
Reelin and mDab1 regulate the development of hippocampal connections
16
Reelin and mDab1 regulate the development of hippocampal connections Victor Borrell, a,b,1,2 Lluís Pujadas, a,b,1 Sergi Simó, a,b,c David Durà, a,b Marta Solé, a,b Jonathan A. Cooper, c Jose A. Del Río, a,b and Eduardo Soriano a,b, ⁎ a Developmental Neurobiology and Regeneration Laboratory, Institute for Research in Biomedicine (IRB)-Barcelona Science Park, Barcelona, E-08028, Spain b Department of Cell Biology and Centro de Investigación Biomédica en Red sobre Enfermedades Neurodegenerativas (CIBERNED), University of Barcelona, Barcelona, E-08028, Spain c Basic Science Division, Fred Hutchinson Cancer Research Center, Seattle, WA 98109, USA Received 8 January 2007; revised 15 June 2007; accepted 16 June 2007 Available online 18 July 2007 We analyze in this study the participation of Reelin and mDab1 in the development of hippocampal connections. We show that mDab1 is present in growth cones and axonal tracts of developing hippocampal afferents. mdab1-deficiency produces severe alterations in the entorhino- hippocampal and commissural connections identical to those described in reeler mice, including innervation of ectopic areas, formation of abnormal patches of fiber termination and a delay in the refinement of projections. Organotypic slice cultures combining tissue from mdab1- mutant and control mice demonstrate that the abnormalities observed in the mutant entorhino-hippocampal projection are caused by mdab1- deficiency in both the projecting neurons and target hippocampal cells. Axonal afferents that innervate the hippocampus react to Reelin by reducing axonal growth, and increasing growth cone collapse and axonal branching. Altogether these results indicate that Reelin and mDab1 participate in the development and refinement of hippocampal connec- tions by regulating axonal extension, targeting and branching. © 2007 Elsevier Inc. All rights reserved. Keywords: mDab1; Reelin; Axonal outgrowth Introduction The correct functioning of the CNS requires both the proper positioning of neurons within layers or nuclei, and the formation of appropriate neuronal connections. The development of axonal pathways depends on the action of guidance molecules on growth cones. Numerous diffusible and membrane-bound molecules play a central role in the guidance and development of neural connections (Mueller, 1999; Chilton, 2006; Huber et al., 2003). Some molecules, such as Netrin-1, participate in both neuronal migration and axonal guidance (Barallobre et al., 2005). The hippocampus (HP) is a laminated structure that receives highly ordered extrinsic and intrinsic inputs, where the different afferent systems terminate in non-overlapping layers (Super and Soriano, 1994). In previous studies we have shown that Cajal–Retzius (CR) cells play a role in the development of hippocampal connections (Borrell et al., 1999b; Del Rio et al., 1997; Super et al., 1998). CR cells, first identified as a special class of pioneer neurons in the marginal zone of the neocortex, are also present in the marginal zone of the HP prior to and during the ingrowth of hippocampal afferents (Del Rio et al., 1996, 1997; Marin-Padilla, 1998; Soriano and Del Rio, 2005; Soriano et al., 1994; Super and Soriano, 1994). CR cells express Reelin, a large extracellular molecule that is critical for lamination of several brain regions and that, when mutated, leads to the reeler phenotype (D'Arcangelo et al., 1995; Rice and Curran, 2001; Tissir and Goffinet, 2003). Shortly after the discovery of Reelin, mDab1–the mouse homolog of Drosophila Disabled protein–was discovered. Mutation of mDab1 leads to migration disorders indistinguishable from those observed in reeler (Howell et al., 1997b, 1997a; Sheldon et al., 1997). mDab1 is an intra- cellular adaptor protein that is phosphorylated in response to Reelin signaling (Bar et al., 2003; Howell et al., 2000; Keshvara et al., 2001; Rice and Curran, 2001; Tissir and Goffinet, 2003). Other molecules shown to participate in the Reelin signaling pathway include the lipoprotein receptors VLDLR and ApoER2, Src family kinases, PI3K, Akt1, GSK3β and MAP1B (Gonzalez- www.elsevier.com/locate/ymcne Mol. Cell. Neurosci. 36 (2007) 158 – 173 Abbreviations: DG, dentate gyrus; EC, entorhinal cortex; f, fimbria; GL, granule cell layer; HF, hippocampal fissure; HP, hippocampus; IML, inner molecular layer; ML, molecular layer; MML, medial molecular layer; NC, neocortex; OML, outer molecular layer; SC, subicular complex; SLM, stratum lacunosum moleculare; SO, stratum oriens; SP, stratum pyramidale; SR, stratum radiatum; WM, white matter. ⁎ Corresponding author. Developmental Neurobiology and Regeneration Laboratory, Institute for Research in Biomedicine (IRB), Barcelona Science Park (S1-A1), Josep Samitier, 1-5, E-08028 Barcelona, Spain. Fax: +34 93 4037116. E-mail address: [email protected] (E. Soriano). 1 These authors contributed equally to this work. 2 Present address: Developmental Neurobiology Laboratory, Instituto de Neurosciencias, University Miguel Hernandez, San Joan d'Alacant, E-03552, Spain. Available online on ScienceDirect (www.sciencedirect.com). 1044-7431/$ - see front matter © 2007 Elsevier Inc. All rights reserved. doi:10.1016/j.mcn.2007.06.006
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Reelin and mDab1 regulate the development of hippocampal connections

www.elsevier.com/locate/ymcne
Mol. Cell. Neurosci. 36 (2007) 158–173Reelin and mDab1 regulate the developmentof hippocampal connections
Victor Borrell,a,b,1,2 Lluís Pujadas,a,b,1 Sergi Simó,a,b,c David Durà,a,b Marta Solé,a,b
Jonathan A. Cooper,c Jose A. Del Río,a,b and Eduardo Sorianoa,b,⁎
aDevelopmental Neurobiology and Regeneration Laboratory, Institute for Research in Biomedicine (IRB)-Barcelona Science Park, Barcelona, E-08028, SpainbDepartment of Cell Biology and Centro de Investigación Biomédica en Red sobre Enfermedades Neurodegenerativas (CIBERNED), University of Barcelona,Barcelona, E-08028, SpaincBasic Science Division, Fred Hutchinson Cancer Research Center, Seattle, WA 98109, USA
Received 8 January 2007; revised 15 June 2007; accepted 16 June 2007Available online 18 July 2007
We analyze in this study the participation of Reelin and mDab1 in thedevelopment of hippocampal connections. We show that mDab1 ispresent in growth cones and axonal tracts of developing hippocampalafferents.mdab1-deficiency produces severe alterations in the entorhino-hippocampal and commissural connections identical to those describedin reeler mice, including innervation of ectopic areas, formation ofabnormal patches of fiber termination and a delay in the refinement ofprojections. Organotypic slice cultures combining tissue from mdab1-mutant and control mice demonstrate that the abnormalities observed inthe mutant entorhino-hippocampal projection are caused by mdab1-deficiency in both the projecting neurons and target hippocampal cells.Axonal afferents that innervate the hippocampus react to Reelin byreducing axonal growth, and increasing growth cone collapse and axonalbranching. Altogether these results indicate that Reelin and mDab1participate in the development and refinement of hippocampal connec-tions by regulating axonal extension, targeting and branching.© 2007 Elsevier Inc. All rights reserved.
Keywords: mDab1; Reelin; Axonal outgrowth
Abbreviations: DG, dentate gyrus; EC, entorhinal cortex; f, fimbria; GL,granule cell layer; HF, hippocampal fissure; HP, hippocampus; IML, innermolecular layer; ML, molecular layer; MML, medial molecular layer; NC,neocortex; OML, outer molecular layer; SC, subicular complex; SLM,stratum lacunosum moleculare; SO, stratum oriens; SP, stratum pyramidale;SR, stratum radiatum; WM, white matter.⁎ Corresponding author. Developmental Neurobiology and Regeneration
Laboratory, Institute for Research in Biomedicine (IRB), Barcelona SciencePark (S1-A1), Josep Samitier, 1-5, E-08028 Barcelona, Spain. Fax: +34 934037116.
E-mail address: [email protected] (E. Soriano).1 These authors contributed equally to this work.2 Present address: Developmental Neurobiology Laboratory, Instituto
de Neurosciencias, University Miguel Hernandez, San Joan d'Alacant,E-03552, Spain.
Available online on ScienceDirect (www.sciencedirect.com).
1044-7431/$ - see front matter © 2007 Elsevier Inc. All rights reserved.doi:10.1016/j.mcn.2007.06.006
Introduction
The correct functioning of the CNS requires both the properpositioning of neurons within layers or nuclei, and the formation ofappropriate neuronal connections. The development of axonalpathways depends on the action of guidance molecules on growthcones. Numerous diffusible and membrane-bound molecules play acentral role in the guidance and development of neural connections(Mueller, 1999; Chilton, 2006; Huber et al., 2003). Somemolecules, such as Netrin-1, participate in both neuronal migrationand axonal guidance (Barallobre et al., 2005). The hippocampus(HP) is a laminated structure that receives highly ordered extrinsicand intrinsic inputs, where the different afferent systems terminatein non-overlapping layers (Super and Soriano, 1994). In previousstudies we have shown that Cajal–Retzius (CR) cells play a role inthe development of hippocampal connections (Borrell et al.,1999b; Del Rio et al., 1997; Super et al., 1998). CR cells, firstidentified as a special class of pioneer neurons in the marginal zoneof the neocortex, are also present in the marginal zone of the HPprior to and during the ingrowth of hippocampal afferents (Del Rioet al., 1996, 1997; Marin-Padilla, 1998; Soriano and Del Rio,2005; Soriano et al., 1994; Super and Soriano, 1994). CR cellsexpress Reelin, a large extracellular molecule that is critical forlamination of several brain regions and that, when mutated, leadsto the reeler phenotype (D'Arcangelo et al., 1995; Rice and Curran,2001; Tissir and Goffinet, 2003). Shortly after the discovery ofReelin, mDab1–the mouse homolog of Drosophila Disabledprotein–was discovered. Mutation of mDab1 leads to migrationdisorders indistinguishable from those observed in reeler (Howellet al., 1997b, 1997a; Sheldon et al., 1997). mDab1 is an intra-cellular adaptor protein that is phosphorylated in response toReelin signaling (Bar et al., 2003; Howell et al., 2000; Keshvaraet al., 2001; Rice and Curran, 2001; Tissir and Goffinet, 2003).Other molecules shown to participate in the Reelin signalingpathway include the lipoprotein receptors VLDLR and ApoER2,Src family kinases, PI3K, Akt1, GSK3β and MAP1B (Gonzalez-

159V. Borrell et al. / Mol. Cell. Neurosci. 36 (2007) 158–173
Billault et al., 2005; D’Arcangelo et al., 1999; Trommsdorff et al.,1999; Bock and Herz, 2003; Beffert et al., 2002; Ballif et al., 2003;Tissir and Goffinet, 2003; Kuo et al., 2005). Moreover, the ERKsignaling pathway has also been implicated in the Reelin pathway(Simo et al., 2007).
Several studies have shown that the development of hippo-campal connections requires the coordinated participation of anumber of molecules (Pozas et al., 2001; Barallobre et al., 2000;Demyanenko et al., 1999; Dahme et al., 1997; Nguyen Ba-Charvetet al., 1999; Stein et al., 1999; Seki and Rutishauser, 1998; Steupet al., 1999, 2000; Chedotal et al., 1998). We have shown thatReelin contributes to the layer-specific targeting of hippocampalafferents and to correct synaptogenesis (Borrell et al., 1999a; DelRio et al., 1997). Reelin has also been shown to positively regulatedendritic growth in the adult brain (Niu et al., 2004). Reelin isthought to be required for LTP and synaptogenesis (Herz andChen, 2006). It has also been reported that Reelin and mDab1 arerequired for synaptic targeting in the retina (Rice et al., 2001) andthat Reelin does not act as a chemorepellent for cortical axons invitro (Jossin and Goffinet, 2001).
