
Measuring bleeding in platelet transfusion trials - Oxford ...
Upload
independentCategory
view
4download
0
PDGFR-β Regulates Proliferation and Migration of Gonocytes
Short title: PDGFR-β regulates gonocytes development
Sabrina Basciani, Gabriele De Luca, Susanna Dolci, Marina Brama, Mario Arizzi,
Stefania Mariani, Giuseppe Rosano, Giovanni Spera and Lucio Gnessi
Department of Medical Physiopathology, University of Rome “La Sapienza”, Rome, Italy (S.B., M.B., M.A., S.M., G.S., L.G.), Department of Medical Sciences, Cardiovascular Research Unit, IRCCS San Raffaele, Roma, Tosinvest Sanità, Rome, Italy (S.B., G.R.), Department of Environment and Primary Prevention, Istituto Superiore di Sanità, Rome, Italy (G.DL.), Department of Public Health and Cell Biology, Section of Anatomy, University of Rome ‘‘Tor Vergata’’, Rome, Italy (S.D.)
Key words: PDGFR, gonocytes, testis, development, spermatogenesis
Corresponding author:
Lucio Gnessi, MD, PhD,
Department of Medical Physiopathology,
Sapienza University,
Policlinico Umberto I,
00161 - Rome, Italy
e-mail: [email protected]
Disclosure Statement: The authors have nothing to disclose.
Endocrinology. First published ahead of print August 7, 2008 as doi:10.1210/en.2008-0349
Copyright (C) 2008 by The Endocrine Society

Abstract
Proliferation and migration of gonocytes, the precursors of spermatogonial stem cells, to the
germline niche in the basal membrane of the seminiferous tubules, are two crucial events that
take place between postnatal days 0.5 (P0.5) and P5.0 in the mouse, and involve a selection of
the cells that are committed to the germline stem cells lineage. Here we show that from
embryonic day 18.0 (E18) and up to P5 the gonocytes express platelet-derived growth factor
receptor β-subtype (PDGFR-β) and that during the same time period the Sertoli cells express
PDGF-B and PDGF-D, both ligands for PDGFR-β. Inhibition of the PDGFR-β tyrosine
kinase activity during the first 5 postnatal days provokes a profound reduction of gonocytes
number through inhibition of their proliferation and induction of apoptosis. Moreover, we
found that PDGFR-β ligands are chemotactic for gonocytes. These data suggest that PDGFR-
β activation has the remarkable capability to drive the selection, survival and migration of the
gonocytes from the center of the seminiferous tubules to the testicular germline niche on the
basal membrane.

INTRODUCTION
In the mouse the fundamental structure of testis cords is established at E11.5 when the primordial germ cells (PGCs), migrated from the allantois, are enclosed by Sertoli cells (1). There, PGCs grow continuously until around E16.5, and then enter G0/G1 arrest. Mitotically quiescent germ cells, called gonocytes, are uniformely round, located in the center of the seminiferous tubules, and separated from the basal membrane (2-4). Between postnatal day (P) 0.5 and P5.0, gonocytes proliferate, extend cytoplasmic processes, and migrate to the basement membrane (2). Migration of gonocytes is likely critical to ensure survival of these cells, as suggested by the observation that gonocytes remaining in the center of testis cords eventually degenerate (5). Gonocytes migrated at the basement membrane, will give rise to spermatogonial stem cells (SSCs), which establish and maintain spermatogenesis in the mature testis. The mechanism of gonocytes proliferation and migration is uncertain, though there are two hypotheses that can be assumed. The first is that it is programmed beforehand, the other hypothesis is that it is regulated by Sertoli cells. Sertoli cells can synthesize several secretory products (6) and some of them have the potential to accomplish this task. Among these products, platelet-derived growth factor (PDGF) seems ideally suited to drive proliferation and migration of gonocytes. The common biological functions of PDGF signaling pathway include regulation of cell proliferation and survival, angiogenesis, cell migration, membrane ruffles and cytoskeletal rearrangements (7). The PDGF family of growth factors, that consists of four gene products PDGF-A, PDGF-B, PDGF-C and PDGF-D, selectively signals through two PDGF receptors (PDGFRs), PDGFR-α and PDGFR-β (8) and participate in a variety of developmental processes. PDGF-B and PDGFR-β are essential for the development of support cells in the vasculature, whereas PDGF-A and PDGFR-α are more broadly required during embryogenesis, with essential roles in numerous contexts, including central nervous system, neural crest and organ development (8, 9).
In the testis, PDGF-A and PDGFR-α are XY-specific factors required for the proliferation and differentiation of fetal and adult Leydig cells (10, 11) and PDGFR-α is a recognized marker of the undifferentiated stem cell which will give rise to the Leydig cells lineage after birth (12). PDGFR-β and a truncated cytosolic variant of PDGFR-β (V1-PDGFR-β) lacking the PDGF binding domain but with retained tyrosine kinase activity and ability to autophosphorylate, have been localized in the gonocytes during the first 5 days after birth in the rat (13-15), mouse and rat purified gonocytes proliferate following PDGF treatment (13, 14, 16), early postnatal rat Sertoli cells produce PDGFs, and peritubular myoid cells (PMC) precursors express PDGFR-β and are chemotactically attracted by its ligand (17). In man, testicular expression of the PDGF system is developmentally controlled through a spatiotemporal pattern of expression (18). In the current study, we used the tyrosine kinase inhibitor imatinib to explore the possible relationship between expression of PDGFR by neonatal gonocytes and proliferation and migration of these cells. Imatinib, a drug that inhibits the tyrosine kinase functions of BCR-ABL, PDGFRs, and c-kit receptor associated with specific diseases, and is a first-line treatment in patients with chronic myeloid leukemia and gastrointestinal stromal tumours (19), was administered during the first 5 days postpartum. This time window was selected to cover the period of postnatal gonocytes development and to avoid the interference with c-kit, the other target of imatinib, whose expression within the seminiferous tubules starts at P6 in mouse (20-23). The outcome of these studies provides substantial new in vivo and in vitro evidence supporting the concept that the expression of PDGR-β by neonatal gonocytes, has a critical role in driving proliferation and migration of these cells toward the basal membrane. PDGR-β inhibition provokes a profound reduction of gonocytes number and of their immediate descendents, undifferentiated spermatogonia, with a delay in the establishment of the seminiferous epithelial cycle.

Materials and Methods
Animals treatment Newborn CD1 male mice (Charles River Labortories, Calco, Italy) were injected intraperitoneally with 50 mg/kg imatinib (provided by Dr. Elisabeth Buchdunger, Novartis Pharmaceuticals, Basel, Switzerland), in 100 μl 0.9% NaCl every 8 hours for 5 days. This treatment schedule was adopted to avoid potential toxic effects by imatinib and to assure the continuous block of the target tyrosine kinase activities (by 8 hours more than 70% of the tyrosine kinase inhibition is loss in intraperitoneally treated mice) (24). Control animals were treated with vehicle alone. Protocols for use of animals were in accordance with the guidelines of the local committe for animal research. Generation of PDGF-B and PDGFR-β mutant mice PDGF-B +/- (25), and PDGFR-β +/- mice (26) were bred as 129Ola/C57BL6J, and 129Sv/C57BL6J hybrids, respectively. Heterozygotes were intercrossed and offspring of different age and genotype were killed and fixed for histological analysis. Because of the lethality of the PDGF-B and PDGFR-β gene targeting at birth, KO mice were studied at E18.5. Histology, immunohistochemistry, cell proliferation and apoptosis assays Testes to be stained by bromodeoxyuridine (BrdU), TdT-mediated dUTP biotin nick end labeling (TUNEL), PDGF-B, PDGF-D, PDGFR-β, SMAD-5, RET and c-kit immunohistochemistry were fixed in Bouin’s solution and paraffin embedded. The reactions were carried out on 5-μm sections. For routine histology, sections were stained by hematoxylin. Histological assessment of the number of tubular cells exhibiting morphology typical of Sertoli cells, spermatogonia, spermatocytes, round and elongated spermatids (27), was performed in P70 animal testis. Cells were counted in at least 30 randomly selected tubular cross-sections from testis tissue from four imatinib treated and four control animals. For BrdU labeling, BrdU (Sigma-Aldrich s.r.l, Milano, Italy) was injected intraperitoneally (100
mg/kg body mass). Injected animals were killed 1 h later and the fixed testes were processed as described (28). Apoptotic cells were detected by TUNEL using the TCAS TdT DAB in situ detection kit (R&D Systems, Minneapolis, MN). Positive cells were detected using rabbit anti-PDGF-B, anti-PDGF-D, anti-c-kit, anti-PDGFR-β C-terminal, anti-RET and goat anti-SMAD-5 antibodies (Santa Cruz Biotechnology, Inc., Santa Cruz, CA). The antibodies, at 1:100 dilution, were incubated overnight at 4°C. Positive reactions were detected by the streptavidin-biotin immunoperoxidase method using a commercial kit (Zymed Laboratories, San Francisco, CA) and 3,3’-diaminobenzidine as chromogenic substrate. Negative controls were done pre-absorbing the primary antibody with the corresponding immunogenic peptide. The preparations were counterstained with hematoxylin and mounted. The staging criteria of the seminiferous tubules in the adult testis were those of Russel et al. (27). Purification and treatment of gonocytes Gonocytes were obtained by sequential collagenase-hyaluronidase-trypsin treatment of 40-50 3-day-old Swiss-CD1 mice as described (29). To remove somatic cell contaminants, cell suspensions were plated for 4 hr on 10-cm-diameter tissue culture dishes in Eagle’s MEM supplemented with glutamine, nonessential amino acids, gentamycin, streptomycin, and penicillin (GIBCO, Grand Island, NY), 2mM sodium lactate, 1 mM sodium pyruvate, and 10% FCS in 5% CO2 umidified atmosphere at 32 °C. Non aderent cells were recovered and plated for an additional 12 h period at the same incubation conditions. Floating cells were collected and separated on a 2-4% BSA gradient. The fractions containing the most gonocytes (as judged by morphology and size) (30) were pooled (0.5 ± 0.03 x 106 gonocytes with a purity of 80-90%). To study the expression of PDGFRs and c-kit and the effect of imatinib on their phosphorylation, 1.25 x 105 cells were suspended and cultured at 32 °C with or without 10 μmol/L imatinib for 2 hours followed by an additional 10 minutes of incubation in the absence or in the presence of 10 ng/mL human recombinant PDGF-BB (Roche, Mannheim, Germany) or for an additional 30 minutes in the absence or in the presence of 100 ng/mL human recombinant kit

