
Alterations in Bcl2/Bax protein levels in platelets form part of an ionomycin-induced process that...
8
Alterations in Bcl-2/Bax protein levels in platelets form part of an ionomycin-induced process that resembles apoptosis DAINA M. VANAGS , 1 S TEN O RRENIUS 1 AND MIGUEL AGUILAR -S ANTELISES 1,2 1 Institute of Environmental Medicine, Division of Toxicology, Karolinska Institute, and 2 Centre for Molecular Medicine, Karolinska Hospital, Stockholm, Sweden Received 26 May 1997; accepted for publication 26 September 1997 Summary. Platelets are physiologically anucleated cells, derived from megakaryocytes, that undergo vesiculation and transformation into small particles when they are stimulated in vitro by ionomycin and other agents. Electron microscopy images suggest a similarity to apoptosis in cells with nuclei, which ends with cell disintegration and formation of apoptotic bodies. By PCR, we have demonstrated mRNA expression of bcl-2, bax a and p53 in highly purified non-stimulated platelets. A side-scatter shift and a decrease in the Bcl-2/Bax protein ratio were observed by flow cytometry analysis after stimulation with ionomycin. The ionomycin-induced modifications were inhibited by the calpain I inhibitor calpeptin and, less effectively, by VAD-cmk, a broad- spectrum caspase inhibitor. However, caspase 3-like activity was very low, with only a twofold increase after ionomycin stimulation, as measured by the cleavage of the fluorogenic peptide substrate DEVD-AMC. Our data indicate that platelets may constitute a natural model for the analysis of cytoplasmic events in apoptosis. Keywords: platelets, apoptosis, Bcl-2, Bax, vesiculation. Most, if not all, animal cells have the ability to self-destruct by apoptosis (Steller, 1995). Cell death by apoptosis is characterized by morphological and biochemical changes that include cytoskeletal alterations, blebbing, chromatin condensation, nuclear fragmentation, phosphatidylserine (PS) exposure and fragmentation into small membrane- enclosed apoptotic bodies (Steller, 1995; Hale et al, 1996). Bcl-2 is the prototype of a gene family whose expression contributes to regulate cell susceptibility to apoptosis (Cory, 1995; Yang & Korsmeyer, 1996). Bcl-2 is associated with enhanced survival whereas Bax is a cell death promoter (Oltvai & Korsmeyer, 1994; Yang & Korsmeyer, 1996). Bcl-2 homologues share two highly conserved regions that serve to form homo- and hetero-dimers with each other (Yin et al, 1994; Sedlak et al, 1995). Bcl-2 and Bax heterodimerization is required for inhibition of apoptosis (Yin et al, 1994) and the ratio of Bcl-2/Bax may determine whether a cell will survive or undergo apoptosis following an apoptotic stimulus (Oltvai et al, 1993; Osorio et al, 1997). p53 is another gene that participates in the regulation of cell death by apoptosis. Different types of DNA damage result in the accumulation of biologically active p53 protein which enhances the tran- scription of several gene products, including the cyclin- dependent kinase inhibitor, p21, which stops the cell cycle, giving the cell an opportunity to repair the DNA damage before re-entering the cell cycle or to undergo apoptosis (Levine, 1997). Constitutive expression of the apoptotic cell death machinery has been analysed by removing the cell nuclei (Jacobson et al, 1994; Schulze-Osthoff et al, 1994). The resultant cytoplasts can be easily induced to present morphological features of apoptosis, except when prepared from cells expressing high levels of Bcl-2. It has been suggested therefore that, at least in some cases, the nucleus is not required for induction of apoptosis (Jacobson et al, 1994; Schulze-Osthoff et al, 1994). Studies with cell-free extracts have also concluded that cytoplasm contains essential factors for triggering apoptosis, although the presence of mitochondria seems to be necessary for the execution process (Earnshaw, 1995). Platelets are anucleated cells derived from megakaryocytes that survive in the peripheral circulation for 8–10 d before being taken up and destroyed by phagocytic cells in the spleen, liver and bone marrow (Thompson & Jakubowski, British Journal of Haematology , 1997, 99, 824–831 824 q 1997 Blackwell Science Ltd Correspondence: Dr M. Aguilar-Santelises, Haematology Section (L8.3), Centre for Molecular Medicine, Karolinska Hospital, S-171 76, Stockholm, Sweden.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Alterations in Bcl2/Bax protein levels in platelets form part of an ionomycin-induced process that...

Alterations in Bcl-2/Bax protein levels in plateletsform part of an ionomycin-induced processthat resembles apoptosis
DAINA M. VANAGS,1 ST EN ORRENIUS1
AND MIGUEL AGUILAR-SANTEL ISES1,2 1Institute of Environmental Medicine,
Division of Toxicology, Karolinska Institute, and 2Centre for Molecular Medicine, Karolinska Hospital, Stockholm, Sweden
Received 26 May 1997; accepted for publication 26 September 1997
Summary. Platelets are physiologically anucleated cells,derived from megakaryocytes, that undergo vesiculationand transformation into small particles when they arestimulated in vitro by ionomycin and other agents.Electron microscopy images suggest a similarity toapoptosis in cells with nuclei, which ends with celldisintegration and formation of apoptotic bodies. ByPCR, we have demonstrated mRNA expression of bcl-2,baxa and p53 in highly purified non-stimulated platelets.A side-scatter shift and a decrease in the Bcl-2/Bax proteinratio were observed by flow cytometry analysis after
stimulation with ionomycin. The ionomycin-inducedmodifications were inhibited by the calpain I inhibitorcalpeptin and, less effectively, by VAD-cmk, a broad-spectrum caspase inhibitor. However, caspase 3-likeactivity was very low, with only a twofold increase afterionomycin stimulation, as measured by the cleavage of thefluorogenic peptide substrate DEVD-AMC. Our dataindicate that platelets may constitute a natural model forthe analysis of cytoplasmic events in apoptosis.
