
Phosphorylation of a major GPI-anchored surface protein of Trypanosoma brucei during transport to...
11
1785 INTRODUCTION African trypanosomes cycle between the mammalian host and the tsetse fly (Vickerman, 1985). In the mammalian host, bloodstream forms of Trypanosoma brucei are covered by a dense uniform coat of variant surface glycoprotein (VSG) molecules. Following ingestion by a tsetse fly, as part of a blood meal, the parasites differentiate into procyclic forms over a period of two to three days. After differentiation, the parasite loses the VSG coat and is covered instead by invariant surface glycoproteins termed procyclins. In T. brucei procyclic forms this coat consists of EP and GPEET forms of procyclins (reviewed by Roditi et al., 1998). The EP procyclins are a group of closely related proteins that are encoded by six or seven genes and contain extensive tandem repeat units of glutamic acid and proline (E and P, respectively, in the single-letter amino acid code) (Roditi et al., 1987; Mowatt and Clayton, 1987, 1988; Mowatt et al., 1989). GPEET procyclin is encoded by two identical genes and contains internal pentapeptide (GPEET) repeats (Mowatt et al., 1989; Koenig et al., 1989). The EP procyclins (Richardson et al., 1988) and the corresponding genes (Roditi et al., 1987; Mowatt and Clayton, 1987, 1988) were first characterized more than a decade ago. In contrast, the expression of GPEET at the protein level has only recently been reported (Ruepp et al., 1997; Treumann et al., 1997; Bütikofer et al., 1997), although the gene was identified much earlier by Mowatt et al. (1989). GPEET was first shown to be co-expressed with EP on the surface of T. brucei procyclic forms (Ruepp et al., 1997). Subsequently, the GPEET protein was isolated and characterized independently by two different laboratories (Treumann et al., 1997; Bütikofer et al., 1997). In common with EP (Field et al., 1991), GPEET was found to be attached to the cell surface via a glycosylphosphatidylinositol (GPI) anchor (Treumann et al., 1997; Bütikofer et al., 1997). The structures of the EP and GPEET GPI moieties are indistinguishable from each other and consist of a GPI core structure that is substituted by heterogeneous sialylated polylactosamine-containing side chains (Ferguson et al., 1993; Treumann et al., 1997). This heavy glycosylation, together with the proposed extended Journal of Cell Science 112, 1785-1795 (1999) Printed in Great Britain © The Company of Biologists Limited 1999 JCS0342 The surface coat of procyclic forms of Trypanosoma brucei consists of related, internally repetitive glycoproteins known as EP and GPEET procyclins. Previously we showed that the extracellular domain of GPEET is phosphorylated. We now show that phosphorylation of this glycosylphosphatidylinositol-anchored surface protein can be induced in vitro using a procyclic membrane extract. Using antibodies that recognize either the phosphorylated or unphosphorylated form of GPEET, we analyzed their expression during differentiation of bloodstream forms to procyclic forms. Unphosphorylated GPEET, together with EP, was detected in cell lysates 2-4 hours after initiating differentiation whereas phosphorylated GPEET only appeared after 24 hours. Surface expression of EP and both forms of GPEET occurred after 24-48 hours and correlated with the detection of phosphorylated GPEET on immuno- blots. Electron micrographs showed that unphosphorylated GPEET was predominantly in the flagellar pocket whereas the phosphorylated form was distributed over the cell surface. In contrast, expression of a membrane-bound human placental alkaline phosphatase in procyclic forms caused the accumulation of dephosphorylated GPEET on the cell surface, while the phosphorylated form was restricted to the flagellar pocket. A GPEET-Fc fusion protein, which was retained intracellularly, was not phosphorylated. We propose that unphosphorylated GPEET procyclin is transported to a location close to or at the cell surface, most probably the flagellar pocket, where it becomes phosphorylated. To the best of our knowledge, this study represents the first localization of phosphorylated and unphosphorylated forms of a GPI- anchored protein within a cell. Key words: Procyclin, Differentiation, Phosphorylation, GPEET, Trypanosome SUMMARY Phosphorylation of a major GPI-anchored surface protein of Trypanosoma brucei during transport to the plasma membrane Peter Bütikofer 1, *, Erik Vassella 2 , Stefan Ruepp 2 , Monika Boschung 1 , Gianluca Civenni 1 , Thomas Seebeck 2 , Andrew Hemphill 3 , Neeloffer Mookherjee 4 , Terry W. Pearson 4 and Isabel Roditi 2 Institutes of 1 Biochemistry and Molecular Biology, 2 General Microbiology, and 3 Parasitology, University of Bern, Bern, Switzerland 4 Department of Biochemistry and Microbiology, University of Victoria, Canada *Author for correspondence (e-mail: [email protected]) Accepted 22 March; published on WWW 11 May 1999
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Phosphorylation of a major GPI-anchored surface protein of Trypanosoma brucei during transport to...

1785
INTRODUCTION
African trypanosomes cycle between the mammalian host andthe tsetse fly (Vickerman, 1985). In the mammalian host,bloodstream forms of Trypanosoma brucei are covered by adense uniform coat of variant surface glycoprotein (VSG)molecules. Following ingestion by a tsetse fly, as part of a bloodmeal, the parasites differentiate into procyclic forms over aperiod of two to three days. After differentiation, the parasiteloses the VSG coat and is covered instead by invariant surfaceglycoproteins termed procyclins. In T. brucei procyclic formsthis coat consists of EP and GPEET forms of procyclins(reviewed by Roditi et al., 1998). The EP procyclins are a groupof closely related proteins that are encoded by six or sevengenes and contain extensive tandem repeat units of glutamicacid and proline (E and P, respectively, in the single-letter aminoacid code) (Roditi et al., 1987; Mowatt and Clayton, 1987,1988; Mowatt et al., 1989). GPEET procyclin is encoded by twoidentical genes and contains internal pentapeptide (GPEET)repeats (Mowatt et al., 1989; Koenig et al., 1989).
The EP procyclins (Richardson et al., 1988) and thecorresponding genes (Roditi et al., 1987; Mowatt and Clayton,1987, 1988) were first characterized more than a decade ago.In contrast, the expression of GPEET at the protein level hasonly recently been reported (Ruepp et al., 1997; Treumann etal., 1997; Bütikofer et al., 1997), although the gene wasidentified much earlier by Mowatt et al. (1989). GPEET wasfirst shown to be co-expressed with EP on the surface of T.brucei procyclic forms (Ruepp et al., 1997). Subsequently, theGPEET protein was isolated and characterized independentlyby two different laboratories (Treumann et al., 1997; Bütikoferet al., 1997). In common with EP (Field et al., 1991), GPEETwas found to be attached to the cell surface via aglycosylphosphatidylinositol (GPI) anchor (Treumann et al.,1997; Bütikofer et al., 1997). The structures of the EP andGPEET GPI moieties are indistinguishable from each other andconsist of a GPI core structure that is substituted byheterogeneous sialylated polylactosamine-containing sidechains (Ferguson et al., 1993; Treumann et al., 1997). Thisheavy glycosylation, together with the proposed extended
Journal of Cell Science 112, 1785-1795 (1999)Printed in Great Britain © The Company of Biologists Limited 1999JCS0342
The surface coat of procyclic forms of Trypanosoma bruceiconsists of related, internally repetitive glycoproteinsknown as EP and GPEET procyclins. Previously weshowed that the extracellular domain of GPEET isphosphorylated. We now show that phosphorylation of thisglycosylphosphatidylinositol-anchored surface protein canbe induced in vitro using a procyclic membrane extract.Using antibodies that recognize either the phosphorylatedor unphosphorylated form of GPEET, we analyzed theirexpression during differentiation of bloodstream forms toprocyclic forms. Unphosphorylated GPEET, together withEP, was detected in cell lysates 2-4 hours after initiatingdifferentiation whereas phosphorylated GPEET onlyappeared after 24 hours. Surface expression of EP and bothforms of GPEET occurred after 24-48 hours and correlatedwith the detection of phosphorylated GPEET on immuno-blots. Electron micrographs showed that unphosphorylatedGPEET was predominantly in the flagellar pocket whereas
the phosphorylated form was distributed over the cellsurface. In contrast, expression of a membrane-boundhuman placental alkaline phosphatase in procyclic formscaused the accumulation of dephosphorylated GPEET onthe cell surface, while the phosphorylated form wasrestricted to the flagellar pocket. A GPEET-Fc fusionprotein, which was retained intracellularly, was notphosphorylated. We propose that unphosphorylatedGPEET procyclin is transported to a location close to or atthe cell surface, most probably the flagellar pocket, whereit becomes phosphorylated. To the best of our knowledge,this study represents the first localization ofphosphorylated and unphosphorylated forms of a GPI-anchored protein within a cell.
