
Regulation of acetylcholinesterase expression by calcium signaling during calcium ionophore A23187-...
16
The International Journal of Biochemistry & Cell Biology 39 (2007) 93–108 Regulation of acetylcholinesterase expression by calcium signaling during calcium ionophore A23187- and thapsigargin-induced apoptosis Hui Zhu a , Wei Gao a , Hua Jiang a , Qi-Huang Jin a , Yu-Fang Shi b , Karl W.K. Tsim c , Xue-Jun Zhang a,∗ a Laboratory of Molecular Cell Biology, Institute of Biochemistry and Cell Biology, Shanghai Institutes for Biological Sciences, Chinese Academy of Sciences, Graduate School of the Chinese Academy of Sciences, 320 YueYang Road, Shanghai 200031, China b Department of Molecular Genetics, Microbiology and Immunology, University of Medicine and Dentistry of New Jersey-Robert Wood Johnson Medical School, USA c Department of Biology and Molecular Neuroscience Center, The Hong Kong University of Science and Technology, Clear Water Bay, Kowloon, Hong Kong, China Received 19 March 2006; received in revised form 6 June 2006; accepted 13 June 2006 Available online 13 July 2006 Abstract We have recently reported that acetylcholinesterase expression was induced during apoptosis in various cell types. In the current study we provide evidence to suggest that the induction of acetylcholinesterase expression during apoptosis is regulated by the mobilization of intracellular Ca 2+ . During apoptosis, treatment of HeLa and MDA-MB-435s cells with the calcium ionophore A23187 resulted in a significant increase in acetylcholinesterase mRNA and protein levels. Chelation of intracellular Ca 2+ by BAPTA- AM (1,2-bis-(2-aminophenoxy)ethane-N,N,N ,N -tetraacetic acid–acetoxymethyl ester), an intracellular Ca 2+ chelator, inhibited acetylcholinesterase expression. A23187 also enhanced the stability of acetylcholinesterase mRNA and increased the activity of acetylcholinesterase promoter, effects that were blocked by BAPTA-AM. Perturbations of cellular Ca 2+ homeostasis by thapsigargin resulted in the increase of acetylcholinesterase expression as well as acetylcholinesterase promoter activity during thapsigargin induced apoptosis in HeLa and MDA-MB-435s cells, effects that were also inhibited by BAPTA-AM. We further demonstrated that the transactivation of the human acetylcholinesterase promoter by A23187 and thapsigargin was partially mediated by a CCAAT motif within the −1270 to −1248 fragment of the human acetylcholinesterase promoter. This motif was able to bind to CCAAT binding factor (CBF/NF-Y). These results strongly suggest that cytosolic Ca 2+ plays a key role in acetylcholinesterase regulation during apoptosis induced by A23187 and thapsigargin. © 2006 Elsevier Ltd. All rights reserved. Keywords: Acetylcholinesterase; Apoptosis; Ca 2+ ; mRNA stability; Transcriptional activity Abbreviations: AChE, acetylcholinesterase, acetylcholine acetyl hydrolase; BAPTA-AM, 1,2-bis-(2-aminophenoxy)ethane-N,N,N ,N - tetraacetic acid–acetoxymethyl ester; TG, thapsigargin; TUNEL, terminal deoxynucleotide transferase-mediated-dUTP nick end labeling ∗ Corresponding author. Tel.: +86 21 54921402; fax: +86 21 54921403. E-mail address: [email protected] (X.-J. Zhang). 1357-2725/$ – see front matter © 2006 Elsevier Ltd. All rights reserved. doi:10.1016/j.biocel.2006.06.012
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Regulation of acetylcholinesterase expression by calcium signaling during calcium ionophore A23187-...

The International Journal of Biochemistry & Cell Biology 39 (2007) 93–108
Regulation of acetylcholinesterase expression by calciumsignaling during calcium ionophore A23187- and
thapsigargin-induced apoptosis
Hui Zhu a, Wei Gao a, Hua Jiang a, Qi-Huang Jin a,Yu-Fang Shi b, Karl W.K. Tsim c, Xue-Jun Zhang a,∗
a Laboratory of Molecular Cell Biology, Institute of Biochemistry and Cell Biology,Shanghai Institutes for Biological Sciences, Chinese Academy of Sciences,
Graduate School of the Chinese Academy of Sciences, 320 YueYang Road, Shanghai 200031, Chinab Department of Molecular Genetics, Microbiology and Immunology,
University of Medicine and Dentistry of New Jersey-Robert Wood Johnson Medical School, USAc Department of Biology and Molecular Neuroscience Center,
The Hong Kong University of Science and Technology, Clear Water Bay, Kowloon, Hong Kong, China
Received 19 March 2006; received in revised form 6 June 2006; accepted 13 June 2006Available online 13 July 2006
Abstract
We have recently reported that acetylcholinesterase expression was induced during apoptosis in various cell types. In the currentstudy we provide evidence to suggest that the induction of acetylcholinesterase expression during apoptosis is regulated by themobilization of intracellular Ca2+. During apoptosis, treatment of HeLa and MDA-MB-435s cells with the calcium ionophoreA23187 resulted in a significant increase in acetylcholinesterase mRNA and protein levels. Chelation of intracellular Ca2+ by BAPTA-AM (1,2-bis-(2-aminophenoxy)ethane-N,N,N′,N′-tetraacetic acid–acetoxymethyl ester), an intracellular Ca2+ chelator, inhibitedacetylcholinesterase expression. A23187 also enhanced the stability of acetylcholinesterase mRNA and increased the activity ofacetylcholinesterase promoter, effects that were blocked by BAPTA-AM. Perturbations of cellular Ca2+ homeostasis by thapsigarginresulted in the increase of acetylcholinesterase expression as well as acetylcholinesterase promoter activity during thapsigargininduced apoptosis in HeLa and MDA-MB-435s cells, effects that were also inhibited by BAPTA-AM. We further demonstrated thatthe transactivation of the human acetylcholinesterase promoter by A23187 and thapsigargin was partially mediated by a CCAATmotif within the −1270 to −1248 fragment of the human acetylcholinesterase promoter. This motif was able to bind to CCAATbinding factor (CBF/NF-Y). These results strongly suggest that cytosolic Ca2+ plays a key role in acetylcholinesterase regulation
during apoptosis induced by A23187 and thapsigargin.© 2006 Elsevier Ltd. All rights reserved.Keywords: Acetylcholinesterase; Apoptosis; Ca2+; mRNA stability; Transcriptional activity
Abbreviations: AChE, acetylcholinesterase, acetylcholine acetyl hydrolase; BAPTA-AM, 1,2-bis-(2-aminophenoxy)ethane-N,N,N′,N′-tetraacetic acid–acetoxymethyl ester; TG, thapsigargin; TUNEL, terminal deoxynucleotide transferase-mediated-dUTP nick end labeling
∗ Corresponding author. Tel.: +86 21 54921402; fax: +86 21 54921403.E-mail address: [email protected] (X.-J. Zhang).
1357-2725/$ – see front matter © 2006 Elsevier Ltd. All rights reserved.doi:10.1016/j.biocel.2006.06.012
zhang xue jun
Note
IF 4.804 2006

f Bioch
94 H. Zhu et al. / The International Journal o1. Introduction
Acetylcholinesterase (AChE) is a type B car-boxylesterase that rapidly hydrolyzes acetylcholine(ACh) at cholinergic synapses and neuromuscu-lar junctions (Taylor & Radic, 1994). It is alsoinvolved in many non-cholinergic functions, suchas cell adhesion, proliferation, and neurite out-growth (Day & Greenfield, 2002; Johnson & Moore,2004; Karpel et al., 1996; Koenigsberger, Chiappa,& Brimijoin, 1997; Whyte & Greenfield, 2003).Three different mature AChE mRNA transcripts,which encode AChE-T (tailed isoform), AChE-E (erythrocytic isoform) and AChE-R (readthroughisoform), have been produced (Massoulie et al.,1998).
The molecular mechanisms regulating ACHE geneexpression have been studied mostly in muscle cells(Angus, Chan, & Jasmin, 2001; Luo, Fuentes, & Taylor,1994; Tung et al., 2004), neurons and hematopoieticcells (Soreq & Seidman, 2001). It is noteworthy that inall three systems, post-transcriptional regulatory mech-anisms seem to play an important role in the induc-tion of AChE expression by stabilizing existing tran-scripts (Chan, Adatia, Krupa, & Jasmin, 1998; Coleman& Taylor, 1996; Deschenes-Furry, Belanger, Perrone-Bizzozero, & Jasmin, 2003; Luo et al., 1994). On theother hand, several regulatory elements within the AChEpromoter have been reported to regulate the transcrip-tional activity of the ACHE gene in Torpedo, mouse,rat and human (Angus et al., 2001; Ben Aziz-Aloya etal., 1993; Chan, Boudreau-Lariviere, Angus, Mankal,& Jasmin, 1999; Ekstrom, Klump, Getman, Karin, &Taylor, 1993; Mutero, Camp, & Taylor, 1995; Siow etal., 2002).
Recent research in our laboratory revealed thatAChE expression was also induced during apoptosisin various cell types, including those cells originatingfrom non-muscle, non-nervous or non-hematopoieticsystems, suggesting that AChE might be a novelregulator of apoptosis (Zhang et al., 2002). Ourhypothesis was in agreement with a recent study byPark, Kim, and Yoo (2004), which elegantly demon-strated that AChE exerted a pivotal role in apop-tosis by participating in the formation of apopto-some. However, the mechanisms that regulate theexpression of AChE during apoptosis have not beenreported.
Elevation of intracellular Ca2+ increases AChE tran-script levels during differentiation of myoblasts tomyotubes (Luo et al., 1994). In cultured chick myotubes,ATP acts via the ATP receptor to stimulate AChE
emistry & Cell Biology 39 (2007) 93–108
expression, which is mediated by protein kinase C andintracellular Ca2+ release (Choi et al., 2003). Amy-loid �-protein (A�), such as A�1–40 and A�25–35,was also demonstrated to increase AChE activity inP19 embryonic carcinoma and retinal cells, via a cal-cium influx mechanism (Melo, Agostinho, & Oliveira,2003; Sberna, Saez-Valero, Beyreuther, Masters, &Small, 1997). These studies suggested that Ca2+-dependent signaling pathways played a role in regu-lating AChE expression or activity in muscle and neu-ron.
