
BCL2-modifying factor promotes germ cell loss during murine ...
Upload
annatrichypanruticampusCategory
view
0download
0
The Combined Functions of Proapoptotic Bcl-2 Family MembersBak and Bax Are Essential for Normal Development of MultipleTissues
Tullia Lindsten1, Andrea J. Ross2, Ayala King1, Wei-Xing Zong1, Jeffrey C. Rathmell1,Helena A. Shiels3, Eugen Ulrich4, Katrina G. Waymire5, Patryce Mahar5, KennethFrauwirth1, Yifeng Chen1, Michael Wei8, Vicki M. Eng6, David M. Adelman7, M. CelesteSimon7, Averil Ma3, Jeffrey A. Golden1, Gerard Evan4, Stanley J. Korsmeyer8, Grant R.MacGregor5, and Craig B. Thompson1,91 Departments of Medicine and Pathology and Laboratory Medicine Abramson Family CancerResearch Institute University of Pennsylvania Philadelphia, Pennsylvania 191042 Graduate Program in Biochemistry, Cell, and Developmental Biology Emory University Schoolof Medicine Atlanta, Georgia 303223 Department of Medicine University of Chicago Chicago, Illinois 606374 Cancer Research Institute UCSF Cancer Center San Francisco, California 941435 Center for Molecular Medicine Emory University School of Medicine Atlanta, Georgia 103226 University Laboratory Animal Resources University of Pennsylvania Philadelphia, Pennsylvania191047 Department of Cell and Developmental Biology Abramson Family Cancer Research Instituteand Howard Hughes Medical Institute University of Pennsylvania Philadelphia, Pennsylvania191048 Departments of Pathology and Medicine Harvard Medical School Dana Farber Cancer InstituteHoward Hughes Medical Institute Boston, Massachusetts 02115
SummaryProapoptotic Bcl-2 family members have been proposed to play a central role in regulatingapoptosis. However, mice lacking bax display limited phenotypic abnormalities. As presentedhere, bak–/– mice were found to be developmentally normal and reproductively fit and failed todevelop any age-related disorders. However, when Bak-deficient mice were mated to Bax-deficient mice to create mice lacking both genes, the majority of bax–/–bak–/– animals diedperinatally with fewer than 10% surviving into adulthood. bax–/–bak–/– mice displayed multipledevelopmental defects, including persistence of interdigital webs, an imperforate vaginal canal,and accumulation of excess cells within both the central nervous and hematopoietic systems. Thus,Bax and Bak have overlapping roles in the regulation of apoptosis during mammaliandevelopment and tissue homeostasis.
Copyright ©2000 by Cell Press9 To whom correspondence should be addressed ([email protected])..
NIH Public AccessAuthor ManuscriptMol Cell. Author manuscript; available in PMC 2011 March 15.
Published in final edited form as:Mol Cell. 2000 December ; 6(6): 1389–1399.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

IntroductionProgrammed cell death (apoptosis) plays a critical role in the regulation of development andtissue homeostasis. In addition, apoptosis plays an important role in the elimination ofdamaged or infected cells (Vaux and Korsmeyer, 1999). Two major apoptotic pathwayshave been identified (Strasser et al., 1995; Green, 2000). The extrinsic cell death pathwayprovides multicellular organisms a means to eliminate cells through instruction. In theextrinsic pathway, apoptosis is initiated through ligand binding to cell surface receptorsexpressed on the cell that will subsequently die. In contrast, a cell-intrinsic death pathwayhas been defined in which apoptosis is initiated within the cell.
The best characterized of the receptors involved in the extrinsic death pathway are membersof the TNF receptor family, such as Fas and TNF-R1 (Nagata, 1997). Once engaged byligand, these receptors initiate the formation of a death-inducing signaling complex, oneessential component of which is the adaptor molecule FADD. FADD in turn recruitsCaspase 8. Upon recruitment to the signaling complex, Caspase 8 undergoes autoproteolyticactivation. Once activated, Caspase 8 can initiate both the activation of additional caspasesand the degradation of intracellular substrates. These events result in the orderly degradationof intracellular contents and cell death.
The cell-intrinsic apoptotic pathway is initiated when the outer mitochondrial membraneloses its integrity, resulting in the release of cytochrome c and other apoptotic regulatoryproteins into the cytosol (Green, 2000). In the cytosol, released cytochrome c binds toApaf-1. Apaf-1 in the presence of cytochrome c and dATP recruits Caspase 9 to form amultimeric complex. Proteolytic activation of Caspase 9 in turn leads to the initiation of acaspase cascade that includes Caspase 3, resulting in the morphologic features of apoptoticcell death.
Bcl-2 proteins are evolutionarily conserved regulators of apoptosis (Adams and Cory, 1998).The Bcl-2 family is composed of both proapoptotic and anti-apoptotic family members. Thebest characterized anti-apoptotic proteins, Bcl-2 and Bcl-xL, appear to directly or indirectlypreserve the integrity of the outer mitochondrial membrane, thus preventing cytochrome crelease and cell death initiation through the cell intrinsic death pathway (Kluck et al., 1997;Vander Heiden et al., 1997; Yang et al., 1997). In contrast, the proapoptotic family membersBax, Bid, and Bak have been shown to promote cyto-chrome c release (Jurgensmeier et al.,1998; Li et al., 1998; Luo et al., 1998; Kluck et al., 1999; Wei et al., 2000). Mice lackingBcl-2, Bcl-xL, Bax, or Bid have been reported (Nakayama et al., 1993; Veis et al., 1993;Knudson et al., 1995; Motoyama et al., 1995; Yin et al., 1999). Bcl-2-deficient micedemonstrate defects in renal development and undergo premature loss of lymphoid cells andmelanocytes, resulting in immunodeficiency and loss of hair pigmentation, respectively(Nakayama et al., 1993; Veis et al., 1993). Animals deficient in Bcl-xL die duringembryogenesis, displaying a massive increase in apoptosis in developing postmitotic cells ofthe embryonic spinal cord, brain stem, and dorsal root ganglia (Motoyama et al., 1995).Extensive apoptosis is also observed within the liver and hematopoietic systems.
Animals lacking the proapoptotic family member Bax are viable, demonstrating that Bax isnot required for development (Knudson et al., 1995). However, male bax–/– mice are steriledue to a developmental arrest in spermatogenesis that is associated with a paradoxicalincrease in the death of spermatogonia. Bax-deficient animals also have increased numbersof sympathetic and motor neurons. During in vitro culture, sympathetic neurons from Bax-deficient animals are resistant to cell death upon trophic factor deprivation.
Although Bcl-2 proteins are thought primarily to be regulators of the intrinsic apoptoticpathway, recent evidence suggests that proapoptotic Bcl-2 family members can act to
Lindsten et al. Page 2
Mol Cell. Author manuscript; available in PMC 2011 March 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

promote cell death in the extrinsic pathway as well. Bid has been shown to be a substrate forCaspase 8 (Li et al., 1998; Luo et al., 1998). Caspase 8 cleavage of Bid results intranslocation of truncated Bid to the mitochondria where it promotes cytochrome c release.Surprisingly, Bid-deficient mice are viable without obvious developmental abnormalities(Yin et al., 1999). However, bid–/– mice display decreased hepatocellular death in responseto in vivo administration of an anti-Fas antibody.
Bak represents another proapoptotic Bcl-2 family member that is closely related to Bax andis roughly comparable to Bax as assayed by in vitro systems (Chittenden et al., 1995; Orthand Dixit, 1997; Tao et al., 1997). In addition, Bak is expressed in essentially all organs,suggesting that it may be a regulator of apoptosis in multiple cell types (Farrow et al., 1995;Kiefer et al., 1995; Krajewski et al., 1996). To investigate this possibility, we generatedBak-deficient mice by gene targeting. Bak-deficient mice failed to demonstrate observabledevelopmental abnormalities. Both male and female animals were capable of reproduction.Bak-deficient mice did not develop any reproducible abnormalities as they aged, and cellsisolated from these mice behaved normally when apoptosis was initiated through either thecell-intrinsic or cell-extrinsic pathways. These data suggest that either Bak is not an essentialregulator of apoptosis or the function of Bak is redundant with that of other broadlyexpressed proapoptotic Bcl-2 family members.
