
High-Throughput Fluorescence Polarization Assay for Chemical Library Screening against...
12
http://jbx.sagepub.com/ Journal of Biomolecular Screening http://jbx.sagepub.com/content/17/3/350 The online version of this article can be found at: DOI: 10.1177/1087057111429372 2012 17: 350 originally published online 7 December 2011 J Biomol Screen Ying Su, Thomas D. Y. Chung, Chaofang Jin, Paul Diaz and John C. Reed Dayong Zhai, Paulo Godoi, Eduard Sergienko, Russell Dahl, Xochella Chan, Brock Brown, Justin Rascon, Andrew Hurder, Anti-Apoptotic Bcl-2 Family Member Bfl-1 High-Throughput Fluorescence Polarization Assay for Chemical Library Screening against Published by: http://www.sagepublications.com On behalf of: Journal of Biomolecular Screening can be found at: Journal of Biomolecular Screening Additional services and information for http://jbx.sagepub.com/cgi/alerts Email Alerts: http://jbx.sagepub.com/subscriptions Subscriptions: http://www.sagepub.com/journalsReprints.nav Reprints: http://www.sagepub.com/journalsPermissions.nav Permissions: What is This? - Dec 7, 2011 OnlineFirst Version of Record - Feb 9, 2012 Version of Record >> by guest on October 25, 2013 jbx.sagepub.com Downloaded from by guest on October 25, 2013 jbx.sagepub.com Downloaded from by guest on October 25, 2013 jbx.sagepub.com Downloaded from by guest on October 25, 2013 jbx.sagepub.com Downloaded from by guest on October 25, 2013 jbx.sagepub.com Downloaded from by guest on October 25, 2013 jbx.sagepub.com Downloaded from by guest on October 25, 2013 jbx.sagepub.com Downloaded from by guest on October 25, 2013 jbx.sagepub.com Downloaded from by guest on October 25, 2013 jbx.sagepub.com Downloaded from by guest on October 25, 2013 jbx.sagepub.com Downloaded from by guest on October 25, 2013 jbx.sagepub.com Downloaded from by guest on October 25, 2013 jbx.sagepub.com Downloaded from
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of High-Throughput Fluorescence Polarization Assay for Chemical Library Screening against...

http://jbx.sagepub.com/Journal of Biomolecular Screening
http://jbx.sagepub.com/content/17/3/350The online version of this article can be found at:
DOI: 10.1177/1087057111429372
2012 17: 350 originally published online 7 December 2011J Biomol ScreenYing Su, Thomas D. Y. Chung, Chaofang Jin, Paul Diaz and John C. Reed
Dayong Zhai, Paulo Godoi, Eduard Sergienko, Russell Dahl, Xochella Chan, Brock Brown, Justin Rascon, Andrew Hurder,Anti-Apoptotic Bcl-2 Family Member Bfl-1
High-Throughput Fluorescence Polarization Assay for Chemical Library Screening against
Published by:
http://www.sagepublications.com
On behalf of:
Journal of Biomolecular Screening
can be found at:Journal of Biomolecular ScreeningAdditional services and information for
http://jbx.sagepub.com/cgi/alertsEmail Alerts:
http://jbx.sagepub.com/subscriptionsSubscriptions:
http://www.sagepub.com/journalsReprints.navReprints:
http://www.sagepub.com/journalsPermissions.navPermissions:
What is This?
- Dec 7, 2011OnlineFirst Version of Record
- Feb 9, 2012Version of Record >>
by guest on October 25, 2013jbx.sagepub.comDownloaded from by guest on October 25, 2013jbx.sagepub.comDownloaded from by guest on October 25, 2013jbx.sagepub.comDownloaded from by guest on October 25, 2013jbx.sagepub.comDownloaded from by guest on October 25, 2013jbx.sagepub.comDownloaded from by guest on October 25, 2013jbx.sagepub.comDownloaded from by guest on October 25, 2013jbx.sagepub.comDownloaded from by guest on October 25, 2013jbx.sagepub.comDownloaded from by guest on October 25, 2013jbx.sagepub.comDownloaded from by guest on October 25, 2013jbx.sagepub.comDownloaded from by guest on October 25, 2013jbx.sagepub.comDownloaded from by guest on October 25, 2013jbx.sagepub.comDownloaded from

Journal of Biomolecular Screening17(3) 350 –360© 2012 Society for Laboratory Automation and ScreeningDOI: 10.1177/1087057111429372http://jbx.sagepub.com
Introduction
The mitochondrial pathway for cell death is governed by members of the Bcl-2 family proteins. Several homologs of Bcl-2 have been identified, with some functioning as block-ers and others as promoters of cell death, constituting a family of more than 27 genes in humans.1–3 Anti-apoptotic Bcl-2 family proteins (Bcl-2, Bcl-X
L, Bcl-W, Bcl-B, Mcl-1,
and Bfl-1) interact with BH3-only pro-apoptotic members (e.g., Bid, Bad, Bim, Puma, Noxa, Hrk, Bmf, Nbk/Bik) to regulate pro-apoptotic multidomain Bcl-2 family proteins Bax and Bak, which trigger mitochondrial outer mem-brane permeability and release of cytochrome c and other pro-apoptotic factors that promote activation of caspases, causing apoptosis.1,3,4
The human genome contains six genes that encode anti-apoptotic members of the Bcl-2 family.2 Among these is Bfl-1 (also known as A1 in mice), a nuclear factor–κB (NF-κB)–inducible member of the Bcl-2 family.5–9 Bfl-1
promotes the survival of immune and inflammatory cells, including mast cells associated with allergy.10 Given that transgenic overexpression of various anti-apoptotic Bcl-2 family proteins in lymphocytes causes autoimmunity,11,12 Bfl-1 may represent a target for certain autoimmune and allergic diseases. In addition, Bfl-1 is highly upregulated in therapy-resistant B-cell chronic lymphocytic leukemia
429372 JBXXXX10.1177/1087057111429372Zhai et al.Journal of Biomolecular Screening
1 Sanford-Burnham Medical Research Institute, Program on Apoptosis and Cell Death Research, La Jolla, CA, USA
2 Sanford-Burnham Medical Research Institute, Conrad Prebys Center for Chemical Genomics, La Jolla, CA, USA
Received Apr 25, 2011, and in revised form Aug 4, 2011. Accepted for publication Oct 4, 2011.
Corresponding Author:John C. Reed, Sanford-Burnham Medical Research Institute, 10901 North Torrey Pines Road, La Jolla, CA 92037, USA Email: [email protected]
High-Throughput Fluorescence Polarization Assay for Chemical Library Screening against Anti- Apoptotic Bcl-2 Family Member Bfl-1
Dayong Zhai1, Paulo Godoi1, Eduard Sergienko2, Russell Dahl2, Xochella Chan2, Brock Brown2, Justin Rascon2, Andrew Hurder2, Ying Su2, Thomas D. Y. Chung2, Chaofang Jin1, Paul Diaz1, and John C. Reed1, 2
Abstract
Overexpression of the anti-apoptotic Bcl-2 family proteins occurs commonly in human cancers. Bfl-1 is highly expressed in some types of malignant cells, contributing significantly to tumor cell survival and chemoresistance. Therefore, it would be desirable to have chemical antagonists of Bfl-1. To this end, we devised a fluorescence polarization assay (FPA) using Bfl-1 protein and fluorescein-conjugated Bid BH3 peptide, which was employed for high-throughput screening of chemical libraries. Approximately 66 000 compounds were screened for the ability to inhibit BH3 peptide binding to Bfl-1, yielding 14 reproducible hits with ≥50% displacement. After dose-response analysis and confirmation using a secondary assay based on time-resolved fluorescence resonance energy transfer (TR-FRET), two groups of Bfl-1-specific inhibitors were identified, including chloromaleimide and sulfonylpyrimidine series compounds. FPAs generated for each of the six anti-apoptotic Bcl-2 proteins demonstrated selective binding of both classes of compounds to Bfl-1. Analogs of the sulfonylpyrimidine series were synthesized and compared with the original hit for Bfl-1 binding by both FPAs and TR-FRET assays. The resulting structure-activity relation analysis led to the chemical probe compound CID-2980973 (ML042). Collectively, these findings demonstrate the feasibility of using the HTS assay for discovery of selective chemical inhibitors of Bfl-1.
