
A diffusion-reaction model of carbon isotope fractionation in foraminifera
Upload
independentCategory
view
0download
0
Vaccine 21 (2003) 3961–3971
Fractionation, structural studies, and immunological characterization ofthe semi-synthetic Quillaja saponins derivative GPI-0100
Dante J. Marciania,∗, Robert C. Reynoldsb, Ashish K. Pathakb,Kyra Finley-Woodmana, Richard D. Mayb
a Galenica Pharmaceuticals Inc., 2800 Milan Court, Suite 118, Birmingham, AL 35211, USAb Southern Research Institute, Birmingham, AL 35205, USA
Received 16 December 2002; received in revised form 4 April 2003
Abstract
Unfractionated GPI-0100 (UFGPI-0100) containing semi-synthetic derivatives of deacylated Quillaja saponins (DS saponins) modifiedat the glucuronic acid residue was resolved by reverse phase low-pressure liquid chromatography (RP-LPLC) into two fractions, RP18-1and RP18-2, with different compositions and adjuvanticity. The fraction RP18-1 contained DS saponin adducts ofN-dicyclohexylurea,and stimulated Th2 immunity with production of IgG1, while the RP18-2 fraction contained the dodecylamide derivatives of DS saponinsand stimulated Th1 immunity with production of IgG2a, IFN-�, IL-2, and CTL. The strong immune stimulatory properties of RP18-2,relative to RP18-1, and the formation of RP18-1/RP18-2 mixed micelles may account for the effective stimulation of Th1 immunity byUFGPI-0100. UFGPI-0100 was free of acylated quillaja saponin components, including the more stable QS-7.© 2003 Elsevier Science Ltd. All rights reserved.
Keywords: Quillaja saponins; Saponin analogs; Adjuvant; GPI-0100
1. Introduction
The bark of the South American treeQuillaja saponariaMolina contains many related triterpenoid saponins (Q.saponins) which have similar immune stimulating activities,but significant differences in their toxicity. These includeQS-7, QS-17, QS-18 and QS-21[1]. Unlike alum, whichstimulates Th2 immunity unsuitable for most intracellularpathogens[2], Q. saponins stimulate Th1 immunity withthe production of cytotoxic T lymphocytes (CTL) againstexogenous antigens[3,4]. Q. saponins are largely composedof an aldehyde-containing triterpenoid genin linked to twooligosaccharide chains, one of which is acylated by twofatty acid residues in tandem. Despite a considerably shorteracyl chain, QS-7 has immune modulating properties similarto those of other Q. saponins but at higher doses[5]. Struc-ture/function relation studies have shown that while the Q.saponins’ aldehyde group delivers the co-stimulatory signalrequired for Th1 immunity[3], the acyl fatty acid moietyis responsible for CTL production and the toxicity of Q.saponins (with the exception of QS-7)[6]. DS saponins,however, do not stimulate Th1 immunity.
∗ Corresponding author. Tel.:+1-205-969-5829; fax:+1-205-970-0058.E-mail address: [email protected] (D.J. Marciani).
Although Q. saponins are effective adjuvants, the es-ter bond between the acylating fatty acid groups and thefucosyl residue is readily hydrolyzed under mild condi-tions. Removal of the acyl groups yields two major DSsaponins that differ in their oligosaccharide structures[7].This deacylation results in the alteration of the Q. saponins’immune stimulating properties and a loss of their toxicity[6]. This occurs whether all the Q. saponins are combinedor isolated[6,8]. At higher doses DS saponins stimulateantigen-specific antibody production, but fail to protectanimals against challenge by certain infectious agents[9].It has been shown that the removal of the labile acylatingfatty acids from Q. saponins followed by the incorporationof a dodecylamide group at the glucuronic acid residueresults in stable semi-synthetic derivatives (unfractionatedGPI-0100 (UFGPI-0100)) with immune stimulating proper-ties similar to those of native saponins[10]. The negligibletoxicity of UFGPI-0100 enables its use at significantlyhigher doses than those needed to stimulate an immuneresponse, without side effects[10]. Thus, the stability andsafety of the GPI-0100 derivative allow for the developmentof commercial vaccines with long shelf lives.
In has recently been claimed that the immune modulatingactivity of UFGPI-0100 was due to the presence of QS-7 inthe final product[11]. Examination of the methods reported
0264-410X/$ – see front matter © 2003 Elsevier Science Ltd. All rights reserved.doi:10.1016/S0264-410X(03)00298-6

3962 D.J. Marciani et al. / Vaccine 21 (2003) 3961–3971
to prepare the putative UFGPI-0100[11] indicates that an in-complete hydrolysis of QS-7 as well as the formation of highamounts of 1,3-dicyclohexylcarbodiimide (DCC) adducts ofDS saponins would be favored[12]. That the modified pro-cedure[11] allowed significant amounts of DCC and its in-termediates, both known to induce T-cell unresponsiveness,to remain in the preparation would explain the poor Th1immune response stimulated by the presumed UFGPI-0100[11,12].
Herein we report the separation of the UFGPI-0100derivatives into two fractions (RP18-1 and RP18-2) byreverse phase low-pressure liquid chromatography (RP-LPLC), and the determination of the structures for someof RP18-2 major components by mass spectrometry. Theimmunological properties of these fractions were comparedto those of alum and Q. saponins. Our results show thatGPI-0100’s Th1 immune stimulatory activity is a propertyof the dodecylamide derivatives.
2. Materials and methods
2.1. Synthesis of UFGPI-0100
UFGPI-0100 was prepared using a Q. saponin extract(Desert King International, San Diego, CA) accordingto the procedure previously described[10] with somemodifications. To the DS saponins (25.0 g, 15 mmol) dis-solved in 250 ml of pyridine, DCC (9.28 g, 45 mmol) andN-hydroxysuccinimide (NHS; 5.18 g, 45 mmol), each dis-solved in 125 ml of pyridine, were added with stirring. Then,a solution of dodecylamine (8.34 g, 45 mmol) in 125 ml ofpyridine was added drop-wise (over a 30 min period), andstirred for 30 h at room temperature. The reaction mixturewas then added to 1.5 l of deionized water and stirred for24 h to destroy excess reagents. The resulting suspensionwas filtered and the clear filtrate was evaporated in a ro-tary evaporator to near dryness. The residue was dissolvedin 1 l of 40 mM acetic acid, stirred for 24 h, and filtered.The clear solution was then diafiltrated in a hollow fibersystem using a 10,000 Da nominal MW cutoff membrane(QSM-02S, A/G Technology Corporation, Needham, MA)against 5 mM acetic acid (10 volumes), followed by deion-ized water until the pH of the diafiltrate was 6–7. Thissolution was then filtered and lyophilized. The resultantUFGPI-0100 was analyzed by HPLC using a Vydac C4column as previously described[10].