Since Bax also displays a widespread tissue distribution and bax–/– mice have relativelylimited phenotypic abnormalities (Oltvai et al., 1993; Knudson et al., 1995), we undertook tobreed bax–/–bak–/– mice. The majority of bax–/–bak–/– mice died perinatally with fewer than10% of the animals reaching adulthood. bax–/–bak–/– adult mice displayed multiplephenotypic abnormalities. Adult bax–/–bak–/– mice retained interdigital webs throughout life,and females had imperforate vaginas. The animals displayed multiple neurologicabnormalities, including deafness, circling behavior, and persistence of a large number ofundifferentiated cells in the periventricular progenitor zones. The hematopoietic system wasalso grossly abnormal, with increased numbers of hematopoietic progenitors and elevatednumbers of granulocytes and lymphocytes. Progressive accumulation of mature B and Tcells led to massive enlargement of the spleen and lymph nodes and infiltration ofparenchymal organs. In in vitro culture, cells from bax–/–bak–/– mice were resistant to deathinduced by neglect or irradiation but could initiate apoptosis in response to Fas cross-linking. Together, these data suggest that Bax and Bak represent redundant proteinsinvolved in the regulation of apoptosis. Simultaneous deficiency in both Bak and Bax resultsin abnormalities associated with the accumulation of cells that normally die either duringdevelopment or during the regulation of tissue homeostasis.
ResultsGeneration of bak–/– Mice
A targeting construct was made that eliminated exons III–VI, which encode the Bcl-2homology (BH) domains 1–3 of the bak gene (Figure 1A). The linearized construct waselectroporated into R1 ES cells, and transfectants were selected by G418 resistance. Fourindependent homologous recombinants were obtained, confirmed by Southern blotting, andinjected into C57BL/6 blastocysts. Chimeric mice were identified and crossed with C57BL/6 mice to generate heterozygous Bak mutants. Heterozygous mice were crossed to generatehomozygous Bak-deficient (bak–/–) mice. Of 77 offspring from initial heterozygote matings,23 were wild type, 30 were heterozygous, and 24 were homozygous mutants. A lack of Bakexpression in the homozygous mutants was confirmed by Western blot analysis (Figure 1B).The bak–/– mice appeared outwardly normal. Autopsy and microscopic evaluation ofmultiple tissues failed to reveal any significant lesions. Assays for cell survival in responseto a variety of apoptotic stimuli revealed no statistical differences in in vitro survival of
Lindsten et al. Page 3
Mol Cell. Author manuscript; available in PMC 2011 March 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

bak–/– cells in comparison to wild-type controls. Both bak–/– males and females were fertileand bred well. In over one year of observation, bak–/– animals have not developed anyreproducible age-related disorders.
Generation of bax–/–bak–/– MiceThe above data suggested that either Bak represents a nonessential gene or that the functionof Bak is redundant with other proapoptotic Bcl-2 family members. To investigate this latterpossibility, we sought to determine whether breeding Bak and Bax mutations together wouldunmask more significant phenotypic abnormalities than those observed in either singledeficient animal. To generate bax–/–bak–/– animals, bak+/– mice were crossed with bax+/–
mice. Offspring that were bax+/–bak+/– were identified and interbred. Genotyping atweaning of offspring from these crosses revealed that bax–/–bak–/– animals were obtained atless than 10% of the expected Mendelian frequency. Furthermore, prior to weaning anumber of dead pups were identified in these litters. Genotyping of these dead animalsshowed that many were bax–/–bak–/–. This suggested that bax–/–bak–/– animals were dying inthe perinatal period. To assess this further, breeding pairs of bax+/–bak–/– animals wereestablished and the offspring of six sequential litters were sacrificed at birth. Of 37 miceproduced by intercrossing bax+/–bak–/– mice, 8 were found to be bax+/+bak–/–, 23 werebax+/–bak–/–, and 6 were bax–/–bak–/–. Careful observation of additional litters from thesematings revealed individual animals that failed to nurse appropriately and which werefrequently found isolated from their siblings and mother in the cage. Genotyping of theseanimals demonstrated that they were bax–/–bak–/– mice. Although frequent observationallowed us to reproducibly observe these animals prior to death, the majority died within 48hr of birth.
Persistence of Interdigital Tissue in Adult bax–/–bak–/– MiceA significant number of bax–/–bak–/– mice survived into adulthood, and these adult animalswere characterized in detail at 12–16 weeks of age. Adult bax–/–bak–/– mice displayedseveral phenotypic abnormalities upon gross examination, the most striking of which is thatthe bax–/–bak–/– mice retained interdigital webs on both their fore and rear paws, as shownin Figure 2A. bax+/+bak+/–, bax+/+bak–/–, and bax–/–bak+/+ mice did not have interdigitalwebs. In Figure 2B, the dorsal and palmar views of paws from a bax+/–bak–/– mouse andfrom a bax–/–bak–/– mouse are shown. The presence of just one copy of wild-type baxcompletely eliminated interdigital webbing. A single wild-type copy of bak also eliminatedthe webbing (data not shown).
Failure to Develop an External Vaginal IntroitisA second visible abnormality of bax–/–bak–/– mice was that all adult females had animperforate vagina (Figure 2C). The presence of an imperforate vagina was not observed ineither bax–/– or bak–/– mice (data not shown). Again, the presence of a single wild-typeallele of either bax or bak was sufficient to reproducibly eliminate this anatomic abnormality(Figure 2C and data not shown).
Neurological Abnormalitiesbax–/–bak–/– mice were all unresponsive to auditory stimulation. In addition, most doublemutant mice displayed circling behavior when exposed to external stress. External stress,such as cage changing, also appeared to elicit seizure activity in a number of bax–/–bak–/–
mice. However, the exencephaly and cranial bone defects that have been observed at birth inCaspase 9–/– and/or Apaf-1–/– (Cecconi et al., 1998; Kuida et al., 1998; Yoshida et al., 1998;Honarpour et al., 2000) mice were not observed in bax–/–bak–/– mice.
Lindsten et al. Page 4
Mol Cell. Author manuscript; available in PMC 2011 March 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Histological Analysis of Brain Tissue from bax–/–bak–/– MiceTo assess the central nervous system of bax–/–bak–/– mice, brains were dissected from thesemice as well as control littermates with other genotypes. Overall, brains of the bax–/–bak–/–
mutant mice were larger than those of other genotypes examined (data not shown). Uponhistological analysis, there appeared to be a generalized increase in the number of neurons inmultiple regions of the brain. However, the most striking abnormality of the bax–/–bak–/–
brains was the large accumulation of small neuronal cells with dense chromatin stainingobserved in the periventricular region (Figure 3A). Similar accumulations of such cells arefound during normal embryogenesis in the same region (equivalent to the ganglioniceminence). This region is known to be populated by neural stem cells and an area whereduring terminal division some daughter cells differentiate into postmitotic neurons whileother daughter cells undergo programmed cell death (Bayer and Altman, 1991; Clarke et al.,2000). In adult bax–/–bak–/– mice, similar populations of small cells were also found in otherlocations where neural stem cells are commonly found, including the hippocampus,cerebellum, and olfactory bulb (data not shown). In addition, similar accumulations of smallcells were found in regions not commonly known to contain neural stem cells in adult mice,such as the dorsal mid-line of the mesencephalon (Figure 3B). A gene dosage effect for thisphenotype was evident as both single bax–/– and single bak–/– brains displayed discreteaccumulations of small cells in the periventricular area (Figure 3A). However, bax–/–bak–/–
mice had a dramatic increase in both the number and distribution of these cells whencompared to animals lacking either Bax or Bak alone.
Given the hematopoietic defects seen elsewhere in the bax–/–bak–/– mice (see below), weexplored the possibility that the ectopic cells in the CNS were of hematopoietic origin.Tissue sections from bax–/–bak–/– brains were stained with anti-CD45, a marker expressedon a broad spectrum of hematopoietic cells. While immunoreactive CD45+ hematopoieticcells were identified within blood vessels, the small cell population in the neuralparenchyma of bax–/–bak–/– mice was CD45– (Figure 3B).