Keywords
cancer and cancer drugs, fluorescence polarization methods, immune system diseases, inflammatory diseases, protein–protein interactions

Zhai et al. 351
(B-CLL), acute myelogenous leukemia with poor progno-sis, large B-cell lymphomas, and some other cancers.7,8,13 Bfl-1 appears to play a critical role in tumor cell survival and chemoresistance, as demonstrated by the observation that Bfl-1 knockdown by short-hairpin RNA (shRNA) can restore apoptosis of drug-resistant leukemia and lymphoma cells in culture,14–16 thus suggesting the importance of Bfl-1 as a therapeutic target for cancer. Bfl-1 has also been shown to be responsible for resistance of malignant cells to chemi-cal inhibitors that neutralize other members of the Bcl-2 family, implying that inhibition of Bcl-2 and Bcl-X
L can be
insufficient to achieve apoptosis when Bfl-1 is highly expressed.14,16–18
Synthetic BH3 peptides bind anti-apoptotic Bcl-2 family proteins with nanomolar affinities, promoting apoptosis.19,20 Nonpeptidyl compounds have been identified that compete with BH3 peptides for binding to anti-apoptotic Bcl-2 fam-ily proteins, mimicking BH3 peptides and creating interest in development of these molecules as potential cancer ther-apeutics.21,22 Of note, systematic analysis of all chemical inhibitors of Bcl-2 thus far described in the literature reveals that few of the synthetic compounds or natural products previously reported bind Bfl-1 with a biologically relevant affinity,21 and none binds selectively to Bfl-1. Thus, the pocket on Bfl-1 that binds BH3 peptides is presumably suf-ficiently different from other Bcl-2 family members that it should be possible to obtain selective inhibitors, either directly from screens of diverse libraries or secondarily through chemical analoging of hits that interact with Bfl-1.
To this end, we have devised a homogeneous fluores-cence polarization assay (FPA) based on binding of recom-binant soluble Bfl-1 protein to synthetic Bid BH3 peptide conjugated with a fluorochrome. In this assay, the rotation of fluorochrome-conjugated peptide ligand is slowed upon binding the Bfl-1 target protein, thus affecting the interac-tion of the fluorochrome with polarized light. High-throughput conditions were optimized, and approximately 66 000 chemical compounds were screened as proof of con-cept. The primary hits were further confirmed using a time-resolved fluorescence resonance energy transfer (TR-FRET) assay. We succeeded in obtaining two groups of Bfl-1-specific inhibitors, including a chloromaleimide series that has been described previously23 and a sulfonylpyrimidine series described here. The active sulfonylpyrimidines were demonstrated to bind selectively to Bfl-1 among the six anti-apoptotic Bcl-2 family proteins. Despite the general challenges of identifying inhibitors of protein–protein inter-actions by high-throughput screening (HTS) and the length of BH3 peptides (~16 amino acids), these findings demon-strate the feasibility of using the FPA reported here for identification of compounds that interact competitively with the BH3 binding site on Bfl-1, which may serve as a
starting point for optimization and eventual generation of drug candidates.
Materials and MethodsProtein Purification
His6-“truncated” Bid (t-Bid) was prepared as previously
reported.24 Glutathione S-transferase (GST)–tagged pro-teins containing Bcl-X
L, Bcl-2, Bcl-W, Bcl-B, Bfl-1, and
Mcl-1 lacking their C-terminal transmembrane domains (~ last 20 amino acids) (“ΔTM”) were expressed from the pGEX 4T-1 plasmid in XL-1 Blue cells (Stratagene, La Jolla, CA) as previously reported.25 Briefly, cells were grown in 2 L of LB with 50 µg/mL ampicillin at 37 °C to an optical density (OD) 600 nm of 1.0, then isopropyl-β-D-thiogalactopyranoside (IPTG; 0.5 mM) was added, and the cultures were incubated at 25 °C for 6 h. Cells were then recovered in 20 mM phosphate buffer (pH 7.4), 150 mM NaCl, 1 mM dithiothreitol (DTT), 1 mM EDTA, and 1 mM phenylmethylsulfonyl fluoride (PMSF), followed by soni-cation. Cellular debris was sedimented by centrifugation at 25 000 g for 20 min, and the resulting supernatants were incubated with 10 mL of glutathionine-Sepharose (Amersham Biosciences, Piscataway, NJ) at 4 °C for 2 h. The resin was washed three times with 20 mM phosphate buffer (pH 7.4), 150 mM NaCl, and 1 mM DTT, and then 10 mM of reduced glutathione dissolved in 50 mM Tris-HCl (pH 8.0) was used to elute the GST fusion proteins.
Fluorescence Polarization AssaysFPAs were performed as described previously using vari-ous Bcl-2 family proteins and fluorescein isothiocyanate (FITC)–conjugated Bid BH3 peptide.25 Briefly, Bfl-1 pro-teins were incubated in 384-well black round-bottom plates with 5 or 2.2 nM FITC–Bid BH3 peptide (FITC-Ahx-EDIIRNIARHLAQVGDSMDR) in a total volume of 20 µL in the dark. Fluorescence polarization was measured using a LJL Analyst HT plate reader (Molecular Devices Corp., Sunnyvale, CA) in phosphate-buffered saline (PBS) or 25 mM Bis-Tris buffer containing 1 mM of TECP, 0.005% Tween-20 (pH 7.0). IC
50 determinations were performed using a sig-
moidal dose–response nonlinear regression model (Y = Bottom + (Top – Bottom)/1 + 10(LogEC50 – X)*Hill slope) with GraphPad Prism software (GraphPad, Inc., San Diego, CA).
High-Throughput ScreeningApproximately 66 000 compounds (10 mM stock solution in DMSO) from the National Institutes of Health (NIH) Small Molecule library were plated into black, 384-well,

352 Journal of Biomolecular Screening 17(3)
round-bottom plates (Greiner Bio-One, Frickenhausen, Germany) using a BioMek FX Laboratory Automation Workstation (Beckman Coulter, Fullerton, CA). The ThermoScientific Matrix WellMate bulk dispenser (Thermo Fisher Scientific, Hudson, NH) was then used to add GST–Bfl-1, followed by FITC–Bid BH3 peptide, both diluted in 25 mM Bis-Tris, 1 mM TCEP, 0.005% Tween-20 (pH 7.0). The first column of the screening plate was posi-tive controls (16 wells), and the second column of the plate was negative controls (16 wells). The remaining wells con-tained ~20 µM compound, 3 nM GST–Bfl-1, and 2.2 nM FITC–Bid BH3 peptide in a total volume of 20 µL. After a 4-h incubation at room temperature in the dark, fluores-cence polarization was measured using an Analyst HT Multi-Mode Plate Reader (Molecular Devices), with an excitation filter at 485 nM and an emission filter at 530 nM. Data analysis was performed using CBIS software (ChemInnovation Software Inc, San Diego, CA).