Calcium is an important second messenger and intra-cellular Ca2+ plays an important role in the modu-lation of apoptosis (Demaurex & Distelhorst, 2003;Groenendyk, Lynch, & Michalak, 2004; Mattson &Chan, 2003). Overload of Ca2+ has even been sug-gested to be the final common pathway of all types ofcell death (Rizzuto et al., 2003). Our previous studydemonstrated that AChE expression was induced uponinduction of apoptosis by various stimuli in variouscell types (Zhang et al., 2002), further suggesting thatthe induction of AChE expression is an universal prop-erty of apoptotic cells. We hypothesize that AChEexpression during apoptosis is related to Ca2+ mobiliza-tion.
The calcium ionophore A23187 induces apoptosis byincreasing cytosolic Ca2+ in various cell types (Azmi,Dhawan, & Singh, 1996; Petersen, Castilho, Hansson,Wieloch, & Brundin, 2000). The sarcoplasmic retic-ulum Ca2+-ATPase pump inhibitor thapsigargin (TG)depletes Ca2+ within the endoplasmic reticulum (ER),resulting in a transient increase in cytoplasmic Ca2+
levels (Sagara & Inesi, 1991), and perturbations in cel-lular Ca2+ homeostasis have been also been reportedto induce apoptosis in both normal and malignant cells(Krebs, 1998). In the current study, we found that, incultured cells, the expression of AChE was markedlyincreased in both A23187-induced or TG-induced HeLaand MDA-MB-435s cell apoptosis, an effect which wasinhibited by BAPTA-AM, a Ca2+ chelator. In addition,A23187 and TG increased the activity of a 2.2 kb humanAChE promoter. The promoter sequence of ACHE genehas been functionally characterized (Ben Aziz-Aloyaet al., 1993; Siow et al., 2002; Wan, Choi, Siow, &Tsim, 2000). A CCAAT motif within the −1270 to−1248 fragment of the human AChE promoter wasshown to be activated by A23187 and TG induction.Moreover, we suggested, for the first time, that the
CCAAT binding factor (CBF/NF-Y) might be a poten-tial transcriptional factor mediating AChE transcrip-tional activity during apoptosis induced by A23187 andTG.
f Bioch
2
2
1ib(b(crsvHsi
2a
poCoamaMcnTt3r
2r
(rtpoTTTdA
H. Zhu et al. / The International Journal o
. Materials and methods
.1. Cell culture and treatments
HeLa and MDA cells were maintained in RPMI640 (GIBCO-BRL, Gaithersburg, MD) with 10% heat-nactivated fetal bovine serum (GIBCO-BRL, Gaithers-urg, MD). Apoptosis was induced by A23187 (2 �M)Calbiochem, La Jolla, CA) and TG (2 �M) (Cal-iochem, La Jolla, CA) in HeLa cells and A231872 �M) and TG (0.5 �M) in MDA cells. Given that MDAells were much more sensitive to TG than HeLa cells,elatively lower concentrations of TG were used in ourtudy. Apoptosis was assessed by morphology obser-ation (rounding, shrinkage and detachment of cells),oechst staining (chromatin condensation), TUNEL
taining (DNA fragmentation), and cleaved caspase-3mmunocytochemistry staining.
.2. Double staining with immunocytochemistrynd TUNEL reaction
Immunocytochemistry analysis protocol has beenreviously described (Zhang et al., 2002). AChE mon-clonal antibody was from BD Biosciences (San Jose,A) with the epitope mapping at the 411–609 peptidef the human AChE C-terminus. The AChE monoclonalntibody detected endogenous levels of a 68 kDa frag-ent of human AChE. Cleaved caspase-3 (Asp 175)
ntibody was from Cell Signaling Technology (Beverly,A); Rhodamine coupled anti-mouse IgG and FITC
oupled anti-rabbit IgG were from Santa Cruz Biotech-ology (Santa Cruz, CA). Cell pellets were suspended inUNEL reaction mixture (Roche Diagnostics Corpora-
ion Indianapolis, IN) for 1 h and stained with Hoechst3258. Stained cells were analyzed with a Nikon fluo-escence microscope (Nikon Inc.).
.3. Semiquantitative RT-PCR and two stepeal-time quantitative RT-PCR
Total RNA was extracted using TRIzol ReagentInvitrogen, Carlsbad, CA) and was subjected toeverse transcription using M-MLV Reverse Transcrip-ase (Promega, Madison, WI). Reverse transcriptionroducts were used for semiquantitative RT-PCR withligonucleotide pairs specific for AChE and GAPDH.he primer pair (5′-CGG GTC TAC GCC TAC GTC
TT GAA CAC CGT GCT TC-3′; 5′-CAC AGGCT GAG CAG CGA TCC TGC TTG CTG-3′) wasesigned to generate a 482 bp fragment of the tailedChE-T mRNA, including exons E1–E2–E3–E4–E6,emistry & Cell Biology 39 (2007) 93–108 95
according to Homo sapiens acetylcholinesterase (YTblood group) (ACHE), transcript variant E4–E6, mRNA,NM 000665. The primer pair (5′-CGG GTC TAC GCCTAC GTC TTT GAA CAC CGT GCT TC-3′; 5′-ATG GGT GAA GCC TGG GCA GGT G-3′) wasdesigned to generate two different fragments, 399 and478 bp, to detect AChE mRNA encoding AChE-E (ery-throcytic isoform, including exons E1–E2–E3–E4–E5)and AChE-R (readthrough isoform, including exonsE1–E2–E3–E4–4′–E5), respectively. Primers for house-keeping gene GAPDH (5′-CCA CCC ATG GCA AATTCC ATG GCA-3′; 5′-TCT AGA CGG CAG GTC AGGTCC ACC-3′) were used as an internal control, with anexpected product size of 588 bp.
Real-time PCR was performed with a PE BiosystemsGeneAmp 5700 sequence detection system (PE Biosys-tems, Foster City, CA) using a SYBR green I PCR kit (PEBiosystems, Foster City, CA). Each reaction contained2.5 �l of the 10× SYBR green buffer, 200 nM dATP,dGTP, and dCTP; 400 nM dUTP, 2 mM MgCl2, 0.25 U ofuracil N-glycosylase (UNG), 0.625 U of Amplitaq GoldDNA polymerase (PE Biosystems, Foster City, CA),100 nM AChE primers (5′-AGC CGA GGC TGT GGTCCT GCA TTA CA-3′; 5′-CGC ATC AGT CGC TGGGCG AAG ATT TT-3′) or 25 nM GAPDH primers (asnoted above); 1 �l 1:10 dilution of cDNA; and water to afinal volume of 50 �l (n = 3/sample). AChE mRNA lev-els relative to GAPDH internal control was determinedby the following formula: 2−(CtAChE − CtGAPDH)Time X,where the “Ct” was defined as the cycle number atwhich fluorescence crossed the threshold and “TimeX” was the indicated time point. �-Actin was used inMDA cells as the internal control, with primers (5′-GCG GGA AAT CGT GCG TGA CAT T-3′; 5′-GATGGA GTT GAA GGT AGT TTC GTG-3′). AChEmRNA levels were determined utilizing the formula:2−(CtAChE − Ct�-actin)Time X.
2.4. Western blotting
Immunoblotting protocol has been previouslydescribed (Zhang et al., 2002). Anti-�-actin mAb wasfrom Sigma (St. Louis, MO); HRP-conjugated sec-ondary antibody was Santa Cruz Biotechnology. Theimmunoreactive protein was visualized using a chemi-luminescence detection kit (ECL, Santa Cruz Biotech-nology).
2.5. Transient transfections and luciferase assays
The 2.2 kb DNA fragment of the human AChE pro-moter (Ben Aziz-Aloya et al., 1993) was subcloned

f Bioch
96 H. Zhu et al. / The International Journal ointo BglII and HindIII sites of the pGL3 basic vector(Promega, Madison, WI) with a downstream tagged fire-fly luciferase gene, pAChE-Luc (Wan et al., 2000). Adistal CCAAT binding mutation within the −1270 to−1248 fragment of the human AChE promoter was cre-ated using a PCR-based mutagenesis procedure with theforward (F) and reverse (R) end primers (F, 5′-ACGGTA CCG AGC TCT TAC GC-3′; R, 5′-CGG GTACCG CCG ATA TCA GGG GGT GTC-3′). The PCRproduct, including the CCAAT binding mutation, wasreplaced in the wild type pAChE-Luc with two KpnI sites to construct Distal CCAAT binding mutant. Aproximal CCAAT binding mutation within the −594 to−571 fragment of the human AChE promoter was cre-ated with the forward (F) and reverse (R) end primers (F,5′-TAG CCC GGG CTC GAG ATC T-3′; R, 5′-TAC CGGAA TGC CAA GCT T-3′) and the following primers(F, 5′-CAG AGA ATT CAT GAG AAG GGG CA-3′; R,5′-TCA TGA ATT CTC TGG CAC CGA GCG-3′). Theproximal CCAAT binding mutation was incorporate inthe wild type pAChE-Luc to construct proximal CCAATbinding mutant. All polymerase chain reaction-derivedconstructs were confirmed by sequencing.
HeLa and MDA cells were cotransfected with fireflyluciferase reporter construct pAChE-Luc, Distal CCAATbinding mutant or Proximal CCAAT binding mutant,with Renilla luciferase reporter plasmid pRL-SV40(Promega, Madison, WI) used as the internal controlutilizing LipofectAMINE regent (Invitrogen, Carlsbad,CA). Apoptosis was induced, 24 h after transfection,by A23187 or TG (Calbiochem, Darmstadt, Germany).Luciferase activity assays were performed with a Dual-Luciferase Reporter Assay System (Promega, Madison,WI), and activity was measured with a luminometer BGP(MGM). Firefly luciferase activity was normalized toRenilla luciferase activity in each well (n = 3/transfectionexperiment).
