
Progressive pulmonary fibrosis is mediated by TGF-beta isoform 1 but not TGF-beta3
Upload
independentCategory
view
0download
0
~ - ~:i:~",~l
E L S E V I E R Clinica Chimiea Acta 235 (1995) 11-31
Active and acid-activatable TGF- /in human sera, platelets and plasma
David J. Grainger a, David E. Mosedale a, James C. Metcalfe *a, Peter L. Weissberg b, Paul R. Kemp a
aDepartment of Biochemistry, University of Cambridge, Tennis Court Road, Cambridge, CB2 1QW, UK
bSchool of Clinical Medicine, University of Cambridge, Hills Road, Cambridge, CB2 2QQ, UK
Received 3 August 1994; revision received 28 October 1994; accepted 11 November 1994
Abstract
Assays; which measure active and latent forms of transforming growth factor/~ (TGF-~) separately in human serum and plasma are required to investigate the biological role of TGF-/~ in a variety of human diseases. We have developed an enzyme-linked immunosorbent assay (ELISA) using two polyclonal antibodies against TGF-/~ which rapidly determines the amount of active plus acid-activatable, latent TGF-/~ forms ((a+I)TGF-/~) present in human serum and plasma in the range 4 pmolfl to 2000 pmol/I. To measure active TGF-~ alone, we have developed a second ELISA using the extracellular domain of the TGF-/~ type II receptor as the capture reagent which detects active TGF-/3 in serum and plasma samples in the range 20 pmol/1 to 4000 pmol/l. Both assays detect TGF-/~I and TGF-/~3 with similar sensitivity, are > 10-foht less sensitive to TGF-/~2 and are not affected by a range of other peptide growth factors. "De mean (a+I)TGF-/~ present in human serum was 330 pmol/l but the range was very large ( < 4 pmol/l to 1400 pmol/l). The mean active TGF-/~ present was 230 pmol/l (range <20 pmol/l to 1400 pmol/l) and the proportion of the (a+I)TGF-/~ present which was active [a/(a+l)] varied from < 10% to 100%. The concentration of (a+I)TGF-~ and the proportion of TGF-/3 which was active were very similar in the serum and platelet-poor plasma prepared from the same whole blood sample. The clot formed during serum preparation retained all of the TGF-/~ which was detected by the (a+I)TGF4/ELISA in the corresponding platelet releasate, although the PDGF in platelets was released into the serum. In contrast, platelet- poor plaLsma contained no detectable PDGF demonstrating that the (a+I)TGF-/~ assayed in the plasma was not due to platelet degranulation after bleeding. Serum active TGF-/~ and (a+I)TGF-/~ concenrations therefore provide a reliable estimate of these forms of TGF-/~ pre- sent in plasma.
* Corresponding author, Tel: +44 223 333633; Fax: +44 223 333636.
0009-8981/95/$09.50 © 1995 Elsevier Science B.V. All rights reserved SSDI 0009-8981 (94)05995-5

12 D.J. Grainger et al./ Clinica Chimica Acta 235 (1995) 11-31
Keywords: Active TGF-fl; Latent TGF-fl; ELISAs; Human Serum; Human platelets; Human plasma
1. Introduction
The transforming growth factor/3 (TGF-fl) superfamily consists of a large number of related peptides which have pleiotropic effects on a wide range of cell types both in vitro and in vivo (reviewed in Ref. [1]). For example, TGF-fl either inhibits or stimulates proliferation of various cell types depending on the experimental condi- tions [2-4]. TGF-fl is also a potent regulator of extracellular matrix production and differentiation in a variety of cell types, including osteoblasts [5] and vascular smooth muscle cells [6]. TGF-fl is involved in a variety of physiological and pathological processes, including wound healing [7], ulceration [8] and several fibrotic disease states, such as glomerular nephritis [9] and cirrhosis of the liver [10]. On a more speculative basis, a role for TGF-fl has been proposed in osteoporosis [11], breast carcinoma [12,13] and other types of cancer [14], and in the development of atherosclerosis [15-17] and restenosis after coronary angioplasty [15,17].
The archetypal member of the family, TGF-fll, is a disulphide-linked 25 kDa dimer of two 112 amino acid peptides. It is extremely highly conserved in primary sequence (identical in man, pig, monkey and cow and a single amino acid substitu- tion in the mouse protein) and has a distinctive structure known as the TGF-fl knot [18]. Two closely related gene products have been identified in man (TGF-fl2 and TGF-fl3) and the heterodimer TGF-fll.2 has been isolated from human platelets [19]. However, no distinct biological functions for the different isoforms, in vivo or in vitro, have been reported. TGF-fls are synthesised as precursor latent forms [20] which do not bind to TGF-fl receptors and have no known biological activity. The precursor form can be cleaved by the serine protease, plasmin [15,21,22], and activa- tion of TGF-fl occurs by dissociation of the 25 kDa dimer from the rest of the com- plex (the latency associated peptide; LAP). Latent forms of the TGF-/3s composed of the mature TGF-fl dimer, LAP and the latent TGF-fl binding protein (LTBP) have also been characterised [34]. These forms of latent TGF-fl are denoted as large latent TGF-fl complexes whereas latent TGF-fls composed only of the mature pep- tide and LAPs are denoted as small latent TGF-fl complexes.
Analysis of the involvement of TGF-fl in a range of pathologies would be aided by the development of assays of the concentrations of both active and latent TGF-/3 forms rapidly in serum, plasma and other biological fluids. Active TGF-~ has previously been assayed in biological fluids using a receptor binding assay, in which the TGF-fl in the sample competes with a known amount of radiolabelled TGF-fl for binding to cells expressing TGF-fl receptors [23]. Although these assays are ac- curate, they are time-consuming and depend on a source of radiolabelled TGF-fl. Active TGF-fl has also been bioassayed by the inhibition it causes of DNA synthesis in mink lung (MvLu) epithelial cells [24]. However, we have found that this bioassay is unsuitable for measuring active TGF-fl in human sera or plasma because both in-

D.J. Grainger et aL / Clinica Chimica Acta 235 (1995) 11-31 13
hibited IVlvLu cell DNA synthesis by a mechanism independent of the TGF-/~ they contained.
In contrast to bioassays, enzyme-linked immunosorbent assays (ELISAs) are potentiallly well suited to assays of large numbers of samples in population studies (for exarnple, see Ref. [25]). We have utilised commercially available polyclonal an- tisera which do not distinguish between the active (a) and acid-activatable, latent (I)TGF-/3 forms to measure (a+I)TGF-/~ in biological fluids. To measure active TGF- /~ we haw~ used the extracellular domain of the TGF-/3 type II receptor, which binds only active TGF-/~, as the capture reagent for a second ELISA. Both the active and (a+I)TGF-/~ ELISAs utilise the same detection antibody; they measure TGF-/31 and TGF-#3 with equal sensitivity, but detect TGF-/~2 at least 10-fold less sensitively. The assays can therefore be used together to estimate the proportion [a/(a+l)] of TGF-# which is active in biological fluids.
We have begun to characterise active and (a+I)TGF-B in the blood of volunteer donors. TGF-/~I, predominantly in the latent form, has been identified previously in human platelets [20]. We have investigated whether the (I)TGF-/~ stored in platelets is released when blood is allowed to clot in vitro and whether active and (a+I)TGF-/~ are present in serum and platelet-poor plasma.
2. Materials and methods
2.1. Materials The antibodies to TGF-B used for the ELISAs were BDA19 (a chicken polyclonal
IgY antibody which neutralises TGF-/3 activity) and BDA47 (an affinity purified rabbit polyclonal IgG antibody), both obtained from R&D Systems (Oxford, UK). Goat anti-rabbit IgG coupled to horseradish peroxidase was obtained from Sigma Chemical Co. (Poole, UK). Standards were obtained of purified porcine TGF-~I (Peninsula, St. Helens, UK) and of recombinant human TGF-BI (Amersham Inter- national and R&D systems). To refer the ELISA data obtained with these TGF-~ls to the interim international standard, bovine TGF-/~I (89/516) was obtained from the National Institute of Biological Standards and Control (Potters Bar, UK). TGF- /32 and "I?GF-/~3 isoforms were obtained from R&D systems. The TGF-/3 standards were dissolved in 25 mmol/1 Tris/HCl pH 7.4 containing 50/zg/ml fatty acid free bovine serum albumin (FAF-BSA) to give 5 izg/ml stock solutions. The concentra- tion of the standard TGF-/~ solutions was checked against the bioassay of DNA syn- thesis in MvLu epithelial cells ([24]; see below). Both TGF-~ standards gave an EDs0 for inhibition of DNA synthesis in the MvLu bioassay of between 2 and 3 pmol/1 which agrees well with the previously reported value of 2 pmol/1 [24].
Platelet-derived growth factor (PDGF) AA and BB homodimers and epidermal growth factor (Bachem Inc., Saffron Walden, UK) were dissolved in 25 mmol/l Tris/HCl (pH 7.4) containing 1% FAF-BSA to give 0.3 ~mol/l stock solutions. Basic fibroblast growth factor (0.56 #mol/1) interleukin 1/3 (0.59 /~mol/1), transforming growth factor ct (1.8 1/~mol/l), interferon 7 (0.59/~mol/1) and insulin-like growth fac- tor I (0.59 ttmol/1; all from Bachem Inc.) were dissolved in sterile MilliQ water to give stock solutions of the concentrations indicated. Angiotensin II and endothelin

14 D.J. Grainger et al. / Clinica Chimica ,4cta 235 (1995) 11-31
I (Sigma Chemical Co.) were dissolved in sterile MilliQ water to give 10 ttmol/1 stock solutions.