Keywords: platelets, apoptosis, Bcl-2, Bax, vesiculation.
Most, if not all, animal cells have the ability to self-destructby apoptosis (Steller, 1995). Cell death by apoptosis ischaracterized by morphological and biochemical changesthat include cytoskeletal alterations, blebbing, chromatincondensation, nuclear fragmentation, phosphatidylserine(PS) exposure and fragmentation into small membrane-enclosed apoptotic bodies (Steller, 1995; Hale et al, 1996).
Bcl-2 is the prototype of a gene family whose expressioncontributes to regulate cell susceptibility to apoptosis (Cory,1995; Yang & Korsmeyer, 1996). Bcl-2 is associated withenhanced survival whereas Bax is a cell death promoter(Oltvai & Korsmeyer, 1994; Yang & Korsmeyer, 1996). Bcl-2homologues share two highly conserved regions that serve toform homo- and hetero-dimers with each other (Yin et al,1994; Sedlak et al, 1995). Bcl-2 and Bax heterodimerizationis required for inhibition of apoptosis (Yin et al, 1994) andthe ratio of Bcl-2/Bax may determine whether a cell willsurvive or undergo apoptosis following an apoptotic stimulus(Oltvai et al, 1993; Osorio et al, 1997). p53 is another genethat participates in the regulation of cell death by apoptosis.
Different types of DNA damage result in the accumulation ofbiologically active p53 protein which enhances the tran-scription of several gene products, including the cyclin-dependent kinase inhibitor, p21, which stops the cell cycle,giving the cell an opportunity to repair the DNA damagebefore re-entering the cell cycle or to undergo apoptosis(Levine, 1997).
Constitutive expression of the apoptotic cell deathmachinery has been analysed by removing the cell nuclei(Jacobson et al, 1994; Schulze-Osthoff et al, 1994). Theresultant cytoplasts can be easily induced to presentmorphological features of apoptosis, except when preparedfrom cells expressing high levels of Bcl-2. It has beensuggested therefore that, at least in some cases, the nucleusis not required for induction of apoptosis (Jacobson et al,1994; Schulze-Osthoff et al, 1994). Studies with cell-freeextracts have also concluded that cytoplasm containsessential factors for triggering apoptosis, although thepresence of mitochondria seems to be necessary for theexecution process (Earnshaw, 1995).
Platelets are anucleated cells derived from megakaryocytesthat survive in the peripheral circulation for 8–10 d beforebeing taken up and destroyed by phagocytic cells in thespleen, liver and bone marrow (Thompson & Jakubowski,
British Journal of Haematology, 1997, 99, 824–831
824 q 1997 Blackwell Science Ltd
Correspondence: Dr M. Aguilar-Santelises, Haematology Section(L8.3), Centre for Molecular Medicine, Karolinska Hospital, S-17176, Stockholm, Sweden.

825Bcl-2, Bax and p53 Expression in Platelets
q 1997 Blackwell Science Ltd, British Journal of Haematology 99: 824–831
1988). Although platelets do not have a nucleus, theycontain metabolically stable mRNA and are capable oflimited protein synthesis (Newman et al, 1988; Djaffar et al,1991; Santoso et al, 1993). After stimulation with collagen,thrombin or calcium ionophore, platelets undergo eventssimilar to those occurring in nucleated cells duringapoptosis, including PS exposure (Dachary-Prigent et al,1993), spectrin cleavage (Fox et al, 1987) and fragmentationinto membrane enclosed microvesicles (Owens, 1994). Wehave analysed platelets for the expression of Bcl-2, Bax andp53, to find out if platelets may constitute a physiologicalmodel to explore nucleus-independent mechanisms ofapoptosis. We have also attempted to correlate this expres-sion with vesiculation and density changes observed inplatelets when stimulated by ionomycin. Our results indicatethat platelets express apoptosis-related genes and thatplatelet activation results in Bcl-2/Bax modulation, alongwith cytoplasmic events which resemble apoptosis.
MATERIALS AND METHODS
Reagents. Apyrase, prostaglandin E1, ionomycin, rabbitIgG, saponin and bovine serum albumin were from Sigma(St Louis, Mo., U.S.A.). Dynabeads mRNA direct kit wasacquired from Dynal (Oslo, Norway). Primers for bcl-2,baxa and p53 amplification were synthesized by Scandina-vian Gene Synthesis (Koping, Sweden). G3PDH primerswere from Clontech (Palo Alto, Calif., U.S.A.). RNAsin waspurchased from Promega (Madison, Wis., U.S.A.), DTT,Moloney murine leukaemia virus (M-MLV) reverse tran-scriptase and first-strand buffer were from GIBCO BRL(Gaithersburg, Md., U.S.A.). Random primers (pd(N)6),ultrapure dNTP and agarose NA were obtained fromPharmacia (Uppsala, Sweden). Taq polymerase and theDNA mw marker VI were from Boehringer Mannheim(Bromma, Sweden). Bcl-2 (N19) rabbit ab, Bcl-2 controlpeptide (N19P), Bax (P19) rabbit ab and Bax controlpeptide (P19P) were purchased from Santa Cruz Biotech-nology (Santa Cruz, Calif., U.S.A.). FITC-conjugated swineanti-rabbit Ig was bought from Dakopatts (Copenhagen,Denmark). Fluorescent monodisperse carboxylated micro-spheres were bought from Polysciences Inc. (Warrington,Pa., U.S.A.). VAD-cmk (Z-Val-Ala-Asp-chloromethylketone;a cell-permeable broad-spectrum caspase inhibitor) wasfrom Enzyme Systems Progress (Dublin, Calif., U.S.A.) andcalpeptin (benzyloxycarbonyl-L-leucylnorleucinal, a cell-permeable peptide calpain I inhibitor) was from Biomol(Plymouth Meeting, Pa., U.S.A.).