Competitive Peptide Displacement Assays for Assessing Compound SelectivityMethods for competitive peptide displacement assays were similar to previous publications.25 Briefly, empirically opti-mized amounts of various GST fusion proteins (3 nM for Bfl-1; 20 nM for Bcl-2, Bcl-X
L, Bcl-W, and Bcl-B; and 10 nM
for Mcl-1) were incubated with the compounds at various concentrations for 5 min at room temperature in 25 mM Bis-Tris, 1 mM TCEP, 0.005% Tween-20 (pH 7.0). Then, 2.2 nM FITC-Bid (for Bfl-1), 10 nM FITC-Bim (for Bcl-2, Bcl-X
L, Bcl-W, and Bcl-B), or 5 nM FITC-Bak (for Mcl-1)
BH3 peptide was added, resulting in a total volume of 20 µL per well. DMSO concentration (resulting from com-pound addition) did not exceed 0.2% (vol:vol). Fluorescence polarization was measured after 20 min. IC
50 determina-
tions were generated by fitting the experimental data using a sigmoidal dose–response nonlinear regression model with GraphPad Prism software (GraphPad, Inc.).
Time-Resolved Fluorescence Resonance Energy Transfer AssaysFor TR-FRET assays, GST–Bfl-1 and anti-GST-terbium (Invitrogen, Carlsbad, CA) were mixed together with the FITC–Bid BH3 peptide in 20 mM Bis-Tris containing 0.005% Tween-20 in 384-well plates in a total volume of 20 µL per well. After incubation at room temperature for 10 min, 2 µL of compound-containing solutions was added to the reaction mixtures containing 3 nM Bfl-1, 2.2 nM FITC–Bid BH3 peptide, and 2 nM anti-GST-terbium for 30 min at room temperature. TR-FRET signals were mea-sured with a SpectraMax M5 plate reader (Molecular Devices) using the following settings: excitation at 330 nm,
emission for FITC signal at 490 nm, and emission for ter-bium signal at 520 nm. IC
50 determinations were generated
by fitting the experimental data using a sigmoidal dose–response nonlinear regression model with GraphPad Prism software (GraphPad, Inc.).
Mitochondria Purification and Protein Release AssaysHeLa cells were pelleted by centrifugation and then washed once in HM buffer (10 mM HEPES [pH 7.4], 250 mM mannitol, 10 mM KCl, 5 mM MgCl
2, 1 mM EGTA), con-
taining 1 mM PMSF and a mixture of protease inhibitors (Roche Molecular Biochemicals, Indianapolis, IN). The cell pellet was then homogenized in HM buffer by 60 strokes of a dounce homogenizer, using a B-type pestle. The homogenate was centrifuged twice at 600 g for 5 min to remove nuclei and debris. The resulting supernatant was centrifuged at 10 000 g for 10 min, and the resulting mito-chondria-containing pellet was washed twice with HM buffer.
For mitochondrial protein release assays, 10 µL of mitochondria (50 µg) was added to a final volume of 50 µL HM buffer containing compounds, with or without t-Bid or Bcl-2 family proteins, at 30 °C for 15 min. The reactions were further incubated at 30 °C for 40 to 60 min, and then mitochon-dria were pelleted by centrifugation and the supernatants were collected, boiled in Laemmli sample buffer, and analyzed by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE)/immunoblotting using anti-SMAC antibody.26
Compound SynthesisCID 1151859 (MLS-0009480). A mixture of 4,4,4-triflu-
oro-1-phenylbutane-1,3-dione (216 mg, 1.0 mmol) and ethyl 2-(carbamimidoylthio)acetate (194 mg, 1.2 mmol) in ethanol (10 mL) was placed in a sealed tube and heated at 100 °C for 5 h. The resulting solution was cooled and reduced to dryness via rotary evaporation, and the residue was purified via silica gel chromatography (EtOAc:hexanes) to yield the pyrimidine intermediate, ethyl 2-((4-phenyl-6-(trifluoromethyl)pyrimidin-2-yl)thio)acetate. To this product (0.17 mg, 0.5 mmol) was added mCPBA (230 mg, 1.0 mmol, 77% by weight) in 5 mL of dichloromethane (DCM), and the resulting mixture was stirred at room tem-perature for 20 h. The mixture was diluted with DCM and extracted with water and brine. The organic layer was col-lected and the excess DCM was removed by rotary evapo-ration. The product was isolated by reverse-phase preparative high-performance liquid chromatography (HPLC; H
2O:ACN). 1H NMR (400 MHz, CDCl
3): δ 8.22
(m, 2H), 8.18 (s, 1H), 7.64 (m, 1H), 7.57 (m, 2H), 4.66 (s, 2H), 4.15 (q, J = 17 Hz, 2H), 1.15 (t, J = 17 Hz, 3H). ESI-MS m/z 375 [M + H]+.
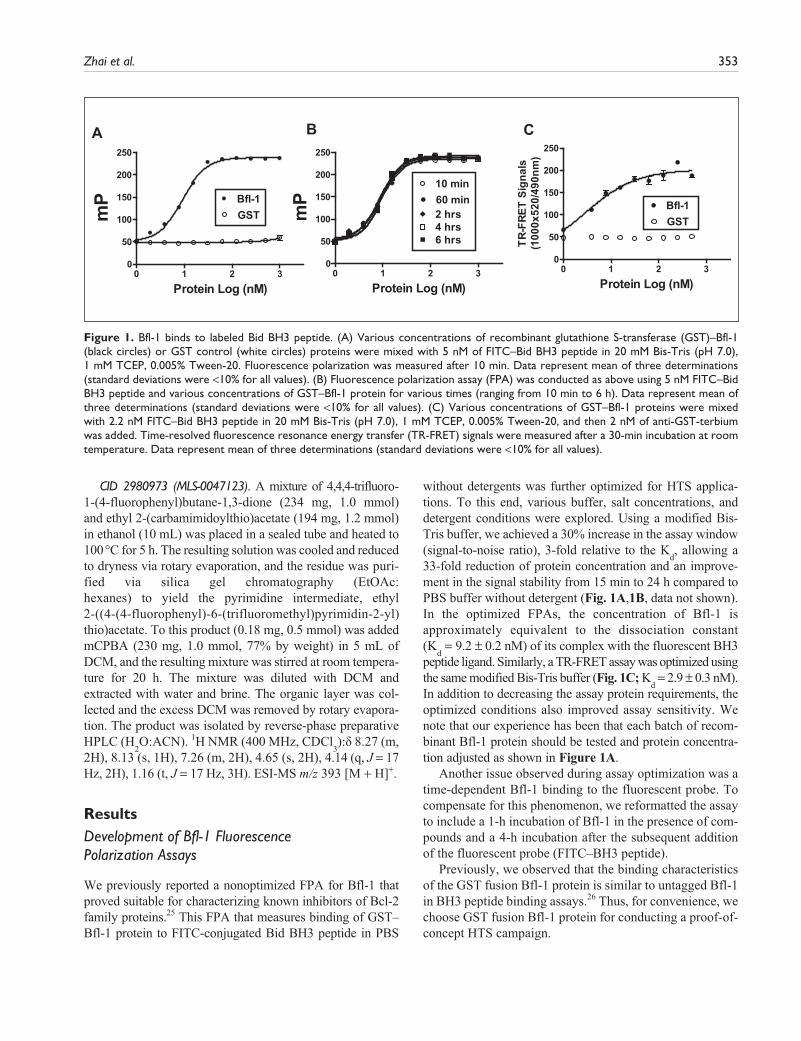
Zhai et al. 353
CID 2980973 (MLS-0047123). A mixture of 4,4,4-trifluoro-1-(4-fluorophenyl)butane-1,3-dione (234 mg, 1.0 mmol) and ethyl 2-(carbamimidoylthio)acetate (194 mg, 1.2 mmol) in ethanol (10 mL) was placed in a sealed tube and heated to 100 °C for 5 h. The resulting solution was cooled and reduced to dryness via rotary evaporation, and the residue was puri-fied via silica gel chromatography (EtOAc: hexanes) to yield the pyrimidine intermediate, ethyl 2-((4-(4-fluorophenyl)-6-(trifluoromethyl)pyrimidin-2-yl)thio)acetate. To this product (0.18 mg, 0.5 mmol) was added mCPBA (230 mg, 1.0 mmol, 77% by weight) in 5 mL of DCM, and the resulting mixture was stirred at room tempera-ture for 20 h. The mixture was diluted with DCM and extracted with water and brine. The organic layer was col-lected and the excess DCM was removed by rotary evapora-tion. The product was isolated by reverse-phase preparative HPLC (H
2O:ACN). 1H NMR (400 MHz, CDCl
3):δ 8.27 (m,
2H), 8.13 (s, 1H), 7.26 (m, 2H), 4.65 (s, 2H), 4.14 (q, J = 17 Hz, 2H), 1.16 (t, J = 17 Hz, 3H). ESI-MS m/z 393 [M + H]+.
