
Palynologische Untersuchungen über die holozäne Vegetations-, Klima- und Siedlungsgeschichte in...
193
Palynologische Untersuchungen über die holozäne Vegetations-, Klima- und Siedlungsgeschichte in Hochasien (Nanga Parbat, Karakorum, Nianbaoyeze, Lhasa) und das Pleistozän in China (Qinling-Gebirge, Gaxun Nur) Dissertation zur Erlangung des Doktorgrades der Mathematisch-Naturwissenschaftlichen Fakultäten der Georg-August-Universität zu Göttingen vorgelegt von Frank Schlütz aus Nieheim Göttingen 1998 Erschienen 1999 in Dissertationes Botanicae, Bd. 315, 183 Seiten.
-
Upload
uni-goettingen -
Category
Documents
-
view
1 -
download
0
Transcript of Palynologische Untersuchungen über die holozäne Vegetations-, Klima- und Siedlungsgeschichte in...

Palynologische Untersuchungen über die holozäne Vegetations-, Klima- und Siedlungsgeschichte in Hochasien (Nanga Parbat, Karakorum, Nianbaoyeze, Lhasa) und das
Pleistozän in China (Qinling-Gebirge, Gaxun Nur)
Dissertation zur Erlangung des Doktorgrades
der Mathematisch-Naturwissenschaftlichen Fakultäten der Georg-August-Universität zu Göttingen
vorgelegt von Frank Schlütz
aus Nieheim
Göttingen 1998
Erschienen 1999 in Dissertationes Botanicae, Bd. 315, 183 Seiten.

Anschrift des Verfassers: Frank Schlütz
Albrecht-v.-Haller Institut für Pflanzenwissenschaften Abteilung für Palynologie und Quartärwissenschaften
Wilhelm-Weber-Str.2 D-37073 Göttingen
Gefördert mit Mitteln der Deutschen Forschungsgemeinschaft (Be 169/13) Dedruckt mit Unterstützung der Deutschen Forschungsgemeinschaft D7 Aktuelle Anschrift: Dr. Frank Schlütz Abteilung für Palynologie und Klimadynamik Albrecht-v.-Haller Institut für Pflanzenwissenschaften Untere Karspüle 2 37073 Göttingen [email protected] http://www.palynologie.uni-goettingen.de/

Inhaltsverzeichnis
1 Einleitung ................................................................................................... 1
2 Arbeitsgebiet und Fragestellungen......................................................... 2
3 Stand der Forschung ................................................................................ 5
4 Material und Methoden ............................................................................. 8
4.1 Profile und Pollendiagramme..................................................... 8
4.2 Probenaufbereitung..................................................................... 8
4.3 Darstellung der Ergebnisse........................................................ 9
4.4 Die palynologische Ansprache ................................................10
4.5 Übersicht der auftretenden Taxa .............................................11
4.6 Vegetation und Pollenniederschlag.........................................27
5 Der Untersuchungsraum ........................................................................30
5.1 Lage und Geologie ....................................................................30
5.2 Klima ...........................................................................................31
5.3 Vegetation ..................................................................................32
5.4 Vegetations- und Florengeschichte.........................................35
5.5 Siedlungsgeschichte.................................................................37
5.5.1 China ............................................................................................... 37
5.5.2 Tibet................................................................................................. 41
5.5.3 Karakorum ....................................................................................... 43
5.6 Ernährung...................................................................................45
5.6.1 Tibet................................................................................................. 45
5.6.2 Karakorum ....................................................................................... 46
6 Ergebnisse und Diskussionen...............................................................48
6.1 Lingfeng im Qinling-Gebirge ....................................................48

6.1.1 Das Untersuchungsgebiet................................................................ 48
6.1.2 Das Profil Lingfeng........................................................................... 52
6.2 Der Gaxun Nur in der Inneren Mongolei..................................61
6.2.1 Das Untersuchungsgebiet................................................................ 61
6.2.2 Das Profil Gaxun Nur – Erste Voruntersuchungen........................... 62
6.3 Das Hochland von Tibet ............................................................67
6.3.1 Südöstliches Tibet: Der Nianbaoyeze .............................................. 67
6.3.1.1 Das Untersuchungsgebiet.......................................................... 67
6.3.1.2 Rezenter Pollenniederschlag ..................................................... 71
6.3.1.3 Die Profile .................................................................................. 73
6.3.1.4 Diskussion ................................................................................. 85
6.3.2 Südliches Tibet: Im Stadtbereich von Lhasa.................................... 91
6.3.2.1 Das Untersuchungsgebiet.......................................................... 91
6.3.2.2 Das Profil Lhasa Stadtbereich ................................................... 93
6.3.2.3 Diskussion ................................................................................. 94
6.3.3 Weitere Untersuchungen in Tibet..................................................... 95
6.3.3.1 Sodenprofile............................................................................... 95
6.3.3.2 Weitere Profile ........................................................................... 96
6.4 Nanga Parbat und Karakorum ..................................................97
6.4.1 Das Untersuchungsgebiet................................................................ 97
6.4.2 Die Profile ...................................................................................... 105
6.4.2.1 Der Nanga Parbat .................................................................... 105
6.4.2.2 Der Karakorum ........................................................................ 109
6.4.3 Diskussion...................................................................................... 116
7 Zusammenfassung ...............................................................................119
7.1 Pleistozän .................................................................................119
7.1.1 Gaxun Nur...................................................................................... 119
7.1.2 Lingfeng ......................................................................................... 119

7.2 Holozän .....................................................................................119
7.2.1 Nianbaoyeze .................................................................................. 119
7.2.2 Lhasa ............................................................................................. 120
7.2.3 Nanga Parbat................................................................................. 120
7.2.4 Karakorum ..................................................................................... 121
7.2.5 Klimageschichtliche Ergebnisse: Ein überregionaler Vergleich...... 121
8 Summary ................................................................................................122
8.1 Pleistocene...............................................................................122
8.1.1 Gaxun Nur (Inner Mongolia)........................................................... 122
8.1.2 Lingfeng (E-China)......................................................................... 122
8.2 Holocene...................................................................................122
8.2.1 Nianbaoyeze (E-Tibet) ................................................................... 122
8.2.2 Lhasa (S-Tibet) .............................................................................. 123
8.2.3 Nanga Parbat (7126 m, NW-Himalaya) ......................................... 123
8.2.4 Karakoram (N-Pakistan) ................................................................ 123
8.2.5 Conclusions ................................................................................... 124
9 Nachwort und Danksagung .................................................................125
10 Literaturverzeichnis...........................................................................127
11 Anhang................................................................................................165
11.1 Index der Pollentypen in Kap. 4.5 ..........................................165
11.2 Liste der Profile........................................................................168
11.3 Liste der Abbildungen .............................................................174
11.4 Verwendete Kürzel und fremdsprachliche Begriffe .............176
11.5 Tabellen ....................................................................................177
11.6 Fototafel ....................................................................................182

1. Einleitung 1
1 Einleitung
Mit den palynologischen Untersuchungen der während der ersten deutsch-chinesischen Expedition im Jahre 1981 gewonnenen Torfprofile begann eine enge Zusammenarbeit zwischen in Hochasien tätigen Geographen und dem Institut für Palynologie und Quartär-wissenschaften der Universität Göttingen (HÖVERMANN & WANG WENYING 1987, BEUG
1987, LEHMKUHL & LIU SHIJIAN 1994a). Daraus ergab sich die vorliegende Arbeit, die durch das Mitwirken von Wissenschaftlern der Universitätsstandorte Göttingen, Marburg, Berlin und Bonn möglich wurde. Die Menge des zur Verfügung gestellten Materials erlaubt es, die räumliche Betrachtung der heutigen Verknüpfung von Klima und Vegetation (TROLL 1939, 1967, 1978, SCHWEINFURTH 1957, MIEHE et al. 1996) in die Vergangenheit auszudehnen und dabei nach den wechselseitigen Beziehungen zwischen naturräumlicher Ausstattung und menschlicher Nutzung zu fragen (SCHWEINFURTH 1983, BEUG & MIEHE 1997, 1999). Die pollenanalytischen Ergebnisse ergänzen das aus anderen Quellen gewonnene Bild über Dauer und Wirkung der menschlichen Einflußnahme (JETTMAR 1993, 1980a, SCHWEINFURTH
1983, JACOBSEN & SCHICKHOFF 1995, KREUTZMANN 1996, HUANG WEIWEN 1994) und sichern es ab. In diesem Zusammenhang sind die Übergangsräume von der Peripherie zum Tibetanischen Plateau, in der vorliegenden Arbeit vertreten durch das nördliche Pakistan (Karakorum, Nanga Parbat) und den Ostrand des Hochlandes von Tibet (Nianbaoyeze), von besonderem Interesse (SCHWEINFURTH 1982, 1986).
Nicht zu übersehen ist der heutige Einfluß des Tibetanischen Plateaus auf das globale Klimageschehen und die seiner allmählichen Hebung zugestandene Bedeutung für die Klimageschichte der letzten Jahrmillionen (BLÜTHGEN & WEISCHET 1980, FLOHN 1968, 1981, KUHLE 1987, 1993, LI JIJUN et al. 1995, LIU TUNGSHENG & DING ZHONGLI 1993, PANT
& RUPA KUMAR 1997, ZHENG BENXING 1989, ZHOU JINSHANG 1987 u. a.).
Ziel der vorliegenden Arbeit ist es, angesichts der Weite des Gebietes gut abgesicherte exemplarische Einblicke in die holozäne Vegetationsgeschichte und die sie bedingenden klimatischen Änderungen und anthropo-zoogenen Einflüsse zu erlangen. Durch die Unter-suchung des Profils „Lingfeng“ aus dem östlich des Tibetanischen Plateaus gelegenen Ge-birge des Qinling kann auch die Entwicklung der Pflanzendecke im letzten Glazial dargelegt werden.

2. Arbeitsgebiet und Fragestellungen 2
2 Arbeitsgebiet und Fragestellungen
Bei den Arbeiten zur holozänen Vegetations- und Siedlungsgeschichte lag einer der räum-lichen Schwerpunkte im Gebirge des Nianbaoyeze am Ostrand des Tibetischen Plateaus, ergänzt um die Untersuchungen zum jüngeren Holozän Südtibets bei Lhasa. Den zweiten Schwerpunkt bildeten der im NW-Himalaja gelegene Nanga Parbat mit dem sich im nördlichen Pakistan anschließenden Gebirgsraum des Karakorum (Abb. 1).
Beide Räume liegen zwischen Gebieten mit feuchtem subtropischen Klima, den meri-dionalen Stromfurchen bzw. dem äußeren Himalaja, und den trockenen Hochlagen Tibets. Sie weisen das den inneren Himalaja und den chinesisch/ostasiatisch-tibetischen Übergangsraum mit seinen typischen Trockentälern kennzeichnende, mäßig feuchte Klima auf (SCHWEINFURTH 1986,1993).
Unterschiede bestehen hier in der Niederschlagsgenese. Im östlichen Tibet bringt über-wiegend der aus südöstlichen Richtungen kommende Sommermonsun die Feuchtigkeit, während die von Norden kommenden winterlichen Schneefälle ausgesprochen gering sind. Im nördlichen Pakistan treten hingegen Winter- und Frühjahrs-Niederschläge zu den sommerlichen Regenfällen des Südwestmonsuns hinzu, die an die aus dem östlichen Mittelmeerraum heranziehenden Tiefdruckstörungen gebunden sind (BÖHNER 1996, WEIERS
1995, ZHAO SONGQIAO 1994).
Der klimatische Übergangscharakter der untersuchten Gebiete bringt es mit sich, daß die Vegetation eine deutlich expositionsabhängige Verteilung zeigt. Im Nianbaoyeze, an der Grenze der Baumverbreitung gelegen, stehen die wenigen Bestände von Fichten an Nordhängen, die Wacholderbäume dagegen an Südhängen. Eine untere, trockenheits-bedingte Baumgrenze fehlt im Nianbaoyeze, findet sich aber unweit in den Flußtälern von Huang He und Minjing (Abb. 7.) Dort sind Trockentäler ähnlich denen des Himalaja ent-wickelt (HOU HSIOHYU 1983, SCHWEINFURTH 1986).
Am Nanga Parbat wird der Talboden entlang des Indus von einer Chenopodiaceen-Wüste eingenommen. Feuchttemperierte Nadelwälder mit Kiefer und Fichte treten erst in höheren Hanglagen bei stärkeren Niederschlägen auf. Zum Inneren des Karakorum hin treten die feuchten Nadelwälder zunächst auf den Südhängen zurück, bis schließlich alle Expositionen von der Artemisia-Steppe beherrscht werden (TROLL 1939, PAFFEN et al. 1956, MIEHE et al. 1996, SCHICKHOFF 1995).
Die pollenanalytischen Untersuchungen sollten nicht allein Auskünfte zur holozänen Vegetationsentwicklung geben, sondern auch mögliche Ursachen für die so augenfällige expositionsdifferenzierte Vegetationsverteilung der Übergangsräume klären. Erste Ergeb-nisse hierzu sind bereits veröffentlicht (SCHLÜTZ 1995, 1997).

2. Arbeitsgebiet und Fragestellungen 3
Abb. 1: Naturräumliche Gliederung Chinas nach ZHAO SONGQIAO (1994) in die Regionen A = Ost-China, B = Nord-China und C = Tibetanisches Plateau (Qinghai-Xizang-Plateau) mit den im Text erwähnten Unterregionen 1 = Südöstliches-Tibet, 2 = Süd-Tibet, 3 = Qaidam-Becken, 4 = Qinling Gebirge und Dabie Shan, 5 = Rotes Becken, 6 = Ordos-Gebiet, 7 = Alaschan Plateau, 8 = Tarim-Becken sowie Lage der Untersuchungsräume (Karakorum/Nanga Parbat, Nianbaoyeze) und einiger Untersuchungspunkte (Lhasa, Lingfeng, Gaxun Nur).

2. Arbeitsgebiet und Fragestellungen 4
Das Profil „Lingfeng“ erschloß den Zeitraum des letzten Glazials. Es stammt aus dem Qinling, jenem Gebirge, das im östlichen China Teil der Wasserscheide zwischen den großen Strömen des Huang He und des Yangzi ist. Zudem verläuft hier die klimatische, tier- und pflanzengeographische sowie kulturelle Grenze zwischen dem Norden und dem Süden Chinas (BLUNDEN & ELVIN 1992, ERKES 1956, ZHAO SONGQIAO 1994). Durch die Lage im Grenzbereich des heutigen Sommermonsuns konnte mit deutlichen klimabedingten Vegetationsveränderungen gerechnet werden (SCHLÜTZ 1998).
Einen gewissen Einblick in noch älterer Abschnitte der Vegetationsgeschichte vermit-teln erste, aber noch sehr vorläufige Untersuchungen an einem etwa 1 Mio. Jahre zurück-reichenden Bohrkern aus der Inneren Mongolei (Profil „Gaxun Nur“).

3. Stand der Forschung 5
3 Stand der Forschung
Die bisher veröffentlichten pollenanalytischen Untersuchungen verteilen sich sehr un-gleichmäßig auf die in der vorliegenden Arbeit behandelten Räume. Zur holozänen Vegetationsgeschichte des Nianbaoyeze und seiner weiteren Umgebung liegt die Arbeit von WANG MANHUA (1987) vor, die mit dem Profil „Hongyuan“ auch das weitflächig vermoorte Becken von Zoige berücksichtigt. Pollendiagramme aus dem selben Gebiet veröffentlichten später auch THELAUS (1992) und FRENZEL (1994). Zur holozänen Entwicklung im nordöstlichen Hochland von Tibet seien die Arbeit von BEUG (1987) und jene über den Qinghai-See (Koko Nor) von KONG ZHAOCHEN et al. (1990) sowie LISTER et al. (1991) genannt. Palynologische Untersuchungen an Sedimenten verschiedener Seen Tibets führten HUANG CIXUAN & LIANG YULIAN (1981) durch. Die zusammenfassenden Darstellungen zum Holozän Chinas von FENG ZHAODONG et al. (1993), JI JINJUN (1996), LI TIANCHI (1988), SUN
XIANGJUN & CHEN YINSHUO (1991a, b), WANG FUBAO & FAN (1987) und ZHOU S. Z. et al. (1991) berücksichtigen auch weniger bekannte Arbeiten. Palynologische Untersuchungen zum Nanga Parbat und Karakorum liegen bislang nicht vor. Aus dem nördlich angrenzenden Hindukusch stammt ein Pollendiagramm von BORTENSCHLAGER & PATZELT
(1978), das etwa 2000 Jahre zurückreicht.
Der Verlauf der holozänen Vegetationsgeschichte läßt sich nach der oben angeführten Literatur zumeist in drei Phasen gliedern. Grob dargestellt folgte auf eine frühholozäne Klimaverbesserung (10000 - 7500 BP) das holozäne Klimaoptimum (7500 - 5000/3000 BP) und anschließend ein Rückgang bis zu heutigen Verhältnissen. Über das Ausmaß der Vegetations- und Klimaänderungen im mittelholozänen Optimum bestehen recht unter-schiedliche Annahmen, ebenso über Beginn und Ausmaß des menschlichen Einflusses auf die Vegetation. In manchen subtropischen Bereichen Chinas mag die Wiedererwärmung schon bis 7500 BP abgeschlossen gewesen sein.
Aus dem Untersuchungsraum des nördlichen Pakistans scheinen bisher keine pollenana-lytischen Untersuchungen vorzuliegen. Ein vorläufiges Pollendiagramm des Autors wurde von JACOBSEN & SCHICKHOFF (1995) im Zusammenhang mit ihren Datierungen zur Siedlungsgeschichte veröffentlicht (vergl. SCHLÜTZ 1998). Aus dem nördlich an den Karakorum anschließenden Wakhan-Tal im Hindukusch haben BORTENSCHLAGER &
PATZELT (1978) ein Diagramm vorgestellt.
Über die bis zu 4 Mio. Jahre alten Sedimente im Tal von Kaschmir gibt es zahlreiche paläobotanische Arbeiten. Eine Zusammenfassung findet sich bei GUPTA (1991). Kritik an den Ergebnissen älterer Arbeiten übte BHATT (1989). Bezüglich der holozänen Vegetations-geschichte sei auf die Untersuchungen von DODIA et al. (1984) hingewiesen. Anhand von Sedimenten eines Sees in Ladakh macht BHATTACHARYYA (1989) einige Angaben zur Vegetation am Ende des letzten Glazials.
Über die Entwicklungen an der Himalaja-Südseite Indiens seit dem Ausklingen des mittleren Holozäns informieren uns die Artikel von SHARMA & CHAUHAN (1988) und SHARMA (1991). Mit dem jüngsten Einfluß des Menschen in einem Hochtal Nepals befaßt sich der Aufsatz von BEUG & MIEHE (1997, 1999).