Key words: Procyclin, Differentiation, Phosphorylation, GPEET,Trypanosome
SUMMARY
Phosphorylation of a major GPI-anchored surface protein of Trypanosoma
brucei during transport to the plasma membrane
Peter Bütikofer1,*, Erik Vassella2, Stefan Ruepp2, Monika Boschung1, Gianluca Civenni1, Thomas Seebeck2,Andrew Hemphill3, Neeloffer Mookherjee4, Terry W. Pearson4 and Isabel Roditi2
Institutes of 1Biochemistry and Molecular Biology, 2General Microbiology, and 3Parasitology, University of Bern, Bern, Switzerland4Department of Biochemistry and Microbiology, University of Victoria, Canada*Author for correspondence (e-mail: [email protected])
Accepted 22 March; published on WWW 11 May 1999

1786
peptide conformation of EP (Roditi et al., 1989) and GPEET(Treumann et al., 1997), may account for their unusualmigration on SDS-PAGE (see for example Bütikofer et al.,1997; Treumann et al., 1997). The biosynthesis of GPIstructures in both bloodstream and procyclic formtrypanosomes has been well characterized and occurs in theendoplasmic reticulum (ER) (McConville and Ferguson, 1993;Vidugiriene and Menon, 1995). After translocation of thepolypeptide precursor into the ER, attachment of the GPIanchor is mediated by a C-terminal GPI attachment signalpeptide via a reaction involving a transamidase (Udenfriendand Kodukula, 1995). Recently it has been shown in transgenicprocyclic trypanosomes that the deletion of the GPI anchorattachment signal of VSG resulted in reduced rates of transportof the truncated protein to the cell surface and its release as asoluble protein (Bangs et al., 1997). Despite the absence of aGPI anchor, the truncated protein seemed to fold properly butaccumulated in the ER (McDowell et al., 1998). These dataindicate that the addition of a GPI anchor may provide a signalfor rapid protein transport from the ER to the cell surface inprocyclic trypanosomes. Similar observations have been madebefore in mammalian cells and yeast by inhibiting GPI anchorbiosynthesis or mutating the GPI attachment signal peptide(Moran and Caras, 1992; Delahunty et al., 1993; Field et al.,1994; Doering and Schekman, 1996). Perhaps the GPI anchoracts as a specific ligand that directs the protein to vesicles inthe secretory pathway or, alternatively, provides some physicalproperty that forms the basis for selection of GPI-anchoredproteins for rapid transport to the plasma membrane.
Although the two forms of procyclin are structurally related,they appear to have different roles: Ruepp et al. (1997)generated mutants with no EP genes and only one copy of aGPEET gene and found that these cells did not show an alteredphenotype in culture, but had a reduced ability to establishheavy infections in tsetse fly midguts. These results indicatethat EP is not required in culture but is important for survivalof trypanosomes in the insect host, possibly by mediatingparasite-fly interactions (Ruepp et al., 1997). In contrast, thelast GPEET gene could not be deleted unless another copy ofGPEET was introduced into a different locus (Ruepp et al.,1997). Furthermore, it has been reported that GPEETexpression may change with the time that procyclic forms arekept in culture; in long-term cultures of T. brucei 427, GPEET,and not EP, often represents the major form of procyclin(Treumann et al., 1997; Bütikofer et al., 1997; Roditi et al.,1998). These results suggest that, at least in certain strains,GPEET may play an important role in optimal cell growth.
In our earlier work on GPEET procyclin (Bütikofer et al.,1997) we reported that a polyclonal antibody raised against thesynthetic pentapeptide repeat (GPEET)3C coupled to keyholelimpet haemocyanin only poorly recognized GPEET unless theprotein was first treated with alkaline phosphatase. Theseresults suggested that native GPEET might be phosphorylated(Bütikofer et al., 1997). The phosphorylation of GPEET wasproposed to occur on the Thr in one or several of the 6 identical(E)TGPE(E) repeat units (Bütikofer et al., 1997; underlinedamino acids are essential, and those in parenthesis stimulatephosphorylation by casein kinase II; Pinna, 1990). Veryrecently, the phosphorylation of GPEET has been confirmedusing chemical and mass spectrometric methods (Mehlert etal., 1999).
At present, relatively little is known about thephosphorylation of extracellularly oriented proteins. Inmammalian cells (Kübler et al., 1982, 1983; Sonka et al., 1989;Volonté et al., 1994; Apasov et al., 1996; Merlo et al., 1997)and parasites (Lester et al., 1990; Tomavo et al., 1992;Hermoso et al., 1994; Sacerdoti-Sierra and Jaffe, 1997)phosphorylation of membrane surface proteins and solubleextracellular proteins has been proposed to occur through theaction of protein kinases localized either on the cell surface orreleased into the extracellular medium. Several types of ecto-kinases have been identified that phosphorylate bothendogenous and exogenous substrates on serine/threonine ortyrosine residues. However, the roles of these enzymes, and thefunctional consequences of cell surface proteinphosphorylation, are still unclear.
Here we report the results of our studies on the expressionof GPEET procyclin during differentiation from bloodstreamforms to procyclic forms of T. brucei 427. Particular attentionwas paid to the distribution of unphosphorylated andphosphorylated forms of GPEET in procyclic cells in anattempt to localize the site of the phosphorylation reaction.
MATERIALS AND METHODS
MaterialsUnless otherwise stated, all reagents were of analytical grade and werefrom Fluka (Buchs, Switzerland), Sigma (St Louis, MO), or Merck(Darmstadt, Germany). [γ-32P]ATP was purchased from NEN LifeSciences Products (Geneva, Switzerland). Acrylamide solution wasfrom National Diagnostics (Hull, UK), alkaline phosphatase from calfintestine (grade II) from Boehringer (Mannheim, Germany) andProtein G-Sepharose from Pharmacia (Dübendorf, Switzerland).
AntibodiesPolyclonal antibodies against GPEET procyclin, termed K1, wereraised against a synthetic peptide, (GPEET)3C, conjugated to keyholelimpet haemocyanin (Ruepp et al., 1997). The anti-EP procyclinmonoclonal antibody TRBP1/247 (mAb 247) has been describedpreviously (Richardson et al., 1986, 1988). The anti-GPEET mAb5H3 was produced as follows: 3 BALB/c mice were immunized byintraperitoneal injection with 107 live EP-deficient Nour 6C procyclicforms (Ruepp et al., 1997) in complete Freund’s adjuvant. After asecond injection in incomplete Freund’s adjuvant three weeks later,sera from the three mice were tested in ELISA against glutaraldehyde-fixed intact procyclic forms and the two mice with the highestantibody titres were selected. A final intravenous injection of 107 Nour6C trypanosomes in the tail vein was followed three days later byremoval of the spleens. Cell fusion, one-step HAT selection andcloning of hybridomas were performed using ClonaCellTM (Stem CellTechnologies Inc., Vancouver, BC) according to the manufacturer’sintructions. One thousand clones were selected and after 3 days ofgrowth in individual microwells their supernatants were screened byELISA on glutaraldehyde-fixed intact Nour 6C cells in order to detectsurface-binding mAbs (Richardson et al., 1986). After screening onhuman transferrin (to eliminate non-specific mAbs) 123 anti-Nour 6CmAbs were selected. The mAbs were then tested in ELISA usingHPLC-purified GPEET from Nour 6C procyclic forms as antigen.Four mAbs were positive. These were tested on immunoblots oflysates of both T. brucei 427 wild-type and Nour 6C procyclic forms.All 4 mAbs detected antigen as a broad dark band of 22-32 kDa inboth wild-type and EP-deficient parasites. The mAb 5H3 was selectedas it gave a very strong signal. The isotype (IgG1) of mAb 5H3 wasdetermined by antigen capture ELISA using a kit from American
P. Bütikofer and others

1787Phosphorylation and transport of GPEET
Qualex (La Mirada, CA). Affinity-purified polyclonal antibodiesagainst VSG variant 221 were obtained from M.L. Cardoso deAlmeida (Sao Paulo, Brazil). The goat anti-human IgG (Fc specific)antibody conjugated to horseradish peroxidase was from Sigma.Horseradish peroxidase-conjugated and FITC-conjugated rabbit anti-mouse and swine anti-rabbit antibodies were purchased from Dako(Copenhagen, Denmark).