2.2. Cloning and expression of the extracellular domain of the TGF-13 type H receptor The extracellular domain of the TGF-~ type II receptor was amplified from the
vector H2 3FF [26] using the polymerase chain reaction (PCR). The vector DNA was linearised with Not I, precipitated and resuspended at 10 ng//~l. Amplification was carried out in a 50/zl reaction containing 2.5/~1 DNA, 5/zl 10 x TAQ buffer (LKB Pharmacia; Uppsala, Sweden), 250 ng of each oligonucleotide primer (GAAT- T C C C A T G G G T C ~ T G C T C and GAATrCGTCAGGATrGCTGGTGTT; Wellcome Protein and Nucleic Acid Chemistry Facility, University of Cambridge), I U TAQ polymerase and a mixture of dATP, dTTP, dCTP and dGTP to give a final concentration of 200 #mol/l for each nucleotide. The sample was overlaid with 50 #1 paraffin oil. The reaction was carried out using a thermal cycler (PREM; Cambridge, UK) for 30 cycles (denaturing at 94°C for 1 min, annealing at 55°C for 2 min, elongation at 72°C for 2 min). The 450 bp fragment produced was purified by electrophoresis in low gel temperature agarose, digested with EcoRI and cloned into the glutathione-S-transferase fusion vector pGEX 2T (LKB Pharmacia). Vectors carrying inserts in the required orientation were identified by plasmid map- ping. The sequence of the insert was checked by subcloning the 450 bp EcoRI frag- ment from the chosen clone (pGT1 C) into Bluescript KS+ followed by double strand sequencing. The sequence showed a single base change (C to A at position +13 from the initiation codon) compared with the published sequence [26] which introduces a Leu to Met mutation in the protein.
2.3. Purification of the fusion protein An overnight culture of Escherichia coli TG 1 containing pGT 1C was diluted 1:100
into fresh 2YT medium (500 ml) containing 270/~mol/1 ampicillin and grown to an ODr00 of 0.5. Production of the fusion protein was induced by addition of 1 mmol/l isopropylthiogalactoside and the cells were harvested 5 h later by centrifugation. The bacteria were resuspended in 50 ml phosphate-buffered saline (PBS; 150 mmol/l NaCI, 2 mmol/1 Na2HPO4, 4 mmol/1 Na2HPO4, pH 7.3) containing 1% Triton X-100 and 1 mmol/1 PMSF and lysed by sonication for 5 min. The lysate was centri- fuged (10 000 x g; 5 min) and the fusion protein was purified from the supernatant by the one-step purification method of Smith and Johnson [27]. FPLC of the purified glutathione-binding proteins on a Superdex 200 HR column in 20 mmol/1 ammonium bicarbonate (pH 8.0) demonstrated that > 95% of the protein present was the desired 43 kDa TGF-fl receptor fusion protein.
2.4. ELISA to measure (a+I)TGF-f3 Maxisorp 96-well ELISA plates (Gibco; Uxbridge, UK) were coated with the cap-
ture antibody by incubating with 50/~1 BDA19 anti-TGF-/~ chicken IgY (40/zg/ml) diluted in Tris-buffered saline (TBS; 137 mmol/l NaCI, 50 mmol/l Tris/HCl, pH 7.4) and shaking the plates until dry by evaporation at room temperature (approximately 12 h). The plates were washed 3 x 3 min with PBS, blocked with 350/~1 3% FAF-

D.J. Grainger et al./ Clinica Chimica Acta 235 (1995) 11-31 15
BSA in TBS for 1 h, washed 3 x 3 min with TBS and incubated for 2 h with 100 t~l of test samples or dilutions of a TGF-/3 stock solution for calibration. The stan- dard TGF-~I stock solutions were diluted in TBS to concentrations between 0.4 pmol/l and 4000 pmol/l: purified porcine TGF-/3 was used for calibration except where indicated. The plates were washed (3 x 3 min) with TBS + 3% FAF- BSA + 0.1% Triton X-100 (wash buffer) and incubated with 20 #1 detection antibody (BDA47; anti-TGF-/3 rabbit IgG) at 1/xg/ml in wash buffer for 1 h. The plates were rinsed with wash buffer (3 x 3 min) and incubated with an antibody against rabbit IgG conjugated to horseradish peroxidase (Sigma A-6154) at 1:2500 dilution in wash buffer for 1 h. After washing (3 x 3 min with wash buffer) the plates were incubated for 15 rain with the chromogenic substrate orthophenylenediamine (Sigma) accor- ding to the manufacturer's instructions. The reaction was stopped by addition of an equal volume of 3 mol/l HCI and the absorbances were read on an ELISA plate reader (Titertek Multiscan; Flow Laboratories, High Wycombe, UK) within 15 min of stopping the reaction. Absorbances were converted into quantities of TGF-/~ pro- tein using the calibration curve from the TGF-/3 standard.
2.5. ELISA to measure active TGF-B This ELISA was performed as for the ELISA to assay (a+I)TGF-~ except: (i) the
ELISA ]plates were coated with the purified TGF-/3 receptor fusion protein using 20 t~l of a 50 /zg protein per ml solution in TBS and (ii) the detection reagent (BDA47) was used at 5 ~g/ml.
2.6. Mink lung epithelial cell DNA synthesis bioassay for active TGF-O A modification of the method described by Danielpour et al. [24] was used.
Briefly, the mink lung epithelial cells (MvLu; American Type Culture Collection; passage 49-60) were subcultured at 1:5 dilution in DMEM + 10% FCS. After 24 h the med!ium was replaced with DMEM + 10% FCS containing the test sample (< 1% v/v) or purified porcine TGF-/~ standards diluted in TBS, in the presence and absence of neutralising antiserum to TGF-/3 (BDA19) at 10/~g/ml. DNA synthesis was determined by a 1 h pulse of 6-[3H]thymidine (5 #Ci/ml; Amersham Interna- tional) added 23 h after the medium containing the test sample or TGF-/3 standard. TGF-O activity was calculated from the inhibition of DNA synthesis which was reversed by TGF-~ neutralising antibody, using a calibration curve obtained with dilutions of the TGF-~ standard to convert the inhibition of DNA synthesis into concentrations of active TGF-/3.
2. 7. Preparation of conditioned culture medium, human platelets, platelet-poor plasma and serum
Medium (DMEM + 20% FCS) was conditioned for 24 h on cultures of adult human aortic vascular smooth muscle cells (VSMCs) obtained by enzymatic disper- sion of aortic media as described previously [16].
Peripheral venous blood (20 ml) was collected from male volunteers (aged 23-54); 10 ml were aliquoted immediately into tubes containing 1.1 ml of sterile 3.8% (w/v) trisodium citrate in MilliQ water at room temperature. The samples were centrifuged

16 D.J. Grainger et al./ Clinica Chimica Acta 235 (1995) 11-31
(250 x g; 15 min) to remove red blood cells. Apyrase (Sigma) was added to the platelet-rich plasma to a final concentration of 100 mg/1 to prevent platelet degranulation; PMSF (1 mmol/l) and aprotinin (1 mg/1) were added to prevent pro- teolytic activation or degradation of TGF-/~. These samples were centrifuged (700 x g; 15 min) and the supernatant platelet-poor plasma was separated from the platelet pellet. The platelet-poor plasma was kept at room temperature until assayed by ELISAs within 2 h of preparation or was stored in 0.5-ml aliquots at -80°C. The platelet pellets were resuspended in l0 ml (i.e. the original volume of blood) of a buf- fered saline solution (145 mmol/l NaC1, 5 mmol/1 KCI, 10 mmol/l glucose, 10 mmol/l MgSO4, 0.5 mmol/l EGTA, 1 mmol/l PMSF, 1 mg/l aprotinin, l0 mmol/1 HEPES, pH 7.4) and re-centrifuged as before. The washed platelet pellet was resuspended in 10 ml of buffered saline solution and the platelet concentration was determined by haemocytometer. Platelets were lysed by ultrasonication until < 10% of unlysed platelets were detected by haemocytometer. Human platelet suspensions were also obtained from the Blood Transfusion Service, Cambridge, UK. The platelets were collected by centrifugation (3000 × g; 3 min) and approximately 0. l g of platelets was resuspended in 0.5 ml MilliQ water and lysed by three cycles of freeze-thawing. The membrane fragments were removed by centrifugation (14 000 x g; l0 min) and the supernatant was mixed with an equal volume of 2 x TBS.
The remaining l0 ml of freshly drawn blood samples were dispensed immediately into polypropylene tubes and allowed to clot at room temperature for 2 h. The clot- ted samples were centrifuged (1000 x g; 4 min), the serum was removed and either stored on ice until assayed within 2 h or stored at -80°C until assayed. The clot was washed three times by centrifugation (1000 × g; 4 min) in 5 ml of 150 mmol/l phos- phate buffer (pH 7.0) and the third wash was retained for TGF-/~ assays. The washed clot was dissolved in 5 ml of 150 mmol/l phosphate buffer (pH 2.0) for 30 min, then neutralised by addition of 5 ml of 150 mmol/l phosphate buffer (pH 12.0). The sam- ples were assayed for TGF-/3 immediately or stored in 1-ml aliquots at -80°C.
All blood-derived samples were stored at -80°C and were not thawed until assayed. The initial freeze-thaw cycle resulted in less than 10% loss of active or (a+I)TGF-B activity in the ELISAs. However, three additional freeze-thaw cycles of samples containing TGF-B in active or acid-activatable, latent forms were sufficient to cause loss of approximately 90% activity.
