
Juvenile Administration of Methylphenidate Attenuates Adult Hippocampal Neurogenesis

Regulation of Neurogenesisby Interkinetic Nuclear Migrationthrough an Apical-Basal Notch GradientFilippo Del Bene,1 Ann M. Wehman,1 Brian A. Link,2,* and Herwig Baier1,*1Department of Physiology, Programs in Neuroscience, Genetics, and Developmental Biology, University of California,
San Francisco, 1550 4th Street, San Francisco, CA 94158-2722, USA2Department of Cell Biology, Neurobiology, and Anatomy, Medical College of Wisconsin, 8701 Watertown Plank Road, Milwaukee,
WI 53226, USA*Correspondence: [email protected] (B.A.L.), [email protected] (H.B.)
DOI 10.1016/j.cell.2008.07.017
SUMMARY
The different cell types in the central nervous systemdevelop from a common pool of progenitor cells. Thenuclei of progenitors move between the apical andbasal surfaces of the neuroepithelium in phase withtheir cell cycle, a process termed interkinetic nuclearmigration (INM). In the retina of zebrafish mikre oko(mok) mutants, in which the motor protein Dynac-tin-1 is disrupted, interkinetic nuclei migrate morerapidly and deeply to the basal side and more slowlyto the apical side. We found that Notch signaling ispredominantly activated on the apical side in bothmutants and wild-type. Mutant progenitors are,thus, less exposed to Notch and exit the cell cycleprematurely. This leads to an overproduction ofearly-born retinal ganglion cells (RGCs) at the ex-pense of later-born interneurons and glia. Our dataindicate that the function of INM is to balance theexposure of progenitor nuclei to neurogenic versusproliferative signals.
INTRODUCTION
The enormous neuronal diversity present in the vertebrate cen-
tral nervous system (CNS) arises from an apparently homoge-
neous progenitor population (Edlund and Jessell, 1999; Pearson
and Doe, 2004). The mechanisms that generate the correct num-
ber and relative proportion of each cell type are incompletely un-
derstood. Which factors influence, at any given time, how many
progenitors are undergoing a final neurogenic mitosis, producing
two neurons; how many are dividing asymmetrically, producing
one neuron and one progenitor; and how many are staying in
a proliferative state, producing two new progenitors?
Neurons arise during development from a pseudostratified co-
lumnar epithelium composed of mitotically active neuroepithelial
cells. These cells have an elongated shape, with cytoplasmic
connections to both the apical (ventricular) and basal surfaces.
Their nuclei occupy different levels within the epithelium depend-
ing on the phase of the cell cycle. Mitotic (M phase) nuclei are
located in close proximity to the apical surface, while nuclei un-
dergoing DNA synthesis (S phase) are displaced more basally.
This characteristic movement of nuclei within CNS neuroepithe-
lia was first predicted more than 70 years ago based on histolog-
ical observations (Sauer, 1935) and was termed interkinetic
nuclear migration (INM) (reviewed by Baye and Link, 2008). Re-
cently, the dynamics of INM have been shown to correlate with
neurogenic cell divisions within the retina, such that neuroepithe-
lia with more basal nuclear movements are biased to generate
postmitotic daughters (Baye and Link, 2007a). The precise
developmental function of INM, however, has remained elusive.
The retina consists of six neuronal and one glial cell types,
which differentiate in a stereotyped yet overlapping birth order
(Cepko et al., 1996; Marquardt and Gruss, 2002; Poggi et al.,
2005b). An extensive number of cell lineage studies have de-
scribed that retinal progenitors are multipotent, giving rise to all
of the major neuronal and glial cell types (Holt et al., 1988; Turner
and Cepko, 1987; Turner et al., 1990; Wetts and Fraser, 1988).
Together, these observations have led to the proposal that reti-
nal progenitors transit through a series of competence states
in a fixed order, and, during each one, they are intrinsically
able to generate one or a small subset of cell types (Livesey
and Cepko, 2001).
In Drosophila, Notch signaling, through lateral inhibition, main-
tains neighboring cells in a multipotent, proliferative state,
whereas downregulation of Notch is the prerequisite of neuronal
differentiation (Chitnis, 1995). This mechanism appears to be
largely conserved in the vertebrate retina (Jadhav et al., 2006;
Nelson et al., 2006; Perron and Harris, 2000). Thus, as the pro-
genitors produce the various cell types according to their com-
petence state, Notch ensures at each step that a subset of the
progenitors is retained for consecutive waves of neurogenesis.
A missing piece in the puzzle is the mechanism that controls
the dosage of Notch signaling such that a fraction of progenitors
exit the cell cycle and generate the appropriate number of each
cell type.
Here, we report a zebrafish mutant in which retinal progenitors
exit the cell cycle prematurely and retinal neurogenesis is accel-
erated. The progenitor population is quickly depleted and gives
rise to an imbalanced ratio of early cell fates versus late ones,
Cell 134, 1055–1065, September 19, 2008 ª2008 Elsevier Inc. 1055
reminiscent of Notch loss of function or a neurogenic gain of
function. To our initial surprise, we discovered that it is caused
by disruption of the motor protein Dynactin-1. We show that
this mutation leads to defects in specific aspects of INM without
affecting the intrinsic competence of the progenitors or their re-
sponsiveness to extrinsic cues. Furthermore, we demonstrate
the existence of a Notch gradient within the neuroepithelium,
with high levels in the apical domain. These results suggest
that INM regulates the duration and level of exposure of progen-
itor nuclei to neurogenic signals.
RESULTS
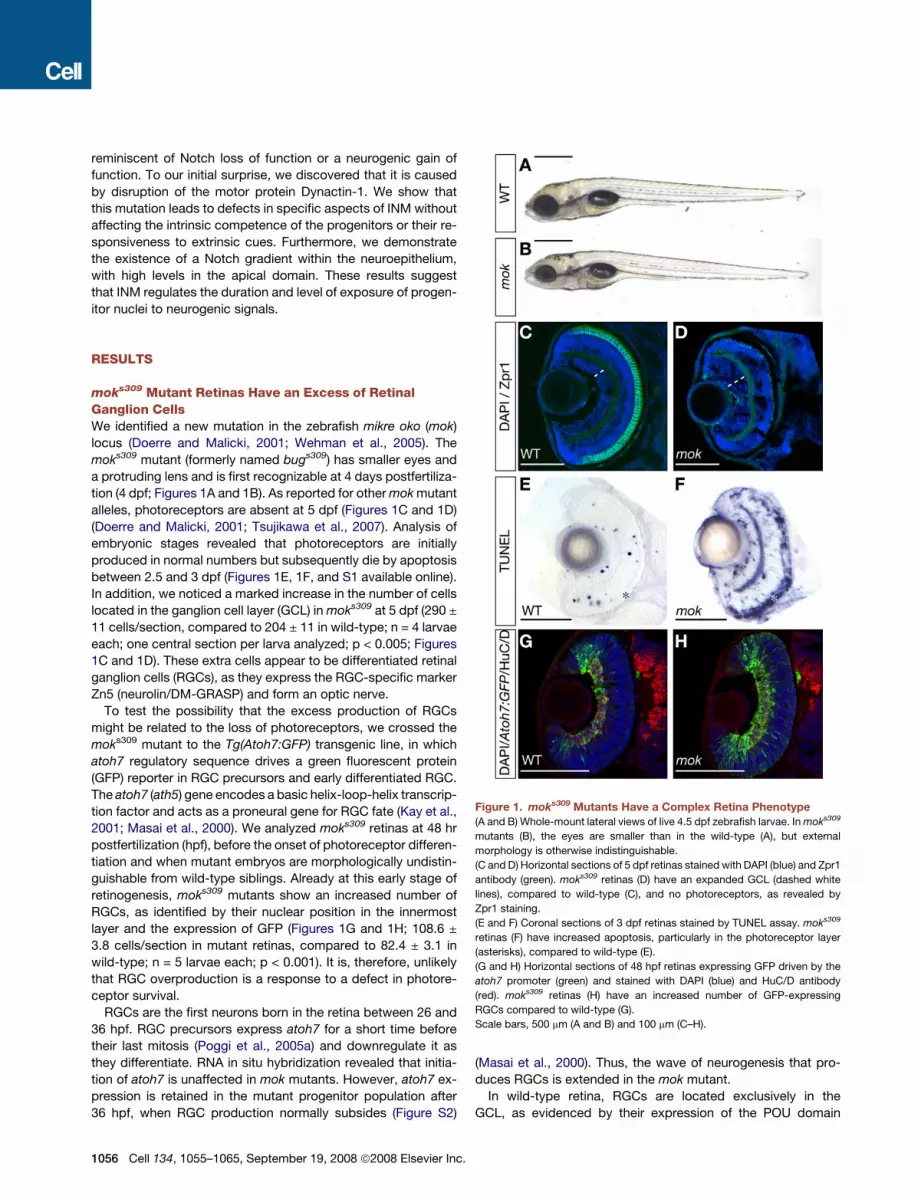
moks309 Mutant Retinas Have an Excess of RetinalGanglion CellsWe identified a new mutation in the zebrafish mikre oko (mok)
locus (Doerre and Malicki, 2001; Wehman et al., 2005). The
moks309 mutant (formerly named bugs309) has smaller eyes and
a protruding lens and is first recognizable at 4 days postfertiliza-
tion (4 dpf; Figures 1A and 1B). As reported for other mok mutant
alleles, photoreceptors are absent at 5 dpf (Figures 1C and 1D)
(Doerre and Malicki, 2001; Tsujikawa et al., 2007). Analysis of
embryonic stages revealed that photoreceptors are initially
produced in normal numbers but subsequently die by apoptosis
between 2.5 and 3 dpf (Figures 1E, 1F, and S1 available online).