ligand [(KL) PeproTech, London, United Kingdom]. Cells were centrifuged and frozen at –80 C° until protein extraction. Western blot Tissues or cell proteins were extracted, 80 μg of protein extract per lane were loaded and separated on 7.5% polyacrylamide gel and transferred to nitrocellulose membranes (31). A positive control cell extract of a human breast cancer cell line (MCF-7, American Type Culture Collection, Rockville, MD), for the proteins found to be absent in gonocytes protein extract was included on the same gel. Membranes were incubated with 1:300 dilution of the following antibodies: anti-PDGFR-α, anti-PDGFR-β, anti-phospho-PDGFR-α (anti-p-PDGFR-α), anti-p-PDGFR-β, anti-p-c-kit, anti-c-kit and anti-β-actin (Santa Cruz Biotechnology). Thereafter, the membranes were incubated with horseradish peroxidase-conjugated secondary antibodies (1:10,000 dilution; Santa Cruz Biotechnology) for 1 hour at room temperature. Bound antibodies were detected by enhanced chemiluminescence (Amersham Pharmacia Biotech, Piscataway, NJ). Secondary antibodies alone served as negative controls. Protein bands were quantifid by densitometric analysis (Kodak Digital Science 1D Image analysis software, Eastman Kodak Co., Rochester, NY) after normalizing individual bands to the respective β-actin bands. Reverse transcription and PCR To study mRNA expression for Stra8, Rbm, Dazl, Lsh and CRABP, polyadenylated mRNA extraction and semiquantitative reverse transcription-PCR (RT-PCR) were done on frozen testis as described (31). PCR reactions were optimized for the number of cycles to ensure product intensity within the linear phase of amplification. Primers used are listed in Supplementary Table 1. Quantitation of the signals was done by densitometric analysis (Kodak Digital Science) after normalizing individual bands to the respective β-actin bands. Chemotaxis assay Chemotaxis was measured using a 48-well micro chamber (NeuroProbe, Inc., Baltimore, MD) with polycarbonate filters with 12μm pores (NeuroProbe, Inc.). In PDGFR inhibition
experiments, cells were preincubated for 30 min with 10 μM imatinib in DMEM, while the controls were incubated in DMEM only. PDGF-B and PDGF-D growth factors dilutions in DMEM were filled into the lower wells and covered by the filter. Purified gonocytes (2.5x104) in 50 μl DMEM were filled in the upper wells. After a 6-h incubation at 34°C in 5% CO2, the filter was carefully removed and cells on the upper side eliminated by rinsing with cold PBS and scraping over a rubber wiper. The cells migrated on the lower side of the filter were fixed in methanol and stained with Mayer's hematoxylin. Growth factors were tested in triplicate for each concentration. Control wells with DMEM only in the bottom well were applied for each experiment. The number of cells migrated in 12 random fields per well were counted (400 x magnification), and the average number of cells per high-power field (HPF) was calculated. The migration index was expressed as the ratio of migration observed in the presence of PDGFs or PDGFs plus imatinib and divided by the migration observed with the medium alone. Sperm counts Epididymes were removed from five imatinib treated and five control mice at P35, P70, P90 and P120, individually housed for 5 days prior to be killed. The cauda region from each epididymes was removed and quickly transferred into a culture dish containing 2 ml of DMEM 10% FCS at 37°C and then transferred into another dish with 200 μl of DMEM 10% FCS. Four incisions were made in the cauda epididymes to allow spermatozoa to disperse for 30 min at 37°C. Sperm concentration was determined with a hematocytometer on a sample of the epididymal sperm suspension. Assessment of serum hormone levels Blood samples were drawn by cardiocenthesis. After centrifugation, serum samples were stored at –80 °C until assayed. Testosterone (T) levels were determined by radioimmunoassay (DiaSorin S.p.A., VC, Italy) according to the manufacturer instructions. Serum FSH was measured by Dr. A. F. Parlow, National Institute of Diabetes and Digestive and Kidney Diseases, National Hormone and Peptide Program, Torrance, CA, by double-antibody RIA using mouse FSH standard (AFP-5308D), iodinated rat FSH