Absence of Developmental Defects in Other Parenchymal OrgansDespite the abnormalities observed in the central nervous system and reproductive systemsof bax–/–bak–/– mice, most other organs appeared to be formed normally duringdevelopment. No gross anatomical or histologic abnormalities were observed in the kidney,liver, lung, heart, pancreas, or bladder in either bak–/– or bax–/– bak–/– animals. To confirmthat these internal organs were also functioning at physiologic levels, standard bloodchemistries were determined for bax–/–bak–/– mice and appropriate littermate controls. Noconsistent abnormalities in electrolytes, blood glucose, renal function, or hepatic functiontests were found in either bak–/– or bax–/–bak–/– animals (data not shown). bax–/– micedisplay defective spermatogenesis that is associated with a developmental arrest ofspermatogonia. Histology of the testes from bax–/–bak–/– male mice was indistinguishablefrom that observed in bax–/– littermates (data not shown).
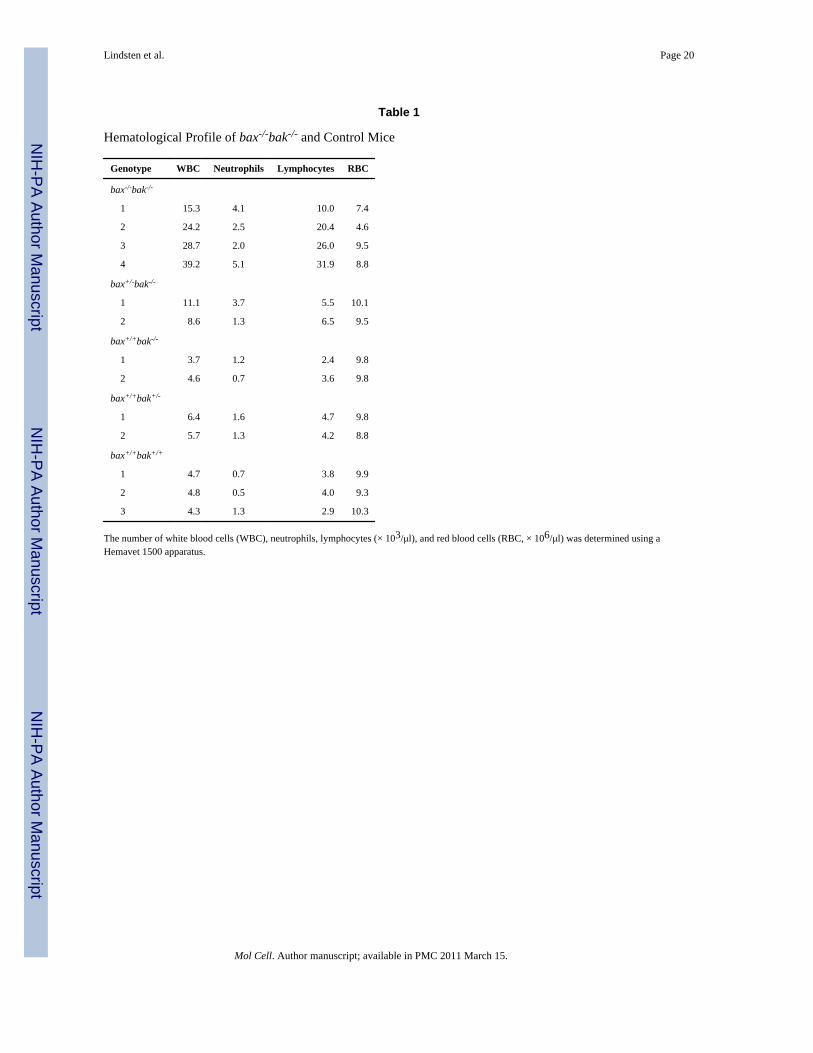
Increased Numbers of Myeloid and Lymphoid Cells in bax–/–bak–/– AnimalsBlood cell counts revealed markedly elevated white blood cell counts in bax–/–bak–/– mice(Table 1). Statistically significant increases in both myeloid cells and lymphoid cells werepresent. In addition, bax–/–bak–/– animals displayed mild anemia and thrombocytopenia thatwere not observed in either bax–/– or bak–/– animals. Hematopoietic colony assays revealedconsistent and statistically significant increases in the number of myeloid colony formingunits (CFU)/106 bone marrow cells (Table 2). A gene dosage effect also appeared to bepresent as the total numbers of CFU increased in rough proportion to the total number ofmutant bak and bax alleles present. Mild elevations in erythroid and megakaryocyte CFUwere also observed.
Lindsten et al. Page 5
Mol Cell. Author manuscript; available in PMC 2011 March 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Massive Lymphoid Accumulation in bax–/–bak–/– MiceThe majority of the increase in the white blood cell count of bax–/–bak–/– animals was due toan elevation in the number of lymphocytes. bax–/–bak–/– mice had increased numbers oflymphocytes not only within the circulation but also within the peripheral lymphoid organs.As described previously (Knudson et al., 1995), animals deficient in Bax had spleens andlymph nodes that were slightly enlarged over wild-type mice (Figure 4A). However,bax–/–bak–/– mice have a significant further enlargement of the peripheral lymphoid organs.Dramatic splenomegaly was observed in all bax–/–bak–/– mutant mice examined, and Figure4B shows a histologic section of the spleen from a bax–/–bak–/– mouse. The bax–/–bak–/–
spleen had an expanded red pulp region that, by histological analysis, contained a largeincrease in the number of plasma cells and histiocytes. A pronounced hyperplasia of thewhite pulp was also observed.
In addition to lymphoid accumulation in the peripheral lymphoid organs, lymphocyticinfiltration of parenchymal organs was also observed. Lymphocyte infiltration in the liverwas periportal as well as periarterial (Figure 4B). The kidney also displayed perivascularlymphocytic infiltration. Several older bax–/–bak–/– mice also displayed varying degrees ofglomerulonephropathy characterized by hypercellularity.
Lymphocytes that Accumulated in bax–/–bak–/– Mice Had a Memory Cell PhenotypeTo determine the phenotypic properties of the lymphocytes that accumulate in bax–/–bak–/–
mice, lymphocytes from the peripheral lymph nodes and spleens were phenotyped by flowcytometry. Lymph nodes from both control and bax–/–bak–/– animals had similar frequenciesof CD4 and CD8 T cells (Figure 5A). However, the T cells that accumulate in thebax–/–bak–/– animals were skewed toward a memory cell phenotype. The T cells were smallin size as measured by forward angle light scatter, lacked expression of activation markerssuch as CD25, had increased expression of the memory cell marker, CD44, and decreasedexpression of the peripheral addressin, CD62L (Figure 5A). The percentages of B cells werealso found to be similar in bax–/–bak–/– lymph nodes as compared to control lymph nodes.However, the B220+ B cells present in the lymph node were skewed toward a B220bright
IgD– phenotype, indicating the presence of an increased number of class-switched ormemory B cells (Figure 5B). Similar data were obtained in spleen (data not shown). T and Bcells isolated from either bax–/– or bak–/– single-deficient mice did not display increasedpercentages of cells with a memory phenotype.
Lymphocytes from bax–/–bak–/– Mice Were Resistant to Death by NeglectOf the defects observed in adult bax–/–bak–/– mice, only the lymphocyte accumulation wasnot also observed in animals sacrificed at birth. Thus, the combined deficiency of Bax andBak led to the accumulation of lymphocytes as the animals aged. The increase inlymphocyte number could occur either as a consequence of increased production orincreased survival. The small size and lack of expression of activation markers such asCD25 on lymphocytes in both lymphoid and nonlymphoid organs of the bax–/–bak–/–
animals suggest that the cells accumulate as a result of a defect in apoptosis. To address thisissue, splenocytes from bax–/–bak–/– and control mice were isolated, and cell survival insuspension culture was analyzed over the ensuing 4 days. Figure 5C shows the percentage oflive cells as determined by propidium iodide staining at daily intervals. Splenocytes frombax–/–bak–/– mice showed markedly enhanced survival, demonstrating a defect in death byneglect. In contrast, cells from Bak-deficient animals and animals expressing both Bax andBak underwent apoptosis when placed in single cell culture. The ability of bax–/– lymphoidcells to undergo death by neglect has been previously analyzed and found to be similar towild-type mice (Knudson, et al., 1995 and data not shown).