In addition, we noticed a marked increase in the number of cells
located in the ganglion cell layer (GCL) in moks309 at 5 dpf (290 ±
11 cells/section, compared to 204 ± 11 in wild-type; n = 4 larvae
each; one central section per larva analyzed; p < 0.005; Figures
1C and 1D). These extra cells appear to be differentiated retinal
ganglion cells (RGCs), as they express the RGC-specific marker
Zn5 (neurolin/DM-GRASP) and form an optic nerve.
To test the possibility that the excess production of RGCs
might be related to the loss of photoreceptors, we crossed the
moks309 mutant to the Tg(Atoh7:GFP) transgenic line, in which
atoh7 regulatory sequence drives a green fluorescent protein
(GFP) reporter in RGC precursors and early differentiated RGC.
The atoh7 (ath5) gene encodes a basic helix-loop-helix transcrip-
tion factor and acts as a proneural gene for RGC fate (Kay et al.,
2001; Masai et al., 2000). We analyzed moks309 retinas at 48 hr
postfertilization (hpf), before the onset of photoreceptor differen-
tiation and when mutant embryos are morphologically undistin-
guishable from wild-type siblings. Already at this early stage of
retinogenesis, moks309 mutants show an increased number of
RGCs, as identified by their nuclear position in the innermost
layer and the expression of GFP (Figures 1G and 1H; 108.6 ±
3.8 cells/section in mutant retinas, compared to 82.4 ± 3.1 in
wild-type; n = 5 larvae each; p < 0.001). It is, therefore, unlikely
that RGC overproduction is a response to a defect in photore-
ceptor survival.
RGCs are the first neurons born in the retina between 26 and
36 hpf. RGC precursors express atoh7 for a short time before
their last mitosis (Poggi et al., 2005a) and downregulate it as
they differentiate. RNA in situ hybridization revealed that initia-
tion of atoh7 is unaffected in mok mutants. However, atoh7 ex-
pression is retained in the mutant progenitor population after
36 hpf, when RGC production normally subsides (Figure S2)
1056 Cell 134, 1055–1065, September 19, 2008 ª2008 Elsevier Inc
(Masai et al., 2000). Thus, the wave of neurogenesis that pro-
duces RGCs is extended in the mok mutant.
In wild-type retina, RGCs are located exclusively in the
GCL, as evidenced by their expression of the POU domain
Figure 1. moks309 Mutants Have a Complex Retina Phenotype
(A and B) Whole-mount lateral views of live 4.5 dpf zebrafish larvae. In moks309
mutants (B), the eyes are smaller than in the wild-type (A), but external
morphology is otherwise indistinguishable.
(C and D) Horizontal sections of 5 dpf retinas stained with DAPI (blue) and Zpr1
antibody (green). moks309 retinas (D) have an expanded GCL (dashed white
lines), compared to wild-type (C), and no photoreceptors, as revealed by
Zpr1 staining.
(E and F) Coronal sections of 3 dpf retinas stained by TUNEL assay. moks309
retinas (F) have increased apoptosis, particularly in the photoreceptor layer
(asterisks), compared to wild-type (E).
(G and H) Horizontal sections of 48 hpf retinas expressing GFP driven by the
atoh7 promoter (green) and stained with DAPI (blue) and HuC/D antibody
(red). moks309 retinas (H) have an increased number of GFP-expressing
RGCs compared to wild-type (G).
Scale bars, 500 mm (A and B) and 100 mm (C–H).
.

transcription factors Pou4f2 (Brn3b) and Pou4f3 (Brn3c) (Xiao
et al., 2005). In mok mutants, however, some of the extra
RGCs are found in the inner nuclear layer (INL) (Figures 2B, 2F,
and S3). In Tg(atoh7:GFP) fish, mature RGCs, as well as cells
that were competent earlier to produce RGCs (but followed a
different cell fate), remain fluorescently labeled for several days
after the atoh7 promoter is turned off, due to perdurance of
GFP (Masai et al., 2003). In wild-type retina, GFP-labeled cells
accumulate predominantly in the GCL. In moks309 mutants, by
contrast, cells in the enlarged GCL, as well as cells located out-
side the GCL (Figures 2A and 2E), are strongly GFP labeled. This
finding indicates that a greater number of progenitors in mok
mutants become competent to produce RGCs.
Bipolar and Muller Glia Cells Are Absent or SeverelyReduced in moks309 RetinasIn the retina, all neuronal cell types and Muller glia cells arise from
a common progenitor pool (Holt et al., 1988; Turner and Cepko,
1987; Wetts and Fraser, 1988). Therefore, we speculated that
overproduction of RGCs could lead to depletion of the progenitor
pool available for the genesis of later-born cells. To test this
hypothesis, we investigated the presence of markers for other
cell types. Muller glia are absent or strongly reduced in number
in mutant retinas at 5 dpf, as revealed by immunohistochemical
staining for glutamine synthetase (GS) (Figures 2C and 2G). The
Figure 2. moks309 Retinas Have Increased
RGCs and Decreased Bipolar and Muller
Glia Cells Due to Premature Neurogenesis
(A–H) Horizontal 5 dpf retina sections. GFP ex-
pression under the control of atoh7 (A and E) and
Brn3c (Pou4f3) (B and F) promoters reveals an in-
crease in RGCs in moks309 (E and F) as compared
to the wild-type (A and B), with some RGCs ectop-
ically located outside the GCL (yellow arrowheads
in [E] and [F]). The Muller glia marker GS (C and G)
and the bipolar cell marker PKCa (D and H) show
fewer immunoreactive cell bodies (yellow arrow-
heads) in moks309 (G and H) as compared to
wild-type (C and D).
(I–P) Horizontal 50 hpf retina sections stained for
IdU (green, injected at 26 hpf), BrdU (red, injected
at 38 hpf), and DAPI (blue). moks309 retinas (M–P)
show more IdU-positive and BrdU-negative cells
than wild-type, indicating that a larger number of
progenitors had exited the cell cycle between 26
and 38 hpf.
Scale bars, 100 mm.
same pattern was observed for bipolar
cells, as shown by decreased PKCa (Fig-
ures 2D and 2H). We confirmed this result
by in situ hybridization for vsx1, which
encodes a transcription factor marking
bipolar cells at early stages of differentia-
tion (Passini et al., 1997). vsx1 RNA is
decreased in moks309 mutant retinas (Fig-
ure S3). In contrast, no differences were
observed in numbers of horizontal or
amacrine cells, based on expression ofGAD65/67, parvalbumin, and an amacrine-specific transgenic
reporter (Kay et al., 2001) (Figure S4). As reported above, photo-
receptors are eliminated by apoptosis in the moks309 mutants,
but their specification and number seem unaffected at earlier
stages (Tsujikawa et al., 2007). Together, these results show
that the moks309 mutation causes a severe depletion of bipolar
cells and Muller glia.
