
Propofol Dose-Dependently Reduces Tumor Necrosis Factor??Induced Human Umbilical Vein Endothelial...
7
ANESTHETIC PHARMACOLOGY INTERNATIONAL SOCIETY FOR ANAESTHETIC PHARMACOLOGY SECTION EDITOR JAMES G. BOVILL Propofol Dose-Dependently Reduces Tumor Necrosis Factor--Induced Human Umbilical Vein Endothelial Cell Apoptosis: Effects on Bcl-2 and Bax Expression and Nitric Oxide Generation Tao Luo, MD*, Zhengyuan Xia, MD, PhD*‡, David M. Ansley, MD, FRCP‡, Jingping Ouyang, MD†, David J. Granville, PhD§, Yinping Li, PhD†, Zhong-Yuan Xia, MD*, Qing-Shan Zhou, MD, PhD*, and Xian-Yi Liu, MD* *Department of Anesthesiology, Renmin Hospital, Wuhan University; †Department of Pathophysiology, Faculty of Medicine, Wuhan University, Wuhan, People’s Republic of China; ‡Centre for Anesthesia & Analgesia, Department of Pharmacology & Therapeutics, The University of British Columbia, Vancouver; and §The James Hogg iCAPTURE Centre for Cardiovascular and Pulmonary Research, The University of British Columbia, Vancouver, British Columbia, Canada We investigated whether propofol can inhibit tumor necrosis factor (TNF)--induced apoptosis in cultured human umbilical vein endothelial cells (HUVECs). Iso- lated HUVECs were cultured in Dulbecco’s modified Eagle medium supplemented with 20% bovine calf se- rum. HUVECs in untreated and propofol control groups were cultured at 37°C for 24.5 h. HUVECs in the TNF treatment groups were initially cultured for 30 min in the presence of TNF or various concentrations of propofol, respectively, which were then cultured for 24 h with the addition of TNF at 40 ng/mL in the me- dium. Apoptosis was detected using terminal deoxy- nucleotidyl transferase-mediated dUTP nick-end label- ing (TUNEL) and confirmed by electron microscopy. The antiapoptotic Bcl-2 and proapoptotic Bax protein expressions were measured by immunocytochemical analysis. TNF stimulation resulted in a reduced Bcl-2/ Bax ratio and increased apoptotic index (AI: percentage of apoptotic cells) in HUVECs. Propofol, at concentra- tions 12 M, significantly (P 0.001) and dose- dependently attenuated TNF-induced increase in AI and decrease in Bcl-2/Bax ratio. This was accompanied by increases in nitric oxide production. There is an in- verse correlation between the ratio of Bcl-2/Bax expres- sion and AI (P 0.0009). These results suggest that propofol, at clinical relevant concentrations, can reduce TNF-induced HUVEC apoptosis. (Anesth Analg 2005;100:1653–9) T he vascular endothelial monolayer serves as a bar- rier between the bloodstream and the vascular wall. Apoptotic endothelial cell death may criti- cally disturb the integrity of the endothelial monolayer and thereby contribute to vascular injury and atheroscle- rosis. Apoptosis has become increasingly recognized as a mechanism of cell death during myocardial ischemia reperfusion injury (IRI), although the relative contribu- tion of necrosis and apoptosis to total cardiac cell loss during IRI remains controversial. Endothelial cell apo- ptosis has been shown to precede myocyte cell apoptosis in the setting of myocardial IRI (1). The latter study suggests that circulatory proapoptotic inflammatory cy- tokines (such as tumor necrosis factor [TNF]-) and re- active oxygen species (ROS), that are increased during myocardial IRI and atherosclerosis, promote myocyte apoptosis subsequent to the induction of endothelial cell apoptosis. Propofol, 2,6-diisopropylphenol, an IV anesthetic drug frequently used during cardiac surgery and in postoperative sedation (2), enhances red blood cell and tissue antioxidant capacity both in vitro and in Supported in part by a Scientific Achievement Award (982038-5, to ZX) from Hubei Province, China; in part by an internal research funding (to TL and ZX) from Renmin Hospital of Wuhan Univer- sity, China; and in part by a Jean Templeton Hugill funding (to DMA) from the Centre for Anesthesia & Analgesia, Department of Pharmacology & Therapeutics, The University of British Columbia, Canada. DJG is a Tier II Canada Research Chair and a Michael Smith Foundation for Health Research Scholar. ZX and DMA contributed equally to this study. Accepted for publication November 1, 2004. Address correspondence and reprint requests to Dr. Zhengyuan Xia, Centre for Anesthesia & Analgesia, Department of Pharmacology & Therapeutics, The University of British Columbia, Vancouver, BC, V6T 1Z3, Canada. Address e-mail to [email protected]. DOI: 10.1213/01.ANE.0000150945.95254.D8 ©2005 by the International Anesthesia Research Society 0003-2999/05 Anesth Analg 2005;100:1653–9 1653
-
Upload
independent -
Category
Documents
-
view
5 -
download
0
Transcript of Propofol Dose-Dependently Reduces Tumor Necrosis Factor??Induced Human Umbilical Vein Endothelial...

ANESTHETIC PHARMACOLOGY INTERNATIONAL SOCIETY FOR ANAESTHETIC PHARMACOLOGYSECTION EDITOR
JAMES G. BOVILL
Propofol Dose-Dependently Reduces Tumor NecrosisFactor-�-Induced Human Umbilical Vein Endothelial CellApoptosis: Effects on Bcl-2 and Bax Expression and NitricOxide GenerationTao Luo, MD*, Zhengyuan Xia, MD, PhD*‡, David M. Ansley, MD, FRCP‡,Jingping Ouyang, MD†, David J. Granville, PhD§, Yinping Li, PhD†, Zhong-Yuan Xia, MD*,Qing-Shan Zhou, MD, PhD*, and Xian-Yi Liu, MD*
*Department of Anesthesiology, Renmin Hospital, Wuhan University; †Department of Pathophysiology, Faculty ofMedicine, Wuhan University, Wuhan, People’s Republic of China; ‡Centre for Anesthesia & Analgesia, Department ofPharmacology & Therapeutics, The University of British Columbia, Vancouver; and §The James Hogg iCAPTURE Centrefor Cardiovascular and Pulmonary Research, The University of British Columbia, Vancouver, British Columbia, Canada
We investigated whether propofol can inhibit tumornecrosis factor (TNF)-�-induced apoptosis in culturedhuman umbilical vein endothelial cells (HUVECs). Iso-lated HUVECs were cultured in Dulbecco’s modifiedEagle medium supplemented with 20% bovine calf se-rum. HUVECs in untreated and propofol controlgroups were cultured at 37°C for 24.5 h. HUVECs in theTNF treatment groups were initially cultured for 30 minin the presence of TNF or various concentrations ofpropofol, respectively, which were then cultured for24 h with the addition of TNF at 40 ng/mL in the me-dium. Apoptosis was detected using terminal deoxy-nucleotidyl transferase-mediated dUTP nick-end label-ing (TUNEL) and confirmed by electron microscopy.
