacteria were isolated from liveor dead fish inoculated with these mutants, all of which showed
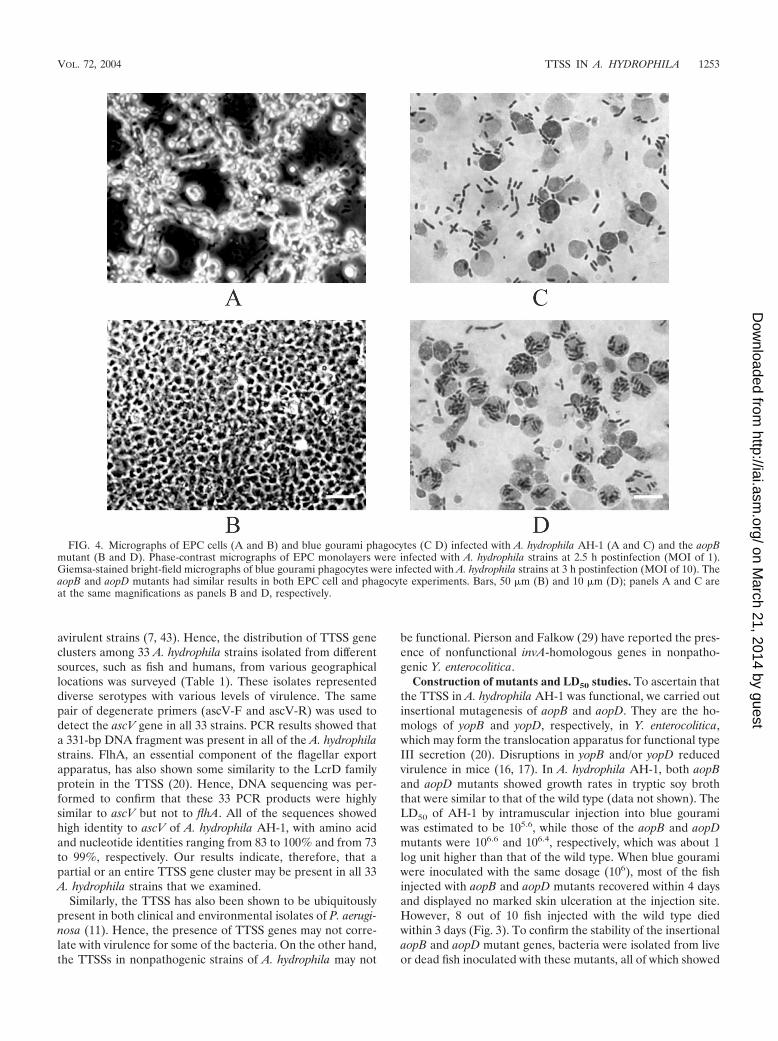
FIG. 4. Micrographs of EPC cells (A and B) and blue gourami phagocytes (C D) infected with A. hydrophila AH-1 (A and C) and the aopBmutant (B and D). Phase-contrast micrographs of EPC monolayers were infected with A. hydrophila strains at 2.5 h postinfection (MOI of 1).Giemsa-stained bright-field micrographs of blue gourami phagocytes were infected with A. hydrophila strains at 3 h postinfection (MOI of 10). TheaopB and aopD mutants had similar results in both EPC cell and phagocyte experiments. Bars, 50 �m (B) and 10 �m (D); panels A and C areat the same magnifications as panels B and D, respectively.
VOL. 72, 2004 TTSS IN A. HYDROPHILA 1253
on March 21, 2014 by guest
http://iai.asm.org/
Dow
nloaded from

resistance to chloramphenicol. PCR with appropriate primersfor these mutants also confirmed the stability of these mutants.These results strongly indicate that the TTSS plays an impor-tant role in the pathogenesis of A. hydrophila.