A Greater Number of Progenitors Exit the CellCycle in mok MutantsFactors that delay cell-cycle exit in the retina give rise to an ex-
cess of later-born cell types, while prematurely forcing progeni-
tor cells to become postmitotic increases the generation of
RGCs (Ohnuma et al., 2002). Therefore, we suspected that the
cell-fate switch observed in moks309 would be accompanied by
abnormalities in the timing of cell-cycle exit. To confirm this,
we performed double-labeling experiments with two thymidine
analogs, IdU and BrdU. We allowed IdU and BrdU incorporation
for periods longer than a full cell cycle (12 hr) in order to label the
total population of proliferative cells. We injected IdU into the
developing embryos at 26 hpf, followed by BrdU at 38 hpf, and
finally fixed the embryos at 50 hpf. Cells that are positively
labeled for IdU, but not BrdU, had undergone the last S phase
between 26 and 38 hpf, when RGCs are generated in wild-
type. moks309 retinas show an �25% increase in IdU-positive
Cell 134, 1055–1065, September 19, 2008 ª2008 Elsevier Inc. 1057

and BrdU-negative cells compared to wild-type retinas (Figures
2I–2P; 33.5 ± 1.9 cells/section in mutant, compared to 25.2 ± 1.1
in wild-type; n > 8 larvae each; p < 0.005). This result was con-
firmed by similar double-labeling experiments using BrdU, in-
jected at 28 hpf, and the mitotically active cell marker PCNA (pro-
liferating cell nuclear antigen). Embryos were fixed at 40 hpf and
stained for PCNA (Figure S5). Taken together, our data demon-
strate that more cells leave the cell cycle during the first wave
of neurogenesis in moks309, thus biasing the neurons to adopt
the RGC fate.
To investigate the cell autonomy of the mutation, we analyzed
mutant/wild-type chimeras following blastomere transplanta-
tions at the 1000 cell stage. If mok acted nonautonomously,
then mutant clones in a wild-type environment should generate
retinal cell types in normal proportions. Conversely, if mok func-
tioned intrinsically in progenitor cells to promote neurogenesis,
then mutant clones should produce an excess of RGCs and
fewer INL cells regardless of their genetic environment. Clonal
analysis showed that the latter possibility is correct. While
wild-type clones in a wild-type retina give rise to neurons located
in the GCL, INL, and ONL in a 3:5:2 ratio, respectively, mutant
clones produce these cells in a 5:3:2 ratio (Figures 3A, 3B, and
3E; n = 1447 wild-type and 649 mutant cells counted in total).
We conclude that mok acts cell autonomously.
We further observed that wild-type clones in moks309 mutant
retinas tend to be excluded from the GCL and produce fewer
RGCs (2:6:2 ratio; n = 786 cells counted; Figures 3C and 3E)
than when they develop in a wild-type environment (2:6:2 ratio
for mutant cells in wild-type versus 3:5:2 for wild-type cells in
wild-type). Mutant clones in moks309 retinas show an increase
of RGCs (4:5:1 ratio; n = 223 cells counted; Figures 3D and 3E)
compared to wild-type clones in wild-type. These results sug-
gest that cell-fate switches of genotypically mutant progenitors
are even more dramatically biased toward the RGC fate when
they are in a wild-type environment than when they are sur-
rounded by other mutant cells (5:3:2 versus 4:5:1). This is prob-
ably due to intercellular feedback regulation within the develop-
ing retina. For instance, newborn RGCs limit the production of
additional RGCs by secreting the GDF11, which suppresses ex-
pression of atoh7 in uncommitted progenitors (Kim et al., 2005).
Consistent with this view, wild-type progenitors are more likely to
generate RGCs when transplanted into an environment in which
RGCs are absent, as in lakritz zebrafish mutants, which carry
a null mutation in atoh7 (Poggi et al., 2005a). Our transplantation
data suggest that such negative feedback attenuates the fate
switch in moks309 retinas.
The mok Gene Encodes Dynactin-1To gain insight into the molecular basis of the cell-fate regulation
defect, we positionally cloned the mok gene. The moks309 phe-
notype is perfectly linked to a nonsense point mutation in Dynac-
tin-1 (Dnct1, p150Glued), introducing a stop codon at amino acid
867 and a complete deletion of the C-terminal third of this pro-
tein. This region contains important protein-protein interaction
domains responsible for the binding of Dnct1 to other dynactin
subunits like Arp1 and p25 (Schroer, 2004). In Drosophila, the
glued allele that carries a similar truncation in the dnct1 ortholog
is not incorporated in the dynactin complex (McGrail et al., 1995).
1058 Cell 134, 1055–1065, September 19, 2008 ª2008 Elsevier Inc.
Western blot of whole-embryo extracts showed that Dnct1
protein is weakly detected in both mutants and wild-type imme-
diately after fertilization and for the first 3 days of development,
indicating that it is maternally supplied (Figure 4B). In moks309
mutants, Dnct1 protein is no longer detectable at 4 dpf (Fig-
ure 4A). Consistent with these data, the expression of dnct1
Figure 3. Cell Transplantation Analysis Reveals that mok Acts Cell
Autonomously
(A–D) Representative sections of 5 dpf chimeric retinas. The transplanted cells
are clearly identified by the expression of H2A-GFP marker (green) in their
nuclei. Cell transplantation shows that moks309 mutant clones in wild-type
host retinas (B) have a higher propensity to generate neurons located in the
GCL compared to control (A). Conversely, wild-type clones in moks309 host ret-
inas preferentially generate INL neurons (C). moks309 mutant clones in moks309
host retinas are shown for comparison (D). Dashed lines indicate the outer limit
of the GCL. Scale bars, 100 mm.
(E) Quantification of the transplantation results showing the distribution of
clones in the three retinal nuclear layers. ***p < 0.001; *p < 0.01. Error bar
indicates SEM.

RNA is ubiquitous and weak at early stages (data not shown) and
is enriched at 3 dpf in most of the larval CNS, including the GCL
and part of the INL (Figures 4C and 4D). To demonstrate that
dnct1 is the gene affected in the moks309 mutation, we designed
a splicing morpholino oligonucleotide (MO) to disrupt dnct1
Figure 4. Expression of Dnct1 and Its Function in INM
(A) Western blotting of extracts from 4 dpf embryos shows that Dnct1 is unde-
tectable in moks309.
(B) Time course analysis of Dnct1 expression by western blot in wild-type
zebrafish (numbers on top indicate hours after fertilization).
(C and D) Whole-mount in situ hybridization shows dnct1 enriched in the head
and eye region (C) and in the notochord (D). Scale bars, 100 mm.
(E and F) Coronal sections of 2 dpf retinas stained for the mitotic marker PH3.
In moks309 (F), a number of mitotic cells are sparsely located throughout the
retina, while, in the wild-type (E), they are confined within 2–3 cell diameters
from the ventricular surface. Dashed lines demarcate the apical (right) and
basal (left) domains. Asterisks indicate mitotic cells residing in the developing
lens.
(G) Scatter plot of wild-type (gray triangles) and moks309 mutant (black
squares) circles, showing the maximum basal distance of nuclei during INM
from 30–48 hpf.
(H) Histogram showing that these populations are statistically different
(p = 0.001, Wilcoxon two-sample test). Error bar indicates SEM.
C
function in wild-type embryos (Draper et al., 2001). MO-injected
embryos phenocopied moks309 mutants, including thickening of
the GCL and absence of differentiated photoreceptors (Fig-
ure S6). In conclusion, we predict that moks309 is a null or strong
hypomorph of dnct1.
INM Is Perturbed in moks309 Mutant EmbryosDynactin mediates the interaction of the dynein motor with many,
if not all, of its cargoes and allows the motor to traverse the mi-
crotubule lattice over long distances by increasing its processiv-
ity (King and Schroer, 2000). Loss-of-function analysis during
Drosophila eye development revealed that dynactin is required
for correct nuclear migration and maintenance of nuclear posi-
tion within postmitotic photoreceptors (Fan and Ready, 1997;
Whited et al., 2004). A similar nuclear positioning defect is ob-
served in zebrafish photoreceptor cells that carry a different
mok mutation (Tsujikawa et al., 2007). Therefore, we speculated
that, in moks309 mutants, the INM of neuroepithelial cells could
be altered and that this could, in turn, be responsible for the
effects on cell-cycle exit. To test this hypothesis, we analyzed
the positions of neuroepithelial nuclei during late G2/M phase us-
ing phosphorylated histone H3 (PH3) as a marker. PH3-positive
nuclei in wild-type are located close to the apical (ventricular)
surface of the neural retina at all time points analyzed (24, 36,
and 48 hpf; Figure 4E and data not shown; 47/47 cells in two ret-
inas at 48 hpf). In contrast, about 40% of PH3-positive nuclei are
positioned ectopically toward the basal side in moks309 mutants
(Figure 4F; 25/63 cells in three retinas at 48 hpf).