2.2. Reverse phase-LPLC
Prior to RP-LPLC, UFGPI-0100 was partially puri-fied by flash chromatography. About 2 g of UFGPI-0100dissolved in 30 ml of 97% methanol were applied to achromatography column (∼2.5 cm i.d.) packed with 20 gof LiChroprep RP-18 gel (40–63�m, EM Science, Gibb-stown, NJ) equilibrated with 97% methanol. GPI-0100 in
methanol was passed through the gel, and the gel waswashed with 30 ml of 97% methanol. The pooled solutionswere brought to dryness in a rotary evaporator to yield thepretreated UFGPI-0100. Chromatographic fractionationswere performed using a LP Combi Flash Sm 50 Foxy 200LC system (ISCO Inc., Lincoln, NE). Fourteen milliliterof methanol were added to 1 g of pretreated UFGPI-0100dissolved in 6 ml of deionized water. After 1 h, the solutionwas filtered through a 0.45�m filter and applied to a re-verse phase Lobar LiChroprep RP-18 B (319 mm× 25 mm)column (EM Science) equilibrated with 70% methanol inwater using a flow rate of 10 ml/min. The column was theneluted at a flow rate of 20 ml/min with a gradient from 70to 100% methanol in water for 40 min, and the effluent wasmonitored at 214 nm. Fractions containing the major chro-matographic peaks were pooled, analyzed by HPLC[10],and the solvent was removed in a rotary evaporator. Thedry material was then dissolved in about 10 ml of water andlyophilized.
2.3. LC–MS
An electrospray ion trap mass spectrometer (LCQ, Finni-gan MAT, San Jose, CA) coupled online with a HP1100 LCsystem (Hewlett-Packard, CA) was used for sample anal-ysis. A 2.1 mm × 250 mm Vydac C4 column was elutedfor 30 min with a 30–48% aqueous acetonitrile gradient in0.1% acetic acid, followed by 15 min of a 48–70% acetoni-trile gradient in 0.1% acetic acid. Chromatographic elutionwas performed using a flow rate of 0.2 ml/min and the ef-fluent was monitored at 214 nm. The ion trap mass spec-trometer was operated with an automated gain control inorder to maintain a constant level of ions in the trap. ALC–MS run was first performed in a full scan mode, andcompounds of interest were then subjected to LC–MS–MSanalysis.
2.4. Chemical analysis
The residual pyridine, NHS, DCC, dodecylamine, andmethanol contents of GPI-0100 preparations were deter-mined by gas chromatography (GC) by DHL AnalyticalLaboratories (Birmingham, AL). The protein content of theGPI-0100 preparation was determined by amino acid anal-ysis of samples hydrolyzed in 6N HCl. Endotoxin (EU/mg)was determined using the Limulus Amebocyte Lysate PY-ROCHOME assay (Associates of Cape Cod, Falmouth,MA).
2.5. Immunological studies
2.5.1. Adjuvants, antigens, cell lines and other reagentsAlHydroGel (2% alum) was purchased from Accurate
Chemical and Scientific Corp. (Westbury, NY). Ovalbumin(OVA; grade V) and concanavalin A were purchased from

D.J. Marciani et al. / Vaccine 21 (2003) 3961–3971 3963
Sigma (St. Louis, MO). Horseradish peroxidase (HRP) con-jugates of goat anti-mouse IgG1, IgG2a and IgG2b wereobtained from Southern Biotechnology Associates (Birm-ingham, AL). 3,3′,5,5′-Tetramethylbenzidine (TMB) waspurchased from Dako Corporation (Carpinteria, CA). EL4and E.G7-OVA mouse lymphoma cell lines were obtainedfrom the American Type Culture Collection (Rockville,MD). Cell culture media was obtained from GIBCO BRL(Gaithersburg, MD).
2.5.2. Immunization protocolsFemale BALB/c and C57BL/6 mice (6–8 weeks old) were
obtained from Charles River Laboratories (Raleigh, NC).Antigen/adjuvant mixtures were prepared in polypropylenetubes and incubated overnight at room temperature prior toinjection. Groups of six animals were immunized subcuta-neously (s.c.) on days 1, 15, and 29 with phosphate-bufferedsaline (PBS) or with 5�g OVA, alone or in the presenceof various doses of the adjuvants as indicated in the text.Animals were bled on day 43 and sera were prepared andfrozen for subsequent enzyme-linked immunosorbent assays(ELISA). Because the E.G7-OVA cell line was derived fromEL4 cells of C57BL/6 origin, this mouse strain was used forthe CTL studies.
2.5.3. Immunological assaysThe titers for IgG1, IgG2a, and IgG2b were determined in
pooled sera by ELISA as previously described[10], exceptthat the plates were coated with 50�g/ml OVA rather than5�g/ml. Lymphoproliferation and CTL assays were per-formed in triplicate using pooled spleen cells isolated frommice sacrificed 2 weeks after the third immunization[10].For CTL assays spleen cells (2×106 per well) from controland immunized mice were added to 24-well plates contain-ing 1 ml of complete medium plus 1× 105 E.G7-OVA cellsirradiated with 20,000 R. After 6 days at 37◦C in humidi-fied 5% CO2 cells were harvested and washed to yield theeffector (E) population. Target cells (T) consisted of EL4and E.G7-OVA cells that had been incubated for 1 h with300�Ci of 51Cr-labeled NaCrO4 (ICN Biomedicals, CostMesa, CA) and washed. Effectors and targets were added invarious E:T ratios to round-bottom 96-well microtiter plates(each well containing 1× 104 target cells), centrifuged for30 s at 200× g and incubated at 37◦C in a humidified 5%CO2 incubator. After 6 h, plates were harvested with the Ska-tron Supernatant Collection System (Sterling, VA) and theradioactivity in the supernatants was counted in a gammacounter and expressed as counts per minute (cpm). Con-trols for the CTL assay consisted of target cells lysed with2% Triton X-100 (maximum release) and target cells treatedwith medium only (spontaneous release). The percent lysisfor each E:T ratio was calculated by the formula:
lysis(%)
= experimental(cpm)−spontaneous release(cpm)
maximum release(cpm)−spontaneous release(cpm).