Delayed cytotoxic effect of aopB and aopD mutants on EPCcells. EPC cells were infected with AH-1 and the aopB andaopD mutants. Upon infection with A. hydrophila AH-1, theEPC cells underwent a series of cytopathic changes similar tothose of another strain, PPD134/91, as described in an earlierstudy (40). The cells infected with AH-1 progressively de-tached from one another, elongated to form long spindles,became rounded, and eventually detached from one anotherand also from the well. At 2.5 h postinfection, approximately50% of the rounded EPC cells remained attached to the tissueculture plate (Fig. 4A), while EPC cells infected with either theaopB or aopD mutant showed no significant morphologicalchanges (Fig. 4B) compared to the uninfected control (datanot shown). An uninfected monolayer of EPC cells appearedas a smooth sheet with the cells adhering tightly to their neigh-bors. EPC cells infected with the aopB or aopD mutant startedto show morphological changes at 3 h postinfection. By 5.5 hpostinfection, only 50% of the rounded EPC cells remained.Mutation of aopB or aopD therefore delayed cytotoxic changesto the EPC cells. There may be other factors mediating cyto-toxicity in A. hydrophila. Virulence factors such as aerolysinand hemolysin (45) and serine protease (32) have been re-ported to be involved in the cytotoxicity for different culturedcells. This may explain the delayed onset of the cytotoxic effectof aopB and aopD mutants of AH-1.
Phagocytosis assay. Phagocytes are the primary defense bar-riers in any host, and bacteria have to either avoid or overcomethe phagocyte-mediated killing to establish themselves in the
host. We carried out a microscopic examination of phagocytesinfected with AH-1 and the mutants. After 3 h of infection withAH-1, most of the bacteria were outside the phagocytes, witha few of them inside (Fig. 4C). On the other hand, most of theaopB and aopD mutants were inside the phagocytes (Fig. 4D).These results may indicate that the wild-type (AH-1) bacteriamay avoid or inhibit phagocytosis but that the mutants fail toavoid phagocytosis, thereby becoming ingested.
To analyze further whether the wild-type bacteria resistphagocytosis, an internalization assay was carried out. Afterinfection of the phagocytes with AH-1 and the aopB and aopDmutants for 3 h, the monolayer was treated with gentamicin for1.5 h to kill all of the extracellular bacteria. The results clearlyshowed that the mutants had a 4- to 6-times-higher ingestionrate than the wild type (Fig. 5). Wild-type AH-1 was resistantto phagocytosis, with only 0.24% of bacteria within the phago-cytes. However, the aopD and aopB mutants showed increasedphagocytosis, with 1.1 and 1.5% of bacteria within the phago-cytes, respectively. These results indicate that mutations inaopB and aopD affect the phagocytosis and may play an im-portant role in antiphagocytosis.
In Y. enterocolitica, YopB and YopD, the homologs of AopBand AopD, have also been shown to be essential for cytotox-icity and antiphagocytosis (16, 17). They may form the pore onthe host cell membrane to deliver effectors such as YopE andYopH, which cause cytotoxicity and antiphagocytosis, to therespective target host cells (14, 28, 33, 38). From the results ofcell culture assays, we speculate that AopB and AopD of AH-1may acting in a manner similar to those of Y. enterocolitica. TheTTSS of AH-1 may also use AopB and AopD as transloconcomponents to deliver similar effectors into host cells, mediat-ing antiphagocytosis and cytotoxicity. The natures of these
FIG. 5. Phagocytosis assay. The percent phagocytosis of gourami phagocytes was calculated after infection with wild-type A. hydrophila (AH-1)(WT) or mutants (aopB and aopD). Results are expressed as the representative mean � standard error of the mean from duplicate wells intriplicate experiments. Asterisks indicate a significant difference from results obtained with A. hydrophila AH-1 (P � 0.05).
1254 YU ET AL. INFECT. IMMUN.
on March 21, 2014 by guest
http://iai.asm.org/
Dow
nloaded from

effectors produced by AH-1 are being investigated. These re-sults will help in a better understanding of the roles played bythe TTSS in A. hydrophila pathogenesis.