2.8. Bioassays of PDGF PDGF was bioassayed by its mitogenic activity on human VSMCs derived by
explant as described previously [28]. VSMCs were made quiescent by incubation in serum-free DMEM for 48 h. Samples of serum or platelet-poor plasma were added at a final concentration in DMEM of 5% or 20%, respectively. DNA synthesis was assayed by [3H]thymidine (Amersham International; 5 /zCi/ml) incorporation between 12 h and 36 h after addition of the test samples to the cells. The proportion of DNA synthesis due to PDGF was estimated by the addition of polyclonal antibody (50 mg/l) which neutralises all forms of PDGF [29] to replicate cell samples.
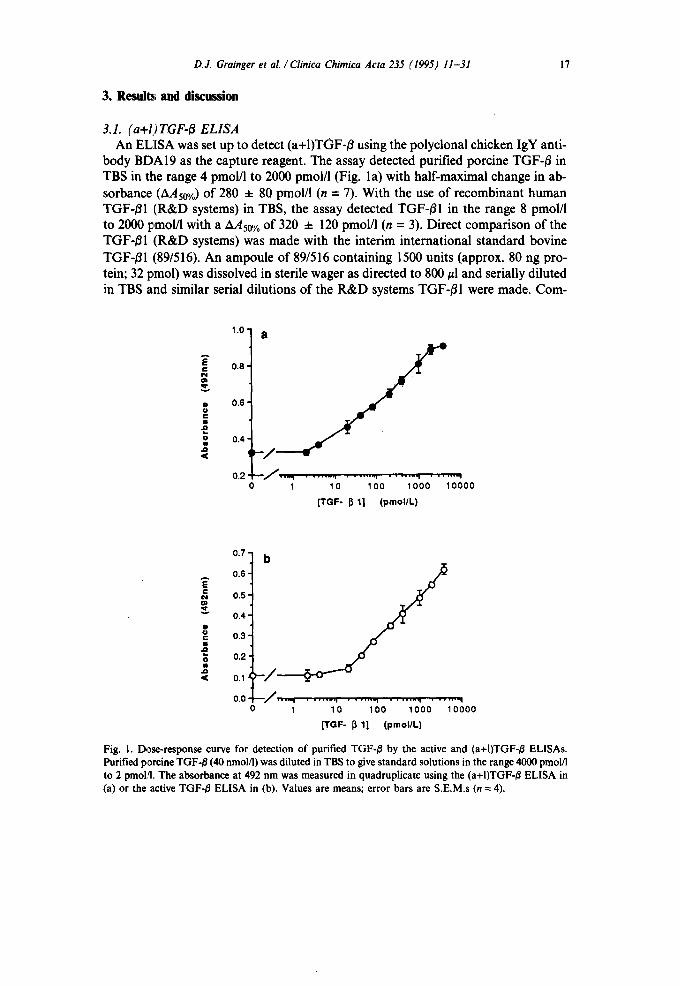
D.J. Grainger et al . / Clinica Chimica Acta 235 (1995) 11-31 17
3. Results and discussion
3,1. (a+l) TGF-~ ELISA An EL]SA was set up to detect (a+I)TGF-/3 using the polyclonal chicken IgY anti-
body BDA19 as the capture reagent• The assay detected purified porcine TGF-/3 in TBS in the range 4 pmol/l to 2000 pmol/l (Fig. la) with half-maximal change in ab- sorbance (AAs0%) of 280 ± 80 pmol/l (n = 7). With the use of recombinant human TGF-/51 (R&D systems) in TBS, the assay detected TGF-/31 in the range 8 pmol/l to 2000 pmol/1 with a AAs0 % of 320 ± 120 pmol/l (n = 3). Direct comparison of the TGF-/S1 (R&D systems) was made with the interim international standard bovine TGF-/31 (89/516). An ampoule of 89/516 containing 1500 units (approx. 80 ng pro- tein; 32 pmol) was dissolved in sterile wager as directed to 800/zl and serially diluted in TBS and similar serial dilutions of the R&D systems TGF-/31 were made. Com-
1 .0 -
E 0,8-
~ •
~ 0.6- c I I
o m
. o
a
0.4-
0.2 ~ • ---., o 1
. . . . . . I . . . . . . - 1 . . . . . . ,1 . . . . . . ,
10 1 O0 1000 10000
[TGF- ~ 1] (pmollL)
0.7
0.6 E c ¢~ 0.5
=., 0.4"
0
o 0.3 I
0.2 0 t
.Q
,< 0.1
0.0 1
. . . . . . • i . . . . . . - I . . . . . . . i . . . . . . . q
10 100 1000 10000
[TGF- ~ 1] (pmol/L)
Fig. 1. Dose-response curve for detection of purified TGF-/~ by the active and (a+I)TGF-/3 ELISAs. Purified porcine TGF-/~ (40 nmol/1) was diluted in TBS to give standard solutions in the range 4000 pmol/I to 2 pmoi,q. The absorbance at 492 nm was measured in quadruplicate using the (a+I)TGF-/5 ELISA in (a) or the active TGF-/5 ELISA in (b). Values are means; error bars are S.E.M.s (n = 4).

18 D.J. Grainger et al./ Clinica Chimica Acta 235 (1995) 11-31
parison of the calibration curves showed that a nominal 1.0 pmol of R&D TGF-BI had an activity of 130 ± 8 units.
To test the specificity of the capture antibody in the total TGF-B assay it was replaced with non-immune chicken IgY (R&D systems). The change in absorbance in the presence of 4000 pmol/l of purified-porcine TGF-/~I was less than 5%, in- dicating that TGF-B binding under the assay conditions was specific to the capture agent.
To test whether the ELISA detected acid activatable, latent forms of TGF-/3, a sample of human platelets from the blood bank was lysed and assayed before and after acid-activation of the TGF-B [20,30]. The latent TGF-B was converted to active TGF-B by addition of 5% v/v 150 mmol/l sodium phosphate buffer at pH 2.0 for 5 min, then neutralised by addition of 5% v/v 150 mmol/l sodium phosphate buffer at pH 12.0 [31]. Control samples were treated with 10% v/v 150 mmol/l sodium phos- phate buffer at pH 7.0. MvLu cell bioassay of the untreated and acid-treated platelet lysate showed that the amount of active TGF-B was increased 5. l-fold after acid acti- vation of the latent TGF-/3, indicating that approximately 80% of the TGF-~ present in the unactivated sample was in the acid-activatable, latent form. When assayed by the TGF-/~ ELISA, the control aliquot contained 680 4- 80 pmol/1 TGF-/~ (n = 3) and the acid-activated aliquot contained 600 4- 120 pmol/1 TGF-B (n = 3). These re- suits show that the (a+I)TGF-/3 ELISA does not distinguish between active and acid- activatable, latent TGF-/~ forms in human platelets.
The precise conditions for activation of the small and large complexes of latent TGF-B have not been characterised and there is some evidence for the existence of two pools of latent TGF-B which differ in the conditions required for activation. We therefore define (I)TGF-/~ as that pool of latent TGF-/3 which is acid-activatable by the treatment described above (i.e. exposure to pH 2.0 for 5 min before neutralisa- tion to pH 7.0 without overshoot). Longer exposure to pH 2.0 did not significantly affect the concentration of activated TGF-B and it remains to be determined which form(s) of latent TGF-/~ are activated under the defined conditions.
3.2. Active TGF-B ELISA A second ELISA was established to measure active TGF-B in the presence of
latent TGF-B using a truncated TGF-B type II receptor protein fused to glutathione- S-transferase as the capture reagent. This assay detected purified porcine TGF-BI in TBS in the range 20 pmol/1 to 4000 pmol/l (Fig. lb) with a AA50% of 680 ± 160 pmol/l (n = 4) and recombinant human TGF-B in TBS in the range 40 pmol/l to 4000 pmol/l with a AAso% of 720 ± 120 pmol/l (n = 3). To test the specificity of the trun- cated receptor fusion protein as the capture agent, it was replaced with glutathione- S-transferase. The change in absorbance in the presence of 4000 pmol/l of purified porcine TGF-fll was less than 5%, indicating that TGF-B binding was specific to the capture agent under the assay conditions.