(AFP-4251A) and guinea pig anti-mFSH (AFP 1760191; final dilution, 1:200,000). Statistical analysis Data are expressed as mean values ± SEM. Statistical comparisons were made by Student’s t test for single statistical comparisons of different groups. The results were analyzed for statistically significant differences using ANOVA for multiple comparisons. P < 0.05 was considered significant.
Results
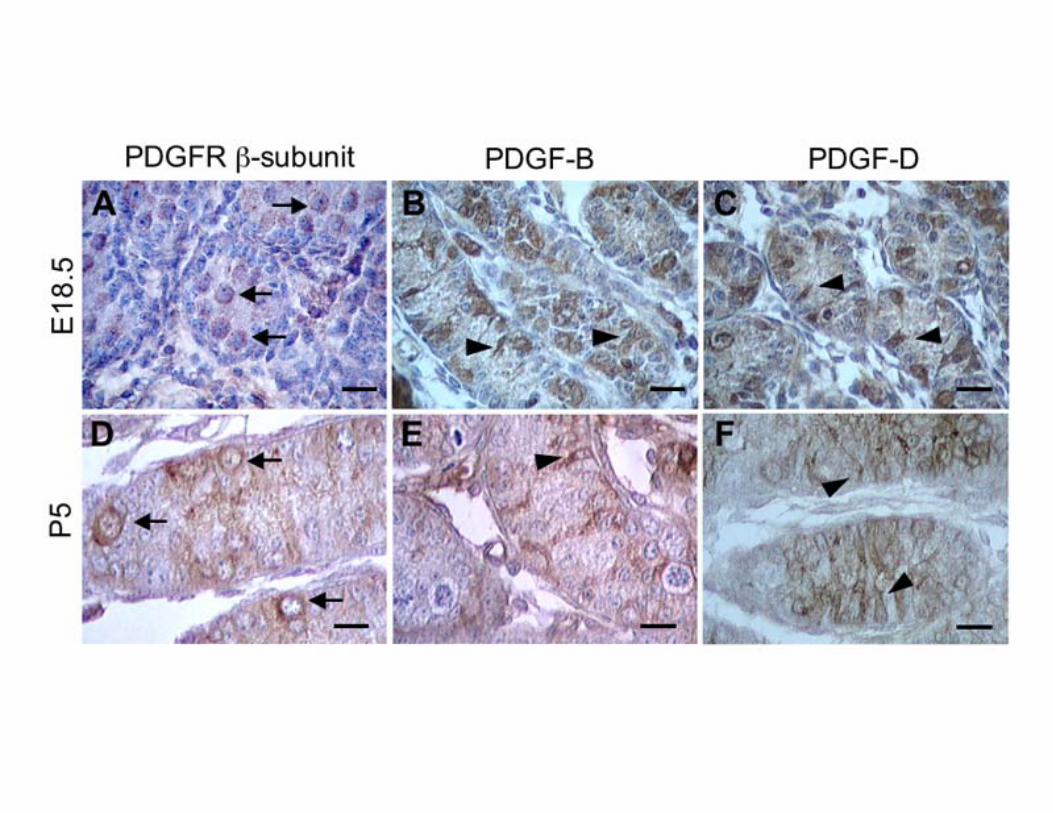
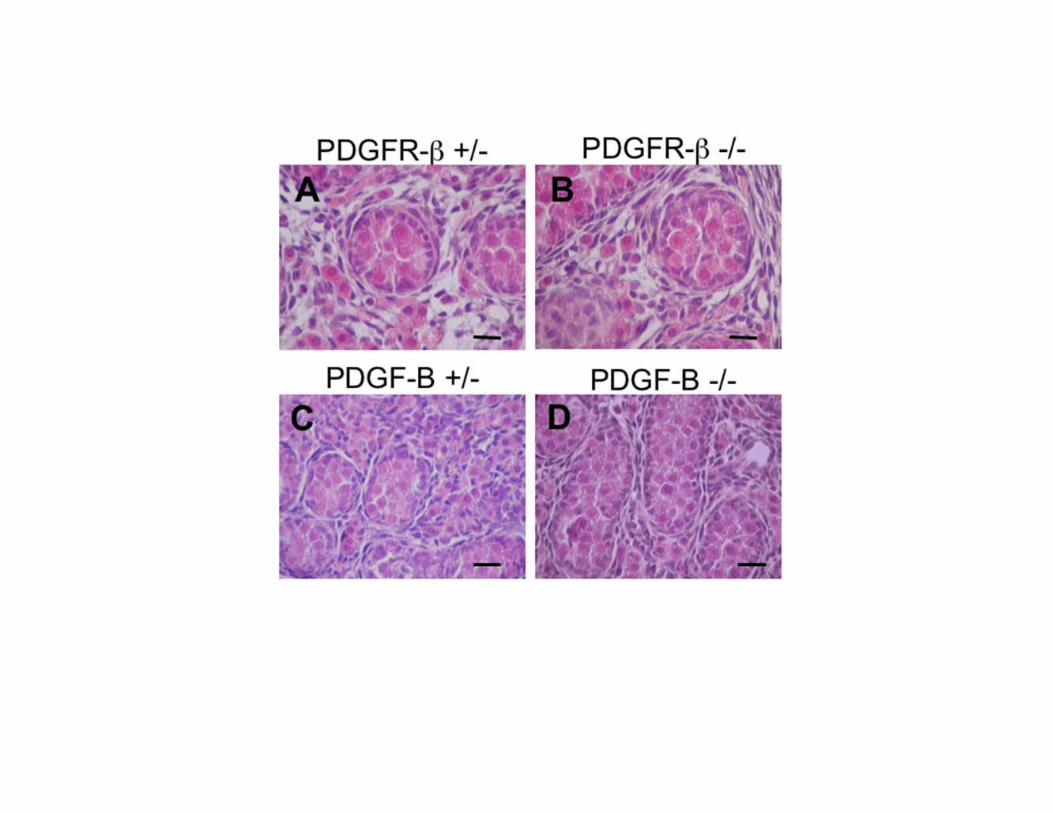
Gonocytes express PDGFR-β and Sertoli cells express PDGF-B and PDGF-D We first examined the localization of PDGFR-β, PDGF-B and PDGF-D by immunohistochemistry. For the analysis of the PDGFR β-subunit, we used an antibody directed against the C-terminal portion of the molecule that does not distinguish between PDGFR-β and the truncated form of PDGFR-β lacking the ligand binding domain, V1-PDGFR-β. At E18.5 the gonocytes were positive for the PDGFR β-subunit (Fig. 1A). The positive staining correlates well with gonocytes morphology (32). After birth, the number of centrally located positive cells decreases progressively and at P5, they are found on the basal membrane (Fig. 1D). After P5 the positivity is drastically reduced (data not shown). We also found that during the same time period the Sertoli cells express PDGF-B (Fig. 1B, E) and PDGF-D (Fig. 1C, F), both ligands for PDGFR-β. These findings suggest an involvement of PDGFR β-subunit in the development of the gonocytes. PDGFR-β and PDGF-B null mutations do not influence the prenatal development of the testis To address whether PDGFR-β activation is involved in gonocytes development, we analyzed the testis of PDGFR-β and PDGF-B null mutant mice (25, 26). The gene targeting of PDGFR-β, involves the kinase domain of the receptor sequence coding gene, entailing the V1-PDGFR-β as well. Due to the lethal phenotype of the homozigous mutants at E17.0-E19.0, our analysis was restricted to male mice recovered at E18.5. We did not find significant alterations in the microscopic
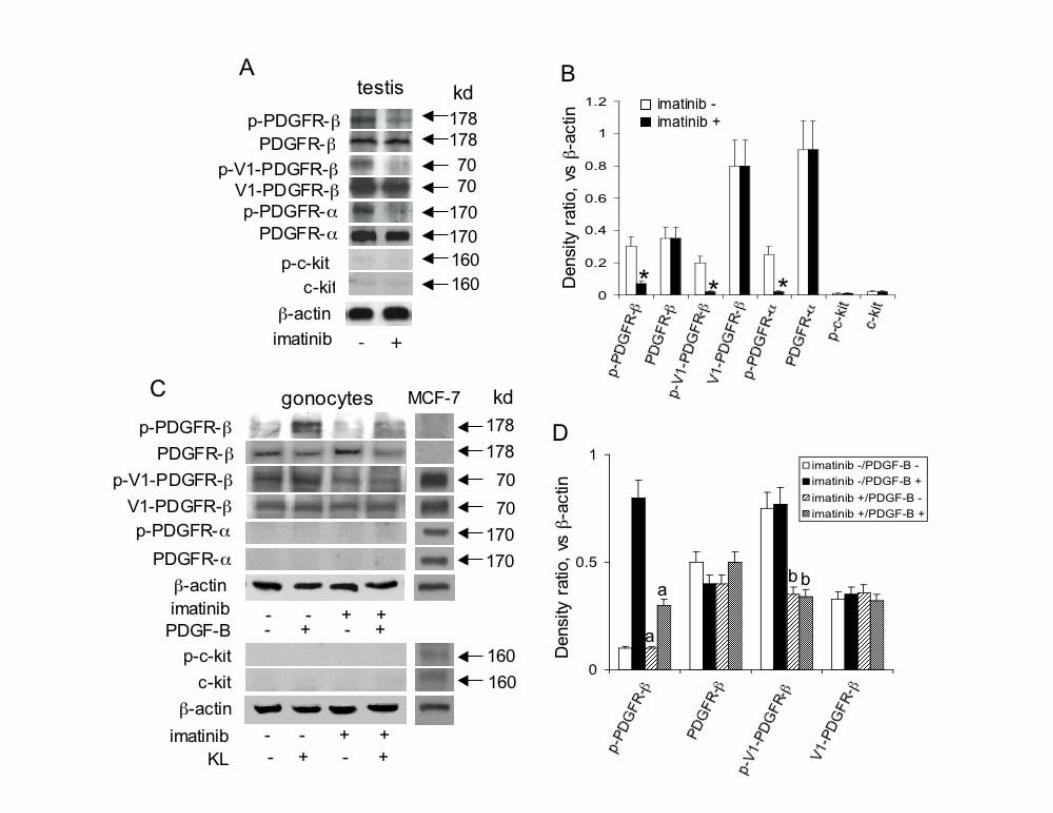
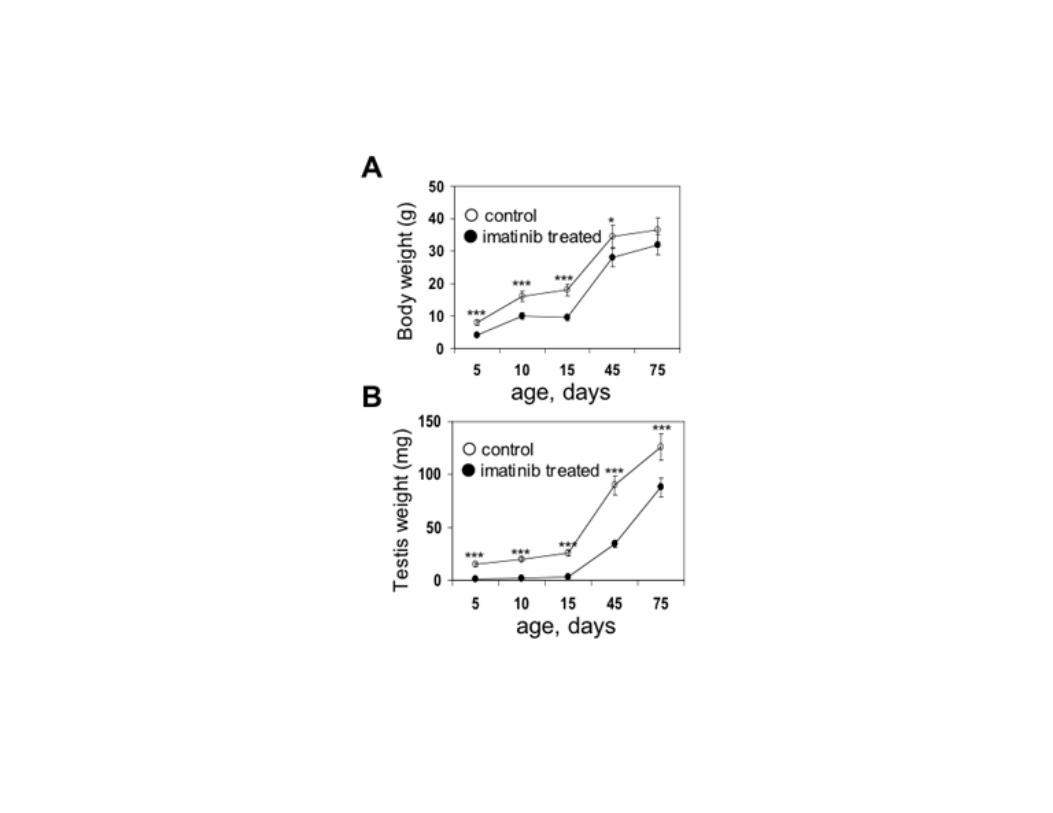
structure of the testis of PDGFR-β and PDGF-B -/- animals compared to heterozigous littermates (Fig. 2). This result extends what reported by Brennan et al. which found no overt defects in early XY gonads development at 11.5-13.5 dpc in PDGFR-β -/- mice (10), and rules out a critical role for the β-receptor of PDGF either wild type or truncated and for PDGF-B during testicular prenatal development. Imatinib inhibits the phosphorylation of PDGFR-β expressed by gonocytes To examine whether PDGFR-β signaling loss would affect the seminiferous tubules morphology postnatally, we administered the PDGFRs tyrosine kinase inhibitor imatinib mesylate to male mice from P1 to P5. We first examined the PDGFRs and c-kit protein expression and the effect of imatinib on PDGFRs and c-kit phosphorylation in the testes of treated animals and in isolated gonocytes by Western blot. Testis (Fig. 3A) and purified gonocytes (Fig. 3B) expressed the PDGFR-β protein whose phosphorylation was inhibited by imatinib. We also found a 70 kDa protein, most likely corresponding to V1-PDGFR-β (15), whose phosphorylation was inhibited by imatinib as well, in whole testis protein extracts (Fig. 3A, C) and in isolated gonocytes (Fig. 3B, D). The identity of the 70 kDa band with V1-PDGFR-β was also confirmed by the constitutive phosphorylation of the band not significantly increased by the treatment of the cells with the PDGFR-β ligand PDGF-B (Fig. 3B). PDGFR-α was found in testis extracts but not in isolated cells. According to previous data (20-23), c-kit was almost undetectable in the whole testis protein extracts and completely absent in gonocytes. These findings confirm that PDGFR-β and a V1-PDGFR-β-like protein are expressed in mouse gonocytes, show that the concentration and dosing schedule of imatinib were appropriate for in vivo inhibition of testicular receptor phosphorylation, and rule out a direct involvement of either c-kit or PDGFR-α kinase activity in gonocytes development. Inhibition of PDGFR-β activity during the first 5 postnatal days reduces testis size The animals treated with imatinib were smaller than untreated controls, and had smaller testes (Fig. 4). The testicular size reduction was more