ResultsDevelopment of Bfl-1 Fluorescence Polarization Assays
We previously reported a nonoptimized FPA for Bfl-1 that proved suitable for characterizing known inhibitors of Bcl-2 family proteins.25 This FPA that measures binding of GST–Bfl-1 protein to FITC-conjugated Bid BH3 peptide in PBS
without detergents was further optimized for HTS applica-tions. To this end, various buffer, salt concentrations, and detergent conditions were explored. Using a modified Bis-Tris buffer, we achieved a 30% increase in the assay window (signal-to-noise ratio), 3-fold relative to the K
d, allowing a
33-fold reduction of protein concentration and an improve-ment in the signal stability from 15 min to 24 h compared to PBS buffer without detergent (Fig. 1A,1B, data not shown). In the optimized FPAs, the concentration of Bfl-1 is approximately equivalent to the dissociation constant (K
d = 9.2 ± 0.2 nM) of its complex with the fluorescent BH3
peptide ligand. Similarly, a TR-FRET assay was optimized using the same modified Bis-Tris buffer (Fig. 1C; K
d = 2.9 ± 0.3 nM).
In addition to decreasing the assay protein requirements, the optimized conditions also improved assay sensitivity. We note that our experience has been that each batch of recom-binant Bfl-1 protein should be tested and protein concentra-tion adjusted as shown in Figure 1A.
Another issue observed during assay optimization was a time-dependent Bfl-1 binding to the fluorescent probe. To compensate for this phenomenon, we reformatted the assay to include a 1-h incubation of Bfl-1 in the presence of com-pounds and a 4-h incubation after the subsequent addition of the fluorescent probe (FITC–BH3 peptide).
Previously, we observed that the binding characteristics of the GST fusion Bfl-1 protein is similar to untagged Bfl-1 in BH3 peptide binding assays.26 Thus, for convenience, we choose GST fusion Bfl-1 protein for conducting a proof-of-concept HTS campaign.
A B
0 1 2 30
50
100
150
200
250
GSTBfl-1
Protein Log (nM)
mP
0 1 2 30
50
100
150
200
250
60 min10 min
2 hrs4 hrs6 hrs
Protein Log (nM)m
P
0 1 2 30
50
100
150
200
250
GSTBfl-1
Protein Log (nM)
TR-F
RET
Sign
als
(100
0x52
0/49
0nm
)
C
Figure 1. Bfl-1 binds to labeled Bid BH3 peptide. (A) Various concentrations of recombinant glutathione S-transferase (GST)–Bfl-1 (black circles) or GST control (white circles) proteins were mixed with 5 nM of FITC–Bid BH3 peptide in 20 mM Bis-Tris (pH 7.0), 1 mM TCEP, 0.005% Tween-20. Fluorescence polarization was measured after 10 min. Data represent mean of three determinations (standard deviations were <10% for all values). (B) Fluorescence polarization assay (FPA) was conducted as above using 5 nM FITC–Bid BH3 peptide and various concentrations of GST–Bfl-1 protein for various times (ranging from 10 min to 6 h). Data represent mean of three determinations (standard deviations were <10% for all values). (C) Various concentrations of GST–Bfl-1 proteins were mixed with 2.2 nM FITC–Bid BH3 peptide in 20 mM Bis-Tris (pH 7.0), 1 mM TCEP, 0.005% Tween-20, and then 2 nM of anti-GST-terbium was added. Time-resolved fluorescence resonance energy transfer (TR-FRET) signals were measured after a 30-min incubation at room temperature. Data represent mean of three determinations (standard deviations were <10% for all values).

354 Journal of Biomolecular Screening 17(3)
High-Throughput Screening
The performance of the optimized assay was evaluated by performing multiple replicate tests in a 384-well format using either FITC–Bid BH3 peptide alone (assay mini-mum) or with Bfl-1 protein (assay maximum). The assay demonstrated robust performance, with a Z′ factor of 0.83 (Fig. 2A). Moreover, the Z′ factor remained >0.7 after overnight incubation (data not shown).
The assay was screened against the entire NIH small-mol-ecule library, which at the time consisted of ~66,000 com-pounds, searching for chemical inhibitors that displaced FITC–Bid BH3 peptide from GST–Bfl-1 protein, resulting in 44 primary positive results, with an average Z′ factor of ~0.75 ± 0.04 (Fig. 2B). “Hits” were defined as compounds that induced a ≥50% decrease in fluorescence polarization without increasing fluorescence intensity by ≥25% (thus eliminating compounds that interfered with the assay due to their autofluorescence). After correcting for fluorescence interferences, the primary screen then yielded 20 hits. These hits were cherry picked from assay plates and retested, result-ing in 14 confirmed hits, representing a hit rate of 0.021%.
A TR-FRET secondary assay used the same molecu-lar interaction between the FITC–Bid BH3 peptide and the GST–Bfl-1 protein, except terbium-conjugated anti-GST served as the fluorescent donor for exciting the FITC BH3 peptide. Of 20 compounds active in the pri-mary FPA-based screen, 14 demonstrated inhibitory activity in the TR-FRET assay. These were the same 14 com-pounds that showed reproducible activity in the FPA (above).
Compound dose–response curves were generated using both fluorescence polarization (FP) and TR-FRET assays for the 14 validated hits. Compounds that did not display a sigmoidal dose–response curve or that increased fluores-cence intensity (some compounds increased fluorescence intensity at concentrations higher than the screening con-centration) were eliminated. Finally, seven compounds appeared to be promising based on dose–response results from both the FP and TR-FRET assays (IC
50 values sum-
marized in Table 1). The potency of these screening hits varied, with IC
50 values as low as <1 µM to as high as
approximately 30 µM, depending on whether measured by FPA or TR-FRET.
Z’ factor=0.83
A BResult Distribution
FPA (% Inhibition)
1600
1400
1200
1000
800
600
400
200C
ount
0
-30 -15 0 15 30 45 60 75 90 105
0 50 100 150 2000
50
100
150
200
250
Bfl-1+FITC-BidFITC-Bid+Bid BH3 peptide
Well #
mP
Bfl-1+FITC-Bid+Bid peptideHits
Figure 2. Compound library screening. (A) Glutathione S-transferase (GST)–Bfl-1 (10 nM) was incubated with 5 nM FITC–Bid BH3 peptide without (black squares) or with 50 nM of unlabeled Bid BH3 peptide (black circles). Alternatively, FITC–Bid peptide was added to wells without Bfl-1 protein (first column, 16 wells, white circles). Fluorescence polarization was measured after 10 min. Data represent fluorescence polarization (mP) measured in individual wells, constituting 176 replicates for each condition in a 384-well format. The Z′ score was calculated. (B) For chemical library screening, GST–Bfl-1 (3 nM) was incubated with FITC–Bid BH3 peptide (2.2 nM) with DMSO alone or with 20 µM of the National Institutes of Health (NIH) library compounds (blue). Fluorescence polarization was measured as previously described. Blue bars represent fluorescence polarization assay (FPA) results for compound library screening, indicating the number of compounds (y-axis) that displayed a specific percentage inhibition (x-axis). Red bars indicate control wells included in all plates, which comprised either FITC–BH3 Bid peptide alone (right) or FITC–Bid BH3 peptide plus Bfl-1 protein (left). The box shows the result for “hits,” where ≥50% inhibition of FPA signal is considered positive.