Lindsten et al. Page 6
Mol Cell. Author manuscript; available in PMC 2011 March 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Impaired Radiation- and Etoposide-Induced but Normal Fas-Induced Death in bax–/–bak–/–
MiceOne of the classic assays for apoptosis is the radiation-induced death of thymocytes (Wyllie,1980). Irradiation of thymocytes has been shown to initiate apoptosis in a p53-dependentfashion (Clarke et al., 1993; Lowe et al., 1993). Cell death appears to result from activationof the cell-intrinsic pathway involving mitochondria (Kuida et al., 1998). Neither bax–/– norbak–/– animals displayed defects in radiation-induced apoptosis (Figure 6A and Knudson etal., 1995). Thymocytes from bax–/–bak–/– animals displayed enhanced survival compared tolittermate controls when exposed to gamma irradiation. Thymocyte cell death followingtreatment with the chemotherapeutic agent etoposide is also reported to be p53-dependent.Thymocytes from bax–/–bak–/– were found to be resistant to treatment with etoposide(Figure 6B), whereas bax–/– and bak–/– single deficient thymocytes were sensitive to thistreatment. In contrast, when thymocytes from bax–/–bak–/– and control mice were subjectedto cross-linking of the Fas receptor, bax–/–bak–/– thymocytes showed sensitivity similar tothat of the control thymocytes (Figure 6C). This indicates that, at least in this tissue, the cell-extrinsic pathway is not dependent on the proapoptotic activity of either Bax or Bak.
DiscussionThe phenotype of bax–/–bak–/– mice provides a clear demonstration of the importance ofproapoptotic Bcl-2 proteins in the regulation of both mammalian development and tissuehomeostasis. The combined deficiency of Bax and Bak has a significantly more severe anddiverse phenotype than deficiency in either gene alone. Bak deficiency alone has nodemonstrable phenotype that we have as yet been able to uncover. Bax-deficient animals areviable and their major developmental defect is male sterility that is associated with adevelopmental arrest of the spermatogonia compartment with a resultant increase inapoptotic germ cell death (Knudson et al., 1995). Although slight increases in sympatheticneurons, motor neurons, and lymphocytes have been described in the absence of Bax, bax–/–
mice appear to be behaviorally normal, they fail to develop any significantlymphoproliferative complications, and the females are capable of reproduction (Knudson etal., 1995; Deckwerth et al., 1996). In contrast, bax–/–bak–/– animals had significantdisruptions in the development of the digits, female reproductive tract, and the centralnervous system. bax–/–bak–/– mice appear to have increased number of hematopoieticprecursors and develop a progressive accumulation of mature lymphocytes. Lymphocytesthat accumulate in bax–/–bak–/– mice displayed defects in death by neglect and were alsoresistant to radiation- and etoposide-induced apoptosis. However, the ability of bax–/–bak–/–
thymocytes to initiate apoptosis in response to Fas signal transduction was comparable tothat of wild-type thymocytes. Together, the data indicate that Bax and Bak have largelyoverlapping roles in regulating cell death initiated by activators of the intrinsic cell deathpathway. However, at least in some tissues, the extrinsic cell death pathway can initiateapoptosis in the absence of both Bax and Bak.
The phenotype of bax–/–bak–/– mice overlaps the phenotypes described for mice lackingindividual genes involved in the intrinsic cell death pathway. In fact, the bax–/–bak–/– micein most respects have a more severe phenotype than those described for Apaf-1–/– orCaspase 9–/– mice (Cecconi et al., 1998; Kuida et al., 1998; Yoshida et al., 1998; Honarpouret al., 2000). The accumulation of lymphoid tissue and the failure of vaginal developmentobserved in bax–/–bak–/– mice has not been described in adult Apaf-1–/– or Caspase 9–/–
mice. The resorption of interdigital webs was completed during embryogenesis in bothApaf-1–/– and Caspase 9–/– mice. In addition, the adult mice deficient in Caspase 9 orApaf-1 that have been analyzed do not demonstrate any morphologic abnormalities in theircentral nervous systems. In contrast, all of the bax–/–bak–/– mice that we have analyzed
Lindsten et al. Page 7
Mol Cell. Author manuscript; available in PMC 2011 March 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

demonstrate persistence of increased numbers of neuronal cells in the periventricularprogenitor regions.
Like Apaf-1–/– and Caspase 9–/– mice, the majority of bax–/–bak–/– mice died perinatally.However, unlike the Caspase 9–/– and Apaf-1–/– mice, we have not seen any evidence ofexencephaly in the bax–/–bak–/– animals, nor have the defects in cranial bone formationreported for Apaf-1–/– mice been observed in the bax–/–bak–/– mice. Caspase 9–/– animalshave been reported to have obstruction of the ventricles as a result of expansion in the sizeof the periventricular progenitor zones (Kuida et al., 1998). Although the periventricularprogenitor zones persist into adulthood in the bax–/–bak–/– mice, hydrocephalus does notappear to be an explanation for the perinatal death of bax–/–bak–/– animals. In thebax–/–bak–/– mice, death seemed to occur because of a failure of the newborn animals tonurse. bax–/–bak–/– pups were frequently found still alive but neglected by their mother andseparated from the rest of their litter. Attempts at foster mothering these animals have failed,suggesting that the bax–/–bak–/– animals have a defect in suckling or social behavior. Thiscould result from the neuronal developmental abnormalities such as those that lead to theloss of response to auditory stimulation and the circling behavior observed in adults. Theseabnormalities are commonly seen when the development of neuroepithelial structures isabnormal (Alagramam et al., 1999). Abnormalities with optic and inner ear developmenthave been reported in Apaf-1–/– mice (Honarpour et al., 2000). Alternatively, abnormalitiesin cranial bone formation can also result in hearing and behavioral abnormalities, and thismight result from a milder form of the cranioschisis seen in Apaf-1–/– mice (Yoshida et al.,1998). Regardless, suckling is a complex behavioral trait, and the failure of bax–/–bak–/–
mice to suckle effectively does not appear to have a simple, readily apparent mechanisticbasis.
The developmental and homeostatic defects in the bax–/–bak–/– mice indicate that Bax andBak are redundant regulators of the intrinsic cell death pathway. This redundancy offunction is not observed in the other genes involved in the intrinsic cell death pathway,including Apaf-1, Caspase 9, and Caspase 3. The failure of bax–/–bak–/– mice to develop theexencephaly observed in Apaf-1–/– and Caspase 9–/– mice could result from additionalredundancy among proapoptotic Bcl-2 family members in developing neural tissue or fromcompensatory changes in the expression of anti-apoptotic Bcl-2 family members within thecentral nervous system. Alternatively, lack of exencephaly may result from the ability ofApaf-1 and Caspase 9 to act as amplifiers of the cell death decision downstream ofmitochondrial cytochrome c release and somehow compensate for the lack of Bax and Bakduring the developmental elimination of excess neural cells.
The ability of some Apaf-1–/– and Caspase 9–/– mice to develop normally has beeninterpreted to suggest that there must be redundant apoptotic pathways that serve to causethe elimination of interdigital webs, remove excess neuroprogenitors, and prevent theaccumulation of lymphoid cells (Kuida et al., 1998; Honarpour et al., 2000). The fact thatthese excess accumulations of cells persist in bax–/–bak–/– mice suggests that theseadditional mediators of cell death are dependent on Bax and/or Bak function. It has recentlybeen described that other proteins in addition to cytochrome c can be released frommitochondria that can promote apoptosis. These include AIF, which upon release appears totranslocate to the nucleus and promote DNA degradation (Susin et al., 1999), and Smac/DIABLO, which can block the ability of IAP proteins to act as caspase inhibitors (Du et al.,2000; Verhagen et al., 2000). The Bax- and/or Bak-dependent release of additionalproapoptotic proteins such as AIF and Smac/DIABLO from mitochondria could potentiallyinduce cell death in the absence of a functional cytochrome c/Apaf-1/Caspase 9 complex.