2.6. Electrophoretic mobility shift assays (EMSAs)
Nuclear extracts of HeLa cells were prepared accord-ing to Dignam et al. (Dignamm, Martin, Shastry,& Roeder, 1983) with some modifications. Elec-trophoretic mobility shift assays were performed withnuclear extract protein (4 �g) in binding buffer (10 mMTris–HCl, 1 mM MgCl2, 0.5 mM EDTA, 50 mM NaCl,0.5 mM DTT, 4% glycerol, pH 7.5) and poly (dI–dC)(1 �g). Binding reactions were incubated (20 min, room
temperature) with 20,000 cpm (0.1–0.5 ng) double-stranded oligonucleotides end-labeled with [�-32P]ATP using T4 polynucleotide kinase (Invitrogen). Anexcess of unlabeled double-stranded oligonucleotidesemistry & Cell Biology 39 (2007) 93–108
was used as competitors, where indicated. Double-stranded oligonucleotides corresponding to the distalCCAAT binding site and the proximal binding site were5′-ACC CCC TAA TTG GGG CGG TAC CC-3′ and 5′-TGC CAG ACA TTG GAT GAG AAG GG-3′, named WtCCAAT Dist and Wt CCAAT Prox, respectively. Mutantdouble-stranded oligonucleotides corresponding to thedistal CCAAT binding site were 5′-ACC CCC TGATAT CGG CGG TAC CC-3′, named Mut CCAAT Dist.The consensus sequences of the CCAAT motif oligonu-cleotides were as follows: C/EBP� (5′-TGC AGA TTGCGC AAT CTG CA-3′) (sc-2525; Santa Cruz), CCAATprotein 1 (CP1) (5′-AGA CTT TGT AAA AAG ACTAAA CAA TTT CAA CTC ACG AAA C-3′) (Elizondo,Corchero, Sterneck, & Gonzalez, 2000), CCAAT protein2 (CP2) (5′-GTT TTA CTC GGT AGA GCA AGC ACAAAC CAG-3′) (Elizondo et al., 2000), CCAAT transcrip-tion factor (CTF-2) (5′-TAC CTT ATT TTG GAT TGAAGC CAA TAT GAT AAT-3′) (Elizondo et al., 2000),C/EBP (5′-CTAGGGCTTGCGCAATCTATATTCG-3′)(Elizondo et al., 2000), CCAAT transcription factor(CTF-1) (5′-CCT TTG GCA TGC TGC CCA ATA TG-3′) (Roy & Lee, 1995), and CBF transcription factor(CBF wt) (5′-AGA CCG TAC GTG ATT GGT TAATCT CTT-3′) (sc-2591; Santa Cruz). Mutant oligonu-cleotides of the consensus binding site for the CBFtranscription factor were (CBF mut) (5′-AGA CCG TACGAA ATA CGG GAA TCT CTT-3′) (sc-2592; SantaCruz). Antibody supershift assays were carried out inthe same buffer, with further addition of 2 �l of anti-CBF-A/NF-YB (sc-13045X), anti-CBF-B/NF-YA (sc-10779 X), anti-CBF-C/NF-YC (sc-13044X) antibodies(Santa Cruz, CA), and anti-Oct 4 antibody (Santa Cruz,CA). The reaction was electrophoresed on a 4%, 0.5×TBE native polyacrylamide gel then autoradiographedto detect specific DNA–protein complex formation.
2.7. Chromatin immunoprecipitation assay
Assays were performed using a chromatin immuno-precipitation assay kit (Upstate Biotech, Lake Placid,NY) according to manufacturer’s protocol. Cells werefixed with 1% formaldehyde (15 min, 37 ◦C). Cell pel-lets were resuspended in SDS lysis buffer and soni-cated using an ultrasonic homogenizer VP-5S (TAITEC,Tokyo, Japan). After centrifugation, the supernatantwas diluted in chromatin immunoprecipitation dilutionbuffer then incubated (overnight, 4 ◦C) with anti-CBF-
A/NF-YB, CBF-B/NF-YA, and CBF-C/NF-YC antibod-ies (Santa Cruz Biotechnology). Immune complexeswere recovered by the addition of 60 �l of salmon spermDNA/protein A-agarose slurry, followed by incubation
f Bioch
(si(T5tnpPDa
2
t*
d
3
3e
fnnsagiSMtasmiTAioalcic
Ad
H. Zhu et al. / The International Journal o
2 h, 4 ◦C). Beads were washed with both low and highalt buffers, a LiCl buffer and a Tris/EDTA buffer andmmune complexes were eluted with an elution buffer1% SDS, 100 mM NaHCO3, and 1 mM dithiothreitol).o reverse DNA cross-linking, elutes were added toM NaCl (8 �l) and incubated (4 h, 65 ◦C), followed by
reatment with proteinase K. DNA was recovered by phe-olchloroform extraction and ethanol precipitation andellets were resuspended in 50 �l of Tris/EDTA buffer.CR was carried out for 35 cycles using 5 �l of sampleNA solution, and PCR products were separated on 2%
garose gels in 1× Tris–acetate/EDTA.
.8. Statistical analyses
Each experiment was repeated a minimum of threeimes and results were expressed as means ± S.E.M.*P < 0.005 and *P < 0.05 were calculated using Stu-ent’s two-tailed t test.
. Results
.1. A23187 and TG triggered induction of AChExpression during apoptosis in HeLa and MDA cells
The HeLa and MDA-MB-435s cell lines are derivedrom cervical epithelia and human breast ductal carci-omas, respectively. Living HeLa and MDA cells doot express AChE obviously. In order to establish ourystem, we treated HeLa and MDA cells with A23187nd TG. A23187, a Ca2+ ionophore, equilibrates Ca2+
radients across membranes and can cause a rapid risen intracellular Ca2+ levels (Bartlett, Luethy, Carlson,ollott, & Holbrook, 1992; Waser, Mesaeli, Spencer, &ichalak, 1997). Thapsigargin (TG), by inhibiting of
he sarcoplasmic reticulum Ca2+-ATPase pump, causestransient increase in cytoplasmic Ca2+ from ER Ca2+
tores, and a later influx of Ca2+ from the extracellularilieu, leading to the induction of apoptotic cell death
n various cell types (Jackisch et al., 2000; Kaneko &sukamoto, 1994; McColl et al., 1998). In our system,23187 and TG are both capable of inducing apoptosis
n HeLa cells and MDA cells assessed by morphol-gy observation, Hoechst staining, TUNEL staining, andctivated caspase-3 staining. Fig. 1 shows that at 48 h fol-owing A23187 and TG treatments, apoptosis of HeLaells was detected, as characterized by rounding, shrink-ng, chromatin condensation, DNA fragmentation and
aspase-3 activation.AChE expression during apoptosis induced by23187 and TG was also investigated. Fig. 1B and Cemonstrate the correlation between changes in cell mor-
emistry & Cell Biology 39 (2007) 93–108 97
phology, chromatin condensation, DNA fragmentationand AChE protein expression in HeLa cells. Apoptoticcells with condensed nuclei were positive for both AChEand TUNEL staining while living cells were negativefor both. Additionally, A23187 and TG induced AChEexpression in cleaved caspase-3 staining positive cells(Fig. 1D and E). Similar results were noted in MDA cells(data not shown). Our data demonstrated that AChE pro-tein expression was significantly increased in HeLa andMDA cells when they were induced to undergo apoptosisby A23187 and TG.
Semiquantitative RT-PCR confirmed our previousreport that the 482 bp tailed AChE-T fragment, but notthe AChE-E and AChE-R fragments, was induced dur-ing apoptosis (Yang, He, & Zhang, 2002; Zhang et al.,2002) (Fig. 2). Although transcriptional activation of theACHE gene is often associated with a shift in its splicingpattern, leading to accumulation of the rare AChE-R vari-ant under psychological, chemical, and physical stresses(Kaufer, Friedman, Seidman, & Soreq, 1998; Shohamiet al., 2000), AChE-R variant accumulation has beennoted mostly in mammalian brain neurons, with theshift dependant on neuronal activity (Soreq & Seidman,2001). AChE-R mRNA transcript has been suggested torepresent a general nervous system stress-response ele-ment (Shohami et al., 2000). In various cell apoptosis,we found that AChE-T variant, which is the major tran-script, increased obviously (Yang et al., 2002; Zhang etal., 2002).