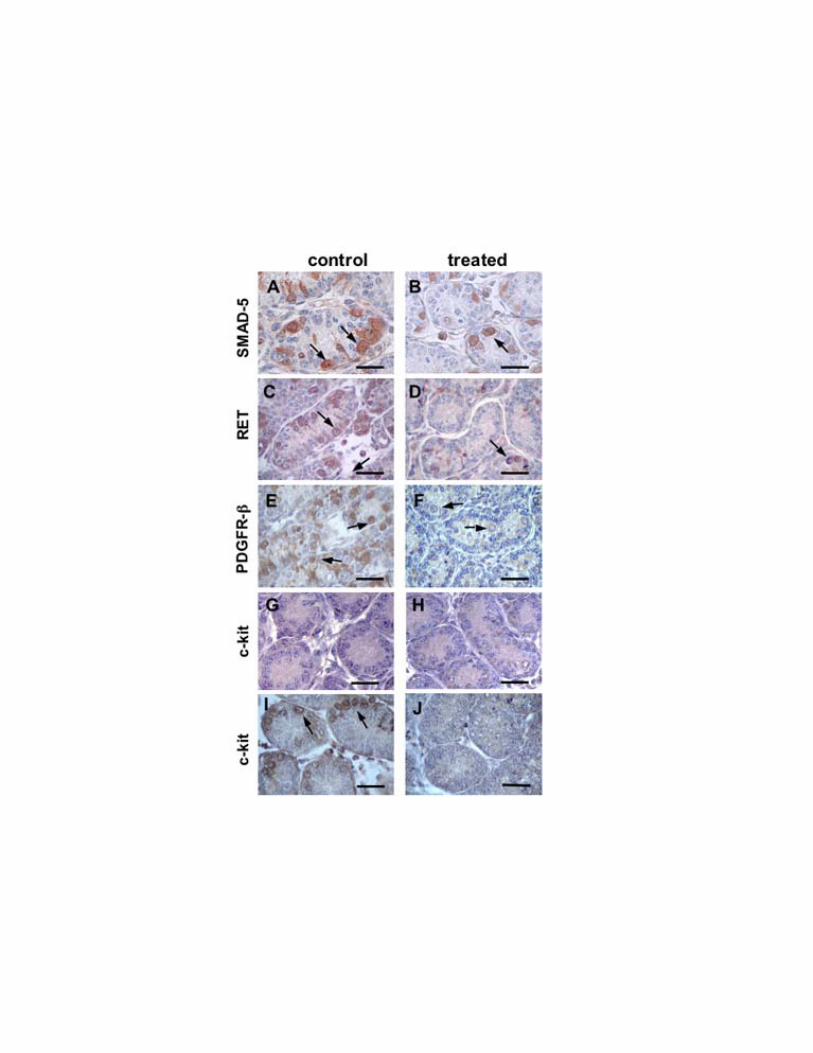
pronounced than the average reduction of the body size. During treatment and at follow-up the animals were healthy and behaved normally. Treated animals were also subjected to histopathologic analysis. No major finding was noted at autopsy. After the interruption of treatment, the weight of the animals increased and by the age of 10 weeks it matched the controls (Fig. 4A). On the contrary the testicular size was not recovered (Fig. 4B). We conclude that the gross phenotypic consequences of imatinib administration during the first 5 post-natal days are a transient growth retardation and testicular underdevelopment. Inhibiting PDGFR-β in the neonatal period reduces gonocytes number and their migration to the basal membrane, causes a depletion of SSCs and delays the initiation of spermatogenesis At P5, microscopic analysis of the testis of animals treated with imatinib, showed a delay in maturation of the germ line and a significant reduction of gonocytes and spermatogonia (Fig. 5). This was observed by analysis of expression of SMAD-5, a downstream signal mediator for several bone morphogenetic proteins which specifically marks mitotic spermatogonia in postnatal life (33, 34), RET receptor tyrosine kinase, whose expression is limited to SSCs (35), PDGFR-β and c-kit. Almost all gonocytes in control mice had migrated to the seminiferous tubule basal membrane. The majority were SMAD-5, RET and PDGFR-β positive (Fig. 5A, C, E). Imatinib treated animals had only few SMAD-5 (Fig. 5B) and RET positive cells (Fig. 5D) on the basal membrane while the center of the seminiferous tubules contained negative gonocytes. The anti-PDGFR-β antibody stained the few gonocytes still located in the center of the seminiferous tubule and rare cells on the basal membrane (Fig. 5F). c-kit, whose localization in the spermatogonia within the seminiferous tubules starts at P6 in mouse, was absent in both control (Fig. 5G) and treated (Fig. 5H) animals. At P8, c-kit positive spermatogonia were completely absent in treated mice (Fig. 5J) and abundantly seen in controls (Fig. 5I). We conclude that inhibition of PDGFR-β signaling during the first 5 days after birth, reduces the number of gonocytes and their migration to the basal membrane.
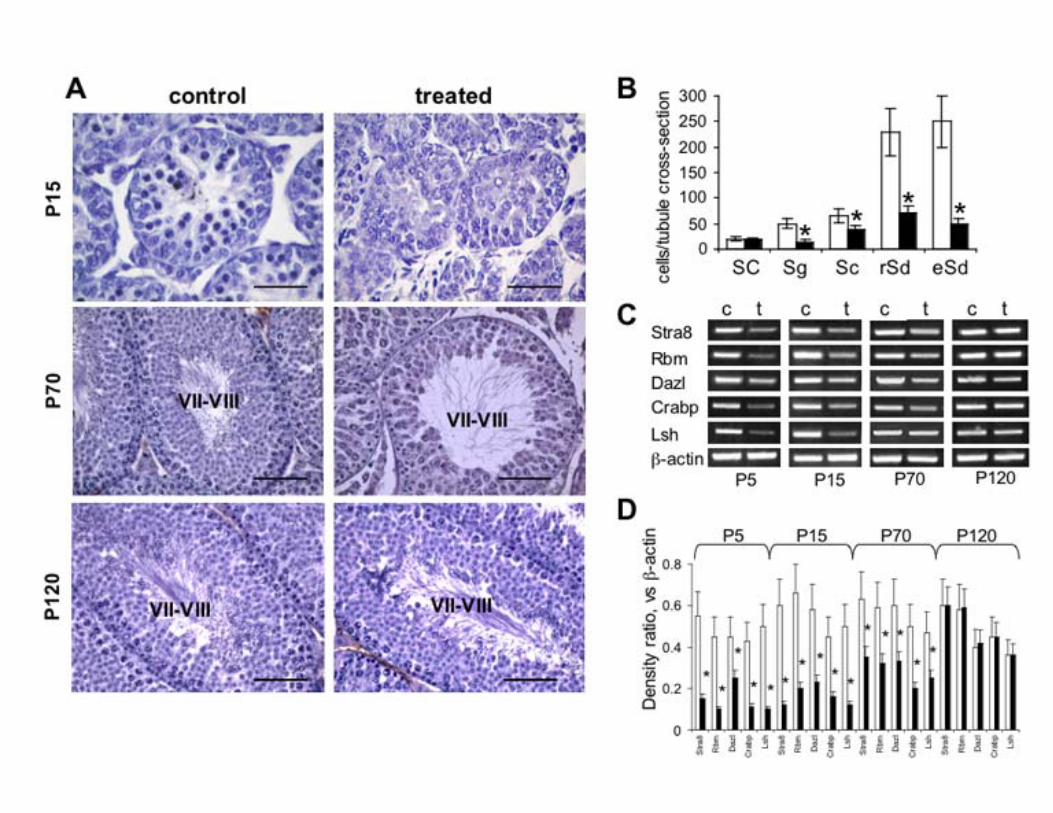
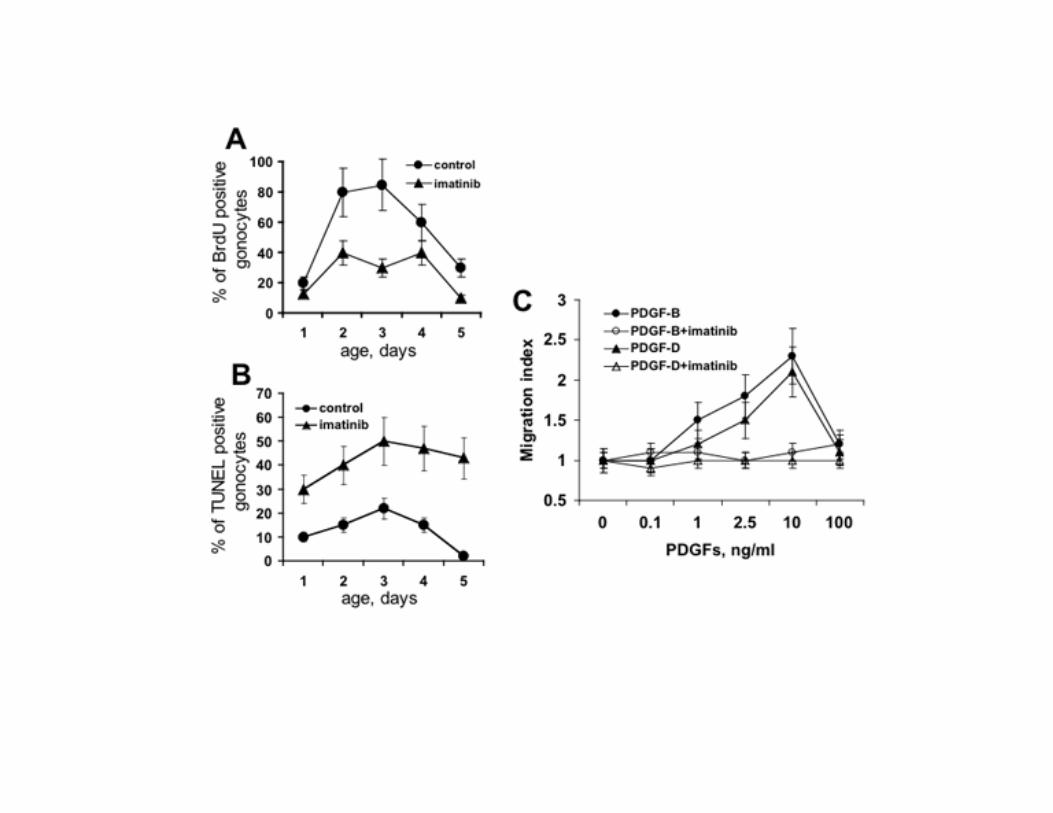
To evaluate whether the inhibition of the PDGFR-β activity in the early postnatal period could influence the development of spermatogenesis at later ages, we studied the histology of the testis at P15, P70 and P120 (Fig. 6A). By P15 the testes of control mice contained differentiated germ cells such as spermatocytes. In contrast, no differentiated germ cells were observed in testes from treated animals. At P70 the seminiferous tubules of imatinib treated mice showed a qualitatively normal spermatogenesis. However, the number of spermatogonia and of differentiated germ cells products (spermatocytes, round spermatids and elongated spermatids) per tubule cross-section was severely reduced (Fig. 6B). At older ages the spermatogenesis was completely recovered. No obvious defects in Sertoli cells, Leydig cells and PMC were seen at any age. To evaluate the pattern of germ cells development at molecular level, we compared the expression of spermatogonia-specific genes (36) between controls and imatinib treated animals at P5, P15, P70 and P120 (Fig. 6C, D). In imatinib treated animals, at P5, the spermatogonia marker Stra8 was reduced 3.7-fold, and other spermatogonia-selective transcripts, including RNA-binding-motif protein (Rbm), Dazl, lymphoid-specific helicase (Lsh) and cellular retinoic-acid-binding protein (CRABP) were reduced 3-fold, 2.1-fold, 3.5-fold and 3.9-fold, respectively. At P15, Stra8, Rbm, Dazl, Lsh and CRABP were reduced 2.6-fold, 2.4-fold, 1.9-fold, 2.3-fold and 2.9-fold respectively. At P70, Stra8, Rbm, Dazl, Lsh and CRABP were reduced 1.8-fold, 1.8-fold, 1.8-fold, 2.5-fold and 1.8-fold. At later ages these differences were no longer detectable. Thus, in agreement with the microscopic appearance, spermatogonia-specific genes are greatly underrepresented in the testes of imatinib treated animals during early postnatal and pubertal development followed by a progressive recovery at later ages. Silencing of PDGFR-β inhibits proliferation and induces apoptosis of gonocytes, and PDGFR-β ligands chemotactically attract isolated cells Cell proliferation was examined by labelling in vivo with BrdU (Fig. 7A). BrdU incorporation in gonocytes was profoundly reduced in imatinib treated animals compared with