To confirm that the active TGF-fl ELISA did not detect acid-activatable, latent TGF-fl, samples of human platelet TGF-fl before and after acid activation were assayed. The active TGF-fl ELISA gave 160 ± 40 pmol/l (n = 3) in the unactivated sample and 640 4- 80 pmol/1 (n = 3) TGF-B in the acid-activated sample, consistent
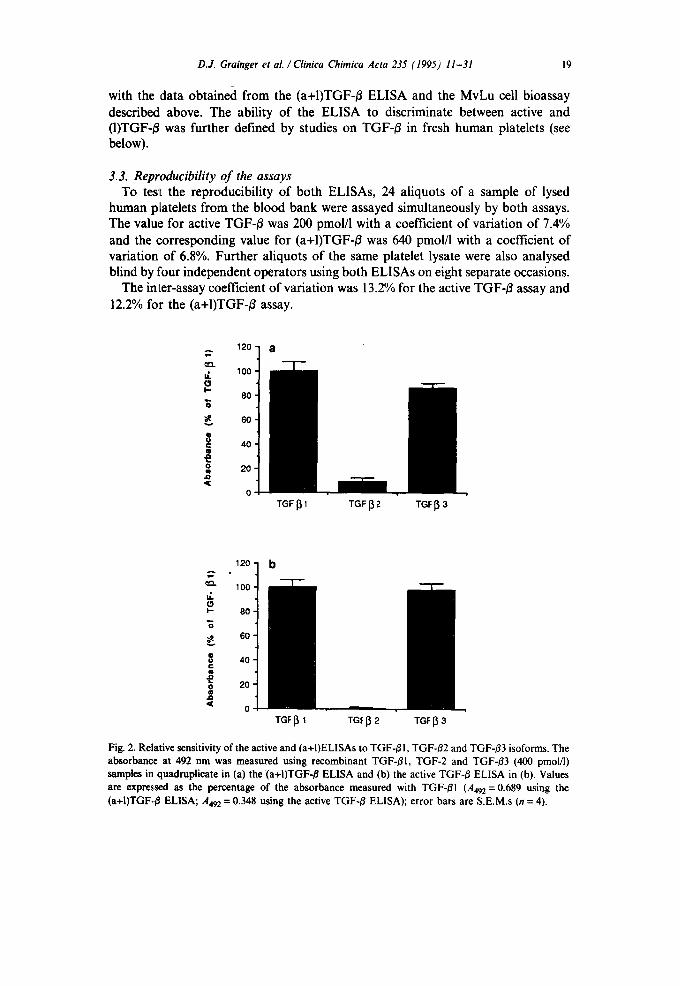
D.J. Grainger et al . / Clinica Chimica Acta 235 (1995) 11-31 19
with the ,data obtained from the (a+I)TGF-/3 ELISA and the MvLu cell bioassay described above. The ability of the ELISA to discriminate between active and (I)TGF-/~ was further defined by studies on TGF-B in fresh human platelets (see below).
3.3. Reproducibility of the assays To test the reproducibility of both ELISAs, 24 aliquots of a sample of lysed
human platelets from the blood bank were assayed simultaneously by both assays. The value for active TGF-~ was 200 pmol/l with a coefficient of variation of 7.4% and the corresponding value for (a+I)TGF-~ was 640 pmol/1 with a coefficient of variation of 6.8%. Further aliquots of the same platelet lysate were also analysed blind by four independent operators using both ELISAs on eight separate occasions.
The inter-assay coefficient of variation was 13.2% for the active TGF-/~ assay and 12.2% for the (a+I)TGF-/~ assay.
120 -
¢3. ,,, 100 -
0 I.-
8 0 -
~ 60- Q U ." 4 0 - u
~ 20-
0- TGF I~ 1 TGF 13 2 TGF 13 3
120
~ " 1 oo
(.-1 i.- 8o
0
g so
~ 4o c
g 2o
o TGF I~ 1 TGf 13 2 TGF I~ 3
Fig. 2. Relative sensitivity of the active and (a+I)ELISAs to TGF-/~I, TGF-~2 and TGF-/~3 isoforms. The absorbance at 492 nm was measured using recombinant TGF-/31, TGF-2 and TGF-~3 (400 pmol/l) samples in quadruplicate in (a) the (a+I)TGF-/~ ELISA and (b) the active TGF-/~ ELISA in (b). Values are expressed as the percentage of the absorbance measured with TGF-~I (A492 = 0.689 using the (a+I)TGF../3 ELISA; A492 = 0.348 using the active TGF-/~ ELISA); error bars are S.E.M.s (n = 4).
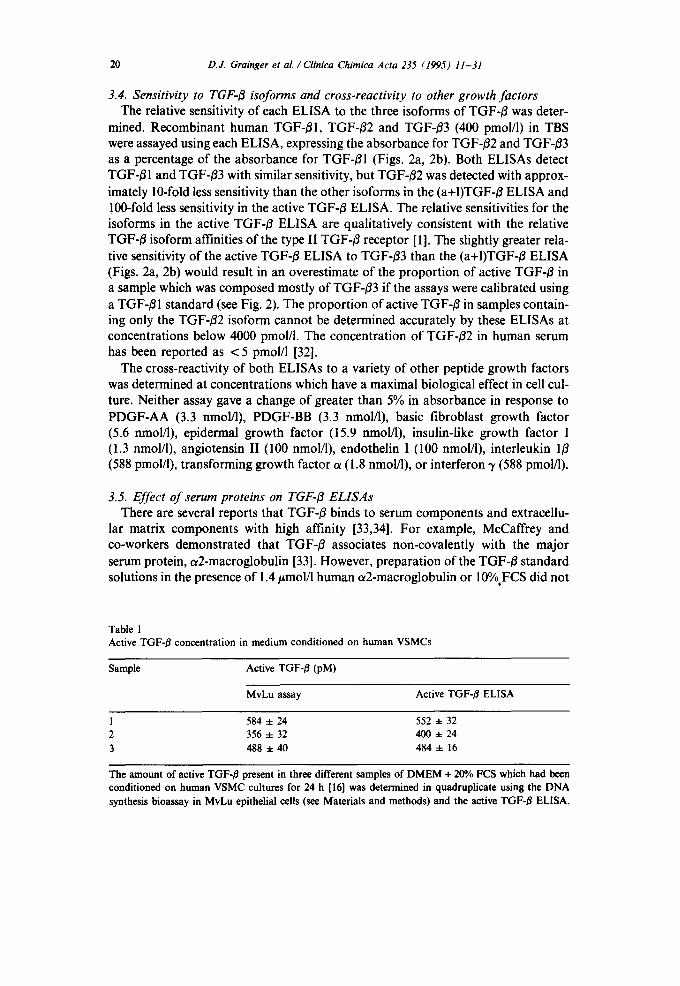
20 D.J. Grainger et aL /Clinica Chimica Acta 235 (1995) 11-31
3.4. Sensitivity to TGF-O isoforms and cross-reactivity to other growth factors The relative sensitivity of each ELISA to the three isoforms of TGF-B was deter-
mined. Recombinant human TGF-B1, TGF-B2 and TGF-B3 (400 pmol/1) in TBS were assayed using each ELISA, expressing the absorbance for TGF-/32 and TGF-B3 as a percentage of the absorbance for TGF-B1 (Figs. 2a, 2b). Both ELISAs detect TGF-/~I and TGF-O3 with similar sensitivity, but TGF-/~2 was detected with approx- imately 10-fold less sensitivity than the other isoforms in the (a+I)TGF-B ELISA and 100-fold less sensitivity in the active TGF-/~ ELISA. The relative sensitivities for the isoforms in the active TGF-B ELISA are qualitatively consistent with the relative TGF-B isoform affinities of the type II TGF-/3 receptor [1]. The slightly greater rela- tive sensitivity of the active TGF-O ELISA to TGF-#3 than the (a+I)TGF-/~ ELISA (Figs. 2a, 2b) would result in an overestimate of the proportion of active TGF-/3 in a sample which was composed mostly of TGF-/33 if the assays were calibrated using a TGF-Ol standard (see Fig. 2). The proportion of active TGF-~ in samples contain- ing only the TGF-B2 isoform cannot be determined accurately by these ELISAs at concentrations below 4000 pmol/l. The concentration of TGF-~2 in human serum has been reported as < 5 pmol/l [32].
The cross-reactivity of both ELISAs to a variety of other peptide growth factors was determined at concentrations which have a maximal biological effect in cell cul- ture. Neither assay gave a change of greater than 5% in absorbance in response to PDGF-AA (3.3 nmol/l), PDGF-BB (3.3 nmol/1), basic fibroblast growth factor (5.6 nmol/1), epidermal growth factor (15.9 nmol/1), insulin-like growth factor I (1.3 nmol/l), angiotensin II (100 nmol/l), endothelin I (100 nmol/l), interleukin 1~ (588 pmol/l), transforming growth factor tx (1.8 nmol/l), or interferon 3' (588 pmol/l).
3.5. Effect o f serum proteins on TGF-~ E L I S A s There are several reports that TGF-/~ binds to serum components and extracellu-
lar matrix components with high affinity [33,341. For example, McCaffrey and co-workers demonstrated that TGF-~ associates non-covalently with the major serum protein, ot2-macroglobulin [33]. However, preparation of the TGF-B standard solutions in the presence of 1.4 #mol/l human ct2-macroglobulin or 10% FCS did not
Table 1 Active TGF-B concentration in medium conditioned on human VSMCs
Sample Active TGF-B (pM)
MvLu assay Active TGF-~ ELISA
1 584 ± 24 552 ± 32 2 356 ± 32 400 ± 24 3 488 ± 40 484 ± 16
The amount of active TGF-B present in three different samples of DMEM + 20% FCS which had been conditioned on human VSMC cultures for 24 h [16] was determined in quadruplicate using the DNA synthesis bioassay in MvLu epithelial cells (see Materials and methods) and the active TGF-B ELISA.

D.J. Grainger et al . / Clinica Chimica Acta 235 (1995) 11-31 21
affect the AAs0o/o by more than 10% compared with the AAso% for the standard TGF-/3 solutions diluted in TBS in either ELISA. We conclude that any non- covalent interactions formed between TGF-~ and tx2-macroglobulin or with com- ponents of FCS do not prevent active TGF-B from binding to the type II TGF-B receptor in the active TGF-¢/ELISA or to the capture antibody in the (a+I)TGF-B ELISA, nor do they inhibit binding by the detection antibody. It has been noted in a previous report that purified TGF-B and ot2-macroglobulin may not interact in the same way as endogenous serum TGF-~ and a2-macroglobulin [35].