3. Stand der Forschung 6
Wichtige Hinweise über den wechselnden Einfluß des Sommermonsuns bieten Unter-suchungen in Indien und dem westlichen Tibet. Bezüglich Indiens sei auf AGRAWAL (1991, 1992), AGRAWAL & GUPTA (1988), BHATIA & SINGH (1988), BRYSON & SWAIN (1981), HEGDE (1977), SINGH et al. (1990), VISHNU-MITTRE (1979) verwiesen. Im westlichen und nordwestlichen Tibet geben die Untersuchungen an Seesedimenten, durchgeführt von AVOUAC et al. (1996), LI YUANFANG et al. (1994) und RHODES et al. (1996), Aufschluß über Veränderungen der Monsunintensität. Besonders zu erwähnen sind ferner die Arbeiten von VAN CAMPO et al. (1996), FAN HUI et al. (1996) und FONTES et al. (1996), die mit unter-schiedlichen Methoden die holozäne Entwicklung des Bangong Co-Beckens betreffen. Eine Synthese ihrer Ergebnisse geben GASSE et al. (1996). Demnach soll auf eine kurzzeitige Zunahme des Sommermonsuns im frühen Holozän eine das Mittelholozän hindurch anhaltende Stärkung erfolgt sein, die sich bis an den Übergang zum jüngeren Holozän hielt. Die im Laufe des Holozäns zunehmende Bedeutung mediterraner Störungen für die Niederschlagsverhältnisse in Vorderasien betonen ZEIST & BOTTEMA (1977, 1988).
Den zahlreichen Arbeiten über den Klimaablauf des Pleistozäns östlich des Tibeta-nischen Plateaus liegen überwiegend Profile aus den teilweise sehr mächtigen Lößabla-gerungen zugrunde (WU ZIRONG & GAO FUQING 1985). Besonders die Paläoböden werden zur Rekonstruktion der Klimaentwicklung herangezogen. Die Resultate werden zumeist mit der Isotopenstratigraphie der Tiefsee korreliert (AN ZHISHENG et al. 1991a, b, c, 1995, BRONGER & HEINKELE 1989, FORMAN 1991, KUKLA & AN ZHISHENG 1989, SUN DONGHUAI
et al. 1996, ZHANG PINGZHONG et al. 1995, ZHANG XIAOYE et al. 1994, ZHENG HONGHAN
1985). Von besonderem Interesse für die Untersuchungen am Profil „Lingfeng“ sind die Arbeit von PACHUR et al. (1994) und der Versuch einer Rekonstruktion des letztglazialen Klimas von ZHANG LANSHENG (1984).
Wohl außer Zweifel steht, daß, über längere Zeiträume betrachtet, Entstehung und Hebung des Tibetanischen Hochlandes sich auf Klima, Flora und Vegetation ausgewirkt haben (LI JIJUN 1991a, b, LI JIJUN et al. 1995, LIN ZHENYAO & WU XIANGDING 1987, WU
XIANGDING & LIN ZHENYAO 1988, ZHENG BENXING 1989). Auf eine damit verbundene Entstehung des Monsuns gehen besonders LIU TUNGSHENG & DING ZHONGLI (1993) ein. Verschiedene Autoren (u. a. ROLPH et al. 1993, WU XIANGDING & LIN ZHENYAO 1988, ZHENG BENXING 1988) sehen eine zunehmende Trockenheit und Kälte Tibets und des nordwestlichen Chinas als Folge dieser Hebung. Die damit einhergehenden geringen Niederschlagsmengen werden als Grund für die relativ begrenzte spätpleistozäne Vergletscherung und das anhaltende Austrocknen der Seen angesehen (ZHENG BENXING
1988, 1989, ZHANG LANSHENG 1988, ZHANG QINGSONG et al. 1994).
HUANG CIXUAN & LIANG YULIAN (1981) und LI BINGYUAN et al. (1985) nehmen sogar für den vergleichsweise kurzen Zeitraum des Holozäns eine enge Beziehung zwischen Hebung und Klimaänderungen an. Wahrscheinlicher erscheint aber die Ansicht anderer Autoren, wonach der Einfluß der geringen Hebungsraten (nach LIU TUNGSHENG et al. 1996: 0,1 - 1 cm/a) auf das Klima des Holozän zu vernachlässigen ist (CUI ZHIJIU et al. 1995, SUN
XIANGJUN & CHEN YINSHUO 1991a, VAN CAMPO & GASSE 1993). FORT (1996) geht sogar von einem gegenwärtigen Absinken des Plateaus aus, während die umgebenden Gebirge ansteigen. Eine räumliche Differenzierung der Bewegungsvorgänge ist für das gesamte Quartär festzustellen (LI JIJUN et al. 1979), entsprechend der unterschiedlichen Entstehungs-

3. Stand der Forschung 7
geschichte der beteiligten tektonischen Einheiten (LI TINGDONG et al. 1986, WANG SHUJI &
SUN YAN 1987, YANG YICHOU et al. 1982).
Für eine Beurteilung zukünftiger Vegetationsänderungen sei auf die Untersuchungen zum Wuchsverhalten der alpinen Matten Tibets unter Erwärmung (ZHANG YANQING &
WELKER 1996) und auf die Beobachtungen zur Sekundärvegetation und Biomasseent-wicklung bei anthropo-zoogenem Einfluß im zentralen Himalaja Indiens hingewiesen (RIKHARI et al. 1992, 1993, RAM 1992).

4. Material und Methoden – Profile und Pollendiagramme 8
4 Mater ial und Methoden
4.1 Profile und Pollendiagramme
Insgesamt lagen etwa 40 Profile mit Kernlängen zwischen 0,1 und 21 m vor. Die Profile wurden erbohrt (Dachnowski-Sonde, Russischer Kammerbohrer) oder an Aufschlüssen entnommen. Nach Voruntersuchungen wurden 22 Profile für eine eingehende Bearbeitung ausgewählt. Der untere Abschnitt der Profile enthält zumeist stark minerogenes Material (Ton- bis Kiesfraktion). Gruslagen finden sich manchmal auch in höheren Profilabschnitten. Vereinzelt herrschen auf ganzer Profillänge schluffig bis sandige Sedimente vor. Auswertbare palynologische Analysen sind dann auf einzelne mehr organogene Lagen beschränkt.
Die Pollendichte war meist gering, die Erhaltung aber fast immer befriedigend oder gut. Hinweise auf eine selektive Zersetzung fanden sich nur selten. Eine Ausnahme bildete das lößartige Material aus dem obersten Bereich des Profils „Lingfeng“ denn wegen des bei Trockenheit ungenügenden Luftabschlusses kommt es im Löß oft zu selektiver Pollenzersetzung. Zudem ist infolge des äolischen Transportes die Herkunft des Pollens nur schwer abzuschätzen. Hinzu kommt die Gefahr bioturbater Vermischung. Die geringen Pollenmengen im Löß lassen sich zwar durch Verarbeitung großer Probenmengen anreichern (KE MANHONG 1994), die zuvor angesprochenen Probleme werden dadurch aber nicht beseitigt. Entsprechend ist bei der Interpretation Zurückhaltung geboten.
4.2 Probenaufbereitung
Die Probenmenge betrug bei Niedermoortorfen etwa 2 cm3, bei Material mit hohem minero-genem Anteil 5 cm3. Durch die Zugabe von Salzsäure wurden Karbonate entfernt und die für Pollenkonzentrationsmessungen zugefügten Tabletten mit Lycopodium-Sporen aufgelöst. Nach einer Behandlung mit kochender 10%iger Kalilauge und dem Absieben der gröberen Fraktion wurde der verbleibende Teil mehrtägig mit kalter Flußsäure (73%) behandelt. Nach der darauf folgende Azetolyse (BEUG 1957) wurden die Proben im Ultraschallbad mit Nylonsieben einer Maschenweite von 5x8 µm gesiebt und anschließend in Glycerin aufgenommen.
Um den Zeitaufwand bei den Auszählungen der oft sehr pollenarmen Proben in einem vertretbaren Rahmen zu halten, konnte auf eine Anreicherung des Pollens durch das Sieben im Ultraschallbad nicht verzichtet werden. Bei vergleichenden Zählungen ungesiebter und gesiebter Proben konnte kein Verlust kleiner und dünnwandiger Pollenkörner (z. B. Juniperus-Typ) nach der Ultraschall-Behandlung festgestellt werden. In einigen Fällen zeigten aber die kleinen, langgestreckten Pollenkörner aus der Familie der Apiaceen einen Rückgang von 5 auf 1 %.
Die mikroskopische Ansprache der Sporomorphen (Pollen, Sporen, Algen) erfolgte überwiegend bei 500facher Vergrößerung. Zur Bestimmung des Gräserpollens nach

4. Material und Methoden – Darstellung der Ergebnisse 9
BEUG (1961) und bei schwierigen Pollenformen wurden ein Immersionsobjektiv und eine Phasenkontrasteinrichtung (Neofluar 100fach, Apertur 1,3) benutzt.
Im Laufe der Untersuchungen wurde begonnen, die Pollenkonzentration zu ermitteln. Die notwendige, aber bei Niedermoortorfen oft fehlerbelastete Volumenbestimmung erfolgte vor der oben beschriebenen Aufbereitung durch Messung des Auftriebs der gefrorenen Proben in Wasser (BARTENS 1990). Mit der Zugabe einer bekannten Menge von Lycopodium-Sporen in Tablettenform konnte anhand der palynologischen Auszählungen die Anzahl der PK im Gesamtvolumen und daraus die Pollenkonzentration (Pollenkörner/cm3) berechnet werden (BENNINGHOFF 1962, STOCKMARR 1971).
Zur Untersuchung des Profils „Lingfeng“ stellte Prof. B. FRENZEL (Hohenheim) die bereits nach dem Schweretrennungsverfahren aufbereitete Proben zur Verfügung; zur Auf-bereitung dieser Proben siehe EISELE et al. (1994). In Göttingen wurden die Mehrzahl dieser Proben zusätzlich im Ultraschallbad gesiebt.
Zur Interpretation der palynologischen Ergebnisse war eine ausreichende Zahl absoluter Altersbestimmungen unerläßlich. Die Datierungen (Radiokohlenstoffmethode) beschränkten sich zunächst auf ausgesuchte Proben in Basisnähe. Nach Vorliegen ausreichender palynologischer Ergebnisse folgten weitere Datierungen. Bei dem verwendeten Material handelte es überwiegend um Torfe oder minerogen-torfiges Sediment. Die physikalischen Messungen wurden in bewährter Weise durch Prof. M. A. GEYH in Hannover durchgeführt. Zum Profil „Lingfeng“ liegen aus Hannover außerdem U/Th-Datierungen sowie von chinesischer Seite Radiokarbondatierungen vor (PACHUR et al. 1994).
Im allgemeinen beschränken sich die Angaben zur Stratigraphie auf das Notwendigste. Für eine sichere Ansprache der Sedimente und als Ergänzung der pollenanalytischen Ergeb-nisse wurden vereinzelt Profilabschnitte unterschiedlicher Länge mit 5%iger Kalilauge aufgeschlossen. Die Untersuchungen der nach Fraktionsgrößen (0,25 mm, 0,8 mm) ausgesiebten Bestandteile erfolgte mit einer Stereolupe.
4.3 Darstellung der Ergebnisse
Den in Form von Prozentdiagrammen wiedergegebenen palynologischen Ergebnissen sind Stratigraphiesäulen und Altersangaben vorangestellt. Die Prozentwerte der Pollenkurven beziehen sich auf die Grundsummen der jeweiligen Proben. Die Grundsumme ergibt sich unter Ausschluß von Cyperaceen, Wasserpflanzen und unbestimmbaren Pollenformen aus der Gesamtzahl der gezählten Pollenkörner (PK). Der Pollen von Bäumen und Sträuchern wird als Gehölzpollen (GhP) zusammengefaßt, der übrige Teil der Grundsumme als Nicht-gehölzpollen (NGhP). In den Profilen vom Nanga Parbat und von Lingfeng wurde der GhP als Grundsumme gewählt.
Zur Darstellung der Diagramme wurden institutseigene Programme benutzt (STEFFAN &
DUPONT 1988). In einigen Fällen erfolgte eine Weiterverarbeitung mit CorelDraw. An die Stratigraphiesäule schließt sich ein Block mit den schwarz hinterlegten Kurven des GhP an. Im zweiten Block sind das Verhältnis vom GhP zum NGhP und die Anteile einiger Gruppen

4. Material und Methoden – Die palynologische Ansprache 10
dargestellt. Auf die Kurven der Cyperaceen und des NGhP folgen Algen (Pediastrum, Botryococcus) sowie Sporen (Farne, Lebermoose). Angaben über die Höhe der Grundsumme und Pollenkonzentrationen stehen am rechten Ende der Diagramme.
Wo der Anhang „-Typ“ verwendet wurde, tritt die betreffende Pollen- oder Sporenform nicht nur in der namengebenden Sippe auf. Das Kürzel „p. p.“ (pro parte) weist auf die Abtrennung von Formen aus dem genannten Taxon hin. So wird z. B. die Bezeichnung Apiaceae p. p. verwendet, da der zu den Apiaceen gehörende Bupleurum-Typ gesondert aufgeführt ist. Wenn systematisch tiefer stehende Taxa gelegentlich nur unvollständig abgetrennt werden konnten oder es sinnvoller erschien, sie rechnerisch im höher stehenden Taxon zu belassen, sind sie als schwarze Balken rechts von der Kurve des höheren Taxon aufgeführt (z. B. Caryophyllaceae, Melandrium-Typ, Cerastium-Typ).
4.4 Die palynologische Ansprache
Viele der gefundenen Sporomorphen sind aus dem Spätpleistozän und Holozän Mittel-europas bekannt. Entsprechend konnte die umfangreiche Vergleichssammlung und Foto-kartei des Instituts für Palynologie und Quartärwissenschaften in Göttingen genutzt werden. Neben mitteleuropäischen Arten enthält diese Vergleichssammlung auch mediterrane, afrikanische und sog. tertiäre Taxa, von denen manche in den untersuchten Profilen auftraten. Die speziell für vegetationsgeschichtliche Untersuchungen in Asien bereits vor Jahren begonnene Erweiterung der Sammlung konnte im Laufe der eigenen Arbeiten um etwa 230 Präparate ergänzt werden.
Die Benennung der palynologischen Taxa orientiert sich an MOORE et al. (1991) sowie BEUG (1961). Ergänzungen und Abweichungen sind im Kapitel 4.4 angegeben. Für eine Ansprache unbekannter Sporomorphen wurden auch der Schlüssel von FAEGRI &
IVERSEN (1989) sowie die Nordwesteuropäische Pollenflora benutzt (PUNT et al. 1976, PUNT
& CLARKE 1980, 1981, 1984, PUNT et al. 1988, PUNT & BLACKMORE 1991). Zudem liegt für den asiatischen Teil der Göttinger Vergleichssammlung ein von Prof. Dr. H.-J. BEUG
ausgearbeiteter unveröffentlichter Bestimmungsschlüssel vor.
Veröffentlichte Pollenfloren, die Teile des Untersuchungsraumes abdecken, liegen in sehr unterschiedlicher Qualität vor. Den nordwestlichen Himalaja betreffen die Arbeiten von GUPTA & SHARMA (1986), NAIR (1965) und RAO & SHUKLA (1975). Umfangreicher sind das gut bebilderte Werk von WANG FUHSIUNG et al. (1995) und die Pollenfloren einer chinesischen AUTORENGRUPPE (1982), von HUANG TSENGCHIENG (1972), YING TSUNSHEN et al. (1993) sowie die japanische Arbeit von NAKAMURA (1980a, b).
Da manche der in Frage kommenden Pflanzensippen Vertreter im afrikanischen Teil des altweltlichen Trockengürtels haben, erwiesen sich auch die reich bebilderten Werke von BONNEFILLE & RIOLLET (1980), EL GHAZALI (1993), REILLE (1992, 1995) und SEKINA &
MOORE (1995) als hilfreich. Für die Ansprache der Sporen von Farnpflanzen standen die umfangreichen Darstellungen von ZHANG YULONG et al. (1990) und HUANG
TSENGCHIENG (1981) sowie die Artikel von NAYAR (1963) und NAYAR & DEVI (1964a, b, c,

4. Material und Methoden – Übersicht der auftretenden Taxa 11
1965, 1966, 1967, 1968a, b) zur Verfügung. Definitionen der Fachtermini finden sich zudem in der Zusammenstellung von PUNT et al. (1994).
4.5 Übersicht der auftretenden Taxa
Im Folgenden sind die meisten der in den Diagrammen aufgeführten Taxa bzw. Pollen-formen zusammengestellt. Eine pollenmorphologische Beschreibung ist dabei weniger beabsichtigt. Kurze Angaben zu den botanischen Vertretern sollen die Diskussion der Dia-gramme (Kap. 6) erleichtern, und der Vegetationskundige mag eine gewisse Vorstellung von den sich hinter den Namen verbergenden pflanzensystematischen Sippen bekommen.
Die Anordnung der Pflanzengruppen orientiert sich an ENGLER (1954, 1964). Bei der Einordnung dort nicht aufgeführter Sippen halfen die Arbeiten von MARTENSEN &
PROBST (1990), POLUNIN & STAINTON (1990), WANG FUHSIUNG et al. (1995) und ZHANG
YULONG et al. (1990). Dank der Übersetzungshilfe durch Herrn Dr. SHI NING (Göttingen)
konnte auch das chinesische Wörterbuch von HOW FOONCHEW et al. (1982) herangezogen werden. Entsprechend der Reihenfolge im Diagramm werden Algen und Sporenpflanzen an den Schluß gestellt.
Familien und höhere Sippen sind durch Unterstreichung, Pollentaxa durch fette Aus-zeichnung hervorgehoben (alphabetische Liste im Anhang). Anstelle der Familienendung „-ceae“ wird im übrigen Teil der Arbeit „-ceen“ geschrieben. Wo sich Angaben auf Ver-gleichspräparate beziehen wird dies durch das Kürzel „VP“ kenntlich gemacht. Wo es notwendig erschien, sind der Sammler (PERSONENNAME) und die Herbarnummer (kursiv) der zugehörigen Pflanzenbelege angegeben.
GYMNOSPERMAE – Nacktsamer
Die in der ganzen nördlichen Hemisphäre verbreitete Familie der Pinaceae ist mit zahl-reichen Arten am Aufbau der Gebirgsnadelwälder Chinas, Tibets und des Himalaja beteiligt. Vielfach fehlt es noch an geeignetem Material für Vergleichspräparate. Soweit möglich wurde nach der Wanddicke und Form der Luftsäcke zwischen PK der Gattungen Picea und Abies unterschieden. Der Formenreichtum im Profil „Lingfeng“ machte eine weitere Aufteilung erforderlich. Diese erfolgte nach Abschluß der eigentlichen Auszählungen anhand der pollenmorphologischen Beobachtungen und Messungen. Zu Picea wurden alle PK mit der von Picea excelsa bekannten Form der Luftsäcke gestellt. Hierunter fallen auch PK mit Wandstärken von über 4 µm. Solche dickwandigen zu Picea gestellten PK wurden vereinzelt auch in Profilen aus dem Nianbaoyeze beobachtet. Beim Picea omorica-Typ sind die Luftsäcke etwas vom Pollenmittelteil abgesetzt. Eine Beschreibung gibt BEUG (1961). Die namengebende Art P. omorica kommt nur in Bosnien und Serbien vor (KRÜSSMANN
1983). Welche der zahlreichen chinesischen Picea-Arten einen vergleichbaren Pollentyp haben, ist unbekannt. PK, die in Wandaufbau, Wanddicke und Form der Luftsäcke zwischen dem Picea omorica-Typ und Abies stehen, wurden als Picea omorica/Abies-Typ bezeichnet. In der Gattung Pinus kann zwischen einem Diploxylon-Typ und dem Haploxylon-Typ

4. Material und Methoden – Übersicht der auftretenden Taxa 12
unterschieden werden. Sie repräsentieren die Arten der jeweiligen Untergattung. Im Dia-gramm „Lingfeng“ wurden beide Typen aufgeführt. In den meisten Diagrammen war die Zahl der bestimmbaren PK sehr gering, so daß auf eine Unterscheidung verzichtet wurde. Vereinzelte durch Wanddicke und relativ große Luftsäcke abweichende PK wurden nicht gesondert ausgewiesen. Nachweise von Larix liegen mit einer Ausnahme nur für das Profil „Lingfeng“ vor. Die pollenmorphologisch vergleichbare Gattung Pseudotsuga kann nach Art und Lage ihrer heutigen Vorkommen ausgeschlossen werden (BEUG 1961, KRÜSSMANN
1983). Die Gattung Cedrus ist in Asien nur durch C. deodara (im westlichen Himalaja) vertreten (MEUSEL & SCHUBERT 1971a). Zur Kennzeichnung des Pollens siehe BEUG (1961). CHEN CUNGEN (1987) gibt für China sieben Tsuga-Arten an. Ihr Pollen trat in den Proben nur sporadisch auf und dürfte in den meisten Fällen durch Windtransport aus weiter entfernten Gebieten stammen. Anfängliche Versuche einer pollenmorphologischen Gliederung wurden nicht weiterverfolgt.
Viele Cupressaceae und manche Taxodiaceae besitzen den gleichen einfach gebauten Pollentyp wie die im Untersuchungsraum wichtige Gattung Juniperus (BERTSCH 1961). Im Diagramm „Lhasa Stadtbereich“ und in den Profilen aus dem südöstlichen Tibet wurde diesem Umstand durch die Bezeichnung Juniperus-Typ Rechnung getragen, da Cupressus
torulosa sowie Thuja orientalis in Lhasa gepflanzt werden (WARDLE 1981) und weil Cupressus gigantea im Yarlung Zangbo-Tal natürliche Vorkommen hat (CHEN WEILIE
1981). In den übrigen Fällen dürften die Pollenfunde vom Juniperus-Typ fast ausschließlich von Vertretern der Gattung Juniperus stammen, deren Sektion Sabina in der chinesischen Literatur meist als Gattung geführt wird, zu der aber auch manchmal Arten der Sektion Oxycedrus gestellt werden. So sehr sich die Wacholderarten in Ökologie und Wuchsform unterscheiden und eine pollenmorphologische Gliederung daher wünschenswert wäre, so ist ihr Pollen doch sehr einheitlich. Eine einfache Untergliederung nach der Wanddicke oder der Anordnung und Häufigkeit der Skulpturelemente scheint nicht möglich zu sein (BORTENSCHLAGER 1990, JOCHIMSEN 1986). Die Größe der Skulpturelemente liegt unter 1 µm (BORTENSCHLAGER 1990, DUHOUX 1982), ihrer Form entsprechend sind sie als Microgemmae oder besser als Microclavae anzusprechen (BERTSCH 1961, BEUG 1961). Bei den eigenen Untersuchungen erschienen manche dieser Skulpturelemente „hohl“ (vergl. dazu die elektronenmikroskopische Aufnahme des Mikrotomschnittes eines PK von Sequoia sempervirens bei GULLVÅG 1966, Tafel 14 a). Dies war bei 1250facher Vergrößerung im einfachem Durchlicht und auch im Phasenkontrast zu erkennen. Dunkle „Flecken“ bei der LO-Analyse (NILSSON & PRAGLOWSKI 1992) deuteten bei sonst solide erscheinenden Elementen auf sehr kleine „Hohlräume“ hin. Vereinzelt wurden auch Skulpturelemente von mehr als 1 µm Durchmesser gefunden. Diese waren in der Regel hohl und nicht selten von ungleichmäßiger Gestalt. Es bleibt zu prüfen, ob sich die angesprochenen Merkmale für eine Differenzierung eignen.