Preparation of platelets. Venous blood was obtained fromhealthy donors and mixed with acid-citrate-dextrose (1 volumeanticoagulant : 6 volumes blood). Platelet-rich plasma (PRP)was prepared by centrifugation at 200 g for 11 min at roomtemperature in the presence of 1 U/ml apyrase and 1 mM
prostaglandin E1 (PGE1). Platelets in the upper half of the PRPwere carefully harvested and centrifuged at 1500 g for 15 min.This new pellet was washed twice in pH 6·5 HEPES-Tyrode’sbuffer (136 mM NaCl, 2·68 mM KCl, 1 mM MgCl2, 0·36 mM
NaH2PO4, 11·9 mM NaHCO3, 3·4 mM HEPES, 0·1% glucose,0·35% BSA) containing 50 IU/ml heparin, 1 U/ml apyrase,
1 mM PGE1 and 2 mM CaCl2. Platelets were then resuspended inpH 7·4 HEPES-Tyrode’s buffer containing 1 mM CaCl2 andexamined microscopically to select a platelet:leucocyte ratio>50 000:1. An aliquot of platelets (108/ml) was incubatedwith 7 mM ionomycin for 10 min at 378C without stirring, toensure maximal microparticle formation. In some experimentsthe platelets were preincubated with 100 mg/ml calpeptin or20 mM VAD-cmk for 30 min. After isolation and treatment,non-stimulated and stimulated platelets were pelleted, fixedwith 4% paraformaldehyde in PBS for 10 min, and washedtwice with PBS.
Electron microscopy. Platelets on micropore filters werefixed with 4% glutaraldehyde in 0·1 M phosphate buffer, pH7·3 for 3 h, post-fixed for 1 h in 2% osmium tetroxide inphosphate buffer, and dehydrated with ethanol. Plateletswere critical-point dried, mounted on stubs, and sputter-coated with gold before examination with a Philips 505scanning electron microscope.
RNA extraction, cDNA preparation and PCR. Platelets werepelleted at 48C for mRNA extraction. mRNA was extractedfrom 109 platelets by using the Dynabeads mRNA direct kit,according to the manufacturer’s instructions. Poly Aþ RNAwas detached from Dynabeads oligo (dt)25, ethanol pre-cipitated, denatured and reverse transcribed with randomhexanucleotides, to obtain 40 ml of cDNA. Thereafter, 1 ml ofcDNA was amplified in a 20 ml PCR reaction mixture, withcycling conditions optimized for each pair of primers (TableI). The number of cycles and annealing temperature usedwere: 35 cycles and 608C for glyceraldehyde-3-phosphatedehydrogenase (G3PDH), 35 cycles and 728C for bcl-2, 35cycles and 578C for p53 and 29 cycles and 668C, two cyclesand 678C, two cycles and 688C, and two cycles and 698C forbaxa (Aguilar-Santelises et al, 1996). The constant domain(Cb) of the T-cell receptor (TCR) was amplified for 25 cycles,608C. cDNA from the membrane glycoprotein IIIa (GP IIIa)was amplified during 35 cycles, with 458C as the annealingtemperature. Amplifications with all primers, except thosefor GP IIIa, were performed with 1·5 mM MgCl2. GP IIIaamplification was done with 2·5 mM MgCl2. Additional13 min at 728C and 13 min at 48C were employed at theend of the amplifications. The PCR products were thenresolved by agarose gel electrophoresis and photographed.
Table I. Primers utilized for PCR amplification.
S G3PDH 50TGAAGGTCGGAGTCAACGGATTTGGT30 983 bpAS G3PDH 50CATGTGGGCCATGAGGTCCACCAC30
S Cb 50GTGCACCTCCTTCCCATT30 140 bpAS Cb 50GTCGCTGTGTTTGAGCCATCAGAA30
S GP IIIa 50GTCCTCCAGCTCATTGTTGATGC30 1086 bpAS GP IIIa 50CATCACTGAGAGCAGGACCACCAGG30
S bcl-2 50CGACGACTTCTCCCGCCGCTACCGC30 319 bpAS bcl-2 50CCGCATGCTGGGGCCGTACAGTTCC30
S baxa 50GCTCTGAGCAGATCATGAAGACAG30 488 bpAS baxa 50CACAAAGATGGTCACGGTCTGC30
S p53 50TCTGTGACTTGCACGTACTC30 640 bpAS p53 50CACGGATCTGAAGGGTGAAA30
Flow cytometry. PFA-fixed platelets (108/ml) were resus-pended in PBS containing 0·1% saponin and 0·2% BSA topermeabilize the cells and to block non-specific staining. Thisbuffer was used for subsequent incubations and rinses duringthe whole staining. Platelets were aliquoted and stained forimmunofluorescence with rabbit ab against Bcl-2 or Bax plusswine anti-rabbit FITC-conjugate as secondary ab. Controlswith cells alone, cells with the conjugate alone or with anirrelevant rabbit IgG ab plus the conjugate, were run inparallel. Platelets were also stained with Bcl-2 (N19) and Bax(P19) rabbit ab, preincubated overnight with their specificpeptides (N19P and P19P, respectively), to confirm thespecificity of the staining. Stained samples were analysed byflow cytometry (FACS) using a Becton Dickinson Vantageapparatus equipped with a Coherent Enterprise 488 nm laser.Fluorescent monodisperse carboxylated microspheres with1·72 mm diameter were used to calibrate the instrument.10 000 cells were acquired for each determination, with a ratelower than 400 events/s. Results are presented as specificmean fluorescence intensity (MFI) which was obtained bysubtracting MFI values in the presence of the irrelevant abplus the FITC conjugate from MFI values obtained in thepresence of the specific ab plus the FITC-conjugate.
Phosphatidylserine (PS) exposure. 5 × 106 control or stimu-lated platelets were used to measure plasma clotting time,using Russell’s viper venom (RVV) as indirect measurementof PS exposure (Fadok et al, 1992; Vanags et al, 1996).