Here we analyze the development of entorhino-hippocampalconnections and the pattern of commissural projections in mdab1knock-out mice. We show that the mdab1 gene is expressed in theentorhinal cortex (EC) and HP and that mDab1 protein is expressed inentorhinal and commissural afferents. We demonstrate that theabsence of mDab1 leads to several alterations in the development ofthe entorhino-hippocampal pathway, including a delay in theappearance of the topographic patterns of projection, and theformation of ectopic patches of axonal termination. Some of theseabnormalities are transient, whereas others persist into adulthood.Using mutant/wild-type mixed entorhino-hippocampal organotypicco-cultures, we demonstrate that the absence of mDab1 protein inboth the entorhinal projecting neurons and in the target HP contributesto the abnormalities observed in mdab1-mutants. Finally, we showthat Reelin induces axonal collapse of CA3 axons and increasesbranching of HP neurons. Similarly, Reelin also affects the outgrowthof EC and dorsal root ganglia (DRG) axons in explant cultures,indicating that Reelin modulates axonal growth and branching.
Results
Pattern of mdab1 mRNA expression and immunoreactivity in thedeveloping hippocampal system
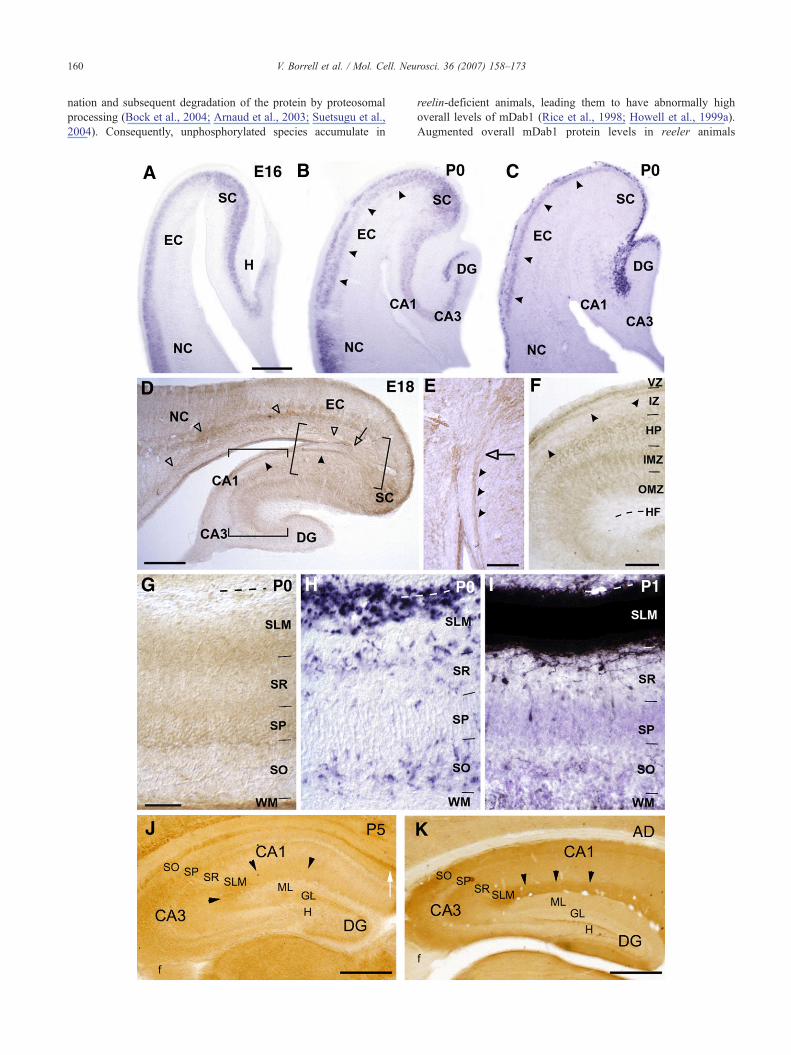
To understand the role of mDab1 in hippocampal developmentwe first analyzed its expression at the gene and protein levels. Inagreement with previous studies (Sheldon et al., 1997; Rice et al.,1998), mdab1 expression was detected by in situ hybridization atE14–18 in the cortical plate of the entire developing cerebral cortex.In the EC, mdab1 expression was also detected in the cortical plate,which contains postmitotic neurons, whereas in the developing HP,expression was confined to the hippocampal plate, where post-mitotic pyramidal neurons accumulate (Fig. 1A). Lower expressionlevels were detected in the dentate gyrus (DG) (Fig. 1A). In newbornmice, mdab1mRNAwas detected in postmitotic neurons of the EC,including upper and lower layers (layers II–III), which also expressreelin (Figs. 1B and C). In the P0 HP, expression was detected indeveloping pyramidal and granule cells (Fig. 1B). At later stages(P5–Adult) the pattern ofmdab1 expression remained essentially thesame although levels of expression decreased with age, except inlayer II of the EC (data not shown).
At E14, mDab1 immunostaining in the HP was mainly confinedto the fimbria, containing developing commissural axons. At E16–E18, axonal bundles were strongly labeled in the fimbria, inter-mediate zone (especially in the CA1 and subicular areas), andangular bundle (Figs. 1D and E). The hippocampal plate and theinner marginal zone were diffusely labeled, whereas the outermarginal zone was mDab1-negative at E16. Starting at E18, thelower aspect of the outer marginal zone became mDab1-positive(Figs. 1D and F). In the DG, the emerging granule cell layer wasimmunoreactive. mDab1-positive cells were observed in thedentate ventricular zone and along the migratory pathway towardsthe developing DG (Fig. 1D).
At P0, the overall level of mDab1 immunostaining increasedwith respect to earlier stages. Numerous commissural axon bundleswere labeled in the fimbria and white matter. The stratum lacu-nosum moleculare (SLM) was highly immunoreactive in its loweraspect, where fiber bundles could be identified. However, mDab1could not be detected in the upper aspect of the SLM (Fig. 1G). Inorder to ascertain whether entorhinal afferents terminating in theSLM are mDab1-positive, we compared the pattern of mDab1immunolabeling with that of entorhino-hippocampal innervation inthe CA1 area. mDab1 staining coincided with entorhinal axons, butonly in the portion of the SLM closest to the stratum radiatum (SR)(Figs. 1G and I). Interestingly, the absence of an mDab1 signal inthe upper aspect of the SLM correlated with the presence ofReelin-expressing CR cells (Figs. 1G and H), suggesting thatmDab1 protein levels in entorhinal axons that innervate the SLMmay be locally downregulated by Reelin (Rice et al., 1998; Arnaudet al., 2003).
At P5–P8, in the HP proper, the stratum oriens (SO) and whitematter were strongly labeled, with axonal bundles being recogniz-able. In addition, the SLM displayed strong and homogeneousimmunoreactivity, with axonal fascicles visible in area CA3. TheSR displayed diffuse, weak immunostaining, whereas the stratumlucidum (SL) was mDab1-negative. In the DG, diffuse stainingwas observed in the medial molecular layer (MML), whereas nosignal was visible in the inner or outer molecular layers (Fig. 1J).At P15, axon fascicles were mDab1 positive in the hippocampalcommissure, fimbria, and in the hippocampal white matter. The HPproper had diffuse immunostaining in the SO, SR, and SLM. In theDG, mDab1 immunolabeling was observed in the granule celllayer and in the medial and outer molecular layers (data notshown).
In the adult brain, axonal fascicles were immunopositive in thehippocampal commissure, but axonal staining was fainter in thefimbria and hippocampal white matter. The SO and SLM showedstrong staining, whereas in the SR the staining was weaker. Thestratum lucidum was devoid of staining, as observed during develop-ment. In the adult DG, the molecular layer (ML) was mDab1-positive in its middle and outer portions (Fig. 1K). We thusconclude that mDab1 is expressed in projection neurons of theentorhinal cortex and hippocampus and that mDab1 protein islocalized in developing axonal tracts and termination layers.
mDab1 protein is present in the developing axons that innervatethe HP
To evaluate the involvement of mDab1 in the development ofhippocampal connections, we first analyzed whether mDab1 pro-tein in axons was regulated by Reelin in vivo. Phosphorylation ofmDab1 protein occurs after Reelin signaling, and induces ubiquiti-
160 V. Borrell et al. / Mol. Cell. Neurosci. 36 (2007) 158–173
nation and subsequent degradation of the protein by proteosomalprocessing (Bock et al., 2004; Arnaud et al., 2003; Suetsugu et al.,2004). Consequently, unphosphorylated species accumulate in
reelin-deficient animals, leading them to have abnormally highoverall levels of mDab1 (Rice et al., 1998; Howell et al., 1999a).Augmented overall mDab1 protein levels in reeler animals
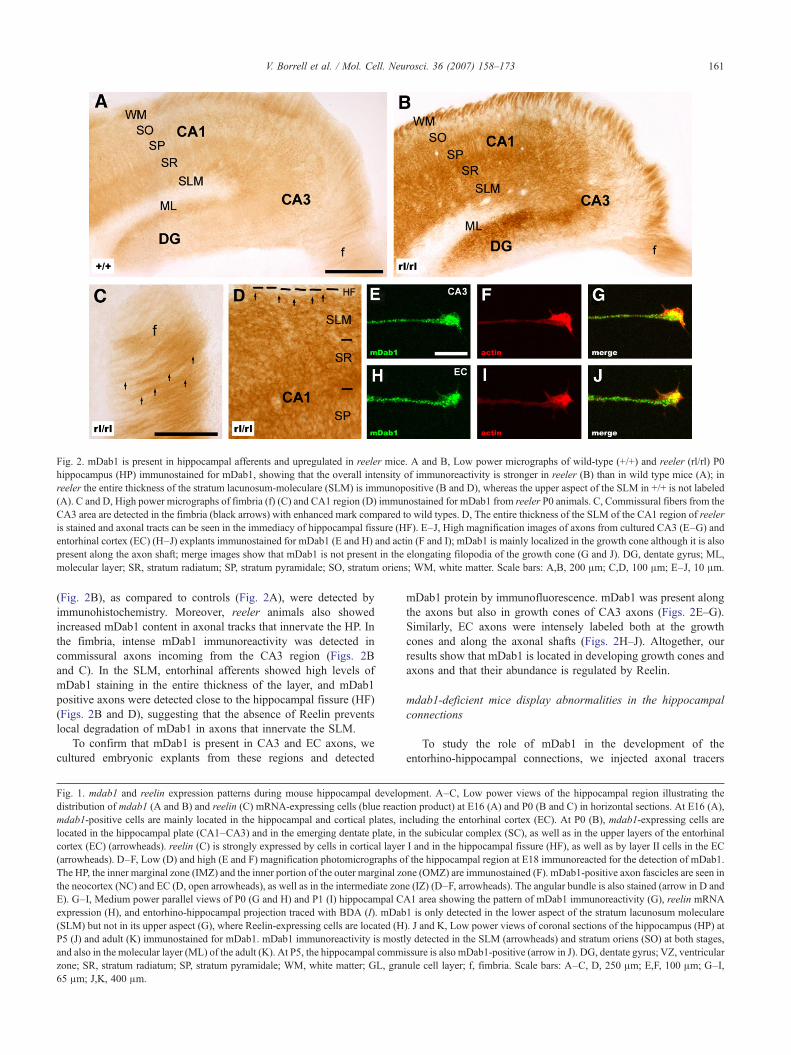
Fig. 2. mDab1 is present in hippocampal afferents and upregulated in reeler mice. A and B, Low power micrographs of wild-type (+/+) and reeler (rl/rl) P0hippocampus (HP) immunostained for mDab1, showing that the overall intensity of immunoreactivity is stronger in reeler (B) than in wild type mice (A); inreeler the entire thickness of the stratum lacunosum-moleculare (SLM) is immunopositive (B and D), whereas the upper aspect of the SLM in +/+ is not labeled(A). C and D, High power micrographs of fimbria (f) (C) and CA1 region (D) immunostained for mDab1 from reeler P0 animals. C, Commissural fibers from theCA3 area are detected in the fimbria (black arrows) with enhanced mark compared to wild types. D, The entire thickness of the SLM of the CA1 region of reeleris stained and axonal tracts can be seen in the immediacy of hippocampal fissure (HF). E–J, High magnification images of axons from cultured CA3 (E–G) andentorhinal cortex (EC) (H–J) explants immunostained for mDab1 (E and H) and actin (F and I); mDab1 is mainly localized in the growth cone although it is alsopresent along the axon shaft; merge images show that mDab1 is not present in the elongating filopodia of the growth cone (G and J). DG, dentate gyrus; ML,molecular layer; SR, stratum radiatum; SP, stratum pyramidale; SO, stratum oriens; WM, white matter. Scale bars: A,B, 200 μm; C,D, 100 μm; E–J, 10 μm.
