
Kainate Activation of Horizontal, Bipolar, Amacrine, and Ganglion Cells in the Rabbit Retina

Cell Reports
Report
Kainate Receptors Coexist in a FunctionalComplex with KCC2 and Regulate ChlorideHomeostasis in Hippocampal NeuronsVivek Mahadevan,1 Jessica C. Pressey,1 Brooke A. Acton,1 Pavel Uvarov,5 Michelle Y. Huang,1 Jonah Chevrier,1
Andrew Puchalski,1 Caiwei M. Li,1 Evgueni A. Ivakine,2 Matti S. Airaksinen,5 Eric Delpire,3 Roderick R. McInnes,2,4
and Melanie A. Woodin1,*1Department of Cell & Systems Biology, University of Toronto, Toronto, ON M5S 3G5, Canada2Hospital for Sick Children Research Institute, Toronto, ON M5G 1X8, Canada3Department of Anesthesiology, Vanderbilt University School of Medicine, Nashville, TN 37232, USA4Lady Davis Institute, Jewish General Hospital, McGill University, Montreal, QC H3T 1E2, Canada5Institute of Biomedicine, Anatomy, University of Helsinki, 00014 Helsinki, Finland
*Correspondence: [email protected]://dx.doi.org/10.1016/j.celrep.2014.05.022
This is an open access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/3.0/).
SUMMARY
KCC2 is the neuron-specific K+-Cl� cotransporterrequired for maintaining low intracellular Cl�, whichis essential for fast inhibitory synaptic transmissionin the mature CNS. Despite the requirement ofKCC2 for inhibitory synaptic transmission, under-standing of the cellular mechanisms that regulateKCC2 expression and function is rudimentary. Weexamined KCC2 in its native protein complex in vivoto identify key KCC2-interacting partners thatregulate KCC2 function. Using blue native-polyacryl-amide gel electrophoresis (BN-PAGE), we deter-mined that native KCC2 exists in a macromolecularcomplex with kainate-type glutamate receptors(KARs). We found that KAR subunits are requiredfor KCC2 oligomerization and surface expression.In accordance with this finding, acute and chronicgenetic deletion of KARs decreased KCC2 functionand weakened synaptic inhibition in hippocampalneurons. Our results reveal KARs as regulators ofKCC2, significantly advancing our growing under-standing of the tight interplay between excitationand inhibition.
INTRODUCTION
Hyperpolarizing GABAergic synaptic transmission in the mature
CNS depends upon a low concentration of intracellular Cl� [Cl�]iKCC2 is the neuron-specificmember of the K+-Cl� cotransporter
gene family that primarily extrudes Cl� from neurons, making it
essential for inhibitory synaptic transmission (Acton et al.,
2012; Blaesse et al., 2009; Rivera et al., 1999). Physiological
levels of neuronal activity can regulate KCC2 in a Ca2+-depen-
dent manner to induce inhibitory synaptic plasticity, which plays
a key role in the delicate balance between inhibition and excita-
tion (Fiumelli and Woodin, 2007; Lamsa et al., 2010; Woodin
1762 Cell Reports 7, 1762–1770, June 26, 2014 ª2014 The Authors
et al., 2003). However, aberrant KCC2 regulation results in
increased neuronal Cl� and contributes toward the pathophysi-
ology of numerous neurological disorders including epilepsy,
autism, and neuropathic pain (Coull et al., 2005; Kahle et al.,
2008; Tyzio et al., 2014; Woo et al., 2002).
KCC2 membrane expression and function are regulated by
multiple posttranslational mechanisms, including alterations in
phosphorylation state, oligomerization, association with lipid
rafts, and cleavage by proteases (Blaesse et al., 2006; Lee
et al., 2011; Puskarjov et al., 2012; Rinehart et al., 2009; Wata-
nabe et al., 2009). Recently, we made an important addition to
this list ofmechanisms that regulate KCC2 function by identifying
a KCC2-interacting protein termed Neto2 (Ivakine et al., 2013).
We found that Neto2 is required to maintain KCC2 abundance
in neurons and for efficient KCC2-mediated Cl� transport.
Thus, the KCC2-Neto2 interaction is vital for normal synaptic in-
hibition in mature neurons.
Neto2 is a CUB domain containing transmembrane protein
that also acts as an auxiliary subunit of native kainate-type gluta-
mate receptors (KARs). Neto2 regulates both the kinetics and
synaptic localization of KAR subunits (Copits et al., 2011; Tang
et al., 2012; Wyeth et al., 2014; Zhang et al., 2009). KARs are
unique ionotropic glutamate receptors that perform multiple
functions during synaptic transmission and plasticity (Lerma
and Marques, 2013). They regulate GABAergic release from
presynaptic terminals (Rodrıguez-Moreno et al., 1997), mediate
slow excitatory currents postsynaptically (Castillo et al., 1997),
and are involved in mossy fiber-pyramidal neuron long-term
potentiation in the CA3 area (Contractor et al., 2001).
Our identification of the Neto2-KCC2 interaction, coupled with
the previous demonstrations that Neto2 is an auxiliary subunit of
KARs, led us to ask whether KCC2 and KARs coexist in a macro-
molecular complex. In particular, we examined the role of GluK2
subunits that were previously shown to interact with Neto2
(Copits et al., 2011; Tang et al., 2011; Zhang et al., 2009). In
this study, we have made a surprising discovery that native
oligomeric KCC2 coexists in an ensemble with the GluK2 KAR
subunit in the CNS. Moreover, we determined that KARs
are required to maintain both KCC2 oligomerization and the

Figure 1. KCC2 Interacts with GluK2 KARs
in Mouse Brain and in Heterologous Cells
(A) Native KCC2 complexes fromC12E9-solubilized
whole-brain membrane fractions immunoprecipi-
tated with anti-KCC2 and immunoblotted with the
antibodies indicated at right (KCC2, GluK2/3,
Neto2). Representative example of three inde-
pendent replicates. IP, immunoprecipitate; I, input
fraction (1% of IP); U. unbound fraction (1% of IP);
O. oligomer; M. monomer; also see Figure S1A.
(B) (Bi) Coimmunoprecipitation experiments per-
formed in HEK293 cells transfected with KCC2 and
KAR subunits, solubilized in RIPA buffer, immu-
noprecipitated with anti-KCC2, and immuno-
blotted with the antibodies indicated at right
(KCC2, GluK2/3, myc); also see Figures S1B and
S1C. Representative example of three to four in-
dependent biological replicates (Bii) Quantitation
of the bound fractions to KCC2 was performed by
measuring the band intensity of the immunopre-
cipitated fraction compared with total input (10%)
using ImageJ software.
(C) Confocal images of DIV 12-14 cultured mouse
hippocampal neurons immunostained for endog-
enous KCC2 (left, green) and GluK1/2 (middle,
red), demonstrating that the two proteins are co-
localized (right, yellow). Representative of confocal
images obtained from 26 neurons over four inde-
pendent experiments performed using eight cov-
erslips. (Scale bars, 10 mm.) Bottom inset is a
magnification from the primary dendrite indicated
in the box.
All summary figures represent mean ± SEM. *p <
0.05, **p < 0.01, ***p < 0.001.
expression of this transporter in the membrane. When we per-
formed an electrophysiological characterization of KCC2 func-
tion following KAR subunit disruption, we found neurons had
a depolarized reversal potential for GABA (EGABA). Hence, our
findings represent a regulation of KCC2 function and fast synap-
tic inhibition by components of excitatory transmission.
RESULTS
KCC2 and GluK2 KARs Interact In Vivo and In VitroWe have recently discovered that KCC2 binds to the single-pass
CUB domain protein Neto2, and that this interaction is required
for efficient Cl� extrusion in hippocampal neurons (Ivakine
et al., 2013). Several groups have previously established that
Neto2 is a critical auxiliary subunit of native KARs, including
GluK2 (Copits et al., 2011; Tang et al., 2011; Wyeth et al.,
2014; Zhang et al., 2009). This led us to hypothesize that KARs
Cell Reports 7, 1762–177
might be a putative candidate that
could interact with KCC2. In order to
determine whether KCC2 interacts
with KAR subunits in vivo, we performed
a coimmunoprecipitation assay from
whole-brain native membrane prepara-
tions. We found that anti-KCC2 anti-
bodies coimmunoprecipitated GluK2/3
primarily from wild-type mice in comparison to KCC2b+/�
mice, indicating the existence of a KCC2-KAR complex in vivo
(Figures 1A and S1A; n = 3). To determine whether KCC2 can
interact with KARs independent of exogenous Neto2, we per-
formed coimmunoprecipitation experiments in HEK293 cells
transfected with KCC2 and KAR subunits alone. In this assay,
we found that KCC2 could coimmunoprecipitate GluK2, but
not GluK1 (Figures 1B and S1B; n = 4). We also performed the
experiment in the reverse direction and found that GluK2, but
not GluK1, could also robustly coimmunoprecipitate KCC2 (Fig-
ure S1C; n = 3). Based on the interaction of KCC2 and GluK2 in
these coimmunoprecipitation experiments, we hypothesized
that these two proteins would colocalize in neurons. We tested
this hypothesis by performing immunofluorescent staining of
endogenous proteins using antibodies specific for KCC2
and GluK1/2 in cultured hippocampal neurons, followed by
quantitative colocalization. Hippocampal neurons showed
0, June 26, 2014 ª2014 The Authors 1763