Lindsten et al. Page 8
Mol Cell. Author manuscript; available in PMC 2011 March 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Bax and Bak have also been proposed as important mediators of the cell-extrinsic deathpathway, acting as molecules that interact with the caspase substrate, Bid, to inducecytochrome c release from mitochondria (Eskes et al., 2000; Wei et al., 2000). At least indeveloping thymocytes, neither Bax nor Bak is required for Fas-induced death. Fas signaltransduction also induces thymocyte death in Apaf-1–/– and Caspase 9–/– mice (Kuida et al.,1998; Yoshida et al., 1998). Thus, communication between the extrinsic and intrinsicpathways is not essential for Fas-mediated death in all tissues. Alternatively, additionalproapoptotic Bcl-2-related proteins may also interact with Bid to carry out this function.
In summary, these data demonstrate the importance of proapoptotic Bcl-2 family membersin the developmental involution of interdigital webs and of neuronal cells in theperiventricular progenitor zones. The abnormal development of the female reproductive tractand the absence of hearing support previous studies that suggested that the function of Bcl-2family genes may be important in the development of these complex tissues (Ishii et al.,1996; Rodriguez et al., 1997). Finally, the progressive accumulation of lymphoid cellsdemonstrates the importance of these genes in the control of tissue homeostasis in adults. Acommon feature of many of the excess cells that accumulate in the bax–/–bak–/– mice is thatthese are cells thought to be eliminated in normal animals as a result of the loss of cellsurvival signaling. Cell death induced by growth factor withdrawal (death by neglect) can beinhibited in multiple tissues by targeted transgene expression of anti-apoptotic Bcl-2proteins. The data presented here demonstrate that death by neglect can also be impaired bythe simultaneous deficiency in two proapoptotic Bcl-2 family members. Unlike other centralapoptotic proteins, Bax and Bak appear to play overlapping roles in regulating theelimination of excess cells during both embryonic development and tissue homeostasis.
Experimental ProceduresGeneration of Knockout Mice
The genomic organization for the bak mouse gene has been described (Ulrich et al., 1997).A targeting construct was made that replaced exons III–VI with the 1.2 kb Xho I–Bam HIfragment of pMCIneo Poly(A) (Stratagene, La Jolla, CA). This deletion removed 235 aafrom the 254 aa coding region of Bak, including the entire BH1, BH2, and BH3 domains.The 59 flank of the targeting construct contained the 4.3 kb Bgl II–SalI fragment, and the 39flank contained the 1.2 kb BglII–XhoI fragment, as depicted in Figure 1A. The bak targetingconstruct was electroporated into R1 ES cells (Nagy et al., 1993). Electroporation, selection,and maintenance of ES cells have been previously described (Shiels et al., 2000). Theresulting expanded ES cell clones were screened for homologous recombination by Southernblotting. Four independent ES cell lines were identified and injected as described (Shiels etal., 2000). The generation of Bax mutant mice has been previously described (Knudson etal., 1995). bax–/–bak–/– mice were generated from matings of bax+/–bak+/– parents. Theseparental mice were generated from outcrosses of B6.129X1 (N9) bax+/– mice mated withoutcrosses of (129S3 × 129X1) × C57BL/6 bak+/– mice. Offspring were genotyped andbax+/–bak+/– mice set up in breeding pairs. In more recent breedings, set up to maximize forthe yield of double mutant mice, breedings were also set up with mice that werebax+/–bak–/–. Mice were genotyped by PCR of DNA samples from tail snips. The wild-typebak allele was identified using sense primer 5′-GGTGTCCACACTAGAGAACTACTC andanti-sense primer 5′-GAGCCATGAAGATGTTTAGC. The mutant bak allele was identifiedusing a neomycin gene sense primer (5′-GCCCGGTTCTTTTTGTCAAGACCGA). Theanti-sense primer for the mutant bak allele was the same as that one used for the wild-typeallele. The wild-type bax allele was identified using sense primer 5′-TGATCAGAACCATCATG and anti-sense primer 5′-GTTGACCAGAGTGGCGTAGG.To detect the bax null allele, a neomycin gene-specific primer (5′-
Lindsten et al. Page 9
Mol Cell. Author manuscript; available in PMC 2011 March 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

CCGCTTCCATTGCTCAGCGG) was used in combination with the wild-type bax anti-sense primer described above.
Western Blot AnalysisSingle cell suspensions from thymus and spleen of mice were prepared and lysed in RIPAbuffer (10 mM Tris [pH 8], 140 mM NaCl, 1% deoxycholate, 0.1% SDS, 1% Triton X-100,0.025% NaN3 with the addition of a protease inhibitor cocktail [Roche, Indianapolis, IN]).The protein concentration of each lysate was determined by a colorimetric assay (BCAProtein Assay, Pierce, Rockford, IL), and 20 mg of total protein was resolved on a 12%precast SDS–polyacrylamide gel (Novex, San Diego, CA). After transfer to nitrocellu-loseusing standard techniques, the Western blot was probed with a polyclonal anti-Bak antibody(Upstate Biotechnology, Lake Placid, NY).
Gross and Histopathological AnalysisAfter euthanasia, heart blood was collected and analyzed in a Hemavet 1500 apparatus(CDC Technologies Inc., Oxford, CT). A variety of tissues were harvested and fixed in 10%phosphate-buffered formalin (ACCRA Lab, Swedesboro, NJ). Brains were freed from theskull bone before fixation. Tissues were embedded, sectioned, mounted, and stained withhematoxylin-eosin using standard techniques. Brains were sectioned coronally and stainedwith cresyl violet. Photographs of slides were taken using an ES 800 Nikon microscope anda MicroMAX-5MHz-1300Y digital camera (Princeton Instruments, Inc. Trenton NJ) andanalyzed with the MetaMorph Imaging System (Universal Imaging Corporation, WestChester, PA). Single cell suspensions were also made from lymphoid tissues for furtheranalysis. For immunohistochemistry, 10 μm thick sections were deparaffinized, andimmunohistochemistry was performed using standard protocols (Kendler and Golden,1996). The primary antibody was anti-CD45RB (PharMingen, San Diego, CA), whichrecognizes B cells, most subsets of T cells, macrophages, and dendritic cells, used at 1 μg/ml.
Flow CytometryFlow cytometry was done on a FACS Calibur, and data was analyzed using Cell Questsoftware (Becton Dickinson; Mountain View, CA). Antibodies used include: CD4-CyChrome (clone H129.19; Phar-Mingen, San Diego, CA); CD8-APC (clone 53–6.7;PharMingen); CD44-CyChrome (clone IM7; PharMingen); CD62L-APC (clone MEL-14;PharMingen); B220-APC (clone RA3–6B2; PharMingen); Thy1.2-PE (clone OX-7;PharMingen); and IgD-FITC (clone 11–26c.2a; Phar-Mingen). Cell viabilities weredetermined by propidium iodide exclusion (PI; Molecular Probes, Eugene, OR).
Lymphocyte Death AssaysSplenocytes were isolated from mice with different genotypes. 1 × 105 cells were plated ineach well of 96-well microtiter plates. Cells were cultured for 0, 1, 2, 3, or 4 days in RPMI1640 medium supplemented with 10% FBS, 100 U/ml penicillin, 100 μg/ml streptomycin, 4mM L-glutamine, 5.5 × 10–5 M β-mercaptoethanol, and 10 mM Hepes solution. Thymocytedeath assay was performed as described (Kuida et al., 1998). Briefly, thymocytes wereisolated from mice with different genotypes. One million cells were treated with anti-Fasantibody (Jo2, PharMingen) plus 30 μg/ml cycloheximide (Sigma), γ radiation (500 rad,Gammacell), indicated doses of etopo-side (Clontech, Palo Alto, CA) or complete mediumonly. Cell viabilities were determined by PI exclusion.
Lindsten et al. Page 10
Mol Cell. Author manuscript; available in PMC 2011 March 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Bone Marrow Hematopoietic Progenitor AssaysBone marrow was harvested as described (Adelman et al., 1999). Briefly, the femur andtibia were dissected free from the surrounding muscle and tendons. Marrow was expungedwith PBS (without Ca2+ or Mg2+), supplemented with 2% FBS and 1% pen/strep. Red cellswere lysed and the remaining cells were counted using a Coulter Z2 Counter. 2 × 104 cellswere plated in “complete” methylcellulose containing 15% FBS, 50 ng/ml rmSCF, 10 ng/mlrmIL-3, 10 ng/ml rhIL-6, 3 U/ml rhEPO, 10 μg/ml bovine insulin, 200 μg/ml humantransferrin, 1% BSA, and 10–4 M β-ME (Stem Cell Technologies). Cultures were set up intriplicate and cultured at 37°C in air plus 5% CO2 for 5 days. Colony forming units (CFU)were scored based on colony morphology and cytological staining. CFU-GEMM, CFU-M,CFU-GM, and CFU-G were scored as myeloid colonies; CFU-E were scored as erythroidcolonies; and CFU-Meg were scored as megakaryocyte colonies.