Time-lapse analysis of single-cell INM revealed a significant
increase in the maximal-basal position and a faster than normal
apical-to-basal movement of the nuclei in mutant cells (Movies
S1 and S2; Figures 4G and 4H; Table S1). Similar results were
obtained in dnct1-MO-injected embryos. The basal-to-apical
migration velocity was also reduced in the absence of dnct1 func-
tion, although the effect in this direction was less pronounced.
The net result is that interkinetic nuclei in mok mutants migrate
faster to the basal surface and further basally, take longer to re-
turn, and often enter mitosis before they have reached the apical
domain. In the wild-type retina, the depth of INM is an approxi-
mate predictor of a neurogenic cell division upon return to the
apical side (Baye and Link, 2007a). Therefore, we hypothesized
that this net change in INM promotes production of neurons.
An alternative hypothesis posits that cell polarity may be dis-
rupted in moks309 mutants. Indeed, previous work established
that intrinsic cell polarity is essential for the relationship between
nuclear position and neurogenesis (Baye and Link, 2007a). How-
ever, analysis of a series of apical and basal markers (laminin,
ZO-1, aPKCz) showed a correct localization in the mutant ret-
inas. We also investigated the distribution of a GFP-tagged
Par3 protein (Pard3), which partially overlaps with the adherens
junction-associated actin bundles (Wei et al., 2004). Apical local-
ization of Pard3-GFP was indistinguishable from wild-type
(Figure S7). These findings are consistent with a recent study
showing that polarity of photoreceptors is intact in mok mutants
(Tsujikawa et al., 2007). Cumulatively, these observations do not
support the interpretation that apical-basal polarity defects
underlie the displaced mitosis and neurogenic phenotypes in
mok retina.
ell 134, 1055–1065, September 19, 2008 ª2008 Elsevier Inc. 1059

A Notch Gradient Creates Distinct Signaling Milieusalong the Apical-Basal AxisNotch activation is known to delay neurogenesis (Furukawa
et al., 2000; Gaiano et al., 2000; Morrison et al., 2000; Scheer
et al., 2001). We observed that notch1a RNA is enriched in the
apical domain of the neuroepithelium in zebrafish (Figure 5A),
extending earlier reports in chick (Murciano et al., 2002). In addi-
tion, the Notch receptor ligands DeltaB and DeltaC are mostly
found in the basal half of the developing neuroretina (Figures
5B and 5C). Reflecting the localization of its transcript, DeltaC
protein is restricted to the basal half of the developing neurore-
tina both in wild-type and mutant embryos (Figures 5E and 5F).
Thus, Notch receptor and Delta ligands appear to be preferen-
tially expressed on opposite sides of the neuroepithelium.
We asked whether the Notch localization gradient leads to an
apical-to-basal gradient in Notch signaling. Upon binding of
ligand, Notch receptor undergoes a proteolytic cleavage. The cy-
tosolic fragment (Notch intracellular domain [NICD]) translocates
to the nucleus, where it modulates the transcription of target
genes. A specific anti-NICD antibody has been used in mouse
to detect the pattern of Notch activation in the developing brain
(Tokunaga et al., 2004). Using this antibody in mouse retina sec-
tions, we detected Notch activation only in nuclei located at the
apical surface, consistent with data in zebrafish (Figure 5J).
her4 is one target gene of the Notch/Delta pathway that is upre-
gulated by NICD and involved in neurogenesis (Takke et al.,
1999). Using the Tg(her4.1:dRFP) transgenic line, in which
a short-lived form of red fluorescent protein (dRFP) is expressed
under the control of the her4 promoter (Yeo et al., 2007), we mon-
itored her4 expression in vivo by time-lapse analysis (Movie S3;
Figures 5D and S8). In 13/15 cells moving from basal to apical,
dRFP fluorescence increased during the 2 hr observation period,
while the fluorescence decreased in 11/15 cells moving apical to
basal. This suggests that Notch signaling is activated as the
nuclei move into the apical compartment and downregulated in
nuclei that move in the opposite direction.
Dynactin-Driven INM Serves to Expose ProgenitorNuclei to the Notch GradientThe apical distribution of Notch, together with the dynactin-
driven, basally directed migration of interkinetic nuclei, offers
a mechanism by which the rate of neurogenesis could be regu-
lated. Consistent with our model, the microtubule minus ends,
marked by centrioles, are located at the apical surface (Figures
5G, 5H, and 5K), and microtubules are primarily oriented parallel
to the AB axis (Figure 5I). Using anti-Dnct1 antibodies, we ob-
served cytoplasmic punctate staining with enrichment at the
apical surface both in zebrafish wild-type and mouse retinal neu-
roepithelia, but not in moks309 mutants (Figures 5G, 5H, and 5K).
BBS4 protein localization is dependent on dynein motor function
and disrupted in moks309 mutants (Tsujikawa et al., 2007) (Fig-
ures 5L and S9). Thus, Dnct1 likely exerts its influence on neuro-
genesis by controlling INM within this graded signaling environ-
ment. Nuclei moving further basally appear to downregulate
Notch, while nuclei that remain closer to the ventricular surface
during INM retain high Notch activity levels.
An alternative way by which microtubule-associated motors
could affect Notch/Delta signaling in the neuroepithelial cells is
Figure 5. A Gradient of Notch Signaling
along the Apical-Basal Axis of the Develop-
ing Retina
(A–C) Coronal sections of 26 hpf retinas showing
mRNA expression levels of components of the
Notch/Delta signaling pathway. In situ hybridiza-
tion shows higher levels of notch1a close to the
apical surface of the retina (A) and deltaB and
deltaC close to the basal surface (B and C).
(D) Optical section of a 33 hpf retina expressing
her4:dRFP and H2A-GFP transgenes in a mosaic
manner.
(E and F) Coronal sections of 26 hpf retinas stained
with anti-DeltaC antibody, showing punctate cyto-
plasmic staining distributed in the basal half of the
tissue both in wild-type and moks309 retinas.
(G and H) Coronal sections of 26 hpf retinas
stained with anti-Dnct1 antibody, showing an en-
richment at the apical surface in wild-type retinas
(G), which is virtually absent in mutants (H).
(I) Coronal section of 26 hpf retina; a-tubulin stain-
ing reveals the parallel orientation of microtubules
to the apical-basal axis.
(J–L) Sections of mouse retina. Activated Notch1
antibody labels a subset of nuclei at the apical sur-
face (J). Anti-Dnct1 and anti-BBS4 antibodies
show a cytoplasmic, punctated staining enriched
at the apical surface. g-tubulin staining (G, H, K, L)
reveals the apical localization of the centrioles in
retinal progenitors. In (D)–(L), DAPI (blue) stains
the nuclei. In all panels, apical surface is on the left.
Scale bars, 25 mm (A–I) and 50 mm (J–L).