2.5.4. Determination of IFN-γ and IL-2Mice were sacrificed 2 weeks after the third immuniza-
tion (day 43). Spleens were harvested, pooled in groups,and single cell suspensions were prepared as described[10]and cultured (2× 105 per well) in duplicate in 96-well flatmicrotiter plates (Costar/Corning, Corning, NY) with eithermedium (no stimulus or background), 3�g/ml Con A, orOVA concentrations ranging from 0.5 to 10�g/ml. Cultureswere incubated at 37◦C in humidified air containing 5%CO2 for 18, 24, 48 or 72 h, and the supernatants were re-moved and frozen at−20◦C for subsequent assays. IFN-�and IL-2 were determined using the mouse IFN-� ELISAkit from Endogen (Woburn, MA) and the IL-2 OptEIATM
kit from BD-Pharmingen (San Diego, CA). The latter wasmodified for use with Milk Diluent/Blocking Solution fromKirkegaard & Perry Laboratories (Gaithersburg, MD), theblocking reagent for the IL-2 determination. Plates were readat 450 nm using a molecular devices plate reader.
3. Results
3.1. Chromatographic fractionation of GPI-0100 andLC–MS
Flash chromatography of UFGPI-0100 removed a 5–10%preparation contaminant that interfered with the RP-LPLC.The RP-LPLC of the pretreated UFGPI-0100 yielded asmall frontal peak F-1, which contained residual reactants,followed by two peaks, RP18-1 and RP18-2 (Fig. 1). TheRP18-1 solubility in water was lower than that of RP18-2,precipitating with time at room temperature for severalhours. The RP18-1 and RP18-2 fractions approximatelycorresponded to 12 and 33% of the pretreated UFGPI-0100.Some small peaks of the UFGPI-0100 (Fig. 2A) had aretention time (RT) close to QS-7 (∼14 min) but with a dif-ferent molecular weight (∼1670 Da). The HPLC profile ofRP18-1 showed six main peaks with RT from 20 to 32 minand molecular weight that corresponded to those of DSsaponin adducts ofN-dicyclohexylurea (Fig. 2B, Table 1)as proposed[11]. The RP18-2 peak, which representedone-third of UFGPI-0100, contained the major HPLC peaks
Table 1Molecular massesa for DCC adducts of DS saponins (RP18-1) and theircorresponding dodecylamide derivatives (RP18-2)
RP18-1 RTb m/z [M + H]+DCC
RP18-2RTb
m/z [M + H]+dodecyl
26.10 1896 32.28 185726.40 1763 32.44 172426.66 1881 32.52 184227.16 1749 32.91 171028.72 1733 34.90 169429.35 1720 35.25 1680
a Molecular mass of protonated molecular ions [M + H]+ (m/z).b Retention time (RT) (min).

3964 D.J. Marciani et al. / Vaccine 21 (2003) 3961–3971
Fig. 1. Reverse phase-LPLC fractionation of UFGPI-0100 on a LiChroprep RP-18 column. The fractions between the arrows were pooled and processedas described in the text.
with RT between 32 and 48 min. LC–MS analysis showedthat the main peaks were dodecylamide derivatives of DSsaponins, with molecular weight ranging from 1681 to1992 Da (Fig. 2C, Table 1). Structure determination ofthree of RP18-2 main components derived from proto-nated molecular and main fragment ions,m/z [M + H]+,confirmed their dodecylamide derivative nature. The peakwith a molecular weight of 1841 Da corresponded to thedodecylamide derivative of deacylated QS-17 and QS-18saponins (Fig. 3A), while the peak with a molecular weightof 1679 Da corresponded to the dodecylamide derivativeof deacylated QS-21 saponin (Fig. 3B). A peak with amolecular weight of 1709 Da apparently corresponds to anew Q. which saponin (Fig. 3C). A HPLC peak with RT45.84 min was found to have a molecular weight of 1991 Dathat corresponds to the dodecylamide derivative of deacy-lated QS-7 saponin (not shown). Chemical analysis of theRP18-2 fraction showed residual chemical concentrationsbelow GC limits of detection (Table 2).
3.2. Immunological studies
Similar to Q. saponins, UFGPI-0100 stimulates humoraland T-cell immunity with CTL production, but its immune
Table 2Chemical analysis of fraction RP18-2
Specification Content
Pyridinea <0.049%N-Hydroxysuccimidea <0.012%Dicyclohexylcarbodiimidea <0.029%Dodecylaminea <0.029%Methanola <0.038%Proteinb ∼0.00�g/mlEndotoxinc <0.125 EU/mg
a Values for chemical contaminants are expressed as below the limitof sensitivity of the GC instruments using a 20 mg sample.
b No amino acids were detected above the negative control amino acidlevels.
c 1 EU = 100 pg endotoxin.
stimulatory activity is 5–10 times lower than that of nativesaponins[10]. To determine the immune stimulating activityof the different GPI-0100 species and rule out residual QS-7as the source of immune stimulatory activity, several im-munological parameters were determined for UFGPI-0100as well as its two main fractions: RP18-1 and RP18-2.