TrypanosomesThe parental procyclic strain T. b. brucei 427 (Cross and Manning,1973) and the derivative clone Nour 6C (Ruepp et al., 1997) werecultured at 27°C in SDM-79 medium containing 5% heat-inactivatedfetal bovine serum (Brun and Schönenberger, 1979). T. b. brucei 427has the full complement of procyclin genes (six copies of EP and twocopies of GPEET). The deletion mutant Nour 6C has no EP genes andonly one copy of GPEET (Ruepp et al., 1997). Neo 2A (Neo) cellsare G418-resistant trypanosomes in which a neomycin-resistancegene has replaced the two EP genes in the Pro A locus (Ruepp et al.,1997). Bloodstream forms of the same strain (MITat 1.2; 221) werecultured at 37°C and 5% CO2 in HMI-9 medium containing 10% heat-inactivated fetal bovine serum (Carruthers and Cross, 1992).
Differentiation of trypanosomes from bloodstream formsto procyclic formsBloodstream form trypanosomes were grown to a density of 5×105
cells/ml as described above. Subsequently, the cells were pelletedby brief centrifugation, the culture supernatant was discarded, andthe cells were resuspended in DTM medium supplemented with 10mM glycerol, 5.5 mM L-proline and 15% fetal bovine serum(Overath et al., 1986) at a density of 1-2×106 cells/ml.Differentiation was induced by adding citrate and cis-aconitate (eachat a final concentration of 3 mM; added from 0.3 M stock solutionsin 25 mM Hepes, pH 7.3) and transferring the cells to 27°C (Overathet al., 1983). Procyclic forms derived from a differentiatedpopulation were cultured in DTM medium at densities between 1-2×106 cells/ml and 1×107 cells/ml.
Transfection of procyclic trypanosomes with humanplacental alkaline phosphatase (PLAP)Construction of recombinant plasmidsThe expression vector pGAPRONE-∆164 containing the neomycinresistance gene as a selectable marker (Furger et al., 1997) wasdigested with BamHI and HindIII to release the coding region ofGARP (glutamic acid/alanine-rich surface protein of Trypanosomacongolense). Subsequently, the multiple cloning site of pBluescriptbetween BamHI and HindIII was ligated into pGAPRONE-∆164 andthe EcoRI PLAP insert (GenBank sequence m12551) from theeukaryotic expression vector pCMVβ (a kind gift from N. Fasel,University of Lausanne) was subcloned into the EcoRI site.
Transfection of cellsT. b. brucei 427 procyclic forms were transfected by electroporationusing 10 µg plasmid DNA digested with SpeI and NotI to release theinsert. Transfected trypanosomes were grown in SDM-79 containing10% fetal bovine serum for 18 hours and resistant cells weresubsequently selected with 25 µg/ml of G418 (Gibco BRL, Basel,Switzerland). Clones were obtained by limiting dilution of G418-resistant parasites in SDM-79 medium supplemented with 20% fetalbovine serum.
Expression of GPEET-Fc fusion proteinConstruction of recombinant plasmidsA polymerase chain reaction (PCR) with the plasmid pBluescript-GPEET (Ruepp et al., 1997) and the primers ABC-H (TA AAG CTTATG GCA CCT CGT T; the synthetic restriction sites used in cloningare underlined) and Pro MscI (TTT CAG CGT TGC ATG GCC AGGTT) was performed to produce a truncated form of GPEET that lacked
the GPI anchor attachment site. A second PCR reaction with theplasmid pJFE14/B61-Fc (generously provided by S. Davis,Regeneron Pharmaceuticals Inc., Tarrytown, NY) using the primersFc SrfI (ATC GGG GGC CCG GGC GAG CC) and Fc BamHI (GCGGA TCC TCA TTT ACC CGG AG) was performed to amplify theFc part of a human IgG1. The PCR product from pBluescript-GPEETwas digested with MscI and HindIII, the PCR product frompJFE14/B61-Fc was digested with SrfI and BamHI and both wereligated into HindIII- and BamHI-digested pEP-∆164-Pur (Ruepp etal., 1997) giving rise to the plasmid pGPEET-Fc-∆164-Pur.
Transfection of cellsT. b. brucei 427 procyclic forms were transfected by electroporationusing 5 µg plasmid DNA digested with the appropriate enzymes torelease the insert. Selection was performed with phleomycin,hygromycin B, nourseothricin and puromycin as described previously(Ruepp et al., 1997) in order to target integration to a specific locus(ProA). The isolation of clones by limiting dilution was performed asdescribed in the previous section.
Extraction of trypanosomes and isolation of GPEET byoctyl-Sepharose chromatographyProcyclic trypanosomes were washed and sequentially extracted withchloroform:methanol (2:1, by vol.), chloroform:methanol:water(10:10:3; by vol.), 9% (v/v) butan-1-ol in water (butanol extract) and0.1% (w/v) Triton X-100 in 20 mM Tris-HCl, pH 7.4 (Triton extract)(Bütikofer et al., 1997). Aliquots of the extracts were stored at −20°C.GPEET procyclin was purified from the butanol extract bychromatography on octyl-Sepharose (Bütikofer et al., 1997).
Phosphorylation of GPEET with 32P in vitroWashed trypanosomes were lysed by incubation in 5 ml buffer (20mM Hepes, pH 7.4, 0.1 mM EDTA, 1 mM DTT) for 15 minutes onice and pressing the suspension three times through a 27G needle.After centrifugation for 30 minutes at 10,000 rpm in a Sorvall SS34rotor, the crude membrane pellet was resuspended in 1 ml of high-saltbuffer (50 mM Hepes, pH 7.4, 0.5 mM EDTA, 5 mM MgCl2, 2 mMDTT, 2 M KCl) and washed once with the same buffer. Membraneswere finally washed twice in phosphorylation buffer (50 mM Hepes,pH 7.2, 50 mM KCl, 0.5 mM EDTA, 5 mM MgCl2, 2 mM DTT). [γ-32P]ATP was added to 200 µl of membranes (equaling 2×107 cells)and incubated for 5 minutes at 30°C. The phosphorylation reactionwas stopped by adding a mixture of chloroform:methanol (1:1; byvol.) to yield a final concentration of chloroform:methanol:water of10:10:3 (by vol.). Phosphorylated GPEET was isolated by extractionand octyl-Sepharose chromatography as described above.
Treatment with alkaline phosphataseTrypanosomes or cell extracts were incubated in the presence orabsence of 10 µg alkaline phosphatase from calf intestine (140 U/mg)in 50 µl of 50 mM diethanolamine, pH 9.8, containing 1 mM MgCl2,for 16-18 hours at 37°C.
Analysis of PLAP activity in trypanosomesProcyclic forms (1×106 cells) were washed with 10 mM Tris, pH 7.4,containing 140 mM NaCl, resuspended in 50 µl of the same buffer,and added to 150 µl assay solution containing 1.33 M diethanolamine,pH 9.8, 0.67 mM MgCl2, and 13.3 mM p-nitrophenylphosphate.Conversion of p-nitrophenylphosphate to p-nitrophenol was measuredspectrophotometrically at 405 nm in microtiter plates over a period of5 minutes at room temperature.
SDS-PAGE and immunoblottingSDS-PAGE was performed under reducing conditions (Laemmli,1970) on 12% polyacrylamide gels. For autoradiography, gels werefixed, soaked in Amplify (Amersham), dried, and exposed to KodakX-Omat S films (INTEGRA Biosciences AG, Wallisellen,

1788
Switzerland) at −70°C. Proteins were transferred from polyacrylamidegels onto polyvinylidene difluoride membranes (Millipore Corp.,Bedford, MA) by electroblotting. The membranes were blocked for 1hours with 5% (w/v) milk powder in blotting buffer (10 mM Tris, pH7.4, containing 140 mM NaCl), incubated with the first antibodydiluted in blocking solution followed by the horseradish peroxidase-conjugated second antibody diluted in blocking solution. The washingsteps in between were carried out in blotting buffer containing 0.05%(v/v) Tween-20. All incubations were performed at room temperature.Antigen binding was visualized with an enhanced chemiluminescencedetection kit (Pierce Chemical Co., Rockford, IL), according to theinstructions of the manufacturer.