1060 Cell 134, 1055–1065, September 19, 2008 ª2008 Elsevier Inc.
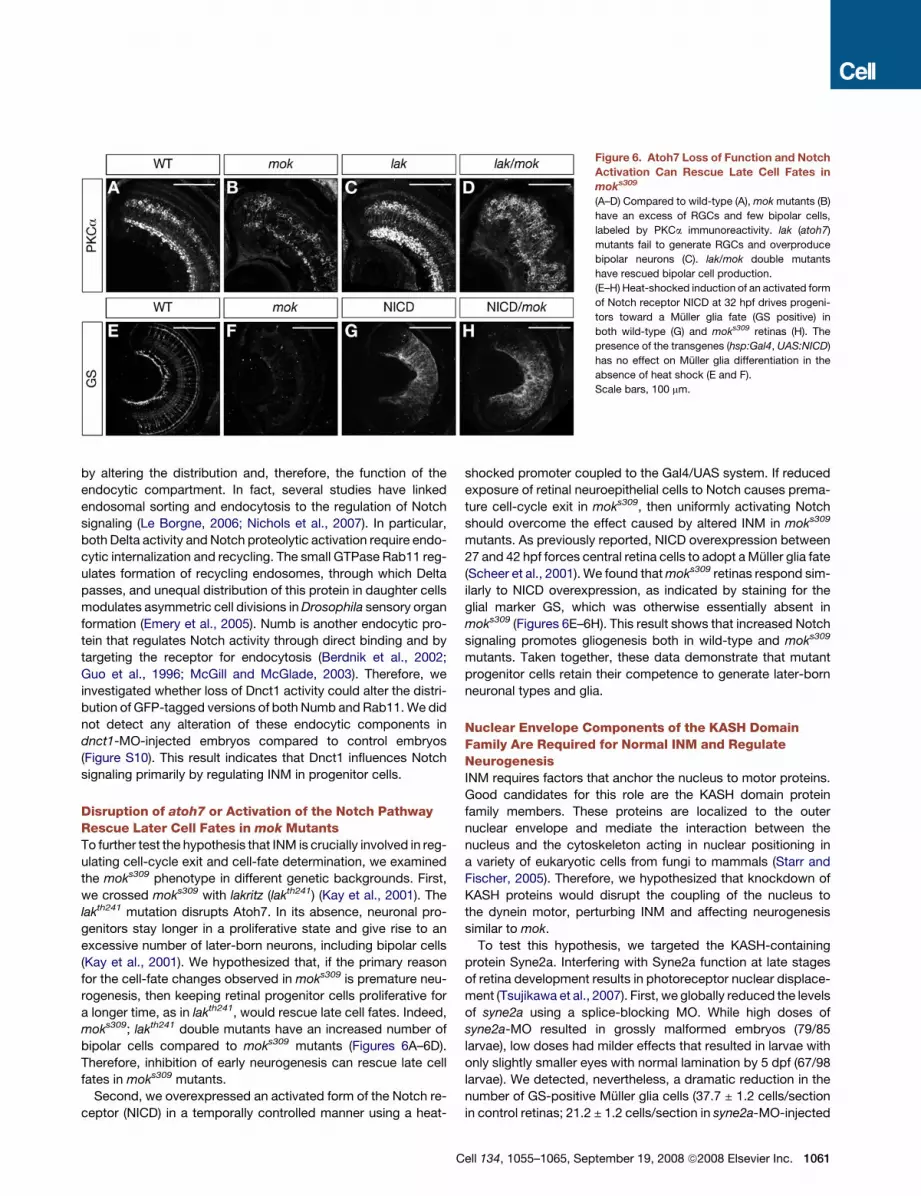
Figure 6. Atoh7 Loss of Function and Notch
Activation Can Rescue Late Cell Fates in
moks309
(A–D) Compared to wild-type (A), mok mutants (B)
have an excess of RGCs and few bipolar cells,
labeled by PKCa immunoreactivity. lak (atoh7)
mutants fail to generate RGCs and overproduce
bipolar neurons (C). lak/mok double mutants
have rescued bipolar cell production.
(E–H) Heat-shocked induction of an activated form
of Notch receptor NICD at 32 hpf drives progeni-
tors toward a Muller glia fate (GS positive) in
both wild-type (G) and moks309 retinas (H). The
presence of the transgenes (hsp:Gal4, UAS:NICD)
has no effect on Muller glia differentiation in the
absence of heat shock (E and F).
Scale bars, 100 mm.
by altering the distribution and, therefore, the function of the
endocytic compartment. In fact, several studies have linked
endosomal sorting and endocytosis to the regulation of Notch
signaling (Le Borgne, 2006; Nichols et al., 2007). In particular,
both Delta activity and Notch proteolytic activation require endo-
cytic internalization and recycling. The small GTPase Rab11 reg-
ulates formation of recycling endosomes, through which Delta
passes, and unequal distribution of this protein in daughter cells
modulates asymmetric cell divisions in Drosophila sensory organ
formation (Emery et al., 2005). Numb is another endocytic pro-
tein that regulates Notch activity through direct binding and by
targeting the receptor for endocytosis (Berdnik et al., 2002;
Guo et al., 1996; McGill and McGlade, 2003). Therefore, we
investigated whether loss of Dnct1 activity could alter the distri-
bution of GFP-tagged versions of both Numb and Rab11. We did
not detect any alteration of these endocytic components in
dnct1-MO-injected embryos compared to control embryos
(Figure S10). This result indicates that Dnct1 influences Notch
signaling primarily by regulating INM in progenitor cells.
Disruption of atoh7 or Activation of the Notch PathwayRescue Later Cell Fates in mok MutantsTo further test the hypothesis that INM is crucially involved in reg-
ulating cell-cycle exit and cell-fate determination, we examined
the moks309 phenotype in different genetic backgrounds. First,
we crossed moks309 with lakritz (lakth241) (Kay et al., 2001). The
lakth241 mutation disrupts Atoh7. In its absence, neuronal pro-
genitors stay longer in a proliferative state and give rise to an
excessive number of later-born neurons, including bipolar cells
(Kay et al., 2001). We hypothesized that, if the primary reason
for the cell-fate changes observed in moks309 is premature neu-
rogenesis, then keeping retinal progenitor cells proliferative for
a longer time, as in lakth241, would rescue late cell fates. Indeed,
moks309; lakth241 double mutants have an increased number of
bipolar cells compared to moks309 mutants (Figures 6A–6D).
Therefore, inhibition of early neurogenesis can rescue late cell
fates in moks309 mutants.
Second, we overexpressed an activated form of the Notch re-
ceptor (NICD) in a temporally controlled manner using a heat-
shocked promoter coupled to the Gal4/UAS system. If reduced
exposure of retinal neuroepithelial cells to Notch causes prema-
ture cell-cycle exit in moks309, then uniformly activating Notch
should overcome the effect caused by altered INM in moks309
mutants. As previously reported, NICD overexpression between
27 and 42 hpf forces central retina cells to adopt a Muller glia fate
(Scheer et al., 2001). We found that moks309 retinas respond sim-
ilarly to NICD overexpression, as indicated by staining for the
glial marker GS, which was otherwise essentially absent in
moks309 (Figures 6E–6H). This result shows that increased Notch
signaling promotes gliogenesis both in wild-type and moks309
mutants. Taken together, these data demonstrate that mutant
progenitor cells retain their competence to generate later-born
neuronal types and glia.
Nuclear Envelope Components of the KASH DomainFamily Are Required for Normal INM and RegulateNeurogenesisINM requires factors that anchor the nucleus to motor proteins.
Good candidates for this role are the KASH domain protein
family members. These proteins are localized to the outer
nuclear envelope and mediate the interaction between the
nucleus and the cytoskeleton acting in nuclear positioning in
a variety of eukaryotic cells from fungi to mammals (Starr and
Fischer, 2005). Therefore, we hypothesized that knockdown of
KASH proteins would disrupt the coupling of the nucleus to
the dynein motor, perturbing INM and affecting neurogenesis
similar to mok.
To test this hypothesis, we targeted the KASH-containing
protein Syne2a. Interfering with Syne2a function at late stages
of retina development results in photoreceptor nuclear displace-
ment (Tsujikawa et al., 2007). First, we globally reduced the levels
of syne2a using a splice-blocking MO. While high doses of
syne2a-MO resulted in grossly malformed embryos (79/85
larvae), low doses had milder effects that resulted in larvae with
only slightly smaller eyes with normal lamination by 5 dpf (67/98
larvae). We detected, nevertheless, a dramatic reduction in the
number of GS-positive Muller glia cells (37.7 ± 1.2 cells/section
in control retinas; 21.2 ± 1.2 cells/section in syne2a-MO-injected
Cell 134, 1055–1065, September 19, 2008 ª2008 Elsevier Inc. 1061

retinas; n = 4 in each group; p < 0.001; Figures 7A and 7B), similar
to the phenotype observed in moks309 retinas.
To separate the global effects of syne2a knockdown on embry-
onic development from specific effects in the developing retina,
we overexpressed a dominant-negative, GFP-tagged Syne2a
KASH domain in a mosaic fashion under control of a heat-
shocked promoter (Tsujikawa et al., 2007). Plasmid DNA was in-
jected at the 2–4 cell stage, and heat shock was applied at 23 hpf.
Figure 7. Disruption of the Nuclear Anchor to Dynactin Phenocopies
the moks309 Mutation
(A and B) Expression of the Muller glia marker GS is reduced in the retina of
larvae injected with a syne2a-MO (B) compared to control MO-injected larvae
(A). Scale bars, 100 mm.
(C and D) Representative examples of sections of 5 dpf retinas overexpressing
a control vector (C) or a dominant-negative Syne2a (KASH, [D]) under the con-
trol of a heat-shock promoter. The cells that express the constructs are iden-
tified by the expression of GFP marker (green). Clones of cells that express the
syne2a dominant-negative construct preferentially generate GCL neurons.
Scale bars, 50 mm.