The antiapoptotic Bcl-2 and proapoptotic Bax proteinexpressions were measured by immunocytochemicalanalysis. TNF stimulation resulted in a reduced Bcl-2/Bax ratio and increased apoptotic index (AI: percentageof apoptotic cells) in HUVECs. Propofol, at concentra-tions �12 �M, significantly (P � 0.001) and dose-dependently attenuated TNF-induced increase in AIand decrease in Bcl-2/Bax ratio. This was accompaniedby increases in nitric oxide production. There is an in-verse correlation between the ratio of Bcl-2/Bax expres-sion and AI (P � 0.0009). These results suggest thatpropofol, at clinical relevant concentrations, can reduceTNF-induced HUVEC apoptosis.
(Anesth Analg 2005;100:1653–9)
T he vascular endothelial monolayer serves as a bar-rier between the bloodstream and the vascularwall. Apoptotic endothelial cell death may criti-
cally disturb the integrity of the endothelial monolayer
and thereby contribute to vascular injury and atheroscle-rosis. Apoptosis has become increasingly recognized as amechanism of cell death during myocardial ischemiareperfusion injury (IRI), although the relative contribu-tion of necrosis and apoptosis to total cardiac cell lossduring IRI remains controversial. Endothelial cell apo-ptosis has been shown to precede myocyte cell apoptosisin the setting of myocardial IRI (1). The latter studysuggests that circulatory proapoptotic inflammatory cy-tokines (such as tumor necrosis factor [TNF]-�) and re-active oxygen species (ROS), that are increased duringmyocardial IRI and atherosclerosis, promote myocyteapoptosis subsequent to the induction of endothelial cellapoptosis.
Propofol, 2,6-diisopropylphenol, an IV anestheticdrug frequently used during cardiac surgery and inpostoperative sedation (2), enhances red blood celland tissue antioxidant capacity both in vitro and in
Supported in part by a Scientific Achievement Award (982038-5,to ZX) from Hubei Province, China; in part by an internal researchfunding (to TL and ZX) from Renmin Hospital of Wuhan Univer-sity, China; and in part by a Jean Templeton Hugill funding (toDMA) from the Centre for Anesthesia & Analgesia, Department ofPharmacology & Therapeutics, The University of British Columbia,Canada. DJG is a Tier II Canada Research Chair and a Michael SmithFoundation for Health Research Scholar.
ZX and DMA contributed equally to this study.Accepted for publication November 1, 2004.Address correspondence and reprint requests to Dr. Zhengyuan Xia,
Centre for Anesthesia & Analgesia, Department of Pharmacology &Therapeutics, The University of British Columbia, Vancouver, BC, V6T1Z3, Canada. Address e-mail to [email protected].
DOI: 10.1213/01.ANE.0000150945.95254.D8
©2005 by the International Anesthesia Research Society0003-2999/05 Anesth Analg 2005;100:1653–9 1653

vivo (3,4). We (5) recently demonstrated that propo-fol enhances myocardial antioxidant capacity andresults in improved postischemic cardiac function inthe isolated rat heart, in a dose-dependent manner.In addition, several studies show that aging en-hances the sensitivity of human endothelial cellstoward apoptotic stimuli (6). Interestingly, propo-fol, when applied at a clinically achievable largeconcentration, enhances the ischemic tolerance ofmiddle-aged rat hearts (7). This finding promptedus to postulate that propofol may produce a cardio-protective effect that is attributable to its ability toenhance endothelial cell resistance toward apoptoticstimuli.
We hypothesized that propofol could inhibit TNF-�-induced human umbilical vein endothelial cells(HUVECs) apoptosis by resuming a proper ratio of theantiapoptotic Bcl-2 protein over the proapoptotic Baxprotein expression and that the propofol antiapoptoticeffect is related to its antioxidant capacity and itsability to enhance the generation of nitric oxide (NO),an important endothelial cell survival factor.
MethodsHUVECs were isolated according to the method ofJaffe et al. (8). Cells were cultured in Dulbecco’s mod-ified Eagle medium (Gibco) supplemented with 20%bovine calf serum (Gibco), maintained at 37°C in5% CO2, and used at passage 2–3 to avoid “age-dependent” variations in levels of apoptosis.
When the cells were at 70% confluence, the culturedHUVECs were divided into 7 groups: HUVECs inuntreated group (control) and propofol treatment con-trol (P25) group were further cultured at 37°C for 24 h,respectively, in the absence (control) or presence of 25�M propofol (AstraZeneca China, Beijing) in the me-dium. TNF-� was not applied in the two controlgroups throughout the experiment. HUVECs in theTNF-� (TNF) group and TNF-� plus propofol treat-ment groups were initially cultured for 30 min in thepresence of zero (TNF group), 12.5 (P12.5 � TNFgroup), 25 (P25 � TNF group), 50 (P50 � TNF group),and 100 (P100 � TNF group) �M propofol, respec-tively. The cells were then further cultured for 24 hwith the addition of TNF at 40 ng/mL into the me-dium. The concentration of TNF used to induce apo-ptosis in the present study was chosen on the basis ofpreviously published literature (9) in addition to pre-liminary studies. Experiments were repeated a mini-mum of eight times per group. Our preliminary studyresults indicated that intralipid, the solvent for propo-fol, at concentrations needed to dissolve up to 50 or100 �M propofol did not have a significant effect onHUVEC apoptotic cell death. Therefore, we did notinclude intralipid groups in the ensuing studies.