Immunofluorescence microscopy and flow cytometry ofdifferentiating cellsLabelling of permeabilized cellsDifferentiating trypanosomes were fixed with acetone for 10 minutes(Tolson et al., 1989). EP and GPEET procyclins were detected usingthe anti-EP mAb 247 (Richardson et al., 1986, 1988) at a dilution of1:200 or rabbit anti-GPEET polyclonal antiserum (K1; Ruepp et al.,1997) at a dilution of 1:400. DNA was stained with the intercalatingdye 4:6-diamidino-2-phenylindole.
Surface labelling of cellsDuring the differentiation of bloodstream forms to procyclic forms,aliquots of the cell suspension were fixed in 2% formaldehyde and0.05% glutaraldehyde at 4°C (Roditi et al., 1989). The different formsof procyclin on the surface of differentiating trypanosomes werelabelled with mAb 247 (1:500), mAb 5H3 (1:1000) or K1 antiserum(1:500) followed by FITC-conjugated second antibodies used atdilutions of 1:400. Fluorescent staining on the trypanosome surfacewas quantified with a FACSCalibur flow cytometer (BectonDickinson, Mountain View, CA) as described elsewhere (Vassella andBoshart, 1996).
LR-white embedding and immunogold labeling oftrypanosomesTrypanosomes were washed by three rounds of centrifugation andresuspension in PBS (8 mM Na2HPO4, 1.5 mM KH2PO4, pH 7.2, 137mM NaCl, 2.6 mM KCl), and cells were fixed in freshly preparedPBS/3% paraformaldehyde/0.05% glutaraldehyde for 30 minutes atroom temperature. Trypanosomes were then washed three times inPBS and incubated in PBS containing 50 mM glycine for 1 hours at4°C. After extensive washing in PBS by several rounds ofcentrifugation, specimens were dehydrated using a graded series ofethanol (50-70-90-100%; 5 minutes each) at −20°C, and embedded inLR-White resin at −15°C, with four changes of fresh resin over aperiod of three days. The resin was polymerized at 58°C for 24 hours.Sections were cut using a Reichert & Jung ultramicrotome, and werepicked up onto 200 mesh Formvar-carbon-coated nickel grids(PLANO GmbH, Marburg, Germany). Loaded grids were stored at4°C for a maximum of 48 hours.
Prior to antibody labeling of the sections, electron microscopy(EM) grids were incubated in EM-blocking buffer (PBS containing50 mM glycine and 0.5% (w/v) bovine serum albumin) overnight at4°C. All subsequent steps were performed at room temperature.Sections were rinsed in PBS and incubated for 1 hours with either themAb 5H3 or the K1 antiserum, both diluted 1:400 in EM blockingbuffer. After washing with five changes of PBS, for 2 minutes each,the respective goat anti-mouse and goat anti-rabbit antibodiesconjugated to 10 nm gold particles (Amersham) were applied at adilution of 1:5 in PBS containing 0.5% bovine serum albumin for 45minutes. After six washes with PBS, for 5 minutes each, thespecimens were briefly rinsed with distilled water and air-dried.Finally, grids were stained with lead citrate and uranyl acetate(Hemphill and Croft, 1997) and viewed on a Phillips 300 transmissionelectron microscope operating at 60 kV.
RESULTS
Phosphorylation of GPEET in vitroThe major form of GPEET can be extracted from delipidatedT. brucei 427 cell pellets by 9% (v/v) butan-1-ol and migrateson SDS-PAGE with an apparent molecular mass of 22-32 kDa(Bütikofer et al., 1997). Experiments showing that the anti-GPEET antiserum K1 reacted weakly with the major form ofGPEET unless it was treated with alkaline phosphatase,suggested that native GPEET might be phosphorylated on oneor several threonine residues in the GPEET repeat units(Bütikofer et al., 1997). To study if GPEET could also bephosphorylated in vitro, trypanosome membrane extracts wereincubated with [γ-32P]ATP. This resulted in the incorporationof radioactivity into a large number of proteins as revealed bySDS-PAGE and autoradiography (results not shown). Thelabeled material was subsequently delipidated using organicsolvents and GPEET was extracted into 9% (v/v) butan-1-oland purified by octyl-Sepharose chromatography. Thisprocedure has been applied before to obtain essentially purepreparations of GPEET procyclin (Bütikofer et al., 1997;Treumann et al., 1997) or EP procyclins (Ferguson et al.,1993). Analysis of purified GPEET by SDS-PAGE andautoradiography showed that the label was efficientlyincorporated into a band with an apparent molecular mass of22-32 kDa (Fig. 1A, top panel). The radioactivity could beremoved nearly quantitatively by treatment of purified GPEETwith alkaline phosphatase (Fig. 1A, top panel). When the samesamples were analyzed by SDS-PAGE and immuno-blotting,the K1 antiserum showed strong reactivity with alkalinephosphatase-treated GPEET whereas the untreated form waspoorly recognized (Fig. 1A, middle panel). In contrast, mAb5H3, which was raised against native GPEET, reacted verystrongly with the phosphorylated form of GPEET butonly weakly with alkaline phosphatase-treated, i.e.dephosphorylated, GPEET (Fig. 1A, bottom panel). Theseresults demonstrate that GPEET can be phosphorylated in vitrousing a procyclic T. brucei 427 membrane preparation andthat the two antibodies preferentially react with theunphosphorylated (K1 antiserum) or phosphorylated (mAb5H3) form of GPEET.
P. Bütikofer and others
Fig. 1. Phosphorylation of GPEET in vitro. Trypanosomemembranes were incubated in the presence of [γ-32P]ATP, washed,and GPEET was isolated by sequential extraction andchromatography on octyl-Sepharose. Radiolabelled GPEET from thebutanol extract was incubated in the absence (-) or presence (+) ofalkaline phosphatase and analyzed by SDS-PAGE followed byautoradiography (top panel) or immuno-blotting using the K1antiserum (middle panel) or mAb 5H3 (bottom panel).

1789Phosphorylation and transport of GPEET
We have previously reported that the K1 antiserum alsoreacts on immuno-blots with a minor form of GPEET (possiblya doublet) with an apparent molecular mass of ~20 kDa (Rueppet al., 1997; Bütikofer et al., 1997) which is present in wholecell lysates, but resists extraction by 9% butan-1-ol. This formof GPEET reacted strongly with the K1 antiserum withoutpretreatment with alkaline phosphatase (Fig. 2A, lane a), butwas not recognized by mAb 5H3 (Fig. 2B, lane a). Aftertreatment of the cells with alkaline phosphatase, both the 20and 22-32 kDa bands were strongly labeled by the K1antiserum and an additional band at ~18 kDa appeared (Fig.2A, lane b) which may represent a proteolytic degradationproduct of the 20 kDa band. This band was also not recognizedby mAb 5H3 (Fig. 2B, lane b). The nature of these minor formsof GPEET, including their phosphorylation status, is currentlyunder further investigation.
Expression of GPEET during the differentiation ofbloodstream form trypanosomes into procyclicformsThe replacement of the VSG surface coat by EP procyclinduring the differentiation of T. brucei bloodstream forms intoprocyclic forms is well-established (Roditi et al., 1989;Ziegelbauer et al., 1990, 1993; Vassella and Boshart, 1996;Matthews and Gull, 1997). The recent identification of GPEETin procyclic forms (Ruepp et al., 1997; Treumann et al., 1997;Bütikofer et al., 1997; this report) prompted us to study itsexpression during the differentiation of bloodstream forms of
T. brucei MITat 1.2 to procyclic forms. Differentiation wasinduced by the addition of citrate/cis-aconitate to the culturemedium together with a drop in culture temperature from 37°Cto 27°C. Trypanosomes were collected after given timepointsand analyzed by SDS-PAGE and immuno-blotting usingantibodies against VSG, EP and GPEET. We found that VSGwas almost completely lost from trypanosomes 24 hours afterinitiating differentiation (Fig. 3, top panel). In contrast, thesynthesis of EP could be detected by 4 hours and reachedmaximal levels 24-48 hours after triggering differentiation(Fig. 3, second panel). The time course of release of VSG andexpression of EP in our study is comparable to that reportedbefore for another member of the MITaR 1 lineage, MITat 1.4(Overath et al., 1983; Roditi et al., 1989).