controls. To determine whether the loss of gonocytes was due to apoptosis, we used TUNEL labelling, which showed an increase in gonocytes TUNEL staining in imatinib treated animals (Fig. 7B). BrdU incorporaton and TUNEL staining in testicular cells other than gonocytes were unaffected by imatinib (data not shown). We conclude that decreased proliferation and increased apoptosis are the main mechanisms of the gonocytes number reduction caused by PDGFR β-subunits activity inhibition. We next investigated whether PDGF chemotactically attracts gonocytes. Both PDGF-B and PDGF-D were attractants for the isolated cells (Fig. 7C). These results indicate that the reduction of gonocytes on the basal membrane after imatinib administration, in addition to induction of apoptosis and reduction of proliferation, could be ascribed to a defect of cells migration. PDGFRs inhibition does not alter serum levels of hormones Serum T and FSH levels were measured to determine whether the pituitary-testis axis had contributed to the testicular phenotype. We found no significant difference in serum levels of T and of FSH measured at P5 between control and imatinib-treated mice (T: 2.0 + 0.1 vs 1.9 + 0.2 ng/ml, mean + SEM of four animals; FSH: 6.15 + 2.0 vs 6.77 + 1.6 ng/ml, mean + SEM of four animals). These results indicate that the testicular phenotype after imatinib administration is due to the inhibition of PDGFRs rather than an endocrine disorder. Inhibition of PDGFR-β during the early postnatal days provokes a transient reduction of epidydimal sperm count To test whether the microscopic phenotype of the testis of imatinib treated mice would affect their reproductive potential, we examined epidydimal sperm count of the animals (Table 1). In line with the testicular histology, at P35 and P70, the number of spermatozoa from the epididymis of the treated mice was reduced by 86% and 67%. At P90 and P120, the number of spermatozoa matched that of controls. These findings indicate that PDGFR-β inhibition during the first 5 postnatal days results initially in a significant reduction of sperm count followed by an almost complete recovery.
Discussion Our studies provide evidence that activation of PDGFR-β is required for proliferation and migration of gonocytes in the early postnatal period, and that its silencing during the first five postnatal days causes a profound reduction of spermatogonia which is recovered at later ages. We could detect expression of PDGFR-β in the gonocytes and of PDGFR-β ligands, PDGF-B and PDGF-D, in Sertoli cells in prenatal and early postnatal testis by immunohistochemistry, placing the system in the correct spatio/temporal position for stimulating proliferation of gonocytes as well as for promoting their migration. Previous in vitro studies suggested a role for PDGF-B and PDGFR-β during prenatal mouse testis development (37-39). However, examination of testis from PDGFR-β -/- mice at E13.5 by Brennan et al. (10) and our analysis of PDGFR-β and PDGF-B null mice gonads at E18.5, revealed no overt defects, ruling out a critical role for this receptor during prenatal testis development. This imply that the effect of PDGF-B and PDGFR-β inhibition seen in in vitro and ex vivo experiments on prenatal testis could be masked in vivo by other systems. The persistence of PDGFR-β in the gonocytes and of PDGF-B and PDGF-D in Sertoli cells during the first 5 days after birth led us to focus our study on their potential roles in the postnatal testicular development. We could not use the PDGFR-β or PDGF-B -/- models because of the lethality of the targeting of these genes right before birth (25, 26), thus, we administered the PDGFRs antagonist imatinib in mice from P1 to P5. This time interval was selected to cover exactly the period of neonatal gonocyte proliferation and migration and to avoid the confounding effect on c-kit inhibition that may take place at later times. Indeed, Nurmio et al. reported that imatinib administration in P5-P7 rat, blocked or delayed migration of gonocytes from the center of the seminiferous tubules to the basement membrane, delayed the formation of the germ-line stem cell pool, reduced proliferation of type A spermatogonia normally observed at P8 and induced germ cell apoptosis (40). They attributed all these events to the inhibition of c-kit. However, it is now well established that c-kit is expressed in primordial germ cells (41)