4. Material und Methoden – Übersicht der auftretenden Taxa 13
Die Gattung Ephedra (Ephedraceae) ist mit zahlreichen ihrer insgesamt ca. 40 Arten in den Trockengebieten Innerasiens vertreten. Eine Auswahl der zum Ephedra distachya-Typ und Ephedra fragilis-Typ gehörenden Arten geben BEUG (1961) und LANG (1994).
ANGIOSPERMAE – Bedecktsamer
Die Mitglieder der Juglandaceae sind überwiegend streng an Windbestäubung angepaßte Bäume. Pollen der Gattung Juglans unterscheidet sich u. a. durch die heteropolar verteilten Poren von dem der Gattung Pterocarya. Doch kommt bei einem Teil der PK in beiden Sippen die Porenanordnung der jeweils anderen Gattung vor. Bei den Juglans-Funden handelt es sich ziemlich sicher um PK von J. regia, die sich durch ihre relativ hohe Zahl extraäquatorialer Poren vom Pollen der in China verbreiteten Sekt. Cardiocaryon DODE
(J. cathayensis u. a., siehe SCHAARSCHMIDT 1988) abgrenzen lassen (VP, BOS & PUNT 1991, HUANG TSENGCHIENG 1972, NAKAMURA 1980a, b, STONE & BROOME 1975, WANG
FUHSIUNG et al. 1995). Juglans und Pterocarya kommen besonders in sommergrünen Laub- und Auwäldern auf frischen, nährstoffreichen Böden vor und ertragen kurz anhaltende Über-schwemmungen (SCHAARSCHMIDT 1988). Im Karakorum wird J. regia als Fruchtbaum angebaut und ist auch als Bauholz sehr geschätzt (HUGHES 1984, PAFFEN et al. 1956, SCHWEINFURTH 1957). Bei Lhasa wird J. cathayensis genutzt (WARDLE 1981).
Weiden (Salicaceae) spielen in der alpinen und subalpinen Vegetation in Tibet und im Himalaja eine wichtige Rolle. Neben der für Salix üblichen reticulaten Pollenform traten im Material vom Nianbaoyeze auch PK vom Salix oritrepha-Typ auf. Bei diesen ist das Reticulum durch Verrucae teilweise bis fast vollständig maskiert. Ein entsprechendes VP von S. oritrepha liegt vor. Unter den 22 untersuchten Salix-Arten aus Gansu fand WANG
JINGQUAN (1985) eine solche Skulptur sowohl bei S. oritrepha als auch bei den tricol-poroidaten PK von S. microtricha und S. sclerophylla. Nach den Abbildungen bei KEZINA (1988) scheint auch ein Vertreter der manchmal von Salix abgetrennten, wind-bestäubten nordostasiatischen Gattung Chosenia hierher zu gehören (HEYWOOD 1982, FUKAREK 1994). Pollen von Populus war nur im Profil „Lhasa Stadtbereich“ vorhanden.
Die Betulaceae stellen in der Gattung Betula mit B. utilis einen der wichtigsten Laub-holzvertreter in den Hochlagen im Himalaja und in Tibet. Man findet B. utilis sowohl als Baum in den hochmontanen Abies-Wäldern Nepals und an der Ostabdachung des Tibe-tanischen Plateaus als auch als Krummholz an der Waldgrenze des Nanga Parbat (MIEHE
1991, TROLL 1939, WINKLER 1994). Weitere häufige Arten sind u. a. B. platyphylla, B. platyphylla var. szechuanica, B. albo-sinensis, B. alnoides und B. cylindricum. (CHEN
CUNGEN 1987, CHEN GUICHEN et al. 1994, LI WENHUA et al. 1981, SCHMIDT-VOGT 1987, POLUNIN & STAINTON 1990, THELAUS 1992). Nach den wenigen verfügbaren Messungen haben die PK von B. utilis, B. platyphylla und B. albo-sinensis Durchmesser von über 30 µm. Die PK von B. platyphylla var. japonica, B. platyphylla var. mandschurica und B. alnoides sind mit Größen von durchschnittlich 23 - 26 µm etwas kleiner (frdl. persönliche Mitteilung Prof. Dr. H.-J. BEUG; NAKAMURA 1980a, WANG FUHSIUNG et al. 1995). Einen Bericht über die Probleme bei der Artansprache von Birkenpollen gibt MÄKELÄ (1996). Eigene

4. Material und Methoden – Übersicht der auftretenden Taxa 14
Größenmessungen wurden am „Profil 3“ aus dem Nianbaoyeze vorgenommen (vergl. Kap. 6.3.1.3) Nachweise von Alnus und Corylus blieben selten. Die wenigen PK von Carpinus waren triporat und entsprachen denen von C. orientalis. Benannt wurden sie nach der im Un-tersuchungsraum vorkommenden Art als Carpinus cordata-Typ.
Die Fagaceae sind in allen Profilen durch Pollen von Quercus repräsentiert, der wohl immer aus dem Regional- oder Fernflug stammt. Die Einzelfunde von Fagus im Profil „Lingfeng“ entsprachen etwa unserer Rotbuche und könnten damit, außer von F. longi-
petiolata, von jeder der übrigen 10 Buchenarten Chinas stammen (HONG BIGONG & AN
SHUQING 1993, PRAGLOWSKI 1982).
Pollenfunde aus den Gattungen der Unterfamilie Ulmoideae (Ulmaceae) wurden als Ulmus-Typ zusammengefaßt. Vertreten sind die Gattungen Ulmus, Zelkova und Hemiptelea. Pentaporate PK zur Gattung Ulmus und tetraporate PK zu Zelkova zu stellen, wäre falsch. Sind doch z. B. bei U. parvifolia, die bei Lhasa steht, und bei U. pumila, dem oft einzigen Gehölz wassernaher Standorte in den Wüstengebieten Zentralasiens, 90 % bzw. 35 % des Pollen tetraporat. Bäume der Gattung Zelkova sind ein Element der Bergwälder Nordchinas und Japans. Z. serrata und Z. sinica besitzen z. B. tetra- und pentaporate PK. Bei Untersuchungen an eigenem Material der auf der Mittelmeerinsel Kreta endemischen Z. abelicea (JAHN & SCHÖNFELDER 1995) waren 65 % der PK pentaporat und etwa 3 % hexa-porat. NAKAGAWA et al. 1998 beschreiben die PK von Z. abelicea hingegen als überwiegend pentaporat und teilweise tetraporat und Z. sicula (SO-Sizilien) als pentaporat mit auch hexa-poraten PK. Bei der in Nord- und Nordostchina bis Korea beheimateten Hemiptelea davidii überwiegen hexa- über heptaporate PK (FUKAREK 1993, NAKAMURA 1980a, WANG
FUHSIUNG et al. 1995, WARDLE 1981, XIN YIQUN 1993, ZAVADA 1983).
Bei den wenigen Funden des Cannabis-Typs (Cannabaceae) wird es sich wohl kaum um solche des auf feucht-schattige Standorte angewiesenen Hopfens – in Ostasien Humulus
scandens (H. japonicus) – sondern um PK von Hanfpflanzen handeln (ENGLER 1964, NAKAMURA 1980b, WANG FUHSIUNG et al. 1995).
Die in den Profilen „Shukan 2“ und „Lhasa Stadtbereich“ als Urtica (Urticaceae) bestimmten PK entsprechen dem Urtica dioica-Typ, zu dem in NW-Europa auch U. urens gehört (PUNT & MALOTAUX 1984).
Der Einzelfund von Thesium (Santalaceae) im Profil „Shukan 2“ ist in sofern interes-sant, als die Vertreter der Gattung Thesium hier Zeiger für stark übernutzte Weiden sind (frdl. persönliche Mitteilung Prof. Dr. G. MIEHE).
Die Halbschmarotzer aus der Gattung Arceuthobium (Loranthaceae) befallen verschiedene Nadelbäume. Vom Mittelmeerraum bis in den westlichen Himalaja findet man die Wacholdermistel A. oxycedri auf Juniperus-Arten. Die im Himalaja auf Pinus wallichiana schmarotzende A. minutissimum ist wohl eine der kleinsten dikotylen Pflanzen. Äußerlich ist von ihr nur die winzige Blütenknospe sichtbar (ENGLER 1964, FUKAREK 1993, SCHÜTT et al. 1992). Nach den Beobachtungen von HANDEL-MAZZETTI
(1936) in Südwest-China besiedelt A. chinense neben Pinus und Abies besonders die Gattung

4. Material und Methoden – Übersicht der auftretenden Taxa 15
Keteleeria, deren stark befallene Jungpflanzen absterben. Vertreter der Gattung Juniperus sind nach seinen Angaben frei von Arceuthobium. Selbst über die Verbreitung von A. oxycedri in Europa ist zu wenig bekannt, um eine Arealkarte zeichnen zu können (FASSL
1996). Nach Norden bleibt sie weit hinter der Verbreitung von Juniperus communis zurück, befällt diese Art aber im mediterranen Raum. Wie bei anderen Misteln (Viscum, Loranthus) scheinen also auch in der Gattung Arceuthobium die klimatischen Ansprüche enger als die der Wirte zu sein (TUBEUF 1923), womit Pollenfunden ein entsprechender Zeigerwert zukommt. Die PK sind hexacolpat und echinat. Lange, schmale Colpi stehen abwechselnd mit kurzen, breiten Colpi. Abbildungen von A. chinense finden sich bei der AUTORENGRUPPE (1982), von A. oxycedri in REILLE (1992). Pollen von Arceuthobium kann auch in Deutschland auftreten (SCHLÜTZ 1997).
Die Familie der Polygonaceae ist pollenmorphologisch reichhaltig und zudem häufig vertreten. Rheum, der Rhababer, hat sein Verbreitungszentrum in den Gebirgen Zentral-asiens und Chinas. Die PK wurden nach vorliegenden VP unter Berücksichtigung der für Rheum officinalis von LEEUWEN et al. (1988) beschriebenen Form der Endoapertur bestimmt. Der Rumex-Typ enthält die Gattungen Rumex (Ampfer) und Oxyria (Säuerling). Davon abgetrennt wurde der Eurumex-Typ, dem bei LEEUWEN et al. (1988) etwa R. crispus, die R. sanguineus- und R. conglomeratus-Gruppe entsprechen; hierher gehört auch R. nepalensis (VP). Die Vertreter dieses Typs siedeln oft an Gräben, in Ufergesellschaften und Lägerfluren.
Sektionen der Gattung Polygonum s. l. (Knöterich) werden verschiedentlich als eigene Gattungen eingestuft (HARALDSON 1978, HEDBERG 1946, HONG SUKPYO 1991). Der häufige Polygonum bistorta-Typ vertritt die Sektion Bistorta. Hierher gehören der im Karakorum durch seine Zwergspalierstrauch-Matten stellenweise aspektbildende P. affine (LLOYD &
LLOYD 1968, PAFFEN et al. 1956) sowie P. amplexicaule (VP, HEDBERG 1946), die oft aus Tibet erwähnten Arten P. viviparum und P. macrophyllum, ebenso wie der aus dem Nianbao-yeze und Qinling-Gebirge belegte P. sphaerostachyum (VP) (BJÖRK 1993, CHANG D. H. S. 1981, HUANG RONGFU 1987, ZHENG DU & YANG QINYE 1985). Die PK der bis in 4300 m vorkommenden Art P. vacciniifolium sind überwiegend tricolpat mit auch außerhalb der Polregion verzweigten Stäbchen (VP), vermitteln somit zum Polygonum alpinum-Typ (POLUNIN & STAINTON 1990, HARALDSON 1978). Die von GUPTA & SHARMA (1986) unter P. vacciniifolium abgebildeten PK zeigen vermutlich P. plebeium. Zum Polygonum
aviculare-Typ zählt der bis etwa 2000 m Höhe vorkommende Polygonum plebeium. P. aviculare (Vogelknöterich) selbst erreicht im Nordwesthimalaja Höhen von 4000 m. P. paronychioides (VP, HEDBERG 1946) wird für die Strauchsteppen am Nanga Parbat angegeben (SCHWEINFURTH 1957). Der Polygonum aviculare-Typ trat besonders in den Profilen aus dem Karakorum und von Lhasa auf. Polygonum sibiricum (HONG SUKPYO 1991: Knorringia sibirica) läßt sich nach Skulptur und Form deutlich von den übrigen tri-colporaten PK der Polygonaceae abtrennen. Das Material für das VP verdanke ich Herrn Dr. DICKORÉ, Göttingen. Es stammt von südlich des Yarlung Zangbo aus 5060 m Höhe (Nagarze-Lhozak, Nr. 9875). Im Talboden bei Lhasa bevorzugt P. sibiricum nasse Plätze,

4. Material und Methoden – Übersicht der auftretenden Taxa 16
nach HILBIG (1995) ist die Art in der Mongolei Teil der Salzvegetation.. Für den Nordwest-himalaja gibt MANI (1978) die Art aus Höhen zwischen 3300 - 5000 m an. Der Polygonum
alpinum-Typ (tricolpat, dickwandig) kennzeichnet die Sektion Aconogonon, ausgenommen die Arten P. campanulatum und P. pangiantum (HONG SUKPYO 1992).
Pollen aus der Sektion Persicaria ist eureticulat. Nach den Aperturen wurde zwischen dem pericolpaten Polygonum amphibium-Typ und dem periporaten P. persicaria-Typ unterschieden. Polygonum amphibium tritt im Himalaja bis in 4000 m auf (GUPTA &
SHARMA 1986). Zum selben Pollentyp wie P. persicaria gehören im Nordwesthimalaja P. hydropiper, P. lapathifolium und P. orientale (GUPTA & SHARMA 1986, HEDBERG 1946, LEEUWEN et al. 1988). GUPTA & SHARMA geben auch P. chinense als periporat an, doch sind die PK dieser Art nach HEDBERG (1946) und HONG SUKPYO & HEDBERG (1990) tricolpat. Das Auftreten des Polygonum amphibium- und P. persicaria-Typs zeigt im allgemeinen feucht-nasse Standortverhältnisse an. Im Profil „Lhasa Stadtbereich“ waren beide Typen nicht selten, in „Shukan 2“ wurde der P. persicaria-Typ gefunden. Neben diesen periaperturaten PK treten auch tricolpate, eureticulate Formen auf. Nach dem vorliegenden VP wurden PK ohne Columellae in den Lumina zu Polygonum glaciale gestellt (Nianbao-yeze).
Unter dem Namen Koenigia wurden alle auftretenden periaperturat-echinaten PK der Gattung zusammengefaßt. Es traten wie bei K. nepalensis sowohl deutlich periporate PK als auch solche mit schwach langgestreckten Aperturen auf. Letztere findet man auch bei K.
filicaulis. Formen mit langen Colpi (K. forrestii) und hexapericolpate PK (P. campanulatum) wurden nicht gefunden (VP, HONG SUKPYO 1992, HONG SUKPYO & HEDBERG 1990).
Besonders häufig traten im Material aus dem Nianbaoyeze PK auf, die wahrscheinlich zur Familie der Polygonaceae zu stellen sind, und daher in einer früheren Arbeit als cf. Poly-gonaceae bezeichnet wurden (SCHLÜTZ 1995). Da bislang weder bestätigende VP beschafft werden konnten, noch diese Annahme stützende Abbildungen in der Literatur bekannt sind, erfolgt bis auf weiteres die Bezeichnung Typ N1.
Polsterförmige Caryophyllaceae kommen bis an die Obergrenze des Pflanzenwuchses vor. Die Arten der heute oft zu Silene gestellten Gattung Melandrium produzieren deutlich reticulaten Pollen vom Melandrium-Typ (z. B. Silene dioica-Typ und S. latifolia ssp. alba bei MOORE et al. 1991). BJÖRK (1993) und HUANG RONGFU (1987) fanden Vertreter von Melandrium in den Kobresia-Matten Tibets. Der Cerastium-Typ tritt außer in der Gattung Cerastium u. a. auch bei Stellaria auf (MOORE et al. 1991) und ist in den Profilen „Batyatum Jut“ und „Alambar“ aus dem Karakorum nicht selten.
Die Familie der Chenopodiaceae ist in den salzreichen Steppen von Zentral- und Ostasien mit zahlreichen Arten vertreten und gebietsweise vegetationsbestimmend (ENGLER 1964). Eine pollenanalytische Artbestimmung scheint aussichtslos (FRENZEL 1993). Zudem sieht der Pollen einiger Arten der Amaranthaceae ähnlich aus (WANG FUHSIUNG et al. 1995). PK mit einer deutlich unterdurchschnittlichen Anzahl von Poren erscheinen im optischen Schnitt polygonal und wurden entsprechend einem VP als Haloxylon-Typ

4. Material und Methoden – Übersicht der auftretenden Taxa 17
bezeichnet (Foto 1 und 2, im Anhang). Derartige PK sind aber wahrscheinlich weit ver-breitet, wie die Abbildungen des Aellenia- und Noaea-Typs bei ZEIST & BOTTEMA (1977) zeigen.
In der Familie Ranunculaceae konnten 5 Pollenformen unterschieden werden. Der Anemone/Ranunculus-Typ enthält im wesentlichen Vertreter der namengebenden Sippen sowie daneben die Gattungen Clematis und Pulsatilla. Einige Arten aus der Untergattung Batrachium lassen sich von den übrigen Ranunculus-Arten pollenanalytisch als Batrachium-Typ unterscheiden. Sie sind in stehenden oder langsam fließenden Gewässern verbreitet (GARCKE 1972). Beim Caltha-Typ ist nach den Angaben von CLARKE et al. (1991) neben der Gattung Caltha auch an Aquilegia und Isopyrum zu denken. Neben den bekannten schwach striaten PK der Gattung Trollius wurden in erheblichem Umfang deutlich striate PK gefunden, wie sie das VP eines Trollius-Beleges aus Xizang (DICKORÉ 9224) zeigt. Die kleine bis mittelhohe Stauden bildenden Arten der Gattung Thalictrum verhalten sich ökologisch recht unterschiedlich und werden teils durch Insekten, teils über den Wind bestäubt. Palynologisch sind sie wohl nicht zu unterscheiden (ENGLER 1964, FUKAREK 1993, FRENZEL 1993). Nach den Untersuchungen von XI YIZHEN (1993) an 62 Aconitum-Arten ist ihr Pollen sehr gleichförmig und eine Artansprache unmöglich. Die Gattung kommt bis in die nivale Stufe vor (HUANG RONGFU 1987). Es ist nicht auszuschließen, daß bei den routinemäßigen Zählungen auch z. T. die sehr ähnlichen PK von Consolida, Delphinium oder Adonis (CLARKE et al. 1991) zu Aconitum gestellt wurden.
Die Nymphaeaceae sind eine Familie von Wasser- und Sumpfpflanzen. Die im Profil „Lhasa Stadtbereich“ gefundenen PK der Gattung Nymphaea wichen in ihrer Skulptur stark vom Nymphaea alba-Typ (JONES & CLARKE 1981) ab. Nach einem vorliegendem VP und den Angaben in der Literatur handelt es sich um PK Nymphaea tetragona (AUTORENGRUPPE
1982, BEUG 1961, NAKAMURA 1980b). Die in Indien, dem Himalaja und in China südlich des Yangzi verbreitete N. stellata gehört nicht hierher (FUKAREK 1993, GUPTA & SHARMA
1986, WANG FUHSIUNG et al. 1995).
Die große Familie der Brassicaceae besitzt in Zentralasien eines ihrer Mannigfaltig-keitszentren. Viele ihrer meist einjährigen Arten kommen in den Hochgebirgen bis an die Vegetationsgrenze vor. Sie besiedeln aber auch die durch Menschen und Tiere gestörten Flächen sowie trockene und salzige Böden (BJÖRK 1993, LERCH 1991, MARTENSEN &
PROBST 1990, SCHWEINFURTH 1957).
Die großen, zur Familie der Crassulaceae gehörenden Gattungen Sedum und Rhodiola
sind im Himalaja und in Ostasien mit vielen Arten vertreten (ENGLER 1964, FUKAREK 1993). Häufig sind sie in den krautreichen Horstgras-Steppen des Karakorum und in der nivalen Stufe Tibets vorzufinden (HUANG RONGFU 1987, LI BOSHENG 1981, MIEHE 1988, MIEHE et al. 1996)
Die Saxifragaceae sind durch die Gattung Saxifraga vertreten. Sie kommt sowohl in der subnivalen Stufe als auch im Unterwuchs von Gebüschen, an sumpfigen Stellen und an Felsen vor (CHANG D. H. S. 1981, HUANG RONGFU 1987, MIEHE 1988). Es wurden der