Apoptosis was detected using DNA in situ terminaldeoxynucleotidyl transferase (TdT)-mediated deoxyuri-dine triphosphate (dUTP)-biotin nick end-labeling(TUNEL) staining as per the manufacturer’s protocol(Boster Biotech, Wuhan, China). In brief, after equili-bration, end-labeling with digoxigenin-11-dUTP byTdT enzyme in buffer was performed for 1 h at 37°C ina humidifying chamber. After treatment with stop/wash buffer, sections were incubated with anti-digoxigenin antibody-peroxidase conjugate, rinsed,and stained with diaminobenzidine tetrahydrochloride.Negative controls were incubated with phosphate buff-ered saline (PBS) instead of TdT enzyme, and positivecontrols were treated with DNase1. Sections were coun-terstained with Mayer’s hematoxylin and mounted. Allexperiments were repeated on at least six independentoccasions with consistent results.
Cells were washed in Dulbecco’s modified Eaglemedium without bovine serum albumin, and cyto-spins were performed (650 rpm for 6 min) on saline-coated slides at 1 � 106 cells/mL. Slides were fixed in2% paraformaldehyde for 15 min at room temperatureand washed 5 times in PBS. Cells were permeabilizedfor 10 min at room temperature in blocking buffer (3%bovine serum albumin in PBS) plus 0.1% Triton X-100followed by blocking of nonspecific binding in block-ing buffer for 1 h at room temperature. They were thenincubated with the primary antibodies (anti-Bcl-21:50 dilution, and, anti-Bax 1:50 dilution) (Boster Bio-tech). After overnight incubation at 4°C, cells werewashed with PBS and incubated for 30 min at roomtemperature with the biotinylated secondary anti-body. After washing, the cells were incubated for30 min in Vectastain Elite ABC reagent (Boster Bio-tech). After another wash, cells were incubated inperoxidase substrate solution until the desired stainintensity was developed. The cells were then washed,dehydrated in increasing concentrations of ethanol,and cover-slipped using permount. Control sectionswere either incubated with the secondary antibodyalone or immunoabsorbed with an excess of blockingpeptide. Random fields (20–30 per slide) were exam-ined at a high magnification (�400) to calculate theprevalence of DNA fragmentation and Bcl-2/Bax ex-pression. The percentage of TUNEL-positive cells(termed apoptotic index, AI) was determined by di-viding the number of positive-staining nuclei by thetotal number of nuclei of the cell and multiplying thatvalue by 100. The percentage of Bcl-2/Bax expressionwas also determined. The density of Bcl-2 and Baxprotein expression was determined using an auto-matic computer-assisted image analyzing system(IBAS-2000; Kontron, Germany), which automaticallymeasures the density of 100 HUVECs from 4 to 6random fields. The density of Bcl-2 and Bax proteinexpression was expressed in arbitrary units.
1654 ANESTHETIC PHARMACOLOGY LUO ET AL. ANESTH ANALGPROPOFOL INHIBITS ENDOTHELIAL CELL APOPTOSIS 2005;100:1653–9
Electron microscopy was performed to confirm thatthe ultrastructural features of apoptosis were presentin cells exposed to TNF. Endothelial cells exposed tothe conditions outlined above were fixed in 2.5% glu-taraldehyde (pH 7.3) buffered with 0.1 mol/L sodiumcacodylate overnight at 4°C and then washed with 0.1mol/L sodium cacodylate buffer for 15 min beforepostfixation with 1% osmium tetroxide buffered with0.1 mol/L sodium cacodylate for 1 h on ice. Afteranother wash with 0.1 mol/L sodium cacodylatebuffer for 15 min, cells were dehydrated with increas-ing concentrations of alcohol. Next, cells were infil-trated with propylene oxide for 15 min, followed by1:1 propylene oxide/epoxy resin for 1 h, 1:2 propyleneoxide/epoxy resin for 2 h, and finally 100% epoxyresin for 2 h. Cells were embedded with fresh epoxyresin into molds and placed in a 60°C oven for 2 h.Ultrathin sections were stained with uranyl acetateand lead citrate and were examined with the use of aHitachi H-600 electron microscope (Hitachi, Japan).
Media of the cultured endothelial cells were col-lected 24 h after their respective treatments. The con-centration of nitrites (NO2
�) and nitrates (NO3�), sta-
ble end products of NO, was determined by the Griessreaction, as follows. After deproteination by a solutionof zinc sulfate, samples were incubated with cadmiumgranules to reduce nitrate to nitrite. The total nitritewas measured at 540-nm absorbance by diazotizationwith Griess reagent (Boshide Biotech Ltd., Wuhan).Nitrite concentrations were calculated by comparisonwith a standard calibration curve with sodium nitrite.
Results were expressed as mean � sd. Data weretested for normal distribution by the Kolmogorov-Smirnov test. Significance was evaluated using analy-sis of variance followed by Tukey’s post hoc test. Thecorrelation relationships were evaluated by the Pear-son test. P � 0.05 was considered significant.
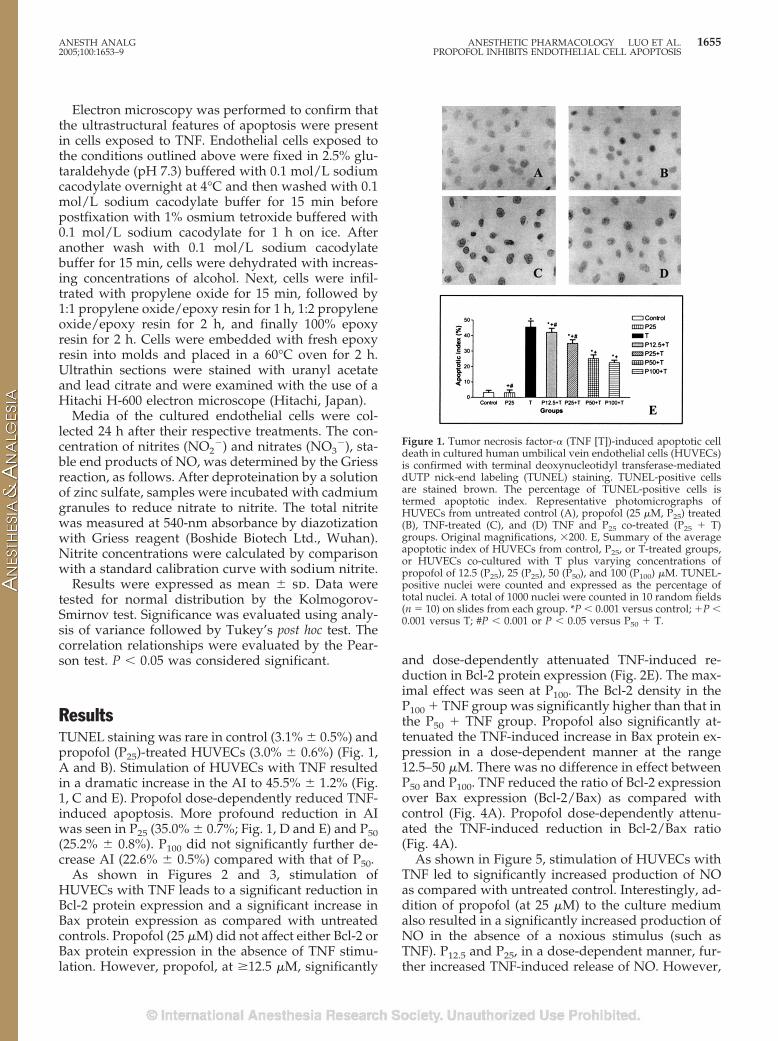
ResultsTUNEL staining was rare in control (3.1% � 0.5%) andpropofol (P25)-treated HUVECs (3.0% � 0.6%) (Fig. 1,A and B). Stimulation of HUVECs with TNF resultedin a dramatic increase in the AI to 45.5% � 1.2% (Fig.1, C and E). Propofol dose-dependently reduced TNF-induced apoptosis. More profound reduction in AIwas seen in P25 (35.0% � 0.7%; Fig. 1, D and E) and P50(25.2% � 0.8%). P100 did not significantly further de-crease AI (22.6% � 0.5%) compared with that of P50.