3.2. Effect of calcium on AChE expression duringapoptosis induced by A23187 and TG in HeLa andMDA cells
We next examined whether the induction of AChEexpression and the increase in intracellular Ca2+ werecausally related. BAPTA-AM, an intracellular Ca2+
chelator, significantly reduced Ca2+ concentrations,maintaining it at low levels in the cytoplasm (Muallem,Khademazad, & Sachs, 1990). BAPTA-AM also pre-vented the dramatic increase in cytosolic Ca2+ medi-ated by A23187 and TG (Bartlett et al., 1992; He etal., 2002). Pre-treatment of HeLa and MDA cells withBAPTA-AM (10 �M, 45 min) inhibited the rapid risein cytosolic Ca2+ induced by A23187 and inhibited thetransient rise in cytosolic Ca2+ caused by TG (data notshown), in agreement with previous studies. Treatmentwith A23187 resulted in an increase in AChE mRNA
levels at 6 h, an early stage of apoptosis while HeLaand MDA cells did not possess obvious apoptotic mor-phologic features. However, pre-treatment of both twocell lines with BAPTA-AM (10 �M, 45 min) followed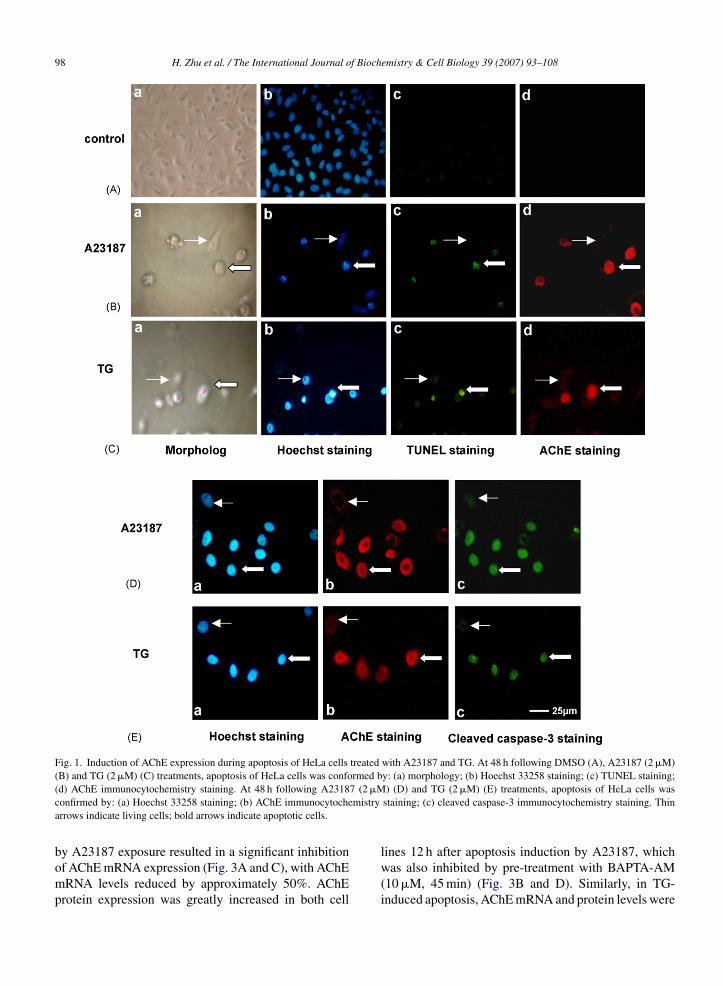
98 H. Zhu et al. / The International Journal of Biochemistry & Cell Biology 39 (2007) 93–108
Fig. 1. Induction of AChE expression during apoptosis of HeLa cells treated with A23187 and TG. At 48 h following DMSO (A), A23187 (2 �M)(B) and TG (2 �M) (C) treatments, apoptosis of HeLa cells was conformed by: (a) morphology; (b) Hoechst 33258 staining; (c) TUNEL staining;
7 (2 �Mmistry
(d) AChE immunocytochemistry staining. At 48 h following A2318confirmed by: (a) Hoechst 33258 staining; (b) AChE immunocytochearrows indicate living cells; bold arrows indicate apoptotic cells.
by A23187 exposure resulted in a significant inhibitionof AChE mRNA expression (Fig. 3A and C), with AChEmRNA levels reduced by approximately 50%. AChEprotein expression was greatly increased in both cell
) (D) and TG (2 �M) (E) treatments, apoptosis of HeLa cells wasstaining; (c) cleaved caspase-3 immunocytochemistry staining. Thin
lines 12 h after apoptosis induction by A23187, whichwas also inhibited by pre-treatment with BAPTA-AM(10 �M, 45 min) (Fig. 3B and D). Similarly, in TG-induced apoptosis, AChE mRNA and protein levels were

H. Zhu et al. / The International Journal of Bioch
FA14
sBAtA
3dM
Aua
FwAsw
ig. 2. Semiquantitative RT-PCR analysis of the induction of tailedChE-T mRNA. HeLa cells were treated with DMSO (control, lane) or A23187 (2 �M) for 6 h (lane 2), 12 h (lane 3), 24 h (lane 4), and8 h (lane 5).
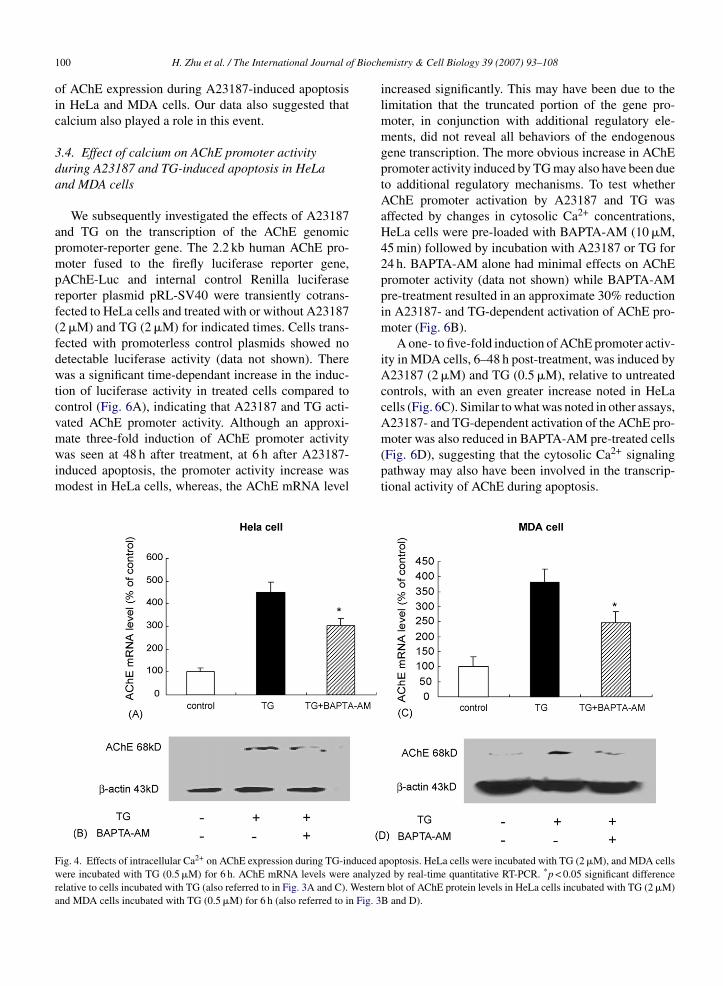
ignificantly increased at 6 h, and pre-treatment withAPTA-AM (10 �M, 45 min) also inhibited TG-inducedChE expression (Fig. 4). These data strongly suggested
hat cytosolic Ca2+ plays an important role in regulatingChE expression during apoptosis.
.3. Effect of calcium on AChE mRNA stabilityuring A23187-induced apoptosis of HeLa andDA cells
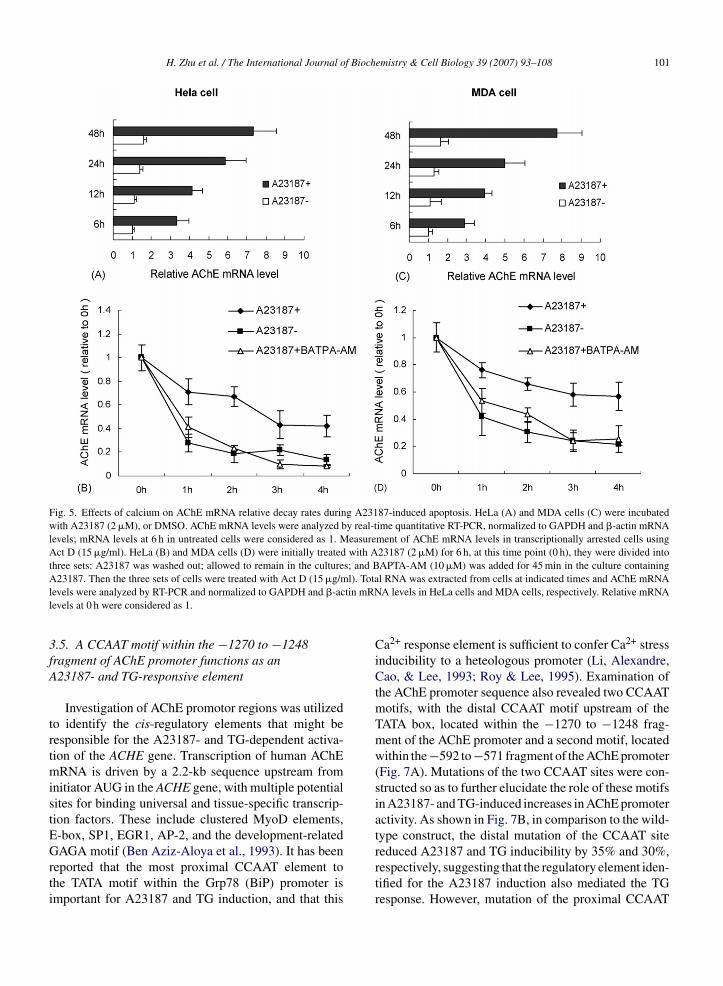
AChE mRNA levels increased dramatically in23187 treated HeLa and MDA cells compared withntreated controls at indicated time points (Fig. 5And C). The small increase in AChE mRNA levels in
ig. 3. Effects of intracellular Ca2+ on AChE expression during A23187-induere normalized to GAPDH and �-actin mRNA levels in HeLa (A) and MDA23187 (2 �M, 6 h) and (cross-hatched bars) pre-incubated with BAPTA-AM
ignificant difference relative to cells incubated with A23187. (B) and (D) ACith DMSO, A23187 (2 �M) or pre-incubated with BAPTA-AM (10 �M, 45
emistry & Cell Biology 39 (2007) 93–108 99
untreated cells may have resulted from apoptosis, whichoccurred in a few cells, with time. We further investi-gated the effects of A23187 on AChE mRNA stabilityso as to clarify whether elevated AChE mRNA levelsduring apoptosis resulted from an increase in the rate ofAChE transcription or an increase in the AChE mRNAstability, or both. To do so, we employed a classicaltechnique involving the measurement of AChE mRNArelative decay rates in transcriptionally arrested cells byusing actinomycin D (Act D) in the absence or pres-ence of A23187. The effect of BAPTA-AM on AChEmRNA decay rates was also examined. GAPDH and�-actin mRNA levels were used as internal controls inHeLa and MDA cells, respectively. Since there is a lowlevel of AChE mRNA in untreated cells, it is difficultto analyze AChE mRNA decay rates in the presence ofAct D without A23187 pre-treatment. HeLa and MDAcells were first treated with A23187 (2 �M) for 6 h. Asshown in Fig. 5B and D, the removal of A23187 as wellas the addition of BAPTA-AM with A23187 resulted in
an increase in the decay rate of AChE mRNA levels,suggesting that AChE mRNA stability was enhanced byA23187 and that the post-transcriptional mRNA stabil-ity is an important mechanism accounting for elevationced apoptosis. Real-time quantitative RT-PCR of AChE mRNA levelscells (C), respectively. (Open bars) DMSO treatment (6 h), (filled bars)(10 �M, 45 min) then incubated with A23187 (2 �M, 6 h). **p < 0.005hE protein levels were analyzed by Western-blot after 12-h treatmentmin) prior to A23187 (2 �M).
f Bioch
100 H. Zhu et al. / The International Journal oof AChE expression during A23187-induced apoptosisin HeLa and MDA cells. Our data also suggested thatcalcium also played a role in this event.