UFGPI-0100 and its RP18-2 fraction elicited a Th1 an-tibody profile with a dose dependent stimulation of IgG1,IgG2a and IgG2b titers in BALB/c mice (Fig. 4). In con-trast, the RP18-1 fraction stimulated a Th2 antibody profilesimilar to that obtained with alum, i.e. an increase in IgG1with no significant increase in IgG2a and IgG2b titers(Fig. 4). In effect, the IgG1/IgG2a index determined forincreasing doses of these adjuvants shows a decrease forthe UFGPI-0100 and RP18-2 and an increase for RP18-1(Table 3), that corresponds to Th1 and Th2 antibody in-duction respectively. The stimulation of humoral immunityby RP18-1 was found to be about 10 times lower than thatof UFGPI-0100 or RP18-2. Q. saponins stimulated a Th1
Table 3Effect of UFGPI-0100, RP18-1 and RP18-2 over the induction of anti-OVAIgG1 and IgG2a production
Adjuvant IgG1 titer IgG2a titer IgG1/IgG2a
Alum (200�g) 70000 10 7000Q. saponins (10�g) 60000 1200 50
UFGPI-0100a (10�g) 70000 10 7000UFGPI-0100 (50�g) 110000 100 1100UFGPI-0100 (100�g) 110000 1100 100UFGPI-0100 (250�g) 230000 2900 79
RP18-1 (10�g) 24000 20 1200RP18-1 (50�g) 24000 20 1200RP18-1 (100�g) 39000 20 1950RP18-1 (250�g) 57000 40 1425
RP18-2 (10�g) 63000 80 788RP18-2 (50�g) 150000 3000 50RP18-2 (100�g) 260000 10000 26RP18-2 (250�g) 260000 11000 24
a Unfractionated GPI-0100 (UFGPI-0100).
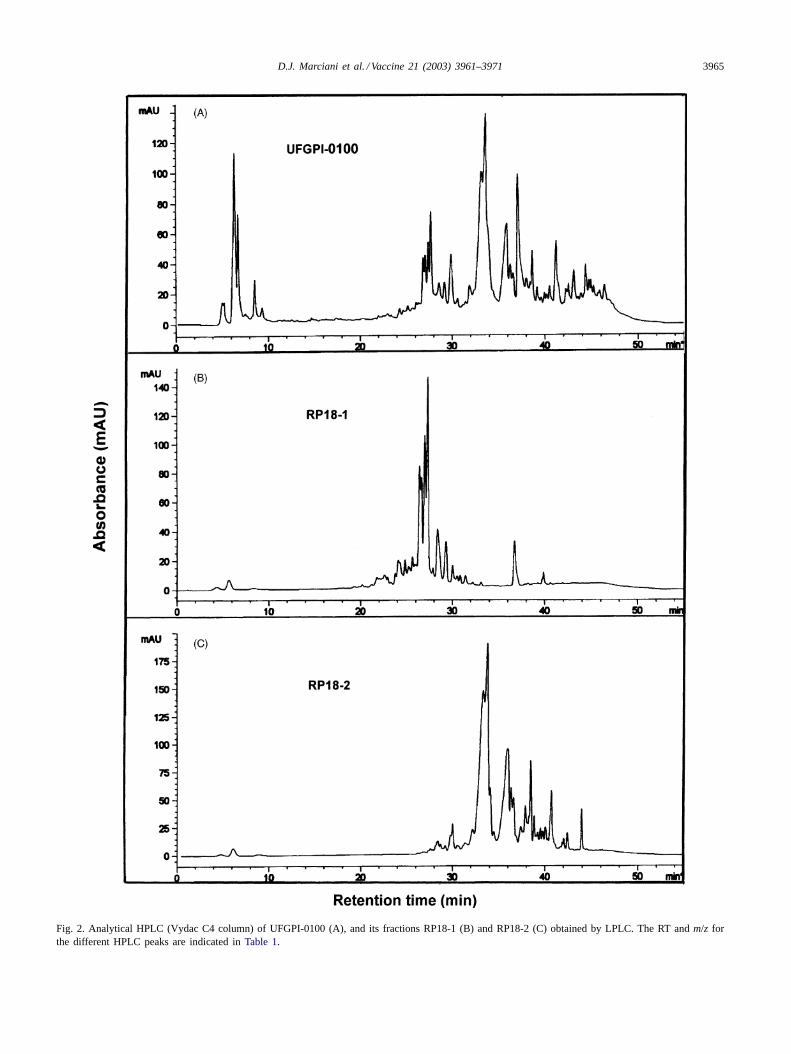
D.J. Marciani et al. / Vaccine 21 (2003) 3961–3971 3965
Fig. 2. Analytical HPLC (Vydac C4 column) of UFGPI-0100 (A), and its fractions RP18-1 (B) and RP18-2 (C) obtained by LPLC. The RT andm/z forthe different HPLC peaks are indicated inTable 1.
