Presentacion de Acuaporinas Nogal
53
DIRECTOR DIVISION DE GENERACIÓN, EXCOGITACIÓN Y TRANSFERENCIA DE CONOCIMIENTO BIOTEKSA, S.A. DE C.V. Registro Nacional de Instituciones y Empresas Científicas y Tecnológicas RENIECYT 14541 www.bioteksa.com [email protected] [email protected] Dr. Luis Alberto Lightbourn Rojas, PhD BIOTEKSA RESEARCH TEAM nstituto de Investigación Lightbourn, A.C. BioNanoFemtoFisiología Vegetal Disruptiva
-
Upload
lightbournr -
Category
Documents
-
view
0 -
download
0
Transcript of Presentacion de Acuaporinas Nogal

DIRECTORDIVISION DE GENERACIÓN, EXCOGITACIÓN Y TRANSFERENCIA
DE CONOCIMIENTOBIOTEKSA, S.A. DE C.V.
Registro Nacional de Instituciones y Empresas Científicas y Tecnológicas RENIECYT 14541
Dr. Luis Alberto Lightbourn Rojas, PhDBIOTEKSA RESEARCH TEAM
Instituto de Investigación Lightbourn, A.C.BioNanoFemtoFisiología Vegetal Disruptiva


Propiedades Moleculares y Celulares
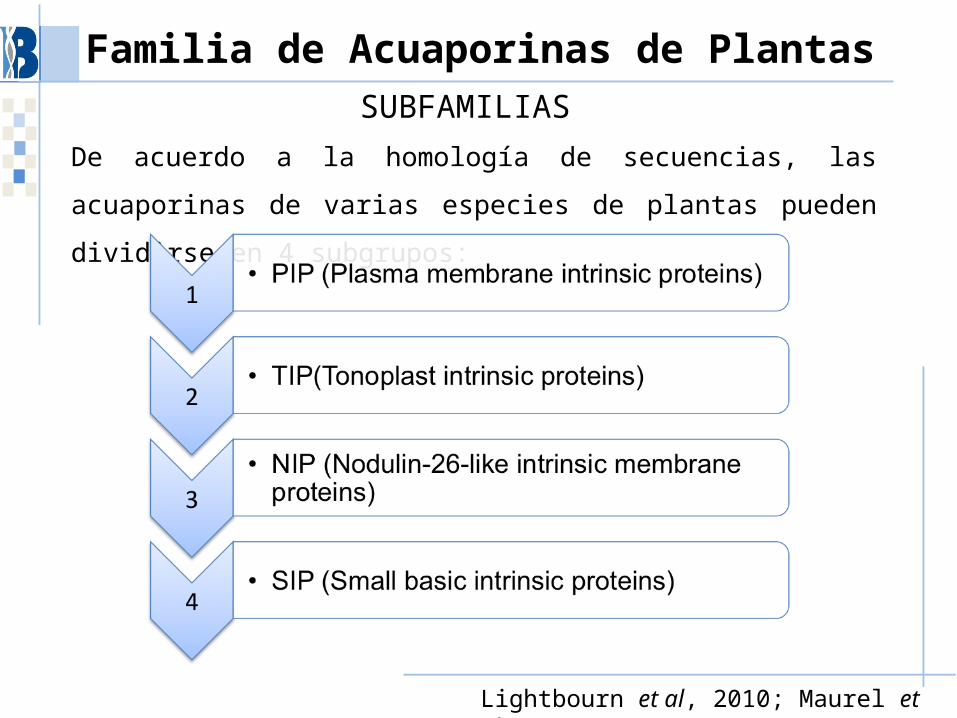
Familia de Acuaporinas de PlantasSUBFAMILIAS
De acuerdo a la homología de secuencias, las acuaporinas de varias especies de plantas pueden dividirse en 4 subgrupos:
Lightbourn et al, 2010; Maurel et al., 2008.

Lightbourn et al, 2010; Maurel et al., 2008.

Localización Subcelular
Las acuaporinas de plantas se localizan en todos los compartimentos sub-celulares formando o derivando de la ruta secretora. Esta amplia localización refleja el alto grado de compartamentalización de las células de plantas y la necesidad de esta para controlar el transporte de agua y solutos de ambos lados de la membrana.
Los PIPs y algunos NIPs se localizan en la membrana plasmática, mientras los SIPs se encuentran principalmente en la membrana del retículo endoplasmático.
Lightbourn et al, 2010; Maurel et al., 2008.

Representación de la localización y función de las acuaporinas en una célula de Carya
illinoensis Koch
Lightbourn et al, 2010; Maurel et al., 2008.