4. Material und Methoden – Übersicht der auftretenden Taxa 18
Saxifraga stellaris-, Saxifraga oppositifolia- und der Saxifraga granulata-Typ unter-schieden. Zur Pollenmorphologie und zu Artangaben siehe MOORE et al. (1991) und VERBEEK-REUVERS (1980).
Der Pollen der Rosaceae spielt in dem Untersuchungsmaterial eine nicht geringe Rolle. Zum Potentilla-Typ gehören neben Potentilla auch Gattungen wie Fragaria, Sibbaldia und z. T. auch Rosa (EIDE 1981, EIKELAND 1979, REITSMA 1966). Wegen der großen Bedeutung von Potentilla (Dasiphora) fruticosa in den Gebüschformationen wird ihr Typ zum Gehölzpollen (GhP) gerechnet. P. anserina ist in Tibet in großer Zahl auf gestörten Flächen wie z. B. Schaflagerplätzen anzutreffen (BJÖRK 1993, HERMANNS 1949). In den Kobresia-Matten treten häufig P. stenophylla, P. nivea und P. multifida auf (CHANG D. H. S. 1981, HUANG RONGFU 1987). Sträucher von Sibiraea (-Typ) kommen gemeinsam mit P. fruticosa vor oder bilden mit Hippophaë Galeriegehölze (BJÖRK 1993, HUANG RONGFU 1987, MIEHE
1991, ZHANG JINGWEI et al. 1988, ZHENG DU & YANG QINYE 1985). Als Rosaceae p. p. wurden die übrigen tricolporaten Pollenformen dieser Familie zusammengefaßt. Hierzu zählen u. a. die Gattungen Sibbaldia, Neillia, Sorbus und Cotoneaster sowie Arten aus der Gattung Rosa.
In der Gattung Sanguisorba produzieren Pflanzen der insektenbestäubten Art S. offi-
cinalis (Großer Wiesenknopf) mit einer Chromosomenzahl von 2n = 28 PK, die oft als hexazonocolporat bezeichnet werden. Pflanzen mit 2n = 56 zeigen hingegen trizonocol-porate PK. Die Aperturen haben ein Operculum fast von der Breite der Mesocolpia, so daß die Ränder der 3 Colpi weit auseinander liegen und sechs Colpi vortäuschen. Dieses Phänomen ist viel diskutiert (EIDE 1981, ERDTMAN & NORDBORG 1961a, FAEGRI & IVERSEN
1989, REITSMA 1966, MOORE et al. 1991). Im „Profil 14“ vom Nianbaoyeze, im Material von Malinganko und im Profil „Lingfeng“ traten PK wie von 2n = 56 Pflanzen in großer Zahl auf. Während sich bei diesen die Enden der Colpenränder nur etwas annähern (Pontoperculum sensu ERDTMAN 1952), waren in denselben Proben immer auch PK vorhanden, bei denen die Colpenränder sich treffen und vereint polwärts laufen (Foto 3 und 4, im Anhang). Beide Formen treten bei der Art Sanguisorba filiformis auf (VP-Material von DICKORÉ), die an sumpfigen Stellen sehr häufig sein kann (BJÖRK 1993, HANDEL-MAZZETTI 1936, THELAUS 1992).
Viele Sippen der Fabaceae haben ihre Hauptentfaltung in den gemäßigten Breiten, besonders in Gebieten mit ausgedehnten Trockenzeiten. Für Xizang geben LI PEIQIONG & NI
ZHICHENG (1981) 41 Gattungen mit 254 Arten als wild vorkommend und weitere 11 Gattungen mit 16 Arten als kultiviert an. Die mit jeweils etwa 300 Arten kaum über-schaubaren Gattungen Astragalus und Oxytropis haben ihren Verbreitungsschwerpunkt in den innerasiatischen Trockengebieten, wo sie Bestandteil der landschaftsbestimmenden Igeldornpolster-Formationen sind. Der größte Teil des unter Fabaceae p. p. summierten Pollens dürfte aus diesen Sippen stammen. Zum Hedysarum-Typ gehören nach MOORE et al. (1991) die Gattungen Hedysarum und Onobrychis. Vertreter von Hedysarum kommen sowohl in Salix oritrepha-Gebüschen als auch an steilen Erosionsrinnen vor (HUANG
RONGFU 1987, TROLL 1967). Anhand weniger VP aus den großen Gattungen Caragana und Thermopsis wurde ein Caragana-Typ aufgestellt.

4. Material und Methoden – Übersicht der auftretenden Taxa 19
Von der Familie der Geraniaceae konnten die Gattungen Geranium und Erodium nachgewiesen werden. Sie sind pollenmorphologisch leicht trennbar (STAFFORD & BLACK-MORE 1991). Übergänge von Erodium zu Geranium, von denen BORTENSCHLAGER (1967) berichtet, wurden nicht beobachtet.
Zur Familie der Zygophyllaceae gehören viele Xerophyten und Halophyten, die Charak-terpflanzen der Wüsten und Salzsteppen sind. Bei den 25 Arten der Gattung Tribulus handelt es sich meist um einjährige, oftmals niederliegende Kräuter. Ihre Früchte besitzen oft starke Stacheln und werden als Trampelkletten von Tieren verschleppt (FUKAREK 1993, KINZEL 1982). Eine pollenmorphologische Artdiagnose scheidet wohl aus (BONNEFILLE &
RIOLLET 1980, EL GHAZALI 1993, PRAGLOWSKI 1987, REILLE 1992, 1995, YUNUS &
NAIR 1989). Foto 5 (im Anhang) zeigt ein PK aus dem Profil von Lhasa. Vertreter der artenarmen Gattung Nitraria trifft man in Salzwüsten an. Es sind kleine, oft dornige Sträucher. PolIen von Nitraria fanden KONG ZHAOCHEN et al. (1990) im Qinghai-See (Kuku Nur) und SHILOVA (1984) in Oberflächenproben aus der Mongolei. Bei den eigenen Unter-suchungen waren die tricolpaten, striaten, durch Porenform und dicke Endexine kenntlichen PK für die Proben aus dem Gaxun Nur typisch (Abb. bei KONG ZHAOCHEN & DU NAIQIU
1981, REILLE 1995).
Die Familie der Thymelaeaceae ist mit den Gattungen Thymelaea (incl. Stellera) und Daphne in Eurasien weit verbreitet. Pollen vom Thymelaea-Typ ist Arten beider Gattungen eigen. Die kleinen Büsche von Daphne kommen ähnlich Pistacia in Trockentälern vor. In der alpinen Stufe bilden einige Arten dichte Bestände an Steilhängen und offenen Flußbänken (HUANG RONGFU 1987, SCHWEINFURTH 1957, TROLL 1939).
Die Pollenfunde der Familie der Onagraceae dürften von der Gattung Epilobium (incl. Chamaenerion) stammen. An feuchten Standorten des nordöstlichen Tibet wächst z. B. E. palustre (HUANG RONGFU 1987).
Die Gattung Myriophyllum (Tausendblatt) stellt eine fast kosmopolitisch verbreitete Gruppe submerser Wasserpflanzen stehender oder langsam fließender Gewässer aus der Familie der Haloragaceae (FUKAREK 1993). Neben dem in den Profilen „Lhasa Stadtbereich“ und „Lingfeng“ recht häufigen Myriophyllum spicatum-Typ wurde vereinzelt auch der Myriophyllum verticillatum-Typ gefunden (ENGEL 1980a, PRAGLOWSKI 1970). M. spicatum wird von BJÖRK (1993) aus dem Becken von Zoige (Osttibet) erwähnt. Vereinzelt traten auch PK des Myriophyllum alternifolium-Typs auf.
Hippuris vulgaris, der einzige Vertreter der Hippuridaceae, ist auf der Nordhalbkugel von den gemäßigten Breiten bis weit in die Arktis hinein verbreitet. Wie Myriophyllum ist Hippuris eine Pflanze meist seichter, stehender Gewässer. In Tibet gedeiht Hippuris auch als Sumpfpflanze (BJÖRK 1993, FUKAREK 1993, ZHANG JINGWEI et al. 1988). ZHAO KUIYI et al. (1981) gibt ihn für Xizang aus Altarmen an. Zur Beschreibung des Pollens siehe ENGEL (1980b).

4. Material und Methoden – Übersicht der auftretenden Taxa 20
Beim derzeitigen Wissensstand konnte von den Apiaceae p. p. oder Umbelliferae nur ein weit gefaßter Bupleurum-Typ mit den Gattungen Bupleurum, Pleurospermum, Trachy-
dium, Astoma, Physospermum und Smyrnium abgetrennt werden (ZEIST & BOTTEMA 1977, PUNT 1984). Zumindest Bupleurum, Pleurospermum und Trachydium sind aus dem Himalaja und Tibet bekannt (RIKHARI et al. 1993, MANI 1978, SHAN RENHUA & SHE MENGLAN 1981). Während unsere einheimischen Arten des Hasenohrs (Bupleurum) meist einjährige Kräuter oder Stauden sind, findet man in trockenen Gebieten auch Halbsträucher und Dornpolster-pflanzen in dieser Gattung (ENGLER 1964, KLÖTZLI 1993). Von sumpfigen Flächen des Zoige-Beckens im östlichen Tibet geben BJÖRK (1993) und THELAUS (1992) die Art Chamaesium paradoxum an.
Die Gattung Rhododendron stellt die größte Sippe in der Familie der Ericaceae dar. Ihr Auftreten reicht von Bäumen in der Nebelwaldstufe bis hin zu Sträuchern der alpinen Stufe. Aus Xizang kennt man etwa 170 Arten. Ihren Verbreitungsschwerpunkt hat die Gattung im Bereich der meridionalen Stromfurchen (FANG RUIZHENG & MING TIANLU 1981, LI
WENHUA 1993, MIEHE 1991). Ihr Pollen ist im unteren Teil des Diagramms „Lingfeng“ recht häufig, in den Profilen vom Nanga Parbat und aus dem Nianbaoyeze nur spärlich vertreten.
Ein großer Teil der fast 500 Primula-Arten (Primulaceae) hat seine Heimat in der alpinen Vegetation der asiatischen Hochgebirge. In Xizang wachsen etwa 110 Arten (CHEN
FENGHUAI & HU QIMING 1981, FUKAREK 1994). Die wenigen Pollenfunde wurden zum Primula farinosa-Typ oder zum Primula veris-Typ gestellt (MOORE et al. 1991). Auch Androsace bevorzugt hohe Lagen. Die oftmals in Felsspalten verankerten Polster sind ein wichtiger Bestandteil in der alpinen und subnivalen Vegetation (CHANG D. H. S. 1981, LI
BOSHENG et al. 1985, MIEHE 1988, 1991, QIN ZHIYE & XIE WENZHONG 1980).
Kräuter, Halbsträucher und kleine Sträucher aus der Familie der Plumbaginaceae kommen besonders in den Trockengebieten Vorder- und Zentralasiens sowie des Mittel-meergebietes vor. Ihre Unterfamilie Plumbaginoideae ist durch Pollen vom Plumbago-Typ (ERDTMAN 1952) gekennzeichnet (FUKAREK 1993, MARTENSEN & PROBST 1990). Dieser Pollen-Typ tritt im Profil „Lhasa Stadtbereich“ auf und dürfte von Ceratostigma stammen. Der Pollen der sechs chinesischen Arten dürfte wie die vorliegenden fossilen PK colpat sein; manche Präparate von C. willmottianum zeigen auch pericolpate PK (NILSSON &
PRAGLOWSKI 1992, NOWICKE & SKVARLA 1977, WU ZHENGYI et al. 1981). Zur Unterscheidung vom Linum austriacum-Typ (PUNT & BREEJEN 1981) vergl. TURNER &
BLACKMORE (1984). Typisch für die Unterfamilie der Limonioideae ist der Armeria-Typ (ERDTMAN 1952). In den Profilen aus dem Karakorum wurden sowohl die feinreticulate als auch die grobreticulate Form gefunden. Als Pollenspender kommen hier wohl nur die Dornpolster bildenden Arten der Gattung Acantholimon in Frage (VP, KLÖTZLI
1993, MIEHE et al. 1996, NOWICKE & SKVARLA 1977).
Die Familie der Oleaceae ist in den Diagrammen aus dem Karakorum und vom Nanga Parbat durch die Gattung Fraxinus (Esche) vertreten. Zum Fraxinus excelsior-Typ zählt neben der bis in den Nordwesthimalaja vorkommenden Gemeinen Esche F. excelsior

4. Material und Methoden – Übersicht der auftretenden Taxa 21
(GUPTA & SHARMA 1986, NAIR 1965) auch F. angustifolia. Der Fraxinus floribunda-Typ wurde anhand eines VP von Herbarmaterial aus China aufgestellt und kommt dem F. ornus-
Typ sehr nahe (BEUG 1964, PUNT et al. 1991). In der trockenen Gehölzvegetation Afghanistans und des Himalaja tritt häufig F. xanthoxyloides auf (MEHER-HOMJI 1973, SCHWEINFURTH 1957, POLUNIN & STAINTON 1990, STORRS & STORRS 1990). Als Angehöriger der Sektion Fraxinus (FITSCHEN 1987) dürfte ihr Pollen zum Fraxinus
excelsior-Typ gehören.
Die Gattung Pistacia (Anacardiaceae) tritt in Form kleiner Bäume und Sträucher in den Halbwüsten, Steppen und Trockentälern des westlichen Himalaja und östlichen Tibets auf (PAFFEN et al. 1956, TROLL 1939, SCHWEINFURTH & CHEN WEILIE 1984, ZHANG JINGWEI et al. 1981, MEUSEL & SCHUBERT 1971a). Nachweise liegen vom Nanga Parbat und aus dem Nianbaoyeze („Profil 3“) vor.
Die Familie Elaeagnaceae ist in den Diagrammen durch Pollen der Gattung Hippophaë vertreten. H. rhamnoides bildet unter den unterschiedlichsten klimatischen Bedingungen entlang der Flüsse auf Schotterbänken dichte Gebüsche. Daneben kommen weitere, aber palynologisch nicht abtrennbare Arten vor (HUANG RONGFU 1987, LU RONGSEN 1992, PRACH 1994, ROUSI 1971, SORSA 1971).
Die Familie der Gentianaceae (Enziangewächse) ist im Untersuchungsraum mit mehreren Gattungen vertreten. In Xizang besitzen die Gattungen Gentiana (99 Arten), Swertia (22) und Lomatogonium (12) gemeinsam etwa 85 % aller Arten der Familie (HE
TINGNONG 1981). Sie sind in Kobresia-Matten, Feuchtwiesen, im Unterwuchs der alpinen Gebüschen und in der subnivalen Stufe verbreitet (BJÖRK 1993, CHANG D. H. S. 1981, HUANG RONGFU 1987). Sämtliche zu den Gentianaceae gezählten PK waren striato-reticulat. Der Swertia-Typ steht für die Gattungen Swertia und Lomatogonium. Nach Ausbildung der Porenregion stammt in manchen Profilen fast die Hälfte des untersuchten Pollens aus der Gattung Lomatogonium. Vereinzelt ließen sich PK der kleinen Gattung Tripterospermum zuordnen (VP). Alle übrigen Funde wurden als Gentianaceae p. p. zusammengefaßt. Nicht striato-reticulate PK, wie von manchen Swertia-Arten bekannt, traten nicht auf (VP, NILSSON 1964, 1967, JONSSON 1973).
Der Galium-Typ gehört zu den Rubiacaeae, einer der formenreichsten Familien der Blütenpflanzen. Der Typ scheint auf die Gruppe der Rubieae mit Gattungen wie Galium, Asperula und Rubia beschränkt zu sein (ENGLER 1964, WANG FUHSIUNG et al. 1995). Galium wird von feucht-nassen Lägerfluren angegeben (SCHWEINFURTH 1957, TROLL 1939).
Viele der Lamiaceae oder Labiatae sind an die trockenen Bedingungen der Steppen-gebiete und Pionierstandorte der Flußtäler angepaßt; andere sind mesophytisch. Die Familie kommt weltweit mit über 220 Gattungen und 5600 Arten vor, ist dabei aber vom Mittelmeerraum bis nach Zentralasien besonders reich entfaltet (MANI 1978, MARTENSEN &
PROBST 1990). In der vorliegenden Arbeit wurden drei Pollentypen unterschieden, denen nach MOORE et al. (1991) und FAEGRI & IVERSEN (1989) die folgenden im Himalaja vorkommenden Gattungen angehören (KLOTZ 1989, MANI 1978, POLUNIN & STAINTON
1990): Prunella-Typ (Nepeta, Glechoma), Mentha-Typ (Mentha, Thymus, Origanum,

4. Material und Methoden – Übersicht der auftretenden Taxa 22
Clinopodium, Salvia) und Stachys-Typ (Stachys, Lamium, Leonurus, Ajuga, Scutellaria). Wie die übrigen Gattungen der Stachyeae (ENGLER 1964) weisen auch die Vertreter der wichtigen Gattung Phlomis tricolpate PK auf. Während aber P. spectabilis zum Stachys-Typ zählt, fehlt anderen Arten der Gattung ein Suprareticulum (VP).
Die Scrophulariaceae sind im Untersuchungsgebiet durch die Gattung Pedicularis reichlich vertreten. Pollenfunde waren sehr selten. Nach Zahl und Anordnung der Colpi wurde zwischen dem Pedicularis palustris-Typ und P. oederi-Typ unterschieden (BEUG
1961). Andere Pedicularis-Arten gehören dem Rhinanthus-Typ (MOORE et al. 1991) an. Sehr ähnlich ist der Viola palustris-Typ (Violaceae). Die wenigen übrigen, zu den Rachenblütlern gerechneten Funde, wurden als Scrophular iaceae p. p. bezeichnet.
Arten des Wasserschlauchs, Utricularia, wie in der Nordhemisphäre U. vulgaris und in Tibet U. aurea, besiedeln stehende bis langsam fließende Gewässer (ENGLER 1964, OBERDORFER 1990, ZHANG JINGWEI et al. 1988). Nachweise der Gattung liegen aus der „Teichphase“ des Profils „Lhasa Stadtbereich“ vor.
Die Plantaginaceae sind an gestörte Wuchsorte (z. B. mit starker Trittbelastung) angepaßt. Von Plantago lanceolata, dem P. alpina-Typ und vom P. major/media-Typ erreicht nur der letztere manchmal höhere Werte, so z. B. in den Diagrammen „Batyatum
Jut“ und „Lhasa Stadtbereich“. Zur Pollenmorphologie der Gattung siehe MOORE et al. (1991), CLARKE & JONES (1980a) und auch UBERA et al. (1988).
Sträucher der Gattung Lonicera (Caprifoliaceae) wachsen in Gebüschen im Bereich der Kobresia-Matten sowie in alpinen Steppen. In der Hochgebirgshalbwüste Tibets und in den Trockentälern treten sie im Galeriebuschwerk entlang der Wasserläufe auf (BJÖRK 1993, CHANG D. H. S. 1981, CHEN GUICHEN et al. 1994, HUANG RONGFU 1987, MIEHE 1991, SCHWEINFURTH 1957, WARDLE 1981).
PK der Gattung Valeriana (Valerianaceae, Baldriangewächse) sind in Mitteleuropa durch auf Verrucae sitzende Stacheln gekennzeichnet (CLARKE & JONES 1980b, WAGENITZ
1956); darüber hinaus gibt es aber auch Arten ohne Verrucae (CLARKE 1978). Solche PK ähneln denen mancher Arten der Dipsacaceae (CLARKE & JONES 1981). Der Pollen beider Familien ist an dem Halo um die Aperturen zu erkennen. Fast immer wurden neben PK ohne Verrucae auch solche mit Verrucae gefunden, so daß auch erstere die Familie der Valer ianaceae repräsentieren dürften.
Zu den größten Pflanzenfamilien gehören die Asteraceae. Ihre Hauptentfaltung haben sie in außertropischen Gebieten mit Trockenzeiten (MARTENSEN & PROBST 1990). In Xizang gibt es fast 90 Gattungen mit mehr als 500 Sippen. Von den Gattungen Aster, Artemisia, Senecio, Saussurea und Cremanthodium sind jeweils über als 30 Arten bekannt (SHI ZHU &
CHEN YILIN 1981). In der Unterfamilie der Cichorioideae (= Liguliflorae) zeichnen sich bis auf wenige Ausnahmen die altweltlichen Vertreter durch fenestrate PK (ENGLER 1964, BLACKMORE 1982, 1984, TOMB 1975). Im Untersuchungsraum sind u. a. die Gattungen Crepis, Dubyaea, Hieracium, Lactuca, Leontodon, Picris, Scorzonera, Taraxacum und Tragopogon verbreitet (MANI 1978, POLUNIN & STAINTON 1990, PAFFEN et al. 1956, TROLL
1939). Nach SCHWEINFURTH (1957) sind Scorzonera und Taraxacum für humose