(E) Quantification of the KASH overexpression results, showing the distribution
of clones in the three retinal nuclear layers. *p < 0.01, **p < 0.005. Error bar
indicates SEM.
(F) A model of the mechanism that couples INM with graded Notch activation.
1062 Cell 134, 1055–1065, September 19, 2008 ª2008 Elsevier Inc
KASH-expressing clones preferentially generated RGC neurons
at the expense of INL and ONL cells (Figures 7C–7E). These re-
sults are not the consequence of reduced INL or ONL cell survival,
because similar overexpression experiments after 48 hpf did not
cause immediate cell loss (Tsujikawa et al., 2007). These results
reinforce the notion that interfering with INM, either through the
motor protein complex or the nuclear anchor, perturbs cell-cycle
exit and neurogenesis.
DISCUSSION
We show here that a mutation in Dnct1, a microtubule-motor-as-
sociated protein, perturbs INM in a selective fashion and results
in an overproduction of early-born neurons in the zebrafish ret-
ina. The progenitor nuclei move more quickly and deeply in the
basal direction and more slowly apically. We further established
that antineurogenic Notch signals are enriched on the apical side
of the neuroepithelium in both mutants and wild-type. Combined
with previous observations that progenitors whose nuclei mi-
grate deep are more likely to produce postmitotic neuronal
daughters following their return to the apical side (Baye and
Link, 2007a), our studies suggest a mechanism by which INM
cooperates with an apical-basal Notch gradient to select pro-
genitors for cell-cycle exit and apportion cell fates (Figure 7F).
The phenotype of the mok mutant is reminiscent of manipula-
tions of cell-fate determinants, such as overexpression of bHLH
proneural genes. Therefore, we asked whether the mok mutation
altered the intrinsic competence of progenitors. The bHLH tran-
scription factor Atoh7 functions as a proneural factor to set neu-
rogenic competency of early retinal cell types and is essential for
RGC fate. Disruption of atoh7 eliminates RGCs and increases the
number of progenitors that remain in the cell cycle (Brown et al.,
2001; Kay et al., 2001; Wang et al., 2001). As a consequence of
this enlarged progenitor pool, bipolar cells and glia are increased
in number in the zebrafish lak (atoh7) mutant. Conversely, over-
expression of atoh7 results in an excess of RGCs, resembling
the mok phenotype (Hatakeyama and Kageyama, 2004; Vetter
and Brown, 2001; Yan et al., 2005). In lak; mok double mutants,
glia and bipolar neurons, which are severely reduced in mok
single mutants, develop similarly to lak single mutants, demon-
strating that mok mutant progenitors retain their potential to
generate later-born neurons. Photoreceptors, which develop
normally in lak single mutants, however, are absent in the double
mutants, as they are in mok single mutants, due to a related but
independent function of Dnct1 in photoreceptor nuclear posi-
tioning (Tsujikawa et al., 2007). In conclusion, Dnct1 does not
affect the neurogenic competence of retinal progenitor cells.
Inhibition of Notch (Austin et al., 1995) or disruption of GDF11
(Kim et al., 2005) mimic some aspects of the mok phenotype.
Notch prevents progenitors from leaving the cell cycle and differ-
entiating prematurely. Its activation during the time progenitors
that are competent to produce RGCs leads to a depletion of
this cell type. GDF11, on the other hand, is secreted by differen-
tiated RGCs to negatively regulate the number of new RGCs
produced by suppressing atoh7. As a consequence, progenitors
are more likely to generate RGCs when they are transplanted into
an environment in which RGCs are absent, as in lak mutants
(Poggi et al., 2005a). We made a complementary observation
.

by transplanting wild-type cells into mok mutants. These clones
tend to form fewer RGCs, suggesting that the negative feedback
signals are intact in the mutant environment. Moreover, we show
that constitutive activation of Notch in mok mutants blocks RGC
production and leads to an overproduction of glia, as it does in
wild-type. Together, these results demonstrate that mok mutant
cells are still able to produce and respond to extrinsic regulators
of cell fate.
A Notch gradient along the apical-basal axis of the neuroepi-
thelium is likely to play a key role in neurogenesis. Notch
mRNA is increased on the apical side, whereas Delta mRNA
and protein are enriched basally. This results in a gradient of
Notch transcriptional activity, as demonstrated by the higher
concentration of Notch ICD in the apical domains and the in-
creased her4 expression in cells whose nuclei move into the
high-Notch environment. We have found no evidence that Notch
signaling, neuroepithelial polarity, or the Notch gradient itself are
altered in mok mutants. Distribution of the endocytic pathway
components Rab11 and Numb, which, in other contexts, regu-
late Notch, is unaltered. Moreover, an independent manipulation
of the dynein/dynactin-dependent component of INM (disruption
of the nuclear anchor protein Syne2a) perturbs cell-fate deci-
sions similarly to the mok mutation.
Between mitoses, a progenitor nucleus moves twice through
a Notch spatial gradient. If the nucleus stays close to the apical
side, it will encounter high Notch levels throughout the cell cycle,
and both of its daughters are likely to remain proliferative. On the
other hand, if the nucleus is translocated more basally, Notch
activity is reduced, predisposing the progenitor to produce one
or two daughter neurons during its subsequent mitosis. It follows
that, in mok mutants, the balance of neurogenic versus prolifer-
ative divisions is shifted by decreasing exposure to Notch across
the progenitor population. Given that both INM and Notch signal-
ing compartments are ubiquitous features of CNS neuroepithelia
(Frade, 2002) as well as nonneuronal epithelia (Bort et al., 2006),
it seems likely that this mechanism is widely employed during
embryonic development, growth, and regeneration.
EXPERIMENTAL PROCEDURES
Immunohistochemistry and In Situ Hybridization
Retinal sections were stained using standard protocols (Kay et al., 2001). E12
CD1 mice embryos (gift from Florence Lee) were fixed in 4% PFA and pro-
cessed for cryosectioning according to standard protocols. The full list of pri-
mary and secondary antibodies is given in the Supplemental Data. Staining for
g-tubulin required the treatment of the samples for 5 min in acetone at �20�C
prior to blocking. Fixation and staining for a-tubulin were performed as de-
scribed (Wehman et al., 2007). Apoptosis was detected by whole-mount
TUNEL assay using the ApoTag kit (Chemicon). Whole-mount in situ hybridiza-
tion was performed according to standard protocols, and a list of antisense
probes is reported in the Supplemetal Data.
BrdU/IdU Incorporation
Cell proliferation was assayed by BrdU/IdU incorporation as previously de-
scribed (Burns and Kuan, 2005; Kay et al., 2001) with the following modifica-
tion. Embryos were injected with 10 mM BrdU or IdU solutions into the yolk
and grown until fixation.
Cell Transplantation Analysis
Chimeric embryos were generated using standard methods (Ho and Kane,
1990). Donor embryos were Tg(h2afv:GFP)kca6 to easily identify transplanted
clones. Both donor embryos and chimeras were allowed to develop until 5
dpf to identify mutants. Chimeric larvae were then fixed and processed for im-
munohistochemistry.
DNA and MO Injections
All injections were performed at the 1 cell stage unless specified otherwise.
dnct1-MO were injected at a concentration of 20 mM. Sequence: 50-ctgagg
gacggccggtctgtggagg. syne2a-MO were injected at 500 mM (high dose) or
50 mM (low dose). The sequence has been described previously (Tsujikawa
et al., 2007). Expression of Syne2a-KASH domain Pard3-GFP, EGFP-Numb,
and EGFP-Rab11 was carried out as described (Geldmacher-Voss et al.,
2003; Muto et al., 2006; Reugels et al., 2006; Tsujikawa et al., 2007). The
pard3-EGFP construct was injected at 2–4 cell stages to achieve sparse ex-
pression. EGFP-Numb and EGFP-rab11 were subcloned in the Tol2-based
vector and injected with transposase RNA to achieve uniform expression
(Kawakami, 2004).
Nuclear Labeling to Record INM
To label nuclei of retinal progenitor cells, plasmid DNA encoding the histone
H2B-GFP fusion protein was microinjected into 1–4 cell-stage embryos to
label nuclei in a mosaic fashion throughout the embryo (Koster and Fraser,
2001; Meng et al., 1999). Embryos were derived from incrosses of heterozy-
gous moks309 pairs. All embryos were grown in 0.003% 1-phenyl-2-thiourea
(PTU) to block pigmentation. At 22 hpf, small pieces of tail tissue were used
to PCR genotype either wild-type or mutant embryos prior to imaging. Follow-
ing imaging, all embryos were allowed to develop and were scored for their
phenotypes in order to validate genotyping results.