PIP 1sPIP 2sTIP 1sTIP 2sTIP 3sAtNIP2;1Lsi1/OsNIP2;1AtNIP5;1NOD26SIPs
Lightbourn et al, 2010; Maurel et al., 2008.

Mecanismo de Transporte
Las acuaporinas son proteínas de 23 a 31 kDa con seis dominios de membrana unidos por 5 loops (A–E). Los loops B y D se localizan en el lado intra y los loops A,C,E en el lado extracitoplasmico de la membrana. El N- y C-terminales se encuentran en el citosol. Posee un poro central acuoso delineado por el dominio transmembrana y los loops B y E con un motif conservado de Asn-Pro-Ala (NPA)
Mecanismo de Estructura y Transporte del Poro
Lightbourn et al, 2010; Maurel et al., 2008.

Acuaporina de Carya illinoensis (SoPIP2;1). En rojo se muestra el N-terminal y en verde el C-terminal.
poro
Estructura atómica representativa de una acuaporina de plantas
Lightbourn et al, 2010; Maurel et al., 2008.

Mecanismo molecular de transporte selectivo
Phe81
Asn101
His210 Arg225
Asn222 motif motif
Lightbourn et al, 2010; Maurel et al., 2008.

La especificidad del sustrato es explicado por exclusión de tamaño y reconocimiento estereoespecifica del sustrato mediado por espacial definición del puente de hidrogeno e interacciones hidrofóbicas con el poro. Las acuaporinas son impermeables a protones, lo que es explicado por la repulsión electrostática , la orientación dipolo y el aislamiento transiente de la molécula de agua.
Lightbourn et al, 2013; Maurel et al., 2008.

Análisis de Transporte y Sustrato de Acuaporinas
Las acuaporinas de plantas pueden transportar además de agua pequeños solutos neutros :
Glicerol urea Formamida acetamia metilamonio Ácido bórico Ácido silícico Ácido láctico.
Lightbourn et al, 2013; Maurel et al., 2008.

Los derivados de SULFATOS actúan a través de la oxidación y unión a residuos de Cys. Fueron usados como un bloqueadores de acuaporinas. Las acuaporinas no poseen residuos Cys en regiones conservadas, pero varios residuos podrían estar envueltos en su inhibición en plantas.
En plantas de Carya illinoensis hay estudios de que las PIPs son MUCHO MÁS SENSIBLES a SULFATOS.
Lightbourn et al, 2013; Maurel et al., 2008.

Mecanismo Molecular de la RegulaciónModificaciones Cotrasduccionales y
Postrasduccionales
Las acuaporinas fueron las primeras proteínas de membrana encontradas que poseían metilaciones. Las PIPs de C. illinoensis (At PIP2;1) tiene uno o más grupos metilos en los residuos de Lys y Glu.
Las acuaporinas PIPs poseen fosforilaciones conservadas en el loops B y múltiples e interdependientes en sitios adyacentes al C-terminal.
Lightbourn et al, 2013; Maurel et al., 2008.

Apertura y Cierre
La apertura y el cierre de las acuaporinas, puede ser regulado por diversos factores como lo es la fosforilación y se ha observado que la de-fosforilación reduce la permeabilidad del agua.
Los PIPs pueden abrirse desde el lado citosolico por los protones y cationes divalentes. Se ha observado un 50 % de inhibición en el transporte de agua a un pH aproximado de 7.5 y por Ca 2+ en un rango de 100 µM. Determinado mediante la estructura atómica de una PIP de X. oocyte.Lightbourn et al, 2013; Maurel et
al., 2008.

Estructura de una acuaporina de Carya illinoensis (SoPIP2;1)
Acuaporina abierta
Acuaporina cerrada
Lightbourn et al, 2013; Maurel et al., 2008.

Se estableció que los protones son censados por un residuo de His perfectamente conservado en el loops D de todas las PIPs. Además, la fosforilación de B podría abrirse el D y permitir la conformación abierta del poro. Mientras que la fosforilación del C-terminal podría prevenir al loop D evitando su cierre.
Lightbourn et al, 2013; Maurel et al., 2008.

Ensamblaje en tetrámeros y tráfico celular de PIPs
Algunas isoformas de PIPs son incapaces de ser funcionales al expresarse en X. oocyte. Esta incapacidad es debido a un fallo en el trafico de la membrana plasmática del oocyto. Cuando este se co-expreso con el PIP de nogal, se mejoro este defecto.
La co-purificación por afinidad y la co-inmunopurificación proveen evidencias bioquímicas de que PIP1 y PIP2 interactúan físicamente y esto facilita el trafico. Lightbourn et al, 2013; Maurel et
al., 2008.