The active TGF-/~ concentration was measured in three samples of medium (DMEM containing 10% FCS) conditioned for 24 h on human VSMCs which pro- duce active TGF-B [16,17]. The values obtained with the active TGF-B ELISA were compared with those obtained using the MvLu cell bioassay (Table 1). The results obtained by the two assays were not statistically different for any of the three sam- ples tested (P = 0.88, 0.48 and 0.99, Student's unpaired t-test). We conclude that the ELISA gives values for active TGF-B concentrations in conditioned medium which are closely consistent with the MvLu cell bioassay used previously. Where possible it is important to demonstrate consistency between the active TGF-/3 ELISA and the bioassay for conditioned media and other biological fluids. For example, it has recently been reported that direct addition of conditioned media to ELISA microwells can lead to inaccurate measurement of TGF-/~ for reasons that are not fully understood [36]. Protocols which activate and concentrate TGF-Bs to partially purify the samples and exchange the buffer were recommended [36].
Other factors which might interfere with the assays are any peroxidases present in serum which bind to the capture reagents. To test for peroxidases, the capture antibody in the (a+I)TGF-/3 assay was replaced with non-immune chicken IgY and the trun,:ated receptor fusion protein in the active TGF-/3 assay was replaced with glutathione-S-transferase, as described previously. The change in absorbance in either assay was less than 5% in the presence of either DMEM containing 10% FCS
Table 2 Active and, (a+I)TGF-/3 concentrations in human sera
Donor TGF-O(pmol/l)
Unactivated serum Acid-activated serum
Active (a+l) Active (a+l)
A < 40 240 240 240 B 120 120 120 120 C 200 320 320 320 D 240 240 240 240
Serum saraples from four male donors were assayed in a single experiment for active and (a+I)TGF-/~ by the ELISAs before and after acid activation as described in the text. All samples were assayed in quadruplicate.
22 D.J. Grainger et al. /Clinica Chimica Acta 235 (1995) 11-31
or human serum from donors A, E, K or N in Table 2. These data indicated that any peroxidase activity in FCS or human serum did not significantly affect the assays of active or (a+I)TGF-/3.
3.6. Assays of TGF-/3 in human serum The above experiments suggested that the ELISAs could be used to measure TGF-
13 in human serum and the use of the assays for sera was therefore characterised. It
E g~
0.5
.o <
A E Q.
z a
1
0.4
0.3
0.2
0.1 0
10000
8000
6000
4000
2000
0
. . . . . . i . . . . . . . . i 100 1 0 0 0
[TGF- ~ 1] (pmo l /L )
C o n t r o l
0.9
~ 0.8
c o,7
• 0.6 {:
o.s
.D 0.4
0.3 . . . . . . i . . . . . . . . i 100 1 0 0 0
[TGF- ~ 1] (pmo l /L )
+TGF +TGF + s e r u m + s e r u m
+ ant ibody + ant ibody
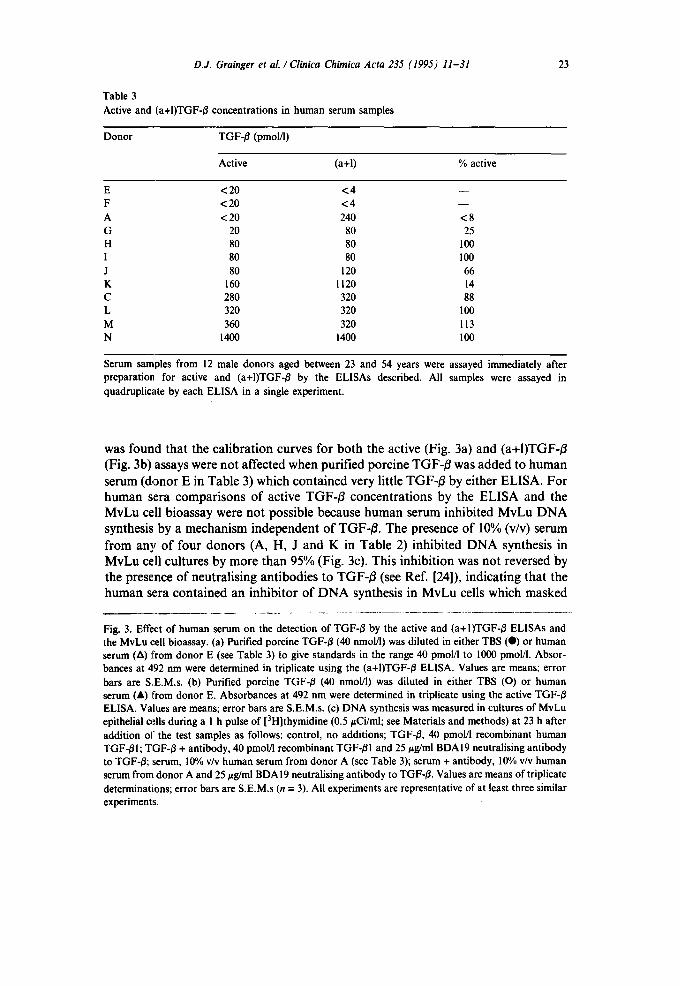
D.J. Grainger et al . / Clinica Chimica Acta 235 (1995) 11-31
Table 3 Active and (a+I)TGF-B concentrations in human serum samples
23
Donor TGF-B (pmol/l)
Active (a+l) % active
E <20 <4 - - F <20 <4 - - A < 20 240 < 8 G 20 80 25 H 80 80 100 1 80 80 100 J 80 120 66 K 160 1120 14 C 280 320 88 L 320 320 100 M 360 320 I 13 N 1400 1400 100
Serum samples from 12 male donors aged between 23 and 54 years were assayed immediately after preparation for active and (a+I)TGF-/3 by the ELISAs described. All samples were assayed in quadruplicate by each ELISA in a single experiment.
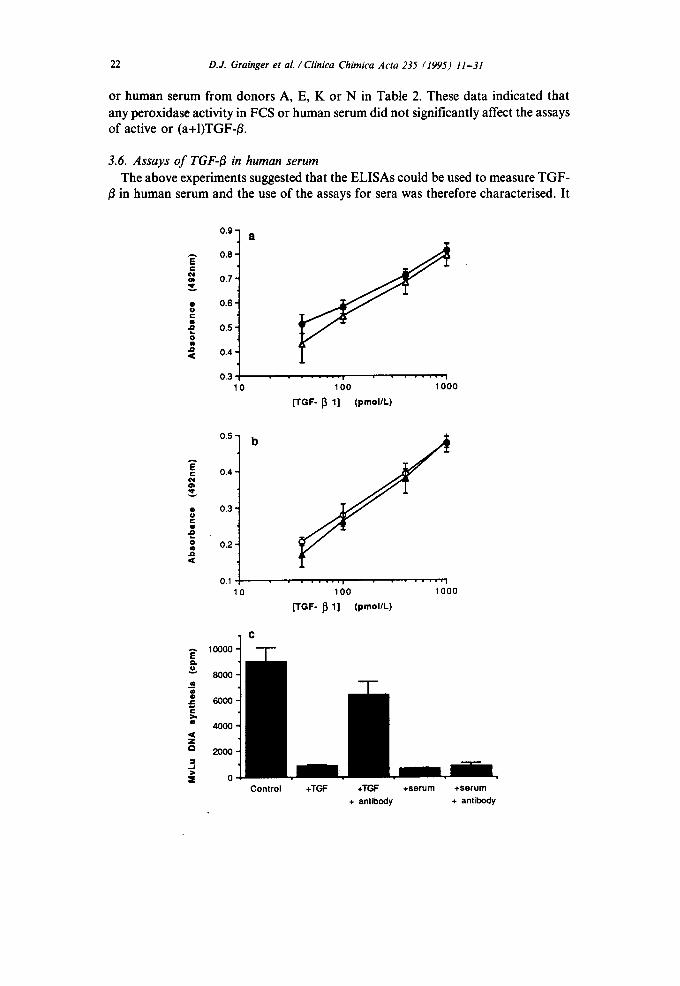
was found tha t the ca l i b r a t i on curves for b o t h the ac t ive (Fig. 3a) and ( a + I ) T G F - B
(Fig. 3b) assays were n o t a f fec ted w h e n pur i f ied po rc ine TGF- /~ was added to h u m a n
se rum ( d o n o r E in T a b l e 3 ) w h i c h c o n t a i n e d very lit t le TGF- /~ by e i ther E L I S A . F o r
h u m a n sera c o m p a r i s o n s o f ac t ive T G F - ~ c o n c e n t r a t i o n s by the E L I S A and the
M v L u cell b ioassay were no t poss ib le because h u m a n se rum inh ib i ted M v L u D N A
synthesis by a m e c h a n i s m i n d e p e n d e n t o f T G F - B . T h e presence o f 10% (v/v) se rum
f r o m any o f f o u r d o n o r s (A, H, J and K in T a b l e 2) inh ib i ted D N A synthesis in
M v L u cell cu l tures by m o r e t h a n 95% (Fig. 3c). Th is inh ib i t ion was no t reversed by
the presence o f neu t ra l i s ing an t ibod ie s to T G F - B (see Ref . [24]), ind ica t ing tha t the
h u m a n sera c o n t a i n e d an inh ib i to r o f D N A synthesis in M v L u cells which m a s k e d
. . . . . . . . . . . . . . . . . . . . . . . . . . . .