Figure 2. Native KCC2 and KARs Exist in the
Same Oligomeric Complex
(A) One-dimensional BN-PAGE separation of
native KCC2 and GluK2/3 complexes from P30
mouse brain solubilized with Digitonin or C12E9; gel
separations were immunoblotted with the anti-
bodies indicated above (KCC2, GluK2/3). This blot
is a representative example of three independent
biological replicates. Native KCC2 is present as a
putative tetramer (blue arrow head), and as higher-
order protein complexes (green arrow heads).
Native GluK2/3 migrates in a similar molecular
weight range as native KCC2 (dotted yellow lines);
also see Figure S2A.
(B) (Bi) Antibody-shift assay followed by two-
dimensional BN-PAGE separation using C12E9-
solubilized whole-brain membrane fractions,
incubated with antibodies for HA or GluK2/3;
samples resolved first in a 5% 1D-BN-PAGE, and
individual lanes from the first dimension were
separated in a 6% 2D-SDS-PAGE); gel separa-
tions were immunoblotted with anti GluK2/3
antibody. (Bii and Biii) Similar to (Bi), but using
digitonin-solubilized hippocampal membrane
fractions and antibodies targeted to immunoglob-
ulin G or GluK2/3. Samples resolved in a 4% 1D-
BN-PAGE and 6% SDS-PAGE. Gel separations
were immunoblotted with KCC2 and Neto2 anti-
bodies respectively. Bottom panels in Bi, Bii, Biii
represent densitometric profiles and area under
the densitogram to indicate similar loading; red
arrowheads denote antibody-induced shifts in the
proteins indicated. Representative example of
three independent biological replicates; also see
Figures S2B–S2D.
immunofluorescence for both endogenous KCC2 and KARs,
with a partial colocalization of these two proteins (Figure 1C;
n = 26). We then performed an intensity correlation analysis to
quantitate the colocalization and calculated a Pearson’s correla-
tion coefficient of 0.61 ± 0.03 between the GluK1/2 and KCC2
immunofluorescent signals, indicating �60% colocalization of
these proteins. Thus, we have discovered a surprising protein
interaction between KCC2 and the predominant kainate receptor
subunit, GluK2.
Native KCC2 Exists in a Hetero-Oligomeric Ensemblewith Native KARsFunctional KCC2 predominantly exists as an oligomer in the
mature brain (Blaesse et al., 2006; Ivakine et al., 2013; Uvarov
et al., 2009). Consistently, we observed that native KCC2 exist
as a putative tetramer and in higher-order complexes above
400 kDa (Figures 2A and S2A; n = 3), in P30 whole-brain mem-
brane lysates using a one-dimensional blue native polyacryl-
amide gel electrophoresis (1D-BN-PAGE). Similar to native
KCC2, we also observed higher-order complexes of comparable
molecular weights that contained native GluK2/3 (Figures 2A and
S2A). Next, to determine whether native KCC2 exists in an
ensemble with KAR subunits, we employed an antibody-shift
assay coupled with two-dimensional blue native polyacrylamide
1764 Cell Reports 7, 1762–1770, June 26, 2014 ª2014 The Authors
gel electrophoresis (2D-BN-PAGE). The 2D-BN-PAGE strategy
has been previously employed to examine the native assemblies
of AMPA receptor multimeric complex (Schwenk et al., 2009).
Using this approach, we first verified that the addition of
the GluK2/3 antibody could shift GluK2/3 to higher molecular
weights (Figure 2Bi; n = 3). We observed that this antibody-
induced shift in GluK2/3 also shifted a population of native
KCC2 in hippocampal preparations (Figure 2Bii; n = 3). Using
the same experimental strategy, we found that KCC2 antibodies
could also shift a population of GluK2/3 in hippocampal prepara-
tions (Figure S2C; n = 3), a finding that we also observed
in digitonin-solubilized cortical membrane preparations (Fig-
ure S2D; n = 2). As a positive control for this assay, we probed
for Neto2, because this protein interacts with both KCC2
and GluK2 (Ivakine et al., 2013; Tang et al., 2011; Zhang et al.,
2009). As expected, we found that Neto2 could be shifted
with both GluK2/3 (Figure 2Biii) and KCC2 antibodies (Fig-
ure S2D). We confirmed the specificity of these interactions in
this assay by repeating the experiments using antibodies for
the transferrin receptor and observed that antibodies to this
receptor did not shift GluK2/3 (Figure S2B). Thus, we established
that functional oligomeric KCC2 coexists in a hetero-oligomeric
complex with the predominant KAR subunit GluK2 in hippocam-
pus and cortex.

Figure 3. GluK1/2-Null Hippocampal
Neurons Have an Increased Monomeric:
Oligomeric KCC2 Ratio
(A) Representative immunoblots of C12E9-solubi-
lized native lysates prepared in the presence of
25 mM iodoacetamide to prevent the formation of
nonspecific disulphide bonds between KCC2
monomers during membrane extraction, from
wild-type and GluK1/2-null hippocampi; resolved
in a standard 6% SDS-PAGE; immunoblotted with
the antibodies indicated at left (KCC2, NKCC1,
GluK2/3, ATP1A3, Tuj1). Also see Figure S3Ai.
(B) Samples obtained from the same preparation
as in (A) were resolved in the absence of DTT
in nondenaturing PFO-PAGE conditions. Red
arrowhead indicates the oligomeric KCC2 band
migrating above �400 kDa that is predominantly
reduced in GluK1/2�/� lysates; blots shown in (A)
and (B) are representative of three independent
biological replicates, see Figures S3Aii and S4A.
(C) Summary figures showing levels of (Ci) KCC2
monomers, (Cii) KCC2 oligomers, (Ciii) total KCC2,
and (Civ) monomer:oligomer KCC2 ratio in GluK1/
2-null hippocampal homogenates relative to that of
wild-type.
(D) (Di) Example confocal microscopic immuno-
fluorescent images of cultured hippocampal neu-
rons from wild-type and GluK1/2-null mice stained
with anti-KCC2 antibody (red; n = 49 neurons).
(Scale bars, 24 mm.) (Dii) Summary of the average
fluorescence intensity of somatic KCC2 in wild-
type and GluK1/2-null neurons.
All summary figures represent mean ± SEM. *p <
0.05, **p < 0.01, ***p < 0.001.
KARs Regulate the Assembly or Stability of Native KCC2OligomersIf oligomeric KCC2 exists in a complex with GluK2, and oligo-
meric KCC2 is the functional form of this transporter, this raises
the possibility that KARs may play an important role in the regu-
lation of KCC2 oligomers. To test this possibility, we examined
the ratio of KCC2 monomers to oligomers in native membranes
prepared from wild-type and GluK1/2-null hippocampal lysates
under conditions preventing the formation of nonspecific disul-
phide bonds (Uvarov et al., 2009). In an SDS-PAGE, we observed
an increase in monomeric KCC2 and a decrease in oligomeric
KCC2 in GluK1/2-null hippocampi (Figures 3A and S3Ai; n = 3).
We also examined the levels of the neuron-specific pump
ATP1A3 and another KCC2-family member NKCC1, and
observed no significant difference between wild-type and
GluK1/2-null lysates (Figure 3A). We reasoned that the dena-
turing gel running conditions of SDS-PAGE could impede a
robust quantification of KCC2 oligomeric levels, so we subse-
Cell Reports 7, 1762–177
quently chose to resolve changes in the
monomer:oligomer ratio using a previ-
ously established nondenaturing PFO-
PAGE (Uvarov et al., 2009). Using these
conditions, we observed a 2-fold increase
in monomeric KCC2 in GluK1/2-null
hippocampi (Figures 3B, 3Ci, and S3Aii;
n = 3) as expected from the previous SDS-PAGE result. In addi-
tion, we also observed a significant decrease in oligomeric KCC2
levels above �400 kDa in GluK1/2-null hippocampi compared
with wild-type levels (Figures 3B, 3Cii, and S3Aii), and no signifi-
cant change in total KCC2 levels under the same conditions (Fig-
ure 3Ciii). We verified that there is no change in total KCC2 levels
by an additional standard approach by preparing the samples in
the absence of iodoacetamide, and resolved them under strong
denaturing conditions, indicating that there is no net change in
total KCC2 levels (Figure S3Aiii; n = 3). Additionally, we verified
that the increases inmonomeric KCC2 levels were not accompa-
nied by changes in KCC2 gene expression, by examining the
relative KCC2 mRNA abundance using quantitative real-time
PCR. We found no significant differences between wild-type
and GluK1/2-null hippocampi prepared from postnatal day 30
mice (Figure S3B; n = 3). Put together, these results demonstrate
that the presence of GluK1/2 determines the monomer:oligomer
ratio of KCC2 (Figure 3B and 3Civ) at the posttranscriptional
0, June 26, 2014 ª2014 The Authors 1765

Figure 4. GluK1/2-Null Neurons Have
Deceased KCC2 Expressed at the Surface
(A) (Ai) Representative immunoblots of KCC2
monomers and oligomers from the surface and
internal fraction of wild-type and GluK1/2-null
neurons. The first two lanes correspond to bio-
tinylated surface proteins (50 mg) recovered from
the neutravidin beads. The last two lanes corre-
spond to unbiotinylated internal proteins (5 mg)
recovered from the supernatant. (Aii–v) Summary
figures showing levels of surface KCC2 mono-
mers, surface KCC2 oligomers, total surface KCC2
(normalized to surface levels of neuron-specific
ATP1A3), and internal KCC2 (normalized to b-actin
levels) in GluK1/2-null homogenates relative to that
of wild-type (n = 5).
(B) (Bi) Example confocal microscopic immuno-
fluorescent images from a single confocal plane of
live cultured hippocampal neurons overexpressing
KCC2-HA from wild-type and GluK1/2-null mice
(also see Figure S4B). Neuronal transfection is
indicated by GFP fluorescence, KCC2-HA is indi-
cated by red fluorescent signal. Summary of the
(Bii) average number of anti-HA puncta/neuron,
(Biii) percentage of anti-HA puncta proximal to the
surface (Biv) mean puncta size (surface area, mm2),
and (Bv) puncta intensity (in arbitrary units x1000) in
wild-type and GluK1/2-null neurons. (Bvi) Total
immunofluorescence of live imaging of wild-type
and GluK1/2-null neurons. n = 23 neurons.
All summary figures represent mean ± SEM. *p <
0.05, **p < 0.01, ***p < 0.001.
level. Moreover, by showing that loss of KAR subunits induces a
significant reduction in KCC2 oligomers, particularly above
�400 kDa, we further strengthen our two major claims: (1) the
existence of a KAR:KCC2 hetero-oligomeric complex in the hip-
pocampus, and (2) KARs promote the assembly or the stability of
KCC2 oligomers within the complex.
KARs Maintain KCC2 Surface Expression in NeuronsIt has been previously demonstrated by several groups that
immature neurons are characterized by predominantly mono-
meric KCC2 that exists intracellularly (reviewed in Chamma
et al., 2012). Based on these previous demonstrations, we exam-
ined whether there were differences in the expression patterns of
KCC2 between wild-type and GluK1/2�/� neurons. We first
1766 Cell Reports 7, 1762–1770, June 26, 2014 ª2014 The Authors
examined total KCC2 levels by per-
forming immunofluorescent staining of
cultured hippocampal neurons with anti-
KCC2 antibody. We found a significant
increase in endogenous KCC2 immuno-
reactivity in the soma of GluK1/2�/�
neurons compared to wild-type neurons
(wild-type, n = 47; GluK1/2�/�, n = 49;
p < 0.001; Figure 3D). However, for
KCC2 to be functional it needs to be
expressed in the membrane. Next, we
examined the membrane expression
pattern of KCC2 by performing a surface biotinylation assay at
4�C. We found a significant decrease in monomeric (p =
0.029), oligomeric (p = 0.029), and total (p = 0.008) KCC2 levels
in the surface of GluK1/2�/� neurons compared to wild-type
neurons, with a corresponding increase in internal monomeric
KCC2 (p = 0.008; n = 5; Figure 4A). To visualize KCC2 expres-
sion, we performed live immunofluorescence of KCC2 contain-
ing an extracellular HA tag under nonpermeabilizing conditions
at 37�C. We have previously demonstrated that this KCC2-HA
chimeric protein traffics to the membrane and is functional (Ac-
ton et al., 2012). We found that in GluK1/2�/� neurons there
was a significant decrease the number of anti-HA puncta (Fig-
ure 4Bii; n = 23), with no significant differences in either the
size or intensity of the puncta (Figures 4Biv and 4Bv; n = 23),