In common with EP, the expression of GPEET is an earlyevent in the differentiation process. The K1 antiserum detecteda band of ~20 kDa apparent molecular mass within 4 hoursafter initiating differentiation (Fig. 3, third panel). This bandrepresents the minor unphosphorylated form of GPEET (Fig.2A; see also Ruepp et al., 1997; Bütikofer et al., 1997).Twenty-four hours after triggering differentiation, the K1antiserum also reacted with the 22-32 kDa major form ofGPEET (Fig. 3, third panel). In contrast, mAb 5H3 showed noreactivity with any protein in differentiating cells for the first6 hours after initiation of differentiation (Fig. 3, bottom panel).Only after 24-48 hours did mAb 5H3 react with the 22-32 kDaband.
Immunofluorescence microscopy of acetone-fixed (i.e.permeabilized) cells showed that 6 hours after initiation ofdifferentiation, 10-20% of the cells bound the anti-EP mAb 247and anti-GPEET antiserum (K1); after 17 hours, 60-70% of thecells were EP- and GPEET-positive. After 48 hours essentiallyall cells were labeled with mAb 247 and K1 antiserum, as wellas with mAb 5H3, although we observed occasional cellsexpressing either EP or GPEET but not both (results notshown).
The expression of EP and GPEET during the differentiationof T. brucei bloodstream forms to procyclic forms was furtherstudied by flow cytometry of surface-labeled cells. The resultsshowed that neither EP or GPEET were expressed on the cellsurface for the first 12 hours after initiating differentiation (Fig.4). In contrast, after 24-48 hours both the anti-EP antibody andthe two anti-GPEET antibodies bound to similar numbers ofcells, indicating that the cell surface expression of EP andGPEET occurred simultaneously. After 5 and 26 days in
Fig. 2. Detection of multiple forms of GPEET in trypanosomemembranes. T. brucei 427 procyclic trypanosomes were washed,incubated in the absence (lanes a) or presence (lanes b) of alkalinephosphatase and analyzed by SDS-PAGE followed by immuno-blotting using the K1 antiserum (A) or mAb 5H3 (B).
Fig. 3. Expression of GPEET during the differentiation of T.brucei bloodstream forms to procyclic forms. Bloodstreamform trypanosomes were resuspended in DTM medium anddifferentiation into procyclic forms was induced by addingcitrate and cis-aconitate and incubating the cells at 27°C for72 hours. Control cells were incubated at 27°C for 72 hours inthe absence of citrate and cis-aconitate. At given time points,aliquots of the cell suspensions were removed and the cellswere washed twice in isotonic buffer. Equal amounts of cellswere analyzed by SDS-PAGE and immuno-blotting using theanti-VSG, anti-EP (mAb 247), K1 antiserum and mAb 5H3 asindicated. The lanes are labeled with the time (in hours) afterinitiating differentiation; lanes c0 and c72 contain the controlcells after 0 hours and 72 hours, respectively, of incubation.

1790
continuous culture, the maximal intensity of the labelingslightly decreased for the K1 antiserum and mAb 5H3 whilethat for the anti-EP mAb 247 remained constant (Fig. 4).
Electron microscopy of GPEET in procyclic formtrypanosomesThe distribution of the phosphorylated and unphosphorylatedforms of GPEET was investigated using immunogold labelingof sections of LR-White-embedded trypanosomes. On sectionscontaining T. brucei 427 wild-type cells, labeling with the K1antiserum was restricted mainly to the flagellar pocket, areassurrounding the pocket, and the flagellum attachment zone(Fig. 5A-C). Only a few gold particles could be found on otherparts of the cell. In contrast, immunogold labeling with mAb5H3 resulted in extensive staining all over the trypanosomesurface (Fig. 5D). In addition, the flagellar pocket andvesiculated structures surrounding the flagellar pocket werealso labeled. Taken together, these results suggest that bothforms of GPEET are present in the region of the flagellarpocket, but that the majority of surface GPEET isphosphorylated.
Expression of human PLAP in procyclictrypanosomesIn order to study if the expression of a membrane-boundphosphatase would affect GPEET phosphorylation andtransport, procyclic forms of T. brucei 427 were transfectedwith a cDNA encoding a GPI-anchored form of humanplacental alkaline phosphatase (PLAP). Three clones,designated PLAP 1.2, 1.5 and 2.2, were selected on the basisof antibiotic resistance; all of them expressed alkaline
phosphatase activity. Integration of the PLAP DNA wasanalyzed by Southern blotting and occurred into the Pro A(PLAP 1.2), Pro C (PLAP 1.5) and Pro B (PLAP 2.2) loci.Transformants which expressed PLAP (PLAP+) had the samegeneration time as wild-type trypanosomes and could be keptin culture for several months without any noticeablemorphological changes compared to the parental line (resultsnot shown). Interestingly, however, we noticed that the amountof PLAP expressed in the three clones progressively decreasedduring prolonged culture times. Concomittant with thedecrease in PLAP activity, the cells expressed lessdephosphorylated and more phosphorylated GPEET (resultsnot shown).
Membrane association of PLAP was studied by phasepartitioning in Triton X-114 and chromatography on octyl-Sepharose. Enzyme activity was monitored as the protein couldnot be detected using antibodies that were commerciallyavailable. More than 95% of PLAP activity was recovered inthe detergent-rich phase after phase partitioning of PLAP+ cellsin Triton X-114. In addition, after solubilization of PLAP+ cellswith 0.1% Triton X-100 and chromatography on octyl-Sepharose, alkaline phosphatase activity quantitatively boundto the column and co-eluted together with GPI-anchored EPand GPEET upon elution of the column with an increasing 1-propanol gradient (results not shown). These data areconsistent with the attachment of PLAP to the membrane viaa GPI moiety. Direct identification of the PLAP GPI anchorusing radiolabeled GPI precurors was not successful and maybe due to the low amounts of PLAP expressed in trypanosomesrelative to EP and GPEET. The low level of expression of GPI-anchored PLAP may result from a poor recognition of themammalian GPI anchor attachment signal in trypanosomes. Asimilar observation has been made for the converse case, i.e.the expression of VSG in mammalian cells (Moran and Caras,1994).
GPEET phosphorylation in PLAP+ procyclictrypanosomesThe effects of PLAP expression on the phosphorylation ofGPEET were studied by SDS-PAGE and immuno-blottingusing the K1 antiserum and mAb 5H3. Neo 2A is a derivativeof T. brucei 427 in which 2 procyclin genes have been replacedby the neomycin resistance gene (Ruepp et al. 1997). Incontrast to the results obtained with wild-type cells (Fig. 2A)and the Neo 2A controls, the K1 antiserum reacted stronglywith the 22-32 kDa form of GPEET in all three PLAP+ celllines (Fig. 6A). Once again, the K1 antiserum also bound tothe 20 kDa minor form of GPEET and, in addition, recognizedthe form at 18 kDa in PLAP 1.2 cells (Fig. 6A). The appearanceof the 18 kDa form was variable and could be detected in theother PLAP+ clones on some blots (results not shown).Conversely, mAb 5H3 only recognized the 22-32 kDa form ofGPEET in Neo 2A controls but not in any of the PLAP+ cells(Fig. 6B).
The cellular distribution of phosphorylated andunphosphorylated GPEET in PLAP-expressing trypanosomeswas subsequently studied by electron microscopy.Unphosphorylated GPEET (detected by the K1 antiserum) wasnow distributed all over the surface of the cell (Fig. 7A). Incommon with wild-type cells, the K1 antiserum also stronglylabelled the region of the flagellar pocket. In contrast, however,
P. Bütikofer and others
Fig. 4. Flow cytometry analysis of surface-labeled bloodstreamforms during differentiation to procyclic forms. At given time pointsafter triggering differentiation, trypanosomes were fixed, labeledwith antibodies against EP (mAb 247), phosphorylated (GPEET-P;mAb 5H3) or unphosphorylated GPEET (GPEET; K1 antiserum) andanalyzed by flow cytometry. The histograms of fluorescence intensitywere scaled to similar peak heights for better comparison.