Fig. 3. Effect of human serum on the detection of TGF-B by the active and (a+I)TGF-B ELISAs and the MvLu cell bioassay. (a) Purified porcine TGF-B (40 nmolfl) was diluted in either TBS (0) or human serum (A) from donor E (see Table 3) to give standards in the range 40 pmol/I to 1000 pmol/l. Absor- bances at 492 nm were determined in triplicate using the (a+I)TGF-/~ ELISA. Values are means; error bars are S.E.M.s. (b) Purified porcine TGF-B (40 nmol/l) was diluted in either TBS (O) or human serum (A) from donor E. Absorbances at 492 nm were determined in triplicate using the active TGF-B ELISA. Values are means; error bars are S.E.M.s. (c) DNA synthesis was measured in cultures of MvLu epithelial oflls during a 1 h pulse of [3H]thymidine (0.5 #Ci/ml; see Materials and methods) at 23 h after addition of the test samples as follows: control, no additions; TGF-/~, 40 pmol/l recombinant human TGF-BI; TGF-B + antibody, 40 pmol/l recombinant TGF-BI and 25 #g/ml BDAI9 neutralising antibody to TGF-B; serum, 10% v/v human serum from donor A (see Table 3); serum + antibody, 10% v/v human serum from donor A and 25/zg/ml BDAI9 neutralising antibody to TGF-B. Values are means of triplicate determinations; error bars are S.E.M.s (n = 3). All experiments are representative of at least three similar experiments.

24 D.J. Grainger et al . / Clinica Chimica Acta 235 (1995) 11-31
any effect of TGF-/$. The MvLu cell bioassay cannot therefore be used to determine the concentration of active TGF-/~ in unfractionated 10% human serum samples.
Alternative approaches were therefore required to validate the ELISA assays for direct use with human serum. The main requirement was to determine whether human sera contain non-TGF-/3 components which significantly affect the TGF-~3 concentrations estimated by either assay. Overestimated values of TGF-/$ would be obtained if a serum component was bound specifically or non-specifically by the cap- ture agent in either assay and was also recognised by the detection antibody or by the antibody to rabbit IgG linked to horseradish peroxidase. Alternatively, under- estimated values would result if a serum component competed with TGF-~3 for the capture agent in either assay but was not recognised by the detection antibody. In a previous study in which TGF-/~ in unfractionated serum (after transient acidifica- tion) was determined by a radio-receptor assay, it was found that components in the serum interfered with the assay [35]. This resulted in a dilution curve which was not parallel to the standard dilution curve and estimates of TGF-~ were 20 to 40 times lower than those obtained by acid-ethanol extraction of the same samples. Thus it is possible that serum components which result in either overestimated or under- estimated TGF-/3 values in our ELISAs would also interfere with other assays (receptor binding or radioimmunoassays) used to validate serum TGF-~ concentra- tions estimated by the ELISAs. We therefore required a more rigorous test for in- terfering components in serum. This was achieved by determining whether the concentrations of active and (a+I)TGF-~ concentrations in sera were internally con- sistent before and after activation of (I)TGF-/3 by acid treatment. Only under very implausible circumstances would consistent accounting of active and (a+I)TGF-/3 be obtained in the presence of serum components which interfered with either or both assays.
ELISAs of active and (a+I)TGF-fl concentrations were performed on the sera from four male donors before and after the sera were acidified to pH 2.0 and neutralised to pH 7.0 as described for the lysed human platelet samples. For each of the sera in Table 2, there was no difference within the accuracy of the assays be- tween the amount of (a+I)TGF-fl before and after acid treatment. Furthermore, after acid treatment the amount of active TGF-fl was not significantly different from the amount of (a+I)TGF-fl. These results imply that it is very unlikely that the sera tested contained components which interfered with either TGF-~ ELISA since they would cause significant imbalances in the quantitative accounting of the amounts of active and (a+I)TGF-/$ before and after acid treatment. The use of acid treatment of the sera and re-assay of the active and (a+I)TGF-fl concentrations therefore pro- vides an important internal control for the TGF-fl assays when used directly for sera or complex biological fluids.
The sera from 12 male donors (aged 23-54 years) were assayed for active and (a+I)TGF-fl by the ELISAs (Table 3). The mean (a+I)TGF-fl concentration was 330 pmol/1, but the variation was very large (range less than 4 pmol/l to 1400 pmol/l). Similarly, the mean active TGF-fl concentration was 230 pmol/1 and the range was from less than 20 pmol/l to 1400 pmol/l. The proportion of the (a+I)TGF-fl present which was active ranged from < 10% to 100% with a mean of 73% for the samples
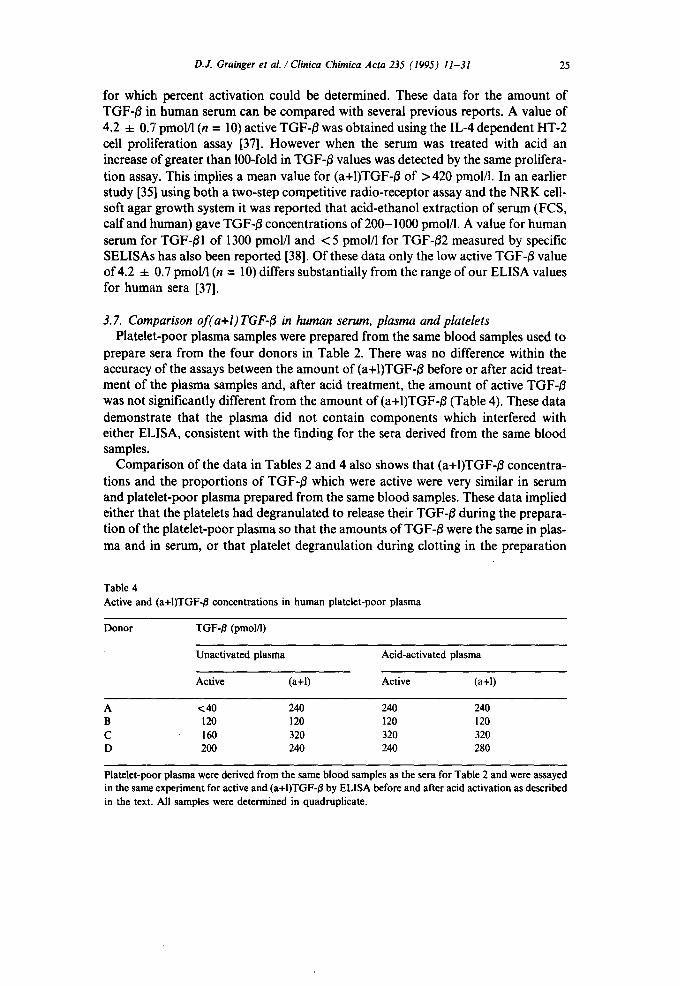
D.J. Grainger et al./ Clinica Chimica Acta 235 (1995) 11-31 25
for which percent activation could be determined. These data for the amount of TGF-B in, human serum can be compared with several previous reports. A value of 4,2 ± 0.7' pmol/1 (n = 10) active TGF-# was obtained using the IL-4 dependent HT-2 cell proliferation assay [37]. However when the serum was treated with acid an increase of greater than 100-fold in TGF-/3 values was detected by the same prolifera- tion assay. This implies a mean value for (a+I)TGF-B of > 420 pmol/1. In an earlier study [35] using both a two-step competitive radio-receptor assay and the N RK cell- soft agar growth system it was reported that acid-ethanol extraction of serum (FCS, calf and human) gave TGF-/3 concentrations of 200-1000 pmol/l. A value for human serum for TGF-BI of 1300 pmol/1 and < 5 pmol/1 for TGF-/~2 measured by specific SELISAs has also been reported [38]. Of these data only the low active TGF-B value of 4.2 ± 0.7 pmol/l (n = 10) differs substantially from the range of our ELISA values for human sera [37].
3. 7. Comparison of(a+l)TGF-I~ in human serum, plasma and platelets Platelet-poor plasma samples were prepared from the same blood samples used to
prepare sera from the four donors in Table 2. There was no difference within the accuracy of the assays between the amount of (a+I)TGF-/~ before or after acid treat- ment of the plasma samples and, after acid treatment, the amount of active TGF-/~ was not ,fignificantly different from the amount of (a+I)TGF-~ (Table 4). These data demonstrate that the plasma did not contain components which interfered with either ELISA, consistent with the finding for the sera derived from the same blood samples.
Comparison of the data in Tables 2 and 4 also shows that (a+I)TGF-~ concentra- tions and the proportions of TGF-/~ which were active were very similar in serum and platelet-poor plasma prepared from the same blood samples. These data implied either that the platelets had degranulated to release their TGF-/3 during the prepara- tion of the platelet-poor plasma so that the amounts of TGF-B were the same in plas- ma and in serum, or that platelet degranulation during clotting in the preparation
Table 4 Active and (a+I)TGF-B concentrations in human platelet-poor plasma
Donor TGF-/~ (pmol/l)
Unactivated plasma Acid-activated plasma
Active (a+l) Active (a+l)
A <40 240 240 240 B 120 120 120 120 C 160 320 320 320 D 200 240 240 280
Platelet-poor plasma were derived from the same blood samples as the sera for Table 2 and were assayed in the same experiment for active and (a+I)TGF-/~ by ELISA before and after acid activation as described in the text. All samples were determined in quadruplicate.