Zhai et al. 355
Characterization of Compound Selectivity for Bfl-1
The seven validated hits from the screening could be divided into three groups based on structural features of the compounds: #1 (MLS-0053105 = CID-1180676) is a chlo-romaleimide, #2 to #5 (#2, MLS-0047123 = CID-2980973; #3, MLS-0009480 = CID-1151859; #4, MLS-0051609 = CID-3244140; #5, MLS-0051509 = CID-3237441) com-prise a sulfonylpyrimidine series; and #6 and #7 MLS-0019296 (CID-5389865) and MLS-0025736 (CID-8423532) are quinolinyl-oxadiazoles. The reactivity of these com-pounds with other targets previously screened and depos-ited into the PubChem database at the time of our discovery revealed that the chloromaleimide showed reactivity in only 1 of 30 assays tested, the sulfonylpyrimidines in 3 of 60 assays, and the quinolinyl-oxadiazoles in none of 29 assays tested. Thus, these three chemical series do not show promiscuous activity with other classes of targets.
To test compound selectivity against all six human anti-apoptotic Bcl-2 family protein members, we established FPAs that monitor the binding of various fluorochrome-conjugated BH3 peptides to the recombinant GST fusion proteins. BH3 peptides were selected based on our prior comparison of binding affinities,27 resulting in pairings for the binding assays as follows: Bfl-1/FITC–Bid BH3 pep-tide, Bcl-W/FITC–Bim BH3 peptide, Bcl-2/FITC–Bim BH3 peptide, Bcl-X
L/FITC–Bim BH3 peptide, and Mcl-1/
FITC–Bak BH3 peptide. Table 2 summarizes the IC50
val-ues of seven validated compounds for each of the six anti-apoptotic Bcl-2 family members. The chloromaleimide compounds showed submicromolar inhibition of Bfl-1,
with over 10 times less potent inhibition of Bcl-W, Bcl-2, and Bcl-X
L and no activity against Bcl-B and Mcl-1, indi-
cating good selectivity for Bfl-1. The sulfonylpyrimidine compounds showed superb selectivity against Bfl-1, with no inhibition of other Bcl-2 family members at concentra-tions as high as 100 µM. One of the quinolinyl-oxadiazole series compounds showed cross-reactivity with Bcl-W, but otherwise this series was also Bfl-1 selective (Table 2).
Bioactivity of Compounds Tested Using Mitochondrial AssayIsolated mitochondria were used to test the bioactivity of compounds under circumstances where recombinant Bfl-1 protein nullifies mitochondrial release of proteins induced by t-Bid protein, similar to our previous report where we analyzed BH3 peptide activity.26 Interestingly, sulfonyl-pyrimidine CID-1151859 caused mitochondria release of SMAC protein in a concentration-dependent manner (Fig. 3A), perhaps reflecting activity against endogenous proteins associated with the mitochondria. Recombinant Bfl-1 protein, but not Bcl-2 or Bcl-X
L, blocked the release
of SMAC protein induced by this compound, consistent with our data indicating selective binding to Bfl-1 (Fig. 3B,3C). In contrast, chloromaleimide compound CID-1180676 did not cause SMAC release from mitochondria (Fig. 3D). We then employed recombinant Bid (t-Bid) protein as an inducer of SMAC release from mitochondria and tested the effects of compounds with respect to neutralization by recombinant anti-apoptotic Bcl-2 proteins Bfl-1 and Bcl-X
L.
The chloromaleimide compound CID-1180676 negated Bfl-1 protection against t-Bid-induced release of SMAC
Table 1. IC50
Values of Bfl-1 Inhibiting Compounds
Compound # PubChem CID # Structure Bfl-1 FPA IC50
(µM) Bfl-1 TR-FRET IC50
(µM)
1 = MLS-0053105 1180676 0.4 0.6
2 = MLS-0047123 2980973 3.3 3.6
3 = MLS-0009480 1151859 2.4 7.0
4 = MLS-0051609 3244140 20.7 7.2
5 = MLS-0051509 3237441 5.9 7.1
6 = MLS-0019296 5389865 29.4 19.57 = MLS-0025736 5423532 6.1 4.5
For fluorescence polarization assays (FPAs), various concentrations of compounds (MLS and CID numbers provided, along with structures) were incubated with glutathione S-transferase (GST)–Bfl-1 (3 nM) in 20 mM Bis-Tris (pH 7.0), 1 mM TCEP, 0.005% Tween-20. Then, 2.2 nM of FITC–Bid BH3 peptide was introduced, and fluorescence polarization was measured after 30 min. IC
50 determinations were generated by fitting the experimental data
using a sigmoidal dose–response nonlinear regression model with GraphPad Prism software. For time-resolved fluorescence resonance energy transfer (TR-FRET), various concentrations of compounds were added to reaction mixtures containing 3 nM of Bfl-1, 2.2 nM of FITC–Bid BH3 peptide, and 2 nM of anti-GST-terbium for 30 min at room temperature (mean ± SEM: n = 3). TR-FRET signals were measured with a SpectraMax M5 plate reader, and IC
50s were generated similar to FPA data (mean ± SEM: n = 3).
N
N
ClO
N
O
Cl
Cl N
N
ClO
N
O
Cl
Cl
O
O
SO
O
N
NF
FF
F
O
O
SO
O
N
NF
FF
F
O
O
SO
O
N
NF
FF
O
O
SO
O
N
NF
FF
O
O
N
N
SO
O
FF
F
O
O
N
N
SO
O
FF
F
O
O
H 2N
NNS
OO
F
F
F
F
F
O
O
H 2N
NNS
OO
F
F
F
F
F
O
N OH
NO
NO
N OH
NO
N
O
N OH
N O
N FO
N OH
N O
N F

356 Journal of Biomolecular Screening 17(3)
CID-1151859 0 1 2 5 10 20 50 100 P
CID-1151859 0 20 20 20 20 20 20
Bfl-1 (µM) 0 0 1 2 5 10 20 P
A
B
CID-1151859 0 20 20 20 20 20 20 20 20
Bcl-2 (µM) 0 0 1 5 20
Bcl-XL (µM) 0 1 5 20 P
C
D
E
CID-1180676 0 1 2 5 10 20 50 P
tBid 0 20 20 20 20 20 20 20 20 20
Bfl-1 (µM) 0 0 1 1 1 1 0 0 0 0
Bcl-XL (µM) 0 0 0 0 0 0 1 1 1 1
CID-1180676 0 0 0 1 5 10 0 1 5 10
Figure 3. Analysis of Bfl-1-binding compounds using a mitochondria-based functional assay. Various concentrations of compound CID-1151859 or CID-1180676 (µM) were preincubated with specific concentrations of purified recombinant t-Bid (20 nM in A–D) (0 vs 20 µM in E), without (A, D) or with (B, C, E) various concentrations of anti-apoptotic Bcl-2 family proteins, including Bfl-1 at 0-20 µM (B), Bcl-X
L at 0-20 µM (C), Bcl-2 at 0-20 µM (C), or 0-1 µM Bfl-1 or 0-1 µM Bcl-X
L (E) in HM buffer, before adding 50 µg of
isolated HeLa mitochondria for 1 h at 30 °C. Samples were centrifuged to generate supernatants that were analyzed by sodium dodecyl sulfate polyacrylamide gel electrophoresis/immunoblotting using the anti-SMAC antibody. The “Pellet” (P) represents intact, untreated mitochondria, run in gels as a control. Data are representative of at least three experiments.