3.4. Effect of calcium on AChE promoter activityduring A23187 and TG-induced apoptosis in HeLaand MDA cells
We subsequently investigated the effects of A23187and TG on the transcription of the AChE genomicpromoter-reporter gene. The 2.2 kb human AChE pro-moter fused to the firefly luciferase reporter gene,pAChE-Luc and internal control Renilla luciferasereporter plasmid pRL-SV40 were transiently cotrans-fected to HeLa cells and treated with or without A23187(2 �M) and TG (2 �M) for indicated times. Cells trans-fected with promoterless control plasmids showed nodetectable luciferase activity (data not shown). Therewas a significant time-dependant increase in the induc-tion of luciferase activity in treated cells compared tocontrol (Fig. 6A), indicating that A23187 and TG acti-vated AChE promoter activity. Although an approxi-
mate three-fold induction of AChE promoter activitywas seen at 48 h after treatment, at 6 h after A23187-induced apoptosis, the promoter activity increase wasmodest in HeLa cells, whereas, the AChE mRNA levelFig. 4. Effects of intracellular Ca2+ on AChE expression during TG-induced awere incubated with TG (0.5 �M) for 6 h. AChE mRNA levels were analyzrelative to cells incubated with TG (also referred to in Fig. 3A and C). Westernand MDA cells incubated with TG (0.5 �M) for 6 h (also referred to in Fig. 3
emistry & Cell Biology 39 (2007) 93–108
increased significantly. This may have been due to thelimitation that the truncated portion of the gene pro-moter, in conjunction with additional regulatory ele-ments, did not reveal all behaviors of the endogenousgene transcription. The more obvious increase in AChEpromoter activity induced by TG may also have been dueto additional regulatory mechanisms. To test whetherAChE promoter activation by A23187 and TG wasaffected by changes in cytosolic Ca2+ concentrations,HeLa cells were pre-loaded with BAPTA-AM (10 �M,45 min) followed by incubation with A23187 or TG for24 h. BAPTA-AM alone had minimal effects on AChEpromoter activity (data not shown) while BAPTA-AMpre-treatment resulted in an approximate 30% reductionin A23187- and TG-dependent activation of AChE pro-moter (Fig. 6B).
A one- to five-fold induction of AChE promoter activ-ity in MDA cells, 6–48 h post-treatment, was induced byA23187 (2 �M) and TG (0.5 �M), relative to untreatedcontrols, with an even greater increase noted in HeLacells (Fig. 6C). Similar to what was noted in other assays,A23187- and TG-dependent activation of the AChE pro-
moter was also reduced in BAPTA-AM pre-treated cells(Fig. 6D), suggesting that the cytosolic Ca2+ signalingpathway may also have been involved in the transcrip-tional activity of AChE during apoptosis.poptosis. HeLa cells were incubated with TG (2 �M), and MDA cellsed by real-time quantitative RT-PCR. *p < 0.05 significant difference
blot of AChE protein levels in HeLa cells incubated with TG (2 �M)B and D).
H. Zhu et al. / The International Journal of Biochemistry & Cell Biology 39 (2007) 93–108 101
Fig. 5. Effects of calcium on AChE mRNA relative decay rates during A23187-induced apoptosis. HeLa (A) and MDA cells (C) were incubatedwith A23187 (2 �M), or DMSO. AChE mRNA levels were analyzed by real-time quantitative RT-PCR, normalized to GAPDH and �-actin mRNAlevels; mRNA levels at 6 h in untreated cells were considered as 1. Measurement of AChE mRNA levels in transcriptionally arrested cells usingAct D (15 �g/ml). HeLa (B) and MDA cells (D) were initially treated with A23187 (2 �M) for 6 h, at this time point (0 h), they were divided intothree sets: A23187 was washed out; allowed to remain in the cultures; and BAPTA-AM (10 �M) was added for 45 min in the culture containingA l). Total tin mRNl
3fA
trtmistEGrti
23187. Then the three sets of cells were treated with Act D (15 �g/mevels were analyzed by RT-PCR and normalized to GAPDH and �-acevels at 0 h were considered as 1.
.5. A CCAAT motif within the −1270 to −1248ragment of AChE promoter functions as an23187- and TG-responsive element
Investigation of AChE promotor regions was utilizedo identify the cis-regulatory elements that might beesponsible for the A23187- and TG-dependent activa-ion of the ACHE gene. Transcription of human AChE
RNA is driven by a 2.2-kb sequence upstream fromnitiator AUG in the ACHE gene, with multiple potentialites for binding universal and tissue-specific transcrip-ion factors. These include clustered MyoD elements,-box, SP1, EGR1, AP-2, and the development-related
AGA motif (Ben Aziz-Aloya et al., 1993). It has beeneported that the most proximal CCAAT element tohe TATA motif within the Grp78 (BiP) promoter ismportant for A23187 and TG induction, and that this
l RNA was extracted from cells at indicated times and AChE mRNAA levels in HeLa cells and MDA cells, respectively. Relative mRNA
Ca2+ response element is sufficient to confer Ca2+ stressinducibility to a heteologous promoter (Li, Alexandre,Cao, & Lee, 1993; Roy & Lee, 1995). Examination ofthe AChE promoter sequence also revealed two CCAATmotifs, with the distal CCAAT motif upstream of theTATA box, located within the −1270 to −1248 frag-ment of the AChE promoter and a second motif, locatedwithin the−592 to−571 fragment of the AChE promoter(Fig. 7A). Mutations of the two CCAAT sites were con-structed so as to further elucidate the role of these motifsin A23187- and TG-induced increases in AChE promoteractivity. As shown in Fig. 7B, in comparison to the wild-type construct, the distal mutation of the CCAAT site
reduced A23187 and TG inducibility by 35% and 30%,respectively, suggesting that the regulatory element iden-tified for the A23187 induction also mediated the TGresponse. However, mutation of the proximal CCAAT
102 H. Zhu et al. / The International Journal of Biochemistry & Cell Biology 39 (2007) 93–108
Fig. 6. Effects of calcium on AChE promoter activity during A23187- and TG-induced apoptosis. HeLa and MDA cells were cotransfected withpAChE-Luc and pRL-SV40. HeLa cell transfectants (A) were incubated with DMSO (un-induced control) or induced by A23187 (2 �M) or TG(2 �M). MDA cells transfectants (C) were incubated with DMSO (un-induced control) or induced by A23187 (2 �M) or TG (0.5 �M). HeLacells transfectants (B) were incubated with DMSO (un-induced control), A23187 (2 �M) and TG (2 �M), or A23187 (2 �M) and TG (2 �M) plus
bated w. AChEant diff
BAPTA-AM (10 �M) for 24 h; MDA cells transfectants (D) were incuA23187 (2 �M) and TG (0.5 �M) plus BAPTA-AM (10 �M) for 24 has the fold induction relative to un-induced controls. *p < 0.05 signific
binding site did not reduced A23187 or TG inducibilityas dramatically. Similar responses were seen in MDAcells (Fig. 7C) suggesting that the distal CCAAT elementupstream of the TATA box was important for A23187-and TG-induced AChE promoter activation. Given thatA23187 and TG are known to induce perturbations incellular Ca2+ homeostasis, our data further suggestedthat the distal CCAAT binding motif was involved inCa2+ signal transduction. It is relevant to note that resid-ual levels of A23187 and TG induction were observedwith mutations in the distal CCAAT, suggesting otherelements in the AChE promoter compensated for thefunction of the mutated elements in A23187 and TGresponses.
3.6. Identification of the CCAAT binding factor(CBF/NF-Y) binding to the distal CCAAT element
In order to investigate which binding proteins wereable to bind to the two CCAAT motifs, distal andthe proximal CCAAT site probes (Wt CCAAT Dist
ith DMSO (un-induced control), A23187 (2 �M) and TG (0.5 �M), orpromoter activity was analyzed by luciferase activity and representederence relative to cells incubated with A23187 or TG alone.