26 D.J. Grainger et al./ Clinica Chimica Acta 235 (1995) 11-31
Table 5 (a+I)TGF-# concentrations in human serum, plasma, platelets and acid-treated clots
Donor (a+I)TGF-/$ (pmol/l)
Serum Platelet-poor Platelets Acid-treated clot plasma
E <40 40 1000 960 N 80 80 880 760 B 120 120 1000 1200 D 280 280 1600 1600 A 320 360 1200 1200 C 440 440 1000 720 M 1200 1400 760 760
Serum, platelet-poor plasma and platelets were prepared from blood from seven male donors. Clots were removed from the serum samples by centrifugation, washed, dissolved by acidification and neutralised as described in Materials and methods. TGF-fl was released from platelets by sonication which lysed > 90% of the platelets present. The (a+I)TGF-fl in each sample was assayed by ELISA in quadruplicate. TGF*fl concentrations for platelets and clots are calculated for the volume of blood from which they were derived.
of serum did not release active or (I)TGF-/~ into the serum. The serum and plasma TGF-/~ concentrations would then be similar because the serum and plasma would not contain a significant amount of active or (I)TGF-/3 from platelets which had degranulated after drawing the blood samples.
To examine whether the active or (I)TGF-/$ in the serum and plasma samples was derived from degranulation of platelets after drawing blood, (a+I)TGF-/~ concentra- tions in the sera, acid-extracted clots, platelet-poor plasma and platelets from seven donors were compared (Table 5). The (a+I)TGF-~ concentrations in serum and plas- ma derived from the same blood samples were very similar, consistent with the data in Tables 2 and 4. The average concentration of (a+I)TGF-/$ from the degranulated platelet samples was 1063 pmol/1 and the average platelet concentration by haemocytometer in the platelet preparations was 3.0 x I01~/I, equivalent to an average Of 2100 molecules of TGF-fl per platelet. This may be compared with a pre- vious estimate of 500-2000 molecules of TGF-~ per platelet recovered from 'platelet secretate' [20]. However, the surprising observation was that the (a+I)TGF-~ con- centrations of the degranulated platelets and the acid-extracted clots derived from the same blood samples were very similar. This observation implies that any active or (I)TGF-/3 released by platelets which degranulated in the clots was almost entirely retained within the clot, since quantitative recovery of the (a+I)TGF-/~ was obtained from the clot after acid treatment. The retention of (a+I)TGF-/~ in the clot would account for the close similarity of the (a+I)TGF-/~ concentrations in the sera and plasma and this conclusion was tested further as described below. However, it should be emphasised that the data do not in any way preclude the possibility that platelets contain substantial amounts of latent TGF-/~ form(s) which are not

D.J. Grainger et al./ Clinica Chimica Acta 235 (1995) 11-31 27
detected by the (a+I)TGF-/~ ELISA because they are not activated by the defined acid-activation procedure.
No active TGF-/~ could be detected in the platelet releasate from freshly prepared platelets, unlike the TGF-/3 obtained from blood bank platelets. When active recom- binant htLrnan TGF-~I was added to the platelet releasate containing the highest con- centration of (a+I)TGF-/3 (1600 pmol/1 from donor D), the calibration curve for active TGF-/~ was superimposed on the curve for the recombinant human TGF-/31 in TBS (data not shown). These observations show that the selectivity of the active TGF-/~ assay is at least 50-fold greater for active TGF-31 than for (I)TGF-31.
The mean value for (a+I)TGF-/3 in platelet-poor plasma was 389 ± 177 pmol/l (n 7). Some of the.reported values for TGF-/~ in platelet-poor plasma are similar to those described here. In two separate studies using acid-ethanol extraction of platelet-poor plasma and the MvLu cell bioassay, TGF-3 concentrations of 212 ± 1:32 pmol/l (n = 9) [39] and 244 4- 40 pmol/1 (range >80 to <400 pmol/l; n = 10) 140] were recently reported. Previously, Wakefield et al. [41] reported that human plasma contains significant levels of TGF-3 (60 4- 24 pmol/l; n = 10) and concluded that latent TGF-/~ does circulate in normal individuals. One much lower value of 2.3 pmol/l (range 2.1-2.7 pmol/l; n = 9) for TGF-31 in platelet-poor plasma assayed by a TGF-/~I ELISA on acid-ethanol extracts has also been reported [42].
3.8. PDGF activity in plasma and serum
The similarity of both the active and (a+I)TGF-/~ concentrations in platelet-poor plasma and serum from the same donor (Tables 2, 4 and 5) prompted the question of whether the TGF-/~ had been released by a partial degranulation of platelets when the blood samples were drawn and before the onset of clot formation in the serum samples. Since PDGF is contained in the same platelet a-granules as latent TGF-3,
Table 6 Mitogenic indices of human serum and plasma on human vascular smooth muscle cells
Donor Mitogenic index
Serum Plasma
B 45 0.7 H 52 1.4 C 60 0.9 D 65 1.0 A 83 1.2
DMEM ccmtaining 5% serum or 20% platelet-poor plasma from five male donors was added to quiescent, explant-derived human smooth muscle cells and DNA synthesis was assayed in triplicate by incorporation of [3H]th~anidine between 12 h and 36 h after addition of the samples, as described in Materials and methods. The mitogenic indices are the ratios of 3H counts incorporated in the test cells samples to 3H counts in control cells treated with medium alone (1506 ± 123 counts/vain). The mitogenic indices for the plasma samples were unaffected by neutralising antiserum to PDGF but were reduced by more than 52% for each of the serum samples.

28 D.J. Grainger et al. /Clinica Chimica Acta 235 (1995) 11-31
a bioassay for PDGF activity as a mitogen for human VSMCs was used to determine the extent of platelet degranulation during the preparation of the platelet-poor plas- ma (Table 6).
Platelet-poor plasma had no significant mitogenic activity on human VSMCs measured as a ratio of [3H]thymidine incorporation in the presence or absence of plasma (Table 6) and the ratio was unaffected by neutralising antibody to PDGF. However, addition of 3.3 pmol/l PDGF to the plasma samples caused an increase in the average mitogenic index from 1.0 to 1.6 and this increase was blocked by neutralising PDGF antibody. The platelet-poor plasma samples therefore contained less than 3.3 pmol/1 of active PDGF. In contrast, the human serum samples gave large mitogenic indices of 45-83 for the same cell preparation and at least 52% of the mitogenic activity was reversed by neutralising antibody to PDGF (50 mg/1). This mitogenic activity attributable to PDGF is consistent with previous estimates that PDGF accounts for approximately 50% of platelet derived mitogenic activity of human serum, as assayed on glial cells or fibroblasts [43]. The mitogenic stimulation reversible by neutralising PDGF antibody (50 mg/l) in the serum samples cor- responds to concentrations of human PDGF of greater than 300 pmol/1 and less than 600 pmol/l in the human sera. This value may be compared with a reported concen- tration of PDGF in human serum of 500 pmol/1 by radio-receptor assay [44]. A serum concentration of greater than 300 pmol/l therefore implies degranulation of most of the platelets during clot formation to release PDGF into the serum under conditions in which the TGF-B remains associated with the clot. The undetectable PDGF activity in the plasma samples indicates that the amount of PDGF in the plas- ma corresponds to degranulation of less than 5% of the platelets after bleeding.
Most previous work has shown that normal human plasma contains undetectable levels of PDGF (reviewed in Ref. [45]). However, in one report [44] PDGF in human platelet-poor plasma was estimated at 33 pmol/1 by radio-receptor assay with a cor- responding serum concentration of 500 pmol/l. Thus the preparation of platelet-poor plasma containing little or no detectable PDGF from platelet degranulation during preparation in our experiments is consistent with previous data.
Taken together, these observations strongly imply (i) that the TGF-/3 in platelet- poor plasma and serum does not result from platelet degranulation which occurs on or after taking the blood samples and (ii) that the concentrations of (a+I)TGF-B in serum and plasma are very similar because platelet degranulation on clotting does not release (a+I)TGF-/3 into the serum which can be detected by the (a+I)TGF-B assay. We note that similar (a+I)TGFq3 concentrations in serum were obtained from repeated bleeds from the same donors. For example, donor A gave (a+I)TGF-~ con- centrations of 240, 240, 320, 240 and 280 pmol/l from five bleeds at intervals of at least 7 days. Furthermore, similar proportions of (a+I)TGF-~ were active in re- peated bleeds from the same donors. These observations are consistent with negligi- ble platelet degranulation after the blood samples are drawn since degranulation would be unlikely to be sufficiently controlled to yield reproducible amounts of (a+I)TGF-/3 in sera prepared from separate bleeds.
The data leave open the question of the origin of the TGF-/3 in platelet-poor plas- ma. It is generally assumed that the plasma TGF-B is mainly derived from platelets

D.J. Grainger et al . / Clinica Chimica Acta 235 (1995) 11-31 29
but this has not been demonstrated experimentally. However, the ELISAs described here should facilitate analysis of the mechanisms controlling platelet-poor plasma concentrations of active and (a+I)TGF-/~. They should also facilitate examination of correlations between TGF-/~ concentrations in plasma or serum and various diseases in which TGF-/3 may be implicated.
Acknowh~gements
We thank Mrs Christine Witchell and Dr. Thanasis Manousis for excellent techni- cal assistance. We are grateful to Herbert Lin (Whitehead Institute, Cambridge, USA) for providing a plasmid encoding the full length TGF-/3 type II receptor and Russell Ross (University of Washington, Seattle) for the neutralising antibody to PDGF. The work was supported by a British Heart Foundation group grant to J.C.M. and P.L.W.P.L.W. is a British Heart Foundation Senior Research Fellow. D.J.G. was the recipient of a Wellcome Prize Fellowship.
References
[1] Mass.agu6 J. The transforming growth factor-//family. Annu Rev Cell Biol 1990;6:597-641. [2] Pfeil,~chifter J, D'Sousa SM, Mundy GR. Effects of transforming growth factor-# on osteoblastic
osteosarcoma cells. Endocrinology 1987;121:212-218. [3] Weissberg PL, Grainger DJ, Metcalfe JC. Effects of TGF-//on vascular smooth muscle cell growth.