161V. Borrell et al. / Mol. Cell. Neurosci. 36 (2007) 158–173
(Fig. 2B), as compared to controls (Fig. 2A), were detected byimmunohistochemistry. Moreover, reeler animals also showedincreased mDab1 content in axonal tracks that innervate the HP. Inthe fimbria, intense mDab1 immunoreactivity was detected incommissural axons incoming from the CA3 region (Figs. 2Band C). In the SLM, entorhinal afferents showed high levels ofmDab1 staining in the entire thickness of the layer, and mDab1positive axons were detected close to the hippocampal fissure (HF)(Figs. 2B and D), suggesting that the absence of Reelin preventslocal degradation of mDab1 in axons that innervate the SLM.
To confirm that mDab1 is present in CA3 and EC axons, wecultured embryonic explants from these regions and detected
Fig. 1. mdab1 and reelin expression patterns during mouse hippocampal developdistribution of mdab1 (A and B) and reelin (C) mRNA-expressing cells (blue reactmdab1-positive cells are mainly located in the hippocampal and cortical plates, inlocated in the hippocampal plate (CA1–CA3) and in the emerging dentate plate, incortex (EC) (arrowheads). reelin (C) is strongly expressed by cells in cortical layer(arrowheads). D–F, Low (D) and high (E and F) magnification photomicrographs oThe HP, the inner marginal zone (IMZ) and the inner portion of the outer marginal zothe neocortex (NC) and EC (D, open arrowheads), as well as in the intermediate zonE). G–I, Medium power parallel views of P0 (G and H) and P1 (I) hippocampal Cexpression (H), and entorhino-hippocampal projection traced with BDA (I). mDab(SLM) but not in its upper aspect (G), where Reelin-expressing cells are located (HP5 (J) and adult (K) immunostained for mDab1. mDab1 immunoreactivity is mostand also in the molecular layer (ML) of the adult (K). At P5, the hippocampal commzone; SR, stratum radiatum; SP, stratum pyramidale; WM, white matter; GL, gran65 μm; J,K, 400 μm.
mDab1 protein by immunofluorescence. mDab1 was present alongthe axons but also in growth cones of CA3 axons (Figs. 2E–G).Similarly, EC axons were intensely labeled both at the growthcones and along the axonal shafts (Figs. 2H–J). Altogether, ourresults show that mDab1 is located in developing growth cones andaxons and that their abundance is regulated by Reelin.
mdab1-deficient mice display abnormalities in the hippocampalconnections
To study the role of mDab1 in the development of theentorhino-hippocampal connections, we injected axonal tracers
ment. A–C, Low power views of the hippocampal region illustrating theion product) at E16 (A) and P0 (B and C) in horizontal sections. At E16 (A),cluding the entorhinal cortex (EC). At P0 (B), mdab1-expressing cells arethe subicular complex (SC), as well as in the upper layers of the entorhinalI and in the hippocampal fissure (HF), as well as by layer II cells in the ECf the hippocampal region at E18 immunoreacted for the detection of mDab1.ne (OMZ) are immunostained (F). mDab1-positive axon fascicles are seen ine (IZ) (D–F, arrowheads). The angular bundle is also stained (arrow in D andA1 area showing the pattern of mDab1 immunoreactivity (G), reelin mRNA1 is only detected in the lower aspect of the stratum lacunosum moleculare). J and K, Low power views of coronal sections of the hippocampus (HP) atly detected in the SLM (arrowheads) and stratum oriens (SO) at both stages,issure is also mDab1-positive (arrow in J). DG, dentate gyrus; VZ, ventricularule cell layer; f, fimbria. Scale bars: A–C, D, 250 μm; E,F, 100 μm; G–I,
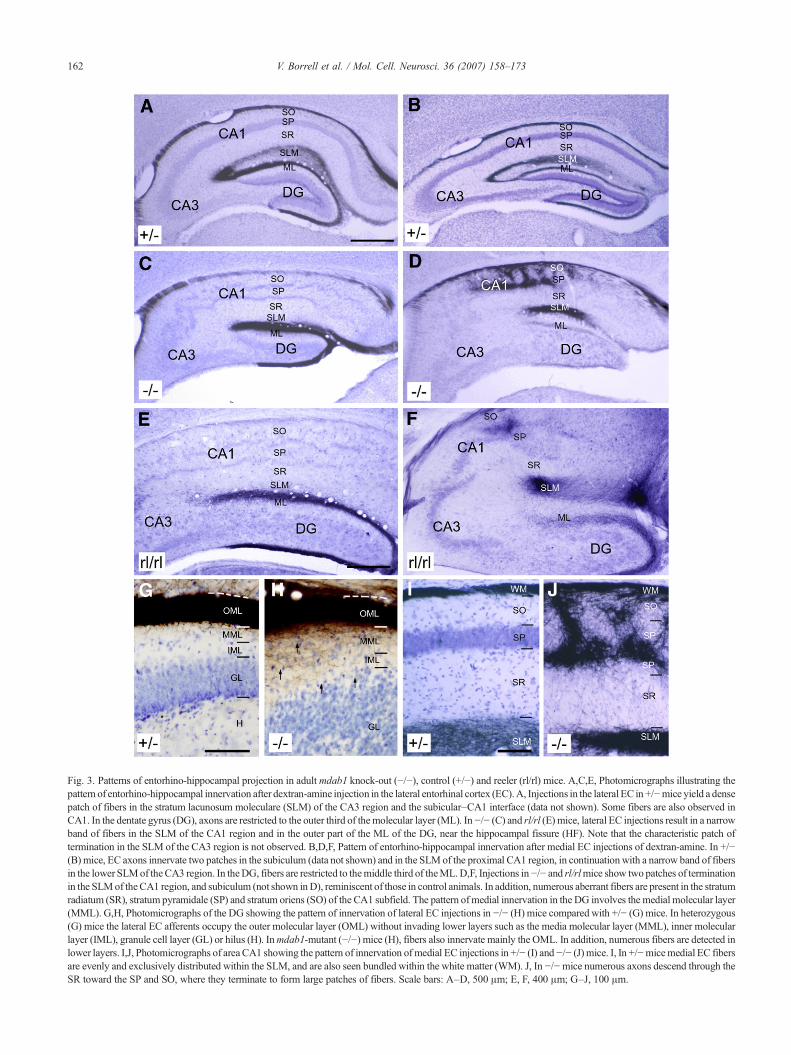
Fig. 3. Patterns of entorhino-hippocampal projection in adultmdab1 knock-out (−/−), control (+/−) and reeler (rl/rl) mice. A,C,E, Photomicrographs illustrating thepattern of entorhino-hippocampal innervation after dextran-amine injection in the lateral entorhinal cortex (EC).A, Injections in the lateral EC in +/−mice yield a densepatch of fibers in the stratum lacunosum moleculare (SLM) of the CA3 region and the subicular–CA1 interface (data not shown). Some fibers are also observed inCA1. In the dentate gyrus (DG), axons are restricted to the outer third of themolecular layer (ML). In−/− (C) and rl/rl (E) mice, lateral EC injections result in a narrowband of fibers in the SLM of the CA1 region and in the outer part of the ML of the DG, near the hippocampal fissure (HF). Note that the characteristic patch oftermination in the SLM of the CA3 region is not observed. B,D,F, Pattern of entorhino-hippocampal innervation after medial EC injections of dextran-amine. In +/−(B) mice, EC axons innervate two patches in the subiculum (data not shown) and in the SLMof the proximal CA1 region, in continuationwith a narrow band of fibersin the lower SLMof theCA3 region. In theDG, fibers are restricted to themiddle third of theML.D,F, Injections in−/− and rl/rlmice show two patches of terminationin the SLMof theCA1 region, and subiculum (not shown inD), reminiscent of those in control animals. In addition, numerous aberrant fibers are present in the stratumradiatum (SR), stratum pyramidale (SP) and stratum oriens (SO) of the CA1 subfield. The pattern ofmedial innervation in the DG involves the medial molecular layer(MML). G,H, Photomicrographs of the DG showing the pattern of innervation of lateral EC injections in −/− (H) mice compared with +/− (G) mice. In heterozygous(G) mice the lateral EC afferents occupy the outer molecular layer (OML) without invading lower layers such as the media molecular layer (MML), inner molecularlayer (IML), granule cell layer (GL) or hilus (H). Inmdab1-mutant (−/−) mice (H), fibers also innervate mainly the OML. In addition, numerous fibers are detected inlower layers. I,J, Photomicrographs of area CA1 showing the pattern of innervation of medial EC injections in +/− (I) and−/− (J) mice. I, In +/−micemedial EC fibersare evenly and exclusively distributed within the SLM, and are also seen bundled within the white matter (WM). J, In −/−mice numerous axons descend through theSR toward the SP and SO, where they terminate to form large patches of fibers. Scale bars: A–D, 500 μm; E, F, 400 μm; G–J, 100 μm.
162 V. Borrell et al. / Mol. Cell. Neurosci. 36 (2007) 158–173

163V. Borrell et al. / Mol. Cell. Neurosci. 36 (2007) 158–173
into the EC of postnatal mutant mice, then compared the observedinnervation patterns with those of wild-type, heterozygous mice aswell as to previous analyses of the reeler phenotype (Borrell et al.,1999a). Since mdab1 −/− and scm/scm mice yielded identicallabeling patterns, both groups were pooled and are hereafter de-scribed together.
As previously described (Super and Soriano, 1994), theentorhinal innervation patterns in P1 and P5 wild-type mice wererestricted to the SLM. The innervation was denser at P5, withhighly defasciculated axons forming elaborate arbors and display-ing numerous collaterals. In the DG, at P1 the entorhinal fiberswere just beginning to innervate the dentate outer molecular layer(OML), whereas at P5 they had already formed a dense innervationin this layer. The heterozygous (+/−) and (+/scm) mice showed thesame innervation pattern reported for wild type mice (Supplemen-tary Figs. 1A and D).
The pattern of entorhinal innervation in mdab1-mutant mice atP1–P5 was very similar to that observed in reeler pups (Borrellet al., 1999a). At dorsal hippocampal levels, most entorhinal fibersof reeler and mdab1-mutants were densely packed near the HF,following rather straight courses and exhibiting few collaterals oraxonal varicosities (Supplementary Fig. 1B and C). Moreover, asin reeler mice, the CA3 area was almost devoid of entorhinalinnervation. In the DG, entorhinal fibers were densely packed inthe OML at P5, and many fibers were seen terminating ectopi-cally in the inner molecular layer (IML) and hilus (SupplementaryFig. 1B and C). In addition, entorhinal axons terminated aberrantlywithin the SR, the SP and SO, especially at ventral levels, formingectopic termination patches (Supplementary Figs. 1E and F).
The innervation pattern from P16 onward was essentially thesame as that in adults, so these ages are hereafter describedtogether.
Tracer injections in the lateral or medial entorhinal areas ofwild-type mice revealed two distinct patterns of entorhinal in-nervation to the HP (Swanson and Cowan, 1977; Ruth et al., 1982;Borrell et al., 1999a). Injections in the lateral EC of either wild typeor heterozygous mice formed a dense patch of profusely branchedfibers in the SLM of the CA3 and in the interface CA1-subiculum(Fig. 3A). Conversely, medial entorhinal injections innervatedselectively the molecular layer of the subiculum, the SLM of areaCA1, and the lower aspect of the SLM in CA3, where they formedelaborated axonal arbors (Fig. 3B and I). In mdab1-mutant mice,innervation from the lateral EC was confined to the upper aspect ofthe SLM in the CA1 and subiculum, forming a continuous band offibers without regional patches of termination (Fig. 3C). Theinnervation of the SLM was narrower in the mdab1-mutants (44+4.6 μm) than in control littermates (119+4 μm), and numerousfibers in the CA1 area were fasciculated, displaying few axonalbranches or varicosities. This pattern of innervation was essentiallysimilar to that previously observed in reeler mice (Fig. 3E) (Borrellet al., 1999a).