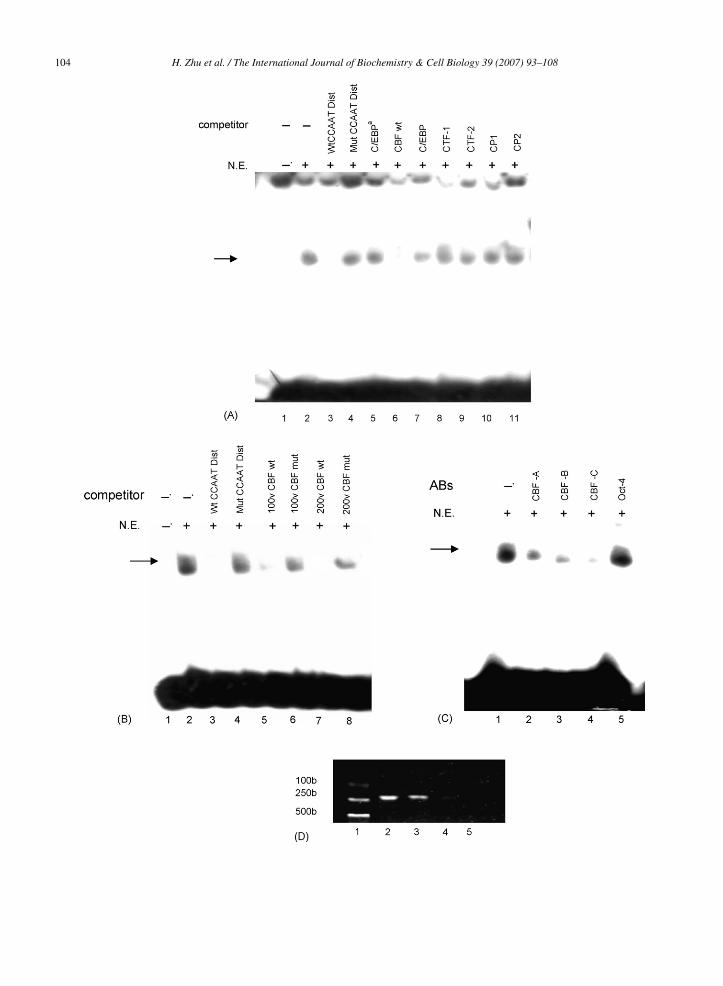
and Wt CCAAT Prox) were used in EMSAs togetherwith nuclear extracts from HeLa cells. The Wt CCAATDist probe was found to form DNA–protein complex(Fig. 8A, lane 2), whereas, Wt CCAAT Prox probe didnot bind to any protein (data not shown). As shownin Fig. 8A, an excess of unlabeled probe that incorpo-rated mutation in the distal CCAAT binding site didnot inhibit the complex formation (Fig. 8A, lane 4).However, an excess of unlabeled probe, specific forthe distal CCAAT site, competed for complex forma-tion with the labeled Wt CCAAT Dist probe (Fig. 8A,lane 3). Given that the distal CCAAT motif found inthe human ACHE gene promoter region could poten-tially represent a binding site for a number of nuclearfactors, including C/EBP�, C/EBP, CTF1, CTF2, CP1,CP2, and CBF/NF-Y, the binding of these nuclear factorswas tested in a competitive EMSA. Our data showed that
an excess of the consensus oligonucleotides of CBF/NF-Y (CBF wt) inhibited the binding of the labeled WtCCAAT Dist probe (Fig. 8A, lane 6), whereas, C/EBP�,C/EBP, CTF1, CTF2, CP1, and CP2 consensus oligonu-
H. Zhu et al. / The International Journal of Biochemistry & Cell Biology 39 (2007) 93–108 103
Fig. 7. A CCAAT motif within the −1270 to −1248 fragment of the AChE promoter is an A23187- and TG-responsive element. (A) The twoCCAAT mutants from the human AChE promoter were constructed as described in Section 2. Wild type and mutated human AChE promoters weretagged with reporter luciferase, pAChE-Luc, Distal CCAAT binding mutant, and Proximal CCAAT binding mutant, respectively. HeLa and MDAcells were cotransfected with pAChE-Luc or Distal CCAAT binding mutant or Proximal CCAAT binding mutant, and pRL-SV40. Transfectantsof HeLa cells (B) were incubated with DMSO (un-induced control) or induced by A23187 (2 �M) or TG (2 �M) for 24 h. Transfectants of MDAc d by A2a on relatc
c5sbmmCDiasoa(A5
ca
ells (C) were incubated with DMSO (un-induced control) or inducenalyzed by the luciferase activity and represented as the fold inductiotransfected with pAChE-Luc and pRL-SV40.
leotides did not abolish binding activity (Fig. 8A, lane, and lanes 7–11). A 100- and 200-fold excess of con-ensus oligonucleotides of CBF wt almost abolished theinding, whereas, CBF/NF-Y consensus mutant (CBFut) could not (Fig. 8B). CBF/NF-Y is a complexinimally composed of three subunits: CBF-A/NF-YB,BF-B/NF-YA, and CBF-C/NF-YC, all required forNA-binding (Mantovani, 1999). To further character-
ze the binding proteins, we performed an interferencessay using specific antibodies against CBF/NF-Y. Ashown in Fig. 8C, the DNA–protein complexes werenly inhibited obviously by anti-CBF antibodies, namelynti-CBF-A/NF-YB (Fig. 8C, lane 2), CBF-B/NF-YAFig. 8C, lane 3), and CBF-C/NF-YC (Fig. 8C, lane 4).dditionally, specific antibodies to Oct-4 (Fig. 8C, lane
) failed to inhibit the binding.We subsequently performed chromatin immunopre-ipitation analysis to examine whether CBF/NF-Y wasble to bind to the AChE promoter in vivo. Protein–DNA
3187 (2 �M) or TG (0.5 �M) for 24 h. AChE promoter activity wasive to un-induced controls. *p < 0.05 significant difference from cells
complexes were immunoprecipitated with antibodiesand the recovered DNA fragments, including the distalCCAAT motif, were monitored by PCR using primers forthe −1312 to −1111 region of the AChE promoter. DNAfragments immunoprecipitated with polyclonal antibod-ies against CBF/NF-Y were amplified by PCR (Fig. 8D,lane 3) as well as the positive control (Fig. 8D, lane 2),whereas, those immunoprecipitated with anti-Oct 4 anti-body (Fig. 8D, lane 4) or without antibody (Fig. 8D, lane5) were not. These results indicated that CBF/NF-Y wasable to bind directly to the distal CCAAT motif in theAChE promoter.
4. Discussion
In our previous study, we reported the novel find-ing that apart from its catalytic function in hydrolyzingacetylcholine, AChE expression was induced in vari-ous cell types upon treatment with different apoptosis
104 H. Zhu et al. / The International Journal of Biochemistry & Cell Biology 39 (2007) 93–108

f Bioch
ioaastemprsOwnscT
rbcporelhom&rpdirgPim
Fcc9wCavioCca
H. Zhu et al. / The International Journal o
nducers (Zhang et al., 2002). Blocking the expressionf AChE with antisense inhibited apoptosis (Zhang etl., 2002), and silencing of the ACHE gene by siRNAbolished the decrease in cell viability (Park et al., 2004),uggesting that AChE could be a novel regulator of apop-osis. Elucidating the regulatory mechanisms of AChExpression during apoptosis may improve therapeuticethods to regulate apoptosis, an important factor in
rominent diseases, including cancers (with apoptosis-esistant tumor cells) and neurodegenerative disorders,uch as Alzheimer’s and Parkinson’s disease (Fadeel,rrenius, & Zhivotovsky, 1999). In the present study,e choose HeLa and MDA cells, two cell lines that doot express AChE strongly in normal states, as our modelystems. We found that intracellular Ca2+ played a criti-al role in the expression of AChE during A23187- andG-induced apoptosis.
We further examined the mechanisms that up-egulated AChE mRNA levels during apoptosis inducedy A23187. A23187 can cause a sustained high intra-ellular free Ca2+ level. Our results indicated that aost-transcriptional mechanism, operating at the levelf transcript stability, appeared to be an importantegulatory step involved in the regulation of AChExpression induced by A23187, and that this regu-atory processes was mediated by cytosolic Ca2+. Itas previously been demonstrated that the regulationf mRNA stability plays a crucial role in the basicechanisms underlying gene expression (Guhaniyogi
Brewer, 2001; Mitchell & Tollervey, 2000). Thisegulatory mechanism involves not only distinct cyto-lasmic factors, but also the contribution of sequenceeterminants located both within and outside the cod-ng regions of mature transcripts, such as AUUUA-ich elements (AREs), identified by their ability to tar-
et host mRNAs towards rapid degradation (Barreau,aillard, & Osborne, 2005). Various exogenous factors,ncluding calcium ionophores, modulate ARE-directedRNA degradation (Ruth, Esnault, Jarzembowski, &
ig. 8. Identification of the CCAAT binding factor (CBF/NF-Y) able to bindonsensus oligonucleotides as competitors. 32P-labeled Wt CCAAT Dist waompetitors, Wt CCAAT Dist (lane 3), Mut CCAAT Dist (lane 4), C/EBPα (l), CP1 (lane 10) and CP2 (lane 11). (B) Interference assays using CBF conas incubated with nuclear extracts from HeLa cells in the presence of comBF wt (lane 5), 100v CBF mut (lane 6), 200v CBF wt (lane 7) and 200vgainst CBF/NF-YC transcription factors. 32P-labeled Wt CCAAT Dist wasarious antibodies, anti-CBF-A/NF-YB (lane 2), anti-CBF-B/NF-YA (lane 3)mmunoprecipitation analysis was performed to monitor the binding of CBFf the AChE promoter, in vivo. Protein–DNA complexes were incubated witBF-C/NF-YC, and isolated by immunoprecipitation (lane 3). Positive controntrols were isolated by immunoprecipitation with Oct 4 IgG (lane 4) or witnalyzed by PCR with the indicated primers, and DNA marker was used as a
emistry & Cell Biology 39 (2007) 93–108 105
Malter, 1999). Interestingly an AUUUA-element hasbeen located in the 3′ untranslated region (UTR) ofAChE mRNA. It has been recently reported that theRNA-binding protein HuD regulated AChE expression,at a post-transcriptional level, by binding directly to theAChE 3′ UTR, containing an ARE element, in nervegrowth factor-treated PC12 cells (Deschenes-Furry et al.,2003). Taylor and colleagues proposed that the ACHEgene was ubiquitously transcribed, and that the tran-script was readily degraded in most tissues, whereas,it was specifically stabilized in the tissues where tran-scripts could be detected (Mutero et al., 1995). The risein cytosolic Ca2+ during A23187-induced apoptosis inHeLa and MDA cells may directly or indirectly blockdestabilizing factors or activate stabilizing factors crit-ical for AChE mRNA stabilization. Further research isneeded to fully elucidate this question.