4. Material und Methoden – Übersicht der auftretenden Taxa 23
Wiesensteppen charakteristisch, Hieracium für stärker genutzte Standorte.
Alle übrigen Asteraceae wurden früher in der Unterfamilie Asteroideae (= Tubuliflorae) vereint und besitzen zumeist echinate, nicht fenestrate PK (ENGLER 1964), die sich in Typen unterteilen lassen. Der Aster-Typ (sensu MOORE et al. 1991, Senecio-Typ bei FAEGRI &
IVERSEN 1989) kommt in den Gruppen Heliantheae, Astereae, Inuleae, Senecioneae und Calenduleae vor (MARTENSEN & PROBST 1990). Im Untersuchungsraum häufig sind Vertreter der Gattungen Anaphalis, Aster, Bidens, Cremanthodium, Galinsoga, Erigeron, Leontopodium, Ligularia, Senecio (SHI ZHU & CHEN YILIN 1981 u. a.). Die zum selben Verwandtschaftskreis zählende windblütige Spitzklette Xanthium (Einzelfund im Profil „Alambar“) hat hingegen PK vom Ambrosia-Typ (BEUG 1964, MOORE et al. 1991). Für viele Arten der Gruppe Anthemideae ist der Anthemis-Typ (sensu MOORE et al. 1991, Achillea-
Typ bei FAEGRI & IVERSEN 1989) typisch. Er tritt u. a. in den Gattungen Anthemis, Achillea, Matricaria, Chrysanthemum und Tanacetum auf. Die zahlreichen Arten von Artemisia wachsen unter recht verschiedenen ökologischen Bedingungen (MIEHE et al. 1997). Ihr vom Wind verbreiteter Pollen hebt sich von dem der anderen Anthemideae ab. Eine pollenmor-phologische Artdiagnose scheint kaum möglich zu sein (FRENZEL 1993, SINGH &
JOSHI 1969). Die Cardueae (Distelgruppe) und Mutisieae (in Asien Ainsliaea und Gerbera) besitzen Vertreter mit großen, dickwandigen PK. Hierzu gehört auch der Saussurea-Typ. Saussurea ist nach SHI ZHU & CHEN YILIN (1981) die größte Gattung der Asteraceae in Xizang. Ihre Vertreter gedeihen in Kobresia-Matten und Steppen. Einige auffällige Arten prägen die Vegetation der subnivalen Stufe (HUANG RONGFU 1987, THELAUS 1992, WANG
JINTING 1981). Aus dem Karakorum liegen PK vor, die wohl dem von ZEIST &
BOTTEMA (1977) an Cousinia spp. und Centaurea xeranthemoides aufgestellten Cousinia-
Typ entsprechen. VP von Cousinia thomsonii (MIEHE 1295) und Cousinia sp. (MIEHE 5291)
aus diesem Bereich zeigen aber einen anderen Pollentyp. Daher wurde für PK vom Cousinia-Typ sensu ZEIST & BOTTEMA (1977) der Name Centaurea/Cousinia-Typ gewählt.
PK aus der Sektion Eupotamogeton der Gattung Potamogeton (Potamogetonaceae) und der Gattung Triglochin (Juncaginaceae) ähneln einander (MOORE et al. 1991). Triglochin
maritima und T. palustre können aber im Phasenkontrast von Eupotamogeton unterschieden werden (BEUG 1961). Dies gilt offenbar auch für den Nordosten Tibets, wo BEUG (1987) in Material aus den Kakitu-Bergen Pollen von Triglochin in großen Menge fand. Nachdem die eigene Bearbeitung mehrerer Profile aus dem Nianbaoyeze und dem Karakorum keinen Nachweis von Triglochin erbrachte, wurden VP von T. maritima
(MIEHE 8303) und T. palustre (leg. SCHLÜTZ, Nepal 1995, det. B. DICKORÉ, Göttingen) angefertigt. Beide VP zeigen hier den Pollentyp von Eupotamogeton, daher die Bezeichnung Potamogeton/Triglochin-Typ.
Die PK vieler Sippen aus der Familie der Liliaceae (s. l.) weisen kaum Merkmale für eine systematische Zuordnung auf (BEUG 1961). Hingegen sind die PK der überwiegend irano-turanischen Gattung Eremurus, die mit wenigen Arten in den Karakorum hineinreicht, leicht kenntlich (Foto 6 und 7, im Anhang). Die pollenmorphologisch nicht unähnliche

4. Material und Methoden – Übersicht der auftretenden Taxa 24
Gattung Aloë kommt im Untersuchungsgebiet nicht vor (AGARWAL 1983, BONNEFILLE &
RIOLLET 1980, DICKORÉ 1995, REILLE 1995, WANG FUHSIUNG et al. 1995).
Die Poaceae besitzen einen relativ einheitlichen Pollentyp. Die umfangreichen Vorar-beiten von FIRBAS (1937) und Oberflächenanalysen ermöglichten BEUG (1961) die Abtren-nung verschiedener Getreide-Typen. Mit der gleichen Problematik beschäftigten sich auch ANDERSEN & BERTELSEN (1972), BONNEFILLE & RIOLLET (1980), KÖHLER & LANGE (1979) und ROWLEY (1960). ZEIST & BOTTEMA (1977) schieden PK ab einer Größe von 40 µm als Cerealia-(Getreide-) Typ aus. Auf die besonderen Gefahren einer Fehlinterpretation eines nur nach der Größe aufgestellten Getreide-Typs im südasiatischen Raum geht VISHNU-MITTRE (1981) ein. Für Indien gibt er Wildgrasarten mit einer Pollenkorngröße von über 50 µm an. Von diesen wächst Heteropogon contortus im Karakorum bis in Höhen von 3500 m (DICKORÉ 1995). In der vorliegenden Arbeit wurden PK mit einem Durchmesser von mehr als 37 µm und entsprechender Apertur im Phasenkontrast untersucht (vergl. BEUG
1961). Fast immer gehörten sie zum Hordeum-Typ, im Profil „Lhasa Stadtbereich“ trat zudem der Triticum-Typ auf. In Poaceae > 37 µm sind der Hordeum-Typ und erhaltungs-bedingt nicht bestimmbare PK enthalten. Während der Hordeum-Typ wegen seiner weiten Verbreitung unter den Wildgräsern nicht ohne weiteres als Hinweis auf Ackerbau zu werten ist, dürften die Funde vom Triticum-Typ bei Lhasa wohl auf Weizenanbau zurückgehen (LIU
YANHUA 1992). Eine pollenmorphologische Untersuchung der Wildgräser bei Lhasa fehlt aber.
Vereinzelt traten PK von der auf ruhigen Wasseroberflächen treibenden Gattung Lemna (Lemnaceae) auf.
Als Sparganium-Typ werden die in der Gattung Sparganium (Sparganiaceae) und von Typha angustifolia (Typhaceae) ausgebildeten Monaden bezeichnet. Zum Typha latifolia-Typ (Diagramm „Lingfeng“) gehören Tetraden von Typha-Arten (BEUG 1961, PUNT 1976).
Die Matten Tibets und des Karakorums werden im wesentlichen von Sauergräsern der Gattung Kobresia und einigen Carex-Arten gebildet. Beide Gattungen gedeihen zudem auch gut an solchen für die Entnahme von Pollenprofilen bevorzugten feucht-nassen Standorten. Deshalb hat der Pollen der Cyperaceae in den Diagrammen fast immer sehr hohe Anteile.
Auf die wenigen Nachweise von Buxus (Buxaceae), Euphorbia (Euphorbiaceae), Acer (Aceraceae), Tilia (Tiliaceae), Campanula-Typ (Campanulaceae), Boraginaceae, Hedera (Araliaceae), Hypericum-Typ, Linum (Linaceae) und Malvaceae soll hier nicht näher eingegangen werden.
CHLOROPHYTA – Grünalgen
Die Gattungen Botryococcus und Pediastrum wurden regelmäßig erfaßt, Staurastrum nur ein einziges Mal. Botryococcus scheint im Gegensatz zu Pediastrum nicht nur im freien Wasserkörper, sondern auch noch unter genügend feuchten Bedingungen zu gedeihen. Entsprechend ist Botryococcus nicht selten auch in den rein torfigen Partien der Profile

4. Material und Methoden – Übersicht der auftretenden Taxa 25
vorhanden. Soweit erkennbar handelte es sich bei den Kolonien von Pediastrum über-wiegend um die Art P. boryanum. Im Profil „Lhasa Stadtbereich“ war die Gattung Pediastrum während einer „Teichphase“ sehr formenreich entwickelt. Ob es sich bei den unterbrochenen und teilweise deutlicher skulpturierten Kolonien um Varietäten von P. boryanum oder um eigene Arten handelte, konnte nicht entschieden werden. Funde der Zieralgengattung Staurastrum - Halbzellen mit dreistrahligem Grundriß - liegen nur aus dem Profil „Alambar“ vor (vergl. Abb. 187/2 in SCHUBERT et al. 1983). Abbildungen und Schlüssel zu den genannten Algengruppen finden sich u. a. bei AGARKAR et al. (1984), BARRIENTOS (1979), KOMAREK & FOTT (1983) sowie SCHUBERT et al. (1983).
4. BRYOPHYTA – Moose
Unter den Moosen bilden u. a. die Hepaticae (Lebermoose) erhaltungs- und bestimmungs-fähige Sporen aus. In fast allen Profilen war der Riccia-Typ (sensu MOORE et al. 1991) vorhanden, der auf offene Böden hinweist. Sporen der Gattung Riella traten im Profil „Lhasa Stadtbereich“ auf. Die Vertreter dieser Gattung sind einjährig und kommen in niederschlagsarmen Gebieten mit jährlichen Regenzeiten vor. Sie siedeln auf dem Schlamm-boden seichter, stehender Gewässer. Meist handelt es sich um relativ konkurrenzarme Salz-wasserstandorte (FREY et al. 1995, FUKAREK et al. 1992). Die fossilen Funde gleichen den von Riella halophila und Riella notarisii bei ERDTMAN (1957) und HOCHULI &
GRONER (1985) abgebildeten Sporen (Foto 8 und 9, im Anhang). Von der besonders in Trockengebieten vorkommende Gattung Targionia liegt ein Nachweis im Profil „Lingfeng“ vor. Die Sporenmorphologie entspricht der von Targionia hypophylla (ERDTMAN 1957). Weitere Sporen mußten wegen unzureichender Bestimmungsmöglichkeit unter der Bezeichnung Hepaticae p. p. zusammengefaßt werden.
Sporen der Gattung Sphagnum (Sphagnaceae) wurden angesichts der überwiegend aus Cyperaceen-Resten aufgebauten Torfe nur sehr selten angetroffen (Profil „Lhasa Stadtbereich“ und „Profil 16“ aus dem Nianbaoyeze). Morphologisch gleichen sie mittel-europäischen Sphagnum-Sporen (ZHANG YULONG & WU PANCHENG 1990).
PTERIDOPHYTA – Sporen-Gefäßpflanzen
Die Benennung von Sporen aus der Familie der Lycopodiaceae folgt der heute üblichen Auftrennung der Gattung Lycopodium. Es wurden der Diphasiastrum-Typ und Huperzia unterschieden. In Proben mit Zusatz von Lycopodium-Sporen (zur Bestimmung der Pollenkonzentration) mögen einzelne fossile Sporen nicht als solche erkannt worden sein.
Die Selaginellaceae werden rezent durch die mehr als 700 Arten umfassende Gattung Selaginella repräsentiert. Die Mehrzahl von ihnen wächst in feuchten tropischen und subtropischen Urwäldern. Daneben gibt es einige xerotherme Vertreter, die Trockenzeiten überdauern (HUANG TSENGCHIENG 1981, FUKAREK et al. 1992). ZHANG YULONG et al. (1990) beschreiben 21 der ungefähr 50 chinesischen Arten. Im Profil „Lhasa Stadt-bereich“ wurden acht Mikrosporen dieser Gattung gefunden, die im Diagramm zusammen

4. Material und Methoden – Übersicht der auftretenden Taxa 26
als Selaginella aufgeführt sind. Einige der Sporen ähnelten Selaginella selaginoides, andere besaßen eine unspezifisch verrucat-tuberculate Oberfläche, wie sie u. a. S. tamariscina zeigt (HUANG TSENGCHIENG 1981, KREMP & KAWASAKI 1972). Eine Spore mit lang ausgezogenen, im optischen Schnitt an Golfschläger erinnernden Baculae könnte von S. superba stammen. Eine Spore mit weit von der Exine abgesetzten Perine dürfte zu S. sanguinolenta zu stellen sein. S. tamariscina und S. sanguinolenta sind aus dem Gebiet von Lhasa bekannt (MIEHE 1988).
Equisetum (Equisetaceae) ist in den Diagrammen „Sirle Bala“ und „Lingfeng“ recht häufig, im „Profil 3“ aus dem Nianbaoyeze gering vertreten. In der Mehrzahl der Fälle stehen Schachtelhalme an feuchten Plätzen (BJÖRK 1993, FUKAREK et al. 1992, ZHANG
YULONG et al. 1990)
Die folgenden Gruppen bilden innerhalb der Pteridophyta die Klasse der sog. Echten Farne, der Filicatae.
Sporen von Botrychium (Ophioglossaceae) wurden in den Profilen „Alambar“, „Ling-feng“ und dem „Profil 14“ aus dem Nianbaoyeze gefunden. Sie gleichen dem Botrychium
lunaria-Typ (MOORE et al. 1991). Viele Arten der Gattung sind empfindlich gegen Beschat-tung und vermögen nur an nährstoffarmen Standorten der Konkurrenz anderer Pflanzen zu widerstehen (FUKAREK et al. 1992, ZHANG YULONG et al. 1990).
Wohl nur die Gattung Schizaea (Schizaeaceae) besitzt striate, monolete Sporen. Die breiten Muri verlaufen ungefähr parallel zur Apertur, die Striae sind schmal. In China gibt es zwei Arten, die an trockenen Standorten auf offenen, sauren Böden wachsen. Aus dem Profil „Lingfeng“ liegt ein Einzelfund vor.
Die überwiegend paläotropische Gattung Plagiogyria (Plagiogyriaceae) besitzt z. T. dickwandige, mit Orbicules besetzte Sporen. Die Sporen wurden vereinzelt im Profil „Ling-feng“ gefunden. Bei Verlust der Orbicules fällt es schwer, die Sporen von denen anderer Gattungen zu unterscheiden. Unter dem Sammelbegriff Filicatae tr ilet p. p. wurden sie mit allen anderen morphologisch unspezifischen trileten Farnsporen zusammengefaßt.
Verschiedene der früher als untergeordnete Taxa zur Familie der Polypodiaceae gestellte Sippen werden heute als eigene Familien geführt (siehe MARTENSEN & PROBST 1990). Sie haben meist monolete Sporen mit einer charakteristisch ausgebildeten Perine. Bei Verlust der Perine, was bei fossilem Material die Regel ist, sind die Sporen kaum bestimmbar und werden gemeinsam mit den monoleten Sporen anderer Farne als Filicatae monolet p. p. bezeichnet. Von den übrigen monoleten Farnsporen ist der Polypodium-Typ durch die Skulptur der Exine zu unterscheiden. Bei den meisten Sippen dieses Typs (Polypodium, Belvisia, Drynaria, Lepidogrammitis, Davalliaceae) bleibt die Skulptur deutlich schwächer als bei Polypodium vulgare (Polypodiaceae). Neben einer Vielzahl tropisch-subtropischer Arten siedeln einige dieser Farne auch in gemäßigten Klimaten (ZHANG YULONG et al. 1990, NAYAR 1963, NAYAR & DEVI 1964c).

4. Material und Methoden – Vegetation und Pollenniederschlag 27
Der Pteris-Typ (Pteridaceae) tritt überwiegend in den Gattungen Pteris (Flügel- oder Saumfarn) und Onychium (Sinopteridaceae) auf. Ihre trileten Sporen besitzen eine auffällig skulpturierte Exine. Beide Gattungen haben in China ihren Verbreitungsschwerpunkt im tropisch-subtropischen Südosten und reichen nördlich bis zum Qinling (NAYAR & DEVI
1966, ZHANG YULONG et al. 1990).
Der einzige Vertreter der Gattung Pteridium (Hypolepidaceae) ist der Adlerfarn P. aqui-
linum. Er ist fast kosmopolitisch an gestörten Standorten zu finden (ENGLER 1954), jedoch sind die Sporen nicht charakteristisch genug um sie sicher von anderen evtl. im Untersuchungsraum auftretenden Farnen abzutrennen, so daß die Bezeichnung Pteridium-Typ gewählt wurde.
4.6 Vegetation und Pollenniederschlag
Die Vegetation eines Gebietes spiegelt sich im Pollenniederschlag wider, wenn auch in oft verzerrter Form. Die Rekonstruktion der früheren Vegetation aus palynologischen Daten wird durch den Vergleich des Pollengehalts von Oberflächenproben mit den rezenten Vegetations- und Klimabedingungen bekanntlich sehr erleichtert.
Im folgenden sind kurz die Ergebnisse ausgewählter Veröffentlichungen dargestellt, die ausschließlich oder ergänzend zu Profilen auch Oberflächenproben berücksichtigen. Die Prozentangaben beziehen sich zumeist auf Gesamtsummen, in denen die fast immer geringen Anteile von Cyperaceen und Sporen enthalten sind.
Die Untersuchungen von WENG CHENGYU et al. (1993) an Oberflächenproben aus dem westlichen Kunlun Shan zeigen eine klare pollenanalytische Unterscheidbarkeit der dort vorkommenden Vegetationseinheiten Fichtenwälder, Wacholderoffenwälder, Steppen, Wüsten, Polsterfluren und alpine Matten. Die Spektren aus den fast vegetationsfreien Gletschervorfeldern sind erwartungsgemäß stark von Fernflug beeinflußt. Die höchsten Picea-Anteile (10 %) entsprechen sogar den Werten einiger Proben aus lichten Fichtenwäldern.
In den von VAN CAMPO et al. (1996) untersuchten Oberflächen- und Pollenfallenproben aus dem Karakorum, dem Kunlun Shan und aus Kashgar erreichte die Summe des Baumpollens selten 2 %. Etwa 50 % aller PK stammten von Artemisia, am zweithäufigsten waren Chenopodiaceen.
Auch Oberflächenproben aus dem Nordosten Chinas weisen je nach Vegetation unter-schiedliche Pollenspektren auf (YUAN SHAOMIN & SUN XIANGJUN 1990). Gleiches gilt für die Waldtypen Sichuans (JARVIS 1993, JARVIS & CLAY-POOLE 1992) und anscheinend auch für die Mongolei (SHILOVA 1984).
BHATTACHARYYA (1989) fand 500 Meter über der Waldstufe im Kaschmir-Tal Pollen von Abies, Picea und Pinus noch mit Anteilen von 40 - 50 %, was wohl mit der geringen lokalen Pollenproduktion an den Untersuchungspunkten zusammenhängt. In über 400 km Entfernung von Wäldern ist Pinus noch mit 5 % vertreten, Cedrus mit 1 %. Über Fernflug

4. Material und Methoden – Vegetation und Pollenniederschlag 28
von Cedrus-Pollen berichten auch BORTENSCHLAGER & PATZELT (1978) aus dem Wakhan-Tal im Hindukusch und BEUG (1987) aus dem Kakitu-Gebirge Nordost-Tibets.
In Kiefernwäldern erreicht Pinus einen Pollenanteil von über 95 % bis fast 99 % am Gesamtpollen Im allgemeinen weisen Werte ab etwa 30 % auf das Vorkommen von Pinus am Untersuchungspunkt hin. In sommergrünen Laubwäldern in 500 m Abstand vom Kiefernwald können aber noch über 50 % des Pollens von Pinus stammen. In der alpinen Wüste Tibets, bei also äußerst geringer Pollenproduktion vor Ort, wurden sogar in 200 km Entfernung noch Pinus-Anteile von bis zu 34 % gefunden (LI WENYI & YAO ZUJU 1990). Eine ähnliche Überrepräsentanz zeigt auch die Gattung Quercus (LI XU & LIU JINLING 1988, JARVIS 1993, JARVIS & CLAY-POOLE 1992).
Die Prozentwerte von Picea sinken schon bei Abständen von 1 bis 2 km vom Wald auf 0,5 bis 5 % des Gesamtpolleneintrages ab (CUI ZHIJIU et al. 1995). Für den Pollen von Larix ist ein ausgesprochen geringer Windtransport anzunehmen (JOCHIMSEN 1986).
Mit nur 5 bis 10 % fallen die Werte von Abies selbst in Proben aus den Abies-Rhodo-
dendron-Wäldern Sichuans erstaunlich niedrig aus. Gleiches gilt für die insektenbestäubte Gattung Rhododendron. Selbst dort, wo sie mit Abies die Wälder beherrscht, hat sie nur Prozentwerte von 4 bis 5 % (LI XU & LIU JINLING 1988, JARVIS 1993, JARVIS & CLAY-POOLE 1992).
Wacholder bilden im Untersuchungsraum sowohl strauchförmige Bestände als auch lichte Wälder. Mit 40 % liegen die Werte von Juniperus dabei im dichten Strauchwerk deutlich über denen des Offenwaldes mit nur 5 bis 10 % des Gesamtpolleneintrages (BHATTACHARYYA 1989, WENG CHENGYU et al. 1993). Pollen vom Juniperus-Typ kann durch Wind über größere Entfernungen transportiert werden (BHATTACHARYYA 1989, HAMILTON & PERROTT 1980).
Die in den trockenen Gebieten Asiens nicht unbedeutenden Familien der Lamiaceen und Fabaceen treten wohl aufgrund von Insektenbestäubung und Blütenbau meist nur mit wenigen Funden im Diagramm auf und erscheinen somit pollenanalytisch nur schwer er-faßbar zu sein. Ähnliches kann auch für insektenbestäubte Baumarten gelten (GUPTA &
BERA 1996). Auf Probleme beim Nachweis für die im Becken von Kathmandu in Shima-
Castanopsis-Wäldern stehende Gattung Shima weisen VISHNU-MITTRE & SHARMA (1984) hin.
Für den Umgang mit unterschiedlicher Pollenproduktion und Pollenverbreitung ver-schiedener Pflanzengruppen bei der Interpretation palynologischer Befunde sei auch auf die Werke von FIRBAS (1949, 1952) und LANG (1994) sowie auf die Arbeit von JOCHIMSEN (1986) verwiesen.
Wie oben schon angedeutet, ist bei Proben, die nach Alter oder Herkunft aus einem vegetationsarmen Umfeld stammen, mit weit transportierten PK zu rechnen (HOROWITZ
1992, WENG CHENGYU et al. 1993, LI WENYI & YAO ZUJU 1990). Die dabei auftretenden Einflußgrößen wie Windverhältnisse, Orographie und lokale Pollenproduktion sind auch in den Gebirgen Europas zu beobachten (DRESCHER-SCHNEIDER 1993, LANG 1993, MARKGRAF
1980). Beim Polleneintrag in Seen können neben dem Wind auch die einmündenden

4. Material und Methoden – Vegetation und Pollenniederschlag 29
Wasserläufe von Bedeutung sein (KVAVADZE 1996, SUN XIANGJUN & WU YUSHU 1988, XU
QINGHAI et al. 1996).
Nach den angeführten Beispielen und dem allgemeinen Wissensstand der Palynologie lassen sich die meisten Vegetationstypen palynologisch charakterisieren. Dem Nachweis einiger Sippen sind aber enge Grenzen gesetzt, und je nach Art und Herkunft des Unter-suchungsmaterials ist der Eintrag von Pollen durch Wind und Wasser zu berücksichtigen.
Eigene Untersuchungen an Oberflächenproben wurden an Material aus dem Nianbao-yeze durchgeführt (Kap. 6.3.1, Abb. 8).