Table 2. Selectivity of Bfl-1 Compounds against Six Human Anti-Apoptotic Bcl-2 Family Proteins
Compound #Bfl-1/F-Bid IC
50 (µM)
Bcl-W/F-Bim IC
50 (µM)
Bcl-2/F-Bim IC
50 (µM)
Bcl-XL/F-Bim
IC50
(µM)Bcl-B/F-Bim IC
50 (µM)
Mcl-1/F-Bak IC
50 (µM)
1 = MLS-0053105 0.4 12.5 12.5 50 >100 >1002 = MLS-0047123 1.5 >100 >100 >100 >100 >1003 = MLS-0009480 2.0 >100 >100 >100 >100 >1004 = MLS-0051609 16.7 >100 >100 >100 >100 >1005 = MLS-0051509 4.0 >100 >100 >100 >100 >1006 = MLS-0019296 23.9 12.5 >100 >100 >100 >1007 = MLS-0025736 4.7 >100 >100 >100 >100 >100
Various concentrations of compounds were mixed with anti-apoptotic Bcl-2 family proteins (Bfl-1, Bcl-W, Bcl-2, Bcl-XL, Bcl-B, Mcl-1). FITC-BH3 pep-tides (Bid, Bim, or Bak as indicated) were then added and fluorescence polarization was measured after 30 min. IC
50 determinations were generated by
fitting the experimental data using a sigmoidal dose–response. Data are the mean of four determinations (standard derivations ≤15%).
from isolated mitochondria, whereas Bcl-XL did not, sug-
gesting selectivity for Bfl-1 (Fig. 3E).
Structure–Activity Relation Analysis of Chemical Inhibitors of Bfl-1During the course of this project, more than 180 analogs of the chloromaleimide and sulfonylpyrimidine class com-pounds were purchased or synthesized, comparing their activity against Bfl-1 using both the FPA and a TR-FRET assay. The structure–activity relation (SAR) analysis of the chloromaleimide series was published recently.23 Here we
describe a brief SAR analysis of the sulfonylpyrimidines (n = 11 compounds tested). Table 3 summarizes the IC
50
values of the compounds. Altogether, the activity of the compounds tested against Bfl-1 varied by as much as approximately one log, ranging from 2.6 µM to 25.1 µM in IC
50 values.
The synthetic chemistry strategy employed to follow up on hit compound CID-1151859 is shown in scheme 1 (Fig. 4). Specifically, 1,3-diketones were reacted with S-substituted carbamimidothioates to afford the corre-sponding S-alkylpyrimidines. Oxidation of these products at 25 °C with mCPBA furnishes the desired sulfonylpyrimidine

Zhai et al. 357
Table 3. Structure–Activity Relation of Sulfonylpyrimidine Analogs
Compound competition with BH3 peptide binding to Bfl-1 was measured as in Table 1. Data from fluorescence polarization assay (FPA) and time-resolved fluorescence resonance energy transfer (TR-FRET) are the mean of three determinations (with standard deviation <15%).
Bfl-1 FP Bfl-1 TR-FRET
Compound # Structure IC50 (uM) IC50 (uM)
CID-1151859 HTS Hit 4.6 2.9
CID-2980973 Probe 2.6 1.8
MLS-0066988 6.8 2.2
MLS-0046458 7.9 4.0
MLS-0051609 25.1 6.4
MLS-0050257 7.2 6.0
MLS-0041503 11.3 5.3
MLS-0039077 14.6 9.2
MLS-0050749 14.7 12.8
MLS-0051774 6.3 15.6
MLS-0008909 20.6 19.6
N
N
CF3
SOO
EtO2C
N
N
CF3
SOO
EtO2C
F
N
N
CF3
SOO
OMe
N
N
CF3
SOO
OMe
OMe
N
N
CF3
SOO
O
N
N
CF3
SOO
EtO2C
N
N
CF3
SOO
S
N
N
CF3
SOO
SN
ONN
SH
N
N
CF3
SOO
N
ONO
H
N
N
CF3
SOO
NOMe
O
N
N
CF3
SOO
NH
OMe
OMeO
O
CF3
ON N
S
CF3
N N
S
CF3
OO
a b
O
EtO EtO
O
Figure 4. Synthetic scheme for production of hit compound CID-1151859. (a) 4,4,4-trifluoro-1-phenylbutane-1,3-dione and ethyl 2-(carbamimidoylthio)acetate (1.2 eq.) were heated in ethanol at 100 °C for 5 h to yield ethyl 2-((4-phenyl-6-(trifluoromethyl)pyrimidin-2-yl)thio)acetate in 68% yield. (b) To this product, 2 eq of mCPBA in CH
2Cl
2 was added, and the mixture was
stirred at room temperature for 20 h. After evaporation of solvent and reverse-phase preparative high-performance liquid chromatography, CID-1151859 was isolated in 82% yield.
analogs. We synthesized and tested a series of compounds, and in vitro data were generated to provide SAR data on the hit series. Initial studies focused on analogs in which the phenyl ring and/or the sulfone substituent were modi-fied, and some of these compounds are shown in Table 3.
For example, a 3-methoxy substituent is tolerated (MLS-0066988 = CID-751044), whereas a 3,4-dimethoxy deriv-ative (MLS-0051609 = CID-3244140) is less potent than the parent molecule. It also appears that replacement of the phenyl ring with a heteroaryl moiety such as furan (MLS-0050257 = CID-3237865) or thiophene (MLS-0041503 = CID-655335 and MLS-0051774 = CID-3245258) is viable. Truncating the sulfonyl side chain to a methylsulfone moiety as in MLS-0066988 (CID-751044), MLS-0050257 (CID-3237865), MLS-0051609 (CID-3244140), and MLS-0051774 (CID-3245258) appears to lead to a modest reduction in potency compared with the lead compound. Homologation of the carbethoxymeth-ylene side chain to the carbethoxyethylene variant (MLS-0046458 = CID-2219262) leads to a slight reduction in potency. More pronounced increases in side chain length, as in compounds MLS-0041503 (CID-655335), MLS-0039077 (CID-2037235), MLS-0050749 (CID-3238559), and MLS-0008909 (CID-654688) leads to further reduc-tion in potency. Ultimately, we selected the 4-fluorophenyl

358 Journal of Biomolecular Screening 17(3)
derivative CID-2980973, which is twice as potent as the best hit compound CID-1151859 as the Bfl-1 sulfonyl-pyrimidine series chemical probe, which has received the designation (http://mli.nih.gov/mli/mlp-probes/?dl_id=1289, ML042 = CID-2980973) from the NIH Molecular Libraries Initiative.
DiscussionBcl-2 family proteins have been implicated in several dis-eases. Bfl-1 (known as A1 in the mouse) is an NF-κB-inducible anti-apoptotic member of the Bcl-2 family, which has been implicated in cancer and allergic diseases. With respect to cancer, defective apoptosis is a hallmark of many human tumors and is frequently responsible for resistance to therapy. Transcriptome profiling of diffuse large B-cell lymphomas (a common type of non-Hodgkin lymphoma) identified Bfl-1 as a gene signature associated with chemo-resistance.15 Short-hairpin RNAs targeting Bfl-1 mRNAs induce apoptosis of certain lymphoblastoid B-cell lines and diffuse large B-cell lymphoma cell lines.15 Thus, selective inhibitors of Bfl-1 could find applications for certain lym-phomas or lymphoid leukemias, most likely those where constitutively high NF-κB activity or overexpression of Bfl-1 is observed.