Following injections in the medial entorhinal area of mdab1-mutant mice, a typical two-patched pattern of axonal terminationcould be observed in the SLM of the CA1 area and subiculum(Fig. 3D). A dense patch of ectopic terminations was observed inCA1, where axons innervated the SR, SO, and the layer in betweenthe double pyramidal layer (Fig. 3J). This feature was neverobserved in control mice, and was reminiscent of that observed inreeler mice (Fig. 3F) (Borrell et al., 1999a).
In the DG of control (+/+ and +/−) mice, lateral entorhinalfibers were detected in the OML, whereas innervation from the
medial EC was largely confined to the MML (Figs. 3A, B and G).In mdab1-mutants, lateral and medial entorhinal afferents termi-nated mostly in the OML and MML, respectively, although in bothcases a great number of ectopic fibers invaded inappropriate layersof the DG (Figs. 3C, D and H).
Analysis of the development of entorhino-hippocampal projec-tions in mdab1-deficient mice reveals numerous axonal alterationsthat are very similar to those observed in reeler mice (Borrell et al.,1999a; Stanfield et al., 1979). Similarly, hippocampal commissuralprojections are deeply altered in mdab1-deficient mice (Supple-mentary Fig. 2), showing the same abnormalities previously des-cribed in the reeler mouse (Borrell et al., 1999b; Stanfield et al.,1979). Altogether, these results suggests that mDab1 participates inthe transduction pathway of the Reelin signal that operates in thedevelopment of the entorhinal and commissural projections.
mDab1 is involved in the outgrowth and positioning ofentorhino-hippocampal fibers
Since mdab1 is expressed in both the HP and the EC, theabnormalities observed in the entorhino-hippocampal projection ofthe mdab1-deficient mice could be due to the lack of mdab1 in eitherthe ingrowing developing axons or in the target region. To discernbetween these possibilities, we prepared mixed organotypic entorhi-no-hippocampal co-cultures using slices obtained from newbornmdab1-mutants and their control littermates. After 7 days in vitro(DIV), homogenic co-cultures (EC+/+/HP+/+, EC+/−/HP+/−, EC−/−/HP−/−) displayed patterns of entorhino-hippocampal innervationsimilar to those described in vivo, supporting the usefulness of thismodel for characterization of mixed co-cultures (EC+/−/HP−/−, EC−/−/HP+/+). In homogenic wild-type (EC+/+/HP+/+) and heterozygous(EC+/−/HP+/−) co-cultures, entorhino-hippocampal fibers formed adense innervation restricted to the OML and SLM (Fig. 4A). Theseco-cultures frequently showed distinct topographic patches ofentorhinal termination that could be observed in the CA1 and CA3areas, depending on whether the injection was in a medial or lateralco-cultured EC (Fig. 4A). In mutant homogenic co-cultures (EC−/−/HP−/−), innervation of the HP proper exhibited severe abnormalities(Fig. 4D). The entorhinal projection to the SLM was compact andrestricted to the upper aspect, with fibers displaying abundant col-laterals and axonal varicosities. Numerous entorhinal fibers des-cended from the SLM toward the SR, where they branched profuselyand formed an ectopic patch of termination in the CA1 (Fig. 4D,whitearrows) and CA3 areas (Fig. 4D, white arrowheads). Similarly, theperforant entorhinal axons innervated the DG near the HF with fiberspacked very tightly, showing several misrouted fibers ectopicallyinnervating the hilus (Figs. 4H and L).
When heterozygous entorhinal slices were co-cultured withmutant HP (EC+/−/HP−/−), a mixed projection pattern displayingcharacteristics of both control and mutant homogenic cultures wereobtained (Fig. 4B). Thus, afferent axons were not packed close tothe HF but innervated the entire thickness of SLM and OML(Fig. 4B). Topographic termination patches were never formed atour time of culture (7 DIV), and misrouted fibers were frequentlyseen in the SR of the CA1 (Fig. 4B, white arrows) and CA3 areas(Fig. 4B, white arrowheads). Moreover, heterozygous entorhinalaxons profusely innervated the mutant hilus (Fig. 4F).
When mutant entorhinal slices were co-cultured with hetero-zygous hippocampal slices (EC−/−/HP+/−), the innervation patternalso showed mixed characteristics of both control and mutanthomogenic co-cultures (Fig. 4C). Thus, entorhinal axons were
164 V. Borrell et al. / Mol. Cell. Neurosci. 36 (2007) 158–173

Fig. 5. Quantification of abnormalities in wild-type, mdab1-deficient, andmixed organotypic slice co-cultures. Abnormalities in organotypic cultureswere quantified by analyzing the percentage of cultures that exhibitedmoderateto severe ectopic innervation in the different regions. The basal percentage ofwild-type homogenic co-cultures with abnormalities (10% to 30%) isattributable to defective biocytin injections. Ectopies in CA1 are merelyattributable to themispositioning of neurons in the targetHP.On the other hand,ectopic innervation of CA3 and hilus is responsible for defects in mdab1 inboth the EC and HP, with the highest percentage of severely misroutedprojection being observed inwild type EC fibers innervatingmutant HP (75%).
165V. Borrell et al. / Mol. Cell. Neurosci. 36 (2007) 158–173
targeted to the SLM, being compact, but ectopic axons descendedto the SR of the CA3 area (Fig. 4C, white arrowheads). Ectopicfibers in the SR of the CA1 area were almost absent (Fig. 4C, whitearrow). Innervation in the OML was mainly restricted to the upperaspect, where axons were tightly packed even though fibersectopically innervated the hilus (Figs. 4G and K).
To support the findings of these abnormalities, the percentage ofcultures with fiber alterations was calculated (Fig. 5). In the CA1area, ectopic axonal terminations were found in homotypic mdab1-mutant co-cultures and in co-cultures containing mdab1-mutanthippocampal slices, but not in mutant entorhinal slices cultured withheterozygous hippocampi, indicating that ectopies in this region arecaused by mispositioning of target neurons (Figs. 4A–D). Incontrast, mdab1 deficiency in either entorhinal axons or targethippocampal neurons results in similar ectopic fibers in the CA3region and in the hilus (Fig. 5, see also Figs. 4E–H). Similarly, in thehilus, the most severe ectopic innervation was achieved in EC+/−/HP−/− co-cultures followed by mdab1-mutant homogenic co-cultures, indicating that heterozygous entorhinal axons grow moreprofusely into the mdab1-mutant HP than when fibers were nullmutant for mdab1 (Figs. 4E–H and 5). Interestingly, compact fiberswere observed in the HF when the EC was deficient in mdab1, butnot in EC+/−/HP−/− co-cultures, indicating that this abnormality is
Fig. 4. In vitro formation of the entorhino-hippocampal projection in mdab1-defcultures from entorhinal cortex (EC) and hippocampus (HP). Entorhino-hippocampwith DAB precipitates. A, In wild type co-cultures (EC+/−/HP+/−), EC afferents termthe outer aspect of the molecular layer (ML), forming topographic patches of terminco-cultures (EC−/−/HP−/−), numerous misrouted fibers ectopically innervate the CAarrowheads), as well as occur in vivo. B, Mixed EC−/−/HP+/− co-cultures exhibitcultures, but in the CA3 region these ectopies are even more abundant. C, Mixedectopies in CA3 are as frequent as in homogenic mutant co-cultures. E–H, High powwild type homogenic co-cultures, the perforant pathway afferents are restricted to thto the hilus from the ML (black arrows) when at least one part of the co-culture is maxons innervate mutant HP (EC+/−/HP−/−) (G). I–L, Severe compaction of fibers nmdab1-defective. Both EC−/−/HP−/− (L) and EC−/−/HP+/− (K) co-cultures, but not Eresults indicate that mDab1 plays a key role in the adhesion properties of EC outgrow
due to the lack of mdab1 in axonal tracks (Figs. 4I–H and 5). Theresults of these in vitro experiments indicate that some abnormalitiesof the entorhino-hippocampal projection in mdab1 knock-out micemay be attributable to the lack of mDab1 protein in the entorhinalprojecting neurons, suggesting that mDab1 has a role in thedevelopment and adhesion properties of entorhinal axonal tracts.
Reelin modulates axonal outgrowth in vitro
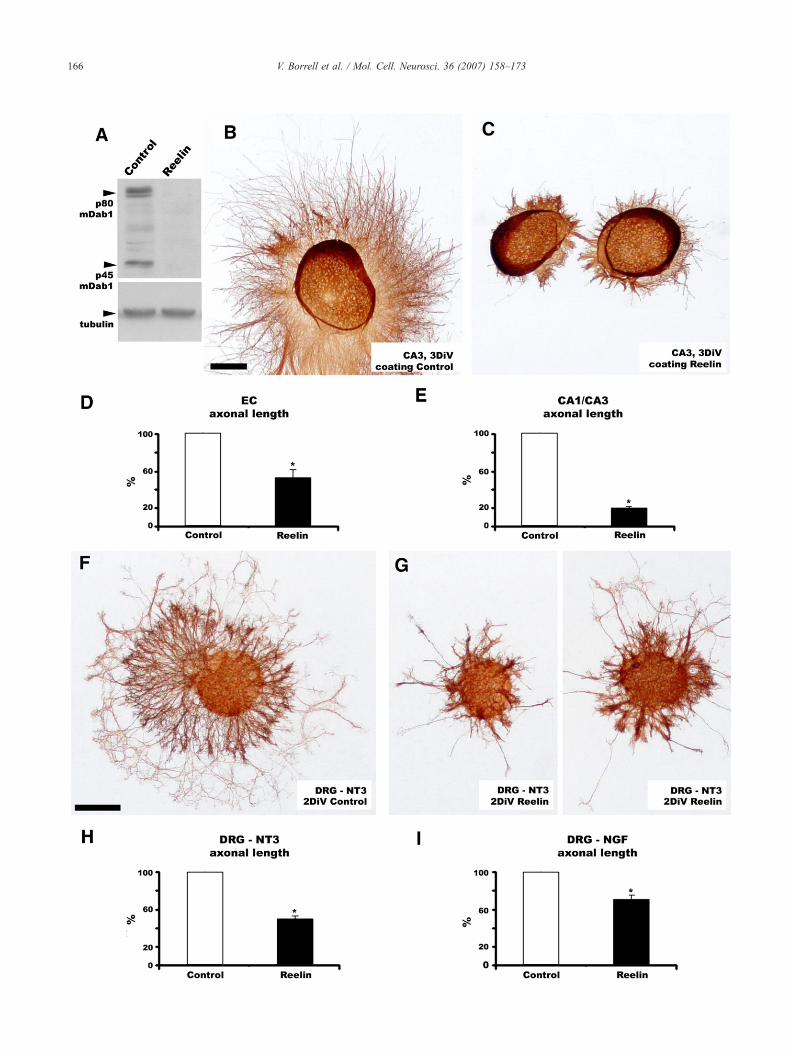
The presence ofmDab1 protein in the hippocampal afferents, andthe phenotypic alterations observed in the hippocampal tracts ofmdab1-deficient mice, indicated to us that the Reelin signalingpathwaymay directly influence axonal outgrowth.We thus analyzedthe effects of Reelin on axonal outgrowth from EC and CA3neurons. We cultured explants from the EC and CA3 regions (P0),and from the DRG (E16), on Reelin-coated coverslips and measuredthe length of axons growing radially from the explants. To test theactivity of Reelin-coated coverslips, we measured the levels ofmDab1 protein in cultured cells by Western Blot (WB). Neuronscultured on Reelin-coated plates showed reduced mDab1 proteincontent and increased phosphorylation, compared to control Mock-coated plates (Fig. 6A and Supplementary Fig. 3A), thus indicatingthat Reelin is functional in the experimental conditions.
In control, mock-treated coverslips, individual axons grew outof the explants for long distances (Figs. 6B and F). However, forcoverslips coated with Reelin, axonal growth was strongly reducedin both CA3 (Figs. 6C and E) and EC (Fig. 6D). Moreover, growingaxons appeared more fasciculated upon Reelin treatment than incontrol conditions (Figs. 6B and C). Interestingly, a similar reduc-tion in axonal growth was observed in DRG explants treated witheither NT3 or NGF (Figs. 6F–I).