Figure 5. Both Genetic Deletion and Acute
Knockdown of GluK2 KAR Subunits Depo-
larizes EGABA
(A) Example IV curves measuring EGABA from
cultured hippocampal neurons from wild-type (left)
and GluK1/2�/� (middle) mice. Summary of EGABA
obtained from all similar IV curves (right).
(B) Example IV curves measuring EGABA from
cultured hippocampal neurons from wild-type
transfected with scrambled shRNA (left) and
shRNA for GluK2 (middle). Summary of EGABA ob-
tained from all similar IV curves.
(C) Summary of EGABA recordings performed in the
absence (control) and presence of the GluK2/5-
KAR inhibitor (5 mM UBP 310). Solid circles are
individual EGABA measurements, open triangles
represent the mean ± SEM. Insets: are raw voltage
clamp traces from example recordings.
Scale bars, 20 pA, 50 ms. Arrow indicates onset of
GABA puff. Dashed vertical lines indicate where
the current amplitudes were obtained for the
generation of the IV curves. All summary figures
represent mean ± SEM. *p < 0.05, **p < 0.01, ***p <
0.001.
indicating an overall decrease in the number of KCC2-HA
puncta, with no difference in the puncta characteristics them-
selves. We then analyzed where the existing puncta were
located and found that there was a significant decrease in the
KCC2-HA puncta that were proximal to the membrane in
GluK1/2�/� neurons (Figure 4Biii and S3; n = 23). We made two
important observations from these biotinylation and immuno-
staining experiments: (1) overall there is an increased total
KCC2 immunoreactivity in soma of GluK1/2�/� neurons (Fig-
ure 3D); and (2) GluK1/2�/� neurons have a decreased mem-
brane expression (Figure 4A). Thus, in addition to promoting
and/or stabilizing KCC2 oligomers, the presence of KAR sub-
units GluK1/2 also promotes and/or stabilizes surface KCC2
levels.
Genetic Deletion and Acute Silencing of KAR SubunitsResult in Depolarized EGABA
Because we observed a decrease in both oligomeric KCC2 and
surface KCC2 in GluK1/2-null neurons, we hypothesized that
these neurons would have aberrant KCC2-mediated Cl� homeo-
Cell Reports 7, 1762–177
stasis. To test this hypothesis, we mea-
sured KCC2 activity by recording the
reversal potential for GABA (EGABA) using
gramicidin-perforated patch clamp re-
cordings. We found that EGABA was depo-
larized in cultured GluK1/2�/� hippocam-
pal neurons (�65.2 ± 1.9 mV; n = 10)
compared to wild-type neurons (�78.6 ±
3.5; n = 7; p = 0.003; Figure 5A), with no
significant difference in either synaptic
conductance (p = 0.5) or maximum cur-
rent amplitude (p = 0.6; Table S2). We
then took a two-step approach to rule
out the possibility that the depolarization
of EGABA in GluK1/2�/� neurons was due to differences in activity
levels between the genotypes. First, we compared the sponta-
neous activity levels using Ca2+ imaging and found no differ-
ences between cultured hippocampal neurons prepared from
wild-type (C57/Bl6 and 129SVE) and GluK1/2�/� mice (Fig-
ure S5A). Second, we used small hairpin RNA (shRNA) to acutely
silence GluK2 in cultured hippocampal neurons; we verified
GluK2 shRNA was effective at silencing GluK2 but not KCC2 us-
ing an in vitro assay (Figure S5B). We found that knocking down
GluK2 in wild-type neurons (C57/Bl6) depolarized EGABA by
15.41 mV compared to neurons transfected with scrambled
shRNA (Figures 5B and S5B; n = 11). Knocking down GluK2 in
129SVE wild-type neurons also significantly depolarized EGABA
(n = 11; p < 0.05). Similar to our results from cultured GluK1/
2�/� hippocampal neurons above, we found no significant differ-
ence in either synaptic conductance (p = 0.6) or maximum cur-
rent amplitude (p = 0.1) for either genotype (Table S2). Last, we
considered the possibility that the depolarization in EGABA we re-
corded following the genetic deletion and acute silencing of KAR
subunits was not due to the loss of the protein, but rather was
0, June 26, 2014 ª2014 The Authors 1767

due to the loss of the GluK2-KAR current. We tested this possi-
bility by recording EGABA in wild-type neurons in the presence
and absence of the GluK2/5-KAR antagonist UBP 310 (5 mM)
(Pinheiro et al., 2013).We found no significant difference in EGABA
following this pharmacological blockade of these receptors (n =
5; p = 0.159; Figure 5C), allowing us to conclude that it is the
protein interaction between GluK2-KARs and KCC2 that is
required to maintain a high KCC2 function.
DISCUSSION
Overall, we have three compelling lines of evidence supporting
the conclusion that KCC2 and KAR subunits coexist in a
complex: (1) in vivo and in vitro coimmunoprecipitation, (2)
antibody-shift assay coupled with 2D-BN-PAGE, and (3) immu-
nofluorescence. Thus, the key K+-Cl� cotransporter required
for fast synaptic inhibition binds to the predominant KAR-type
ionotropic glutamate receptor subunit GluK2 in multiple brain re-
gions including hippocampus. Furthermore, we find that the
GluK2:KCC2 interaction occurs predominantly with oligomeric
KCC2, and the importance of this preferential binding is to main-
tain the functional oligomeric KCC2 complex. Along with our
evidence that both genetic deletion and acute knockdown of
GluK2 result in aberrant KCC2-dependent neuronal Cl� extru-
sion, we have identified kainate receptors as an unexpected
player during neuronal Cl� homeostasis.
Functional KCC2 Exists as a Hetero-OligomericComplex with KARsSeveral studies have established that functional KCC2 predom-
inantly exists as oligomers in mature neurons (Blaesse et al.,
2006; Ivakine et al., 2013; Uvarov et al., 2009; Watanabe et al.,
2009). Here, we report using BN-PAGE that the majority of olig-
omeric KCC2 in mature brain migrates above 400 kDa. The key
finding of this study is that GluK2 is a member of this KCC2-het-
eromeric complex. This finding raises an important question: is
this complex exclusive to KCC2 and GluK2, or do these com-
plexes also include Neto2 and other proteins? This question is
relevant because we recently identified that Neto2 associates
with oligomeric KCC2 (Ivakine et al., 2013). Our current data
demonstrate that, whereas KCC2 and Neto2 interact, Neto2 is
not required for the GluK2:KCC2 interaction because GluK2
and KCC2 can interact in the absence of exogenous Neto2 in
heterologous cells. Despite the fact that Neto2 is not required
for the GluK2:KCC2 interaction, this does not discount the pos-
sibility that these three proteins commonly exist in a heteromeric
complex. Moreover, there is another reason to believe that addi-
tional proteins may also coexist in this heteromeric complex. For
example, the 4.1N protein has been identified as binding part-
ners of both KCC2 (Li et al., 2007) and more recently GluK2
(Copits and Swanson, 2013). Because the 4.1 family of FERM
domain proteins are abundant scaffolds between membrane
and cytoskeletal proteins (Baines et al., 2013), it is possible to
speculate that the GluK2:KCC2 interaction we have identified
could be mediated/stabilized by neuronal FERM domain pro-
teins. Interestingly, while this manuscript was in revision another
group discovered that the dwell time of KCC2 in the vicinity of
excitatory synapses is determined by its interactions with 4.1N
1768 Cell Reports 7, 1762–1770, June 26, 2014 ª2014 The Authors
(Chamma et al., 2013), further strengthening our claims about
the putative participation of the 4.1N protein within the KCC2:
KAR hetero-oligomeric complex.
KCC2 Oligomerization and Surface Expression Dependon GluK2-KARsWe found that GluK1/2-null hippocampal neurons have a
decrease in KCC2 oligomers, particularly above 400 kDa. This
suggests that GluK2 plays a role in either the formation of
KCC2 oligomers and/or regulates their stability, but how might
this happen? The answer may lie in the fact that GluK2-null
hippocampi have an �50% reduction in Neto2 protein levels
(Figures 4A and S4A), and we know that Neto2 is required for
KCC2 oligomerization and the efficacy of KCC2 transport (Iva-
kine et al., 2013). Does this mean that GluK2 regulates KCC2
via only Neto2, or can it directly regulate KCC2 function? Our
current results suggest that GluK2 can also regulate KCC2 inde-
pendently of Neto2. We found that in GluK1/2�/� neurons there
was an increase in KCC2 monomers and a decrease in KCC2
oligomers, with no net change in total KCC2 levels. This is in
contrast to our previous finding that Neto2-null neurons have
an overall decrease in both monomeric and oligomeric KCC2,
which results in a total decrease in KCC2 protein levels (Ivakine
et al., 2013).
Results from our biotinylation experiments allowed us to
conclude that GluK1/2�/� neurons have a decrease in cell-sur-
face KCC2. Thus, in addition to promoting or stabilizing KCC2
oligomers GluK1/2-KARs also maintain and/or stabilize surface
KCC2 levels. We supported these findings by immunostaining
for KCC2; using standard fixed immunofluorescence, we found
an increase in somatic KCC2 levels, whereas live immunofluo-
rescence of KCC2-HA showed a decrease in the number of
anti-KCC2-HA puncta. Although anti-KCC2-HA puncta must
have been present on the surface at some point during the
experiment, we cannot conclude that our puncta analysis repre-
sents only membrane expressed protein. At the temperature
these experiments were performed (37�C), we would expect
some proportion of KCC2-HA would be endocytosed, especially
considering that KCC2 membrane turnover has been reporter to
be relatively high (Lee et al., 2010, but see also Puskarjov et al.,
2012). Thus, the KCC2 puncta we have quantified may represent
a combination of KCC2-HA in the membrane and endocytotic
vesicles. Thus, although we are able to conclude that the loss
of GluK2-KARs decreases KCC2 membrane expression, future
studies should address whether GluK2 also regulates KCC2
membrane turnover. Together, the increase in KCC2 monomers
and decrease in membrane expression suggests that GluK1/
2�/� neurons resemble immature neurons, which have an abun-
dance of cytoplasmic KCC2monomers (Gulyas et al., 2001). Our
electrophysiological data support this observation of an imma-
ture Cl� homeostasis phenotype, where the genetic deletion of
GluK1/2 is reminiscent of immature neurons with poor Cl� extru-
sion, suggesting that kainate receptors are an essential compo-
nent of mature neuronal Cl� homeostasis.
Conclusions and Future SignificanceThe significance of our findings are manifold, but most impor-
tantly: (1) KCC2 and KARs exist in the same macromolecular