5. Der Untersuchungsraum – Lage und Geologie 30
5 Der Untersuchungsraum
Nach einem allgemeinen Überblick werden in den folgenden Abschnitten die Unter-suchungsräume vorgestellt. Die Reihenfolge orientiert sich am Alter der Profile.
5.1 Lage und Geologie
Die Entstehung der Untersuchungsräume in den Gebirgen im Osten Chinas (Qinling), in Tibet (Nianbaoyeze, Lhasa) und im Norden Pakistans (Nanga Parbat, Karakorum) kann bis in das frühe Paläozoikum zurückverfolgt werden (Abb. 1). Zu jener Zeit stießen der heutige Nordrand des Plateaus und Südchina mit dem nördlich von ihnen gelegenen Tarim-Nordchina Block auf rund 3500 km Länge vom östlichen Pamir bis hin zum nördlichen Qinling und Dabei-Gebirge zusammen. Die Anlagerungen weiterer kontinentaler Krustenteile und Inselbögen aus den sich zeitweise auftuenden Meeresarmen (Kunlun-Ozean, Paläo-, Meso- und Neo-Thetys) folgen bis in das Tertiär hinein. Die in den Bereichen der ehemaligen Meeresböden entstandenen Ophiolith-Gürtel zeichnen die Grenzen der sich in ostwestlicher Richtung erstreckenden tektonischen Einheiten nach. Die Verschmelzungs-linie des indischen Subkontinents mit Eurasien – die Indus-Yarlung-Zangbo-Sutur – kann auf der Nordseite des Himalaja durch die Täler beider Flüsse und westlich des Nanga Parbat bis nach Afghanistan hinein verfolgt werden (BENDER 1995b, CHANG CHENGFA 1996, LI
TINGDONG et al. 1986, LI TINGDONG 1996, SHEN XIANJIE 1996, SEARLE 1991, SENGÖR 1986, YANG YICHOU et al. 1982).
Die nordostwärts gerichtete Bewegung (15 cm/a) der von Gondwana abgelösten indischen Platte führte schließlich zur Subduktion unter den eurasiatischen Kontinent. Die ältesten in Indien gefundenen Fossilien von Säugetieren und Pflanzen eurasiatischer Abstammung sowie das Alter der jüngsten marinen Sedimente belegen eine Schließung der Neo-Thetys vor 40 bis 50 Mio. Jahren. Die weiter anhaltende Drift Indiens (5 cm/a) gegen Eurasien führte zur Auffaltung Tibets und des Himalaja (CHANG CHENGFA 1996, KAZMI
1989, LI TINGDONG 1996, MILLER 1982, TRINKLER 1932).
Die Hebungsprozesse verliefen zeitweise so langsam, daß es phasenweise zu Eineb-nungen der Landoberfläche kam. Allgemein wird dem Tibetanischen Hochland ab dem Erreichen der halben heutigen Höhe ein entscheidender Einfluß auf die Entstehung des Monsuns eingeräumt. Nach den unterschiedlichen Ansichten wurde dieses Niveau bereits im Miozän, im Pliozän oder erst während einer schnellen Hebungsphase im Pleistozän erreicht. Die Vorstellung einer schnellen pleistozänen Anhebung rührt daher, daß die Klima-änderungen in Tibet oftmals zum großen Teil auf die zunehmende Höhe des Tibetanischen Plateaus, aber kaum auf die globale pleistozäne Klimaverschlechterung zurückgeführt werden (ANIYA & YOKOYAMA 1991, CHANG CHENGFA 1996, FIELDING 1996, FLOHN 1981, HARRISON et al. 1992, JIANG FUCHU & WU XIHAO 1993, LI BINGYUAN et. al. 1985, LI JIJUN
1991a, LI JIJUN et al. 1979, LI TINGDONG 1996, LIU TUNGSHENG & DING ZHONGLI 1993, PRELL & KUTZBACH 1992, SHEN XIANJIE 1996, SINHA & UPADHYAY 1995, YANG YICHOU et al. 1982, ZHANG QINGSONG et al. 1994, ZHENG BENXING 1989, ZHU CHENG 1995). Die unter-schiedlichen Ansichten werden von FORT (1996) diskutiert.

5. Der Untersuchungsraum – Klima 31
Für die einzelnen Plattenteile Tibets und für die der umgebenden Gebirge werden unter-schiedliche Hebungsraten von meist 0,1 - 1 cm pro Jahr angenommen. Die horizontalen Verschiebungen betragen jährlich bis zu 2 cm (CHANG CHENGFA 1996, FORT 1996, HOLMES
& STREET-PERROTT 1989, LI TINGDONG 1996, LIU TUNGSHENG et al. 1996, ZHANG
QINGSONG et al. 1994, ZHAO YONGTAO 1994, WHITTINGTON 1996, ZEITLER et al. 1982, ZHU
CHENG 1995). In Ergänzung zu den tektonischen Hebungsprozessen führt KUHLE (1993) eine glazialisostatische Vertikalbewegung seit Abschmelzen der letztglazialen Inlandeisbedeckung an. Hingegen gehen FORT (1996) und FIELDING (1996) von einer gegenwärtigen Absenkung des Hochlandes infolge einer ostwestlich gerichtete Dehnung und Verdünnung der Kruste aus.
Die bis heute anhaltende tektonische Tätigkeit ließ Vulkane entstehen und zeigt sich bis in die Gegenwart durch Erbeben und heiße Quellen. Im Raum um Lhasa und im Karakorum erreichen die Beben die Stärke 8, im Gebiet des Nianbaoyeze die Stärke 7 (BENDER 1995c, BUCHROITHNER 1978, CHANG CHENGFA 1996, CHEN ZHIMING 1996, LEHMKUHL 1995a, TENG JIWEN & LIN BANZUO 1984, YANG YICHOU et al. 1981).
5.2 Klima
Nach der am solaren Strahlungsregime orientierten klimatischen Großgliederung der Erde liegen die in der vorliegenden Arbeit untersuchten Gebiete in den Subtropen. Sie sind durch einen relativ hohen Sonnenstand und extreme Strahlungsexposition in den Gebirgen gekennzeichnet.
Die subtropische Zone wird in Eurasien durch das trocken-kalte Tibetanische Plateau in eine monsungeprägte Ostseite und eine mediterrane Westseite gegliedert. Kennzeichnend für den Monsun ist ein ausgeprägter jahreszeitlicher Wechsel der vorherrschenden Windrichtung und ein damit einhergehender Wetterwandel. Im Sommer führen südliche Winde warme, maritime Luftmassen heran, die über dem indischen Subkontinent bis nach Nordpakistan hinein als Südasienmonsun sowie in weiten Teilen des östlichen Chinas ein-schließlich des Ostrandes von Tibet als Ostasienmonsun erhebliche Niederschläge bringen. Das Hochland von Tibet verstärkt mit dem über ihm ausgebildeten Hitzetief das monsunale Wettergeschehen und schirmt zugleich weite Bereiche des nördlichen Chinas vom Nieder-schlag ab. Das südliche Tibet, vom indischen Subkontinent durch den Himalaja getrennt, wird entlang der meridionalen Stromfurchen und im unteren Quertal des Yarlung Zangbo noch vom Südasienmonsun erreicht.
Die Wintermonate sind durch trocken-kalte Winde aus einem Kältehoch über der Mon-golei und Sibirien geprägt. Der Südosten Chinas ist durch die Gebirgsbarrieren von Tibet und des Qinling gegen die klimatischen Auswirkungen dieser Luftmassen zwar abgeschirmt, wird aber trotzdem Winter für Winter von „Kältewellen“ heimgesucht. Der Temperaturabfall während eines solchen Ereignisses beträgt mehr als 10 °C binnen 2 Tagen und kann bis zu 28 °C erreichen.

5. Der Untersuchungsraum – Vegetation 32
Während die innerasiatischen Wüsten und das Tibetanische Hochland im Winter ausgesprochen niederschlagsarm sind, führen Störungen aus der Polarfront im südöstlichen China zu Niederschlägen. Der indische Subkontinent bleibt im Schutze des Himalaja von den Auswirkungen des Wintermonsun weitgehend unbeeinflußt (BLÜTHGEN & WEISCHET
1980, DING YIHUI 1994, FLOHN 1981, LAUER 1993, DOMRÖS & PENG GONGBING 1988, LAUER et al. 1996, ZHANG JIACHENG & LIN ZHIGUANG 1992, ZOU JINSHANG 1987).
Die Untersuchungsräume im Norden Pakistans sind über ihre randmonsunalen Sommer-niederschläge an das Klimageschehen der Ostseite Eurasiens angeschlossen. Im Regen-schatten des Himalaja fällt der größere Anteil der Niederschläge jedoch im Winter und Frühjahr, hervorgerufen durch Störungen aus dem Mittelmeerraum (Abb. 18). Somit verfügen der Karakorum und Teile des inneren Himalaja über relativ feuchte Winter (REIMERS 1992, WEIERS 1995).
Bedingt durch die Wärmerückstrahlung der Gebirgsmassen Tibets steigt die Temperatur auf dem Plateau deutlich über die der freien Atmosphäre in gleicher Höhe über dem Tiefland. So ist die Jahresmitteltemperatur (JMT) Lhasas im Vergleich zu Städten auf gleichem Breitengrad im tief gelegenen Osten Chinas um mehr als 10 °C erhöht (YU
XIAOGAN & SUN SHANGZHI 1981). Dieses Wärmeplus wirkt sich auf die Höhengrenzen der Vegetation und geomorphologische Formungsprozesse aus. Sowohl natürliche Vegetations-einheiten als auch Kulturpflanzen sind zum Plateau hin in höheren Lagen zu finden (DICKORÉ 1995, HAFFNER 1997, HARTMANN 1987, MIEHE 1984, YU XIAOGAN & SUN
SHANGZHI 1981). Im Bereich der meridionalen Stromfurchen findet man die wohl höchst-gelegenen Waldgrenzen der Erde. Wälder von Picea balfouriana kommen an Nordhängen noch in 4400 m Höhe, Bäume von Sabina tibetica auf den Sonnenhängen gar in 4600 m
Höhe vor (LI WENHUA 1993, ZHENG DU & YANG QINYE 1985, ZHENG DU et al. 1981). Einen großräumigen Eindruck hiervon vermitteln die Karten der Waldgrenzen und der Schneegrenzhöhe in der Arbeit von WISSMANN (1961). LEHMKUHL & ROST (1993) gehen von einen letztglazialen und rezenten Anstieg der Schneegrenze zum Tibetanischen Hochland hin um etwa 100 m pro Breitengrad aus.
5.3 Vegetation
Alle Untersuchungsgebiete liegen in der Holarktis. Kennzeichnende Familien dieses Floren-reiches sind u. a. die Pinaceen, Betulaceen, Salicaceen und Ranunculaceen. Die in den Hochgebirgswäldern Chinas, des Himalaja und des Karakorum so wichtigen Gattungen der Pinaceen Picea, Abies, Larix und Pinus sind auf die Holarktis beschränkt. Ebenso Juniperus communis und die Gruppe Caltha palustris agg.
Das Tibetanische Plateau kann als Teil der Zentralasiatischen Florenregion oder auch als eigenständiges Gebiet aufgefaßt werden. Der Gebirgszug des Qinling erstreckt sich dagegen in die Sino-Japonische Florenregion Ostchinas. Diese Region zieht sich in einem schmalen Korridor entlang der Südseite des Himalaja bis in den Karakorum. In der Berg- und Hochgebirgsstufe überwiegen jedoch die holarktischen Arten, und die Endemiten des Westhimalaja zeigen verwandtschaftliche Beziehungen zu Sippen der Holarktis. Am Südfuß des Himalaja beginnt die Paläotropis Indiens. Der größte Teil des Subkontinents gehört zur

5. Der Untersuchungsraum – Vegetation 33
Indischen Florenregion. Der Nordwesten Indiens bildet mit den Wüsten Nordafrikas die Saharo-Sindische Region (ENGLER 1964, MEUSEL & SCHUBERT 1971c, SCHICKHOFF 1993, WALTER & STRAKA 1970).
Mit seinen borealen Nadelwäldern im Nordosten des Landes hat China Anschluß an die Taiga Eurasiens. Nach Süden hin folgt eine Zone sommergrüner Laubwälder, die südlich des Qinling unter Hinzutreten immergrüner Baumarten in eine subtropische Laubwaldzone übergeht. Der äußerste Süden des Landes wird von tropischen Regen- und Monsunwäldern eingenommen (CHEN CUNGEN 1987, HE DAZHANG & DONG HE 1988, HOU HSIOHYU 1983)
Die den Norden der Äußeren Mongolei erreichende Taiga (Larix sibirica, Pinus cembra ssp. sibirica) geht mit einem Waldsteppengürtel in Steppen und Halbwüsten über. Ein Laubwaldgürtel fehlt. Innerhalb des Übergangsgebietes vom Wald zur Steppe sind allein Buschwälder von Ulmus pumila, zumeist auf steinigen südexponierten Steilhängen, gelegentlich anzutreffen. Den Süden des Landes nehmen die steinigen Flächen der Gobi ein, die auf dem Alaschan Plateau der Inneren Mongolei von Sandwüsten abgelöst werden (BARTHEL 1990, HILBIG 1995, HILBIG et al. 1984, HILBIG & KNAPP 1983, WALTER 1968, 1974).
Wo am Nordostrand des Tibetanischen Plateaus mit der Höhe die Niederschläge zuneh-men, treten Nadelwälder an die Stelle der trockenen innerasiatischen Vegetation. In den Hochgebirgswäldern Chinas herrschen im allgemeinen Arten der Gattungen Abies und Picea vor. Darüber kann sich ein subalpiner Larix-Gürtel anschließen. Pinus und Tsuga sind auf tiefere Lagen beschränkt und dringen zumeist nur in den unteren Bereich der Tannen-Fichten-Wälder vor. Unter den trocken-kalten Bedingungen des Hochlandes nördlich vom Huang He fehlen Tannen. Die üppig entwickelten Nadelwälder im Bereich der meridionalen Stromfurchen gehören der Sino-Himalajischen Florenregion an. Ein charakteristisches Geoelement dieser Region ist die Gattung Rhododendron, die mit ca. 60 % ihrer weltweit rund 850 bekannten Arten in den Gebirgsräumen Südwestchinas, Burmas und des östlichen Himalaja ihr Verbreitungszentrum hat.
Das Gebiet um Lhasa liegt im Übergangsbereich zur Tibetanischen Florenregion, die durch Arten wie Stipa purpurea, Ceratoides compacta, Sophora moorcroftiana, Kobresia pygmaea und Carex moorcroftii (ZHENG DU 1983) von der Florenregion Zentralasiens abgegrenzt werden kann. Westlich der Wälder der meridionalen Stromfurchen folgen alpine Matten, die im wesentlichen von Arten der Gattung Kobresia gebildet werden. Diese weichen zum Inneren des Plateaus hin bei Niederschlägen von weniger als 400 mm vor Stipa
purpurea-Steppen zurück, die ihrerseits bei weniger als 100 mm Niederschlag in eine wüstenhafte Vegetation mit Carex moorcroftii und Arten von Ceratoides übergehen und zusammen mit dieser den größten Teil des abflußlosen Qiangtang-Plateaus einnehmen (CHANG D. H. S. 1981, 1983, CHEN WEILIE 1981, FANG RUIZHENG & MING TIANLU 1981,

5. Der Untersuchungsraum – Vegetation 34
HUANG RONGFU 1987, LI MINGSEN 1981, LI WENHUA 1993, LI WENHUA et al. 1981, WANG
JINTING 1981, ZHENG DU 1983, 1985, ZHENG DU et al. 1981).
Vom Durchbruchstal des Yarlung Zangbo im Südosten bis zu dem des Indus im Nord-westen erstrecken sich die Gebirgszüge des Himalaja über eine Entfernung von rund 2500 km. Der 7000 bis 8000 m hohe Hauptkamm des Zentralen Himalaja trennt die in 5000 m Höhe gelegenen trockenen, alpinen Steppen Tibets von den tropischen Regen-wäldern der Südseite. Die Niederschlagssummen nehmen auf der Himalaja-Südseite von über 4500 mm im Osten auf unter 400 mm im Westen ab. Am Gebirgsfuß treten dabei an die Stelle des immergrünen tropischen Regenwaldes (Dipterocarpus turbiantus, Mesua ferrea, Cycas pectinata) nach Westen zu Dipterocarpaceen-Wälder mit der laubwerfenden Shorea robusta.
Am nordwestlichen Himalaja werden das Vorland und die unteren Gebirgshänge von trockenheitsresistenten Pflanzengesellschaften eingenommen. Der geringe monsunale Ein-fluß und der zunehmende Anteil der Frühjahrs- und Winterniederschläge schaffen Klima-bedingungen, unter denen Arten aus dem Verwandtschaftskreis der mediterranen Flora gedeihen. Quercus baloot zeigt morphologische Verwandtschaftsbeziehungen zur iberischen Q. rotundifolia und stellt eine konvergente Entwicklung zu Q. ilex dar. Cedrus deodara vertritt im westlichen Himalaja eine Gattung, deren übrige Arten in Gebirgen des Mittelmee-raumes wachsen. Die in ihrem Artenbestand mediterran getönten Wälder von C. deodara und Q. baloot gedeihen dort, wo der Anteil der Sommerniederschläge am Jahresniederschlag nicht mehr als 40 % beträgt. Die östlichsten Vorkommen solcher Wälder liegen luvseitig zum Sommermonsun im Chenab-Tal östlich des Kaschmir-Beckens. Die bis nach Bhutan hinein auftretende himalajische Pinus roxburghii markiert den äußersten östlichen Rand des mediterranen Einflußbereiches. Sie kommt unter vergleichbaren sommerregenreichen, semihumiden bis semiariden Bedingungen vor, wie die ihr nahe verwandte P. canariensis auf den Kanarischen Inseln.
Die Baum- und Strauchvegetation der trockenen Talstufen des westlichen Himalaja enthält Arten aus dem Mittelmeerraum wie z. B. Punica granatum und Capparis spinosa. Oleander, Ölbaum und Pistazie sind durch Nerium odorum, Olea ferruginea, O. cuspidata, Pistacia khinjuk und Pistacia mutica vertreten. Gemeinsam mit ihnen kommen Akazien und Euphorbien vor. Nicht wenige der trockenheitsliebenden Sippen treten auch im Südosten Chinas auf oder besitzen dort verwandte Arten. Ihr Vorkommen ist mit Annäherung an den Monsun aber auf die Talböden der tief eingeschnittenen trockenheißen Täler begrenzt. Oft ist im feuchten Osthimalaja eine Disjunktion ihrer Areale zu beobachten. Zu dem genannten Sippeninventar tritt am Südostrand des Tibetanischen Plateaus noch Opuntia hinzu. Das Knie des Huang He zwischen Nianbaoyeze und dem Becken von Zoige markiert die Nordgrenze der klimatischen Trockentäler (HANDEL-MAZZETTI 1936, HOU HSIOHYU 1983, MANI 1978, MEHER-HOMJI 1973, MEUSEL & SCHUBERT 1971a, b, c, MIEHE 1984, NEGI 1991, SCHICKHOFF 1993, SCHWEINFURTH 1956, 1957, 1986, SCHWEINFURTH & CHEN WEILIE 1984, THOMAS 1994, TROLL 1967, WU ZHENGYI et al. 1981, ZHANG JINGWEI et al. 1981).