Reelin modulates hippocampal axonal collapse, branching andadhesion
We further investigated the effects of Reelin on axonal growthcones. After 3DIV, CA3 explants were treated with supernatantscontaining Reelin (Fig. 7B) or with control Mock supernatants(Fig. 7A) for 1 h, and then stained with Phalloidin. We found thatReelin is also biologically active in these conditions (Supple-mentary Fig. 3B). We found that incubation with Reelinsignificantly increased the percentages of collapsed growth cones(i.e., those with round-tipped or “pencil-like” shapes lackingfilopodia and lamellipodia) (Fig. 7C). The increase in collapsedgrowth cones was moderate and never reached percentages oftypical chemorepellents such as Class III Semaphorins (Pozas etal., 2001).
icient organotypic slice co-cultures. A–D, Low power micrographs of co-al axons were labeled with biocytin and visualized by immunohistochemistryinated in the entire thickness of the stratum lacunosummoleculare (SLM) andation, reminiscent of the in vivo phenotype. D, In mdab1-mutant homogenic1 area (white arrows) whereas other axons switch to the CA3 region (whitesimilar ectopies in the CA1 and CA3 regions as do mutant homogenic co-EC−/−/HP+/− co-cultures rarely exhibit ectopies in the CA1 area, but severeer micrographs of the hilar region of the DG from organotypic co-cultures. Ine ML (E). In contrast, misrouted fibers from the perforant pathway descendeddab1-deficient (F–H). The most severe ectopies are formed when wild typeear the hippocampal fissure (HF) is only observed when the EC axons areC+/−/HP+/− (I) or EC+/−/HP−/− (J), exhibit compacted fibers in the HF. Theseing axons. Scale bar: A–D, 500 μm; E–H, 100 μm; I,J, 50 μm; K,L, 50 μm.
166 V. Borrell et al. / Mol. Cell. Neurosci. 36 (2007) 158–173

167V. Borrell et al. / Mol. Cell. Neurosci. 36 (2007) 158–173
We then analyzed the effect of Reelin on axonal growth inhippocampal primary cultures. We found that, in contrast to thecontrols, neurons cultured on Reelin-coated coverslips tended tofasciculate in dendritic and axonal bundles, suggesting that hippo-campal neurons grow better on neuronal surfaces than on Reelinsubstrates (Figs. 7D and E). We also quantified the influence ofReelin on axonal branching in hippocampal neurons. Low-densityhippocampal cultures were cultured for 3DIV in a collagen-basedgel matrix containing Reelin (Fig. 7G) or Control Mock media(Fig. 7F). Individual neurons were recorded, and the total axonallength and number of branching points were calculated. As shownin Fig. 7H, Reelin increased both the number of axonal branchingpoints per neuron (by 48.0%) and the total axonal arbor length(by 22.8%) as compared to controls.
Discussion
Reelin is essential for proper neuronal migration during develop-ment of the cerebral cortex (D’Arcangelo et al., 1995; Ogawa et al.,1997; Nakajima et al., 1997). Phenotypic analyses of mutant mice,and biochemical studies, have revealed that the Reelin signalingcascade involves phosphorylation of the adapter protein mDab1(Howell et al., 1997a, 1997b, 2000; Sheldon et al., 1997; Rice andCurran, 2001). We have previously reported that Reelin is alsoinvolved in the development of the hippocampal connections,and that reeler mice display a number of abnormalities in theentorhinal and commissural projections (Del Rio et al., 1997;Borrell et al., 1999a, 1999b). These studies prompted the presentanalysis of the role of mDab1 in the development of hippocampalconnections.
mDab1 is expressed in developing hippocampal axons
The mdab1 gene was first discovered in Drosophila as amodifier of the Abl cytoplasmic tyrosine kinase (Gertler et al.,1989). In the present study we have shown that mdab1 mRNA isexpressed in the EC and HP during mouse development, includingthe stages at which hippocampal connections are first established.Around birth and at postnatal stages, mdab1 is expressed in theupper layers of the EC, where projecting neurons are located, andin the hippocampal plate, where commissural-projecting neuronsare located. It is worth mentioning that these postmitotic neuronsexpress the mdab1 gene long after completion of neuronal migra-tion. We have also shown that developing entorhino-hippocampalaxons express mDab1 protein in vitro and in vivo, not only in theaxonal shaft but also in the growth cones, in agreement withprevious observations (Howell et al., 1999b, 1997a). These datasupport the notion that mDab1 may also function in thedevelopment of hippocampal connections. Furthermore, we haveshown that entorhinal axons running through the upper aspect of
Fig. 6. Reelin reduces axonal outgrowth. A, Western blot showing mDab1 levels inthat Reelin coatings reduce mDab1 protein levels, as detected with the B3 antibodyReelin-coated (B) or in Control Mock-coated (A) coverslips. In Reelin-treated covobserve due to increased fasciculation. D,E, Quantification of length of axons growReelin-treated coverslips for 3 DIV. Histograms summarize the data (expressed in pmean±S.E.M. (⁎, pb0.05). F,G, β-III-tubulin immunostaining of DRG explantsControl Mock-coated (A) coverslips. In Reelin-treated coverslips the axonal outgroDRG explants supplemented with NT3- or NGF-containing media (G and H, respecsummarize the data (expressed in percentages) obtained in independent/separate200 μm; F–G, 200 μm.
the SLM of the HP display reduced levels of mDab1 protein, whichis coincident with the presence of Reelin-expressing CR cells inthis sublayer. Previous studies have shown that mDab1 is degradedin the proteasome after activation by Reelin, and that in reelermice, mDab1 protein is upregulated (Arnaud et al., 2003; Bocket al., 2004; Suetsugu et al., 2004; Howell et al., 1999a; Rice et al.,1998). Our observations that entorhinal axons in the SLM of thereeler HP express high levels of mDab1 not only demonstrate thatthese fibers contain mDab1 in the target layer, but also suggest thatentorhinal axons respond to Reelin by downregulating mDab1protein. Moreover, the presence of mDab1 in growth cones sug-gests a direct role for the Reelin/mDab1 signaling pathway in axongrowth and/or guidance.
Alterations in hippocampal connections in mdab1 mutant mice aresimilar to those in reeler mice
Our study also shows that the patterns of termination ofhippocampal afferents in mdab1-mutants are largely similar tothose described for reelermice (Borrell et al., 1999a, 1999b). For theentorhino-hippocampal pathway, these abnormalities include anincrease in ectopic fibers at inappropriate layers, both in the HPproper and the DG, the formation of aberrant projections from themedial EC to the SR and SO of the CA1 region, and an abundance ofmisrouted fibers. The main alterations observed in the commissuralpathway are the invasion of the SLM in area CA3, and theintermingling of fibers with granule cells in the DG and hilus. Theseresults suggest that mDab1 plays a role in the correct patterning ofhippocampal afferent termination, and indicate that Reelin modu-lates these axonal events through the adaptor mDab1 (Tissir andGoffinet, 2003; Rice et al., 1998; Rice and Curran, 2001).
Mechanisms of action of the Reelin–mDab1 signaling pathway inaxonal growth
Some alterations in reeler mice (e.g. mispositioning of corticalinterneurons) have been reported to be secondary to the alteredlayering of neurons targeted by the mutation (Pla et al., 2006;Yabut et al., 2007). To ascertain whether the abnormal patterns ofentorhino-hippocampal projections in mdab1-mutants were due tothe lack of mDab1 in the developing axons or in the target HP, weestablished mixed combinations of organotypic co-cultures. Inthese co-cultures, the pattern of entorhinal termination was amixture between those observed in control and mutant homogenicco-cultures. Accordingly, the presence of mDab1 in either theprojecting EC or the target HP was not sufficient to rescue themutant phenotype, and in both cases mixed phenotypic patternswere obtained (Fig. 8). The mispositioning of target neurons inmdab1-deficient HP is responsible for the ectopies observed inCA1, and influences those of CA3 and hilar regions. Surprisingly,
neurons cultured onto Control Mock- and Reelin-coated plates for 24 h. Note. B,C, β-III-tubulin immunostaining of CA3 explants cultured for 3 DIV inerslips the axonal outgrowth is reduced, and individual axons are difficult toing out of the EC (C) or CA1/CA3 (D) explants grown in Control Mock- orercentages) obtained from independent/separate experiments. Data representcultured for 3 DIV with NT3 containing media in Reelin-coated (B) or inwth is reduced. H,I, Quantification of the length of axons growing out of thetively) in Control Mock- or Reelin-treated coverslips for 3 DIV. Histogramsexperiments. Data represent mean±S.E.M. (⁎, pb0.05). Scale bar: B–C,
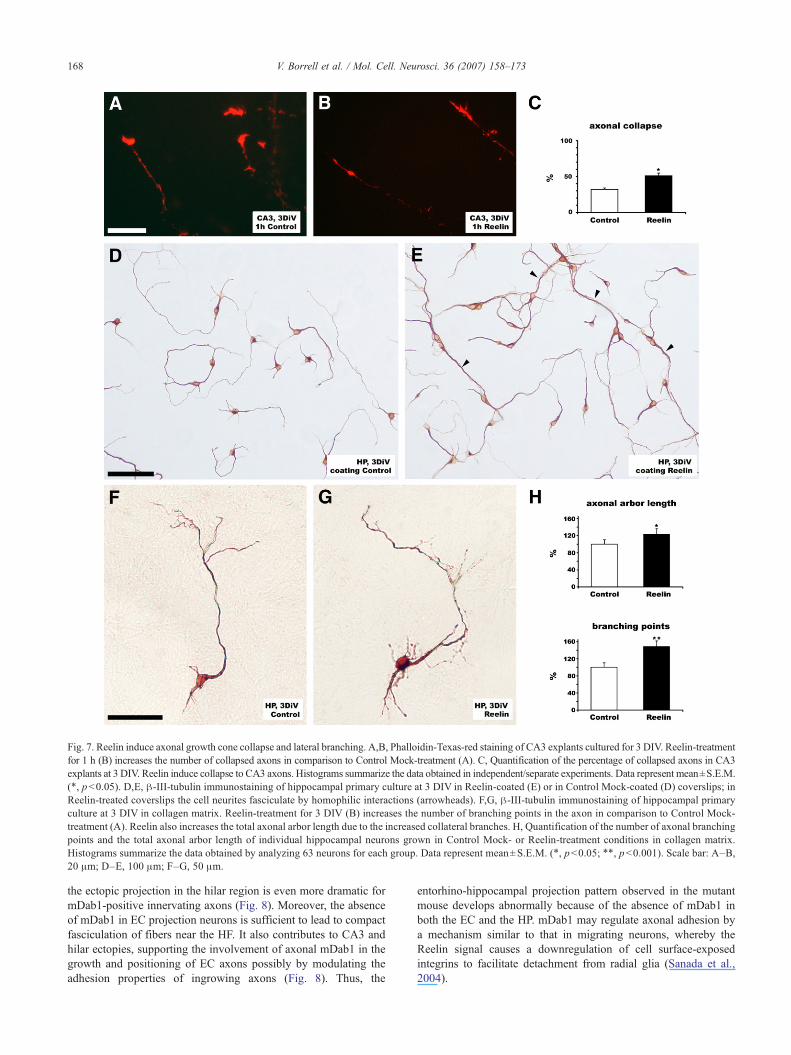
Fig. 7. Reelin induce axonal growth cone collapse and lateral branching. A,B, Phalloidin-Texas-red staining of CA3 explants cultured for 3 DIV. Reelin-treatmentfor 1 h (B) increases the number of collapsed axons in comparison to Control Mock-treatment (A). C, Quantification of the percentage of collapsed axons in CA3explants at 3 DIV. Reelin induce collapse to CA3 axons. Histograms summarize the data obtained in independent/separate experiments. Data represent mean±S.E.M.(⁎, pb0.05). D,E, β-III-tubulin immunostaining of hippocampal primary culture at 3 DIV in Reelin-coated (E) or in Control Mock-coated (D) coverslips; inReelin-treated coverslips the cell neurites fasciculate by homophilic interactions (arrowheads). F,G, β-III-tubulin immunostaining of hippocampal primaryculture at 3 DIV in collagen matrix. Reelin-treatment for 3 DIV (B) increases the number of branching points in the axon in comparison to Control Mock-treatment (A). Reelin also increases the total axonal arbor length due to the increased collateral branches. H, Quantification of the number of axonal branchingpoints and the total axonal arbor length of individual hippocampal neurons grown in Control Mock- or Reelin-treatment conditions in collagen matrix.Histograms summarize the data obtained by analyzing 63 neurons for each group. Data represent mean±S.E.M. (⁎, pb0.05; ⁎⁎, pb0.001). Scale bar: A–B,20 μm; D–E, 100 μm; F–G, 50 μm.