and in differentiated type A spermatogonia (20, 22) but not in gonocytes or in the germline stem cells both in mouse and in rat (42, 23). Accordingly, c-kit has been shown to be critical for the survival, migration and proliferation of primordial germ cells (43, 44) and survival and proliferation of type A spermatogonia (45, 46). The pattern of successive expression, disappearance and reexpression of c-kit indicates that the timing of administration of a compound with multiple targets, such as imatinib, is critical to establish the identity of the affected molecule. The time interval selected by Nurmio et al. (40), spans the period of the initial expression of c-kit by spermatogonia but also the period of expression of PDGFR-β by gonocytes. Thus, we strongly suspect that, while the reduced proliferation of spermatogonia following imatinib administration from P5 to P7 may be due to c-kit inhibition, the effect of imatinib on gonocytes, should be assigned to the PDGFR-β blockage probably occurring during the first day of treatment. To better characterize the protein kinases targeted by imatinib and to evaluate the extent of imatinib inhibition of their signaling, we analyzed expression and phosphorylation status of PDGFR-β, c-kit and PDGFR-α proteins in testis and purified gonocytes by Western blotting. PDGFR-β, PDGFR-α and a faint c-kit signal were found in the testis protein extracts, while gonocytes expressed PDGFR-β solely. These results, coupled with the immunohistochemical findings, confirm that during the first 5 postnatal days, within the testicular tubule, the Sertoli cells are the source of the PDGFRs ligands and gonocytes express PDGFR-β. The lack of c-kit and PDGFR-α expression in gonocytes confirms that PDGFR-β is the only target of imatinib in this cell type. In agreement with previous studies, the PDGFR-α and c-kit immunoreactive bands found in the whole testis protein extracts may be of Leydig cell origin (11, 20), and a portion of PDGFR-β may originate from Leydig cells and PMC (16, 47). Interestingly, the antibody that recognized the conventional form of PDGFR-β, also detected an immunoreactive band of ∼ 70 kDa with ability to autophosphorylate in both testis and purified gonocytes. We did not determine the sequence of this protein, however, the fact that the protein migrates in the predicted
position, autophosphorylates and is recognized by the same antibody used to identify V1-PDGFR-β in rat gonocytes (15), suggests that it may be homologous to V1-PDGFR-β. Thus, the inhibition of gonocytes derived proteins phosphorylation after in vivo administration of imatinib relates to PDGFR-β and to a V1-PDGFR-β-like protein. Inhibition of PDGFR signaling had immediate deleterious effects in the mice testis. The small testes observed following imatinib treatment persisted into adulthood, confirming that PDGFR-mediated actions are required for testicular development. Based on histological, immunohistochemical and molecular examination of the testis of imatinib treated animals, we found that the number of gonocytes was severely reduced and that this reduction is reflected in an impoverishment of spermatogonia. The decrease of gonocytes proliferation following silencing of PDGFR-β may contribute to the testicular phenotype and to the overall reduction of the testicular size that occurs in a background of normal testosterone. Indeed, Culty et al. have shown that PDGF is able to activate the proliferation of isolated rat gonocytes in a dose dependent manner (13, 14). They predicted that PDGF might act as physiological regulator of gonocytes development. Our data demonstrate, for the first time, the involvement of the PDGFR-β/PDGFR-β ligands system in gonocytes development in vivo. An alternative explanation for the reduced testis volume, with qualitatively normal spermatogenesis, could be a reduction of Sertoli cell number. However, the lack of PDGFRs and c-kit expression by Sertoli cells seems to exclude this possibility. Accordingly, Sertoli cell proliferation and number during imatinib administration, were normal. Other potential targets of imatinib action, Leydig cells and PMC, revealed no overt defects as well. This was not unexpected, since the Leydig cell defect seen in PDGF-A -/- mice take place at puberty (11) and PDGFR-α targeting inhibits specification and expansion of fetal Leydig cells in E13.5 mouse (10). Concerning the PMC, it has been noted that the most rapid differentiation of PMC in the mouse is between P7 and P14 and coincides with the first appearance of meiotic stages in the germinal epithelium (48). Thus, the lack of effect of imatinib administration from P1 to P5

on Leydig cells and PMC can be explained with the coincidence of imatinib administration with a period of quiescence in the developmental process of these cells. The migration of the gonocytes from the center of the seminiferous tubule to the basal membrane was reduced by imatinib, indicating that the final location of gonocytes depends on PDGFR-β activation. This was reinforced by the demonstration that both ligands for PDGFR-β, PDGF-B and PDGF-D, attract chemotactically gonocytes in vitro. It is well established that, in addition to mitogenic stimulation, activation of PDGFR-β can induce chemotaxis in a variety of cells (49). Previous studies in the rat, claimed that KL and its receptor c-kit could be involved in gonocytes migration (50). However, the migratory activity was indirectly inferred by the development of pseudopods by gonocytes cocultured with Sertoli cells in the presence of KL. In these experimental conditions, the percentage of gonocytes with pseudopods was only slightly increased by KL or decreased by an anti-c-kit antiserum, suggesting that different signals originating from Sertoli cells could be responsible for this phenomenon. Whether the intratubular PDGFR-β/PDGFR-β-ligands system or even the V1-PDGFR-β-like protein activation are important for the migration of gonocytes in vivo cannot be ascertained at this time, without the ability to directly image the migration of such cells in vivo in real time. At present, what we can say is that the chemotactic effect of PDGF-B and PDGF-D on the gonocytes, in vitro, can be clearly assigned only to the activation of the full length PDGFR-β and not to the V1-PDGFR-β-like protein that lacks the ligand binding domain. Not all gonocyes were affected by imatinib. Regardless of treatment, at P5 some gonocytes located to the basal membrane. One possible explanation is that the gonocytes are heterogeneous and the silencing of PDGFR β-subunit signaling influences the fate of a subset of cells. In line with this idea, Orwing et al. (51) reported that neonatal rat gonocytes exhibit a morphological heterogenity closely related to their transplantable stem cell activity. Furthermore, Yoshida et al. showed that a subset of gonocytes differentiate promptly in the first round of spermatogenesis while another set supports the continuity of
steady-state spermatogenesis (52). Accordingly, the first round of spermatogenesis was lacking in imatinib treated animals. This was indicated by the lack of spermatocytes at P15, the reduced sperm output at P35, the time when the first spermatozoa are released (53), and the absence of c-kit, a marker of differentiating spermatogonia, at P8. The inhibition of gonocytes migration and the decreased number of spermatogonia did not cause a permanent defect on sperm production and a progressive recovery of spermatogenesis and sperm cells output were observed. This suggests that the few spermatogonia found in the imatinib treated animals mantain the capacity of colonize the seminiferous tubules. In conclusion, our experiments have identified PDGFR-β as a key player in the pathway that orchestrates the cellular development of the early postnatal testis. Signaling through this receptor is critical for important events that dictate the fate of gonocytes, the precursors of the SSCs, including resumption of proliferation, inhibition of apoptosis and migration. Analogously to what reported in neurons (54), PDGFR activation could act as a mitogen in the early phase of stem cell differentiation to expand the pool of immature gonocytes. Furthermore, PDGFR-β could stimulate the differentiation of gonocytes toward SSCs. Interestingly, activation of PDGFR-β by Piwil2, a protein of the Argonaute family of proteins implicated in germline and stem cell development (55), may play a role in the differentiation of gonocytes toward spermatogonial stem cells (56). Our studies cannot distinguish between the effects of PDGFR-β and of the PDGFR-β variant V1-PDGFR-β. Wang and Culty have recently hypotesized that, in the rat testis, V1-PDGFR-β could be a dominant negative form of PDGFR that may compete with the full-length receptor and in this manner could modulate responses to PDGF (15). Future experiments will clarify the exact role of V1-PDGFR-β in the development of the testis. Whether the PDGFR-β dependent mechanisms of gonocytes development identified in the present study apply in humans remains to be determined. However, it is important to notice that, since PDGFR-β tyrosine kinase inhibition is a major target of anticancer drug development, the administration of these

compounds in children might cause testicular damage. Accordingly, oligozoospermia and spermatogenesis failure in children treated with imatinib have been reported (57, 58).
Acknowledgments We thank Dr. C. Betsholtz and Dr. P. Soriano for providing PDGF-B +/- and PDGFR-β +/- mice. This work was supported in part by the Italian Ministero dell’Università e della Ricerca (MIUR), Progetti di Ricerca di Interesse Nazionale, PRIN 2005.