5. Der Untersuchungsraum – Vegetations- und Florengeschichte 35
5.4 Vegetations- und Florengeschichte
In der Kreidezeit herrschten im Nordosten Chinas unter gemäßigtem bis subtropischem Klima koniferenreiche Wälder mit Arten der Gattungen Glyptostrobus, Metasequoia, Sequoia und auch Thuja vor. In der späten Kreide traten mit den Lauraceen, Aquifoliaceen, den immergrünen Fagaceen, Palmen, Myrtaceen und Sapotaceen subtropische Laubbäume hinzu. Im südlichen China war das Klima trocken-subtropisch, und Laubbäume wie Quercus, Cercidiphyllum, Ficus, Eucalyptus und Aralia dominierten (GUO SHUANGXING
1981, HSÜ JEN 1983).
Außer im trockenen Nordwesten (Tarim-Becken, Qinghai und West-Gansu) konnten im Paläogen in ganz China subtropische Elemente gedeihen. Im östlichen Nordchina waren neben vorherrschenden laubwerfenden Gattungen wie Carpinus und Alnus immergrüne Arten von Quercus, Dryophyllum sowie Koniferen der Gattungen Sequoia, Metasequoia, Glyptostrobus, Taxodium und Torreya verbreitet. Im Süden wurden die Wälder meist von immergrünen Lauraceen (Cinnamomum, Laurus), Fagaceen (Quercus, Cyclobalanopsis, Castanopsis, Lithocarpus) und Palmen gebildet. Auftreten von Ephedra und Schizaea weisen dabei auf trockene Verhältnisse hin (HSÜ JEN 1983). Aus vermutlich eozänen Kohlen von der Südseite des Gangdise Shan wird eine tropische bis subtropische Flora mit Arten von Ficus, Eucalyptus, Eugenia, Myrica, Phrynium, Cassia und Celastrus beschrieben (KONG ZHAOCHEN & DU NAIQIU 1981, SONG ZHICHEN & LIU GENGWU 1981, TAO JUNRONG
1981, XU REN 1981). Die im Bereich der Ophiolith-Zone des Yarlung Zangbo-Tales anstehende Liuqu-Flora aus dem späten Eozän oder frühen Oligozän setzt sich überwiegend aus immergrünen Vertretern der Palmen, Lauraceen, Magnoliaceen, Myrtaceen, Annona-ceen, Moraceen, Tiliaceen, Myricaceen, Fabaceen, Sabiaceen und Rutaceen zusammen, was einen tropischen Regenwald unter heiß-humidem Klima vermuten läßt (GUO SHUANGXING
1988, TAO JUNRONG 1988). Im Oligozän standen in den Gebirgen um das Qaidam-Becken Wälder mit Abies, Picea, Pinus, Cedrus, Betula und Quercus. In den tieferen Lagen wuchsen Magnoliaceen und Proteaceen. Der Unterwuchs wurde von Asteraceen, Brassicaceen und Poaceen gebildet (HSÜ JEN 1983).
Im Neogen nahm die Trockenheit des Qaidam-Beckens weiter zu. In Qinghai und Gansu entwickelten sich allmählich Halbwüsten. Nordwestchina besaß neben gemäßigten Wäldern vor allem Steppen, Halbwüsten und Wüsten. Der Osten war nördlich des Qinling gemäßigt bis subtropisch mit Wäldern und Steppen. Südlich des Qinling und in Tibet wuchsen immer- und sommergrüne subtropische Arten (HSÜ JEN 1983). Mit der zunehmenden Abkühlung im Miozän bildeten sich in den Gebirgen Nordwestchinas Nadelwälder von Abies, Picea, Larix, Pinus, Tsuga und Juniperus heraus. Darunter folgte ein Laubwaldgürtel mit Betula, Ulmus, Salix, Populus, Juglans, Alnus, Corylus und Tilia. Vereinzelt kamen auch Cedrus und Castanea vor. Die weiten Ebenen trugen Steppen, die von Artemisia, Ephedra und Chenopodiaceen beherrscht wurden. In die Steppen des Qaidam-Beckens (Chenopodiaceen, Polygonaceen, Cyperaceen, Ephedra) wanderten aus dem mediterranen Bereich Tamarica-ceen und Zygophyllaceen (u. a. Nitraria) ein. Im Süden des Plateaus wuchsen Arten der

5. Der Untersuchungsraum – Vegetations- und Florengeschichte 36
Gattungen Cedrus, Tsuga, Juniperus, Podocarpus, Juglans, Ulmus, Quercus, Carpinus, Betula, Populus, Salix, Rhododendron, Thermopsis und Sophora (HSÜ JEN 1983, WANG
XIANZENG 1988, XU REN 1981). Tsuga, Melia, Carya, Ilex, Quercus und Lithocarpus waren dort zu Beginn des Pliozäns ebenso verbreitet wie im Kunlun Shan. Die im Transhimalaja sowie nördlich von Gangdise Shan und Nyainqentanglha Shan entwickelten Wälder mit Cedrus und immergrünen hartlaubigen Eichenarten wie Quercus semecarpifolia, Q. pannosa und Q. senescens gingen ab dem späten Pliozän zurück. An ihre Stelle traten sommergrüne Laubgehölze und Kräuter, die im späten Pleistozän einer alpinen Strauchsteppe Platz machten. Noch im späten Pliozän waren Abies-Picea-Wälder im Kunlun Shan weit ver-breitet (GUO SHUANGXING 1985, HSÜ JEN 1983, KONG ZHAOCHEN & DU NAIQIU 1981, SHEN
CAIMING & TANG LINGYU 1996, XU REN 1981).
Vor ca. 2,5 Mio. Jahren zeichnete sich nach den palynologischen Archiven des Qaidam-Beckens (SHEN ZHENSHU et al. 1992) bei einsetzender Lößbildung mit dem Übergang zu einem trocken-kalten Klima der Beginn des Quar tärs ab (FORT 1996, KUKLA & AN
ZHISHENG 1989, LIU TUNGSHENG & DING ZHONGLI 1993). Die Trockenräume im Nordwesten Chinas dehnten sich nach Osten aus. Von Norden her drang die Taiga vor, und sommergrüne Arten gelangten weiter nach Süden (HSÜ JEN 1983). Zwar wurde in den Kaltphasen das Tibetanische Plateau von einer offenen Steppenvegetation mit Vertretern von Artemisia, Chenopodiaceen und Asteraceen eingenommen, doch kennt man aus warmen Abschnitten des frühen Pleistozäns selbst Nachweise subtropischer Bäume wie z. B. von Cedrus. In dieser Zeit verlief die Vegetationszonierung Chinas weitgehend breitenparallel. Noch im mittleren Pleistozän zog sich eine Zone interglazialer Laub- und Nadelwälder von der Küste Ostchinas bis in den Quellbereich des Yarlung Zangbo, doch begann das Plateau sich allmählich als eigenständige vegetationsgeographische Einheit vom übrigen China abzugrenzen. Im späten Pleistozän und Holozän vollzog sich dann der Wandel zur typischen alpinen Matten- und Strauch-Vegetation Tibets (KONG ZHAOCHEN & DU NAIQIU 1981, TONG
GUOBANG & ZHANG JUNPAI 1991).
Die gegen Ende des Pleistozäns einsetzende Erwärmung führte im mittleren Holozän zu einem Klimaoptimum. Das späte Holozän begann mit einer deutlichen Klima-verschlechterung, gefolgt von Schwankungen bis hin zur Gegenwart. Diese Gliederung ergibt sich sowohl aus der Vegetationsentwicklung als auch der Geschichte von Gletschern und Seen. Der Beginn des Holozäns kann nach bisherigen Vorstellungen um 10000 BP angesetzt werden. Etwa zu dieser Zeit begann in Tibet auch die Moorentwicklung. Das Alter des mittleren Abschnitts beläuft sich ungefähr auf 7500 BP bis 3000/2500 BP. Die ab-schmelzenden Gletscher ließen Seestände ansteigen, und die allgemein größere Feuchte und die höheren Temperaturen führten zu verstärkter Bodenbildung und zur Ausbreitung anspruchsvoller Baumarten. Das eigentliche Klimaoptimum endete gegen 5000 bis 4000 BP, danach folgte ein etwas ungünstigerer Zeitraum. Den Beginn des späten Holozäns markieren Vegetationsänderungen, deutliche Gletschervorstöße und die Überlagerung mancher Torfe durch minerogenes Material (HUANG CIXUAN & LIANG YULIAN 1981, JARVIS 1993, QI
HUANG et al. 1981, SHI YAFENG et al. 1993, SUN XIANGJUN & CHEN YINSHUO 1991a, WANG
FUBAO & FAN 1987, WINKLER & WANG PAO K. 1993, ZHOU S. Z. et al. 1991).

5. Der Untersuchungsraum – Siedlungsgeschichte 37
5.5 Siedlungsgeschichte
5.5.1 China
Zu den ältesten in China gefundenen menschlichen Knochen gehören die des Homo erectus aus der Provinz Yunnan (Abb. 2). Sie besitzen ein Alter von etwa 1,7 Mio. Jahren und wurden als Fossilien des Yuanmou-Menschen bekannt. Ein ähnliches Alter weisen Funde aus der Provinz Sichuan auf. Aus dem nördlichen China sind 1,2 Mio. Jahre alte Steinartefakte sowie Knochen mit Brandspuren aus der Nähe der unmittelbar nördlich des Qinling gelegenen Stadt Xian bekannt geworden. Mächtige Aschelagen in einer der Höhlen von Zhoukoudian, der Fundstätte des Peking-Menschen, stammen aus der Zeit zwischen 0,5 und 0,3 Mio. Jahren v. H. und belegen die ausgedehnte Nutzung des Feuers durch den H. erectus (BOSINSKI 1992, JIA LANPO & HUANG WEIWEN 1991, LIU ZECHUN 1990, ZHOU
GUOXING 1988).
Die ältesten Nachweise des Homo sapiens aus den Provinzen Shanxi und Gansu werden in eine Warmphase zwischen 50.000 und 30.000 BP gestellt. Aber auch aus der Zeit des späten Paläolithikums (ab 35000 BP), in den die Kaltphase des letztglazialen Maximums fällt, liegen aus Nordchina zahlreiche Nachweise des Menschen vor. So etwa von Zhoukoudian, vom Lößplateau und vom Südrand des Tarim-Beckens, einer der späteren Routen der „Seidenstraße“ (HUANG WEIWEN 1990, LIU ZECHUN 1990, QI GUOQIN 1989, WU
XINZHI 1988, YOU YUZHU 1985).
Wenn im Süden Chinas auch schon zwischen 8500 - 8000 BP der Anbau von Taro und Cucumber neben der Jagd zum Lebensunterhalt beitrug, so wird doch der Beginn des Neolithikums mit dem Einsetzten des Trockenfeldbaus der Dadiwan-Kultur um 8000 BP gleichgesetzt (ZHAO SONGQIAO 1994). Diese Kultur war auf dem Lößplateau im Einzugsgebiet des Wei He in der Provinz Gansu angesiedelt und scheint in China der Ausgangspunkt des Ackerbaus gewesen zu sein. Zugehörige Fundschichten sind auf 7800 bis 7355 BP datiert (FENG SHENGWU 1985).
Die Nutzung von Reis hat ihren Ursprung südlich des unteren Yangzi. Dort finden sich die ältesten Nachweise dieses Getreides in den Schichten der um 7000 BP bestehenden Kulturen von Homudu (Hemudu) und Lujiao, südlich von Shanghai (SUN XIANGJUN et al. 1981, WU WEITANG 1985). Aus einer mehrjährigen Wildform gingen verschiedene Sorten von einjährigem Kulturreis hervor (OKA 1988).
Vor etwa 5000 - 6000 Jahre begann die Yangshao-Kultur. Sie war besonders am mit-tleren Huang He verbreitet, und die früheste Kultur auf nordchinesischem Boden, deren Träger die Herstellung von Tongefäßen kannten. Geometrische Figuren und stark stilisierte Vögel- und Fischmotive in roten sowie schwarzen Farben zieren Teile der Töpferwaren. Werkzeuge wurden aus Steinsplittern und geschliffenen Steinen gefertigt.
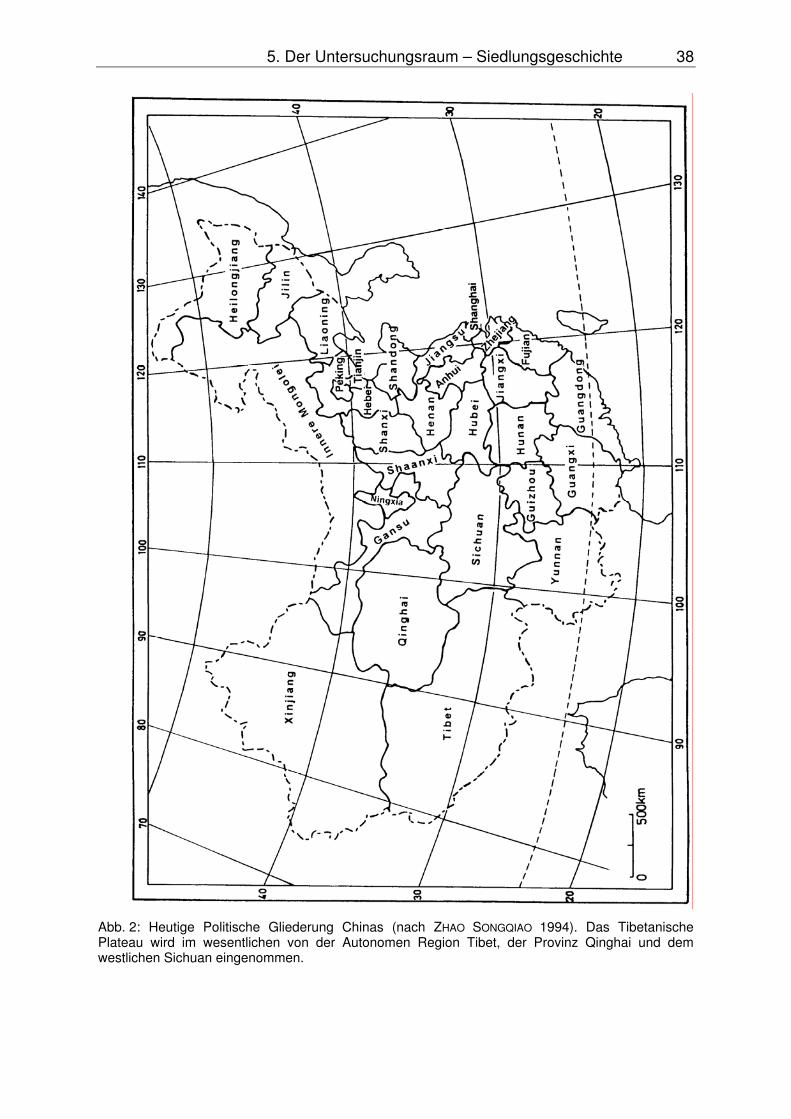
5. Der Untersuchungsraum – Siedlungsgeschichte 38
Abb. 2: Heutige Politische Gliederung Chinas (nach ZHAO SONGQIAO 1994). Das Tibetanische Plateau wird im wesentlichen von der Autonomen Region Tibet, der Provinz Qinghai und dem westlichen Sichuan eingenommen.

5. Der Untersuchungsraum – Siedlungsgeschichte 39
Ein typisches Dorf lag unweit des heutigen Xian. Die Häuser besaßen einen vertieften
Boden und waren von Lehmmauern umgeben. Das Strohdach wurde von Holzpfeilern getragen. Hirse war das Hauptgetreide. Hunde- und Schweinezucht dienten der Fleischproduktion.
Die Longshan-Kultur, die in Henan auf die Yangshao-Kultur folgte, anderenorts aber wohl möglich zeitgleich zu dieser bestand, ist durch äußerst dünnwandige, schwarze Ton-waren mit rechteckigem Profil gekennzeichnet, und verfügte im Gegensatz zur Yangshao-Kultur über die Töpferscheibe. Die Dörfer waren von einem Schutzwall aus gestampftem Lehm umgeben. Zu den Haustieren zählten Schweine, Schafe und Rinder. Die Form der Töpferwaren, die Technik des Lehmstampfens und das Wahrsagen mit Hilfe von ins Feuer geworfenen Knochen deuten Beziehungen zur Shang-Dynastie der Bronzezeit an.
Neben den zwei erwähnten Kulturen gibt es eine ganze Reihe weiterer, weniger unter-suchter Formenkreise. Außer Hirse und Reis sind aus dem Neolithikum auch die Nutzung von Weizen, Gerste und Maulbeere nachgewiesen. Zudem war die Weinherstellung bekannt. Stoffe fertigte man aus Hanf, aber auch Hinweise auf Seide liegen bereits vor. Die Tierhaltung umfaßte Rinder, Schafe, Ziegen, Hunde, Schweine, und Hühner, später auch Pferde (DEXEL 1973, GERNET 1994, HILDEBRAND 1990, ZHAO SONGQIAO 1994).
Die erste Staatsgründung in China vollzog sich vermutlich in der Xia-Dynastie (ca. 2206 - 1766 v. Chr.), deren Einflußbereich das östliche Lößplateau und das nordwestliche Henan umfaßte. Bereits in dieser Zeit begann man mit Baumaßnahmen zur Flutkontrolle und der Herstellung von Werkzeugen aus Bronze. Der eigentliche Beginn der Bronzezeit und des gut organisierten Staatswesens ist aber wohl die Shang-Dynastie (1766 - 1126 v. Chr.), aus der eine Vielzahl bronzener Gegenstände – darunter Waffen, Kultgefäße, Wagenteile und Pferdegeschirre – bekannt sind. Die älteste Form der chinesischen Schrift kennt man von den Knochen und Schildkrötenpanzern dieser Zeit, die einem neolithischen Brauch folgend, zu Orakelzwecken dem Feuer ausgesetzt wurden (GERNET 1994, ZHAO SONGQIAO
1994). Manche Belege sprechen für Beziehungen zwischen dem China der Shang-Dynastie bis in Gebiete im heutigen Iran (BRENTJES 1984).
Die Herrscher der nachfolgenden Westlichen Zhou-Dynastie (1126 - 771 v. Chr.) stammten vom oberen und mittleren Wei He. Ihre Hauptstadt lag südwestlich von Xian. Bewässerung und Düngung verbesserten den Ackerbau. Bohnen, Melonen sowie Soja-bohnen erweiterten das Nahrungsangebot, und der Reisanbau erreichte von Süden kommend den Huang He (GERNET 1994, WU WEITANG 1985, ZHAO SONGQIAO 1994). Nach mehreren Einfällen nördlicher Völkerschaften floh die Regierung gen Osten nach Luoyang (Provinz Henan), wo sie die Östliche Zhou-Dynastie gründete, die bis 249 v. Chr. bestand. In der „Frühlings und Herbst Periode“ (771 - 403 v. Chr.) zerfiel das Land in 140 kleine Fürstentümer. Darauf folgte die Epoche der „Kämpfenden Staaten“ (403 bis 221 v. Chr.). Der Ackerbau dehnte sich auf den Bereich zwischen 25°N und 40°N aus. Das Eisen ist in China seit dem 6. Jh. bekannt, die erste Erwähnung des Gießens stammt aus dem Jahr 513 v. Chr. (GERNET 1994, WAGNER 1993, ZHAO SONGQIAO 1994).

5. Der Untersuchungsraum – Siedlungsgeschichte 40
Qin Shi Huang Di einigte das Land in der Qin-Dynastie (221 - 206 v. Chr.), dehnte den Herrschaftsbereich im subtopischen Süden sowie in den trockenen Gebieten westlich des Lößplateaus aus und ernannte sich selbst zum ersten Kaiser Chinas. Maße, Gewichte und Schrift wurden vereinheitlicht, die Bevölkerung entwaffnet und Bücher verbrannt. Die Hauptstadt Xianyang lag beim heutigen Xian und hatte bereits 1.000.000 Einwohner. In ihr führten die unter hohem Aufwand erbauten Überlandstraßen zusammen. Die hohen Arbeitsbelastungen des Volkes beim Bau des kaiserlichen Mausoleums mit seinen aufwen-digen Grabbeigaben (7000 Terrakottasoldaten), der Paläste, der Überlandstraßen und der Chinesischen Mauer scheinen zu Aufständen geführt zu haben, die zum Ende der Qin-Dynastie beitrugen (GERNET 1994, ZHAO SONGQIAO 1994).
Die Han-Dynastie (206 v. Chr. - 9 n. Chr.) hatte ihren Sitz zunächst in Xian. Es war eine Zeit der Expansion. Mehrere Kriegszüge führten in die Mongolei gegen die Hunnen (Xiong Nu). Deren Vertreibung aus dem Gansu-Korridor eröffnete den Weg in das Tarim-Becken und darüber hinaus in den Pamir und das Ferghana-Becken. Damit war es möglich, entlang der später als „Seidenstraße“ bekannt gewordenen Karawanenwege Handel zu betreiben und politische Kontakte zu knüpfen. Auf die anschließende Xin-Dynastie (9 - 25 n. Chr.) folgte die Späte oder Östliche Han-Dynastie (25 - 220 n. Chr.). Die nördlichen Grenzgebiete wurden gemeinsam von Hunnen und anderen Stämmen angegriffen. Innenpolitische Probleme und Bauernaufstände führten in eine Zeit voller Kriege (220 -280 n. Chr.) in der Millionen von Menschen starben. Das Land zerfiel. Die Wiedervereinigung des Landes in der Westlichen J in-Dynastie (280 - 316 n. Chr.) hatte nur kurzen Bestand und leitete zur Existenz der Östlichen J in-Dynastie und von 16 nördlichen Staaten (316 - 450 n. Chr.) über. Der Norden Chinas wurde von mehreren Stämmen aus den Steppengebieten überrannt. Es folgten zwischen 450 und 589 n. Chr. die Südliche- und die Nördliche-Dynastie (GERNET 1994, YALDIZ 1987, ZHAO SONGQIAO 1994).
Mit der erfolgreichen Wiedervereinigung in der Sui-Dynastie (589 - 618 n. Chr.) be-ginnt das chinesischen Mittelalter. Vorhergehende Unruhen hatten die Bevölkerung so stark dezimiert, daß nun ausreichend Ackerland zur Verfügung stand. Die anschließende Tang-Dynastie (618 - 907 n. Chr.) war zunächst durch militärische und wirtschaftliche Stärke gekennzeichnet. Bewässerungsräder erlaubten die Ausdehnung der Ackerbaugebiete. Die landwirtschaftliche Fläche wurde verdoppelt. In der zweiten Hälfte der Dynastie kam es aber wieder zur kriegerischen Zerteilung des Reiches. Die Tibeter stießen in die Oasen des Tarim-Beckens vor und fielen 763 n. Chr. nach Gansu und Shenxi ein, wo sie die chinesische Hauptstadt plünderten. In der Nördlichen Song-Dynastie (960 - 1126 n. Chr.) trat zwar Ruhe ein, doch gingen weite Teile der Grenzgebiete verloren. Im Süden Chinas herrschte später die Südliche Song-Dynastie (1129 - 1279 n. Chr.). Während dieser Zeit wurde besonders in der Küstenregion und im Bergland Ackerland angelegt. Zu den Anbauprodukten zählten zunehmend Baumwolle, Zuckerrohr und Tee (GERNET 1994, STAHLBERG 1996, ZHAO SONGQIAO 1994).