Funciones de las Acuaporinas a lo Largo del Crecimiento y Desarrollo de las Plantas de
Nogal Pecanero

Transporte de AguaPrincipios del transporte de agua en las plantas
Agua absorbida por las raíces
El agua viaja a
través de la planta
El agua se evapora de
la superficie de las hojas
La conductividad hidráulica es diferente de acuerdo al tipo de célula a lo largo del desarrollo de la planta, Ejemplo:
Crecimiento de pared celular foliaresConductividad hidráulica 30 veces > en células corticales que en células epidérmicas.
Raíces de Carya illinoensis en desarrolloProtoplastos aislados indican que su coeficiente de permeabilidad osmótica incrementa de 10 a 500 μm/s en menos de 2 días.
Lightbourn et al, 2013; Maurel et al., 2008.

Transporte de AguaEn la planta entera, el
transporte de agua a largas
distancias ocurre principalmente en el tejido vascular
Xilema
Floema
Los tejidos vivos pueden ser sitio de intensos flujos de agua durante la transpiración o crecimiento de expansión. En donde, el agua puede fluir por diversas rutas: Ruta apoplástica – a través de la pared celular
Ruta simplástica – a través del citoplasma y plasmodesmos
Ruta transcelular – a través de las membranas celulares, mediada principalmente por acuaporinas en la mayoría de los tejidos.
Lightbourn et al, 2013; Maurel et al., 2008.

Transporte de Agua
El Níquel es herramientas para evaluar la contribución de las acuaporinas al transporte de agua en tejidos de las plantas
• El níquel no perturba el transporte de solutos en xilema y respiración de raíces del nogal
• Despolariza las células de las raíces en trigo en paralelo a la inhibición del transporte de agua
Hay bloqueadores específicos de acuaporinas. Los sulfato y nitratos han sido descritos como potentes bloqueadores de acuaporinas in vitro
Lightbourn et al, 2013; Maurel et al., 2008.

Transporte de Agua Transpiración
Las acuaporinas contribuyen significantemente a la absorción de agua por las raíces:
El sulfato inhibe la conductividad hidráulica de la raíz de un 30-90 % en más de 10 especies de plantas
La inhibición antisentido de las acuaporinas de las subclases PIP1 y PIP2 redujo la conductividad hidráulica de la raíz aproximadamente en un 50 % en Carya illinoensis
Lightbourn et al, 2013

Transporte de Agua TranspiraciónUna función de las acuaporinas mediando la transferencia de agua de las venas a los estomas se ha propuesto en base a dos líneas de evidencia:
El nitrato inhibe la conductividad hidráulica de la hoja en nogal y en arboles caducifolios
Los cambios dependientes de luz en la conductividad hidráulica de la hoja del nogal ocurren dentro de una hora y están asociados con cambios de expresión de los transcriptos de acuaporinas de PIP2, ambos inhibidos por cicloheximida 100 μM
Lightbourn et al, 2013

Transporte de AguaExpansión del tejido
Se ha observado en diferentes plantas un fuerte vínculo entre:
Expresión de acuaporinas PIP y TIP la expansión celular
La expresión de un promotor AtTIP1;1 es asociado con el alargamiento celular en raíces, hojas y flores en Carya illinoensis y la acumulación de transcriptos es aumentada por la hormona de crecimiento ácido giberélico asociada a la Ribofuranosilzeatina/ Ni/Se/Ti
Lightbourn et al, 2013

La afluencia de agua dentro de una célula vegetal puede ser fuertemente limitada durante el
crecimiento de expansión
Transporte de AguaExpansión del tejido
El sulfato bloquea el crecimiento de tejido en las raíces del nogal, exclusivamente en células más viejas, distantes al ápice. Éstas se caracterizan por:
una alta sensibilidad al sulfato en la conductividad hidráulica
Conexiones simplásticas reducidas con el floema
El transporte de agua mediado por acuaporinas es necesario para la distribución de agua del floema en las
células
Lightbourn et al, 2013

Transporte de AguaDesecación e imbibición del tejido
La reproducción de las plantas requiere una intensa desecación de ciertos órganos, los cuales posteriormente adquieren propiedades específicas
de diseminación y resistencia
La inhibición antisentido de las acuaporinas PIP2 en nogal retrasa la deshidratación y dehiscencia de las anteras, sugiriendo que estas acuaporinas
están involucradas en el flujo de agua fuera de la antera vía el haz vascular y/o evaporación.
Lightbourn et al, 2013