Caspase 3-like activity. This was measured by the ability tocleave the peptide substrate Asp-Glu-Val-Asp (DEVD) linkedto amino-4-methylcoumarin (AMC) in a fluorometric assaymodified from Nicholson et al (1995). Platelet lysate andsubstrate were combined in a reaction buffer with 100 mM
HEPES, 10% sucrose, 5 mM DTT, 0·1% CHAPS; pH 7·25.Cleavage was monitored by AMC liberation using 355 nmexcitation and 460 nm emission wavelengths. Fluorescenceunits were converted to pmoles of AMC/min/109 platelets,using a standard curve with increasing doses of AMC againstfluorescence (Thornberry et al, 1992).
Statistical analysis. One-way ANOVA, Mann-Whitney Uand Wilcoxon signed rank test were used to analyse the data.
RESULTS
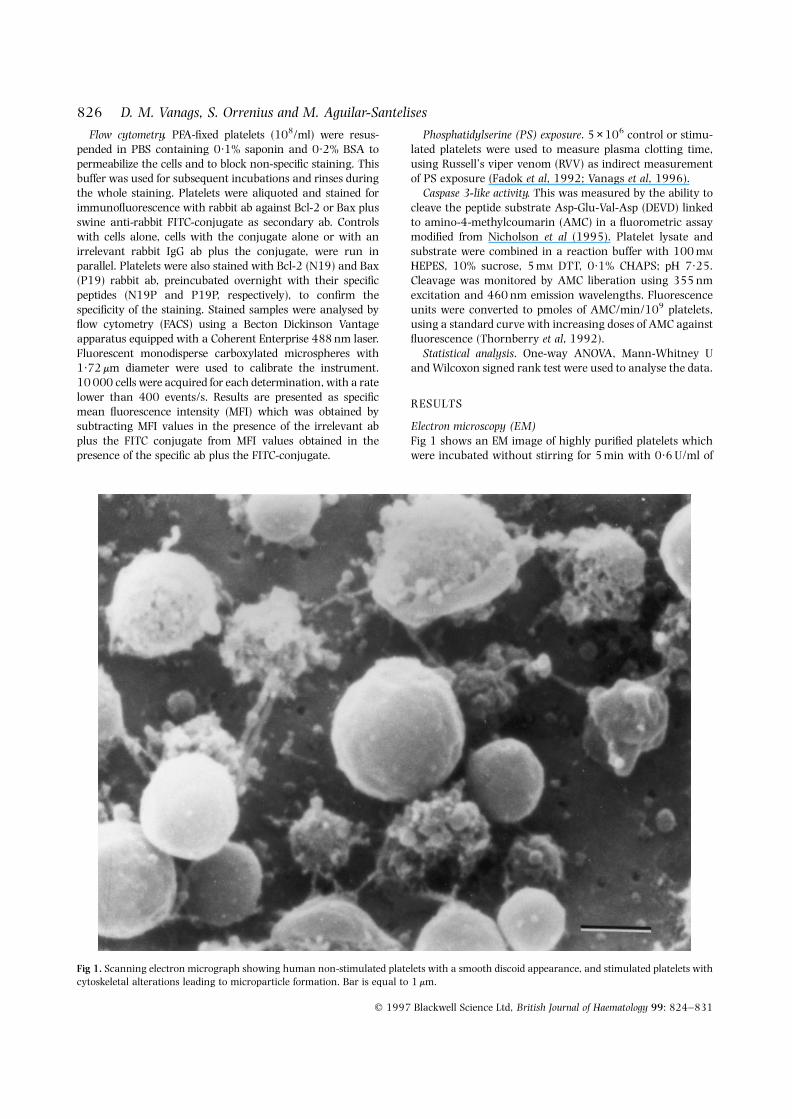
Electron microscopy (EM)Fig 1 shows an EM image of highly purified platelets whichwere incubated without stirring for 5 min with 0·6 U/ml of
q 1997 Blackwell Science Ltd, British Journal of Haematology 99: 824–831
826 D. M. Vanags, S. Orrenius and M. Aguilar-Santelises
Fig 1. Scanning electron micrograph showing human non-stimulated platelets with a smooth discoid appearance, and stimulated platelets withcytoskeletal alterations leading to microparticle formation. Bar is equal to 1 mm.

827Bcl-2, Bax and p53 Expression in Platelets
q 1997 Blackwell Science Ltd, British Journal of Haematology 99: 824–831
thrombin and fixed. The EM photograph was taken withplatelets on Millipore inert teflon filters which do not affectplatelets on their own (Holme et al, 1994). Thrombinstimulation under these conditions rendered both intact andaltered platelets in the same field. Fig 1 shows the contrastbetween activated vesiculating platelets with cytoskeletalmodifications and non-stimulated smooth-shaped platelets.Trypan blue exclusion from the vesicles indicated thatplasma membrane integrity was maintained until fixation(not shown).
Gene expressionWe used RT-PCR to analyse the expression of bcl-2, baxa
and p53 in platelets. All five samples of platelet cDNA werepositive for bcl-2 and baxa, whereas only three of themwere also positive for p53 (Fig 2). The five samples ofplatelet cDNA were positive for the house-keeping geneG3PDH. To ensure that platelet-specific mRNA wasconverted into cDNA, we tested it by PCR amplificationwith primers for the Cb domain of the T-cell receptor andfor the platelet membrane glycoprotein GP IIIa (Jeddi-Tehrani et al, 1994; Newman et al, 1988). As expected,cDNA from the platelets was negative for Cb and positive forGP IIIa, whereas cDNA from the T-cell lines MOLT-3,
MOLT-4, CCRF-CEM and HUT-78 was positive for Cb andnegative for GP IIIa. Both B-cell lines (RL-7 and DAUDI)were negative for Cb and GP IIIa. In addition, all T- and B-cell lines were positive for bcl-2, baxa and p53, exceptDAUDI which was negative for bcl-2, as previously reported(Aguilar-Santelises et al, 1996). These results demonstratethat the mRNA from platelets used to prepare cDNA wasnot contaminated with mRNA from T lymphocytes andthat platelets contain mRNA from apoptosis-related genes(Fig 2).