References
1. Nagano R, Tabata S, Nakanishi Y, Ohsako S, Kurohmar M, Hayashi Y 2000
Reproliferation and relocation of mouse male germ cells (gonocytes) during prespermatogenesis. Anat Rec 258:210-220
2. Clermont, Y, Perey B 1957 Quantitative study of the cell population of the seminiferous tubules in immature rats. Am J Anat 100:241-267
3. Hilscher B, Hilscher W, Bulthoff-Ohnolz B, Kramer U, Birke A, Pelzer H, Gauss G 1974 Kinetics of gametogenesis. I. Comparative histological and autoradiographic studies of oocytes and transitional prospermatogonia during oogenesis and prespermatogenesis. Cell Tissue Res 154:443-470
4. Huckins C, Clermont, Y 1968 Evolution of gonocytes in the rat testis during late embryonic and early post-natal life. Arch Anat Histol Embryol 51:341-354
5. Roosen-Runge EC, Leik J 1968 Gonocyte degeneration in the postnatal male rat. Am J Anat 122:275-300
6. Gnessi L, Fabbri A, Spera G 1997 Gonadal peptides as mediators of development and functional control of the testis: an integrated system with hormones and local environment. Endocr Rev 18:541-609
7. Heldin CH, Westermark B 1999 Mechanism of action and in vivo role of platelet-derived growth factor. Physiol Rev 79:1283–1316
8. Betsholtz C, Karlsson L, Lindahl P 2001 Developmental roles of platelet-derived growth factors. Bioessays 23:494–507
9. Hoch RV, Soriano P 2003 Roles of PDGF in animal development. Development 130:4769–4784
10. Brennan J, Tilmann C, Capel B 2003 Pdgfr-α mediates testis cord organization and fetal Leydig cell development in the XY gonad. Genes Dev 17:800-810
11. Gnessi L, Basciani S, Mariani S, Arizzi M, Spera G, Wang C, Bondjers C, Karlsson L, Betsholtz C 2000 Leydig cell loss and spermatogenic arrest in platelet-derived growth factor (PDGF)-A-deficient mice. J Cell Biol 149:1019-1025
12. Ge R-S, Dong Q, Sottas CM, Papadopoulos V, Zirkin BR, and Hardy P 2006 In search of rat stem Leydig cells: identification, isolation, and lineage-specific development. Proc Natl Acad Sci USA 103:2719-2724
13. Li H, Papadopoulos V, Vidic B, Dym M, Culty M 1997 Regulation of rat testis gonocyte proliferation by platelet-derived growth factor and estradiol: identification of signaling mechanisms involved. Endocrinology 138:1289-1298
14. Thuillier R, Wang Y, Culty M 2003 Prenatal exposure to estrogenic compounds alters the expression pattern of platelet-derived growth factor receptors α and β in neonatal rat testis: identification of gonocytes as targets of estrogen exposure. Biol Reprod 68:867-880
15. Wang Y, Culty M 2007 Identification and distribution of a novel platelet-derived growth factor receptor β variant. Effect of retinoic acid and involvement in cell differentiation. Endocrinology 148:2233-2250
16. Gnessi L, Emidi A, Jannini EA, Carosa E, Maroder M, Arizzi M, Ulisse S, Spera G 1995 Testicular development involves the spatiotemporal control of PDGFs and PDGF receptors gene expression and action. J Cell Biol 131:1105-1121
17. Hasthorpe S, Barbic S, Farmer PJ, Hutson JM 1999 Neonatal mouse gonocyte proliferation assayed by an in vitro clonogenic method. J Reprod Fertil 116:335-344
18. Basciani S, Mariani S, Arizzi M, Ulisse, S, Rucci N, Jannini EA, Della Rocca C, Manicone A, Carani C, Spera, G, Gnessi L 2002 Expression of platelet-derived growth factor-A (PDGF-A), PDGF-B, and PDGF receptor-alpha and -beta during human testicular development and disease. J Clin Endocrinol Metab 87:2310-2319

19. Deininger M, Buchdunger E, Druker BJ 2005 The development of imatinib as a therapeutic agent for chronic myeloid leukemia. Blood 105:2640-2653
20. Manova K, Nocka K, Besmer P, Bachvarova RF 1990 Gonadal expression of c-kit encoded at the W locus of the mouse. Development 110:1057-1069
21. Schrans-Stassen BH, van de Kant HJ, de Rooij DG, van Pelt AM 1999 Differential expression of c-kit in mouse undifferentiated and differentiating type A spermatogonia. Endocrinology 140:5894-5900
22. Sorrentino V, Giorgi M, Geremia R, Besmer P, Rossi P 1991 Expression of the c-kit proto-oncogene in the murine male germ cells. Oncogene 6:149-151
23. Prabhu SM, Meistrich ML, McLaughlin EA, Roman SD, Warne S, Mendis S, Itman C, Loveland KL 2006 Expression of c-Kit receptor mRNA and protein in the developing, adult and irradiated rodent testis. Reproduction 131:489-499
24. le Coutre P, Mologni L, Cleris L, Marchesi E, Buchdunger E, Giardini R, Formelli F, Gambacorti-Passerini C 1999 In vivo eradication of human BCR/ABL-positive leukemia cells with an ABL kinase inhibitor. J Natl Cancer Inst 20:163-168.
25. Soriano P 1994 Abnormal kidney development and hematological disorders in PDGF beta-receptor mutant mice. Genes Dev 16:1888-1896
26. Leveen P, Pekny M, Gebre-Medhin S, Swolin B, Larsson E, Betsholtz C 1994 Mice deficient for PDGF B show renal, cardiovascular, and hematological abnormalities. Genes Dev 16:1875-1887
27. Russell LD, Ettlin RA, Sinha Hikim AP, Clegg ED 1990 Staging for the laboratory mouse. In: Russell LD, Ettlin RA, Sinha Hikim AP, Clegg ED eds Histological and Histopathological Evaluation of the Testis. Vienna, IL: Cache River Press: 119-161
28. Basciani S, Brama M, Mariani S, De Luca G, Arizzi M, Vesci L, Pisano C, Dolci S, Spera G, Gnessi L 2005 Imatinib mesylate inhibits Leydig cell tumor growth: evidence for in vitro and in vivo activity. Cancer Res 65:1897-1903
29. van Dissel-Emiliani FM, de Rooij DG, Meistrich ML 1989 Isolation of rat gonocytes by velocity sedimentation at unit gravity. J Reprod Fertil 86:759-667
30. van Dissel-Emilian FM, De Boer-Brouwer M, De Rooij DJ 1996 Effect of fibroblast growth factor-2 on Sertoli cells and gonocytes in coculture during the perinatal period. Endocrinology 137:647-654
31. Basciani S, Mariani S, Arizzi M, Brama M, Ricci A, Betsholtz C, Bondjers C, Ricci G, Catizone A, Galdieri M, Spera G, Gnessi L 2004 Expression of platelet-derived growth factor (PDGF) in the epididymis and analysis of the epididymal development in PDGF-A, PDGF-B, and PDGF receptor beta deficient mice. Biol Reprod 70:168-177
32. Orth JM 1993 Cell biology of testicular development in the fetus and neonate. In: Desjardins C, Ewing LL eds. Cell and Molecular Biology of the Testis. Oxford: Oxford University Press: 3–42
33. Chang H, Matzuk M 2001 Smad5 is required for mouse primordial germ cell development. Mech Dev 104:61-67
34. Pellegrini M, Grimaldi P, Rossi P, Geremia R, Dolci S 2003 Developmental expression of BMP4/ALK3/SMAD5 signaling pathway in the mouse testis: a potential role of BMP4 in spermatogonia differentiation. J Cell Sci 116:3363-3372
35. Naughton CK, Jain S, Strickland AM, Gupta A, Milbrandt J 2006 Glial cell-line derived neurotrophic factor-mediated RET signaling regulates spermatogonial stem cell fate. Biol Reprod 2:314-321
36. Chen C, Ouyang W, Grigura V, Zhou Q, Carnes K, Lim H, Zhao GQ, Arber S, Kurpios N, Murphy TL, Cheng AM, Hassell JA, Chandrashekar V, Hofmann MC, Hess RA, Murphy KM 2005 ERM is required for transcriptional control of the spermatogonial stem cell niche. Nature 436:1030-1034
37. Uzumcu M, Dirks KA, Skinner MK 2002 Inhibition of platelet-derived growth factor actions in the embryonic testis influences normal cord development and morphology. Biol Reprod 66:745-753