168 V. Borrell et al. / Mol. Cell. Neurosci. 36 (2007) 158–173
the ectopic projection in the hilar region is even more dramatic formDab1-positive innervating axons (Fig. 8). Moreover, the absenceof mDab1 in EC projection neurons is sufficient to lead to compactfasciculation of fibers near the HF. It also contributes to CA3 andhilar ectopies, supporting the involvement of axonal mDab1 in thegrowth and positioning of EC axons possibly by modulating theadhesion properties of ingrowing axons (Fig. 8). Thus, the
entorhino-hippocampal projection pattern observed in the mutantmouse develops abnormally because of the absence of mDab1 inboth the EC and the HP. mDab1 may regulate axonal adhesion bya mechanism similar to that in migrating neurons, whereby theReelin signal causes a downregulation of cell surface-exposedintegrins to facilitate detachment from radial glia (Sanada et al.,2004).
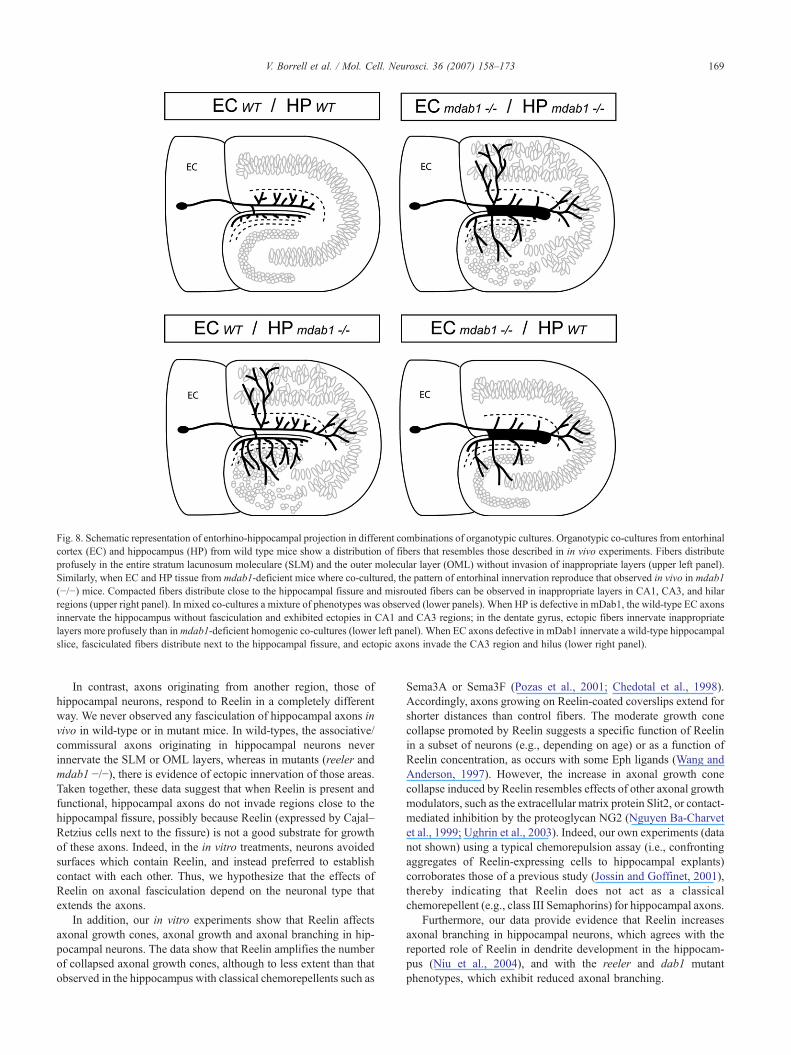
Fig. 8. Schematic representation of entorhino-hippocampal projection in different combinations of organotypic cultures. Organotypic co-cultures from entorhinalcortex (EC) and hippocampus (HP) from wild type mice show a distribution of fibers that resembles those described in in vivo experiments. Fibers distributeprofusely in the entire stratum lacunosum moleculare (SLM) and the outer molecular layer (OML) without invasion of inappropriate layers (upper left panel).Similarly, when EC and HP tissue from mdab1-deficient mice where co-cultured, the pattern of entorhinal innervation reproduce that observed in vivo in mdab1(−/−) mice. Compacted fibers distribute close to the hippocampal fissure and misrouted fibers can be observed in inappropriate layers in CA1, CA3, and hilarregions (upper right panel). In mixed co-cultures a mixture of phenotypes was observed (lower panels). When HP is defective in mDab1, the wild-type EC axonsinnervate the hippocampus without fasciculation and exhibited ectopies in CA1 and CA3 regions; in the dentate gyrus, ectopic fibers innervate inappropriatelayers more profusely than in mdab1-deficient homogenic co-cultures (lower left panel). When EC axons defective in mDab1 innervate a wild-type hippocampalslice, fasciculated fibers distribute next to the hippocampal fissure, and ectopic axons invade the CA3 region and hilus (lower right panel).
169V. Borrell et al. / Mol. Cell. Neurosci. 36 (2007) 158–173
In contrast, axons originating from another region, those ofhippocampal neurons, respond to Reelin in a completely differentway. We never observed any fasciculation of hippocampal axons invivo in wild-type or in mutant mice. In wild-types, the associative/commissural axons originating in hippocampal neurons neverinnervate the SLM or OML layers, whereas in mutants (reeler andmdab1 −/−), there is evidence of ectopic innervation of those areas.Taken together, these data suggest that when Reelin is present andfunctional, hippocampal axons do not invade regions close to thehippocampal fissure, possibly because Reelin (expressed by Cajal–Retzius cells next to the fissure) is not a good substrate for growthof these axons. Indeed, in the in vitro treatments, neurons avoidedsurfaces which contain Reelin, and instead preferred to establishcontact with each other. Thus, we hypothesize that the effects ofReelin on axonal fasciculation depend on the neuronal type thatextends the axons.
In addition, our in vitro experiments show that Reelin affectsaxonal growth cones, axonal growth and axonal branching in hip-pocampal neurons. The data show that Reelin amplifies the numberof collapsed axonal growth cones, although to less extent than thatobserved in the hippocampus with classical chemorepellents such as
Sema3A or Sema3F (Pozas et al., 2001; Chedotal et al., 1998).Accordingly, axons growing on Reelin-coated coverslips extend forshorter distances than control fibers. The moderate growth conecollapse promoted by Reelin suggests a specific function of Reelinin a subset of neurons (e.g., depending on age) or as a function ofReelin concentration, as occurs with some Eph ligands (Wang andAnderson, 1997). However, the increase in axonal growth conecollapse induced by Reelin resembles effects of other axonal growthmodulators, such as the extracellular matrix protein Slit2, or contact-mediated inhibition by the proteoglycan NG2 (Nguyen Ba-Charvetet al., 1999; Ughrin et al., 2003). Indeed, our own experiments (datanot shown) using a typical chemorepulsion assay (i.e., confrontingaggregates of Reelin-expressing cells to hippocampal explants)corroborates those of a previous study (Jossin and Goffinet, 2001),thereby indicating that Reelin does not act as a classicalchemorepellent (e.g., class III Semaphorins) for hippocampal axons.
Furthermore, our data provide evidence that Reelin increasesaxonal branching in hippocampal neurons, which agrees with thereported role of Reelin in dendrite development in the hippocam-pus (Niu et al., 2004), and with the reeler and dab1 mutantphenotypes, which exhibit reduced axonal branching.

170 V. Borrell et al. / Mol. Cell. Neurosci. 36 (2007) 158–173
The formation of specific neural connections is a complex andsequential process implying several molecules. Thus, in addition toclassical chemorepellents and chemoattractants, other extracellularand membrane-anchored molecules regulate and/or modulateprocesses such as axonal extension, fasciculation, axonal branch-ing and synapse formation (Gerrow and El-Husseini, 2006; Akinsand Biederer, 2006; Hayashi et al., 2005; Chilton, 2006; Huberet al., 2003; Mueller, 1999). We propose here that, in addition to itswell-documented role in neuronal migration, Reelin acts as anegative regulator for the growth of mDab1-expressing axons,thereby modulating axonal targeting, adhesion and fasciculation, aswell as influencing axonal branching and synaptogenesis.
Experimental methods
Animals
OF1 mice (Charles River Laboratories) were used for the insitu hybridization and immunocytochemical studies, and for thein vitro explant assays. For tracing studies, two different coloniesof mdab1-mutant mice were used: scrambler homozygous andheterozygous mice were obtained by mating scrambler homo-zygous (scm/scm) males with heterozygous (+/scm) females.mdab1-mutant, heterozygous and wild-type mice were obtainedby mating heterozygous (+/−) mice. Genotypes were determinedas described (Howell et al., 1997b). Since mdab1-mutant mice dieat 3 weeks postnatal, the adult stage could only be analyzed usingscrambler animals, which survive through adulthood. reeler micewere obtained by mating reeler homozygous (rl/rl) males withheterozygous (+/rl) females. The day at which a vaginal plug wasdetected was considered embryonic day 0 (E0), and the day ofbirth was considered postnatal day 0 (P0).
The aforementioned experiments were performed in accordancewith the European Community Council Directive and the NationalInstitutes of Health guidelines for the care and use of laboratoryanimals.
In situ hybridization
Embryos from E14, E16 and E18 stages, and postnatal micefrom stages P0, P5, P10, P15, P21, and adults (two animals each),were perfused with 0.1 M phosphate-buffered (PB) containing 4%paraformaldehyde. The brains were postfixed overnight with thesame solution, cryoprotected with 30% sucrose, and sectionedeither coronally or horizontally at 25–50 μm. mdab1 and reelinantisense riboprobes were labeled with digoxigenin-D-UTP(Boehringer-Mannheim, Germany) by in vitro transcription of acDNA fragment encoding mouse mdab1 (Rice et al., 1998), ormouse reelin (D’Arcangelo et al., 1995) using a T3 polymerase(Ambion). In situ hybridization was performed essentially asdescribed elsewhere (Alcantara et al., 1998; Alvarez-Dolado etal., 1999). Sections were permeabilized in 0.2–0.5% Triton X-100(15 min), treated with 2% H2O2 (15 min), deproteinized with0.2 N HCl (10 min), fixed in 4% paraformaldehyde (10 min), andblocked in 0.2% glycine (5 min). Thereafter, sections wereprehybridized at 60 °C for 3 h in a solution containing 50%formamide, 10% dextran sulfate, 5×Denhardt’s solution, 0.62 MNaCl, 10 mM EDTA, 20 mM PIPES, pH 6.8, 50 mM DTT,250 μg/ml yeast tRNA, and 250 μg/ml denatured salmon spermDNA. Labeled antisense RNA was added to the prehybridizationsolution (500 ng/ml) and hybridization was carried out at 60 °C
overnight. Sections were then washed in 2× SSC (30 min, roomtemperature), digested with 20 μg/ml RNAse A (37 °C, 1 h), andthen washed in 0.5× SSC/50% formamide (4 h, 55 °C) and in 0.1×SSC/0.1% sarkosyl (1 h, 60 °C). Sections were blocked in 10%normal goat serum (2 h) and incubated overnight with an alkalinephosphatase-conjugated antibody to digoxygenin (BoehringerMannheim, 1:2000). After washing, sections were developed withnitroblue tetrazolium (NBT) and 5-bromo-4-chloro-3-indolylphosphate (BCIP), both purchased from Roche, then mountedon gelatin-coated slides and coverslipped with Mowiol.