Conclusions. This is the first report on sequencing and char-acterization of a TTSS gene cluster in A. hydrophila. The prod-ucts of most ORFs of this gene cluster showed high homologyto TTSS proteins of other pathogens, such as A. salmonicida, P.aeruginosa, and Yersinia species, indicating that they may haveevolved from a common ancestor. The detection of ascV byPCR and sequencing analysis with 33 A. hydrophila strainsrevealed that the TTSS may be present in all of the strains weexamined, irrespective of their pathogenic or nonpathogenicnature. Ongoing work is being conducted to see whether thefunctional TTSS is limited to disease but not environmentalisolates of A. hydrophila. Insertional inactivation of aopB oraopD delayed the cytotoxic effect in EPC cells and increaseduptake by phagocytes significantly. LD50 assays of mutants alsoshowed about 1-log-unit increase in LD compared to the wildtype. The above-mentioned biological activities clearly demon-strate that the TTSS is functional in A. hydrophila AH-1.Complementation of aopB and aopD mutants and creation ofmutants with mutations in other TTSS genes will help in un-derstanding their roles in cytotoxicity, antiphagocytosis, andvirulence in fish. The identification of a TTSS in A. hydrophilais an important discovery for unlocking the pathogenesis of thisbacterium. This will allow us to understand the intimate host-bacterium interactions in order to develop suitable strategiesto overcome diseases caused by A. hydrophila.
ACKNOWLEDGMENTS
This work was supported in part by the Biomedical Research Coun-cil of Singapore (BMRC), A*STAR, to K. Y. Leung and by the PlanNacional de I � D grants (Ministerio de Ciencia y Tecnología, Spain)and Generalitat de Catalunya to J. M. Tomas.
We are grateful to Michael Janda, California Department of HealthServices, for providing us some of the A. hydrophila isolates and toX. H. Wang and Y. L. Lau for their help with cell culture and PFGE.
REFERENCES
1. Agger, W. A., J. D. McCormick, and M. J. Gurwith. 1985. Clinical andmicrobiological features of Aeromonas hydrophila-associated diarrhea.J. Clin. Microbiol. 21:909–913.
2. Austin, B., and C. Adams. 1996. Fish pathogens, p. 197–243. In B. Austin, M.Altwegg, P. J. Gosling, and S. W. Joseph (ed.), The genus Aeromonas. JohnWiley and Sons, New York, N.Y.
3. Barton, B. M., G. P. Harding, and A. J. Zuccarelli. 1995. A general methodfor detecting and sizing large plasmids. Anal. Biochem. 226:235–240.
4. Bergman, T., K. Erickson, E. Galyov, C. Persson, and H. Wolf-Watz. 1994.The lcrB (yscN/U) gene cluster of Yersinia pseudotuberculosis is involved inYop secretion and shows high homology to the spa gene clusters of Shigellaflexneri and Salmonella typhimurium. J. Bacteriol. 176:2619–2626.
5. Burr, S. E., K. Stuber, T. Wahli, and J. Frey. 2002. Evidence for a type IIIsecretion system in Aeromonas salmonicida subsp. salmonicida. J. Bacteriol.184:5966–5970.
6. Chakraborty, T., M. A. Montenegro, S. C. Sanyal, R. Helmuth, E. Bulling,and K. N. Timmis. 1984. Cloning of enterotoxin gene from Aeromonashydrophila provides conclusive evidence of production of a cytotoxic entero-toxin. Infect. Immun. 46:435–441.
7. Cornelis, G. R., and F. Van Gijsegem. 2000. Assembly and function of typeIII secretory systems. Annu. Rev. Microbiol. 54:735–774.
8. Deodhar, L. P., K. Saraswathi, and A. Varudkar. 1991. Aeromonas speciesand their association with human diarrheal disease. J. Clin. Microbiol. 29:853–856.
9. Dodd, H. N., and J. M. Pemberton. 1998. Construction of a physical andpreliminary genetic map of Aeromonas hydrophila JMP636. Microbiology144:3087–3096.
10. Dooley, J. S. G., and T. J. Trust. 1988. Surface protein composition ofAeromonas hydrophila strains virulent for fish: identification of a surfacearray protein. J. Bacteriol. 170:499–506.
11. Feltman, H., G. Schulert, S. Khan, M. Jain, L. Peterson, and A. R. Hauser.2001. Prevalence of type III secretion genes in clinical and environmentalisolates of Pseudomonas aeruginosa. Microbiology 147:2659–2669.
12. Finlay, B. B., and S. Falkow. 1997. Common themes in microbial pathoge-nicity revisited. Microbiol. Mol. Biol. Rev. 61:136–169.
13. Galan, J. E., and A. Collmer. 1999. Type III secretion machines: bacterialdevices for protein delivery into host cells. Science 284:1322–1328.