FACS analysisIsolated platelets were stimulated with ionomycin andcompared with non-stimulated platelets by flow cytometry.Ionomycin efficiently stimulates platelets, inducing theformation of membrane-enclosed vesicles or microparticles(Owens, 1994). In our experiments, ionomycin stimulationresulted in increased platelet density, as measured bychanges in SSC that were statistically significant (P<0·05)when compared to SSC from non-stimulated platelets (Fig 3).SSC alterations were blocked by 100 mg/ml calpeptin(P<0·05), but not by 20 mM VAD-cmk (Fig 3). Ionomycin-induced microparticle formation was also blocked bycalpeptin but not by VAD-cmk (not shown). SSC values in
Fig 2. PCR of cDNA obtained from platelets from five different normal donors, four T-cell lines (MOLT-3, MOLT-4, CCRF-CEM and HUT-78) andtwo B-cell lines (RL-7 and DAUDI).

the presence of calpeptin or VAD-cmk were not different fromthe controls (not shown).
Staining of platelets with ab against Bcl-2 (N19) and Bax(P19) produced an increase in mean fluorescence intensity(MFI) for all the samples tested, although there was avariability in the MFI for the different samples (Table II). TheBcl-2 and Bax protein levels were altered after stimulationand a consistent decrease in Bcl-2/Bax ratio was observedwith platelets from six different donors to reach only 60% ofthe values observed with non-stimulated platelets (P<0·05;Table II). The Bcl-2/Bax ratio remained low after ionomycin-induced activation, even in the presence of calpeptin.However, Bcl-2 and Bax protein levels were partiallyinhibited by 100 mg/ml calpeptin (P<0·05) and to a lesserdegree by 20 mM VAD-cmk (Table III). Bcl-2 and Bax levelsafter incubation of platelets with calpeptin or VAD-cmk alonewere not different from the controls (not shown).
The specificity of Bcl-2 and Bax ab was confirmed byblocking the stain with overnight incubation of theantibodies with their specific peptides. Bcl-2 staining wasblocked with an excess of the specific peptide N19Pcorresponding to amino acids 4–21 from human Bcl-2.Bax staining was blocked with an excess of the specificpeptide P19P corresponding to amino acids 43–61 frommurine Bax (Fig 4).
Phosphatidylserine exposureA reduced RVV clotting time indicated significant PSexposure in the samples stimulated with ionomycin, ascompared to the controls (P<0·05), in accordance withprevious reports (Dachary-Prigent et al, 1993). Plateletswere preincubated with 100 mg/ml calpeptin or 20 mM VAD-cmk in order to find out if these inhibitors could prevent PSexposure. However, neither calpeptin nor VAD-cmk modifiedthis effect of ionomycin (Table III). Data obtained for PSexposure after incubation of platelets with calpeptin or VAD-cmk alone were not different from the controls (not shown).
q 1997 Blackwell Science Ltd, British Journal of Haematology 99: 824–831
828 D. M. Vanags, S. Orrenius and M. Aguilar-Santelises
Table II. Bcl-2/Bax staining in platelets.*
Non-stimulated Ionomycin stimulated
Bcl-2 Bax Ratio Bcl-2 Bax Ratio
12·30 05·30 2·32 12·10 9·50 1·2752·70 42·20 1·25 226·00 322·00 0·7019·04 20·99 0·91 94·28 126·73 0·7421·78 33·07 0·66 46·68 225·73 0·21
9·23 10·33 0·89 18·13 21·86 0·8224·90 14·20 1·75 142·20 150·60 0·94
Mean ratio 6 SEM 1·29 6 0·25 0·78 6 0·14*
* Specific MFI, obtained by subtracting MFI values in the presence of the irrelevant ab plus the FITC-conjugate from MFI values in the presence of the specific ab plus the FITC-conjugate.
* P<0·05 as compared to the non-stimulated ratio.
Fig 3. Histogram plot showing logarithmic side scatter from non-stimulated platelets (——), and ionomycin-activated platelets in theabsence (– – –) or in the presence (? ? ? ) of 100 mg/ml calpeptin.
Fig 4. Bcl-2 (A) and Bax (B) staining of platelets with N19 (Bcl-2)and P19 (Bax) rabbit antibodies incubated overnight with (? ? ? ) orwithout (– – –) their specific peptides. Continuous lines correspondto background staining of cells incubated with the irrelevant ab plusthe FITC conjugate.

829Bcl-2, Bax and p53 Expression in Platelets
q 1997 Blackwell Science Ltd, British Journal of Haematology 99: 824–831
Caspase-3-like activityThe occurrence of caspase-3-like activity in platelets wasalso investigated using DEVD-AMC cleavage assay(Nicholson et al, 1995). AMC release was found to be verylow in two experiments with triplicate determinations.Although not statistically significant, the amount found inionomycin-stimulated platelets (1·65 6 0·95 pmol/min/109
platelets) was twice that observed in non-stimulated platelets(0·80 6 0·29 pmol/min/109 platelets). As expected, eventhese low levels of AMC release were reduced to backgroundlevel by 100 nM DEVD-CHO (aldehyde) which selectivelyinhibits caspase-3-like activity (Table IV).
DISCUSSION
Here we report that platelets from human donors expressedbcl-2, baxa and p53 at mRNA level. We also demonstrated thatplatelets contain Bcl-2 and Bax proteins and that the Bcl-2/Bax ratio was altered when platelets were stimulated withionomycin. An elevation of Ca2þ level is linked to plateletactivation and vesiculation in vitro (Holme et al, 1994;Katopodis et al, 1997). Since stimulated platelets also exhibitvolume shrinkage, PS exposure, vesiculation and cell frag-mentation (Bode et al, 1991; Dachary-Pringent et al, 1993;Fox et al, 1987), we propose that they undergo a process whichresembles apoptosis in nucleated cells and that this processincludes changes in the levels of apoptosis-related proteins.