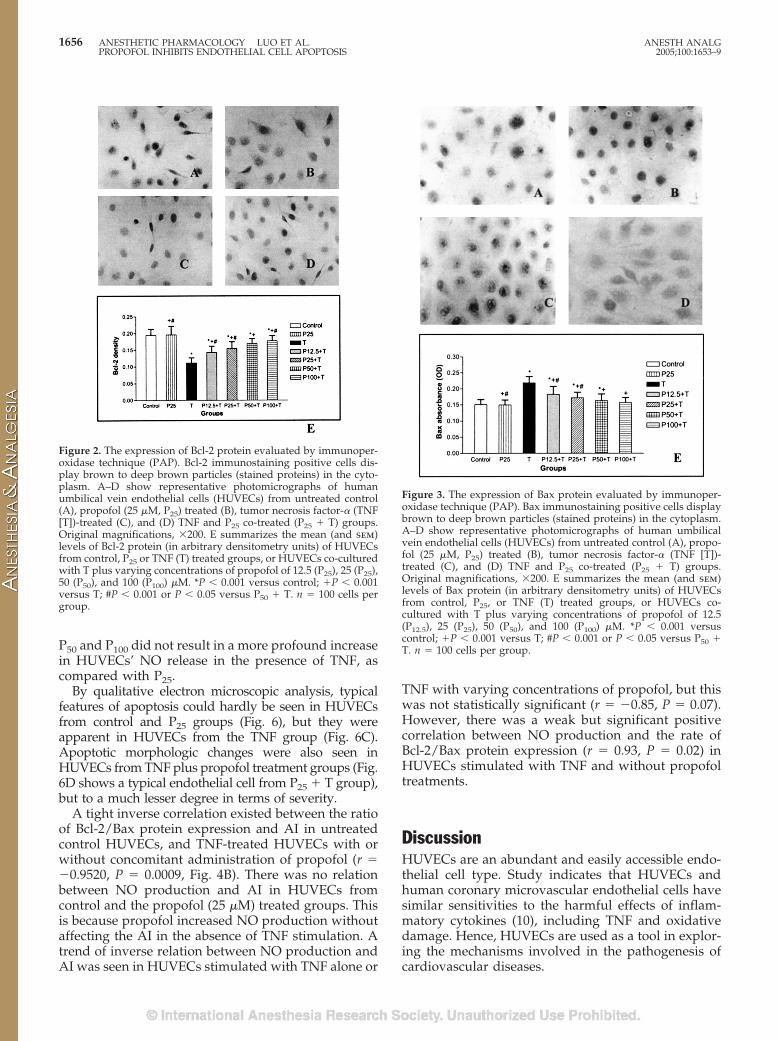
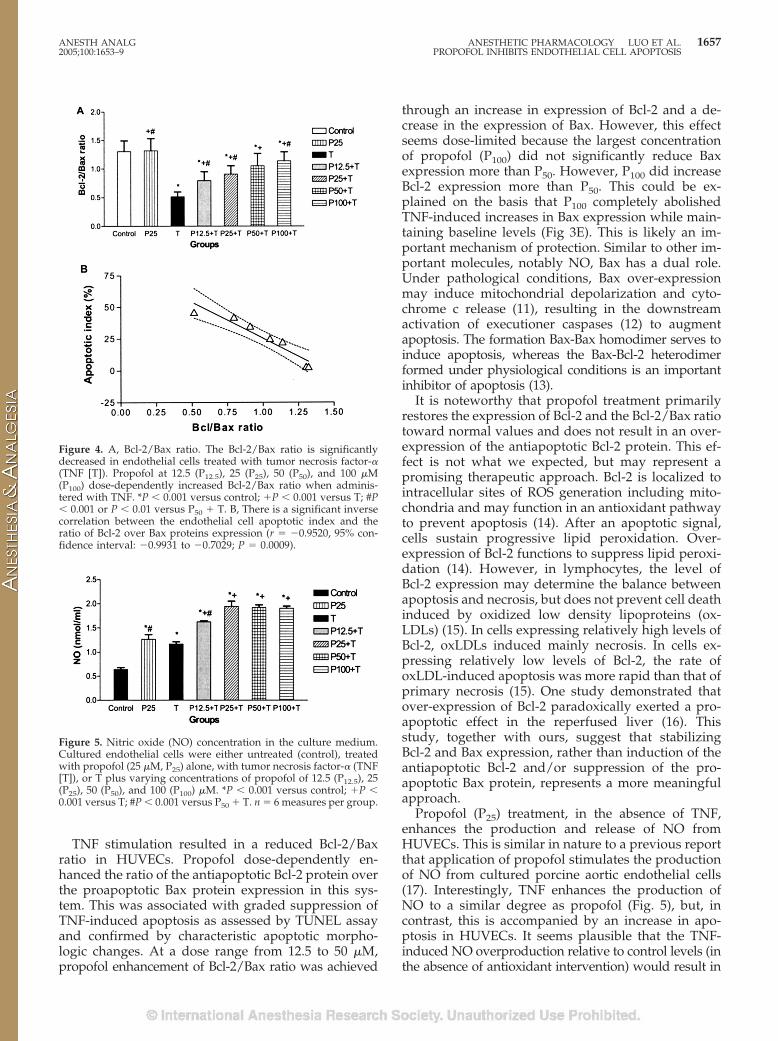
As shown in Figures 2 and 3, stimulation ofHUVECs with TNF leads to a significant reduction inBcl-2 protein expression and a significant increase inBax protein expression as compared with untreatedcontrols. Propofol (25 �M) did not affect either Bcl-2 orBax protein expression in the absence of TNF stimu-lation. However, propofol, at �12.5 �M, significantly
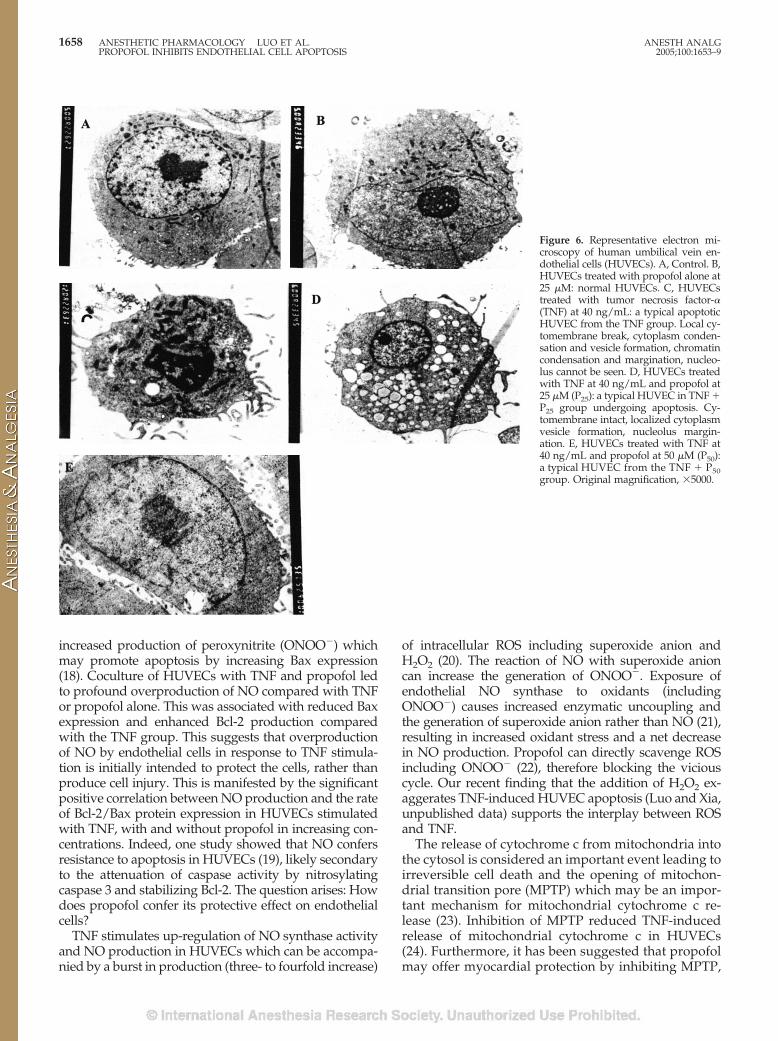
and dose-dependently attenuated TNF-induced re-duction in Bcl-2 protein expression (Fig. 2E). The max-imal effect was seen at P100. The Bcl-2 density in theP100 � TNF group was significantly higher than that inthe P50 � TNF group. Propofol also significantly at-tenuated the TNF-induced increase in Bax protein ex-pression in a dose-dependent manner at the range12.5–50 �M. There was no difference in effect betweenP50 and P100. TNF reduced the ratio of Bcl-2 expressionover Bax expression (Bcl-2/Bax) as compared withcontrol (Fig. 4A). Propofol dose-dependently attenu-ated the TNF-induced reduction in Bcl-2/Bax ratio(Fig. 4A).
As shown in Figure 5, stimulation of HUVECs withTNF led to significantly increased production of NOas compared with untreated control. Interestingly, ad-dition of propofol (at 25 �M) to the culture mediumalso resulted in a significantly increased production ofNO in the absence of a noxious stimulus (such asTNF). P12.5 and P25, in a dose-dependent manner, fur-ther increased TNF-induced release of NO. However,