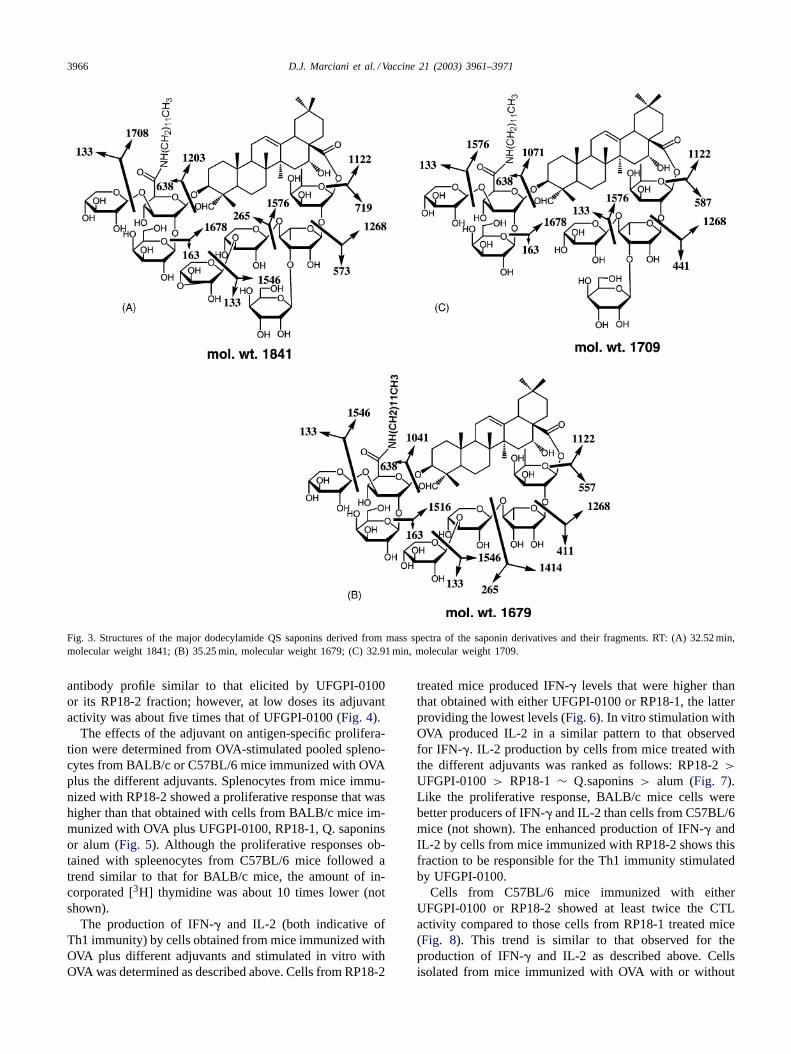
3966 D.J. Marciani et al. / Vaccine 21 (2003) 3961–3971
Fig. 3. Structures of the major dodecylamide QS saponins derived from mass spectra of the saponin derivatives and their fragments. RT: (A) 32.52 min,molecular weight 1841; (B) 35.25 min, molecular weight 1679; (C) 32.91 min, molecular weight 1709.
antibody profile similar to that elicited by UFGPI-0100or its RP18-2 fraction; however, at low doses its adjuvantactivity was about five times that of UFGPI-0100 (Fig. 4).
The effects of the adjuvant on antigen-specific prolifera-tion were determined from OVA-stimulated pooled spleno-cytes from BALB/c or C57BL/6 mice immunized with OVAplus the different adjuvants. Splenocytes from mice immu-nized with RP18-2 showed a proliferative response that washigher than that obtained with cells from BALB/c mice im-munized with OVA plus UFGPI-0100, RP18-1, Q. saponinsor alum (Fig. 5). Although the proliferative responses ob-tained with spleenocytes from C57BL/6 mice followed atrend similar to that for BALB/c mice, the amount of in-corporated [3H] thymidine was about 10 times lower (notshown).
The production of IFN-� and IL-2 (both indicative ofTh1 immunity) by cells obtained from mice immunized withOVA plus different adjuvants and stimulated in vitro withOVA was determined as described above. Cells from RP18-2
treated mice produced IFN-� levels that were higher thanthat obtained with either UFGPI-0100 or RP18-1, the latterproviding the lowest levels (Fig. 6). In vitro stimulation withOVA produced IL-2 in a similar pattern to that observedfor IFN-�. IL-2 production by cells from mice treated withthe different adjuvants was ranked as follows: RP18-2>
UFGPI-0100> RP18-1 ∼ Q.saponins> alum (Fig. 7).Like the proliferative response, BALB/c mice cells werebetter producers of IFN-� and IL-2 than cells from C57BL/6mice (not shown). The enhanced production of IFN-� andIL-2 by cells from mice immunized with RP18-2 shows thisfraction to be responsible for the Th1 immunity stimulatedby UFGPI-0100.
Cells from C57BL/6 mice immunized with eitherUFGPI-0100 or RP18-2 showed at least twice the CTLactivity compared to those cells from RP18-1 treated mice(Fig. 8). This trend is similar to that observed for theproduction of IFN-� and IL-2 as described above. Cellsisolated from mice immunized with OVA with or without

D.J. Marciani et al. / Vaccine 21 (2003) 3961–3971 3967
Fig. 4. Adjuvant effect on the IgG isotype profile. BALB/c mice were immunized s.c. on days 1, 15, and 29 with PBS only, OVA (5�g) alone or incombination with one of the following compounds: Q. saponins (10�g), alum (200�g), or UFGPI-0100, RP18-1 or RP18-2 at the doses indicated inthe figure. IgG isotype titers of pooled sera collected on day 43 were determined by ELISA. IgG1 (), IgG2a (�), and IgG2b ( ).
alum failed to stimulate CTL production. These resultsdemonstrate that the capacity to stimulate CTL productionresides in the RP18-2 fraction of UFGPI-0100.
4. Discussion
Substitution of the fatty acids from aldehyde-containingQ. saponins by an alkylamide chain at the glucuronic acidresidue yields stable derivatives (GPI-0100) that are ca-pable of stimulating Th1 immunity like the Q. saponins,with CTL production against exogenous antigens[10]. Al-though the immune stimulatory activity of UFGPI-0100 andRP18-2 is∼5 times lower than that of Q. saponins, theirnegligible toxicity allows them to be used at high doses toelicit an effective immune response without the side effectsof Q. saponins (e.g. QS-18 and QS-21). Studies in rabbitshave shown that a 7.5 mg dose of UFGPI-0100 had no ef-fects on mortality, clinical or gross pathology (Marciani,
unpublished observations). UFGPI-0100 has a less complexHPLC profile than Q. saponins[10], a result of replacingthe original acyl group, which is heterogeneous in its ter-minal sugar(s)[7], by a dodecylamide residue. However,due to the heterogeneity of the Q. saponins’ oligosaccha-ride chains[4,7], UFGPI-0100 has more components thanthose expected from only two DS saponins. For instance,QS-21 is composed of two isomers that differ in one sugarresidue[13] that are not resolved by HPLC on a Vydac C4column. In addition, two other isomers that share severalstructural characteristics with QS-21 have been reported[14]. That Q. saponins contain minor components with asingle oligosaccharide chain which are capable of formingdodecylamide derivatives may also contribute to the largernumber of derivatives found in UFGPI-0100[15,16].