Confocal Time-Lapse Microscopy
At 26 hpf, labeled embryos were anesthetized with 0.05% Tricane in 0.003%
PTU and embedded in 1.0% low-temperature-melting agarose. Embryos
were placed in a glass bottom culture dish and oriented so that the eye was
facing up. GFP-labeled nuclei and dRFP-labeled cells were imaged on a Nikon
C1 confocal microscope. Transmitted light images were also collected during
the time-lapse to enable accurate measurement of apical and basal surfaces
during nuclear movements. Optical z sections were collected at 2 mm steps
every 12 min for 30–48 hr. These parameters were sufficient to capture M
phase for each cell while reducing photobleaching during the extended time
course. Temperature was maintained throughout all experiments at 28.5�C
using a stage incubator. Nuclear migration velocities were measured as
described (Baye and Link, 2007b). For details, see the Supplemental Data.
SUPPLEMENTAL DATA
The Supplemental Data include Supplemental Experimental Procedures, ten
figures, one table, and three movies and can be found with this article online
at http://www.cell.com/cgi/content/full/134/6/1055/DC1/.
ACKNOWLEDGMENTS
We thank D.Y. Stainier, S. Guo, J. Malicki, J. Lewis, I. Masai, P.A. Raymond,
B.D. Perkins, A.B. Chitnis, N. Katsanis, K.T. Vaughan, R.B. Vallee, and H.A.
Ingraham for fish strains, antibodies, in situ probes, and mouse embryos.
We thank Jeremy Reiter for critically reading the manuscript. F.D.B. was
supported by a Human Frontier Science Program long-term fellowship.
A.M.W. was supported by an American Association of University Women
dissertation fellowship. This work was supported by NIH grants EY013855
(H.B.), EY012406 (H.B.), EY01467 (B.A.L.), and the March of Dimes Foundation
(H.B.).
Received: September 12, 2007
Revised: April 25, 2008
Accepted: July 11, 2008
Published: September 18, 2008
Cell 134, 1055–1065, September 19, 2008 ª2008 Elsevier Inc. 1063

REFERENCES
Austin, C.P., Feldman, D.E., Ida, J.A., Jr., and Cepko, C.L. (1995). Vertebrate
retinal ganglion cells are selected from competent progenitors by the action
of Notch. Development 121, 3637–3650.
Baye, L.M., and Link, B.A. (2007a). Interkinetic nuclear migration and the
selection of neurogenic cell divisions during vertebrate retinogenesis.
J. Neurosci. 27, 10143–10152.
Baye, L.M., and Link, B.A. (2007b). The disarrayed mutation results in cell cycle
and neurogenesis defects during retinal development in zebrafish. BMC Dev.
Biol. 7, 28.
Baye, L.M., and Link, B.A. (2008). Nuclear migration during retinal develop-
ment. Brain Res. 1192, 29–36.
Berdnik, D., Torok, T., Gonzalez-Gaitan, M., and Knoblich, J.A. (2002). The
endocytic protein alpha-Adaptin is required for numb-mediated asymmetric
cell division in Drosophila. Dev. Cell 3, 221–231.
Bort, R., Signore, M., Tremblay, K., Martinez Barbera, J.P., and Zaret, K.S.
(2006). Hex homeobox gene controls the transition of the endoderm to a pseu-
dostratified, cell emergent epithelium for liver bud development. Dev. Biol.
290, 44–56.
Brown, N.L., Patel, S., Brzezinski, J., and Glaser, T. (2001). Math5 is required
for retinal ganglion cell and optic nerve formation. Development 128, 2497–
2508.
Burns, K.A., and Kuan, C.Y. (2005). Low doses of bromo- and iododeoxyuri-
dine produce near-saturation labeling of adult proliferative populations in the
dentate gyrus. Eur. J. Neurosci. 21, 803–807.
Cepko, C.L., Austin, C.P., Yang, X., Alexiades, M., and Ezzeddine, D. (1996).
Cell fate determination in the vertebrate retina. Proc. Natl. Acad. Sci. USA
93, 589–595.
Chitnis, A.B. (1995). The role of Notch in lateral inhibition and cell fate specifi-
cation. Mol. Cell. Neurosci. 6, 311–321.
Doerre, G., and Malicki, J. (2001). A mutation of early photoreceptor develop-
ment, mikre oko, reveals cell-cell interactions involved in the survival and
differentiation of zebrafish photoreceptors. J. Neurosci. 21, 6745–6757.
Draper, B.W., Morcos, P.A., and Kimmel, C.B. (2001). Inhibition of zebrafish
fgf8 pre-mRNA splicing with morpholino oligos: a quantifiable method for
gene knockdown. Genesis 30, 154–156.
Edlund, T., and Jessell, T.M. (1999). Progression from extrinsic to intrinsic
signaling in cell fate specification: a view from the nervous system. Cell 96,
211–224.
Emery, G., Hutterer, A., Berdnik, D., Mayer, B., Wirtz-Peitz, F., Gaitan, M.G.,
and Knoblich, J.A. (2005). Asymmetric Rab 11 endosomes regulate delta recy-
cling and specify cell fate in the Drosophila nervous system. Cell 122, 763–773.
Fan, S.S., and Ready, D.F. (1997). Glued participates in distinct microtubule-
based activities in Drosophila eye development. Development 124, 1497–
1507.
Frade, J.M. (2002). Interkinetic nuclear movement in the vertebrate neuroepi-
thelium: encounters with an old acquaintance. Prog. Brain Res. 136, 67–71.
Furukawa, T., Mukherjee, S., Bao, Z.Z., Morrow, E.M., and Cepko, C.L. (2000).
rax, Hes1, and notch1 promote the formation of Muller glia by postnatal retinal
progenitor cells. Neuron 26, 383–394.
Gaiano, N., Nye, J.S., and Fishell, G. (2000). Radial glial identity is promoted by
Notch1 signaling in the murine forebrain. Neuron 26, 395–404.
Geldmacher-Voss, B., Reugels, A.M., Pauls, S., and Campos-Ortega, J.A.
(2003). A 90-degree rotation of the mitotic spindle changes the orientation of
mitoses of zebrafish neuroepithelial cells. Development 130, 3767–3780.
Guo, M., Jan, L.Y., and Jan, Y.N. (1996). Control of daughter cell fates during
asymmetric division: interaction of Numb and Notch. Neuron 17, 27–41.
Hatakeyama, J., and Kageyama, R. (2004). Retinal cell fate determination and
bHLH factors. Semin. Cell Dev. Biol. 15, 83–89.
Ho, R.K., and Kane, D.A. (1990). Cell-autonomous action of zebrafish spt-1
mutation in specific mesodermal precursors. Nature 348, 728–730.
1064 Cell 134, 1055–1065, September 19, 2008 ª2008 Elsevier Inc.
Holt, C.E., Bertsch, T.W., Ellis, H.M., and Harris, W.A. (1988). Cellular determi-
nation in the Xenopus retina is independent of lineage and birth date. Neuron 1,
15–26.
Jadhav, A.P., Cho, S.H., and Cepko, C.L. (2006). Notch activity permits retinal
cells to progress through multiple progenitor states and acquire a stem cell
property. Proc. Natl. Acad. Sci. USA 103, 18998–19003.
Kawakami, K. (2004). Transgenesis and gene trap methods in zebrafish by
using the Tol2 transposable element. Methods Cell Biol. 77, 201–222.
Kay, J.N., Finger-Baier, K.C., Roeser, T., Staub, W., and Baier, H. (2001). Ret-
inal ganglion cell genesis requires lakritz, a Zebrafish atonal Homolog. Neuron
30, 725–736.
Kim, J., Wu, H.H., Lander, A.D., Lyons, K.M., Matzuk, M.M., and Calof, A.L.
(2005). GDF11 controls the timing of progenitor cell competence in developing
retina. Science 308, 1927–1930.
King, S.J., and Schroer, T.A. (2000). Dynactin increases the processivity of the
cytoplasmic dynein motor. Nat. Cell Biol. 2, 20–24.
Koster, R.W., and Fraser, S.E. (2001). Tracing transgene expression in living
zebrafish embryos. Dev. Biol. 233, 329–346.
Le Borgne, R. (2006). Regulation of Notch signalling by endocytosis and endo-
somal sorting. Curr. Opin. Cell Biol. 18, 213–222.