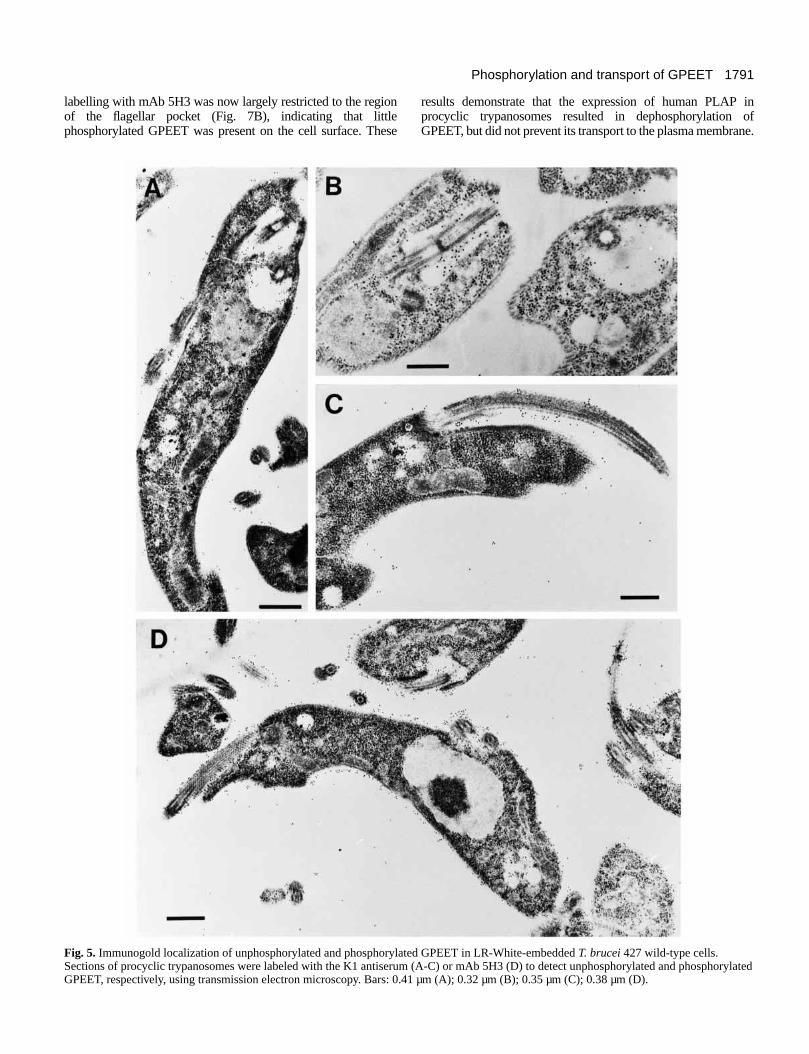
1791Phosphorylation and transport of GPEET
labelling with mAb 5H3 was now largely restricted to the regionof the flagellar pocket (Fig. 7B), indicating that littlephosphorylated GPEET was present on the cell surface. These
results demonstrate that the expression of human PLAP inprocyclic trypanosomes resulted in dephosphorylation ofGPEET, but did not prevent its transport to the plasma membrane.
Fig. 5. Immunogold localization of unphosphorylated and phosphorylated GPEET in LR-White-embedded T. brucei 427 wild-type cells.Sections of procyclic trypanosomes were labeled with the K1 antiserum (A-C) or mAb 5H3 (D) to detect unphosphorylated and phosphorylatedGPEET, respectively, using transmission electron microscopy. Bars: 0.41 µm (A); 0.32 µm (B); 0.35 µm (C); 0.38 µm (D).

1792
Expression of a GPEET-Fc fusion protein inprocyclic form trypanosomesTo investigate whether association with the cell surface was aprerequisite for phosphorylation in T. brucei, the coding regionof the extracellular domain of GPEET was fused to the constant(Fc) region of a human immunoglobulin G gene. The resultingfusion protein is secreted by Drosophila embryo cells (S.
Ruepp, unpublished observation), and it was expected thattransgenic trypanosomes would also secrete GPEET-Fc as itcontains a signal sequence, but no hydrophobic transmembranedomain or signal for GPI anchor addition. In trypanosomes,however, the protein was retained in subcellular compartments,most likely the ER, since it was found to colocalise with BiPusing immunofluorescence microscopy (data not shown). Asimilar observation has recently been made for a truncated formof VSG which cannot receive a GPI anchor (Bangs et al., 1997;McDowell et al., 1998). Immuno-blot analysis was performedusing lysates of procyclic form trypanosomes which expressedboth GPEET-Fc and the endogenous GPI-anchored form ofGPEET (Fig. 8). The K1 antiserum recognized both theunphosphorylated and phosphorylated forms of GPEET as wellas the fusion protein (Fig. 8, lane a). In contrast, mAb 5H3 onlyrecognized the endogenous, phosphorylated form of GPEETand not GPEET-Fc (Fig. 8, lane b). The fact that GPEET-Fc isneither phosphorylated nor secreted is consistent with a kinaseacting on endogenous GPEET at a later stage during transport.
DISCUSSION
The observations that GPEET procyclin contained severalpotential phosphorylation sites for casein kinase II and thatalkaline phosphatase treatment considerably improved the
P. Bütikofer and others
Fig. 6. Effect of expression of PLAP on GPEET phosphorylation inprocyclic trypanosomes. Neo 2A (Neo) cells and PLAP+ cells werewashed twice in buffer containing 10 mM EDTA and analyzed bySDS-PAGE and immuno-blotting using the K1 antiserum (A) ormAb 5H3 (B).
Fig. 7. Immunogold localization of unphosphorylated and phosphorylated GPEET in LR-White-embedded PLAP+ procyclic trypanosomes.Sections of parasites were labeled with the K1 antiserum (A) or mAb 5H3 (B) to detect unphosphorylated and phosphorylated GPEET,respectively, using transmission electron microscopy. Bars: 0.80 µm (A); 0.50 µm (B).

1793Phosphorylation and transport of GPEET
reactivity of the protein with anti-peptide antibodies suggestedthat the polypeptide backbone of native GPEET might bephosphorylated (Bütikofer et al., 1997). This has recently beenconfirmed spectrometrically by Mehlert et al. (1999) who, inaddition, demonstrated that the majority of mature (GPI-anchored) GPEET is phosphorylated on six out of seventhreonine residues. Two antibodies can discriminate betweenthe phosphorylated and unphosphorylated forms of GPEET.Antiserum K1, which was raised against a synthetic peptide(Ruepp et al., 1997), preferentially binds unphosphorylatedminor forms of GPEET (molecular mass ≤20 kDa), but is alsocapable of binding weakly to the major, phosphorylated forms(22-32 kDa; Bütikofer et al., 1997). In contrast, mAb 5H3,which was isolated on the basis of its binding to native GPEET,exclusively binds to phosphorylated forms. Treatment ofpurified GPEET with alkaline phosphatase abolished reactivitywith mAb 5H3, confirming that the epitope recognized by thisantibody contains one or more phosphate groups.
The expression of GPEET and EP procyclins was monitoredsimultaneously in trypanosomes which had been triggered todifferentiate into procyclic forms. Antibodies directed againstthe EP and GPEET internal repeats (mAb 247 and K1antiserum) detected both proteins on immuno-blots or inpermeablised cells within 2-4 hours. The two forms ofprocyclin appeared to be tightly coregulated since, with veryfew exceptions, individual cells were either positive for bothEP and GPEET or negative for both. Despite the fact thatGPEET could be detected very early during differentiation, thedifferent forms of the protein were not expressed with the samekinetics. For the first 6-12 hours, the 20 kDa form of GPEETwas the only form detected in cell lysates. In contrast, thephosphorylated 22-32 kDa forms of GPEET were first detected24 hours after triggering differentiation, which coincided withthe appearance of both EP and GPEET on the surface oftrypanosomes. It still needs to be established what additionalmodifications distinguish the high and low molecular massforms of GPEET. The change in electrophoretic mobilitycannot be due to the phosphate groups alone, since treatmentwith alkaline phosphatase did not convert the larger forms tothe 20 kDa form. Likewise, the expression of human PLAP intrypanosomes, although greatly enhancing the binding of anti-peptide antibodies, had no effect on the migration of GPEET.If one threonine in GPEET is not phosphorylated (Mehlert et
al., 1999), it could theoretically be a site of O-linkedglycosylation. If this is so, however, it cannot be a conventionalhydrogen fluoride-resistant modification as this would havebeen detected in the analysis performed by Mehlert et al.(1999). Another possibility to be considered is that the alteredmobility of the different forms of GPEET reflectsmodifications to the GPI anchor. EP procyclin has the sametype of GPI anchor as GPEET (Treumann et al., 1997), butdoes not exhibit any changes in molecular mass during thecourse of differentiation. It is possible, however, that a slowerrate of maturation of GPEET compared to EP would allow thedetection of postranslational GPI modifications in GPEET thatare not seen in EP.