AcknowledgmentsWe thank the other members of the laboratories for helpful discussions and critique of the manuscript. We thankBarbi Judd for expert technical assistance and Susan Kerns for expert editorial assistance. This work was supportedby grants from the National Institutes of Health, the Howard Hughes Medical Institute, and The Abramson FamilyCancer Research Institute.
ReferencesAdams JM, Cory S. The Bcl-2 protein family: arbiters of cell survival. Science 1998;281:1322–1326.
[PubMed: 9735050]Adelman DM, Maltepe E, Simon MC. Multilineage embryonic hematopoiesis requires hypoxic ARNT
activity. Genes Dev 1999;13:2478–2483. [PubMed: 10521392]Alagramam KN, Kwon HY, Cacheiro NL, Stubbs L, Wright CG, Erway LC, Woychik RP. A new
mouse insertional mutation that causes sensorineural deafness and vestibular defects. Genetics1999;152:1691–1699. [PubMed: 10430593]
Bayer, SA.; Altman, J. Neocortical development. Raven Press; New York: 1991.Cecconi F, Alvarez-Bolado G, Meyer BI, Roth KA, Gruss P. Apaf1 (CED-4 homolog) regulates
programmed cell death in mammalian development. Cell 1998;94:727–737. [PubMed: 9753320]Chittenden T, Harrington EA, O'Connor R, Flemington C, Lutz RJ, Evan GI, Guild BC. Induction of
apoptosis by the Bcl-2 homologue Bak. Nature 1995;374:733–736. [PubMed: 7715730]Clarke AR, Purdie CA, Harrison DJ, Morris RG, Bird CC, Hooper ML, Wyllie AH. Thymocyte
apoptosis induced by p53-dependent and independent pathways. Nature 1993;362:849–852.[PubMed: 8479523]
Clarke DL, Johansson CB, Wilbertz J, Veress B, Nilsson E, Karlstrom H, Lendahl U, Frisen J.Generalized potential of adult neural stem cells. Science 2000;288:1660–1663. [PubMed:10834848]
Deckwerth TL, Elliott JL, Knudson CM, Johnson EM, Snider WD, Korsmeyer SJ. BAX is required forneuronal death after trophic factor deprivation and during development. Neuron 1996;17:401–411.[PubMed: 8816704]
Du C, Fang M, Li Y, Li L, Wang X. Smac, a mitochondrial protein that promotes cytochrome c-dependent caspase activation by eliminating IAP inhibition. Cell 2000;102:33–42. [PubMed:10929711]
Eskes R, Desagher S, Antonsson B, Martinou JC. Bid induces the oligomerization and insertion of Baxinto the outer mitochondrial membrane. Mol. Cell. Biol 2000;20:929–935. [PubMed: 10629050]
Farrow SN, White JH, Martinou I, Raven T, Pun KT, Grinham CJ, Martinou JC, Brown R. Cloning ofa bcl-2 homologue by interaction with adenovirus E1B 19K. Nature 1995;374:731–733. Erratum:Nature 375(6530), 1995. [PubMed: 7715729]
Green DR. Apoptotic pathways: paper wraps stone blunts scissors. Cell 2000;102:1–4. [PubMed:10929706]
Lindsten et al. Page 11
Mol Cell. Author manuscript; available in PMC 2011 March 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Honarpour N, Du C, Richardson JA, Hammer RE, Wang X, Herz J. Adult Apaf-1-deficient miceexhibit male infertility. Dev. Biol 2000;218:248–258. [PubMed: 10656767]
Ishii N, Wanaka A, Ohno K, Matsumoto K, Eguchi Y, Mori T, Tsujimoto Y, Tohyama M.Localization of bcl-2, bax, and bcl-x mRNAs in the developing inner ear of the mouse. Brain Res1996;726:123–128. [PubMed: 8836552]
Jurgensmeier JM, Xie Z, Deveraux Q, Ellerby L, Bredesen D, Reed JC. Bax directly induces release ofcytochrome c from isolated mitochondria. Proc. Natl. Acad. Sci. USA 1998;95:4997–5002.[PubMed: 9560217]
Kendler A, Golden JA. Progenitor cell proliferation outside the ventricular and subventricular zonesduring human brain development. J. Neuropathol. Exp. Neurol 1996;55:1253–1258. [PubMed:8957449]
Kiefer MC, Brauer MJ, Powers VC, Wu JJ, Umansky SR, Tomei LD, Barr PJ. Modulation ofapoptosis by the widely distributed Bcl-2 homologue Bak. Nature 1995;374:736–739. [PubMed:7715731]
Kluck RM, Bossy-Wetzel E, Green DR, Newmeyer DD. The release of cytochrome c frommitochondria: a primary site for Bcl-2 regulation of apoptosis. Science 1997;275:1132–1136.[PubMed: 9027315]
Kluck RM, Esposti MD, Perkins G, Renken C, Kuwana T, Bossy-Wetzel E, Goldberg M, Allen T,Barber MJ, Green DR, Newmeyer DD. The pro-apoptotic proteins, Bid and Bax, cause a limitedpermeabilization of the mitochondrial outer membrane that is enhanced by cytosol. J. Cell Biol1999;147:809–822. [PubMed: 10562282]
Knudson CM, Tung KS, Tourtellotte WG, Brown GA, Korsmeyer SJ. Bax-deficient mice withlymphoid hyperplasia and male germ cell death. Science 1995;270:96–99. [PubMed: 7569956]
Krajewski S, Krajewska M, Reed JC. Immunohistochemical analysis of in vivo patterns of Bakexpression, a proapoptotic member of the Bcl-2 protein family. Cancer Res 1996;56:2849–2855.[PubMed: 8665525]
Kuida K, Haydar TF, Kuan CY, Gu Y, Taya C, Karasuyama H, Su MS, Rakic P, Flavell RA. Reducedapoptosis and cytochrome c–mediated caspase activation in mice lacking caspase 9. Cell1998;94:325–337. [PubMed: 9708735]
Li H, Zhu H, Xu CJ, Yuan J. Cleavage of BID by caspase 8 mediates the mitochondrial damage in theFas pathway of apoptosis. Cell 1998;94:491–501. [PubMed: 9727492]
Lowe SW, Schmitt EM, Smith SW, Osborne BA, Jacks T. p53 is required for radiation-inducedapoptosis in mouse thymocytes. Nature 1993;362:847–849. [PubMed: 8479522]
Luo X, Budihardjo I, Zou H, Slaughter C, Wang X. Bid, a Bcl2 interacting protein, mediatescytochrome c release from mitochondria in response to activation of cell surface death receptors.Cell 1998;94:481–490. [PubMed: 9727491]