Novel small molecules that target Bcl-2 family proteins are now in clinical trials. ABT737 and its orally available derivative ABT263 are the best characterized and show selective and potent activity against Bcl-2, Bcl-X
L, and Bcl-
W, but not Bcl-B, Mcl-1, and Bfl-1.21,28,29 Although impres-sive single-agent responses to ABT737 or ABT263 have been reported in early clinical trials for some types of malignancy,28,29 many tumor cell lines in vitro and most types of solid tumors in patients show resistance to these compounds.17,18,21,29 The failure of tumor and leukemia cell lines to undergo apoptosis in response to ABT737 or ABT263 in culture has been convincingly attributed, in sev-eral cases, to concomitant expression of other members of the Bcl-2 family not targeted by these compounds, includ-ing Bfl-1 in some instances.18,21 In this scenario, Bfl-1-selective compounds could serve as a complement to selective chemical inhibitors of Bcl-2/Bcl-X
L such as
ABT737/ABT263 for cancer treatment.With respect to allergic diseases, expression of Bfl-1 is
observed in mast cells in vivo and induced in cultured mast cells upon ligation of IgE bound to cell surface Fc recep-tors.10 Various gene silencing experiments have suggested that Bfl-1 is required for survival of mast cells following IgE cross-linking, preventing “activation-induced cell death.”10,30 Thus, compounds targeting Bfl-1 may find util-ity for certain allergic diseases.
Bfl-1 is highly expressed in lymphoid tissues in mice.9,31 The endogenous functions of Bfl-1 are largely unknown,
due to difficulties with accomplishing targeted gene ablation in mouse models. Unlike the other anti-apoptotic members of the Bcl-2 family that have all been successfully ablated in mice, the mouse ortholog of Bfl-1 consists of a cluster of four replicated genes (i.e., four copies of the gene, termed A1a, A1b, A1c, and A1d). Thus, to the extent that the Bfl-1 inhibitory compounds described here cross-react with murine A1 orthologs, they may provide a useful tool for investigating the function of A1 proteins in mouse models.
The most potent of the Bfl-1 inhibitory compounds described here are only marginally active against the target protein in vitro, with approximately micromolar IC
50 val-
ues. Inhibiting (or mimicking as is the case here) protein–protein interactions is a challenge for small molecules, especially in the case of Bcl-2 family proteins where the ligand binding site is large, accommodating BH3 peptides of ~16 amino acids in length. However, the ability to iden-tify small compounds (molecular weight [MW] <500) that show micromolar activity bodes well for employing some of these compounds as a starting point for further optimiza-tion. Given the size of the BH3 peptide binding site, it is unlikely that extensive analoging on the sulfonylpyrimidine or chloromaleimide scaffolds will drive potency into the nanomolar level. Rather, by analogy to ABT-737 and ABT-263 (which have MW of ≈813–975 Daltons and yet possess excellent pharmacological properties28), use of technolo-gies such as “chemical fragment” screening by nuclear magnetic resonance (NMR), x-ray crystallography, or sur-face plasmon resonance (SPR) will likely be required to identify complementary chemical moieties that could be covalently linked onto the sulfonylpyrimidine or chloroma-leimide scaffolds to increase the surface area of the BH3 peptide-binding pocket filled by the compounds.
In this report, we demonstrated that Bfl-1-binding com-pounds show activity in a mitochondrial assay that mea-sures Bfl-1-mediated protection of these organelles. The advantage of this bioassay over a cell-based assay is that it avoids many of the variables that affect compound perfor-mance irrespective of target binding, such as cell membrane permeability, serum stability, and plasma protein binding. Interestingly, the sulfonylpyrimidine and chloromaleimide compounds showed different activity profiles in this assay, where sulfonylpyrimidine CID-1151859 induced release of SMAC from isolated mitochondria, whereas chloroma-leimide compound CID-1180676 did not. Given that the exemplary chloromaleimide compound tested here was more potent against Bfl-1 by about 6- to 11-fold (depending on the assay method), the ability of the sulfonylpyrimidine to stimulate release of proteins from mitochondria cannot be explained by its Bfl-1 inhibitory activity. Thus, the sul-fonylpyrimidine compound possesses a bioactivity separate from Bfl-1, although this bioactivity can be neutralized upon addition of Bfl-1 protein (but not Bcl-X
L protein) to

Zhai et al. 359
the mitochondria assay. Because the chloromaleimide did not induce SMAC release from mitochondria, we used recombinant active Bid (a pro-apoptotic Bcl-2 family pro-tein) to stimulate protein release from mitochondria in vitro, demonstrating that recombinant Bfl-1 and Bcl-X
L block the
impact of Bid, with chloromaleimide CID-1180676 (also known as MLS-0053105 and ML041) (http://mli.nih.gov/mli/mlp-probes/?dl_id=1289) neutralizing Bfl-1 but not Bcl-X
L. Thus, we favor the chloromaleimide series as a
starting point for future optimization campaigns.
Acknowledgments
We thank Melanie Hanaii and Tessa Siegfried for manuscript preparation.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research and/or authorship of this article: This work was supported by the NIH (HG005033, MH079863, and CA113318).
References
1. Youle, R. J.; Strasser, A. The BCL-2 Protein Family: Oppos-ing Activities That Mediate Cell Death. Nat. Rev. Mol. Cell. Biol. 2008, 9(1), 47–59.
2. Reed, J. C.; Doctor, K. S.; Godzik, A. The Domains of Apopto-sis: A Genomics Perspective. Sci. STKE 2004, 2004(239), re9.
3. Chipuk, J. E.; Moldoveanu, T.; Llambi, F.; Parsons, M. J.; Green, D. R. The BCL-2 Family Reunion. Mol. Cell. 2010, 37(3), 299–310.
4. van Delft, M. F.; Huang, D. C. How the Bcl-2 Family of Pro-teins Interact to Regulate Apoptosis. Cell. Res. 2006, 16(2), 203–213.
5. Zong, W. X.; Edelstein, L. C.; Chen, C.; Bash, J.; Gelinas, C. The Prosurvival Bcl-2 Homolog Bfl-1/A1 Is a Direct Tran-scriptional Target of NF-kappaB That Blocks TNFalpha-induced Apoptosis. Genes Dev 1999, 13(4), 382–387.
6. Wang, C. Y.; Guttridge, D. C.; Mayo, M. W.; Baldwin, A. S., Jr. NF-kappaB Induces Expression of the Bcl-2 Homologue A1/Bfl-1 to Preferentially Suppress Chemotherapy-Induced Apoptosis. Mol. Cell. Biol. 1999, 19(9), 5923–5929.
7. Lee, H. H.; Dadgostar, H.; Cheng, Q.; Shu, J.; Cheng, G. NF-kappaB-Mediated Up-regulation of Bcl-x and Bfl-1/A1 Is Required for CD40 Survival Signaling in B Lymphocytes. Proc. Natl. Acad. Sci. U. S. A. 1999, 96(16), 9136–9141.
8. Grumont, R. J.; Rourke, I. J.; Gerondakis, S. Rel-dependent Induction of A1 Transcription Is Required to Protect B Cells from Antigen Receptor Ligation-Induced Apoptosis. Genes Dev. 1999, 13(4), 400–411.
9. Choi, S. S.; Park, I. C.; Yun, J. W.; Sung, Y. C.; Hong, S. I.; Shin, H. S. A Novel Bcl-2 Related Gene, Bfl-1, Is Overex-pressed in Stomach Cancer and Preferentially Expressed in Bone Marrow. Oncogene 1995, 11(9), 1693–1698.