14. Hakansson, S., K. Schesser, C. Persson, E. E. Galyov, R. Rosqvist, F.Homble, and H. Wolf-Watz. 1996. The YopB protein of Yersinia pseudotu-berculosis is essential for the translocation of Yop effector proteins across thetarget cell plasma membrane and displays a contact dependent membranedisrupting activity. EMBO J. 15:5812–5823.
15. Handfield, M., P. Simard, M. Couillard, and R. Letarte. 1996. Aeromonashydrophila isolated from food and drinking water: hemagglutination, hemo-lysis, and cytotoxicity for a human intestinal cell line (HT-29). Appl. Environ.Microbiol. 62:3459–3461.
16. Hartland, E. L., A. M. Bordun, and R. M. Robin-Browne. 1996. Contributionof YopB to virulence of Yersinia enterocolitica. Infect. Immun. 64:2308–2314.
17. Hartland, E. L., S. P. Green, W. A. Phillips, and R. M. Robins-Browne. 1994.Essential role of YopD in inhibition of the respiratory burst of macrophagesby Yersinia enterocolitica. Infect. Immun. 62:4445–4453.
18. Hovey, A. K., and D. W. Frank. 1995. Analyses of the DNA-binding andtranscriptional activation properties of ExsA, the transcriptional activator ofthe Pseudomonas aeruginosa exoenzyme S regulon. J. Bacteriol. 177:4427–4436.
19. Howard, S. P., S. Macintyre, and J. T. Buckley. 1996. The genus Aeromonas,p. 267–286. In B. Austin, M. Altwegg, P. J. Gosling, and S. Joseph (ed.),Toxin. John Wiley and Sons, Singapore.
20. Hueck, C. J. 1998. Type III protein secretion systems in bacterial pathogensof animals and plants. Microbiol. Mol. Biol. Rev. 62:379–433.
21. Janda, J. M. 2001. Aeromonas and Plesiomonas, p. 1237–1270. In M. Suss-man (ed.), Molecular medical microbiology. Academic Press, New York,N.Y.
22. Janda, J. M., L. S. Guthertz, R. P. Kokka, and T. Shimada. 1994. Aeromonasspecies in septicemia: laboratory characteristics and clinical observations.Clin. Infect. Dis. 19:77–83.
23. Kokka, R. P., J. M. Janda, L. S. Oshiro, M. Altwegg, T. Shimada, R.Sakazaki, and D. J. Brenner. 1991. Biochemical and genetic characterizationof autoagglutinating phenotypes of Aeromonas species associated with inva-sive and noninvasive disease. J. Infect. Dis. 163:890–894.
24. Leung, K. Y., and R. M. W. Stevenson. 1988. Tn5-induced protease-deficientstrains of Aeromonas hydrophila with reduced virulence for fish. Infect. Im-mun. 56:2639–2644.
25. Merino, S., X. Rubires, A. Aguillar, J. F. Guillot, and J. M. Tomas. 1996. Therole of the O-antigen lipopolysaccharide on the colonization in vivo of thegermfree chicken gut by Aeromonas hydrophila serogroup O:34. Microb.Pathog. 20:325–333.
26. Michiels, T., J. C. Vanooteghem, C. Lambert de Rouvroit, B. China, A.Gustin, P. Boudry, and G. R. Cornelis. 1991. Analysis of virC, an operoninvolved in the secretion of Yop proteins by Yersinia enterocolitica. J. Bac-teriol. 173:4994–5009.
27. Pemberton, J. M., S. P. Kidd, and R. Schmidt. 1997. Secreted enzymes ofAeromonas. FEMS Microbiol. Lett. 152:1–10.
28. Persson, C., R. Nordfelth, A. Holmstrom, S. Hakansson, R. Rosqvist, and H.Wolf-Watz. 1995. Cell-surface-bound Yersinia translocate the protein ty-rosine phosphatase YopH by a polarized mechanism into the target cell.Mol. Microbiol. 18:135–150.
29. Pierson, D. E., and S. Falkow. 1990. Nonpathogenic isolates of Yersiniaenterocolitica do not contain functional inv-homologous sequences. Infect.Immun. 58:1059–1064.
30. Plano, G. V., J. B. Day, and F. Ferracci. 2001. Type III export: new uses foran old pathway. Mol. Microbiol. 40:284–293.