Figure 1. Tumor necrosis factor-� (TNF [T])-induced apoptotic celldeath in cultured human umbilical vein endothelial cells (HUVECs)is confirmed with terminal deoxynucleotidyl transferase-mediateddUTP nick-end labeling (TUNEL) staining. TUNEL-positive cellsare stained brown. The percentage of TUNEL-positive cells istermed apoptotic index. Representative photomicrographs ofHUVECs from untreated control (A), propofol (25 �M, P25) treated(B), TNF-treated (C), and (D) TNF and P25 co-treated (P25 � T)groups. Original magnifications, �200. E, Summary of the averageapoptotic index of HUVECs from control, P25, or T-treated groups,or HUVECs co-cultured with T plus varying concentrations ofpropofol of 12.5 (P25), 25 (P25), 50 (P50), and 100 (P100) �M. TUNEL-positive nuclei were counted and expressed as the percentage oftotal nuclei. A total of 1000 nuclei were counted in 10 random fields(n � 10) on slides from each group. *P � 0.001 versus control; �P �0.001 versus T; #P � 0.001 or P � 0.05 versus P50 � T.
ANESTH ANALG ANESTHETIC PHARMACOLOGY LUO ET AL. 16552005;100:1653–9 PROPOFOL INHIBITS ENDOTHELIAL CELL APOPTOSIS
P50 and P100 did not result in a more profound increasein HUVECs’ NO release in the presence of TNF, ascompared with P25.
By qualitative electron microscopic analysis, typicalfeatures of apoptosis could hardly be seen in HUVECsfrom control and P25 groups (Fig. 6), but they wereapparent in HUVECs from the TNF group (Fig. 6C).Apoptotic morphologic changes were also seen inHUVECs from TNF plus propofol treatment groups (Fig.6D shows a typical endothelial cell from P25 � T group),but to a much lesser degree in terms of severity.