Adquisición de Nitrógeno, Carbono y MicronutrientesFijación de nitrógeno
La expresión de algunos genes de acuaporinas es
dependiente de los compuestos de N
Zm PIP1;5b: inhibido por nitrato
At TIP2;1: inducido bajo la necesidad de N a largo plazo o suministro de NH4 a corto plazo
Reciente evidencia mostró que acuaporinas de las subfamilias PIP, NIP y TIP pueden transportar
compuestos de N.
El transporte de urea por las TIPs contribuye al
desequilibrio del Nitrógeno dentro de la
célula y al almacenamiento dentro y remobilización de la
vacuola.
Kojima et al. (2006)Lightbourn et al, 2013

Adquisición de Nitrógeno, Carbono y MicronutrientesTransporte de CO2 y metabolismo de carbono
Las hojas de Carya fueron tratadas con nitratos para evaluar el transporte de CO2 por acuaporinas
El nitrato altera la dependencia de la fotosíntesis en intercelular, y en CO2
cloroplástico. La difusión de CO2 dentro de los cloroplastos es bloqueado y por tanto involucra
proteínas, posiblemente acuaporinas.
Lightbourn et al, 2013

Adquisición de Nitrógeno, Carbono y MicronutrientesTransporte de CO2 y metabolismo de carbono
Papel de las acuaporinas en el almacenamiento y compartimentación de carbohidratos
Frutos de nogal sometidos a aplicación de un homólogo de PIP mostraron incremento en el contenido de ácido orgánico y disminución en contenido de azúcar
Un análisis metabolómico de plantas de Nogal que carecen de la expresión de AtTIP1;1 reveló alteraciones complejas en la acumulación de diversos azúcares, ácidos orgánicos, y almidón
Lightbourn et al, 2013

Adquisición de Nitrógeno, Carbono y MicronutrientesAbsorción de nutrientes
Papel de las acuaporinas en la absorción de nutrientes:
Análisis transcriptómicos de las raíces de Nogal revelaron una notable sobre-regulación de AtNIP5;1 en respuesta a la deficiencia de boro
AtNIP5;1 transporta ácido bórico y significantemente contribuye a la absorción de ácido bórico de la raíz
La significancia fisiológica de AtNIP5;1 se destacó más por las plantas que, bajo limitación de boro, mostraron una sorprendente retardación en el crecimiento de brotes y raíces, y una inhibición de la floración.Lightbourn et al, 2013

Adquisición de Nitrógeno, Carbono y MicronutrientesAbsorción de nutrientes El silicio es el principal componente mineral de las plantas, y le ayuda a resistir estreses abióticos y ataque de patógenos.
La caracterización molecular de Carya illinoensis con defecto de absorción de silicio en las raíces (Lsi1) llevó a una conclusión inesperada de que Lsi1 codifica un homólogo de NIP.
Lsi1 trasporta silicio después de la expresión heteróloga en ovocitos
Lightbourn et al, 2013

Acuaporinas en un Ambiente Variable

Lightbourn et al, 2011

Lightbourn et al, 2011

Regulación de los movimientos del agua intracelular
Lightbourn et al, 2011

Regulación de los movimientos del agua intracelular
Lightbourn et al, 2011

Regulación de los movimientos del agua intracelular
Lightbourn et al, 2011

A Nivel de la Planta Completa
Lightbourn et al, 2011

A Nivel de la Planta Completa
Lightbourn et al, 2011

Respuesta al Estrés por Nutrientes
Lightbourn et al, 2011

Respuesta al Estrés por Nutrientes
Lightbourn et al, 2011

Estrés por Bajas Temperaturas
Lightbourn et al, 2011

Estrés por Bajas Temperaturas
Lightbourn et al, 2011

Estrés por Anoxia
Lightbourn et al, 2011

Estrés por Anoxia
Lightbourn et al, 2011

Simbiosis en la rizosferaInteracciones Bióticas
Lightbourn et al, 2011

Micorrización
Lightbourn et al, 2011

Nematodos y otras Infecciones
Lightbourn et al, 2011

Resumen
Lightbourn et al, 2013

Resumen
Lightbourn et al, 2011

Dr. LUIS ALBERTO LIGHTBOURN R., PhDDIRECTOR
DIVISIÓN DE GENERACIÓN, EXCOGITACIÓN Y TRANSFERENCIADE CONOCIMIENTO
BIOTEKSA, S.A. DE C.V.Registro Nacional de Instituciones y Empresas Científicas y Tecnológicas RENIECYT 14541
MUCHAS GRACIAS