Both UFGPI-0100 fractions, RP18-1 and RP18-2, con-tain DS saponin derivatives with different immune stimula-tory properties. RP18-1, the smaller and more hydrophilicfraction, elicited Th2 immunity (i.e. primarily IgG1). In

3968 D.J. Marciani et al. / Vaccine 21 (2003) 3961–3971
Fig. 5. Adjuvant effect on the antigen-specific lymphoproliferative response. Female BALB/c mice were immunized s.c. on days 1, 15, and 29 withPBS only, OVA (5�g) alone or in combination with one of the following compounds: Q. saponins (10�g), alum (200�g), or UFGPI-0100, RP18-1 orRP18-2 at the doses indicated in the figure. On day 43, the spleens were removed and pooled. Single cell suspensions prepared and incubated for∼56 hwith 5�g/ml OVA and pulsed with3H thymidine for an additional 16 h. Cells were then harvested and3H thymidine incorporation was determined in aliquid scintillation counter and expressed as cpm. OVA (), alum (�), Q. saponins ( ), UFGPI-0100 ( ), RP18-1 ( ), and RP18-2 ( ).
Fig. 6. IFN-� responses from the spleens of BALB/c mice immunized s.c. on days 1, 15, and 29 with OVA (5�g) alone or in combination with one ofthe following compounds: Q. saponins (10�g), alum (200�g), or UFGPI-0100, RP18-1 or RP18-2 at the doses indicated in the figure. Control animalsreceived PBS only (levels below detection; data not shown). Single cell suspensions of pooled splenocytes were prepared on day 43 and cultured with 2and 10�g/ml of OVA for 24 and 48 h. In vitro IFN-� cytokine levels in spleen cell cultures derived from immunized BALB/c mice were determined byELISA. OVA (�), Q. saponins ( ), alum ( ), UFGPI-0100 ( ), RP18-1 ( ), and RP18-2 (�).

D.J. Marciani et al. / Vaccine 21 (2003) 3961–3971 3969
Fig. 7. IL-2 responses from BALB/c mice immunized s.c. on days 1, 15, and 29 with OVA (5�g) alone or in combination with one of the followingcompounds: Q. saponins (10�g), alum (200�g), or UFGPI-0100, RP18-1 or RP18-2 at the doses indicated in the figure. In vitro IL-2 cytokine levels inspleen cell cultures derived from immunized BALB/c mice were determined by ELISA. Single cell suspensions of pooled splenocytes were prepared on day43 and cultured with 0.5 and 10�g/ml of OVA for 18 and 48 h. OVA (�), alum ( ), Q. saponins ( ), UFGPI-0100 ( ), RP18-1 ( ), and RP18-2 (�).
contrast, the larger and more hydrophobic fraction RP18-2stimulated Th1 immunity (i.e. IgG2a and IgG2b, as wellas elevated levels of IFN-� and IL-2). The significantlystronger immune modulating properties of RP18-2 as com-pared to those of RP18-1 could explain the stimulation ofTh1 immunity by UFGPI-0100. Although the structures ofthe RP18-1 components have not been completely eluci-dated, their inability to stimulate Th1 immunity suggeststhe absence of a co-stimulatory aldehyde group[17,18], or alipophilic group linked to the glucuronic acid residue lead-ing to a micelle formation that limits the aldehyde group’savailability to act as a co-stimulatory agent. Although asmall fraction of Q. saponins have a triterpenoid moietydevoid of aldehyde groups[19], their relative contributionto immune stimulation should be small. From the molec-ular weight distribution RP18-1 seems to be composed ofDCC adducts of DS saponins, which are by-products ofthe amidation reaction. In effect, UFGPI-0100 preparedunder sub-optimal conditions showed a significant increasein the relative amounts of DCC adducts, a result of anexcessive side reaction yieldingN-acylurea by-products[12] (not shown). An explanation for the similar adju-vanticities of UFGPI-0100 and RP18-2 may be mixedmicelles of RP18-1 and RP18-2. Below certain relative
RP18-1:RP18-2 proportions, their components may assem-ble to allow the aldehyde groups to provide co-stimulatorysignals for T-cell activation. Elucidation of the structureof several RP18-2 components showed that they are dode-cylamide derivatives of deacylated QS-7, QS-17, QS-18,QS-21 and other DS saponins. Thus, dodecylamide deriva-tives of DS saponins have the capacity to elicit Th1 im-munity with CTL production, although at higher dosesthan those needed with Q. saponins. The lower adjuvan-ticity of the GPI-0100 species may be due to its increasedhydrophobic properties relative to those of Q. saponins,leading to micelle formation at lower concentrations. Se-questering of GPI-0100 species by micelle formation woulddecrease its availability to stimulate antigen-presentingcells (APC). Likewise, the addition of non-ionic detergentsto GPI-0100 solutions could result in the formation of alarge number of mixed micelles carrying GPI-0100 andenhance the stimulation of APCs. Indeed, a formulationof UFGPI-0100 with Tween 40 resulted in a 10–100-foldincrease in the production of the different anti-OVAIgG isotypes (Marciani et al., unpublished observations).