complex, and (2) an ionotropic glutamate receptor can positively
regulate the function of the predominant neuronal Cl� cotrans-
porter KCC2. These findings have important implications for
both normal physiological functions of neuronal networks and
for pathophysiological conditions that result from dysfunction
of KARs and KCC2. At the physiological level, we have demon-
strated that both genetic deletion and acute knockdown of
GluK2 weakens synaptic inhibition, suggesting that the co-
existence of these proteins provides a nexus for the ongoing
maintenance of the excitatory-inhibitory balance. At the patho-
physiological levels, both KCC2 and KARs are strongly impli-
cated in neurological disorders, including neuropathic pain
(Bhangoo and Swanson, 2013), autism (Tyzio et al., 2014), and
epilepsy (Woo et al., 2002). This raises the possibility that the
disruption of the KCC2:KAR complex may underlie these neuro-
physiological disorders. Understanding the fundamental molec-
ular pathways that regulate the cell intrinsic excitation, inhibition
homeostasis is essential for designing of better therapeutic stra-
tegies for diseases.
EXPERIMENTAL PROCEDURES
Animals and Approvals
All experiments were performed in accordance with approval and guidelines
from the University of Toronto Animal Care Committee and the Canadian
Council on Animal Care. Animals of both sexes were used to prepare hippo-
campal cultures; all other experiments were performed on male mice. The
following animal species were used:
d Wild-type C57/Bl6 (Charles River Laboratories)
d Wild-type 129/SV (Charles River Laboratories)
d GluK1/2�/� maintained on a mixed 129SV/C57Bl6 background
d KCC2+/� maintained on a mixed 129SV/C57Bl6 background
Antibodies
See Table S1 for complete details for all antibodies used in this study.
Biochemistry and Molecular Biology
See Supplemental Experimental Procedures for coimmunoprecipitation anal-
ysis, PFO-PAGE, BN-PAGE, antibody-shift assay, PCR, surface biotinylation.
Hippocampal Cultures and Electrophysiology
Low-density cultures of dissociated mouse hippocampal neurons were
prepared as previously described (Acton et al., 2012). Experiments were
performed after 10–13 days in culture. Gramicidin perforated patch clamp re-
cordings were performed as previously described (Acton et al., 2012). See
Supplemental Experimental Procedures for details on culturing and electro-
physiology, shRNA, and neuronal transfection.
Immunostaining, Confocal Microscopy, Ca2+ Imaging
Live immunostaining was performed as described before (Acton et al., 2012).
See Supplemental Experimental Procedures for details on fixed and live immu-
nostaining, confocal microscopy, and Ca2+ imaging.
Statistics
Results are given as mean ± SEM. See Supplemental Experimental Proce-
dures for details on statistical tests used for individual figures.
SUPPLEMENTAL INFORMATION
Supplemental Information includes Supplemental Experimental Procedures,
five figures, and two tables and can be found with this article online at http://
dx.doi.org/10.1016/j.celrep.2014.05.022.
C
AUTHOR CONTRIBUTIONS
V.M. and M.A.W. designed the study, performed experiments, analyzed data,
and wrote the paper. J.C.P., B.A.A., P.U., M.Y.H., J.C., A.P., and C.M.L. per-
formed experiments. V.M., E.A.I., M.S.A., E.D., R.R.M., and M.A.W. inter-
preted data, critically analyzed, and edited the manuscript.
ACKNOWLEDGMENTS
We thank Dr. Derek Bowie (McGill University) for the GluK2-GFP cDNA, Dr.
Sari Lauri (University of Helsinki) for the GluK1b-myc cDNA, Dr. David B.Mount
for KCC4 cDNA, anti-KCC4 antibody, and Dr. Chris McBain (NIH, Bethesda)
for the GluK1/2�/� mice. We thank Dr. Mike Salter (University of Toronto) for
helpful insight throughout the project. We thank Dr. Nivetha Ramachandran
and Dr. Sakthi Devi Moorthy for technical advice. In addition, we thank Ella
Czerwinska for excellent technical support. This study was supported the
following funding agencies: Canadian Institutes of Health Research (CIHR)
grant to M.A.W.; NIH grant (GM074771) to E.D.; The Academy of Finland
grants to P.U. and M.S.A.
Received: May 24, 2013
Revised: March 31, 2014
Accepted: May 9, 2014
Published: June 5, 2014
REFERENCES
Acton, B.A., Mahadevan, V., Mercado, A., Uvarov, P., Ding, Y., Pressey, J.,
Airaksinen, M.S., Mount, D.B., and Woodin, M.A. (2012). Hyperpolarizing
GABAergic transmission requires the KCC2 C-terminal ISO domain.
J. Neurosci. 32, 8746–8751.
Baines, A.J., Lu, H.C., and Bennett, P.M. (2013). The Protein 4.1 family: Hub
proteins in animals for organizing membrane proteins. Biochim. Biophys.
Acta. 1838, 605–619.
Bhangoo, S.K., and Swanson, G.T. (2013). Kainate receptor signaling in pain
pathways. Mol. Pharmacol. 83, 307–315.
Blaesse, P., Guillemin, I., Schindler, J., Schweizer, M., Delpire, E., Khiroug, L.,
Friauf, E., and Nothwang, H.G. (2006). Oligomerization of KCC2 correlates with
development of inhibitory neurotransmission. J. Neurosci. 26, 10407–10419.
Blaesse, P., Airaksinen, M.S., Rivera, C., and Kaila, K. (2009). Cation-chloride
cotransporters and neuronal function. Neuron 61, 820–838.
Castillo, P.E., Malenka, R.C., and Nicoll, R.A. (1997). Kainate receptors
mediate a slow postsynaptic current in hippocampal CA3 neurons. Nature
388, 182–186.
Chamma, I., Chevy, Q., Poncer, J.C., and Levi, S. (2012). Role of the neuronal
K-Cl co-transporter KCC2 in inhibitory and excitatory neurotransmission.
Front. Cell. Neurosci. 6, 5.
Chamma, I., Heubl, M., Chevy, Q., Renner, M., Moutkine, I., Eugene, E.,
Poncer, J.C., and Levi, S. (2013). Activity-dependent regulation of the K/Cl
transporter KCC2 membrane diffusion, clustering, and function in hippocam-
pal neurons. J. Neurosci. 33, 15488–15503.
Contractor, A., Swanson, G., and Heinemann, S.F. (2001). Kainate receptors
are involved in short- and long-term plasticity at mossy fiber synapses in the
hippocampus. Neuron 29, 209–216.
Copits, B.A., and Swanson, G.T. (2013). Kainate receptor post-translational
modifications differentially regulate association with 4.1N to control activity-
dependent receptor endocytosis. J. Biol. Chem. 288, 8952–8965.
Copits, B.A., Robbins, J.S., Frausto, S., and Swanson, G.T. (2011). Synaptic
targeting and functional modulation of GluK1 kainate receptors by the auxiliary
neuropilin and tolloid-like (NETO) proteins. J. Neurosci. 31, 7334–7340.
Coull, J.A., Beggs, S., Boudreau, D., Boivin, D., Tsuda, M., Inoue, K., Gravel,
C., Salter, M.W., and De Koninck, Y. (2005). BDNF from microglia causes
the shift in neuronal anion gradient underlying neuropathic pain. Nature 438,
1017–1021.
ell Reports 7, 1762–1770, June 26, 2014 ª2014 The Authors 1769