Our results also demonstrated there was a Ca2+-dependent increase in AChE promoter activity dur-ing A23187 and TG-induced apoptosis, indicating thatACHE gene expression was modulated at the transcrip-tional level, with the regulation of AChE promoter activ-ity facilitated by Ca2+-dependent signaling pathways. Inorder to examine the transcriptional regulation of theACHE gene, we focused on a CCAAT motif withinthe −1270 to −1248 fragment of the human AChEpromoter. A23187 and TG are known to disturb Ca2+
homeostasis. The mutation of distal CCAAT bindingsite significantly decreased A23187 and TG-inducedAChE promoter activation, suggesting that the Ca2+ sig-nal transduction was mediated, at least in part, throughthe CCAAT motif. The CCAAT motif is a commonpromoter element present in the proximal promoter ofnumerous mammalian genes transcribed by RNA poly-merase II (Maity & de Crombrugghe, 1998). Mutations
in the CCAAT motifs of several gene promoters altertranscriptional regulation by various inducing agents(Maity & de Crombrugghe, 1998). Li et al. have reportedthat the stimulation of grp78 transcription by A23187to the distal CCAAT element. (A) Interference assays using variouss incubated with nuclear extracts from HeLa cells in the presence ofane 5), CBF wt (lane 6), C/EBP (lane 7), CTF1 (lane 8), CTF2 (lanesensus oligonucleotides as competitors. 32P-labeled Wt CCAAT Distpetitors, Wt CCAAT Dist (lane 3), Mut CCAAT Dist (lane 4), 100vCBF mut (lane 8). (C) Interference assays using specific antibodiesincubated with nuclear extracts from HeLa cells in the presence of
, anti-CBF-C/NF-YC (lane 4) and Oct 4 IgG (lane 5). (D) Chromatin/NF-Y to the ATTGG region within the −1270 to −1248 fragment
h polyclonal antibodies against CBF-A/NF-YB, CBF-B/NF-YA, andols were prepared before immunoprecipitation (lane 2) and negativehout antibody (lane 5). All immunoprecipitated DNA fragments weremolecular weight marker (lane 1).

f Bioch
106 H. Zhu et al. / The International Journal oand TG, following ER Ca2+ discharge, acted througha novel pathway in which a Ca2+ signal was transducedthrough the most proximal CCAAT element to the TATAmotif, achieving specific gene regulatory events (Li etal., 1993). Our result may give another example thatCa2+ signal is transduced through elements containingCCAAT motifs within the promoter regions.
This is, to our knowledge, the first demonstrationthat the CCAAT binding factor (CBF/NF-Y) may bea potential transcriptional factor mediating AChE tran-scriptional activity during apoptosis through a CCAATmotif. CBF/NF-Y was originally identified as a proteincomplex able to bind to the MHC class II conserved Ybox in Ea promoters, the yeast CYC1 U AS CCAATbox, and the α2(I) collagen gene in the proximal pro-moter region (Matsuo et al., 2003). CBF/NF-Y was notonly involved in high-basal-level transcriptional activa-tion (Matsuo et al., 2003), but also contributed to inducedtranscription of the grp78 gene under calcium stressconditions (Roy & Lee, 1995). Having established thatCBF/NF-Y was recruited to the CCAAT motif upstreamof the TATA sequence, within the AChE promoter, weare currently investigating the mechanism by which itregulates the A23187- and TG-induced transcription ofthe ACHE gene. It is possible that the interactions ofCBF/NF-Y with other transcription factors are effectedby A23187 and TG, or that the binding CBF/NF-Y tothe CCAAT motif is influenced by intracellular calciumconcentrations, similar to what has been reported in thegrp78 gene (Roy & Lee, 1995).
It has been suggested that neuronal death inAlzheimer’s disease (AD) is due to apoptosis (Li, Chan,Lai, & Yew, 1997; Su, Deng, & Cotman, 1997). Dysreg-ulation of intracellular calcium signaling also has beenimplicated in the pathogenesis of AD (LaFerla, 2002;Mattson & Chan, 2003). In vivo and in vitro studieshave demonstrated that A� destabilizes neuronal Ca2+
homeostasis, leading to a significant increase in cytoso-lic Ca2+ and triggering neuronal apoptosis (Goodman& Mattson, 1994; Mattson, Tomaselli, & Rydel, 1993).Increased AChE activity has been reported in plaquesand tangles early in the process of amyloid deposition ofAD (Moran, Mufson, & Gomez-Ramos, 1993; Ulrich,Meier-Ruge, Probst, Meier, & Ipsen, 1990), promotingthe assembly of A� into fibrils (Inestrosa, Alvarez, &Calderon, 1996) and increasing the cytotoxicity of thesepeptides (Alvarez et al., 1998). We believe that furtherinvestigation of AChE up-regulation, its targets and func-
tions during apoptosis, will further elucidate apoptosismechanism of action as well as facilitate the developmentof therapeutic approaches that selectively target AChEexpression pathways in the treatment of AD.emistry & Cell Biology 39 (2007) 93–108
Acknowledgements
We thank Dr. Hermona Soreq from The HebrewUniversity of Jerusalem, Israel, for providing humanAChE promoter construct. This work was supported bygrants 2005CB522602, 2002BA711A07, KSCX2-SW-211, NSFC30570920 and 02ZA14118, and ResearchGrants Council of Hong Kong (HKUST 6283/03M toKWKT).
References
Alvarez, A., Alarcon, R., Opazo, C., Campos, E. O., Munoz, F. J.,Calderon, F. H., et al. (1998). Stable complexes involving acetyl-cholinesterase and amyloid-beta peptide change the biochemi-cal properties of the enzyme and increase the neurotoxicity ofAlzheimer’s fibrils. J. Neurosci., 18, 3213–3223.
Angus, L. M., Chan, R. Y., & Jasmin, B. J. (2001). Role of intronicE- and N-box motifs in the transcriptional induction of theacetylcholinesterase gene during myogenic differentiation. J. Biol.Chem., 276, 17603–17609.
Azmi, S., Dhawan, D., & Singh, N. (1996). Calcium ionophore A23187 induces apoptotic cell death in rat thymocytes. Cancer Lett.,107, 97–103.
Barreau, C., Paillard, L., & Osborne, H. B. (2005). AU-rich elementsand associated factors: Are there unifying principles? Nucleic AcidsRes., 33, 7138–7150.
Bartlett, J. D., Luethy, J. D., Carlson, S. G., Sollott, S. J., & Holbrook,N. J. (1992). Calcium ionophore A23187 induces expression ofthe growth arrest and DNA damage inducible CCAAT/enhancer-binding protein (C/EBP)-related gene, gadd153. Ca2+ increasestranscriptional activity and mRNA stability. J. Biol. Chem., 267,20465–20470.
Ben Aziz-Aloya, R., Seidman, S., Timberg, R., Sternfeld, M., Zakut,H., & Soreq, H. (1993). Expression of a human acetylcholinesterasepromoter-reporter construct in developing neuromuscular junc-tions of Xenopus embryos. Proc. Natl. Acad. Sci. U.S.A., 90,2471–2475.
Chan, R. Y., Adatia, F. A., Krupa, A. M., & Jasmin, B. J. (1998).Increased expression of acetylcholinesterase T and R transcriptsduring hematopoietic differentiation is accompanied by parallelelevations in the levels of their respective molecular forms. J. Biol.Chem., 273, 9727–9733.
Chan, R. Y., Boudreau-Lariviere, C., Angus, L. M., Mankal, F. A., &Jasmin, B. J. (1999). An intronic enhancer containing an N-boxmotif is required for synapse- and tissue-specific expression ofthe acetylcholinesterase gene in skeletal muscle fibers. Proc. Natl.Acad. Sci. U.S.A., 96, 4627–4632.
Choi, R. C., Siow, N. L., Cheng, A. W., Ling, K. K., Tung, E. K.,Simon, J., et al. (2003). ATP acts via P2Y1 receptors to stimulateacetylcholinesterase and acetylcholine receptor expression: Trans-duction and transcription control. J. Neurosci., 23, 4445–4456.
Coleman, B. A., & Taylor, P. (1996). Regulation of acetylcholinesteraseexpression during neuronal differentiation. J. Biol. Chem., 271,4410–4416.
Day, T., & Greenfield, S. A. (2002). A non-cholinergic, trophic actionof acetylcholinesterase on hippocampal neurones in vitro: Molec-ular mechanisms. Neuroscience, 111, 649–656.
Demaurex, N., & Distelhorst, C. (2003). Cell biology. Apoptosis—Thecalcium connection. Science, 300, 65–67.
zhang xue jun
Highlight

f Bioch
D
D
E
E
F
G
G
G
H
I
J
J
K
K
K
K
K
L
H. Zhu et al. / The International Journal o
eschenes-Furry, J., Belanger, G., Perrone-Bizzozero, N., & Jasmin,B. J. (2003). Post-transcriptional regulation of acetylcholinesterasemRNAs in nerve growth factor-treated PC12 cells by the RNA-binding protein HuD. J. Biol. Chem., 278, 5710–5717.
ignam, J. D., Martin, P. L., Shastry, B. S., & Roeder, R. G. (1983).Eukaryotic gene transcription with purified components. Meth.Enzymol., 101, 582–598.
kstrom, T. J., Klump, W. M., Getman, D., Karin, M., & Taylor, P.(1993). Promoter elements and transcriptional regulation of theacetylcholinesterase gene. DNA Cell Biol., 12, 63–72.
lizondo, G., Corchero, J., Sterneck, E., & Gonzalez, F. J. (2000).Feedback inhibition of the retinaldehyde dehydrogenase geneALDH1 by retinoic acid through retinoic acid receptor alphaand CCAAT/enhancer-binding protein beta. J. Biol. Chem., 275,39747–39753.
adeel, B., Orrenius, S., & Zhivotovsky, B. (1999). Apoptosis in humandisease: A new skin for the old ceremony? Biochem. Biophys. Res.Commun., 266, 699–717.
oodman, Y., & Mattson, M. P. (1994). Secreted forms of beta-amyloid precursor protein protect hippocampal neurons againstamyloid beta-peptide-induced oxidative injury. Exp. Neurol., 128,1–12.
roenendyk, J., Lynch, J., & Michalak, M. (2004). Calreticulin, Ca2+,and calcineurin—Signaling from the endoplasmic reticulum. Mol.Cells, 17, 383–389.
uhaniyogi, J., & Brewer, G. (2001). Regulation of mRNA stabilityin mammalian cells. Gene, 265, 11–23.
e, Q., Lee, D. I., Rong, R., Yu, M., Luo, X., Klein, M., et al. (2002).Endoplasmic reticulum calcium pool depletion-induced apoptosisis coupled with activation of the death receptor 5 pathway. Onco-gene, 21, 2623–2633.
nestrosa, N. C., Alvarez, A., & Calderon, F. (1996). Acetyl-cholinesterase is a senile plaque component that promotes assem-bly of amyloid beta-peptide into Alzheimer’s filaments. Mol. Psy-chiatry, 1, 359–361.