The distribution of the phosphorylated and unphosphorylatedforms of GPEET within established procyclic formtrypanosomes was determined by transmission electronmicroscopy. In wild-type cells, K1 antiserum detectedunphosphorylated GPEET in vesicles close to the flagellarpocket, in the flagellar pocket itself and at the base of theflagellum, but showed negligeable labelling of the surface.Phosphorylated forms, as detected by mAb 5H3, were alsopresent within the flagellar pocket and, in addition, were spreadover the entire parasite surface. Both forms of GPEET were alsodetected in the flagellar pocket of PLAP+ transformants, but incontrast to the parental cell line, these cells were coated withunphosphorylated GPEET. These results are consistent with amodel in which a kinase present in the flagellar pocket and/orclose to the base of the flagellum converts the unphosphorylatedGPEET to the phosphorylated form. Our finding that GPEET-Fc, which accumulates intracellularly, is not phosphorylated,provides further evidence that phosphorylation does not occurin the ER, but later during the secretion pathway. At present itis not clear whether GPEET must be phosphorylated in orderto be exported, or if this is simply a consequence of its passingthrough the flagellar pocket en route to the plasma membrane.PLAP, which was expressed as a membrane-bound protein, andwould therefore be expected to follow the same secretionpathway as procyclins, might already act on some GPEET inthe flagellar pocket, but probably dephosphorylates the bulk ofit only once it reaches the surface.
Very few ecto-kinases have been identified to date; thosewhich have been localised are either attached to the plasmamembrane (Kübler et al., 1982; Sonka et al., 1989; Lester etal., 1990; Walter et al., 1996; Sacerdoti-Sierra and Jaffe, 1997)or secreted (Sacerdoti-Sierra and Jaffe, 1997). Phosphorylationof GPEET by a putative GPEET kinase clearly occurs duringsecretory transport, possibly in a specific sub-cellular structuresuch as the flagellar pocket, and may additionally occur on thecell surface. We found a GPEET kinase activity in crudepreparations of trypanosome membranes. Furtherfractionation, and the use of appropriate markers for theflagellar pocket, the plasma membrane and other sub-cellularstructures, should allow us to determine its precise localization.It was previously proposed (Mehlert et al., 1999) that GPEET,prior to its transloaction into the ER membrane, might protrudeinto the cytoplasm and be phosphorylated by a casein kinaseII-like enzyme, but we feel that this is unlikely for two reasons.First, there was no apparent labelling of internal structureswhen thin sections were incubated with mAb 5H3. Second, theGPEET-Fc fusion protein, which is retained intracellularly, wasnot detectably phosphorylated.
Fig. 8. Expression of GPEET-Fc in procyclic trypanosomes. T.brucei 427 procyclic forms expressing the GPEET-Fc fusion proteinwere analyzed by SDS-PAGE followed by immuno-blotting usingthe K1 antiserum (lane a) or mAb 5H3 (lane b).

1794
What might be the significance of GPEET phosphorylation?Procyclins are already highly negatively charged by virtue oftheir glutamic acid content and the sialic residues which capthe lactosamine repeats in the GPI anchor. The phosphorylationof Thr residues adds additional negative charges to themolecule. It is interesting to note that promastigote andmetacyclic forms of Leishmania sp. (related trypanosomatidswhich are also transmitted by insects) express GPI-anchoredlipophosphoglycans (reviewed by Turco and Descoteaux,1992). By analogy to procyclins, these are highly negatively-charged, extended molecules, but their backbones consist ofphosphorylated polysaccharide rather than polypeptide repeats.Both types of surface coat would provide these parasites witha charged shield that might protect them against the hydrolyticenvironment in the insects’ intestinal tracts. It is alsoconceivable that the negatively charged surface moleculesparticipate in specific parasite-vector interactions, for exampleby serving as receptors that bind insect factors important forparasite differentiation or, alternatively, for targeting theparasites to the correct host tissue. That EP and GPEETprocyclins can be expressed at different levels in differentstrains of parasites (Treumann et al., 1997; Bütikofer et al.,1997), in different culture conditions and after varyingintervals of culture (Roditi et al., 1998), lends support to theidea that the procyclins, at least, do not have a solely passiverole.
We are grateful to Mike Ferguson (University of Dundee) and JayBangs (University of Wisconsin-Madison) for communicating databefore publication. Jay Bangs is also thanked for providing the anti-BiP antibody. This work was supported in part by Swiss NationalScience Foundation grants 31-050587.97 (to P.B.), 31-050932.97 (toI.R.), 31-46760.96 (to T.S.) and 31-46846.96 (to A.H.), by a grantfrom the Natural Science and Engineering Research Council ofCanada (to T.W.P.) and an award from the Helmut Horten Foundationto I.R. P.B. thanks S. Jungi for the photographs and U. Brodbeck, P.Hofer, and M.S. Bütikofer for continued support and stimulation.
REFERENCES
Apasov, S. G., Smith, P. T., Jelonek, M. T., Margulies, D. H. and Sitkovsky,M. V. (1996). Phosphorylation of extracellular domains of T-lymphocytesurface proteins. J. Biol. Chem. 271, 25677-25683.
Bangs, J. D., Ransom, D. M., McDowell, M. A. and Brouch, E. M. (1997).Expression of bloodstream variant surface glycoproteins in procyclicstage Trypanosoma brucei: role of GPI anchors. EMBO J. 16, 4285-4294.
Brun, R. and Schönenberger, M. (1979). Cultivation and in vitro cloning ofprocyclic culture forms of Trypanosoma brucei in a semi-defined medium.Acta Trop. 36, 289-292.
Bütikofer, P., Ruepp, S., Boschung, M. and Roditi, I. (1997). ‘GPEET’procyclin is the major surface protein of procyclic culture forms ofTrypanosoma brucei brucei strain 427. Biochem. J. 326, 415-423.
Carruthers, V. B. and Cross, G. A. M. (1992). High-efficiency clonal growthof bloodstream- and insect-form Trypanosoma brucei on agarose plates.Proc. Nat. Acad. Sci. USA 89, 8818-8821.
Cross, G. A. and Manning, J. C. (1973). Cultivation of Trypanosoma bruceisspp. in semi-defined media. Parasitology 67, 315-331.
Delahunty, M. D., Stafford, F. J., Yuan, L. C., Shaz, D. and Bonifacino, J.S. (1993). Uncleaved signals for glycosylphosphatidylinositol anchoringcause retention of precursor proteins in the endoplasmic reticulum. J. Biol.Chem. 268, 12017-12027.
Doering, T. L. and Schekman, R. (1996). GPI anchor attachment is requiredfor Gas1p transport from the endoplasmic reticulum in COP II vesicles.EMBO J. 15, 182-191.
Ferguson, M. A. J., Murray, P., Rutherford, H. and McConville M. J.(1993). A simple purification of procyclic acidic repetitive protein anddemonstration of a sialylated glycosyl-phosphatidylinositol membraneanchor. Biochem. J. 291, 51-55.
Field, M. C., Menon, A. K. and Cross, G. A. M. (1991). Aglycosylphosphatidylinositol protein anchor from procyclic stageTrypanosoma brucei: lipid structure and biosynthesis. EMBO J. 10, 2731-2739.
Field, M. C., Moran, P., Li, W., Keller, G. A. and Caras, I. W. (1994).Retention and degradation of proteins containing an uncleaved glycosylphosphatidylinositol signal. J. Biol. Chem. 269, 10830-10837.
Furger, A., Schürch, N., Kurath, U. and Roditi, I. (1997). Elements in the3′ untranslated region of procyclin mRNA regulate expression in insectforms of Trypanosoma brucei by modulating RNA stability and translation.Mol. Cell. Biol. 17, 4372-4380.
Hemphill, A. and Croft, S. L. (1997). Electronmicroscopy in parasitology. InAnalytical Parasitology (ed. M. Rogan and P. Craig), pp. 227-268. Berlin:Springer Verlag.
Hermoso, T., Pérez, J. L., Cornivelli, L. and Hernandez, A. G. (1994).Extracellular phosphorylation in Leishmania major and Leishmaniamexicana during heat shock transformation. Acta Trop. 56, 1-6.
König, E., Delius, H., Carrington, M., Williams, R. O. and Roditi I. (1989).Duplication and transcription of procyclin genes in Trypanosoma brucei.Nucl. Acids Res. 17, 8727-8739.