Fiumelli, H., and Woodin, M.A. (2007). Role of activity-dependent regulation
of neuronal chloride homeostasis in development. Curr. Opin. Neurobiol. 17,
81–86.
Gulyas, A.I., Sık, A., Payne, J.A., Kaila, K., and Freund, T.F. (2001). The KCl
cotransporter, KCC2, is highly expressed in the vicinity of excitatory synapses
in the rat hippocampus. Eur. J. Neurosci. 13, 2205–2217.
Ivakine, E.A., Acton, B.A., Mahadevan, V., Ormond, J., Tang, M., Pressey, J.C.,
Huang, M.Y., Ng, D., Delpire, E., Salter, M.W., et al. (2013). Neto2 is a KCC2
interacting protein required for neuronal Cl- regulation in hippocampal
neurons. Proc. Natl. Acad. Sci. USA 110, 3561–3566.
Kahle, K.T., Staley, K.J., Nahed, B.V., Gamba, G., Hebert, S.C., Lifton, R.P.,
and Mount, D.B. (2008). Roles of the cation-chloride cotransporters in neuro-
logical disease. Nat. Clin. Pract. Neurol. 4, 490–503.
Lamsa, K.P., Kullmann, D.M., and Woodin, M.A. (2010). Spike-timing depen-
dent plasticity in inhibitory circuits. Front. Synaptic Neurosci. 2, 8.
Lee, H.H., Jurd, R., and Moss, S.J. (2010). Tyrosine phosphorylation regulates
the membrane trafficking of the potassium chloride co-transporter KCC2. Mol.
Cell. Neurosci. 45, 173–179.
Lee, H.H., Deeb, T.Z., Walker, J.A., Davies, P.A., andMoss, S.J. (2011). NMDA
receptor activity downregulates KCC2 resulting in depolarizing GABAA recep-
tor-mediated currents. Nat. Neurosci. 14, 736–743.
Lerma, J., and Marques, J.M. (2013). Kainate receptors in health and disease.
Neuron 80, 292–311.
Li, H., Khirug, S., Cai, C., Ludwig, A., Blaesse, P., Kolikova, J., Afzalov, R.,
Coleman, S.K., Lauri, S., Airaksinen, M.S., et al. (2007). KCC2 interacts with
the dendritic cytoskeleton to promote spine development. Neuron 56, 1019–
1033.
Pinheiro, P.S., Lanore, F., Veran, J., Artinian, J., Blanchet, C., Crepel, V.,
Perrais, D., and Mulle, C. (2013). Selective block of postsynaptic kainate
receptors reveals their function at hippocampal mossy fiber synapses. Cereb.
Cortex 23, 323–331.
Puskarjov, M., Ahmad, F., Kaila, K., and Blaesse, P. (2012). Activity-dependent
cleavage of the K-Cl cotransporter KCC2 mediated by calcium-activated
protease calpain. J. Neurosci. 32, 11356–11364.
Rinehart, J., Maksimova, Y.D., Tanis, J.E., Stone, K.L., Hodson, C.A., Zhang,
J., Risinger, M., Pan, W., Wu, D., Colangelo, C.M., et al. (2009). Sites of
regulated phosphorylation that control K-Cl cotransporter activity. Cell 138,
525–536.
Rivera, C., Voipio, J., Payne, J.A., Ruusuvuori, E., Lahtinen, H., Lamsa, K.,
Pirvola, U., Saarma, M., and Kaila, K. (1999). The K+/Cl- co-transporter
1770 Cell Reports 7, 1762–1770, June 26, 2014 ª2014 The Authors
KCC2 renders GABA hyperpolarizing during neuronal maturation. Nature
397, 251–255.
Rodrıguez-Moreno, A., Herreras, O., and Lerma, J. (1997). Kainate receptors
presynaptically downregulate GABAergic inhibition in the rat hippocampus.
Neuron 19, 893–901.
Schwenk, J., Harmel, N., Zolles, G., Bildl, W., Kulik, A., Heimrich, B., Chisaka,
O., Jonas, P., Schulte, U., Fakler, B., and Klocker, N. (2009). Functional prote-
omics identify cornichon proteins as auxiliary subunits of AMPA receptors.
Science 323, 1313–1319.
Tang, M., Pelkey, K.A., Ng, D., Ivakine, E., McBain, C.J., Salter, M.W., and
McInnes, R.R. (2011). Neto1 is an auxiliary subunit of native synaptic kainate
receptors. J. Neurosci. 31, 10009–10018.
Tang, M., Ivakine, E., Mahadevan, V., Salter, M.W., and McInnes, R.R. (2012).
Neto2 interacts with the scaffolding protein GRIP and regulates synaptic abun-
dance of kainate receptors. PLoS ONE 7, e51433.
Tyzio, R., Nardou, R., Ferrari, D.C., Tsintsadze, T., Shahrokhi, A., Eftekhari, S.,
Khalilov, I., Tsintsadze, V., Brouchoud, C., Chazal, G., et al. (2014). Oxytocin-
mediated GABA inhibition during delivery attenuates autism pathogenesis in
rodent offspring. Science 343, 675–679.
Uvarov, P., Ludwig, A., Markkanen, M., Soni, S., Hubner, C.A., Rivera, C., and
Airaksinen, M.S. (2009). Coexpression and heteromerization of two neuronal
K-Cl cotransporter isoforms in neonatal brain. J. Biol. Chem. 284, 13696–
13704.
Watanabe,M.,Wake, H.,Moorhouse, A.J., and Nabekura, J. (2009). Clustering
of neuronal K+-Cl- cotransporters in lipid rafts by tyrosine phosphorylation.
J. Biol. Chem. 284, 27980–27988.
Woo, N.S., Lu, J., England, R., McClellan, R., Dufour, S., Mount, D.B., Deutch,
A.Y., Lovinger, D.M., and Delpire, E. (2002). Hyperexcitability and epilepsy
associated with disruption of the mouse neuronal-specific K-Cl cotransporter
gene. Hippocampus 12, 258–268.
Woodin, M.A., Ganguly, K., and Poo, M.M. (2003). Coincident pre- and post-
synaptic activity modifies GABAergic synapses by postsynaptic changes in
Cl- transporter activity. Neuron 39, 807–820.
Wyeth, M.S., Pelkey, K.A., Petralia, R.S., Salter, M.W., McInnes, R.R., and
McBain, C.J. (2014). Neto auxiliary protein interactions regulate kainate and
NMDA receptor subunit localization at mossy fiber-CA3 pyramidal cell synap-
ses. J. Neurosci. 34, 622–628.
Zhang,W., St-Gelais, F., Grabner, C.P., Trinidad, J.C., Sumioka, A., Morimoto-
Tomita, M., Kim, K.S., Straub, C., Burlingame, A.L., Howe, J.R., and Tomita, S.
(2009). A transmembrane accessory subunit that modulates kainate-type
glutamate receptors. Neuron 61, 385–396.

SUPPLEMENTAL INFORMATION
Kainate receptors co-exist in a functional complex with KCC2 and regulate chloride
homeostasis in hippocampal neurons
Vivek Mahadevan, Jessica C. Pressey, Brooke A. Acton, Pavel Uvarov, Michelle Y. Huang,
Jonah Chevrier, Andrew Puchalski, Caiwei M. Li, Evgueni A. Ivakine, Matti S. Airaksinen, Eric
Delpire, Roderick R. McInnes, Melanie A. Woodin

Supplementary Figure 1, Related to Figure 1
Wildtype KCC2b+/-
O
M
1 2 3 4 5 6
I IP U I IP U
250
130 IP:
KC
C2
60
100
KC
C2
G
luK
2/3
N
eto
2
a
b KCC2
GluK2
GluK1b-myc
- - -
KC
C2
G
luK
1/2
IP:
KC
C2
INP
UT
+ - -
- + -
- - +
+ + -
+ - +
- - -
+ - -
- + -
- - +
+ + -
+ - +
c KCC2
GluK2-FLAG
GluK1b-myc
- + + + - - + - - - - +
KC
C2
G
luK
2
myc
INP
UT
- + + + - - + - - - - +
IP:
Glu
K

b
app
MDa 1.0 0.72 0.48 1.2
Tra
nsfe
rrin
100
Glu
K2/3
Supplementary Figure 2, Related to Figure 2
a
1.05
1.24
0.48
0.72
0.15
0.24
~MDa
GluK2/3
Dig
i
C12E
9
DD
M
0.066
Dig
i
C12E
9
KCC2
DD
M
Pu
tative
tetr
am
eric
KC
C2
Hig
he
r-o
rde
r
ma
cro
mo
lecula
r
KC
C2
kDa
IgG
K
CC
2
100
100
Glu
K2/3
60
60 Neto
2
IgG
K
CC
2
ci
IgG
K
CC
2
130
130
KC
C2
1.0 0.72 0.48
~MDa 1.2
~kDa
cii
IgG
K
CC
2
100
Glu
K2/3
1.0 0.72 0.48
~MDa 1.2
~kDa
100
BN-PAGE (1D)
SD
S-P
AG
E (
2D
)
BN-PAGE (1D) d
1.0 0.72 0.48
~MDa 1.2

Supplementary Figure 3, Related to Figure 3
aii
WT
Glu
K1
/2-/
-
KC
C2
(o
lig
o)
KC
C2
(m
on
o)
WT
Glu
K1
/2-/
-
WT
Glu
K1
/2-/
-
130
400
~kDa
250
WT
Glu
K1/2
-/-
KC
C2
(o
lig
o)
KC
C2
(m
on
o)
130
250
~kDa
400
ai
aiii WT
Glu
K1/2
-/-
KC
C2
(to
tal)
Tu
j1
50
130
~kDa
130
NK
CC
1
(to
tal)
bi bii
WT GluK1/2-/-
KC
C2
G
AP
DH
WT
qR
TP
CR
- K
CC
2 m
RN
A
(% o
f W
T le
ve
ls,
no
rma
lis
ed
to
GA
PD
H)
P = 0.736
0%
20%
40%
60%
80%
100%
120%
Fig. Solubilisation
Sample
processing
Gel
running
S3ai
C12E9 buffer +
25mM IAA
SDS-SB, 100mM
DTT, 37*C
SDS-
PAGE
S3aii
C12E9 buffer +
25mM IAA
PFO-SB
(cold)
PFO-
PAGE
S3aiii
RIPA buffer
without IAA
SDS-SB, 100mM
DTT, 100*C
SDS-
PAGE
aiv

Supplementary Figure 4, Related to Figure 4
Ne
to2
Tu
j1
kDa
60
50
Neto
2 p
rote
in l
eve
ls
no
rma
lis
ed
to
Tu
j1
0
0.2
0.4
0.6
0.8
1
1.2
**
WT
ai aii
b
Supplementary Figure 5, Related to Figure 5
0
20
40
60
80
100
Ac
tive
Neu
ron
s (
%)
ai aii
WT
129SVE
GluK1/2
null
WT
C57Bl6
bii
KCC2
GluK2
Scrambled shRNA
GluK2 shRNA#3
+
+
+
-
+
-
+
+
-
- + +
KCC2
GluK2
β-actin
KCC4
myc
β-actin
KCC4
GluK1-myc
Scrambled shRNA
GluK2 shRNA#3
+
+
+
-
+
-
+
+
-
- + +
bi
*
ns
Glu
K2 l
eve
ls n
orm
ali
se
d t
o a
cti
n
120
100
80
60
40
20
0
GluK2
cDNA+
scrambled
shRNA
GluK2
cDNA+
GluK2
shRNA#3
GluK2
cDNA
Glu
K2
Glu
K2
+ s
cra
mb
led
Glu
K2
+ s
hR
NA
3
Em
pty
GluK2
β-actin

KCC2 and KARs Co-Exist in a Complex Mahadevan et al
Supplemental Material
7
Supplemental Figure 1 (Related to Figure 1): KCC2 interacts with GluK2 KARs in
the mouse brain and in heterologous cells. (a) Higher exposure of Fig. 1a showing reduced levels of KCC2 oligomers
immunoprecipitated in KCC2b+/-
lysate and decreased levels of Neto2 and GluK2/3 co-
precipitated (as indicated by the dotted line yellow boxes). (b) Same experiment as Fig.
1b, but the blots were stripped and re-probed with antibody raised against GluK1/2. This
is especially important because the GluK2/3 antibody that was used in our study was
previously indicated to bind non-specifically to a small proportion of AKAP8 (Zhang et
al., 2009 Neuron 61: 385-396). This experiment reconfirms our observation that GluK2,
but not GluK1 is co-immunoprecipitated with KCC2, regardless of antibody used. (c) Co-
immunoprecipitation experiments performed in HEK-293 cells transfected with KCC2
and KAR subunits (similar to experiment in Fig. 1b), immunoprecipitated with anti-
GluK1 and GluK2, and immunoblotted with the antibodies indicated at right (KCC2,
GluK2/3, myc).
Supplemental Figure 2 (Related to Figure 2): Native KCC2 and KARs exist in the
same oligomeric complex.
(a) Native KCC2 exists as monomers, dimers, trimers and tetramers in addition to higher
order macromolecular complexes. Higher exposure of Fig. 2a, including a third native
detergent dodecyl maltoside (DDM), indicating the existence of multiple higher order
macromolecular assemblies (above ~400kDa) in vivo. (b) Antibody-shift assay
demonstrating that antibodies for the transferrin receptor did not shift GluK2/3. (c)
Antibody-shift assay similar to (Figure 2bii) using digitonin-solubilized hippocampal
membrane fractions, but with antibodies targeted to IgG or KCC2. Representative
example of three independent biological replicates; (d) Antibody-shift assay (similar to
Supp Fig. 2c) with antibodies targeted to KCC2 or IgG, but using digitonin-solubilised
cortical membrane preparations. Representative example of two independent biological
replicates.
Supplemental Figure 3 (Related to Figure 3): GluK1/2-null hippocampal neurons
have an increased monomeric:oligomeric KCC2 ratio. (ai) Higher exposure of Fig. 3a indicating the presence of a higher-order KCC2 oligomer
migrating above ~400kDa, in wild-type lane (red arrow) but undetectable in GluK1/2-/-
lane. (aii) Individual biological triplicates corresponding to Fig. 3b. Lanes used for Fig.
3b are highlighted by dotted lines. (aiii) Total KCC2 levels in RIPA solubilized
hippocampal membranes. (aiv) Schematic representation of the experimental conditions
in figures S3ai, S3aii and S3aiii. (bi) Quantitative RT-PCR of KCC2 mRNA levels
normalized to GAPDH levels, and (bii) semi-quantitative KCC2 mRNA levels from WT
and GluK1/2-/-
hippocampal lysates.
Supplemental Figure 4 (Related to Figures 3 and 4): Neto2 levels are decreased in
GluK1/2-/-
hippocampal lysates. (ai) Analysis of Neto2 protein expression in C12E9 solubilized GluK1/2
-/- hippocampal
membranes. (aii) Summary figure showing a reduction in Neto2 protein levels in
GluK1/2-/-
hippocampal lysates, after normalizing with respective tuj1 level (n=4).
Summary figure represents mean ± sem. *P < 0.05, **P < 0.01, ***P < 0.001. (b)