Motoyama N, Wang F, Roth KA, Sawa H, Nakayama K, Negishi I, Senju S, Zhang Q, Fujii S, et al.Massive cell death of immature hematopoietic cells and neurons in Bcl-x- deficient mice. Science1995;267:1506–1510. [PubMed: 7878471]
Nagata S. Apoptosis by death factor. Cell 1997;88:355–365. [PubMed: 9039262]Nagy A, Rossant J, Nagy R, Abramow-Newerly W, Roder JC. Derivation of completely cell culture–
derived mice from early-passage embryonic stem cells. Proc. Natl. Acad. Sci. USA 1993;90:8424–8428. [PubMed: 8378314]
Nakayama K, Negishi I, Kuida K, Shinkai Y, Louie MC, Fields LE, Lucas PJ, Stewart V, Alt FW, etal. Disappearance of the lymphoid system in Bcl-2 homozygous mutant chimeric mice. Science1993;261:1584–1588. [PubMed: 8372353]
Oltvai ZN, Milliman CL, Korsmeyer SJ. Bcl-2 heterodimerizes in vivo with a conserved homolog,Bax, that accelerates programmed cell death. Cell 1993;74:609–619. [PubMed: 8358790]
Orth K, Dixit VM. Bik and Bak induce apoptosis downstream of CrmA but upstream of inhibitor ofapoptosis. J. Biol. Chem 1997;272:8841–8844. [PubMed: 9082997]
Rodriguez I, Araki K, Khatib K, Martinou JC, Vassalli P. Mouse vaginal opening is an apoptosis-dependent process which can be prevented by the overexpression of Bcl2. Dev. Biol1997;184:115–121. [PubMed: 9142988]
Lindsten et al. Page 12
Mol Cell. Author manuscript; available in PMC 2011 March 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Shiels H, Li X, Schumacker PT, Maltepe E, Padrid PA, Sperling A, Thompson CB, Lindsten T.TRAF4 deficiency leads to tracheal malformation with resulting alterations in airflow to the lungs.Am. J. Pathol 2000;157:679–688. [PubMed: 10934170]
Strasser A, Harris AW, Huang DCS, Krammer PH, Cory S. Bcl-2 and Fas/APO-1 regulate distinctpathways to lymphocyte apoptosis. EMBO J 1995;14:6136–6147. [PubMed: 8557033]
Susin SA, Lorenzo HK, Zamzami N, Marzo I, Snow BE, Brothers GM, Mangion J, Jacotot E,Costantini P, Loeffler M, et al. Molecular characterization of mitochondrial apoptosis-inducingfactor. Nature 1999;397:441–446. [PubMed: 9989411]
Tao W, Kurschner C, Morgan JI. Modulation of cell death in yeast by the Bcl-2 family of proteins. J.Biol. Chem 1997;272:15547–15552. [PubMed: 9182591]
Ulrich E, Kauffmann-Zeh A, Hueber AO, Williamson J, Chittenden T, Ma A, Evan G. Gene structure,cDNA sequence, and expression of murine Bak, a proapoptotic Bcl-2 family member. Genomics1997;44:195–200. [PubMed: 9299236]
Vander Heiden MG, Chandel NS, Williamson EK, Schumacker PT, Thompson CB. Bcl-xL regulatesthe membrane potential and volume homeostasis of mitochondria. Cell 1997;91:627–637.[PubMed: 9393856]
Vaux DL, Korsmeyer SJ. Cell death in development. Cell 1999;96:245–254. [PubMed: 9988219]Veis DJ, Sorenson CM, Shutter JR, Korsmeyer SJ. Bcl-2-deficient mice demonstrate fulminant
lymphoid apoptosis, polycystic kidneys, and hypopigmented hair. Cell 1993;75:229–240.[PubMed: 8402909]
Verhagen AM, Ekert PG, Pakusch M, Silke J, Connolly LM, Reid GE, Moritz RL, Simpson RJ, VauxDL. Identification of DIABLO, a mammalian protein that promotes apoptosis by binding to andantagonizing IAP proteins. Cell 2000;102:43–53. [PubMed: 10929712]
Wei MC, Lindsten T, Mootha VK, Weiler S, Gross A, Ashiya M, Thompson CB, Korsmeyer SJ. tBID,a membrane-targeted death ligand, oligomerizes BAK to release cytochrome c. Genes Dev2000;14:2060–2071. [PubMed: 10950869]
Wyllie AH. Glucocorticoid-induced thymocyte apoptosis is associated with endogenous endonucleaseactivation. Nature 1980;284:555–556. [PubMed: 6245367]
Yang J, Liu X, Bhalla K, Kim CN, Ibrado AM, Cai J, Peng TI, Jones DP, Wang X. Prevention ofapoptosis by Bcl-2: release of cytochrome c from mitochondria blocked. Science 1997;275:1129–1132. [PubMed: 9027314]
Yin XM, Wang K, Gross A, Zhao Y, Zinkel S, Klocke B, Roth KA, Korsmeyer SJ. Bid-deficient miceare resistant to Fas-induced hepatocellular apoptosis. Nature 1999;400:886–891. [PubMed:10476969]
Yoshida H, Kong YY, Yoshida R, Elia AJ, Hakem A, Hakem R, Penninger JM, Mak TW. Apaf1 isrequired for mitochondrial pathways of apoptosis and brain development. Cell 1998;94:739–750.[PubMed: 9753321]
Lindsten et al. Page 13
Mol Cell. Author manuscript; available in PMC 2011 March 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
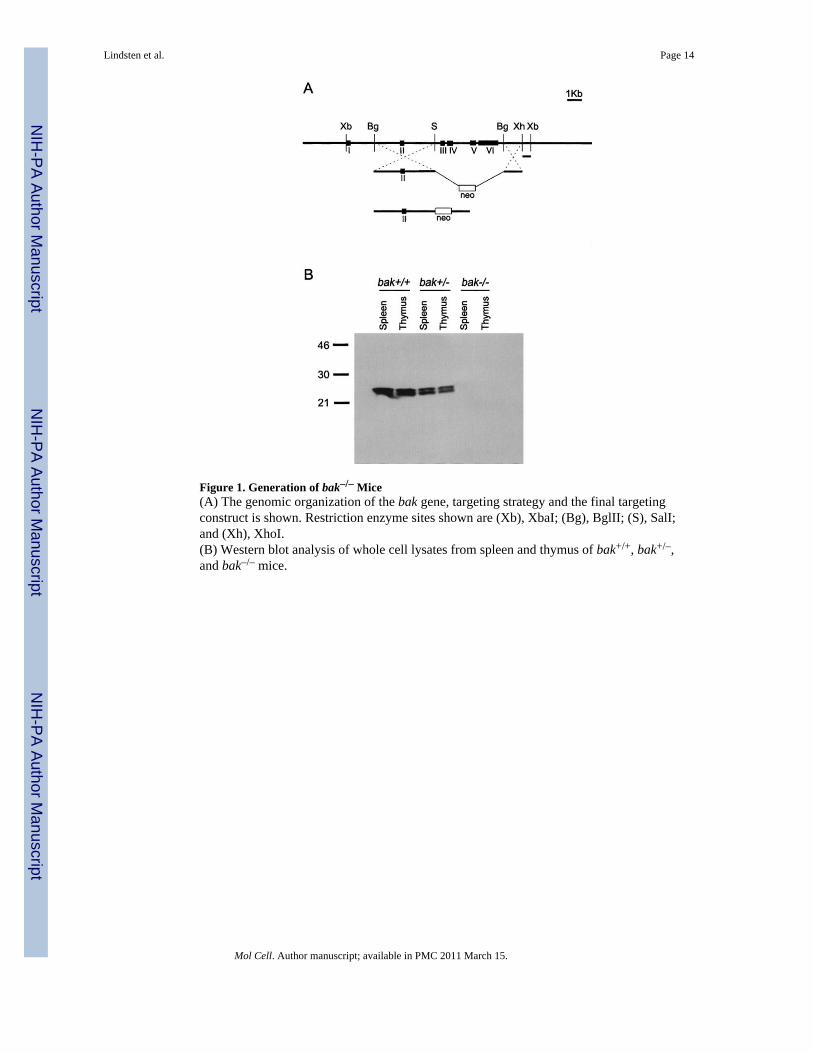
Figure 1. Generation of bak–/– Mice(A) The genomic organization of the bak gene, targeting strategy and the final targetingconstruct is shown. Restriction enzyme sites shown are (Xb), XbaI; (Bg), BglII; (S), SalI;and (Xh), XhoI.(B) Western blot analysis of whole cell lysates from spleen and thymus of bak+/+, bak+/–,and bak–/– mice.
Lindsten et al. Page 14
Mol Cell. Author manuscript; available in PMC 2011 March 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 2. Gross Anatomical Phenotypes of bax–/–bak–/– Mice(A) Photographs of bax+/+bak+/– (a and e), bax+/+bak–/– (b and f), bax–/–bak+/+ (c and g),and bax–/–bak–/– (d and h) mice.(B) Dorsal and ventral views of paws from bax+/–bak–/– (a and c) and bax–/–bak–/– (b and d).(C) Photographs of vaginal openings from bax+/–bak–/– and bax–/–bak–/– mice. Arrows pointto external vaginal region.