ackisch, C., Hahm, H. A., Tombal, B., McCloskey, D., Butash,K., Davidson, N. E., et al. (2000). Delayed micromolar eleva-tion in intracellular calcium precedes induction of apoptosis inthapsigargin-treated breast cancer cells. Clin. Cancer. Res., 6,2844–2850.
ohnson, G., & Moore, S. W. (2004). Identification of a structural siteon acetylcholinesterase that promotes neurite outgrowth and bindslaminin-1 and collagen IV. Biochem. Biophys. Res. Commun., 319,448–455.
aneko, Y., & Tsukamoto, A. (1994). Thapsigargin-induced persis-tent intracellular calcium pool depletion and apoptosis in humanhepatoma cells. Cancer Lett., 79, 147–155.
arpel, R., Sternfeld, M., Ginzberg, D., Guhl, E., Graessmann, A.,& Soreq, H. (1996). Overexpression of alternative human acetyl-cholinesterase forms modulates process extensions in culturedglioma cells. J. Neurochem., 66, 114–123.
aufer, D., Friedman, A., Seidman, S., & Soreq, H. (1998). Acutestress facilitates long-lasting changes in cholinergic gene expres-sion. Nature, 393, 373–377.
oenigsberger, C., Chiappa, S., & Brimijoin, S. (1997). Neuritedifferentiation is modulated in neuroblastoma cells engineeredfor altered acetylcholinesterase expression. J. Neurochem., 69,
1389–1397.rebs, J. (1998). The role of calcium in apoptosis. Biometals, 11,375–382.
aFerla, F. M. (2002). Calcium dyshomeostasis and intracellular sig-naling in Alzheimer’s disease. Nat. Rev. Neurosci., 3, 862–872.
emistry & Cell Biology 39 (2007) 93–108 107
Li, W. P., Chan, W. Y., Lai, H. W., & Yew, D. T. (1997). TerminaldUTP nick end labeling (TUNEL) positive cells in the differentregions of the brain in normal aging and Alzheimer patients. J.Mol. Neurosci., 8, 75–82.
Li, W. W., Alexandre, S., Cao, X., & Lee, A. S. (1993). Transactivationof the grp78 promoter by Ca2+ depletion. A comparative analy-sis with A23187 and the endoplasmic reticulum Ca(2+)-ATPaseinhibitor thapsigargin. J. Biol. Chem., 268, 12003–12009.
Luo, Z., Fuentes, M. E., & Taylor, P. (1994). Regulation of acetyl-cholinesterase mRNA stability by calcium during differentiationfrom myoblasts to myotubes. J. Biol. Chem., 269, 27216–27223.
Maity, S. N., & de Crombrugghe, B. (1998). Role of the CCAAT-binding protein CBF/NF-Y in transcription. Trends Biochem. Sci.,23, 174–178.
Mantovani, R. (1999). The molecular biology of the CCAAT-bindingfactor NF-Y. Gene, 239, 15–27.
Massoulie, J., Anselmet, A., Bon, S., Krejci, E., Legay, C., Morel, N.,et al. (1998). Acetylcholinesterase: C-terminal domains, molec-ular forms and functional localization. J. Physiol. Paris, 92,183–190.
Matsuo, N., Yu-Hua, W., Sumiyoshi, H., Sakata-Takatani, K., Nagato,H., Sakai, K., et al. (2003). The transcription factor CCAAT-binding factor CBF/NF-Y regulates the proximal promoter activityin the human alpha 1(XI) collagen gene (COL11A1). J. Biol.Chem., 278, 32763–32770.
Mattson, M. P., & Chan, S. L. (2003). Calcium orchestrates apoptosis.Nat. Cell Biol., 5, 1041–1043.
Mattson, M. P., Tomaselli, K. J., & Rydel, R. E. (1993). Calcium-destabilizing and neurodegenerative effects of aggregated beta-amyloid peptide are attenuated by basic FGF. Brain Res., 621,35–49.
McColl, K. S., He, H., Zhong, H., Whitacre, C. M., Berger, N. A., &Distelhorst, C. W. (1998). Apoptosis induction by the glucocor-ticoid hormone dexamethasone and the calcium-ATPase inhibitorthapsigargin involves Bc1-2 regulated caspase activation. Mol. CellEndocrinol., 139, 229–238.
Melo, J. B., Agostinho, P., & Oliveira, C. R. (2003). Involvement ofoxidative stress in the enhancement of acetylcholinesterase activityinduced by amyloid beta-peptide. Neurosci. Res., 45, 117–127.
Mitchell, P., & Tollervey, D. (2000). mRNA stability in eukaryotes.Curr. Opin. Genet. Dev., 10, 193–198.
Moran, M. A., Mufson, E. J., & Gomez-Ramos, P. (1993). Colocal-ization of cholinesterases with beta amyloid protein in aged andAlzheimer’s brains. Acta Neuropathol. (Berl.), 85, 362–369.
Muallem, S., Khademazad, M., & Sachs, G. (1990). The route of Ca2+
entry during reloading of the intracellular Ca2+ pool in pancreaticacini. J. Biol. Chem., 265, 2011–2016.
Mutero, A., Camp, S., & Taylor, P. (1995). Promoter elements of themouse acetylcholinesterase gene. Transcriptional regulation duringmuscle differentiation. J. Biol. Chem., 270, 1866–1872.
Park, S. E., Kim, N. D., & Yoo, Y. H. (2004). Acetylcholinesteraseplays a pivotal role in apoptosome formation. Cancer Res., 64,2652–2655.
Petersen, A., Castilho, R. F., Hansson, O., Wieloch, T., & Brundin,P. (2000). Oxidative stress, mitochondrial permeability transitionand activation of caspases in calcium ionophore A23187-induceddeath of cultured striatal neurons. Brain Res., 857, 20–29.
Rizzuto, R., Pinton, P., Ferrari, D., Chami, M., Szabadkai, G., Magal-haes, P. J., et al. (2003). Calcium and apoptosis: Facts and hypothe-ses. Oncogene, 22, 8619–8627.
Roy, B., & Lee, A. S. (1995). Transduction of calcium stress throughinteraction of the human transcription factor CBF with the proximal

f Bioch
108 H. Zhu et al. / The International Journal oCCAAT regulatory element of the grp78/BiP promoter. Mol. CellBiol., 15, 2263–2274.
Ruth, J. H., Esnault, S., Jarzembowski, J. A., & Malter, J. S. (1999). Cal-cium ionophore upregulation of AUUUA-specific binding proteinactivity is contemporaneous with granulocyte macrophage colony-stimulating factor messenger RNA stabilization in AML14.3D10cells. Am. J. Respir. Cell Mol. Biol., 21, 621–628.
Sagara, Y., & Inesi, G. (1991). Inhibition of the sarcoplasmic reticulumCa2+ transport ATPase by thapsigargin at subnanomolar concen-trations. J. Biol. Chem., 266, 13503–13506.
Sberna, G., Saez-Valero, J., Beyreuther, K., Masters, C. L., & Small,D. H. (1997). The amyloid beta-protein of Alzheimer’s diseaseincreases acetylcholinesterase expression by increasing intracellu-lar calcium in embryonal carcinoma P19 cells. J. Neurochem., 69,1177–1184.
Shohami, E., Kaufer, D., Chen, Y., Seidman, S., Cohen, O., Ginzberg,D., et al. (2000). Antisense prevention of neuronal damages fol-lowing head injury in mice. J. Mol. Med., 78, 228–236.
Siow, N. L., Choi, R. C., Cheng, A. W., Jiang, J. X., Wan, D. C., Zhu, S.Q., et al. (2002). A cyclic AMP-dependent pathway regulates theexpression of acetylcholinesterase during myogenic differentiationof C2C12 cells. J. Biol. Chem., 277, 36129–36136.
Soreq, H., & Seidman, S. (2001). Acetylcholinesterase—new roles for
an old actor. Nat. Rev. Neurosci., 2, 294–302.Su, J. H., Deng, G., & Cotman, C. W. (1997). Bax protein expressionis increased in Alzheimer’s brain: Correlations with DNA dam-age, Bcl-2 expression, and brain pathology. J. Neuropathol. Exp.Neurol., 56, 86–93.
emistry & Cell Biology 39 (2007) 93–108
Taylor, P., & Radic, Z. (1994). The cholinesterases: From genes toproteins. Annu. Rev. Pharmacol. Toxicol., 34, 281–320.
Tung, E. K., Choi, R. C., Siow, N. L., Jiang, J. X., Ling, K.K., Simon, J., et al. (2004). P2Y2 receptor activation regulatesthe expression of acetylcholinesterase and acetylcholine receptorgenes at vertebrate neuromuscular junctions. Mol. Pharmacol., 66,794–806.
Ulrich, J., Meier-Ruge, W., Probst, A., Meier, E., & Ipsen, S. (1990).Senile plaques: Staining for acetylcholinesterase and A4 protein:A comparative study in the hippocampus and entorhinal cortex.Acta Neuropathol. (Berl.), 80, 624–628.
Wan, D. C., Choi, R. C., Siow, N. L., & Tsim, K. W. (2000). Thepromoter of human acetylcholinesterase is activated by a cyclicadenosine 3′,5′-monophosphate-dependent pathway in culturedNG108-15 neuroblastoma cells. Neurosci. Lett., 288, 81–85.
Waser, M., Mesaeli, N., Spencer, C., & Michalak, M. (1997). Regula-tion of calreticulin gene expression by calcium. J. Cell Biol., 138,547–557.
Whyte, K. A., & Greenfield, S. A. (2003). Effects of acetyl-cholinesterase and butyrylcholinesterase on cell survival, neuriteoutgrowth, and voltage-dependent calcium currents of embryonicventral mesencephalic neurons. Exp. Neurol., 184, 496–509.
Yang, L., He, H. Y., & Zhang, X. J. (2002). Increased expression of
intranuclear AChE involved in apoptosis of SK-N-SH cells. Neu-rosci. Res., 42, 261–268.Zhang, X. J., Yang, L., Zhao, Q., Caen, J. P., He, H. Y., Jin, Q. H.,et al. (2002). Induction of acetylcholinesterase expression duringapoptosis in various cell types. Cell Death Differ., 9, 790–800.




