Immunohistochemistry
Embryos from E14, E16, and E18 stages, postnatal mice stagesfrom stages P0, P5, P8, P15, and adults were perfused,cryoprotected and sectioned as above. Sections were blocked with10% goat serum for 2 h, and then incubated overnight at 4 °C withan anti-mDab1 rabbit polyclonal antibody (B3; dilution 1:800)(Howell et al., 1997a). Primary antibodies were visualized usingbiotinylated anti-rabbit antibodies (1:200), and a Streptavidin-biotinylated horseradish peroxidase complex (Amersham, LifeScience) (1:400). The peroxidase reaction was developed usingdiaminobenzidine (DAB) and H2O2 as substrates. Sections weremounted, dehydrated and coverslipped. Sections from wild-typeand reeler mice immunoreacted for mDab1 protein detection wereprocessed in bulk (identical incubation conditions and peroxidasereaction times).
Tracing of the entorhino-hippocampal and commissural pathwaysin vivo
Postnatal wild-type, heterozygous,mdab1-mutant mice obtainedfrom either scrambler (scm/scm) ormdab1-mutant (−/−) strains (P0:3 +/+, 7 +/−, 4 +/scm, 3 −/−, 1 scm/scm; P4: 9 +/+, 14 +/scm, 6 scm/scm; P16: 4 +/+, 6 +/−, 1 −/−) and adult mice from the Scramblerstrain (4 +/scm, 2 −/−) were iontophoretically injected withbiotinylated Dextran-Amine (BDA, MW=10.000; MolecularProbes) (+7.6 μA, 2 s On, 1 s Off) in the EC to trace theentorhino-hippocampal projection. Alternatively, P16 (4 +/+, 2+/−,4−/−) and adult mice (2 +/scm, 1 scm/scm) were injected in the CA3and hilus to trace the commissural connections. In addition, a similarcollection of postnatal and adult reeler (rl/rl) mice were also injectedin the EC and in the HP to compare their phenotype with that of themdab1-mutant mice. After 24–48 h of survival, animals wereperfused, cryoprotected and sectioned either coronally or horizon-tally (50 μm thick) as described above. After blocking endogenousperoxidase activity, sections were incubated with ABC complex anddeveloped with a DAB nickel-enhanced reaction (black reactionproduct); sections were counterstained with Nissl stain, dehydratedand coverslipped.
Explant and primary cultures
EC explants were obtained from E16 or P0 OF1 brains asdescribed elsewhere (Chedotal et al., 1998). Tissue pieces werethen embedded on collagen matrix as described elsewhere (Pozaset al., 2001) or plated on coated coverslips (poly-D-lysine/laminin(3 h) with Mock or Reelin containing supernatants as needed (1:10,3 h)). Explants were cultured for 2 days in a 5% CO2, 37 °C, 95%humidity incubator in Neurobasal medium supplemented with 1%horse serum, L-glutamine, NaHCO3, D-glucose, and B-27 supple-

171V. Borrell et al. / Mol. Cell. Neurosci. 36 (2007) 158–173
ment (all from GIBCO Life Technologies). Primary cultures fromhippocampal neurons were obtained from E16 mouse embryos.Brains were dissected in PBS containing 0.6% glucose, and weretrypsinized and mechanically dissociated. Cells were cultured in acollagen matrix or in coated coverslips and cultured for 3 DIV.Mock and Reelin supernatants used for coatings and treatmentswere produced as described (Simo et al., 2007) using a workingconcentration of Reelin of 2 ng/ml.
For immunochemistry and immunofluorescence, explants orprimary cultures were fixed in 4% paraformaldehyde for 40 minand washed several times with PBS 1×. The cultures were thenblocked with 10% goat serum for 2 h. For immunofluorescence,cultures were incubated overnight at 4 °C with an anti-mDab1rabbit polyclonal antibody (B3; dilution 1:800) (Howell et al.,1997a) and/or with Texas-red tagged phalloidin (1:500). Theprimary antibody was visualized using Alexa Fluor 488 F(ab′)2fragment of goat anti-rabbit IgG (H+L) (1:500; Molecularprobes), and the images were captured using an OlympusFluoview FV300 confocal microscope. For immunochemistry,cultures were incubated overnight at 4 °C with mouse anti-βTubulin isotype III (TUJ1) (1:3000; Covance). The primaryantibody was visualized using the same protocol described abovefor immunohistochemistry.
For Western Blot (WB), primary and explant cultures wereprocessed as described (Simo et al., 2007). We used anti-mDab1rabbit polyclonal antibody (B3; dilution 1:2000), anti-phosphotyr-osine (clone 4G10) (1:5000, Upstate), and anti-β Tubulin isotypeIII (TUJ1) (1:50,000; Covance). Secondary antibodies were fromDAKO (dilution 1:2500–1:10,000). For immunoprecipitation, weused anti-mDab1 (ExAlpha).
Outgrowth quantification from explants was performed bymeasuring the lengths of outgrowing axons, defined as the distancefrom the surface of the explant. For each explant, 10 measurementswere made (at least 15 explants per group). Results from three to fiveindependent experiments were standardized by comparing thepercentage of axonal outgrowth in Reelin-treated and controlexplants. The outgrowth of control explants was taken as 100%.
Quantification of axonal length from hippocampal cells wasperformed using IMAT software (developed by UB TechnicalServices), which measures total axonal length (i.e., of the mainaxonal branch plus collaterals). There were 60 neurons per group.We also quantified the number of branching points per neuron.Results were standardized by comparing the percentage of axonalarbor lengths and branching points in Reelin-treated and controlcultures.
Statistical significance was determined using the StatgraphicsPlus 4.0.
Organotypic co-culture experiments
Entorhino-hippocampal co-cultures were prepared from P0–P1pups essentially as described elsewhere (Frotscher and Heimrich,1993; Borrell et al., 1999a; Del Rio et al., 1997). The HP andprospective parietal neocortex were dissected out. Tissue pieceswere cut into transverse slices (325 μm thick) using a McIlwainTissue Chopper, and the slices were maintained in MinimumEssential Medium (MEM) supplemented with L-glutamine (2 mM)at 4 °C. Selected slices were co-cultured using the membraneinterface technique (Stoppini et al., 1991). The phenotype of thepups was determined by the cytoarchitectonics of the HP (Borrell etal., 1999a), and later confirmed by genotyping. Four types of
entorhino-hippocampal co-cultures were prepared: (1) Control co-cultures (EC+/+ or +/−/HP+/+ or +/−) (n=22), in which both slices werefrom wild-type or heterozygous pups; (2)mdab1-mutant co-cultures(EC−/−/HP−/−) (n=19); (3) mixed co-cultures using HP from (−/−)mice and EC from control mice (EC+/−/HP−/−) (n=31); and (4)mixed co-cultures combining control HP and (−/−) EC slices (EC−/−/HP+/−) (n=25). To assess the formation of entorhino-hippocampalconnections, a crystal of biocytin was injected into the entorhinalslice at 6 days in vitro. Co-cultures were fixed 24 h after injection ofthe tracer, with 4% paraformaldehyde in 0.1 M PB. After severalrinses, 40 μm thick notions were obtained, incubated with ABCovernight at 4 °C and developed with a nickel-enhanced DABreaction. Sections were counterstained with Nissl stain.
Acknowledgments
The authors would like to thank D. Goldowitz (Memphis) for thescrambler colony. We would also like to thank T. Yates and G.Y.Qushair for editorial assistance, and S. Maqueda andM. Cuñarro fortechnical assistance. This work was supported by grants SAF2005-00171 and ISCIII (CIBERNED-CB06/05/0098) to E. Soriano;BFU2006-13651 to J.A. Del Río; and National Cancer Institutegrant CA41072 to J.A. Cooper.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.mcn.2007.06.006.
References
Akins, M.R., Biederer, T., 2006. Cell–cell interactions in synaptogenesis.Curr. Opin. Neurobiol. 16, 83–89.
Alcantara, S., Ruiz, M., D'Arcangelo, G., Ezan, F., de Lecea, L., Curran, T.,Sotelo, C., Soriano, E., 1998. Regional and cellular patterns of reelinmRNA expression in the forebrain of the developing and adult mouse. JNeurosci. 18, 7779–7799.
Alvarez-Dolado, M., Ruiz, M., Del Rio, J.A., Alcantara, S., Burgaya, F.,Sheldon, M., Nakajima, K., Bernal, J., Howell, B.W., Curran, T.,Soriano, E., Munoz, A., 1999. Thyroid hormone regulates reelin anddab1 expression during brain development. J. Neurosci. 19, 6979–6993.
Arnaud, L., Ballif, B.A., Cooper, J.A., 2003. Regulation of protein tyrosinekinase signaling by substrate degradation during brain development.Mol. Cell. Biol. 23, 9293–9302.
Ballif, B.A., Arnaud, L., Cooper, J.A., 2003. Tyrosine phosphorylation ofDisabled-1 is essential for Reelin-stimulated activation of Akt and Srcfamily kinases. Brain Res. Mol. Brain Res. 117, 152–159.
Bar, I., Tissir, F., Lambert de Rouvroit, C., De Backer, O., Goffinet, A.M.,2003. The gene encoding disabled-1 (DAB1), the intracellular adaptor ofthe Reelin pathway, reveals unusual complexity in human and mouse.J. Biol. Chem. 278, 5802–5812.
Barallobre, M.J., Del Rio, J.A., Alcantara, S., Borrell, V., Aguado, F., Ruiz,M., Carmona, M.A., Martin, M., Fabre, M., Yuste, R., Tessier-Lavigne,M., Soriano, E., 2000. Aberrant development of hippocampal circuitsand altered neural activity in netrin 1-deficient mice. Development 127,4797–4810.
Barallobre, M.J., Pascual, M., Del Rio, J.A., Soriano, E., 2005. The Netrinfamily of guidance factors: emphasis on Netrin-1 signalling. Brain Res.Brain Res. Rev. 49, 22–47.
Beffert, U., Morfini, G., Bock, H.H., Reyna, H., Brady, S.T., Herz, J., 2002.Reelin-mediated signaling locally regulates protein kinase B/Akt andglycogen synthase kinase 3beta. J. Biol. Chem. 277, 49958–49964.

172 V. Borrell et al. / Mol. Cell. Neurosci. 36 (2007) 158–173
Bock, H.H., Herz, J., 2003. Reelin activates SRC family tyrosine kinases inneurons. Curr. Biol. 13, 18–26.
Bock, H.H., Jossin, Y., May, P., Bergner, O., Herz, J., 2004. Apolipoprotein Ereceptors are required for reelin-induced proteasomal degradation of theneuronal adaptor protein Disabled-1. J. Biol. Chem. 279, 33471–33479.
Borrell, V., Del Rio, J.A., Alcantara, S., Derer, M., Martinez, A., D'Arcangelo,G., Nakajima, K., Mikoshiba, K., Derer, P., Curran, T., Soriano, E., 1999a.Reelin regulates the development and synaptogenesis of the layer-specificentorhino-hippocampal connections. J. Neurosci. 19, 1345–1358.
Borrell, V., Ruiz, M., Del Rio, J.A., Soriano, E., 1999b. Development ofcommissural connections in the hippocampus of reeler mice: evidence ofan inhibitory influence of Cajal–Retzius cells. Exp. Neurol. 156,268–282.
Chedotal, A., Del Rio, J.A., Ruiz, M., He, Z., Borrell, V., de Castro, F., Ezan,F., Goodman, C.S., Tessier-Lavigne, M., Sotelo, C., Soriano, E., 1998.Semaphorins III and IV repel hippocampal axons via two distinctreceptors. Development 125, 4313–4323.
Chilton, J.K., 2006. Molecular mechanisms of axon guidance. Dev. Biol.292, 13–24.
Dahme, M., Bartsch, U., Martini, R., Anliker, B., Schachner, M., Mantei, N.,1997. Disruption of the mouse L1 gene leads to malformations of thenervous system. Nat. Genet. 17, 346–349.
D'Arcangelo, G., Miao, G.G., Chen, S.C., Soares, H.D., Morgan, J.I.,Curran, T., 1995. A protein related to extracellular matrix proteinsdeleted in the mouse mutant reeler. Nature 374, 719–723.