A tight inverse correlation existed between the ratioof Bcl-2/Bax protein expression and AI in untreatedcontrol HUVECs, and TNF-treated HUVECs with orwithout concomitant administration of propofol (r ��0.9520, P � 0.0009, Fig. 4B). There was no relationbetween NO production and AI in HUVECs fromcontrol and the propofol (25 �M) treated groups. Thisis because propofol increased NO production withoutaffecting the AI in the absence of TNF stimulation. Atrend of inverse relation between NO production andAI was seen in HUVECs stimulated with TNF alone or
TNF with varying concentrations of propofol, but thiswas not statistically significant (r � �0.85, P � 0.07).However, there was a weak but significant positivecorrelation between NO production and the rate ofBcl-2/Bax protein expression (r � 0.93, P � 0.02) inHUVECs stimulated with TNF and without propofoltreatments.
DiscussionHUVECs are an abundant and easily accessible endo-thelial cell type. Study indicates that HUVECs andhuman coronary microvascular endothelial cells havesimilar sensitivities to the harmful effects of inflam-matory cytokines (10), including TNF and oxidativedamage. Hence, HUVECs are used as a tool in explor-ing the mechanisms involved in the pathogenesis ofcardiovascular diseases.
Figure 2. The expression of Bcl-2 protein evaluated by immunoper-oxidase technique (PAP). Bcl-2 immunostaining positive cells dis-play brown to deep brown particles (stained proteins) in the cyto-plasm. A–D show representative photomicrographs of humanumbilical vein endothelial cells (HUVECs) from untreated control(A), propofol (25 �M, P25) treated (B), tumor necrosis factor-� (TNF[T])-treated (C), and (D) TNF and P25 co-treated (P25 � T) groups.Original magnifications, �200. E summarizes the mean (and sem)levels of Bcl-2 protein (in arbitrary densitometry units) of HUVECsfrom control, P25 or TNF (T) treated groups, or HUVECs co-culturedwith T plus varying concentrations of propofol of 12.5 (P25), 25 (P25),50 (P50), and 100 (P100) �M. *P � 0.001 versus control; �P � 0.001versus T; #P � 0.001 or P � 0.05 versus P50 � T. n � 100 cells pergroup.
Figure 3. The expression of Bax protein evaluated by immunoper-oxidase technique (PAP). Bax immunostaining positive cells displaybrown to deep brown particles (stained proteins) in the cytoplasm.A–D show representative photomicrographs of human umbilicalvein endothelial cells (HUVECs) from untreated control (A), propo-fol (25 �M, P25) treated (B), tumor necrosis factor-� (TNF [T])-treated (C), and (D) TNF and P25 co-treated (P25 � T) groups.Original magnifications, �200. E summarizes the mean (and sem)levels of Bax protein (in arbitrary densitometry units) of HUVECsfrom control, P25, or TNF (T) treated groups, or HUVECs co-cultured with T plus varying concentrations of propofol of 12.5(P12.5), 25 (P25), 50 (P50), and 100 (P100) �M. *P � 0.001 versuscontrol; �P � 0.001 versus T; #P � 0.001 or P � 0.05 versus P50 �T. n � 100 cells per group.
1656 ANESTHETIC PHARMACOLOGY LUO ET AL. ANESTH ANALGPROPOFOL INHIBITS ENDOTHELIAL CELL APOPTOSIS 2005;100:1653–9
TNF stimulation resulted in a reduced Bcl-2/Baxratio in HUVECs. Propofol dose-dependently en-hanced the ratio of the antiapoptotic Bcl-2 protein overthe proapoptotic Bax protein expression in this sys-tem. This was associated with graded suppression ofTNF-induced apoptosis as assessed by TUNEL assayand confirmed by characteristic apoptotic morpho-logic changes. At a dose range from 12.5 to 50 �M,propofol enhancement of Bcl-2/Bax ratio was achieved
through an increase in expression of Bcl-2 and a de-crease in the expression of Bax. However, this effectseems dose-limited because the largest concentrationof propofol (P100) did not significantly reduce Baxexpression more than P50. However, P100 did increaseBcl-2 expression more than P50. This could be ex-plained on the basis that P100 completely abolishedTNF-induced increases in Bax expression while main-taining baseline levels (Fig 3E). This is likely an im-portant mechanism of protection. Similar to other im-portant molecules, notably NO, Bax has a dual role.Under pathological conditions, Bax over-expressionmay induce mitochondrial depolarization and cyto-chrome c release (11), resulting in the downstreamactivation of executioner caspases (12) to augmentapoptosis. The formation Bax-Bax homodimer serves toinduce apoptosis, whereas the Bax-Bcl-2 heterodimerformed under physiological conditions is an importantinhibitor of apoptosis (13).
It is noteworthy that propofol treatment primarilyrestores the expression of Bcl-2 and the Bcl-2/Bax ratiotoward normal values and does not result in an over-expression of the antiapoptotic Bcl-2 protein. This ef-fect is not what we expected, but may represent apromising therapeutic approach. Bcl-2 is localized tointracellular sites of ROS generation including mito-chondria and may function in an antioxidant pathwayto prevent apoptosis (14). After an apoptotic signal,cells sustain progressive lipid peroxidation. Over-expression of Bcl-2 functions to suppress lipid peroxi-dation (14). However, in lymphocytes, the level ofBcl-2 expression may determine the balance betweenapoptosis and necrosis, but does not prevent cell deathinduced by oxidized low density lipoproteins (ox-LDLs) (15). In cells expressing relatively high levels ofBcl-2, oxLDLs induced mainly necrosis. In cells ex-pressing relatively low levels of Bcl-2, the rate ofoxLDL-induced apoptosis was more rapid than that ofprimary necrosis (15). One study demonstrated thatover-expression of Bcl-2 paradoxically exerted a pro-apoptotic effect in the reperfused liver (16). Thisstudy, together with ours, suggest that stabilizingBcl-2 and Bax expression, rather than induction of theantiapoptotic Bcl-2 and/or suppression of the pro-apoptotic Bax protein, represents a more meaningfulapproach.