Livesey, F.J., and Cepko, C.L. (2001). Vertebrate neural cell-fate determina-
tion: lessons from the retina. Nat. Rev. Neurosci. 2, 109–118.
Marquardt, T., and Gruss, P. (2002). Generating neuronal diversity in the retina:
one for nearly all. Trends Neurosci. 25, 32–38.
Masai, I., Stemple, D.L., Okamoto, H., and Wilson, S.W. (2000). Midline signals
regulate retinal neurogenesis in zebrafish. Neuron 27, 251–263.
Masai, I., Lele, Z., Yamaguchi, M., Komori, A., Nakata, A., Nishiwaki, Y., Wada,
H., Tanaka, H., Nojima, Y., Hammerschmidt, M., et al. (2003). N-cadherin me-
diates retinal lamination, maintenance of forebrain compartments and pattern-
ing of retinal neurites. Development 130, 2479–2494.
McGill, M.A., and McGlade, C.J. (2003). Mammalian numb proteins promote
Notch1 receptor ubiquitination and degradation of the Notch1 intracellular
domain. J. Biol. Chem. 278, 23196–23203.
McGrail, M., Gepner, J., Silvanovich, A., Ludmann, S., Serr, M., and Hays, T.S.
(1995). Regulation of cytoplasmic dynein function in vivo by the Drosophila
Glued complex. J. Cell Biol. 131, 411–425.
Meng, A., Jessen, J.R., and Lin, S. (1999). Transgenesis. Methods Cell Biol. 60,
133–148.
Morrison, S.J., Perez, S.E., Qiao, Z., Verdi, J.M., Hicks, C., Weinmaster, G.,
and Anderson, D.J. (2000). Transient Notch activation initiates an irreversible
switch from neurogenesis to gliogenesis by neural crest stem cells. Cell 101,
499–510.
Murciano, A., Zamora, J., Lopez-Sanchez, J., and Frade, J.M. (2002). Interki-
netic nuclear movement may provide spatial clues to the regulation of neuro-
genesis. Mol. Cell. Neurosci. 21, 285–300.
Muto, A., Arai, K., and Watanabe, S. (2006). Rab11-FIP4 is predominantly
expressed in neural tissues and involved in proliferation as well as in differen-
tiation during zebrafish retinal development. Dev. Biol. 292, 90–102.
Nelson, B.R., Gumuscu, B., Hartman, B.H., and Reh, T.A. (2006). Notch activity
is downregulated just prior to retinal ganglion cell differentiation. Dev. Neuro-
sci. 28, 128–141.
Nichols, J.T., Miyamoto, A., and Weinmaster, G. (2007). Notch signaling–con-
stantly on the move. Traffic 8, 959–969.
Ohnuma, S., Hopper, S., Wang, K.C., Philpott, A., and Harris, W.A. (2002).
Co-ordinating retinal histogenesis: early cell cycle exit enhances early cell
fate determination in the Xenopus retina. Development 129, 2435–2446.
Passini, M.A., Levine, E.M., Canger, A.K., Raymond, P.A., and Schechter, N.
(1997). Vsx-1 and Vsx-2: differential expression of two paired-like homeobox
genes during zebrafish and goldfish retinogenesis. J. Comp. Neurol. 388,
495–505.
Pearson, B.J., and Doe, C.Q. (2004). Specification of temporal identity in the
developing nervous system. Annu. Rev. Cell Dev. Biol. 20, 619–647.

Perron, M., and Harris, W.A. (2000). Determination of vertebrate retinal progen-
itor cell fate by the Notch pathway and basic helix-loop-helix transcription fac-
tors. Cell. Mol. Life Sci. 57, 215–223.
Poggi, L., Vitorino, M., Masai, I., and Harris, W.A. (2005a). Influences on neural
lineage and mode of division in the zebrafish retina in vivo. J. Cell Biol. 171,
991–999.
Poggi, L., Zolessi, F.R., and Harris, W.A. (2005b). Time-lapse analysis of retinal
differentiation. Curr. Opin. Cell Biol. 17, 676–681.
Reugels, A.M., Boggetti, B., Scheer, N., and Campos-Ortega, J.A. (2006).
Asymmetric localization of Numb:EGFP in dividing neuroepithelial cells during
neurulation in Danio rerio. Dev. Dyn. 235, 934–948.
Sauer, F.C. (1935). Mitosis in the neural tube. J. Comp. Neurol. 62, 377–405.
Scheer, N., Groth, A., Hans, S., and Campos-Ortega, J.A. (2001). An instruc-
tive function for Notch in promoting gliogenesis in the zebrafish retina. Devel-
opment 128, 1099–1107.
Schroer, T.A. (2004). Dynactin. Annu. Rev. Cell Dev. Biol. 20, 759–779.
Starr, D.A., and Fischer, J.A. (2005). KASH ’n Karry: the KASH domain family of
cargo-specific cytoskeletal adaptor proteins. Bioessays 27, 1136–1146.
Takke, C., Dornseifer, P., v. Weizsacker, E., and Campos-Ortega, J.A. (1999).
her4, a zebrafish homologue of the Drosophila neurogenic gene E(spl), is a tar-
get of NOTCH signalling. Development 126, 1811–1821.
Tokunaga, A., Kohyama, J., Yoshida, T., Nakao, K., Sawamoto, K., and Okano,
H. (2004). Mapping spatio-temporal activation of Notch signaling during neu-
rogenesis and gliogenesis in the developing mouse brain. J. Neurochem. 90,
142–154.
Tsujikawa, M., Omori, Y., Biyanwila, J., and Malicki, J. (2007). Mechanism of
positioning the cell nucleus in vertebrate photoreceptors. Proc. Natl. Acad.
Sci. USA 104, 14819–14824.
Turner, D.L., and Cepko, C.L. (1987). A common progenitor for neurons and
glia persists in rat retina late in development. Nature 328, 131–136.
Turner, D.L., Snyder, E.Y., and Cepko, C.L. (1990). Lineage-independent de-
termination of cell type in the embryonic mouse retina. Neuron 4, 833–845.
Vetter, M.L., and Brown, N.L. (2001). The role of basic helix-loop-helix genes in
vertebrate retinogenesis. Semin. Cell Dev. Biol. 12, 491–498.
Wang, S.W., Kim, B.S., Ding, K., Wang, H., Sun, D., Johnson, R.L., Klein, W.H.,
and Gan, L. (2001). Requirement for math5 in the development of retinal gan-
glion cells. Genes Dev. 15, 24–29.
Wehman, A.M., Staub, W., Meyers, J.R., Raymond, P.A., and Baier, H. (2005).
Genetic dissection of the zebrafish retinal stem-cell compartment. Dev. Biol.
281, 53–65.
Wehman, A.M., Staub, W., and Baier, H. (2007). The anaphase-promoting
complex is required in both dividing and quiescent cells during zebrafish
development. Dev. Biol. 303, 144–156.
Wei, X., Cheng, Y., Luo, Y., Shi, X., Nelson, S., and Hyde, D.R. (2004). The
zebrafish Pard3 ortholog is required for separation of the eye fields and retinal
lamination. Dev. Biol. 269, 286–301.
Wetts, R., and Fraser, S.E. (1988). Multipotent precursors can give rise to all
major cell types of the frog retina. Science 239, 1142–1145.
Whited, J.L., Cassell, A., Brouillette, M., and Garrity, P.A. (2004). Dynactin is
required to maintain nuclear position within postmitotic Drosophila photore-
ceptor neurons. Development 131, 4677–4686.
Xiao, T., Roeser, T., Staub, W., and Baier, H. (2005). A GFP-based genetic
screen reveals mutations that disrupt the architecture of the zebrafish retino-
tectal projection. Development 132, 2955–2967.
Yan, R.T., Ma, W., Liang, L., and Wang, S.Z. (2005). bHLH genes and retinal
cell fate specification. Mol. Neurobiol. 32, 157–171.
Yeo, S.Y., Kim, M., Kim, H.S., Huh, T.L., and Chitnis, A.B. (2007). Fluorescent
protein expression driven by her4 regulatory elements reveals the spatiotem-
poral pattern of Notch signaling in the nervous system of zebrafish embryos.
Dev. Biol. 301, 555–567.
Cell 134, 1055–1065, September 19, 2008 ª2008 Elsevier Inc. 1065
Copyright © 2022 FDOKUMEN




