Recently, it has been reported that UFGPI-0100 stimu-lates only Th2 immunity and that any Th1 immune stimu-lation is due to residual QS-7 resistance to deacylation and
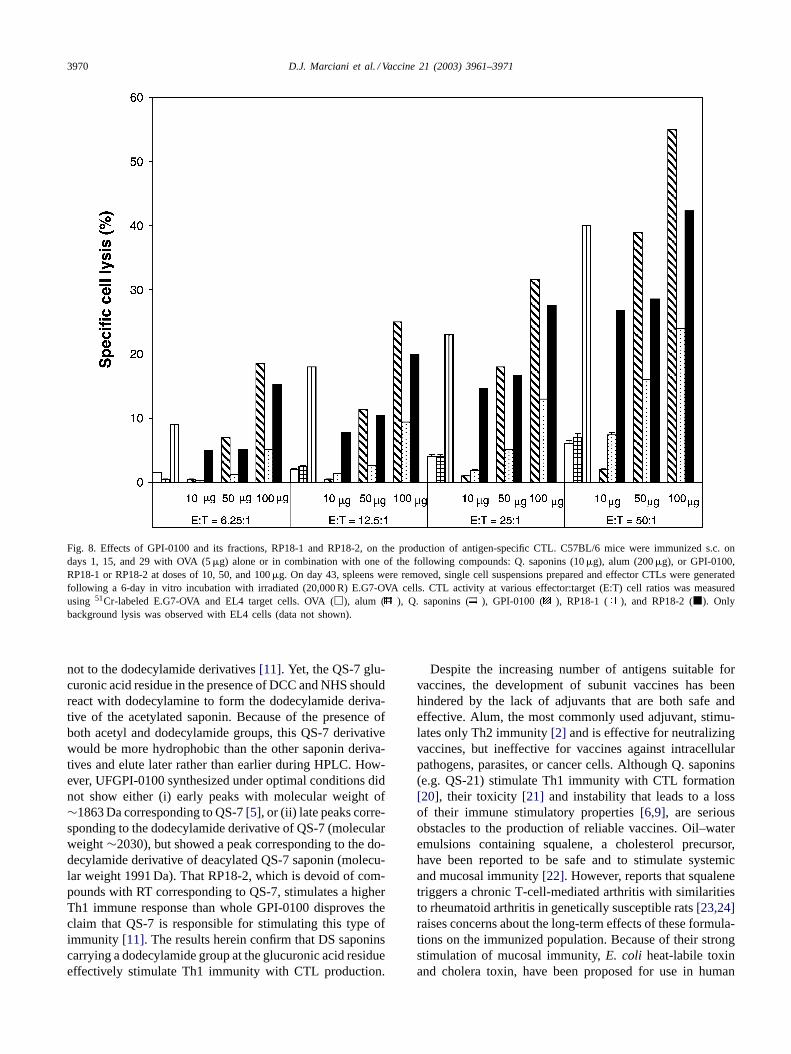
3970 D.J. Marciani et al. / Vaccine 21 (2003) 3961–3971
Fig. 8. Effects of GPI-0100 and its fractions, RP18-1 and RP18-2, on the production of antigen-specific CTL. C57BL/6 mice were immunized s.c. ondays 1, 15, and 29 with OVA (5�g) alone or in combination with one of the following compounds: Q. saponins (10�g), alum (200�g), or GPI-0100,RP18-1 or RP18-2 at doses of 10, 50, and 100�g. On day 43, spleens were removed, single cell suspensions prepared and effector CTLs were generatedfollowing a 6-day in vitro incubation with irradiated (20,000 R) E.G7-OVA cells. CTL activity at various effector:target (E:T) cell ratios was measuredusing 51Cr-labeled E.G7-OVA and EL4 target cells. OVA (�), alum ( ), Q. saponins ( ), GPI-0100 ( ), RP18-1 ( ), and RP18-2 (�). Onlybackground lysis was observed with EL4 cells (data not shown).
not to the dodecylamide derivatives[11]. Yet, the QS-7 glu-curonic acid residue in the presence of DCC and NHS shouldreact with dodecylamine to form the dodecylamide deriva-tive of the acetylated saponin. Because of the presence ofboth acetyl and dodecylamide groups, this QS-7 derivativewould be more hydrophobic than the other saponin deriva-tives and elute later rather than earlier during HPLC. How-ever, UFGPI-0100 synthesized under optimal conditions didnot show either (i) early peaks with molecular weight of∼1863 Da corresponding to QS-7[5], or (ii) late peaks corre-sponding to the dodecylamide derivative of QS-7 (molecularweight∼2030), but showed a peak corresponding to the do-decylamide derivative of deacylated QS-7 saponin (molecu-lar weight 1991 Da). That RP18-2, which is devoid of com-pounds with RT corresponding to QS-7, stimulates a higherTh1 immune response than whole GPI-0100 disproves theclaim that QS-7 is responsible for stimulating this type ofimmunity [11]. The results herein confirm that DS saponinscarrying a dodecylamide group at the glucuronic acid residueeffectively stimulate Th1 immunity with CTL production.
Despite the increasing number of antigens suitable forvaccines, the development of subunit vaccines has beenhindered by the lack of adjuvants that are both safe andeffective. Alum, the most commonly used adjuvant, stimu-lates only Th2 immunity[2] and is effective for neutralizingvaccines, but ineffective for vaccines against intracellularpathogens, parasites, or cancer cells. Although Q. saponins(e.g. QS-21) stimulate Th1 immunity with CTL formation[20], their toxicity [21] and instability that leads to a lossof their immune stimulatory properties[6,9], are seriousobstacles to the production of reliable vaccines. Oil–wateremulsions containing squalene, a cholesterol precursor,have been reported to be safe and to stimulate systemicand mucosal immunity[22]. However, reports that squalenetriggers a chronic T-cell-mediated arthritis with similaritiesto rheumatoid arthritis in genetically susceptible rats[23,24]raises concerns about the long-term effects of these formula-tions on the immunized population. Because of their strongstimulation of mucosal immunity,E. coli heat-labile toxinand cholera toxin, have been proposed for use in human

D.J. Marciani et al. / Vaccine 21 (2003) 3961–3971 3971
vaccines[25,26]. Yet, an experimental influenza vaccinecontainingE. coli heat-labile toxin has recently been linkedto Bell’s palsy, pointing to the risks of using these toxins asimmune enhancers or adjuvants. UFGPI-0100 and particu-larly its fraction RP18-2 appear to fulfill all the requirementsfor safe and effective adjuvants. Because RP18-2 containsonly dodecylamide derivatives from UFGPI-0100 and all theTh1 adjuvanticity resides in this fraction, we propose to here-after apply the term GPI-0100 to the RP18-2 fraction. Unlikemost adjuvants that stimulate Th1 immunity, GPI-0100 has alarge margin of safety. In mice, the ratio of acute toxic doseto effective dose is about 40–50, which allows the GPI-0100dose to be significantly increased to achieve the desiredimmune response with no early onset of toxicity. In effect,it has been recently reported that because GPI-0100 is lesstoxic than QS-21, greater doses of GPI-0100 can be safelyadministered with a commensurate increase in antibodyproduction[27]. That GPI-0100 stimulates strong humoralimmunity, T-cell immunity with CTL production, as wellas mucosal immunity (Michalek, personal communication)would facilitate the development of new subunit vaccineswhere combinations of these immunities will be required,such as tuberculosis, herpes simplex virus, and infectiousagents used as bioterrorism agents. Finally, its stabilityunder a broad range of temperatures, pH, carriers, and anti-gens, would assure the maintenance of a vaccine’s efficacy.