Propofol (P25) treatment, in the absence of TNF,enhances the production and release of NO fromHUVECs. This is similar in nature to a previous reportthat application of propofol stimulates the productionof NO from cultured porcine aortic endothelial cells(17). Interestingly, TNF enhances the production ofNO to a similar degree as propofol (Fig. 5), but, incontrast, this is accompanied by an increase in apo-ptosis in HUVECs. It seems plausible that the TNF-induced NO overproduction relative to control levels (inthe absence of antioxidant intervention) would result in
Figure 4. A, Bcl-2/Bax ratio. The Bcl-2/Bax ratio is significantlydecreased in endothelial cells treated with tumor necrosis factor-�(TNF [T]). Propofol at 12.5 (P12.5), 25 (P25), 50 (P50), and 100 �M(P100) dose-dependently increased Bcl-2/Bax ratio when adminis-tered with TNF. *P � 0.001 versus control; �P � 0.001 versus T; #P� 0.001 or P � 0.01 versus P50 � T. B, There is a significant inversecorrelation between the endothelial cell apoptotic index and theratio of Bcl-2 over Bax proteins expression (r � �0.9520, 95% con-fidence interval: �0.9931 to �0.7029; P � 0.0009).
Figure 5. Nitric oxide (NO) concentration in the culture medium.Cultured endothelial cells were either untreated (control), treatedwith propofol (25 �M, P25) alone, with tumor necrosis factor-� (TNF[T]), or T plus varying concentrations of propofol of 12.5 (P12.5), 25(P25), 50 (P50), and 100 (P100) �M. *P � 0.001 versus control; �P �0.001 versus T; #P � 0.001 versus P50 � T. n � 6 measures per group.
ANESTH ANALG ANESTHETIC PHARMACOLOGY LUO ET AL. 16572005;100:1653–9 PROPOFOL INHIBITS ENDOTHELIAL CELL APOPTOSIS
increased production of peroxynitrite (ONOO�) whichmay promote apoptosis by increasing Bax expression(18). Coculture of HUVECs with TNF and propofol ledto profound overproduction of NO compared with TNFor propofol alone. This was associated with reduced Baxexpression and enhanced Bcl-2 production comparedwith the TNF group. This suggests that overproductionof NO by endothelial cells in response to TNF stimula-tion is initially intended to protect the cells, rather thanproduce cell injury. This is manifested by the significantpositive correlation between NO production and the rateof Bcl-2/Bax protein expression in HUVECs stimulatedwith TNF, with and without propofol in increasing con-centrations. Indeed, one study showed that NO confersresistance to apoptosis in HUVECs (19), likely secondaryto the attenuation of caspase activity by nitrosylatingcaspase 3 and stabilizing Bcl-2. The question arises: Howdoes propofol confer its protective effect on endothelialcells?
TNF stimulates up-regulation of NO synthase activityand NO production in HUVECs which can be accompa-nied by a burst in production (three- to fourfold increase)
of intracellular ROS including superoxide anion andH2O2 (20). The reaction of NO with superoxide anioncan increase the generation of ONOO�. Exposure ofendothelial NO synthase to oxidants (includingONOO�) causes increased enzymatic uncoupling andthe generation of superoxide anion rather than NO (21),resulting in increased oxidant stress and a net decreasein NO production. Propofol can directly scavenge ROSincluding ONOO� (22), therefore blocking the viciouscycle. Our recent finding that the addition of H2O2 ex-aggerates TNF-induced HUVEC apoptosis (Luo and Xia,unpublished data) supports the interplay between ROSand TNF.
The release of cytochrome c from mitochondria intothe cytosol is considered an important event leading toirreversible cell death and the opening of mitochon-drial transition pore (MPTP) which may be an impor-tant mechanism for mitochondrial cytochrome c re-lease (23). Inhibition of MPTP reduced TNF-inducedrelease of mitochondrial cytochrome c in HUVECs(24). Furthermore, it has been suggested that propofolmay offer myocardial protection by inhibiting MPTP,
Figure 6. Representative electron mi-croscopy of human umbilical vein en-dothelial cells (HUVECs). A, Control. B,HUVECs treated with propofol alone at25 �M: normal HUVECs. C, HUVECstreated with tumor necrosis factor-�(TNF) at 40 ng/mL: a typical apoptoticHUVEC from the TNF group. Local cy-tomembrane break, cytoplasm conden-sation and vesicle formation, chromatincondensation and margination, nucleo-lus cannot be seen. D, HUVECs treatedwith TNF at 40 ng/mL and propofol at25 �M (P25): a typical HUVEC in TNF �P25 group undergoing apoptosis. Cy-tomembrane intact, localized cytoplasmvesicle formation, nucleolus margin-ation. E, HUVECs treated with TNF at40 ng/mL and propofol at 50 �M (P50):a typical HUVEC from the TNF � P50group. Original magnification, �5000.
1658 ANESTHETIC PHARMACOLOGY LUO ET AL. ANESTH ANALGPROPOFOL INHIBITS ENDOTHELIAL CELL APOPTOSIS 2005;100:1653–9

at concentrations as small as 11–22 �M (2–4 �g/mL),likely acting indirectly through its antioxidant prop-erties because ROS is a major mediator of MPTPopening (25). Propofol, however, may directly inhibitmyocyte MPTP at concentrations �50 �M (26). Interest-ingly, the most profound inhibition of TNF-inducedHUVEC apoptosis was manifested at propofol concen-trations �50 �M (P50 and P100) in this study, suggestinginhibition of MPTP in HUVECs as one of the mecha-nisms of propofol’s protection.
Our study provides evidence for the first time thatpropofol, a frequently used anesthetic for cardiac sur-gery and for postoperative sedation, attenuates TNF-induced HUVEC apoptosis in a dose-dependent man-ner. Clinically relevant blood concentrations of propofolinclude 0.8–1.0 �g/mL on awakening from propofolanesthesia, 1–2 �g/mL on long-term sedation in theintensive care unit, at least 2.5 �g/mL for satisfactoryhypnosis, and 3–11 �g/mL (approximately 17–62 �M)for maintenance of satisfactory anesthesia (27). Becausethe release of inflammatory cytokines (including TNF) isincreased during and after cardiac surgery and becauseserum from patients with acute coronary syndromesdisplays a proapoptotic effect on endothelial cells (28),application of propofol, in large concentrations (approx-imately 50–60 �M) (3), may prove to be a promisingapproach in reducing peri- and postoperative IRI andrelated cardiovascular symptoms.
References1. Scarabelli T, Stephanou A, Rayment N, et al. Apoptosis of
endothelial cells precedes myocyte cell apoptosis in ischemia/reperfusion injury. Circulation 2001;104:253–6.