Lindsten et al. Page 15
Mol Cell. Author manuscript; available in PMC 2011 March 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 3. Periventricular Accumulations of Small Neuronal Cells in the Brains of bax–/–bak–/–
Mice(A) Histological sections of brain tissue from bax+/+bak+/– (a), bax–/–bak+/+ (b),bax+/–bak–/– (c), and bax–/–bak–/– (d) mice at 25× magnification. Each set is atapproximately the same level and includes the anterior basal ganglia and the corpuscallosum. Arrow points to the periventricular region in all sections.(B) Histological section of the dorsal midline of the mesencephalon from bax–/–bak–/–
mouse at a 400× magnification, stained with anti-CD45RB. Arrows point to CD45+ cells.Arrowhead points to abnormal accumulation of small neuronal cells.
Lindsten et al. Page 16
Mol Cell. Author manuscript; available in PMC 2011 March 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
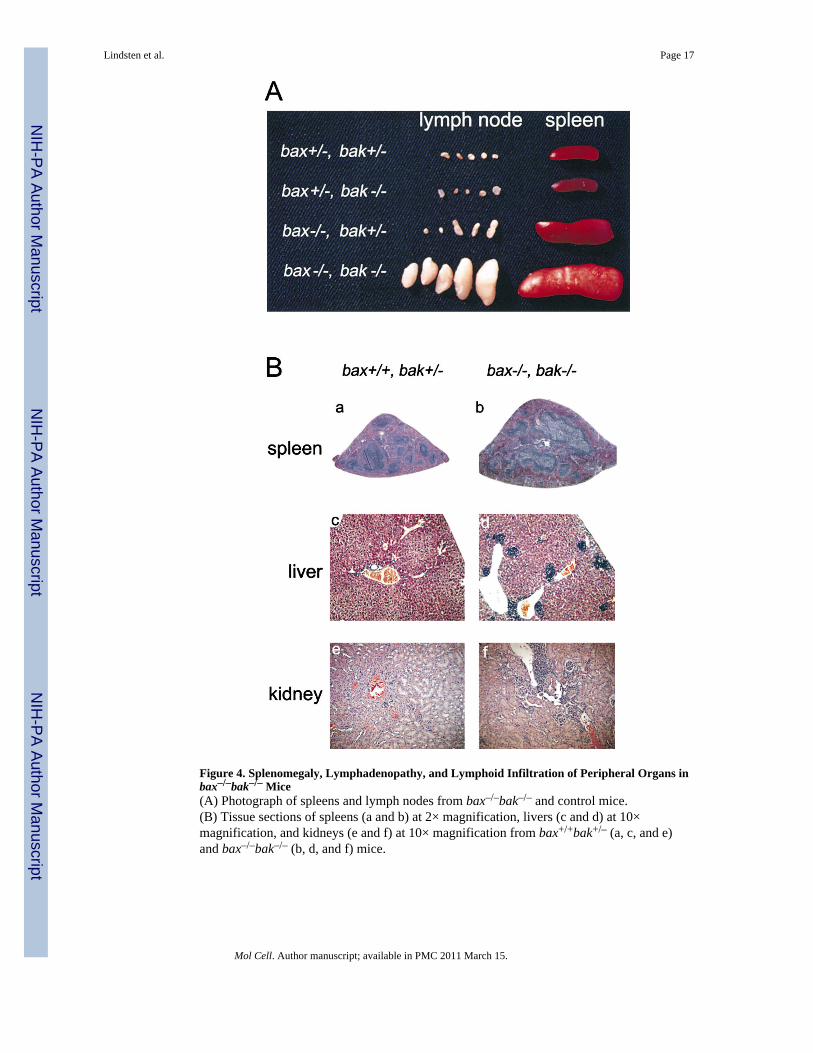
Figure 4. Splenomegaly, Lymphadenopathy, and Lymphoid Infiltration of Peripheral Organs inbax–/–bak–/– Mice(A) Photograph of spleens and lymph nodes from bax–/–bak–/– and control mice.(B) Tissue sections of spleens (a and b) at 2× magnification, livers (c and d) at 10×magnification, and kidneys (e and f) at 10× magnification from bax+/+bak+/– (a, c, and e)and bax–/–bak–/– (b, d, and f) mice.
Lindsten et al. Page 17
Mol Cell. Author manuscript; available in PMC 2011 March 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 5. Peripheral Lymphocytes that Accumulate in bax–/–bak–/– Mice Are Enriched inMemory Cells and Fail to Die by Neglect Peripheral lymphocytes from lymph node cells wereanalyzed by flow cytometry(A and B) CD4 and CD8 expression (A) and Thy1.2 and B220 expression (B). Gated T cellswere also analyzed for the expression of CD44 and CD62L. Gated B cells were alsoanalyzed for expression of IgD.(C) Peripheral lymphocytes from spleens of bax–/–bak–/– (open symbols) and control mice(closed symbols) were cultured for 4 days. Cells were stained with PI and subjected toFACS analysis at the indicated times. Samples were assayed in triplicate.
Lindsten et al. Page 18
Mol Cell. Author manuscript; available in PMC 2011 March 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 6. Thymocyte Death Assays in bax–/–bak–/– Mice(A) Thymocytes from bax–/–bak–/– mice and control mice were treated with 500 rad of γirradiation (closed bars) or left untreated (open bars). Cells were stained with PI andsubjected to FACS analysis after 24 hr of culture. Mean and standard deviations of triplicatesamples from each mouse are presented.(B) Thymocytes were cultured with the indicated amounts of etoposide. Cells were stainedwith PI and subjected to FACS analysis after 24 hr of culture. Mean and standard deviationsof triplicate samples are presented.(C) Thymocytes were cultured with indicated amounts of anti-Fas antibody. Cells werestained with PI and subjected to FACS analysis after 24 hr of culture. Mean and standarddeviations of triplicate samples are presented.
Lindsten et al. Page 19
Mol Cell. Author manuscript; available in PMC 2011 March 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Lindsten et al. Page 20
Table 1
Hematological Profile of bax-/-bak-/- and Control Mice
Genotype WBC Neutrophils Lymphocytes RBC
bax-/-bak-/-
1 15.3 4.1 10.0 7.4
2 24.2 2.5 20.4 4.6
3 28.7 2.0 26.0 9.5
4 39.2 5.1 31.9 8.8
bax+/-bak-/-
1 11.1 3.7 5.5 10.1
2 8.6 1.3 6.5 9.5
bax+/+bak-/-
1 3.7 1.2 2.4 9.8
2 4.6 0.7 3.6 9.8
bax+/+bak+/-
1 6.4 1.6 4.7 9.8
2 5.7 1.3 4.2 8.8
bax+/+bak+/+
1 4.7 0.7 3.8 9.9
2 4.8 0.5 4.0 9.3
3 4.3 1.3 2.9 10.3
The number of white blood cells (WBC), neutrophils, lymphocytes (× 103/μl), and red blood cells (RBC, × 106/μl) was determined using aHemavet 1500 apparatus.
Mol Cell. Author manuscript; available in PMC 2011 March 15.

NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Lindsten et al. Page 21
Table 2
Bone Marrow Hematopoietic Progenitor Assay
Genotype Myeloid Erythroid/Meg Total
bax-/-bak-/-
1 297 98 395
2 250 93 343
3 302 88 390
bax +/- bak -/- 208 111 319
bax +/+ bak -/- 154 104 258
bax+/+bak+/- 172 74 246
bax+/+bak+/+ 144 ± 12 68 ± 5 211 ± 7
Bone marrow cells were isolated from bax-/-bak-/- and control mice and cultured in complete methylcellulose medium for 5 days. CFU werescored based on colony morphology and cytological staining. Myeloid CFU were a combination of macrophage, granulocyte-erythrocyte-megakaryocyte-macrophage, granulocyte-macrophage, and granulocyte CFU. Likewise, the erythroid and megakaryocyte CFU were combined.For the wild-type group, data is expressed as the mean ± standard deviation.
Mol Cell. Author manuscript; available in PMC 2011 March 15.
Copyright © 2022 FDOKUMEN




