References
[1] Kensil CR, Patel U, Marciani D. Separation and characterizationof saponins with adjuvant activity fromQuillaja saponaria Molinacortex. J Immunol 1991;146:431–7.
[2] HogenEsch H. Mechanism of stimulation of the immune responseby aluminum adjuvants. Vaccine 2002;20:S34–9.
[3] Soltysik S, Wu J-Y, Recchia J, Wheeler DA, Newman MJ, et al.Structure/function studies of QS-21 adjuvant: assessment of triterpenealdehyde and glucuronic acid roles in adjuvant function. Vaccine1995;13:1403–10.
[4] Press JB, Reynolds RC, May RD, Marciani DJ. Structure/functionrelationships of immunostimulating saponins. Stud Nat Prod Chem2000;24:131–74.
[5] Kensil CR, Wu JY, Anderson CA, Wheeler DA, Amsden J. QS-21and QS-7: purified saponin adjuvants. Dev Biol Stand 1998;92:41–7.
[6] Marciani DJ, Pathak AK, Reynolds RC, Seitz L, May RD. Alteredimmunomodulating and toxicological properties of degradedQuillajasaponaria Molina saponins. Int Immunopharmacol 2001;1:813–8.
[7] Higuchi R, Tokimitsu Y, Fujioka T, Komori T, Kawasaki T, et al.Structure of desacylsaponins obtained from the bark ofQuillajasaponaria. Phytochemistry 1987;26:229–35.
[8] Kensil C, Soltysik S, Patel U, Marciani DJ. Structure–functionrelationship in adjuvants fromQuillaja saponaria Molina. In: BrownF, Chanock RM, Ginsberg HS, Lerner RA, editors. Vaccine 92. ColdSpring Harbor (NY): Cold Spring Harbor Laboratory Press; 1992.p. 35–40.
[9] Marciani DJ, Ptak RG, Voss TG, Reynolds RC, Pathak AK, etal. Degradation ofQuillaja saponaria Molina saponins: loss of theprotective effects of a herpes simplex virus 1 sub-unit vaccine. IntImmunopharmacol 2002;2:1703–11.
[10] Marciani DJ, Press JB, Reynolds RC, Pathak AK, Pathak V, et al.Development of semisynthetic triterpenoid saponin derivatives withimmune stimulating activity. Vaccine 2000;18:3141–51.
[11] Liu G, Anderson C, Scaltreto H, Barbon J, Kensil CR. QS-21structure/function studies: effects of acylation on adjuvant activity.Vaccine 2002;20:2808–15.
[12] Marciani DJ, Pathak AK, Reynolds RD.Quillaja saponinadjuvants: derivatives formed under sub-optimal conditions. Vaccine2002;20:3237–8.
[13] Soltysik S, Bedore DA, Kensil CR. Adjuvant activity of QS-21isomers. Ann New York Acad Sci 1993;690:392–5.
[14] Nord LI, Kenne L. Separation and structural analysis of saponinsin a bark extract fromQuillaja saponaria Molina. Carbohydr Res1999;320:70–81.
[15] So HS, Yoon HS, Choi DY, Kwon YS, Sung JH, et al. Effect ofa novel saponin adjuvant derived fromQuillaja saponaria on theimmune response to recombinant hepatitis B surface antigen. MolCells 1997;7:178–86.
[16] Guo S, Kenne L, Lundgren LN, Ronnberg B, Sundquist BG.Triterpenoid saponins fromQuillaja saponaria. Phytochemistry1998;48:175–80.
[17] Rhodes J. Covalent chemical events in immune induction:fundamental and therapeutic aspects. Immunol Today 1996;17:436–41.
[18] Chen H, Rhodes J. Schiff base forming drugs. Mechanismsof immune potentiation and therapeutic potential. J Mol Med1996;74:497–504.
[19] Guo S, Kenne L. Structural studies of triterpenoid saponins with newacyl components fromQuillaja saponaria Molina. Phytochemistry2000;55:419–28.
[20] Kensil CR, Wu J-Y, Soltysik S. Structural and immunologicalcharacterization of the vaccine adjuvant QS-21. In: Powell MF,Newman MJ, editors. Vaccine design: the subunit and adjuvantapproach. New York: Plenum Press; 1995. p. 525–41.
[21] Livingston PO, Adluri S, Helling F, Yao TJ, Kensil CR, etal. Phase 1 trial of immunological adjuvant QS-21 with aGM2 ganglioside-keyhole limpet haemocyanin conjugate vaccine inpatients with malignant melanoma. Vaccine 1994;12:1275–80.
[22] O’Hagan DT. Recent advances in vaccine adjuvants for systemic andmucosal administration. J Pharm Pharmacol 1998;49:1–10.
[23] Carlson BC, Jansson AM, Larsson A, Butch A, Lorentzen JC. Theendogenous adjuvant squalene can induce a chronic T-cell mediatedarthritis in rats. Am J Pathol 2000;156:2057–65.
[24] Holm BC, Svelander L, Butch A, Lorentzen JC. The arthritogenicadjuvant squalene does not accumulate in joints, but gives rise topathogenic cells in both draining and non-draining lymph nodes.Clin Exp Immunol 2002;127:430–5.
[25] Glueck R. Pre-clinical and clinical investigation of the safety of anovel adjuvant for intranasal immunization. Vaccine 2002;20:S42–4.
[26] Del Giudice G, Podda A, Rappuoli R. What are the limits ofadjuvanticity? Vaccine 2002;20:S38–41.
[27] Ragupathi G, Coltart DM, Williams LJ, Koide F, Kagan E, et al.On the power of chemical synthesis: immunological evaluation ofmodels for multiantigenic carbohydrate-based cancer vaccines. ProcNatl Acad Sci USA 2002;99:13699–704.
Copyright © 2022 FDOKUMEN




