KCC2 and KARs Co-Exist in a Complex Mahadevan et al
Supplemental Material
Example of analysis performed to determine the location of the puncta relative to the
surface (as determined by the edge of the GFP). Vectors are emanating from the centroid
and passing through individual puncta.
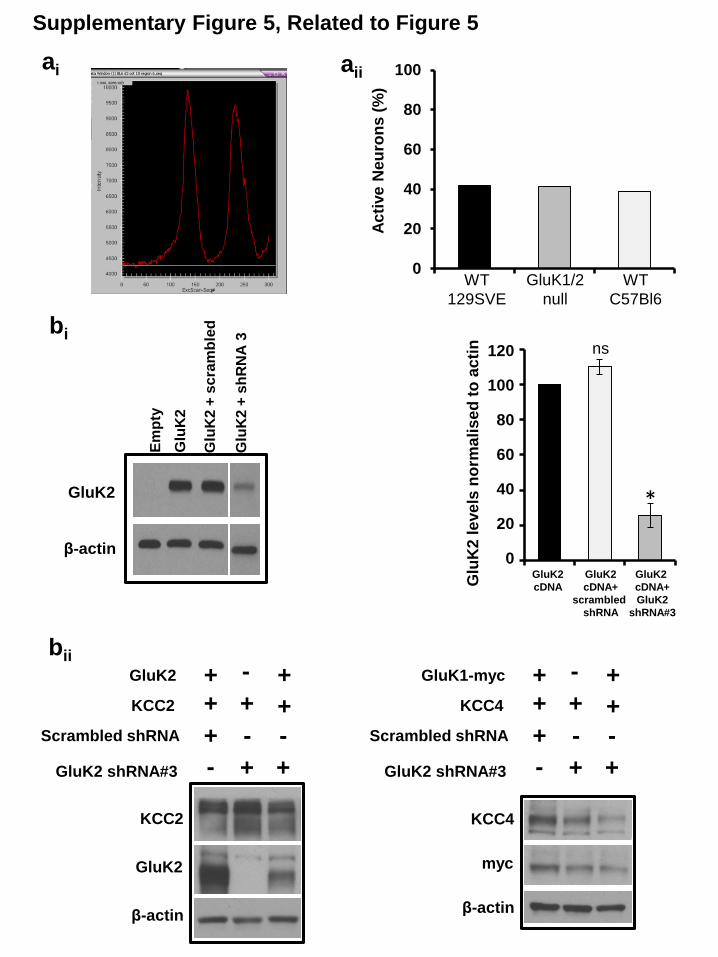
Supplemental Figure 5 (Related to Figure 5): Neuronal activity patterns are the
same in wild-type and GluK1/2 KO neurons. (ai) Sample image of the analysis window used to quantify neuronal activity. When the
Fluo4-AM intensity increased over 2000 arbitrary units during the acquisition period the
neuron was scored as active. (aii) Summary of all analysis similar to (ai) demonstrating
the % of neurons active in two different strains of wild-type neurons (129SVE and
C57/Bl6) and GluK1/2KO neurons. Analysis was performed on: WT 129SVE n = 115;
GluK1/2KO n = 64; WT C57/Bl6 n = 123. N values are a combination of three separate
experimental repetitions. (bi) Efficiency of knockdown of exogenous GluK2 using
scrambled shRNA and mouse-GluK2 shRNA #3 in HEK293 cells. Histogram represents
densitometric analysis of GluK2 levels upon shRNA co-transfection, normalized to
GluK2 transfections alone (n=4). (bi) GluK2 shRNA#3 does not non-specifically silence
exogenous expression of GluK1, KCC2 and KCC4 in HEK293 cells (n=3). GluK2-
shRNA#3 was subsequently used for electrophysiological experiments in Fig 5b.
Summary figure represents mean ± sem. *P < 0.05, **P < 0.01, ***P < 0.001
8

KCC2 and KARs Co-Exist in a Complex Mahadevan et al
Supplemental Material
9
SUPPLEMENTAL EXPERIMENTAL PROCEDURES
Animals & approvals: All experiments were performed in accordance with approval and
guidelines from the University of Toronto Animal Care Committee and the Canadian
Council on Animal Care. Animals of both sexes were used to prepare hippocampal
cultures; all other experiments were performed on male mice. The following animal
species were used:
Wild-type C57/Bl6 (Charles River; Wilmington, MA)
Wild-type 129/SV (Charles River; Wilmington, MA)
GluK1/2-/-
maintained on a mixed 129SV/C57Bl6 background (obtained from
Chris McBain, NIH)
KCC2+/-
maintained on a mixed 129SV/C57Bl6 background (provided by Eric,
Delpire, Vanderbilt Univ)
Antibodies: See Supplemental Table 1 for complete details for all antibodies used in this
study.
RNA extraction, cDNA synthesis, and analysis of KCC2 expression by real-time PCR:
RNA was isolated from hippocampi of ~30 -old male wild-type and GluK1/2−/−
mice
using TRIzol (Invitrogen Life Technologies) according to the manufacturer’s
instructions. cDNA was synthesized using Omniscript reverse transcriptase (Qiagen)
according to the manufacturer’s instructions, using 2 μg of total RNA. 30, 60, 120ng of
cDNA samples were use in quantitative real-time PCR. cDNA samples were amplified in
triplicate using SYBR Green PCR Master Mix (Applied Biosystems) and detected with
ABI/PRISM 7900 HT cycler (Applied Biosystems). Primer Express, version 2.0,
software (Applied Biosystems) was used for primer design: 5′-
GCCATTCACAGCTTCCCCAAACTTG-3′ (KCC2 reverse), 5′-
CCACCTCTGCTGTCTACATCAGCTC- 3′ (KCC2 forward), 5′-
GCAAAGTGGAGATTGTTGCCAT- 3′ [glyceraldehyde-3-phosphate dehydrogenase
(GAPDH) forward], 5′-CCTTGACTGTGCCGTTGAATTT-3′ (GAPDH reverse).
Experiment in Fig. S3 are representative results from 3 independent sample preparations
from each genotype.
Native-Co-IP analysis: All biochemical preparations and centrifugations were performed
at 4 °C. Native-membrane fractions were prepared similarly to previously described
(Ivakine et al., 2013). Mouse brains (P30, male) were homogenized on ice in PBS using a
glass-Teflon homogenizer, followed by a brief low-speed centrifugation. Soft pellets
were resuspended in ice-cold lysis buffer [Tris·HCl, 50 mM, pH 7.4; EDTA, 1 mM;
protease and phosphatase inhibitor mixture (Roche)], homogenized, and centrifuged for
30 minutes at 25,000 × g. The membrane pellets were resuspended in solubilization
buffer (4Xw/v) [Tris·HCl, 50 mM, pH 7.4; NaCl, 150 mM; EDTA, 0.05 mM; 1.5%
C12E9, and protease and phosphatase inhibitor mixture], solubilized for 3 hours on a
rotating platform at 4 °C, and centrifuged for 1 hour at 25,000 × g. For KCC2-
coimmunoprecipitation, membrane fractions (∼0.5 –1.5 mg protein) were incubated with
anti-KCC2 (clone N1/12) (4 hours at 4 °C) with rotation, and subsequently incubated
with 20μl GammaBind IgG beads (1 hour at 4 °C). Following antibody binding to beads,

KCC2 and KARs Co-Exist in a Complex Mahadevan et al
Supplemental Material
10
the beads were washed once with solubilization buffer containing 1.5% C12E9, once with
solubilization buffer minus C12E9. Bound proteins (IP fraction) were eluted with SDS
sample buffer containing DTT (1 hour min at 37 °C) and they were separated along with
1% of input fraction and 1% of unbound fraction in a standard 8% SDS-PAGE, followed
by Western immunoblotting. Experiments in Fig. 1a are representative results from 3
independent biological replicates.
HEK-293-Co-IP analysis: For co-IP experiments, HEK-293 cells were transfected with
KCC2, GluK2, GluK1b-myc (0.25 μg each) using Lipofectamine (Invitrogen) at 70%
confluency. Thirty-six hours after transfection, cells were washed with ice-cold 1× PBS
and lysed in modified RIPA buffer [50 mM Tris·HCl, pH 7.4, 150 mM NaCl, 1 mM
EDTA, 1% Nonidet P-40, 0.1% SDS, 0.5% DOC, and protease inhibitors (Roche)].
Lysed cells were incubated on ice for 30 min and were centrifuged at 15,000 × g for 15
min at 4 °C. Cell lysates or solubilized membrane fractions (∼0.2 – 0.5 mg protein) were
incubated with Rb GluK1 and GluK2/3 antibodies (or anti-KCC2) on a rotating platform
(4 h, 4 °C). Lysates were subsequently incubated with 20μl GammaBind IgG beads (GE
Healthcare) on a rotating platform (1 h at 4 °C). After incubation, beads were washed
twice with modified RIPA buffer, and twice with modified RIPA buffer minus
detergents,. Bound proteins were eluted with SDS sample buffer and subjected to
SDS/PAGE along with 10% of input fraction and immunoblotted. Experiments in Fig.
1b, S1c are representative results from 4 and 3 independent biological replicates
respectively.
PFO-PAGE for resolving monomeric:oligomeric KCC2 ratio: All biochemical
preparations, centrifugations and PFO-PAGE were performed at 4 °C as described
(Uvarov et al., 2009) with modifications. Briefly, the membrane fractions from the
hippocampi of P30 – 60, male, WT and GluK1/2-/-
mice samples were prepared as above
using 1.5% C12E9- based solubilization buffer supplemented with 25mM iodoacetamide
to prevent the formation of artificial di-sulphide bonds. The 2x sample buffer contained
8% (w/v) sodium perfluoro-octanoate (NaPFO, Fluorochem Ltd., Derbyshire, UK),
0.004% bromphenol blue, 20% (v/v) glycerol, and 100 mM Tris base. Proteins were
separated using 5% and 7% Criterion Tris-HCl gels (Bio-Rad) in ice-cold PFO-glycine
buffer (25 mM Tris, 192 mM glycine, 0.5% NaPFO, pH 8.5), followed by Western
immunoblotting. Experiments in Fig. 3a, b, S3b are representative results from 3
independent biological replicates.
Antibody-shift assay & BN-PAGE analysis: All biochemical preparations,
centrifugations and 1D-BN-PAGE were performed at 4 °C. Native-membrane fractions
were prepared similarly as described (Swamy et al., 2006) with minor modifications.
Briefly, mouse brain / cortex / hippocampus (P30) were homogenized on ice in PBS
using a glass-Teflon homogenizer, followed by a brief low-speed centrifugation. Soft
pellets were resuspended in ice-cold lysis buffer [Bis-tris, 20mM, pH7; EDTA, 0.2mM;
sucrose, 300mM; protease and phosphatase inhibitor mixture (Roche)], homogenized,
and centrifuged for 30 minutes at 25,000 × g. The membrane pellets were resuspended in
solubilization buffer (4x w/v) [Bis-tris, 20mM, pH7; 6-aminocaproic acid, 500mM; NaCl,
20mM; EDTA, 0.2mM; glycerol, 10%; iodoacetamide, 25mM; 1% dodecyl-maltoside or