The protein content in platelets is due to proteins that havebeen synthesized in the megakaryocyte, acquired byendocytosis or synthesized in the platelets (Harrison &Cramer, 1993). Our data indicate an increased level of Bcl-2
and Bax after ionomycin-induced activation of platelets.Others have demonstrated synthesis of GM3 ganglioside(Ferroni et al, 1997), HLA class I molecules (Santoso et al,1993) and other platelet proteins (Newman et al, 1988).However, increased protein synthesis will not necessarily bea general response to the activation of platelets. Since diversestimuli result in various degrees of activation, it is alsoprobable that protein synthesis will vary depending on thetype and intensity of stimulation. It has been suggested thatthere may be a higher degree of protein synthesis in newlyformed platelets (Santoso et al, 1993) and other circum-stances, such as platelet heterogeneity, mRNA content, stressand storage conditions, will affect their functional response(Thompson & Jakubowsky, 1988).
Changes in Bcl-2 and Bax protein levels were inhibited bythe calpain inhibitor calpeptin, but not by a broad-spectrumcaspase inhibitor. Calpeptin, but not VAD-cmk, also inhibitedcell shrinkage. However, neither calpeptin nor VAD-cmkaffected the decreased Bcl-2/Bax ratio and the PS exposure.Previous studies have shown that calpain activation cleavescytoskeletal proteins and participates in microparticleformation, hence it is tempting to correlate these events(Fox et al, 1990, 1991). However, studies of PS exposurequantified by annexin binding clearly showed that calpainactivity was not essential for PS exposure, and that PSexposure was not necessarily associated with microparticleformation (Dachary-Prigent et al, 1993). Our results alsoindicate that protease activity was not necessary for PSexposure on platelets.
Calpains are abundant in platelets, and they are activatedwhen platelets are activated by a physiological stimulus or bycalcium ionophore (Kelton et al, 1992; Ariyoshi et al, 1993).In contrast, we found very low levels of caspase-3-likeactivity in a concentrated sample of ionomycin-activatedplatelets. We have shown previously in U937 cells treatedwith TNF that calpain inhibitors were moderately effective atinhibiting the cleavage of intact fodrin (240 kD) into 150 kDand 120 kD fragments. VAD-cmk, on the other hand, did notinhibit the cleavage of intact fodrin to the 150 kD fragmentbut appeared to prevent further breakdown of the 150–120 kD fragment (Vanags et al, 1996). Studies in neuronalapoptosis have demonstrated that a-fodrin can be cleaved bycalpain or caspases, suggesting that different proteasespredominate in various apoptotic systems (Nath et al, 1996).
Table IV. Caspase 3-like-activity in platelets.*
Stimulation Caspase-3-like activity Plus DEVD-CHO
None 0·80 6 0·29 0·11 6 0·09Ionomycin 1·65 6 0·95 0·19 6 0·04
* Expressed as pmol/min AMC released by DEVD-AMC cleavagefrom 109 platelets.
Table III. Effect of protease inhibitors on platelets.
Stimulation Bcl-2* Bax* RVV clotting time†
None 25·5 6 7·3 24·1 6 5·9 111 6 22Ionomycin 105·4 6 36·8‡ 170·8 6 50·4‡ 55 6 5‡Ionomycin plus calpeptin 30·7 6 10·3 51·1 6 18·1 54 6 3‡Ionomycin plus VAD-cmk 92·4 6 29·6‡ 120·7 6 35·4‡ 48 6 2‡
Data are mean 6 SEM from five experiments with different donors.
* Specific MFI.† Inversely proportional to the increase in PS exposure.‡ P<0·05 as compared to non-stimulated controls.

Our finding that caspase-3-like enzymes did not appear toplay a significant role in the generation of apoptosis-likealterations in activated platelets was not surprising sinceplatelets are a good source of active calpain and hencecalpain would be expected to predominate. This is in linewith the possibility that a distinct proteolytic machineryoperates to execute the apoptotic process in different celltypes (Vanags et al, 1996; Xiang et al, 1996).
The Bcl-2/Bax ratio determines the amount of Bcl-2/Baxheterodimers versus Bax/Bax homodimers and, hence, thesusceptibility to apoptosis (Yang & Korsmeyer, 1996; Xianget al, 1996). Bcl-2 gene family expression seems to follow adynamic process that correlates with lineage, stage ofdifferentiation and cell activation. Alterations in the Bcl-2/Bax ratio in platelets after ionomycin stimulation are inagreement with bcl-2 and bax gene regulation accompany-ing apoptosis of nucleated cells. Bcl-2/Bax ratio varies duringlymphocyte development (Gratiot-Deans et al, 1993), and itcan also be modulated by cell surface signalling (Osorio et al,1997). Bcl-2/Bax alterations and other events similar toapoptosis occurring in platelets in the absence of significantcaspase-like activity are also in line with the fact that Baxover-expression causes alterations in mitochondrial functionand cell death without the requirement for caspase-3-likeactivity (Xiang et al, 1996).
Platelet microparticle formation requires gross cytoskele-tal degradation, in association with calpain activation (Foxet al, 1993). Platelet microparticles appear as a consequenceof in vitro stimulation, in vitro ageing or frozen storage (Bodeet al, 1991; Owens, 1994). In vivo formation of plateletmicroparticles may contribute to disseminated intravascularcoagulation and thrombotic complications observed inpatients with heparin-induced thrombocytopenia(Warkentin et al, 1994). However, the physiological con-sequences of platelet microparticle formation are not entirelydefined (Owens, 1994; Holme et al, 1994).