38. Puglianiello A, Campagnolo L, Farini D, Cipollone D, Russo MA, Siracusa G 2004 Expression and role of PDGF-BB and PDGFR-β during testis morphogenesis in the mouse embryo. J Cell Sci 117:1151-1160
39. Ricci G, Catizone A, Galdieri M 2004 Embryonic mouse testis development: role of platelet derived growth factor (PDGF-BB). J Cell Physiol 200:458-467
40. Nurmio M, Toppari J, Zaman F, Andersson AM, Paranko J, Soder O, Jahnukainen K 2007 Inhibition of tyrosine kinases PDGFR and C-Kit by imatinib mesylate interferes with postnatal testicular development in the rat. Int J Androl 4:366-376
41. Lacham-Kaplan O 2004 In vivo and in vitro differentiation of male germ cells in the mouse. Reproduction 128:147–152
42. Ohta H, Yomogida K, Dohmae K, Nishimune Y 2000 Regulation of proliferation and differentiation in spermatogonial stem cells: the role of c-kit and its ligand SCF. Development 127:2125–2131
43. Sette C, Dolci S, Geremia R, Rossi P 2000 The role of stem cell factor and of alternative c-kit gene products in the establishment, maintenance and function of germ cells. Int J Dev Biol 44:599–608
44. Lacham-Kaplan O 2004 In vivo and in vitro differentiation of male germ cells in the mouse. Reproduction 128:147–152
45. Petch LA, Harris J, Raymond VW, Blasband A, Lee DC, Earp HS 1990 A truncated, secreted form of the epidermal growth factor receptor is encoded by an alternatively spliced transcript in normal rat tissue. Mol Cell Biol 10:2973–2982
46. Ohta H, Yomogida K, Dohmae K, Nishimune Y 2000 Regulation of proliferation and differentiation in spermatogonial stem cells: the role of c-kit and its ligand SCF. Development 127:2125–2131
47. Gnessi L, Emidi A, Scarpa S, Palleschi S, Ragano-Caracciolo M, Silvestroni L, Modesti A, Spera G 1993 Platelet-derived growth factor effects on purified testicular peritubular myoid cells: binding, cytosolic Ca2+ increase, mitogenic activity, and extracellular matrix production enhancement. Endocrinology 133:1880-1890
48. Bressler RS, Ross MH 1972 Differentiation of peritubular myoid cells of the testis: effects of intratesticular implantation of newborn mouse testes into normal and hypophysectomized adults. Biol Reprod 6:148-159
49. Ronnstrand L, Heldin CH 2001 Mechanisms of platelet-derived growth factor-induced chemotaxis. Int J Cancer 91:757-762
50. Orth JM, Qiu J, Jester WF Jr, Pilder S 1997 Expression of the c-kit gene is critical for migration of neonatal rat gonocytes in vitro. Biol Reprod 57:676-683
51. Orwig KE, Ryu BY, Avarbock MR, Brinster RL 2002 Male germ-line stem cell potential is predicted by morphology of cells in neonatal rat testes. Proc Natl Acad Sci USA 99:11706-11711
52. Yoshida S, Sukeno M, Nakagawa T, Ohbo K, Nagamatsu G, Suda T, Nabeshima Y 2006 The first round of mouse spermatogenesis is a distinctive program that lacks the self-renewing spermatogonia stage. Development 133:1495-1505
53. Albert M, Roussel C 1983 Changes from puberty to adulthood in the concentration, motility and morphology of mouse epididymal spermatozoa. Int J Androl 5:446-460
54. Erlandsson A, Brannvall K, Gustafsdottir S, Westermark B, Forsberg-Nilsson K 2006 Autocrine/paracrine platelet-derived growth factor regulates proliferation of neural progenitor cells. Cancer Res 66:8042-8048
55. Seto AG, Kingston RE, Lau NC 2007 The coming of age for piwi proteins. Mol Cell 26:603-609
56. Lee JH, Engel W, Nayernia K 2006 Stem cell protein Piwil2 modulates expression of murine spermatogonial stem cell expressed genes. Mol Reprod Dev 73:173-179
57. Seshadri T, Seymour JF, McArthur GA 2004 Oligospermia in a patient receiving imatinib therapy for the hypereosinophilic syndrome. N Engl J Med 351:2134-2135
58. Mariani S, Giona F, Basciani S, Brama M and Gnessi L 2008 Low bone density, slowed growth rate, gynecomastia and decreased inhibin-B/FSH ratio in a boy treated with imatinib during puberty. The Lancet (in press)

Figures Legends
Figure 1. Immunohistochemical analysis of PDGFR β-subunit, PDGF-B, and PDGF-D expression in
E18.5 and P5.0 mice testis. The antibody used to mark PDGFR-β recognizes the C-terminal portion of
the receptor and does not distinguish between PDGFR-β and the truncated form of PDGFR-β, V1-
PDGFR-β. The receptor expression is restricted to gonocytes (arrows) and PDGF-B and PDGF-D are
seen in Sertoli cells (arrowheads). PDGFR β-subunit (A, D), PDGF-B (B, E) and PDGF-D (C, F) in
E18.5 (A, B, C) and P5.0 (D, E, F) testis. Scale bar, 20 μm.
Figure 2. PDGFR-β and PDGF-B gene targeting does not affect testis development before birth.
Hematoxylin/eosin stained sections of PDGFR-β (A) and PDGF-B (C) +/- testis compared with
PDGFR-β (B) and PDGF-B (D) -/- testis at E18.5. Scale bar, 20 μm.
Figure 3. Whole testis and purified mouse gonocytes express PDGFR-β and a V1-PDGFR-β-like
protein and imatinib treatment inhibits their phosphorylation in vivo and in vitro. A) Representative
Westerm blot on protein extract from testes excised from one 3-day-old control mouse and one mouse
treated with 50 mg/kg imatinib 2 hours before sacrifice and (B) densitometric analysis of the bands
expressed as a ratio between the protein being analyzed and β-actin in control ( ) and imatinib
treated ( ) animals. *, P<0.05 versus corresponding control value. Columns, mean ± SEM of four
experiments. C) Representative Western blot on protein extract from gonocytes purified from 3-day-
old mouse testes treated with imatinib in vitro and (D) densitometric analysis of the bands detectable,
expressed as a ratio between the protein being analyzed and β-actin. a, P<0.05 versus PDGF-B treated
cells; b, P<0.05 versus imatinib-/PDGF-B- and imatinib-/PDGF-B+ treated cells (ANOVA). Columns,
mean ± SEM of four experiments. Gonocytes were incubated for 4 hours in medium 0.1% FBS either
in the presence or in the absence of 10 μmol/L imatinib followed by an additional incubation in the
presence or absence of PDGF-BB at 10 ng/mL or KL at 100 ng/mL. The right column of panel C
shows a representative Western blot on protein extract from MCF7 cells used as positive control for
the antibodies that did not react in the gonocytes protein extracs. The filters were probed with anti-p-
PDGFR-α, anti-p-PDGFR-β or anti-p-c-kit antibodies to reveal the status of PDGFRs and c-kit
signaling after treatment. The immunoblots were also probed with anti-PDGFR-α, anti-PDGFR-β,
anti-c-kit or anti-β-actin antibodies. The blots presented in the upper part of panel A, and lower part of
panel C, came from the same blots respectively. The original blots were cut and rearranged for clarity.
Figure 4. Long term effect of imatinib treatment during the first 5 post-natal days on body weight and
testis weight. A) In imatinib treated animals, average body weight was significantly reduced up to P45
(mean ± SEM of 5 animals for each point). At later ages the body weight of the treated animals was

not different from controls. B) Testis weight was significantly decreased in treated animals compared
with controls (mean ± SEM of 10 testes for each point). *, P<0.05; ***, P<0.001
Figure 5. Immunohistochemical staining with anti SMAD-5, anti RET, anti PDGFR β-subunit and
anti-c-kit antibody in one control (A, C, E, G) and one imatinib treated (B, D, F, H) animal at P5.
Immunoreaction with anti c-kit antibody in one control (I) and one imatinib treated (J) animal at P8.
Treated animals were injected intraperitoneally with imatinib, 50 mg/kg, three times a day from P1 to
P5. Arrows indicate examples of positive cells. Similar results were obtained in three different
experiments. Scale bar, 50 μm.
Figure 6. Testicular histology of mice treated with imatinib and pattern of reduction of spermatogonia
specific genes. A) Histology of control and imatinib treated testis at P15, P70 and P120. Roman
numbers indicate the stage of the seminiferous epithelium. Scale bars, 50 μm (P15); 100 μm (P70 and
P120). B) Number of Sertoli cells (SC), spermatogonia (Sg), spermatocytes (Sc), round spermatids
(rSd) and elongated spermatids (eSd) per tubule cross-section in P70 mice treated with imatinib ( )
versus untreated control ( ). Each column is the mean±SEM of the cell count of 30 circular tubules
of testis from four animals. C) Representative RT-PCR analysis of control and imatinib treated
littermates at P5, P15, P70 and P120 are shown for the indicated genes. D) Densitometric analysis of
RT-PCRs expressed as a ratio between the gene being analyzed and β-actin in control ( ) and
imatinib treated ( ) animals. Columns, mean ± SEM of four experiments. The treated mice received
imatinib (50 mg/kg) intraperitoneally three times a day on postnatal days 1-5, while control animals
received vehicle alone. *, P < 0.05 compared with the corresponding control value.
Figure 7. Imatinib administration inhibits proliferation and induces apoptosis and PDGF-B and
PDGF-D are chemotactic for gonocytes. Proliferating cells (A) and apoptotic cells (B) in testis of
imatinib treated and control mice. Each point represents the mean ± SEM of the percentage of BrdU or
TUNEL positive gonocytes of ten HPF (x200) per testis cross-sections of three animals for each age
group. C) Chemotactic effect of the PDGFR-β ligands PDGF-B and PDGF-D on gonocytes from 3-
day-old testis in the absence or in the presence of imatinib. Cell migration is expressed as a migration
index and calculated as the x-fold increase over the negative control (unstimulated cells). Figure
shows the mean ± SEM of three experiments. PDGF-B and PDGF-D showed the maximal number of
migrating cells at 10 ng/ml. The chemotactic effect of PDGFs was inhibited by imatinib 10 μM.
Copyright © 2022 FDOKUMEN




