Kübler, D., Pyerin, W. and Kinzel, V. (1982). Protein kinase activity andsubstrates at the cell surface of intact HeLa cells. J. Biol. Chem. 257, 322-329.
Kübler, D., Pyerin, W., Burow, E. and Kinzel, V. (1983). Substrate-effectedrelease of surface-located protein kinase from intact cells. Proc. Nat. Acad.Sci. USA 80, 4021-4025.
Laemmli, U. K. (1970). Cleavage of structural proteins during the assemblyof the head of bacteriophage T4. Nature 227, 680-685.
Lester, D. L., Hermoso, T. and Jaffe, C. L. (1990). Extracellularphosphorylation in the parasite, Leishmania major. Biochim. Biophys. Acta1052, 293-298.
Matthews, K. R. and Gull, K. (1997). Commitment to differentiation and cellcycle re-entry are coincident but separable events in the transformation ofAfrican trypanosomes from their bloodstream to their insect form. J. CellSci. 110, 2609-2618.
McConville, M. J. and Ferguson, M. A. J. (1993). The structure, biosynthesisand function of glycosylated phosphatidylinositols in the parasitic protozoaand higher eukaryotes. Biochem. J. 294, 305-324.
McDowell, M. A., Ransom, D. M. and Bangs, J. D. (1998).Glycosylphosphatidylinositol-dependent secretory transport inTrypanosoma brucei. Biochem. J. 335, 681-689.
Mehlert, A., Treumann, A. and Ferguson, M. A. J. (1999). Trypanosomabrucei GPEET-PARP is phosphorylated on six out of seven threonineresidues. Mol. Biochem. Parasitol. 98, 291-296.
Merlo, D., Anelli, R., Calissano, P., Ciotti, M. T. and Volonté, C. (1997).Characterization of an ecto-phosphorylated protein of cultured cerebellargranule neurons. J. Neurosci. Res. 47, 500-508.
Moran, P. and Caras, I. W. (1992). Proteins containing an uncleaved signalfor glycosylphosphatidylinositol membrane anchor attachment are retainedin a post-ER compartment. J. Cell Biol. 119, 763-772.
Moran, P. and Caras, I. W. (1994). Requirements forglycosylphosphatidylinositol attachment are similar but not identical inmammalian cells and parasitic protozoa. J. Cell Biol. 125, 333-343.
Mowatt, M. R. and Clayton, C. E. (1987). Developmental regulation of anovel repetitive protein of Trypanosoma brucei. Mol. Cell. Biochem. 7,2838-2844.
Mowatt, M. R. and Clayton, C. E. (1988). Polymorphism in the procyclicacidic repetitive protein gene family of Trypanosoma brucei. Mol. Cell.Biochem. 8, 4055-4062.
Mowatt, M. R., Wisdom, G. S. and Clayton, C. E. (1989). Variation oftandem repeats in the developmentally regulated procyclic acidic repetitiveproteins of Trypanosoma brucei. Mol. Cell. Biochem. 9, 1332-1335.
Overath, P., Czichos, J., Stock, U. and Nonnengaesser, C. (1983).Repression of glycoprotein synthesis and release of surface coat duringtransformation of Trypanosoma brucei. EMBO J. 2, 1721-1728.
Overath, P., Czichos, J. and Haas, C. (1986). The effect of citrate/cis-aconitate on oxidative metabolism during transformation of Trypanosomabrucei. Eur. J. Biochem. 160, 175-182.
Pinna, L. A. (1990). Casein kinase 2: an ‘eminence grise’ in cellularregulation? Biochim. Biophys. Acta 1054, 267-284.
P. Bütikofer and others

1795Phosphorylation and transport of GPEET
Richardson, J. P., Jenni, L., Beecroft, R. P. and Pearson, T. W. (1986).Procyclic tsetse fly midgut forms and culture forms of African trypanosomesshare stage- and species-specific surface antigens identified by monoclonalantibodies. J. Immunol. 136, 2259-2264.
Richardson, J. P., Beecroft, R. P., Tolson, D. L., Liu, M. K. and Pearson,T. W. (1988). Procyclin: an unusual immunodominant glycoprotein surfaceantigen from the procyclic stage of African trypanosomes. Mol. Biochem.Parasitol. 31, 203-216.
Roditi, I., Carrington, M. and Turner, M. (1987). Expression of apolypeptide containing a dipeptide repeat is confined to the insect stage ofTrypanosoma brucei. Nature 325, 272-274.
Roditi, I., Schwarz, H., Pearson, T. W., Beecroft, R. P., Liu, M. K.,Richardson, J. P., Bühring, H.-J., Pleiss, J., Bülow, R., Williams, R. O.and Overath, P. (1989). Procyclin gene expression and loss of the variantsurface glycoprotein during differentiation of Trypanosoma brucei. J. CellBiol. 108, 737-746.
Roditi, I., Furger, A., Ruepp, S., Schürch, N. and Bütikofer, P. (1998).Unravelling the procyclin coat of Trypanosoma brucei. Mol. Biochem.Parasitol. 91, 117-130.
Ruepp, S., Furger, A., Kurath, U., Kunz Renggli, C., Hemphill, A., Brun,R. and Roditi, I. (1997). Survival of Trypanosoma brucei in the tsetse flyis enhanced by the expression of specific forms of procyclin. J. Cell Biol.137, 1369-1379.
Sacerdoti-Sierra, N. and Jaffe, C. L. (1997). Release of ecto-proteinkinases by the protozoan parasite Leishmania major. J. Biol. Chem. 272,39760-30765.
Sonka, J., Kübler, D. and Kinzel, V. (1989). Phosphorylation by cell surfaceprotein kinase of bovine and human fibrinogen and fibrin. Biochim. Biophys.Acta 997, 268-277.
Tolson, D. L., Turco, S. J., Beecroft, R. P. and Pearson, T. W. (1989). Theimmunochemical structure and surface arrangement of Leishmania
donovani lipophosphoglycan determined using monoclonal antibodies. Mol.Biochem. Parasitol. 35, 109-118.
Tomavo, S., Martinage, A. and Dubremetz, J. F. (1992). Phosphorylation ofToxoplasma gondii major surface antigens. Parasitol. Res. 78, 541-544.
Treumann, A., Zitzmann, N., Hülsmeier, A., Prescott, A. R., Almond, A.,Sheehan, J. and Ferguson, M. A. J. (1997). Structural characterisation oftwo forms fo procyclic acidic reprtitive protein expressed by procyclic formsof Trypanosoma brucei. J. Mol. Biol. 269, 529-547.
Turco, S. J. and Descoteaux, A. (1992). The lipophosphoglycan ofLeishmania parasites. Annu. Rev. Microbiol. 46, 65-94.
Udenfriend, S. and Kodukula, K. (1995). How glycosylphosphatidylinositol-anchored membrane proteins are made. Meth. Enzymol. 250, 563-591.
Vassella, E. and Boshart, M. (1996). High molecular mass agarose matrixsupports growth of bloodstream forms of pleomorphic Trypanosoma bruceistrains in axenic culture. Mol. Biochem. Parasitol. 83, 91-105.
Vickermann, K. (1985). Developmental cycles and biology of pathogenictrypanosmes. Br. Med. Bull. 41, 105-114.
Vidugiriene, J. and Menon, A. K. (1995). Biosynthesis ofglycosylphosphatidylinositol anchors. Meth. Enzymol. 250, 513-535.
Volonté, C., Merlo, D., Ciotti, M. T. and Calissano, P. (1994). Identificationof an ectokinase activity in cerebellar granule primary neuronal cultures. J.Neurochem. 63, 2028-2037.
Walter, J., Schnölzer, M., Pyerin, W., Kinzel, V. and Kübler, D. (1996).Induced release of cell surface protein kinase yields CK1- and CK2-likeenzymes in tandem. J. Biol. Chem. 271, 111-119.
Ziegelbauer, K., Quinten, M., Schwarz, H., Pearson, T. W. and Overath,P. (1990). Synchronous differentiation of Trypanosoma brucei frombloodstream to procyclic forms in vitro. Eur. J. Biochem. 192, 373-378.
Ziegelbauer, K., Stahl, B., Karas, M., Stierhof, Y.-D. and Overath, P.(1993). Proteolytic release of cell surface proteins during differentiation ofTrypanosoma brucei. Biochemistry 32, 3737-3742.




