KCC2 and KARs Co-Exist in a Complex Mahadevan et al
Supplemental Material
1% digitonin or 1.5% C12E9; protease and phosphatase inhibitor mixture], solubilized for
3 hours on a rotating platform at 4 °C, and centrifuged for 1 hour at 25,000 × g. Ten to
twenty µg of proteins and native MARK unstained protein standard (Invitrogen) were
mixed with BN-PAGE sample buffer (bis-Tris, 200mM; 6-aminocaproic acid, 1M;
Coomassie blue G250, 5%; sucrose, 30%; iodoacetamide, 25mM) and loaded on a linear
3-15% precast native bis-Tris gel (Invitrogen). Samples were separated in a 1D-BN-
PAGE as described (Swamy et al., 2006), using separate cathode buffer (bis-Tris, 15 mM,
pH7; Tricine, 50 mM; Coomassie blue G250, 0.02%), and anode buffer (bis-Tris 50mM,
pH7). Experiments in Fig. 2a are representative results from 3 independent biological
replicates.
Antibody-shift assay and 2D BN-PAGE analysis were performed as described (Schwenk
et al., 2012; Schwenk et al., 2009) with minor modifications. Briefly, 50μg - 100μg of
digitonin or C12E9 solubilized complexes were pre-incubated for 1 hour with 10μg of Ms-
anti-KCC2(N1/22) / Ms IgG whole molecule; and 5μg Rb-anti-GluK2/3(NL9) / Rb IgG
whole molecule / Rb-anti-HA respectively, prior to the addition of Coomassie blue G250.
1D-BN-PAGE were performed as described above using home-made 4% and 5% bis-tris
gels as described (Swamy et al., 2006). After the completion of the gel run, excised BN-
PAGE lanes were equilibrated in Laemmli buffer containing SDS and DTT for 15
minutes at room temperature to denature the native proteins. After a brief rinse in SDS-
PAGE running buffer, the excised BN-PAGE lanes were placed on a 6% or 8% SDS-
PAGE gel for separation in the second dimension. After standard electroblotting of SDS-
PAGE-resolved samples on nitrocellulose membrane, the blot was cut into two molecular
weight ranges; the top blots were subjected to western blotting analysis with Rb anti-
KCC2, Rb anti-GluK2/3 and the bottom blots with Rb anti-Neto2. Antibody-shift
experiments using hippocampal membranes are representative results from 3 independent
biological replicates, experiments using cortical membranes are representative results
from 2 independent biological replicates.
Surface biotinylation assay for studying surface: internal KCC2 ratio: Biotinylation
studies were performed as previously described (Ng et al., 2009) with modifications.
Briefly, 300μm coronal slices from age-matched (postnatal day 30 – 40) male wild-type
and GluK1/2-null littermate mice (n=5) were cut using a Leica Vibratome in modified
ACSF (180 mM sucrose, 25 mM sodium bicarbonate, 25 mM glucose, 2.5 mM KCl, 1.25
mM sodium phosphate, 2 mM MgCl2, 1 mM CaCl2, 0.4 mM sodium ascorbate, and 3
mM sodium pyruvate, and saturated with 95% O2/5% CO2 (pH 7.4. osmolarity
~295mOsm). Twelve slices from each genotype were recovered in a 50:50 mix of
modified ACSF and normal ACSF (125 mM NaCl, 25 mM sodium bicarbonate, 25 mM
glucose, 2.5 mM KCl, 1.25 mM sodium phosphate, 1 mM MgCl2, 2 mM CaCl2 (pH 7.4.
osmolarity ~295mOsm and saturated with 95% O2/5% CO2) for 30min, followed by a
second recovery in normal ACSF for another 30 min at room temperature.
Slices were incubated in 5 ml of cold-ACSF containing 500 μg/ml EZ-Link Sulfo-NHS-
SS-Biotin (21328, Thermo Scientific) bubbled in 95% O2 5% CO2, with gentle agitation
for 2 hrs at 4◦C. The reaction was stopped by quenching excess biotin in cold-ACSF
containing 100mM Tris and slices were washed twice in cold-ACSF, once in cold-
11

KCC2 and KARs Co-Exist in a Complex Mahadevan et al
Supplemental Material
modified ACSF and snap-frozen immediately on dry ice. Entire cortex region was
dissected under dissecting microscope (Olympus), lysed immediately in modified RIPA
buffer [50 mM Tris·HCl, pH 7.4, 150 mM NaCl, 1 mM EDTA, 1% Nonidet P-40, 0.1%
SDS, 0.5% DOC, protease inhibitors and phosphatase inhibitor mixture (Roche)] and
incubated on ice for 30 min. After thoroughly homogenizing, the samples was
centrifuged, supernatant was collected, and quantified using the BioRad protein
quantification kit. 50 μg of total protein in a total volume of 300 μl (made up using
modified RIPA) was mixed with 200 μl of 50% slurry of Neutravidin beads (29201,
Thermo Scientific) and rotated for 2 hrs at 4◦C. The beads were harvested by
centrifugation and the supernatant corresponding to ~5 μg of unbound fraction was
aliquoted (to measure the internal, unbiotinylated fraction of surface proteins). The beads
were subsequently washed three times in modified RIPA buffer. After the last wash all
solution was thoroughly removed from beads, and the biotin-bound and unbound
fractions were denatured in 6XSDS sample buffer containing DTT at 37◦C for hr before
resolving them on onto 6% SDS-PAGE. Subsequent immunoblot analysis was performed
as described earlier. In a small subset of this experiment, the slices were lysed, and the
neutravidin pull-down were performed in C12E9 based buffer, and we observed similar
results to Fig. 4a (data not shown).
Hippocampal cultures and electrophysiology: Low-density cultures of dissociated
mouse hippocampal neurons were prepared as previously described (Acton et al., 2012),
in accordance to guidelines from the University of Toronto Animal Care Committee and
the Canadian Council on Animal Care. Experiments were performed after 10-13 days in
culture. Cultured neurons were selected for electrophysiology based on three criteria.
Using DIC microscopy we selected neurons: (1) with a healthy oval or pyramidal shaped
cell body; (2) multiple clearly identifiable processes; and (3) a cell body and proximal
dendrites that were relatively isolated (e.g. not in the direct vicinity of other neurons).
Gramicidin (50μg/ml) perforated patch clamp recordings were performed using glass
pipettes as previously described (Acton et al., 2012). Cultured neurons were continuously
perfused with standard extracellular solution (XSC). Recordings started when the series
resistance dropped below 50 MΩ. IV-curves were made by depolarizing the membrane
potential in steps, while simultaneously stimulating GABAergic transmission. A 10μM
GABA puff was applied to the soma. A linear regression of the IPSC/P amplitude was
used to calculate the voltage dependence of IPSC/Ps; the intercept of this line with the
abscissa was taken as EGABA, and the slope of this line was taken as the synaptic
conductance. The maximum current amplitude was taken as the largest absolute current
recorded during the recordings performed for the EGABA measurement.
Electrophysiological values have not been corrected for the liquid junction potential of ~7
mV.
GluK2shRNA and Neuronal transfection: Several (mouse) GluK2 shRNA were
generated in pGIPZ backbone with a GFP reporter. They were screened in HEK-293 to
identify the shRNA with maximum target silencing and least non-specific silencing by
co-expressing with exogenous GluK2 or other negative control plasmids (GluK1, KCC4
(Karadsheh et al., 2004)) at a target:shRNA ratio of 1: 8. GluK2 shRNA clone#3
12

KCC2 and KARs Co-Exist in a Complex Mahadevan et al
Supplemental Material
corresponding to mature antisense – AGAGATTGATCTTCTGTGT (Cat# RMM4431-
200414968, V3LMM_424318, Thermo Scientific), induced efficient target silencing (as
shown in supplemental figure 5b) when compared with scrambled shRNA (#RHS4346,
ThermoScientific). TransfectinTM Lipid Reagent (Biorad) was used to transfect GluK2
shRNA clone#3 or non-silencing shRNA into cultured hippocampal neurons, according
to the manufacturer’s instruction. Experiments in Fig. S5b are representative results from
3 - 4 independent biological replicates.
Fixed immunostaining and confocal microscopy: Fixed immunostaining was performed
by fixing DIV 12-14 cultured hippocampal neurons in 4% paraformaldehyde for 10 min
on ice followed by washing with 2X-PBS. Neurons were then permeabilized with PBS
containing 10% goat serum and 0.5% Triton X-100 for 30 minutes, followed by a 45 min
incubation with anti-GluK1/2 (Abcam) and mouse anti-KCC2 (N1/12) antibodies at 37◦C
to detect endogenous proteins. A small subset of experiments were also performed with
rabbit anti-GluK2/3 (Millipore) and mouse anti-KCC2 (N1/12) antibodies. Finally,
neurons were washed and incubated with Cy3-conjugated goat anti-rabbit and Alexa
Fluor 488-conjugated goat anti-mouse antibodies for 45 minutes at 37◦C. Neurons were
imaged on a Wave FX spinning disc confocal system (Quorum Technologies) with a
Zeiss Observer Z1 inverted microscope. Neurons were selected for imaging using the
same three criteria identified above under the subheading ‘Hippocampal Neurons and
Electrophysiology’. Images were acquired using 3D Image Analysis software (Perkin
Elmer). Images were obtained using a 43x 1.4-NA oil immersion objective.
Colocalization analysis was performed on representative z-stacks selected for analysis
using the ImageJ plugin JACoP (http://rsb.info.nih.gov/ij/plugins/track/jacop.html) (Bolte
and Cordelieres, 2006). Imaging experiments were performed and analyzed in a blinded
manner. Fig 1c is a representative of confocal images obtained from 26 neurons over four
independent experiments performed using 8 coverslips obtained from 4 independent sets
of cultures (of the total number of neurons analyzed (n = 26), 19 were prepared using
anti-GluK1/2, while 5 were prepared with anti-GluK2/3.
Live immunostaining and confocal microscopy: Live immunostaining was performed as
described before (Acton et al., 2012). Briefly, cultured neurons were washed in standard
extracellular solution (XCS), prior to incubating them with mouse monoclonal anti-HA
antibody at 37 °C for 15 minutes. Following incubation with the primary antibody, cells
were again washed in XCS prior to a 15 minutes dark incubation in secondary Alexa
Fluor 555 goat anti-mouse antibody. Images were acquired after 15 minutes of incubation
in the secondary antibody. Neurons were imaged on a Wave FX spinning disc confocal
system (Quorum Technologies) with a Zeiss Observer Z1 inverted microscope using a
60x 1.4NA oil immersion objective. Images were acquired and analyzed using Volocity
3D Image Analysis software (Perkin Elmer). Z-stacks were acquired at a pixel resolution
of 1024xy with a z-step of 0.2 µm (images in Fig. 4c are from singe z-planes). Samples
were acquired sequentially using line-lambda mode at excitation wavelengths of 488 nm
and 568 nm. Anti-HA (Cy3) immunofluorescence puncta intensity was measured under
constant illumination. Puncta were identified using the object identification tool in
Volocity software. We determined the percentage of puncta proximal to the membrane by
determining the location of the puncta relative to the surface of the cell. To do this we
13