In: Cummins P, ed. Growth factors and the cardiovascular system. Dordrecht: Kluwer Acadmeic Publishers, 1993.
[4] Battegay EJ, Raines EW, Seifert RA, Bowen-Pope DF, Ross R. TGF-//induces bimodal prolifera- tion of connective tissue cells via complex control of an autocrine PDGF loop. Cell 1990;63: 515-.524.
lSl Noda M, Rodan GA. Type-# transforming growth factor (TGF-//) regulation of alkaline phospha- tase and other phenotype related mRNAs in osteoblastic rat osteosarcoma cells. J Cell Physiol 1987;133:426-437.
16] Bjorkerud S. Effects of transforming growth factor-//l on human arterial smooth muscle cells in vitro. Arterioscler Thromb 1991;11:892-902.
[7] Mustoe TA, Pierce GF, Thomason A, Gramates P, Sporn MB, Deuel TF. Transforming growth factor-//induces accelerated healing of incisional wounds in rats. Science 1987;237:1333-1336.
18] Connor TB, Roberts AB, Sporn MB, Danielpour D, Dart LL. Correlation of fibrosis and trans- forming growth factor-//type 2 in the eye. J Clin Invest 1989;83:1661-1666.
[9] Border WA, Rouslahti E. Transforming growth factor beta: the dark side of tissue repair. J Clin Invest 1991;90:1-7.
[10] Milani S, Herbst, H, Schuppan D, Stein H, Surrenti C. Transforming growth factors beta 1 and beta 2 are differentially expressed in fibrotic liver disease. Am J Pathol 1991;139:1221-1229.
[11] Finkelman RD, Bell NH, Strong DD, Demers LM, Baylink DJ. Oviarectomy selectively reduces the concentration of transforming growth factor beta in rat bone - - implications for estrogen deficiency-associated bone loss. Proc Natl Acad Sci 1992;89:12190-12193.
[12] Butta A, MacLennan K, Flanders KC et al. Induction of transforming growth factor #1 in human bre~tst cancer in vivo following tamoxifen treatment. Cancer Res 1992;52:4261-4264.
[13] Colltetta AA, Wakefield LM, Howell FV, Danielpour D, Baum M, Sporn MB. The growth inhibi- tion of human breast cancer cells by a novel synthetic progestin involves induction of transforming growth factor beta. J Clin Invest 1991;87:277-283.
[14] Kimchi A, Wang X-F, Weinberg RA, Cheifetz S, Massagu~ J. Absence of TGF-//receptors and growth inhibitory responses in retinoblastoma ceils. Science 1988;240:196-198.

30 D.J. Grainger et al./ Clinica Chimica Acta 235 (1995) 11-31
[15] Grainger DJ, Kirschenlohr HL, Metcalfe JC, Weissberg PL, Wade DP, Lawn RM. Proliferation of human smooth muscle cells promoted by lipoprotein(a). Science 1993;260:1655-1658.
[16] Kirschenlohr HL, Metcalfe JC, Weissberg PL, Grainger DJ. Adult human aortic smooth muscle cells in culture produce active TGF-B. Am J Physiol 1993;265:C571-C576.
[17] Grainger DJ, Weissberg PL, Metcalfe JC. Tamoxifen decreases the rate of proliferation of rat vas- cular smooth muscle cells in culture by inducing production of transforming growth factor ~. Biochem J 1993;294:109-112.
[18] Schlunegger MP, Grutter MG. An unusual feature revealed by the crystal structure at 2.2 A resolu- tion of human transforming growth-beta 2. Nature 1992;358:430-434.
[19] Cbeifetz S, Weatherbee JA, Tsang ML, Anderson JK, Mole JE. The transforming growth factor-B system, a complex pattern of cross-reacting ligands and receptors. Cell 1987;48:409-415.
[20] Wakefield LM, Smith DM, Masui T, Harris CC, Sporn MB. Latent transforming growth factor-B from human platelets. J Biol Chem 1988;263:7646-7654.
[21] Lyons RM, Keski-Oja J, Moses HL. Proteolytic activation of latent transforming growth factor-B from fibroblast conditioned medium. J Cell Biol 1988;106:1659-1665.
[22] Sato Y, Rifkin DB. Inhibition of endothelial cell movement by pericytes and smooth muscle cells: activation of a latent transforming growth factor-Bl-like molecule during co-culture. J Cell Biol 1989;109:309-315.
[23] Masui T, Wakefield LM, Lechner JF, LaVeck MA, Spron MB, Harris CC. Type B transforming growth factor is the primary differentiation-inducing serum factor for normal human bronchial epithelial cells. Proc Natl Acad Sci 1986;83:2438-2442.
[24] Danielpour DD, Dart LL, Flander KC, Roberts AB, Sporn MB. Immunodetection and quantifica- tion of two forms of transforming growth factor beta (TGF-BI and TGF-$2) secreted by cells in culture. J Cell Physiol 1989;138:79-86.
[25] Tanebe M, Teshima S, Hanyu T, Hayashi Y. Rapid and sensitive method for erythropoietin deter- mination in serum. Clin Chem 1992;38:1752-1755.
[26] Lin HY, Wang, X-F, Bg-Eaton E, Weinberg RA, Lodish HF. Expression cloning of the transform- ing growth factor-B type 11 receptor, a functional serine/threonine kinase. Cell 1992;68:775-786.
[27] Smith DB, Johnson KS. Single-step purification of polypeptides expressed in Escherichia coli as fusions with glutathione S-transferase. Gene 1988;67:31-40.
[28] Kocan RM, Moss NS, Benditt EP. Human arterial wall cells and tissues in culture. Methods Cell Biol 1980;21A:153-166.
[29] Ferns GA, Raines E J, Sprugel KH, Motani AS, Reidy MA, Ross R. Inhibtion of neointimal smooth muscle cell accumulation after angioplasty by an antibody to PDGF. Science 1991;253:1129-1133.
[30] Assoian RK, Sporn MB. Type beta transforming growth factor in human platelets: release during platelet degranulation and action on vascular smooth muscle cells. J Cell Biol 1986; 102:1031-1042.
[31] Barnard JA, Lyons RM, Moses HL. The cell biology of transforming growth factor-beta. Biochim Biophys Acta 1990; 1032:79-87.
[32] Danielpur D, Kim KY, Dart LL, Watanabe S, Roberts AB, Spom MB. Evidence for differential regulation of TGF-BI and TGF-B2 expression in vivo by sandwich enzyme-linked immunosorbent assays. Ann NY Acad Sci 1990;593:300-302.
[33] McCaffrey TA, Falcone DJ, Brayton CF, Agarwal LA, Welt FG, Weksler BB. Transforming growth factor-beta activity is potentiated by heparin via dissociation of the TGF-Bhx2-macro- globulin inactive complex. J Cell Biol 1989;109:441-448.
[34] Kanzaki T, Olofsson A, Moren A et al. TGF-BI binding protein: a component of the large latent complex of TGF-BI with multiple repeat sequences. Cell 1990;61:1051-1061.
[35] O'Connor-McCourt MD, Wakefield LM. Latent transforming growth factor-B in serum. J Biol Chem 1987;262:14090-14099.
[36] Danielpour D. Improved sandwich enzyme-linked immunosorbent assays for transforimg growth factor/~-1. J Immunol Methods 1993;158:17-25.
[37] Chao CC, Janoff EN, Hu SX et al. Altered cytokine release in peripheral blood mononuclear cell cultures from patients with chronic fatigue syndrome. Cytokine 1991;3:292-298.

D.J. Grainger et al . / Clinica Chimica Acta 235 (1995) 11-31 31
[38] Dasch JR, Waegell WO, Carroll K, Pace DR, Ellingsworth LE. Capture immunoassays specific for TGF-~-I and TGF-~-2: use in pharmacokinetic studies. Ann NY Acad Sci 1990;593:303-305.
[39] Shirai Y, Kawata S, Ito N e t al. Elevated levels of plasma transforming growth factor beta in patients with hepatocellular carcinoma. Jpn* J Cancer Res 1992;83:676-679.
[40] Anscher MS, Peters WP, Reisenbicber H, Petros WP, Jirtle RL. Transforming growth factor ~ as a predictor of liver and lung fibrosis after autoiogous bone marrow transplantation for advanced brea,~t cancer. N Engl J Med 1993;328:1592-1598.
[41] Wakefield LM, Winokur TS, Hollands RS, Christopherson K, Levinson AD, Sporn MB. Recombi- nant latent transforming growth factor/31 has a longer plasma half-life in rats than active trans- forming growth factor/~1, and a different tissue distribution. J Clin Invest 1990;86:1976-1984.
[421 Anderson J, Briefel G, Jones JM, Ryu IH, McQuire M, Yun YP. Effects of acetate dialysate on tramforming growth factor beta 1, interleukin and O2-microglobulin plasma levels. Kidney Int 1991;40:1110-1117.
[43] Singh JP, Chaiken MA, Styles CD. Phylogenetic analysis of platelet-derived growth factor by radio- receptor assay. J Cell Biol 1982;95:667-671.
[44] Heldin C-H, Westermark B, Wasteson A. Demonstration of an antibody against platelet-derived growth factor. Exp Cell Res 1981;136:255-261.
[45] Ross R, Raines EW, Bowen-Pope DF. The biology of platelet-derived growth factor. Cell 1986;46:155-169.
Copyright © 2022 FDOKUMEN




