D'Arcangelo, G., Homayouni, R., Keshvara, L., Rice, D.S., Sheldon, M.,Curran, T., 1999. Reelin is a ligand for lipoprotein receptors. Neuron 24,471–479.
Del Rio, J.A., Heimrich, B., Super, H., Borrell, V., Frotscher, M.,Soriano, E., 1996. Differential survival of Cajal–Retzius cells inorganotypic cultures of hippocampus and neocortex. J. Neurosci. 16,6896–6907.
Del Rio, J.A., Heimrich, B., Borrell, V., Forster, E., Drakew, A., Alcantara,S., Nakajima, K., Miyata, T., Ogawa, M., Mikoshiba, K., Derer, P.,Frotscher, M., Soriano, E., 1997. A role for Cajal–Retzius cells andreelin in the development of hippocampal connections. Nature 385,70–74.
Demyanenko, G.P., Tsai, A.Y., Maness, P.F., 1999. Abnormalities in neuro-nal process extension, hippocampal development, and the ventricularsystem of L1 knockout mice. J. Neurosci. 19, 4907–4920.
Frotscher, M., Heimrich, B., 1993. Formation of layer-specific fiber projec-tions to the hippocampus in vitro. Proc. Natl. Acad. Sci. U. S. A. 90,10400–10403.
Gerrow, K., El-Husseini, A., 2006. Cell adhesion molecules at the synapse.Front. Biosci. 11, 2400–2419.
Gertler, F.B., Bennett, R.L., Clark, M.J., Hoffmann, F.M., 1989. Drosophilaabl tyrosine kinase in embryonic CNS axons: a role in axonogenesis isrevealed through dosage-sensitive interactions with disabled. Cell 58,103–113.
Gonzalez-Billault, C., Del Rio, J.A., Urena, J.M., Jimenez-Mateos, E.M.,Barallobre, M.J., Pascual, M., Pujadas, L., Simo, S., Torre, A.L, Gavin,R., Wandosell, F., Soriano, E., Avila, J., 2005. A role of MAP1B inReelin-dependent neuronal migration. Cereb Cortex. 15, 1134–1145.
Hayashi, N., Mizusaki, M.J., Kamei, K., Harada, S., Miyata, S., 2005.Chondroitin sulfate proteoglycan phosphacan associates with parallelfibers and modulates axonal extension and fasciculation of cerebellargranule cells. Mol. Cell. Neurosci. 30, 364–377.
Herz, J., Chen, Y., 2006. Reelin, lipoprotein receptors and synapticplasticity. Nat. Rev., Neurosci. 7, 850–859.
Howell, B.W.,Gertler, F.B., Cooper, J.A., 1997a.Mouse disabled (mDab1): a Srcbinding protein implicated in neuronal development. EMBOJ. 16, 121–132.
Howell, B.W., Hawkes, R., Soriano, P., Cooper, J.A., 1997b. Neuronalposition in the developing brain is regulated by mouse disabled-1.Nature 389, 733–737.
Howell, B.W., Herrick, T.M., Cooper, J.A., 1999a. Reelin-induced tryosinephosphorylation of disabled 1 during neuronal positioning. Genes Dev.13, 643–648.
Howell, B.W., Lanier, L.M., Frank, R., Gertler, F.B., Cooper, J.A., 1999b.The disabled 1 phosphotyrosine-binding domain binds to the internal-ization signals of transmembrane glycoproteins and to phospholipids.Mol. Cell. Biol. 19, 5179–5188.
Howell, B.W., Herrick, T.M., Hildebrand, J.D., Zhang, Y., Cooper, J.A.,2000. Dab1 tyrosine phosphorylation sites relay positional signalsduring mouse brain development. Curr. Biol. 10, 877–885.
Huber, A.B., Kolodkin, A.L., Ginty, D.D., Cloutier, J.F., 2003. Signaling atthe growth cone: ligand–receptor complexes and the control of axongrowth and guidance. Annu. Rev. Neurosci. 26, 509–563.
Jossin, Y., Goffinet, A.M., 2001. Reelin does not directly influence axonalgrowth. J. Neurosci. 21, RC183.
Keshvara, L., Benhayon, D., Magdaleno, S., Curran, T., 2001. Identificationof reelin-induced sites of tyrosyl phosphorylation on disabled 1. J. Biol.Chem. 276, 16008–16014.
Kuo, G., Arnaud, L., Kronstad-O'Brien, P., Cooper, J.A., 2005. Absence ofFyn and Src causes a reeler-like phenotype. J. Neurosci. 25, 8578–8586.
Marin-Padilla, M., 1998. Cajal–Retzius cells and the development of theneocortex. Trends Neurosci. 21, 64–71.
Mueller, B.K., 1999. Growth cone guidance: first steps towards a deeperunderstanding. Annu. Rev. Neurosci. 22, 351–388.
Nakajima, K., Mikoshiba, K., Miyata, T., Kudo, C., Ogawa, M., 1997.Disruption of hippocampal development in vivo by CR-50 mAb againstreelin. Proc. Natl. Acad. Sci. U. S. A. 94, 8196–8201.
Nguyen Ba-Charvet, K.T., Brose, K., Marillat, V., Kidd, T., Goodman, C.S.,Tessier-Lavigne, M., Sotelo, C., Chedotal, A., 1999. Slit2-mediatedchemorepulsion and collapse of developing forebrain axons. Neuron 22,463–473.
Niu, S., Renfro, A., Quattrocchi, C.C., Sheldon, M., D'Arcangelo, G., 2004.Reelin promotes hippocampal dendrite development through theVLDLR/ApoER2–Dab1 pathway. Neuron. 41, 71–84.
Ogawa, M., Miyata, T., Nakajima, K., Mikoshiba, K., 1997. The action ofreelin in the layering of cortical neurons in cerebrum. TanpakushitsuKakusan Koso 42, 577–583.
Pla, R., Borrell, V., Flames, N., Marin, O., 2006. Layer acquisition bycortical GABAergic interneurons is independent of Reelin signaling.J. Neurosci. 26, 6924–6934.
Pozas, E., Pascual, M., Nguyen Ba-Charvet, K.T., Guijarro, P., Sotelo, C.,Chedotal, A., Del Rio, J.A., Soriano, E., 2001. Age-dependent effects ofsecreted Semaphorins 3A, 3F, and 3E on developing hippocampalaxons: in vitro effects and phenotype of Semaphorin 3A (−/−) mice.Mol. Cell. Neurosci. 18, 26–43.
Rice, D.S., Curran, T., 2001. Role of the reelin signaling pathway in centralnervous system development. Annu. Rev. Neurosci. 24, 1005–1039.
Rice, D.S., Sheldon, M., D'Arcangelo, G., Nakajima, K., Goldowitz, D.,Curran, T., 1998. Disabled-1 acts downstream of Reelin in a signalingpathway that controls laminar organization in the mammalian brain.Development 125, 3719–3729.
Rice, D.S., Nusinowitz, S., Azimi, A.M., Martinez, A., Soriano, E., Curran,T., 2001. The reelin pathway modulates the structure and function ofretinal synaptic circuitry. Neuron 31, 929–941.
Ruth, R.E., Collier, T.J., Routtenberg, A., 1982. Topography between theentorhinal cortex and the dentate septotemporal axis in rats: I. Medialand intermediate entorhinal projecting cells. J. Comp. Neurol. 209,69–78.
Sanada, K., Gupta, A., Tsai, L.H., 2004. Disabled-1-regulated adhesion ofmigrating neurons to radial glial fiber contributes to neuronal positioningduring early corticogenesis. Neuron 42, 197–211.
Seki, T., Rutishauser, U., 1998. Removal of polysialic acid-neural celladhesion molecule induces aberrant mossy fiber innervation and ectopicsynaptogenesis in the hippocampus. J. Neurosci. 18, 3757–3766.
Sheldon, M., Rice, D.S., D'Arcangelo, G., Yoneshima, H., Nakajima, K.,Mikoshiba, K., Howell, B.W., Cooper, J.A., Goldowitz, D., Curran, T.,1997. Scrambler and yotari disrupt the disabled gene and produce areeler-like phenotype in mice. Nature 389, 730–733.
Simo, S., Pujadas, L., Segura, M.F., Torre, A.L., Del Rio, J.A., Urena, J.M.,Comella, J.X., Soriano, E., 2007. Reelin Induces the detachment of

173V. Borrell et al. / Mol. Cell. Neurosci. 36 (2007) 158–173
postnatal subventricular zone cells and the expression of the Egr-1through Erk1/2 activation. Cereb. Cortex 17, 294–303.
Soriano, E., Del Rio, J.A., 2005. The cells of Cajal–Retzius: still a mysteryone century after. Neuron 46, 389–394.
Soriano, E., Del Rio, J.A., Martinez, A., Super, H., 1994. Organization of theembryonic and early postnatal murine hippocampus. I. Immunocyto-chemical characterization of neuronal populations in the subplate andmarginal zone. J. Comp. Neurol. 342, 571–595.
Stanfield, B.B., Caviness Jr., V.S., Cowan, W.M., 1979. The organization ofcertain afferents to the hippocampus and dentate gyrus in normal andreeler mice. J. Comp. Neurol. 185, 461–483.
Stein, E., Savaskan, N.E., Ninnemann, O., Nitsch, R., Zhou, R., Skutella, T.,1999. A role for the Eph ligand ephrin-A3 in entorhino-hippocampalaxon targeting. J. Neurosci. 19, 8885–8893.
Steup, A., Ninnemann, O., Savaskan, N.E., Nitsch, R., Puschel, A.W.,Skutella, T., 1999. Semaphorin D acts as a repulsive factor for entorhinaland hippocampal neurons. Eur. J. Neurosci. 11, 729–734.
Steup, A., Lohrum, M., Hamscho, N., Savaskan, N.E., Ninnemann, O.,Nitsch, R., Fujisawa, H., Puschel, A.W., Skutella, T., 2000. Sema3C andnetrin-1 differentially affect axon growth in the hippocampal formation.Mol. Cell. Neurosci. 15, 141–155.
Stoppini, L., Buchs, P.A., Muller, D., 1991. A simple method for organotypiccultures of nervous tissue. J. Neurosci. Methods 37, 173–182.
Suetsugu, S., Tezuka, T.,Morimura, T., Hattori,M.,Mikoshiba, K.,Yamamoto,T., Takenawa, T., 2004. Regulation of actin cytoskeleton by mDab1through N-WASP and ubiquitination of mDab1. Biochem. J. 384, 1–8.
Super, H., Soriano, E., 1994. The organization of the embryonic and earlypostnatal murine hippocampus. II. Development of entorhinal, commis-sural, and septal connections studied with the lipophilic tracer DiI.J. Comp. Neurol. 344, 101–120.
Super, H., Martinez, A., Del Rio, J.A., Soriano, E., 1998. Involvement ofdistinct pioneer neurons in the formation of layer-specific connections inthe hippocampus. J. Neurosci. 18, 4616–4626.
Swanson, L.W., Cowan, W.M., 1977. An autoradiographic study of theorganization of the efferent connections of the hippocampal formation inthe rat. J. Comp. Neurol. 172, 49–84.
Tissir, F., Goffinet, A.M., 2003. Reelin and brain development. Nat. Rev.,Neurosci. 4, 496–505.
Trommsdorff, M., Gotthardt, M., Hiesberger, T., Shelton, J., Stockinger,W., Nimpf, J., Hammer, R.E., Richardson, J.A., Herz, J., 1999.Reeler/Disabled-like disruption of neuronal migration in knockoutmice lacking the VLDL receptor and ApoE receptor 2. Cell 97,689–701.
Ughrin, Y.M., Chen, Z.J., Levine, J.M., 2003. Multiple regions of the NG2proteoglycan inhibit neurite growth and induce growth cone collapse.J. Neurosci. 23, 175–186.
Wang, H.U., Anderson, D.J., 1997. Eph family transmembrane ligands canmediate repulsive guidance of trunk neural crest migration and motoraxon outgrowth. Neuron 18, 383–396.
Yabut, O., Renfro, A., Niu, S., Swann, J.W., Marin, O., D'Arcangelo, G.,2007. Abnormal laminar position and dendrite development ofinterneurons in the reeler forebrain. Brain Res. 1104, 75–83.




