Platelet vesicles can transport calpain and other factors inthe circulation, acting as vehicles to concentrate andpotentiate coagulation reactions on damaged vessels (Bloom,1990; Kelton et al, 1992; Pasquet et al, 1996). Platelet-derivedmicroparticles seem to have many of the biological capabilitiesof intact platelets (Owens, 1994). The presence of Bcl-2, Baxand other gene products in circulating platelets raises theinteresting possibility that intact platelets and platelet micro-particles can carry these proteins and release them at distantsites. However, further work is required to explore both thepotential use of platelets as a model for cytoplasmic apoptosisand as carriers of cell death regulatory proteins.
ACKNOWLEDGMENTS
This work was supported by funds from the Swedish Societyof Medicine, the Swedish Medical Research Council (ProjectNo. 03X-2471) and Karolinska Institutet. We are indebted toDr David Fergusson and Ms Janet Bloomfield for expertassistance with the EM photograph.
REFERENCES
Aguilar-Santelises, M., Rottenberg, M.E., Lewin, N., Mellstedt, H. &Jondal, M. (1996) Bcl-2, Bax and p53 expression in B-CLL inrelation to in vitro survival and clinical progression. InternationalJournal of Cancer (Predictive Oncology), 69, 114–119.
Ariyoshi, H., Shiba, E., Sakon, M., Kambayashi, J., Yoshida, K.,Kawashima, S. & Mori, T. (1993) Translocation of human plateletcalpain-I. Biochemistry and Molecular Biology International, 30,63–72.
Bloom, A.L. (1990) Physiology of blood coagulation. Haemostasis,20, (Suppl. 1), 14–29.
Bode, A.P., Orton, S.M., Frye, M.J. & Udis, B.J. (1991) Vesiculation ofplatelets during in vitro aging. Blood, 77, 887–895.
Cory, S. (1995) Regulation of lymphocyte survival by the Bcl-2 genefamily. Annual Review of Immunology, 13, 513–543.
Dachary-Prigent, J., Freyssinet, J.M., Pasquet, J.M., Carron, J.C. &Nurden, A.T. (1993) Annexin V as a probe of aminophospholipidexposure and platelet membrane vesiculation: a flow cytometrystudy showing a role for free sulfhydryl groups. Blood, 81, 2554–2565.
Djaffar, I., Vilette, D., Bray, P.F. & Rosa, J.P. (1991) Quantitativeisolation of RNA from human platelets. Thrombosis Research, 62,127–135.
Earnshaw, W.C. (1995) Apoptosis: lessons from in vitro systems.Trends in Cell Biology, 5, 217–220.
Fadok, V.A., Voelker, D.R., Campbell, P.A., Cohen, J.J., Bratton, D.L. &Henson, P.M. (1992) Exposure of phosphatidylserine on thesurface of apoptotic lymphocytes triggers specific recognition andremoval by macrophages. Journal of Immunology, 148, 2207–2216.
Ferroni, P., Lenti, L., Martini, F., Ciatti, F., Pontieri, G.M. & Gazzaniga,P.P. (1997) Ganglioside content of human platelets: differences inresting and activated platelets. Thrombosis and Haemostasis, 77,548–554.
Fox, J.E.B., Austin, C.D., Reynolds, C.C. & Steffen, P.K. (1991)Evidence that agonist-induced activation of calpain causes theshedding of procoagulant-containing microvesicles from themembrane of aggregating platelets. Journal of BiologicalChemistry, 266, 13289–13295.
Fox, J.E.B., Reynolds, C.C. & Austin, C.D. (1990) The role of calpainin stimulus–response coupling: evidence that calpain mediatesagonist-induced expression of procoagulant activity in platelets.Blood, 76, 2510–2519.
Fox, J.E.B., Reynolds, C.C., Morrow, J.S. & Phillips, D.R. (1987)Spectrin is associated with membrane-bound actin filaments inplatelets and is hydrolysed by the Ca2þ-dependent protease duringplatelet activation. Blood, 69, 537–545.
Fox, J.E.B., Taylor, R.G., Taffarel, M., Boyles, J.K. & Goll, D.E. (1993)Evidence that activation of platelet calpain is induced as aconsequence of binding of adhesive ligand to the integrin,glycoprotein IIb–IIIa. Journal of Cell Biology, 120, 1501–1507.
Gratiot-Deans, J., Ding, L., Turka, L.A. & Nunez, G. (1993) Bcl-2proto-oncogene expression during human T cell development.Journal of Immunology, 151, 83–91.
Hale, A.J., Smith, C.A., Sutherland, L.C., Stoneman, V.E.A.,Longthorne, V.L., Culhane, A.C. & Williams, G.T. (1996)Apoptosis: molecular regulation of cell death. European Journal ofBiochemistry, 236, 1–26.
Harrison, P. & Cramer, E.M. (1993) Platelet a-granules. BloodReviews, 7, 52–62.
Holme, P.A., Solum, N.O., Brosstad, F., Roger, M. & Abdelnoor, M.(1994) Demonstration of platelet-derived microvesicles in blood
q 1997 Blackwell Science Ltd, British Journal of Haematology 99: 824–831
830 D. M. Vanags, S. Orrenius and M. Aguilar-Santelises

831Bcl-2, Bax and p53 Expression in Platelets
q 1997 Blackwell Science Ltd, British Journal of Haematology 99: 824–831
from patients with activated coagulation and fibrinolysis using afiltration technique and Western blotting. Thrombosis andHaemostasis, 72, 666–671.
Jacobson, M.D., Burne, J.F. & Raff, M.C. (1994) Programmed celldeath and Bcl-2 protection in the absence of a nucleus. EMBOJournal, 13, 1899–1910.
Jeddi-Tehrani, M., Grunewald, J., Hodara, V., Andersson, R. &Wigzell, H. (1994) Nonrandom T-cell receptor Jb usage pattern inhuman CD4þ and CD8þ peripheral T cells. Human Immunology,40, 93–100.
Katopodis, J.N., Kolodny, L., Jy, W., Horstman, L.L., De Marchena,E.J., Tao, J.G., Haynes, D.H. & Ahn, Y.S. (1997) Plateletmicroparticles and calcium homeostasis in acute coronaryischemias. American Journal of Hematology, 54, 95–101.