10. Xiang, Z.; Ahmed, A. A.; Moller, C.; Nakayama, K.; Hatakeyama, S.; Nilsson, G. Essential Role of the Prosurvival bcl-2 Homologue A1 in Mast Cell Survival after Allergic Activation. J. Exp. Med. 2001, 194(11), 1561–1569.
11. Andre, J.; Cimaz, R.; Ranchin, B.; Galambrun, C.; Bertrand, Y.; Bouvier, R.; Rieux-Laucat, F.; Trescol-Biemont, M. C.; Cochat, P.; Bonnefoy-Berard, N. Overexpression of the Anti-apoptotic Gene Bfl-1 in B Cells from Patients with Familial Systemic Lupus Erythematosus. Lupus 2007, 16(2), 95–100.
12. Lopez-Hoyos, M.; Carrio, R.; Merino, R.; Buelta, L.; Izui, S.; Nunez, G.; Merino, J. Constitutive Expression of bcl-2 in B Cells Causes a Lethal Form of Lupuslike Autoimmune Dis-ease after Induction of Neonatal Tolerance to H-2b Alloantigens. J. Exp. Med. 1996, 183(6), 2523–2531.
13. Kucharczak, J. F.; Simmons, M. J.; Duckett, C. S.; Gelinas, C. Constitutive Proteasome-Mediated Turnover of Bfl-1/A1 and Its Processing in Response to TNF Receptor Activation in FL5.12 Pro-B Cells Convert It into a Prodeath Factor. Cell Death Differ. 2005, 12(9), 1225–1239.
14. Olsson, A.; Norberg, M.; Okvist, A.; Derkow, K.; Choudhury, A.; Tobin, G.; Celsing, F.; Osterborg, A.; Rosenquist, R.; Jondal, M.; et al. Upregulation of bfl-1 Is a Potential Mechanism of Chemore-sistance in B-Cell Chronic Lymphocytic Leukaemia. Br. J. Cancer 2007, 97(6), 769–777.
15. Brien, G.; Trescol-Biemont, M. C.; Bonnefoy-Berard, N. Down-regulation of Bfl-1 Protein Expression Sensitizes Malignant B Cells to Apoptosis. Oncogene 2007, 26(39), 5828–5832.
16. Morales, A. A.; Olsson, A.; Celsing, F.; Osterborg, A.; Jondal, M.; Osorio, L. M. High Expression of bfl-1 Contributes to the Apoptosis Resistant Phenotype in B-cell Chronic Lympho-cytic Leukemia. Int. J. Cancer 2005, 113(5), 730–737.
17. Deng, J.; Carlson, N.; Takeyama, K.; Dal Cin, P.; Shipp, M.; Letai, A. BH3 Profiling Identifies Three Distinct Classes of Apoptotic Blocks to Predict Response to ABT-737 and Conventional Chemotherapeutic Agents. Cancer Cell. 2007, 12(2), 171–185.
18. van Delft, M. F.; Wei, A. H.; Mason, K. D.; Vandenberg, C. J.; Chen, L.; Czabotar, P. E.; Willis, S. N.; Scott, C. L.; Day, C. L.; Cory, S.; et al. The BH3 Mimetic ABT-737 Tar-gets Selective Bcl-2 Proteins and Efficiently Induces Apop-tosis via Bak/Bax if Mcl-1 Is Neutralized. Cancer Cell. 2006, 10(5), 389–399.
19. Kuwana, T.; Bouchier-Hayes, L.; Chipuk, J. E.; Bonzon, C.; Sullivan, B. A.; Green, D. R.; Newmeyer, D. D. BH3 Domains of BH3-only Proteins Differentially Regulate Bax-mediated Mitochondrial Membrane Permeabilization Both Directly and Indirectly. Mol. Cell. 2005, 17(4), 525–535.
20. Chen, L.; Willis, S. N.; Wei, A.; Smith, B. J.; Fletcher, J. I.; Hinds, M. G.; Colman, P. M.; Day, C. L.; Adams, J. M.;

360 Journal of Biomolecular Screening 17(3)
Huang, D. C. Differential Targeting of Prosurvival Bcl-2 Proteins by Their BH3-only Ligands Allows Complementary Apoptotic Function. Mol. Cell. 2005, 17(3), 393–403.
21. Konopleva, M.; Contractor, R.; Tsao, T.; Samudio, I.; Ruvolo, P. P.; Kitada, S.; Deng, X.; Zhai, D.; Shi, Y. X.; Sneed, T.; et al. Mechanisms of Apoptosis Sensitivity and Resistance to the BH3 Mimetic ABT-737 in Acute Myeloid Leukemia. Can-cer Cell. 2006, 10(5), 375–388.
22. Reed, J. C.; Pellecchia, M. Apoptosis-Based Therapies for Hematologic Malignancies. Blood 2005, 106(2), 408–418.
23. Cashman, J. R.; MacDonald, M.; Ghirmai, S.; Okolotowicz, K. J.; Sergienko, E.; Brown, B.; Garcia, X.; Zhai, D.; Dahl, R.; Reed, J. C. Inhibition of Bfl-1 with N-aryl Maleimides. Bioorg. Med. Chem. Lett. 2010, 20(22), 6560–6564.
24. Zhai, D.; Luciano, F.; Zhu, X.; Guo, B.; Satterthwait, A. C.; Reed, J. C. Humanin Binds and Nullifies Bid Activity by Blocking Its Activation of Bax and Bak. J. Biol. Chem. 2005, 280(16), 15815–15824.
25. Zhai, D.; Jin, C.; Satterthwait, A. C.; Reed, J. C. Comparison of Chemical Inhibitors of Antiapoptotic Bcl-2-Family Pro-teins. Cell Death Differ. 2006, 13(8), 1419–1421.
26. Zhai, D.; Jin, C.; Shiau, C. W.; Kitada, S.; Satterthwait, A. C.; Reed, J. C. Gambogic Acid is an Antagonist of Antiapoptotic
Bcl-2 Family Proteins. Mol. Cancer Ther. 2008, 7(6), 1639–1646.
27. Zhai, D.; Jin, C.; Huang, Z.; Satterthwait, A. C.; Reed, J. C. Differential Regulation of Bax and Bak by Anti-apoptotic Bcl-2 Family Proteins Bcl-B and Mcl-1. J. Biol. Chem. 2008, 283(15), 9580–9586.
28. Tse, C.; Shoemaker, A. R.; Adickes, J.; Anderson, M. G.; Chen, J.; Jin, S.; Johnson, E. F.; Marsh, K. C.; Mitten, M. J.; Nimmer, P.; et al. ABT-263: A Potent and Orally Bioavailable Bcl-2 Family Inhibitor. Cancer Res. 2008, 68(9), 3421–3428.
29. Oltersdorf, T.; Elmore, S. W.; Shoemaker, A. R.; Armstrong, R. C.; Augeri, D. J.; Belli, B. A.; Bruncko, M.; Deckwerth, T. L.; Dinges, J.; Hajduk, P. J.; et al. An Inhibitor of Bcl-2 Family Proteins Induces Regression of Solid Tumours. Nature 2005, 435(7042), 677–681.
30. Xiang, Z.; Moller, C.; Nilsson, G. IgE-Receptor Activation Induces Survival and Bfl-1 Expression in Human Mast Cells but Not Basophils. Allergy 2006, 61(9), 1040–1046.
31. Verschelde, C.; Walzer, T.; Galia, P.; Biemont, M. C.; Quemeneur, L.; Revillard, J. P.; Marvel, J.; Bonnefoy-Berard, N. A1/Bfl-1 Expression Is Restricted to TCR Engage-ment in T Lymphocytes. Cell Death Differ. 2003, 10(9), 1059–1067.




