KCC2 and KARs Co-Exist in a Complex Mahadevan et al
Supplemental Material
used the analysis program to identify the centroid of the cell (based on volume). We then
drew a vector from the centroid to edge of the GFP fluorescence through the puncta. If
the puncta were at the end of the vector they were classified as being proximal to the
surface; any puncta that were part-way along the vector were considered as distal to the
surface. To be considered at the end of the vector we only selected puncta where the
vector was only visible from the intracellular side of the puncta. Imaging experiments
were performed and analyzed in a blinded manner. An example image of this analysis is
found in Fig. S4. Live immunostaining analysis was performed on: wild-type n = 23,
GluK1/2KO n = 22. Each set of n values was obtained from 5 independent sets of
cultures.
Ca2+
imaging. Hippocampal neurons were loaded with the membrane-permeable
fluorescent Ca2+
indicator Fluo4-AM (Invitrogen; Carlsbad, CA, USA) for 30 min at
37°C, 5% CO2. The Fluo4 was dissolved in dimethyl sulfoxide (DMSO) and 20%
pluronic acid to a stock concentration of 1 mM and then diluted to 1 μM in our
extracellular recording solution. Following dye-loading the cells were thoroughly washed
with extracellular recording solution. Neurons were placed into the chamber of an
inverted microscope (Olympus IX71) equipped with an Olympus 0.6 NA × 40 objective.
Fluo4 was excited at 488 nm through a monochromator (Photon Technology
International (Canada) Inc., London, ON), controlled by the ImageMaster software
(Photon Technology International (Canada) Inc., London, ON). Fluorescence emission of
labeled cells at 510 nm was detected with a 16-bit CCD camera (Cascade 650,
Photometrics, Roper Scientific, Tuscon, AZ, USA). Images of 653 × 492 pixels were
accumulated at 500–1000 ms intervals. A neuron that change by 2000 fluorescent units
over a 5 second interval was considered to be active. Imaging experiments were
performed and analyzed in a blinded manner
Statistics: Results are given as mean ± SEM. For electrophysiology data sets and
immunostaining n values represent number of neurons; a minimum of three separate
coverslips were used for each of these data sets. For Western blotting experiments are
representative of a minimum of three separate biological replicates. Statistical
significance was determined using SigmaStat software. Statistical significance was
determined as follows: * p < 0.05, ** p < 0.01, *** p < 0.001. Statistical significance for
Figs 1b, 3a, S3a, 4a, S4a, 4b, 5b, 5c was determined using the Student’s t-test (two-
tailed), while Fig 3b used the paired t-test (two-tailed), 4bii, 4bvi, S5bii used the Mann-
Whitney rank sum test.
14

KCC2 and KARs Co-Exist in a Complex Mahadevan et al
Supplemental Material
SUPPLEMENTAL REFERENCES
Acton, B.A., Mahadevan, V., Mercado, A., Uvarov, P., Ding, Y., Pressey, J., Airaksinen,
M.S., Mount, D.B., and Woodin, M.A. (2012). Hyperpolarizing GABAergic transmission
requires the KCC2 C-terminal ISO domain. J Neurosci 32, 8746-8751.
Bolte, S., and Cordelieres, F.P. (2006). A guided tour into subcellular colocalization
analysis in light microscopy. J Microsc 224, 213-232.
Ivakine, E.A., Acton, B.A., Mahadevan, V., Ormond, J., Tang, M., Pressey, J.C., Huang,
M.Y., Ng, D., Delpire, E., Salter, M.W., et al. (2013). Neto2 is a KCC2 interacting
protein required for neuronal Cl- regulation in hippocampal neurons. Proceedings of the
National Academy of Sciences of the United States of America 110, 3561-3566.
Karadsheh, M.F., Byun, N., Mount, D.B., and Delpire, E. (2004). Localization of the
KCC4 potassium-chloride cotransporter in the nervous system. Neuroscience 123, 381-
391.
Ng, D., Pitcher, G.M., Szilard, R.K., Sertie, A., Kanisek, M., Clapcote, S.J., Lipina, T.,
Kalia, L.V., Joo, D., McKerlie, C., et al. (2009). Neto1 is a novel CUB-domain NMDA
receptor-interacting protein required for synaptic plasticity and learning. PLoS Biol 7,
e41.
Schwenk, J., Harmel, N., Brechet, A., Zolles, G., Berkefeld, H., Muller, C.S., Bildl, W.,
Baehrens, D., Huber, B., Kulik, A., et al. (2012). High-resolution proteomics unravel
architecture and molecular diversity of native AMPA receptor complexes. Neuron 74,
621-633.
Schwenk, J., Harmel, N., Zolles, G., Bildl, W., Kulik, A., Heimrich, B., Chisaka, O.,
Jonas, P., Schulte, U., Fakler, B., et al. (2009). Functional proteomics identify cornichon
proteins as auxiliary subunits of AMPA receptors. Science 323, 1313-1319.
Swamy, M., Siegers, G.M., Minguet, S., Wollscheid, B., and Schamel, W.W. (2006).
Blue native polyacrylamide gel electrophoresis (BN-PAGE) for the identification and
analysis of multiprotein complexes. Science's STKE : signal transduction knowledge
environment 2006, pl4.
Uvarov, P., Ludwig, A., Markkanen, M., Soni, S., Hubner, C.A., Rivera, C., and
Airaksinen, M.S. (2009). Coexpression and heteromerization of two neuronal K-Cl
cotransporter isoforms in neonatal brain. The Journal of biological chemistry 284, 13696-
13704.
15

KCC2 and KARs Co-Exist in a Complex Mahadevan et al
Supplemental Material
16
SUPPLEMENTAL TABLE 1: ANTIBODY INFORMATION
Antibody information Application and corresponding figures
Antibody Product
# Supplier WB IP IF
anti-KCC2
(rabbit)
07-432 Millipore (1:1000)
Figures: 1a, 3a, 3b,
4ai, S2ci, S3a
(2μg)
Figures: 1b (1:1000)
Figures: 3d
anti-KCC2
(mouse)
clone
N1/12
UC
Davis/NIMH/Neur
oMab Facility
(1:500)
Figures: 1b, S1c, 2a,
2bii, S5bii
(5-10 μg)
Figures: 1a,
S2c, S2d
(1:1000)
Figures: 1c, Data
not shown
(similar to 3d)
anti-Neto2
(rabbit)
n/a Tang et al., 2011 (1:1000)
Figures: 1a, 2biii, 4ai,
S2d, S4a
n/a n/a
anti-GluK2/3
(rabbit)
04-921 Millipore (1:1000)
Figures: 1a, 1b, 2a,
2bi, 3a, 4ai, S2cii,
S2b, S2d
(2-5μg)
Figures:
S1c, 2bi, ii,
iii
(1:1000)
Data not shown
(similar to 1c)
anti-GluK1
(rabbit)
07-258 Millipore n/a (2μg)
Figures:
S1c
n/a
anti-GluK1/2
(rabbit)
ab-
66440
abcam (1:1000)
Figures: S1b, S5b
n/a (1:1000)
Figures: 1c
anti-myc
(mouse)
R950-25 Invitrogen (1:1000)
Figures: 1b, S1c,
S5b
n/a n/a
anti-HA
(mouse)
12CA5 Roche n/a n/a (1:100)
Figures: 4b
anti-HA
(rabbit)
PRB-
101P
Covance n/a (5μg)
Figures: 2b
n/a
anti-NKCC1
(mouse)
CloneT4 Developmental
studies hybridoma
bank
(1:1000)
Figures: 3a, S3a
n/a n/a
anti-ATP1a3
(mouse)
NB300-
540
Novus Biologicals (1:2000)
Figures: 3a, 4ai
n/a n/a
anti-Tuj1
(mouse)
MMS-
435P
Covance (1:2000)
Figures: 3a, S3a,
S4a
n/a n/a
anti-
Transferrin
(mouse)
13-6800 Invitrogen n/a (5μg)
Figures:
S2b
n/a
anti-beta actin
(rabbit)
4967 Cell Signaling (1:2000)
Figures: 4ai, S5b
n/a n/a
anti-KCC4
(rabbit)
n/a Karadsheh et al.,
2004 Neuroscience (1:1000)
Figures: S5b
n/a n/a
WB, Western blot; IP, Immunoprecipitation; IF, Immunofluorescence

KCC2 and KARs Co-Exist in a Complex Mahadevan et al
Supplemental Material
17
SUPPLEMENTAL TABLE 2: Synaptic Current Amplitudes and Conductance
Genetic Deletion Acute Knockdown
WT GluK1/2 KO P Scrambled
shRNA
GluK2
shRNA P
GABAergic
Synaptic
Conductance
(pS)
2.53 ± 0.42 2.13 ± .34 0.5 1.65 ± .0.24 1.89 ± 0.34 0.6
Maximum
Current (pA) 65.07 ± 11.01 67.02 ± 19.0 0.6 76.77 ±10.51 104.9 ± 10.45 0.1
Copyright © 2022 FDOKUMEN




