Kelton, J.G., Warkentin, T.E., Hayward, C.P.M., Murphy, W.G. &Moore, J.C. (1992) Calpain activity in patients with thromboticthrombocytopenic purpura is associated with plateletmicroparticles. Blood, 80, 2246–2251.
Levine, A.J. (1997) p53, the cellular gatekeeper for growth anddivision. Cell, 88, 323–331.
Nath, R., Raser, K.J., Stafford, D., Hajimohammadreza, I., Posner, A.,Allen, H., Talanian, R.V., Yuen, P.W., Gilberstsen, R.B. & Wang,K.K.W. (1996) Non-erythroid a-spectrin breakdown by calpainand interleukin 1b-converting-enzyme-like protease(s) in apopto-tic cells: contributory roles of both protease families in neuronalapoptosis. Biochemical Journal, 319, 683–690.
Newman, P.J., Gorski, J., White, G.C., Gidwitz, S., Cretney, C.J. &Aster, R.H. (1988) Enzymatic amplification of platelet-specificmessenger RNA using the polymerase chain reaction. Journal ofClinical Investigation, 82, 739–743.
Nicholson, D.W., Ali, A., Thornberry, N.A., Vaillancourt, J.P., Ding,C.K., Gallant, M., Gareau, Y., Griffin, P.R., Labelle, M., Lazebnik,Y.A., Munday, N.A., Rayu, S.M., Smulson, M.E., Yamin, T.T., Yu,V.L. & Miller, D.K. (1995) Identification and inhibition of the ICE/CED-3 protease necessary for mammalian apoptosis. Nature, 376,37–43.
Oltvai, Z.N. & Korsmeyer, S.J. (1994) Checkpoints of dueling dimersfoil death wishes. Cell, 79, 189–192.
Oltvai, Z.N., Milliman, C.L. & Korsmeyer, S.J. (1993) Bcl-2heterodimerizes in vivo with a conserved homologue, Bax, thataccelerates programmed cell death. Cell, 74, 609–619.
Osorio, L.M., De Santiago, A., Aguilar-Santelises, M., Mellstedt, H. &Jondal, M. (1997) CD6 ligation modulates the Bcl-2/Bax ratio andprotects chronic lymphocytic leukemia B cells from apoptosisinduced by anti-IgM. Blood, 89, 2833–2841.
Owens, M.R. (1994) The role of platelet microparticles inhemostasis. Transfusion Medicine Reviews, 8, 37–44.
Pasquet, J.M., Toti, F., Nurden, A.T. & Dachary-Prigent, J. (1996)Procoagulant activity and active calpain in platelet-derivedmicroparticles. Thrombosis Research, 82, 509–522.
Santoso, S., Kalb, R., Kiefel, V. & Mueller-Eckhardt, C. (1993) Thepresence of messenger RNA for HLA class I in human platelets andits capability for protein biosynthesis. British Journal ofHaematology, 84, 451–456.
Schulze-Osthoff, K., Walczak, H., Droge, W. & Krammer, P.H. (1994)Cell nucleus and DNA fragmentation are not required forapoptosis. Journal of Cell Biology, 127, 15–20.
Sedlak, T.W., Oltvai, Z.N., Yang, E., Wang, K., Boise, L.H., Thompson,C.B. & Korsmeyer, S.J. (1995) Multiple Bcl-2 family membersdemonstrate selective dimerizations with Bax. Proceedings of theNational Academy of Sciences of the United States of America, 92,7834–7838.
Steller, H. (1995) Mechanisms and genes of cellular suicide. Science,267, 1445–1449.
Thompson, C.B. & Jakubowsky, J.A. (1988) The pathophysiology andclinical relevance of platelet heterogeneity. Blood, 72, 1–8.
Thornberry, N.A., Bull, H.G., Calaycay, J.R., Chapman, K.T., Howard,A.D., Kostura, M.J., Miller, D.K., Molineaux, S.M., Weidner, J.R.,Aunins, J., Elliston, K.O., Ayala, J.M., Casano, F.J., Chin, J., Ding,G.J.F., Egger, L.A., Gaffney, E.P., Limjuco, G., Palyha, O.C., Raju,S.M., Rolando, A.M., Salley, J.P., Yamin, T.T., Lee, T.D., Shively, J.E.,MacCross, M., Mumford, R.A., Schmidt, J.A. & Tocci, M.J. (1992)A novel heterodimeric cysteine protease is required for inter-leukin-1b processing in monocytes. Nature, 356, 768–774.
Vanags, D.M., Porn-Ares, M.I., Coppola, S., Burgess, D.H. & Orrenius,S. (1996) Protease involvement in fodrin cleavage and phospha-tidylserine exposure in apoptosis. Journal of Biological Chemistry,271, 31075–31085.
Warkentin, T.E., Hayward, C.P.M., Boshkov, L.K., Santos, A.V.,Sheppard, J.A.I., Bode, A.P. & Kelton, J.G. (1994) Sera frompatients with heparin-induced thrombocytopenia generate plate-let-derived microparticles with procoagulant activity: an explana-tion for the thrombotic complications of heparin-inducedthrombocytopenia. Blood, 84, 3691–3699.
Xiang, J., Chao, D.T. & Korsmeyer, S.J. (1996) Bax-induced cell deathmay not require interleukin 1b-converting enzyme-like proteases.Proceedings of the National Academy of Sciences of the United States ofAmerica, 93, 14559–14563.
Yang, E. & Korsmeyer, S.J. (1996) Molecular thanatopsis: a discourseon the BCL-2 family and cell death. Blood, 88, 386–401.
Yin, X.M., Oltvai, Z.N. & Korsmeyer, S.J. (1994) BH1 and BH2domains of Bcl-2 are required for inhibition of apoptosis andheterodimerization with Bax. Nature, 369, 321–323.




