31. Reed, L. J., and H. Muench. 1938. A simple method of estimating fiftypercent end points. Am. J. Hyg. 27:493–497.
32. Rodriguez, L. A., A. E. Ellis, and T. P. Nieto. 1992. Purification and charac-terization of an extracellular metalloprotease, serine protease and haemoly-sin of Aeromonas hydrophila strain B32: all are lethal for fish. Microb.Pathog. 13:17–24.
33. Rosqvist, R., K. E. Magnusson, and H. Wolf-Watz. 1994. Target cell contacttriggers expression and polarized transfer of Yersinia YopE cytotoxin intomammalian cells. EMBO J. 13:964–972.
34. Rubires, X., F. Saigi, N. Pique, N. Climent, S. Merino, S. Alberti, J. M.Tomas, and M. Regue. 1997. A gene (wbbL) from Serratia marcescensN28b(O4) complements the rfb-50 mutation of Escherichia coli K-12 deriv-atives. J. Bacteriol. 179:7581–7586.
35. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecular cloning: alaboratory manual, 2nd ed. Cold Spring Harbor Laboratory Press, ColdSpring Harbor, N.Y.
36. Secombs, C. J. 1990. Isolation of salmonid macrophages and analysis of theirkilling activity, p. 137–154. In J. S. Stolen, T. C. Fletcher, D. P. Anderson,B. S. Roberson, and W. B. Van Muiswinkel (ed.), Techniques in fish immu-nology. SOS Publications, Fair Haven, N.J.
VOL. 72, 2004 TTSS IN A. HYDROPHILA 1255
on March 21, 2014 by guest
http://iai.asm.org/
Dow
nloaded from

37. Simon, R., U. Priefer, and A. Puhler. 1983. A broad host range mobilizationsystem for in vivo genetic engineering: transposon mutagenesis in gramnegative bacteria. Bio/Technology 1:784–791.
38. Sory, M. P., A. Boland, I. Lambermont, and G. R. Cornelis. 1995. Identifi-cation of the YopE and YopH domains required for secretion and internal-ization into the cytosol of macrophages, using the cyaA gene fusionapproach.Proc. Natl. Acad. Sci. USA 92:11998–12002.
39. Stuber, K., S. E. Burr, M. Braun, T. Wahli, and J. Frey. 2003. Type IIIsecretion genes in Aeromonas salmonicida subsp. salmonicida are located ona large thermolabile virulence plasmid. J. Clin. Microbiol. 41:3854–3856.
40. Tan, E., K. W. Low, W. S. F. Wong, and K. Y. Leung. 1998. Internalizationof Aeromonas hydrophila by fish epithelial cells can be inhibited with atyrosine kinase inhibitor. Microbiology 144:299–307.
41. Thune, R. L., L. A. Stanley, and R. K. Cooper. 1993. Pathogenesis of gram-negative bacterial infections in warm water fish. Annu. Rev. Fish Dis. 3:37–68.
42. Waterfield, N. R., P. J. Daborn, and R. H. ffrench-Constant. 2002. Genomicislands in Photorhabdus. Trends Microbiol. 10:541–545.
43. Winstanley, C., and C. A. Hart. 2001. Type III secretion systems and patho-genicity islands. J. Med. Microbiol. 50:116–126.
44. Wolf, K., and J. A. Mann. 1980. Poikilotherm vertebrate cell lines andviruses: a current listing for fishes. In Vitro 16:168–179.
45. Wong, C. Y., M. W. Heuzenroeder, and R. L. Flower. 1998. Inactivation oftwo haemolytic toxin genes in Aeromonas hydrophila attenuates virulence ina suckling mouse model. Microbiology 144:291–298.
46. Yahr, T. L., L. M. Mende-Mueller, M. B. Friese, and D. W. Frank. 1997.Identification of type III secreted products of the Pseudomonas aeruginosaexoenzyme S regulon. J. Bacteriol. 179:7165–7168.
47. Zhang, Y. L., E. Arakawa, and K. Y. Leung. 2002. Novel Aeromonas hy-drophila PPD134/91 genes involved in O-antigen and capsule biosynthesis.Infect. Immun. 70:2326–2335.
Editor: B. B. Finlay
1256 YU ET AL. INFECT. IMMUN.
on March 21, 2014 by guest
http://iai.asm.org/
Dow
nloaded from




