2. Bryson HM, Fulton BR, Faulds D. Propofol: an update of its usein anaesthesia and conscious sedation. Drugs 1995;50:513–59.
3. Ansley DM, Sun J, Visser WA, et al. High dose propofol en-hances red cell antioxidant capacity during CPB in humans. CanJ Anaesth 1999;46:641–8.
4. Runzer TD, Ansley DM, Godin DV, et al. Tissue antioxidantcapacity during anesthesia: propofol enhances in vivo red celland tissue antioxidant capacity in a rat model. Anesth Analg2002;94:89–93.
5. Xia Z, Godin DV, Chang TK, et al. Dose-dependent protection ofcardiac function by propofol during ischemia and early reper-fusion in rats: effects on 15-F2t-isoprostane formation. CanJ Physiol Pharmacol 2003;81:14–21.
6. Hoffmann J, Haendeler J, Aicher A, et al. Aging enhances thesensitivity of endothelial cells toward apoptotic stimuli: impor-tant role of nitric oxide. Circ Res 2001;89:709–15.
7. Xia Z, Godin DV, Ansley DM. Propofol enhances ischemictolerance of middle-aged rat hearts: effects on 15-F(2t)-isoprostane formation and tissue antioxidant capacity. Cardio-vasc Res 2003;59:113–21.
8. Jaffe EA, Nachman RL, Becker CG, et al. Synthesis of antihemo-philic factor antigen by cultured human endothelial cells. J ClinInvest 1973;52:2745–56.
9. Polunovsky VA, Wendt CH, Ingbar DH, et al. Induction ofendothelial cell apoptosis by TNF�: modulation by inhibitors ofprotein synthesis. Exp Cell Res 1994;214:584–94.
10. Willam C, Koehne P, Jurgensen JS, et al. Tie2 receptor expres-sion is stimulated by hypoxia and proinflammatory cytokines inhuman endothelial cells. Circ Res 2000;87:370–7.
11. Jurgensmeier JM, Xie Z, Deveraux Q, et al. Bax directly inducesrelease of cytochrome c from isolated mitochondria. Proc NatlAcad Sci USA 1998;95:4997–5002.
12. Granville DJ, Shaw JR, Leong S, et al. Release of cytochrome c,Bax migration, Bid cleavage, and activation of caspases 2, 3, 6, 7,8, and 9 during endothelial cell apoptosis. Am J Pathol 1999;155:1021–5.
13. Krajewski S, Krajewska M, Shabaik A, et al. Immunohistochem-ical determination of in vivo distribution of Bax, a dominantinhibitor of Bcl-2. Am J Pathol 1994;145:1323–36.
14. Hockenbery DM, Oltvai ZN, Yin XM, et al. Bcl-2 functions in anantioxidant pathway to prevent apoptosis. Cell 1993;75:241–51.
15. Meilhac O, Escargueil-Blanc I, Thiers JC, et al. Bcl-2 alters thebalance between apoptosis and necrosis, but does not preventcell death induced by oxidized low density lipoproteins. FASEBJ 1999;13:485–94.
16. Oshiro T, Shiraishi M, Muto Y. Adenovirus mediated genetransfer of antiapoptotic protein in hepatic ischemia-reperfusioninjury: the paradoxical effect of Bcl-2 expression in the reper-fused liver. J Surg Res 2002;103:30–6.
17. Petros AJ, Bogle RG, Pearson JD. Propofol stimulates nitricoxide release from cultured porcine aortic endothelial cells. Br JPharmacol 1993;109:6–7.
18. Arstall MA, Sawyer DB, Fukazawa R, et al. Cytokine-mediatedapoptosis in cardiac myocytes: the role of inducible nitric oxidesynthase induction and peroxynitrite generation. Circ Res 1999;85:829–40.
19. Ho FM, Liu SH, Liau CS, et al. Nitric oxide prevents apoptosisof human endothelial cells from high glucose exposure duringearly stage. Cell Biochem 1999;75:258–63.
20. Deshpande SS, Angkeow P, Huang J, et al. Rac1 inhibits TNF-alpha-induced endothelial cell apoptosis: dual regulation byreactive oxygen species. FASEB J 2000;14:1705–14.
21. Zou MH, Shi C, Cohen RA. Oxidation of the zinc-thiolate com-plex and uncoupling of endothelial nitric oxide synthase byperoxynitrite. J Clin Invest 2002;109:817–26.
22. Kahraman S, Demiryurek AT. Propofol is a peroxynitrite scav-enger. Anesth Analg 1997;84:1127–9.
23. Green DR, Reed JC. Mitochondria and apoptosis. Science 1998;281:1309–12.
24. Walter DH, Haendeler J, Galle J, et al. Cyclosporin A inhibitsapoptosis of human endothelial cells by preventing release ofcytochrome C from mitochondria. Circulation 1998;98:1153–7.
25. Sztark F, Ichas F, Ouhabi R, et al. Effects of the anaestheticpropofol on the calcium-induced permeability transition of ratheart mitochondria: direct pore inhibition and shift of the gatingpotential. FEBS Lett 1995;368:101–4.
26. Kowaltowski AJ, Castilho RF, Vercesi AE. Mitochondrial per-meability transition and oxidative stress. FEBS Lett 2001;495:12–5.
27. Short TG, Aun CS, Tan P, et al. A prospective evaluation ofpharmacokinetic model controlled infusion of propofol in pae-diatric patients. Br J Anaesth 1994;72:302–6.
28. Valgimigli M, Agnoletti L, Curello S, et al. Serum from patientswith acute coronary syndromes displays a proapoptotic effecton human endothelial cells: a possible link to pan-coronarysyndromes. Circulation 2003;107:264–70.
ANESTH ANALG ANESTHETIC PHARMACOLOGY LUO ET AL. 16592005;100:1653–9 PROPOFOL INHIBITS ENDOTHELIAL CELL APOPTOSIS


![A-740003 [N-(1-{[(Cyanoimino)(5-quinolinylamino) methyl]amino}-2,2-dimethylpropyl)-2-(3,4-dimethoxyphenyl)acetamide], a Novel and Selective P2X7 Receptor Antagonist, Dose-Dependently](https://static.fdokumen.com/doc/165x107/63441f69596bdb97a9085093/a-740003-n-1-cyanoimino5-quinolinylamino-methylamino-22-dimethylpropyl-2-34-dimethoxyphenylacetamide.jpg)

