5. Der Untersuchungsraum – Siedlungsgeschichte 41
Im späten 12. Jh. hatte Temujin die Mongolen vereint und erhielt 1206 n. Chr. den Titel Tschingis Khan. Sein Mongolen-Reich (1206 - 1368 n. Chr.) umfaßte bereits 1215 n. Chr. weite Teile Chinas nördlich des Huang He und dehnte sich in den folgenden 10 Jahren über Zentralasien bis zum heutigen Iran und ins nordwestliche Indien aus. Kublai Khan (1260 - 1294 n. Chr.), Sohn Tschingis Khans, benannte das Reich in Yuan-Dynastie um und verlegte den Hauptsitz aus der Inneren Mongolei nach Dadu, dem heutigen Peking. Seine nomadisch geprägten Generäle ließen Äcker in Weideland überführen. Nach dem Tode Kublai Khans nahmen Ackerbau sowie Industrie wieder zu (GERNET 1994, HAMBLY 1995, ZHAO SONGQIAO 1994).
Auf die mongolische Yuan-Dynastie folgte die Ming-Dynastie (1368 - 1644 n. Chr.). Zu ihrem Herrschaftsbereich gehörten neben den subtropischen Teilen des heutigen Chinas und den Gebieten im trockenen Nordwesten auch die Randgebiete des Tibetanischen Plateaus. Die von Nordosten kommenden Mandschu eroberten China und errichteten die Qing-Dynastie (1644 - 1911 n. Chr.). Ihr Herrschaftsbereich umfaßte die Mongolei, Xinjiang, Tibet und Taiwan. Die erhobenen Bevölkerungszahlen stiegen von 44 Mio. im Jahr 1753 auf 380 Mio. im Jahr 1887. Auf die Republik von China (1911 - 1949) folgte die heutige Volksrepublik China.
5.5.2 Tibet
Eine Zusammenstellung prähistorischer Funde aus Tibet gibt HUANG WEIWEN (1994). Danach werden die ältesten der als menschliche Werkzeuge eingestuften Steinfunde - die Unterscheidung zwischen Artefakten und natürlichen Verwitterungserscheinungen ist nicht immer unproblematisch - in das mittlere Paläolithikum gestellt. Für drei Fundorte liegen mit Radiokarbondatierungen von 30000, 14000 und 11000 BP Alter vor, die sie ins Spätpaläolithikum stellen. Sie liegen in vergleichsweise geringen Höhen von nur 3500 m und tiefer, und zwar im und am Qaidam-Becken sowie am Qinghai Hu. Die beiden Fundplätze jüngeren Alters deuten auf eine frühe postglaziale Besiedlung Tibets aus Richtung des Lößplateaus über den Kunlun Paß entlang der Route der heutigen Autostraße hin. Daneben gibt es aber auch Hinweise für eine Besiedlung von Süden her (CUI ZHIJIU et al. 1995, HUANG WEIWEN 1994).
Von den zahlreichen Fundstätten des Mesolithikums und Neolithikums liegen nur wenige 14C - Datierungen vor. Eine Siedlung in 2580 m Höhe in Qinghai am Huang He besitzt ein Alter von 6745 ± 85 BP. Am Kunlun Paß wurde eine Feuerstelle in 4040 m Höhe auf 3500 BP datiert. Nahe Qamdo (3100 m) ergaben sich für Steinwerkzeuge, Tonwaren und Tierknochen Altersbestimmungen zwischen etwa 5000 und 3900 BP. Während bei Qamdo Gerste nachgewiesen werden konnte, stammen die zahlreichen Knochenfunde, ebenso wie die vom Huang He, noch von Wildtieren (HUANG WEIWEN 1994, LI BINGYUAN et al. 1985, YU XIAOGAN & SUN SHANGZHI 1981). Die höchsten Fundplätze in 5200 m Höhe werden in das frühe bis mittlere Holozän gestellt und als Hinweis auf günstige Klimabedingungen gesehen. Die dort gefundenen Werkzeuge

5. Der Untersuchungsraum – Siedlungsgeschichte 42
weisen sie als Jagdlager aus (LI BINGYUAN et al. 1985, WANG FUBAO & FAN 1987, WU
XIANGDING & LIN ZHENYAO 1988).
Nur 5 km südlich von Lhasa fand man in situ Töpferwaren, Steingeräte und Tierknochen des späten Neolithikums (HUANG WEIWEN 1994). Zur Zeit der Han-Dynastie, im 2. Jh. n. Chr., wurde nahe Zetang in 3400 m Höhe auf einer Terrasse des Yarlung Zangbo Gerste angebaut (CUI ZHIJIU et al. 1995). Aus schriftlichen Quellen ist bekannt, daß zu Beginn des 6. Jh. n. Chr. die Pflüge bei der Feldbestellung von zwei Yaks gezogen wurden, und daß ein Bewässerungsystem bestand (ZHANG RONGZU 1989).
Bereits während der Han-Dynastie (206 v. Chr. - 220 n. Chr.) nahmen die Tibeter Vor-stöße in das chinesische Kernland vor. In einem Gegenfeldzug zwischen 60 - 62 n. Chr. unterwarf die Han-Dynastie den Nordosten Tibets und legte im Bereich zwischen Huang He und Qinghai Hu Militärkolonien an. Im Gansu-Korridor, dem Zugang zum Tarim-Becken, und den Tälern am Nordostrand des Tibetanischen Plateaus betrieben die Chinesen Ackerbau.
Im Jahr 629 n. Chr. gründete Songtsen Gampo (620 - um 649 n. Chr.) die erste tibetische Monarchie mit Sitz in Lhasa. Wohl spätestens seit dieser Zeit wird bei Lhasa Ackerbau betrieben. Aus dem von ihm unterstützten Buddhismus und der ansässigen Bonreligion entwickelte sich im Laufe der Jahrhunderte der Lamaismus. Das erste buddhistische Kloster wurde um 779 n. Chr. gegründet. Erst unter Trisong Detsen (um 754 - um 797 n. Chr.) wird die Quellenlage mit der Schaffung einer tibetischen Schrift verbessert (HAMBLY 1995, HERMANNS 1949, ZHAO SONGQIAO 1994).
Auf die in Zentralasien stärker werdende tibetische Macht reagierten die Chinesen ab 670 n. Chr. mit Gegenoffensiven. 694 n. Chr. konnten sie die Tibeter zunächst schlagen, doch folgten schon 700 n. Chr. Einfälle der Tibeter nach Gansu. Weitere Militäraktionen beider Seiten folgten. 763 n. Chr. plünderten die Tibeter die Hauptstadt der Tang-Dynastie, das heutige Xian. Zu einem Friedensschluß kam es 822 n. Chr. Mit der Ermordung des Herrschers Langdarma (838-842 n. Chr.) begann der Machtniedergang Tibets. Ihr Einfluß im Gansu-Korridor ging verloren und der Süden Tibets zerfiel in eine Anzahl feudaler Fürstentümer. Im Nordosten kam es im 11. Jh. zur Gründung des Reiches Si-hia. Dessen Macht reichte bis in das Ordos-Gebiet im großen Bogen des Huang He. Doch bereits 1209 fielen die Mongolen in das Hochland ein. 1226 kam es zu einem mongolischen Vernichtungsfeldzug und ganz Tibet wurde Teil der Yuan-Dynastie (GERNET 1994, HAMBLY
1995, KWANTEN 1979, YALDIZ 1987).
Während der Ming-Dynastie (1368 - 1644 n. Chr.) errichtete man entlang der Grenze zu Tibet chinesische Militärkolonien. In der Qing-Dynastie (1644 - 1911 n. Chr.) drangen immer neue Siedler aus dem dicht bevölkerten China in die Grenzgebiete ein und brachen Weiden zu Ackerland um.
Seit 1509 zogen Mongolen aus dem unruhigen Ordos-Gebiet zum Qinghai Hu. 1633 errichteten sie hier ein eigenes Reich. Bis 1642 hatten sie Zentraltibet besetzt und die ansässigen Herrscher geschlagen. Das Land wurde darauf hin dem fünften Dalai Lama
(1617 - 1680) geschenkt. Hieraus ging der lamaistische Kirchenstaat Tibet hervor. Nach der

5. Der Untersuchungsraum – Siedlungsgeschichte 43
Eroberung Tibets (1720) gliederte die Qing-Dynastie den seit mindestens 2000 Jahren von Tibetern bewohnten fruchtbaren Südosten des Landes, die Region Kham, an ihre Provinz Sichuan an. Im Jahr 1928 gründete die Republik China die Provinz Qinghai. Nachdem das ursprünglich tibetische Kham vorübergehend den Status einer Provinz (Sikang) erhielt, bildet es heute, wie bereits zur Zeit der Qing-Dynastie, den Westen der Provinz Sichuan (EBERHARD 1948, HERMANNS 1949, GRUSCHKE 1997, WU SHUHUI 1995, 1996).
5.5.3 Karakorum
Im Karakorum kann man vielleicht ähnlich wie im Pamir-Gebiet mit der Anwesenheit des Menschen seit dem Ausklingen der letzten Eiszeit rechnen. Die vom Pamir bis in den Osten Tibets verbreitete Hissar-Kultur bestand schon um 5000 v. Chr. oder früher (zur geogra-phischen Lage vergl. Abb. 18). Ihre Werkzeuge deuten auf die Zerlegung von Jagdbeute hin (BRENTJES 1991, JETTMAR 1993). Felsbilder, die in weiten Teilen Zentralasiens entlang ehemaliger Verkehrswege gefunden werden, treten auch am Indus und im Karakorum auf, wo sie auf ein Eindringen der Okunev-Kultur Südsibiriens seit dem Beginn des 3. Jtsd. v. Chr. hinweisen (JETTMAR 1984, JETTMAR & THEWALT 1985, POHLE 1997).
In Kaschmir und südlich des Himalaja ist das Paläolithikum durch Werkzeugfunde belegt. Hinweise auf den Gebrauch des Feuers reichen bis 10350 BP zurück. In dieser Zeit begann im Westen Indiens das Mesolithikum (10000 - 4000 BP). Vielleicht wurde schon in dieser das Feuer in den trockenen Gebieten zum Abbrennen der harten mehrjährigen Horstgräser benutzt, um bessere Weidemöglichkeiten für Schaf und Ziege zu schaffen, wie dies aus historischer Zeit bekannt ist. Das Neolithikum umfaßt in Kaschmir etwa die Zeit von 6000 - 4000 BP. Das Chalkolithikum setzt gebietsweise schon um 4500 BP ein und dauerte bis 3000 BP an (WHITEHOUSE & WHITEHOUSE 1975).
Töpferwaren aus Afghanistan sind auf etwa 6450 BP datiert. Hinweise auf domestizierte Schafe und Ziegen reichen noch weiter zurück (WHITEHOUSE & WHITEHOUSE 1975). Von den Gebirgsrändern Belutschistans, westlich des Indus im Süden des heutigen Pakistans, kennt man alte Siedlungen mit Häusern aus Schlammziegeln. Zu Beginn des 7. Jtsd. v. Chr. begann hier der Getreideanbau (z. B. Triticum dicoccum). Seit etwa 4000 v. Chr. nahm die Zahl der Siedlungen in Belutschistan und im Indus-Tal allmählich zu. Im frühen 3. Jtsd. v. Chr. entwickelte sich am Indus eine der ersten großen Stadtkulturen der Welt. Nach Beginn der Bronzezeit erreichte sie um 2600 v. Chr. ihre volle Entfaltung. Diese sogenannte Indus-Kultur erstreckte sich über 1500 km von der Mündung des Indus bis an den Sutleji nördlich von Harappa. Die Städte verfügten über ein rechtwinkeliges Straßennetz und staatlich kontrollierte Getreidespeicher (BEAZLEY 1985, DANI 1986, LAMB 1995, MEDER
1979, SARASWAT 1991, VISHNU-MITTRE 1979, WHITEHOUSE & WHITEHOUSE 1975). Der Einfluß der Indus-Kultur reichte über die Gebirge des Karakorum und Hindukusch hinweg bis an den Oxus (Amudarja) im damaligen Baktrien, wo sich eine Kolonie mit einem groß angelegten Kanalsystem befand (BRENTJES 1984, JETTMAR 1993).

5. Der Untersuchungsraum – Siedlungsgeschichte 44
Ab dem 2. Jtsd. v. Chr. wanderten in mehreren Wellen Arier von Nordwesten her in das Stromgebiet des Indus ein. Sie waren vermutlich ein Hirtenvolk, das auch schon den Ackerbau kannte. Die hochentwickelte Indus-Kultur wurde zurückgedrängt, doch bestanden einige ihrer Städte weiter. Nun begann der Anbau der bis dahin unbekannten Getreidesorten Reis und Hirse (BEAZLEY 1985, GLASENAPP 1996, WHITEHOUSE & WHITEHOUSE 1975).
Mit der Ausdehnung der persischen Herrschaft um 550 v. Chr. kam das Gebiet zwischen den Flüssen Jaxartes (heute Syrdarja, mündet in den Aralsee) und Indus zum Archämenidenreich. Zu dessen aktiver Landwirtschaftspolitik gehörte der Bau von Bewäs-serungsanlagen. Mit dem Einfall Alexander des Großen begann in diesem Teil Asiens die Zeit der Makedonen. Die Macht Alexander des Großen erstreckte sich um 325 v. Chr. bis nach Indien hinein. Nach einem Friedensschluß im Jahre 304 v. Chr. fielen die Gebiete südlich des Hindukusch an das indische Maurya-Reich. Nach dessen Niedergang entstanden um 200 v. Chr. griechisch-baktrische Staaten, deren Macht sich von der Küste des Arabischen Meeres bis zum Jaxartes und nach Osten bis in das Tarim-Becken erstreckte (HAMBLY 1995, DANI & BERNARD 1994, JOHNSON 1995).
Die zunehmende Macht der Hunnen, die im 3. Jh. v. Chr. ihren Höhepunkt hatte, veran-laßte nicht nur den Baubeginn der Chinesischen Mauer, sondern auch die Abwanderung mehrerer Stämme aus den Steppengebieten Innerasiens. Diese erreichten gegen 160 v. Chr. den Jaxartes und fielen schon bald in die südlich davon gelegenen griechisch-baktrischen Herrschaftsgebiete ein. Die ursprünglich nomadischen Skythen ließen sich als Saken im Bereich von Nordwestindien bis in das östliche Afghanistan hinein nieder. Auf sie folgte mit den Yüetschi ein weiterer, besser als Tocharer bekannter Nomadenstamm, der sich zunächst im Bereich des Hindukusch festsetzte und mit dem Kuschan-Reich eine der einflußreichsten Mächte in Zentralasien schuf, deren Herrschaftsbereich sich um 75 n. Chr. in die nordindische Ebene bis zum Ganges ausdehnte. Dabei besetzten sie auch das bis dahin tibetische Ladakh. Die vom westlichen Tarim-Becken bis zur Mündung des Indus reichende Machterstreckung der Kuschan bot den Reisenden bei Unruhen auf der „Seidenstraße“ einen sicheren Anschluß an den Seeweg. Eine dieser wichtigen Südverbindungen führte vom Tarim-Becken durch das Hunza-Tal im Karakorum zum Indus.
Nach dem Sturz des letzten Herrschers im Jahr 227 n. Chr. trat an die Stelle der Kuschan das indische Gupta-Reich. In dieses fielen die inzwischen aus der Mongolei vertriebenen Hunnen ab 505 n. Chr. ein, wurden aber schon 20 Jahre später wieder zurückgedrängt (ENOKI et al. 1994, HAMBLY 1995, JOHNSON 1995, PURI 1994).
Nachdem sich das tangzeitliche China und Tibet bereits seit Mitte des 7. Jh. n. Chr. rivalisierende Kämpfe um die Machtstellung in den Gebieten westlich des Tarim-Becken lieferten, kam es angesichts der zunehmenden Bedrohung durch die Araber im frühen 8. Jh. zur chinesischen Invasion nach Chitral, und den Tibetern gelang die Einnahme Gilgits. Doch waren beide Erfolge nur von kurzer Dauer (BURKI 1991, GERNET 1994, MÜLLER-STELLRECHT 1979, 1980, RAUNIG 1978).
Etwa 1221 n. Chr. überquerten die Mongolen auf dem Weg zum Indus den Hindukusch. Der zweite Sohn Tschingis Khans erhielt u. a. den Hindukusch und die südlich daran

5. Der Untersuchungsraum – Ernährung 45
anschließenden Gebiete als Lehen (KWANTEN 1979, HAMBLY 1995). Auf den Untergang dieses Tschaghatai-Lehens im frühen 14. Jh. folgte ein Khanat, dessen Armee in den 30iger Jahren des 15. Jh. bis nach Kaschmir und in die Nähe von Lhasa vordrang (HAMBLY 1995).
Ab 1320 n. Chr. übernahmen die von Norden her eindringenden Rais-Könige die Macht in Gilgit. Dem in Chitral ansässigen Herrscherhaus der Kator gelang es während politischer Unruhen im 17. Jh. seine Macht auf Yasin auszudehnen. Wiederholte Vorstöße von Chitral aus gegen das östlich von Yasin gelegene Gilgit führten im 19. Jh. schließlich zur Besetzung des Gebietes durch Kaschmir und damit zum Einfluß der anglo-indischen Kolonialmacht. 1947 schloß sich die muslemische Bevölkerung des Karakorum an das neugegründete Pakistan an. Hingegen wurde Kaschmir unter seinem hinduistischen Maharaja Teil Indiens (MÜLLER-STELLRECHT 1979, 1980, BURKI 1991).
5.6 Ernährung
5.6.1 Tibet
Die Nahrungsgrundlage der Nomaden Tibets sind das Fleisch ihrer Schafe und zugekauftes Getreide. Von den Yaks schlachten sie nur die alten Tiere. Pferd, Wildhalbesel und Schwein werden nicht verzehrt. Das Fleisch wird halbgar gekocht oder in schmale Streifen zerschnitten an der Luft getrocknet. Zur Wurstherstellung füllt man Därme mit Blut, Fleisch, Fett und Mehl. Gelegentlich erlegt man Hirsch, Reh, Gazelle, Gemse und Moschustier. Vor der Einführung des Gewehres war die Jagd in dem deckungsarmen Gelände wenig ergiebig. Heute sind hingegen von den ehemals großen Wildtierherden nur noch Restbestände zu beobachten (DICKORÉ 1995, HERMANNS 1949, JETTMAR 1993, MIEHE 1991, PETZSCH &
PIECHOCKI 1986).
Über den Beginn der Tierhaltung auf dem Plateau ist wenig bekannt. Die Domestikation des Schafes (Großart Ovis ammon) vollzog sich vor über 10000 Jahren im vorderen Orient. Die Größe der Chromosomensätze und cytologische Untersuchungen sprechen für eine Abstammung von den heute im Mittelmeerraum bis in den Iran verbreiteten Mufflons (BENECKE 1994). Seit dem 6. Jtsd. v. Chr. sind Schafe aus dem Nordwesten Indiens, aus Turkmenistan und China belegt (SCHOLZ 1995). Die Funde von Ovis spec. am Huang He (6745 BP) stammen von einer Wildform (HUANG WEIWEN 1994). Nach HERMANNS (1949) liegt die Wiege des Nomadentums Tibets in A mdo, dem heutigen Qinghai. Vermutlich hielt man zunächst Schafe, zeigt doch das Schriftzeichen für Tibeter in der Shang-Dynastie (1766 - 1126 v. Chr.) einen Menschen hinter einem Schaf. Die Hirtenkultur könnte vom Pamir-Hindukusch-Gebiet entlang des Nordrandes von Kunlun Shan und Altun Shan in den Nordosten des Tibetanischen Hochlandes gelangt sein. Von hier zogen die Nomaden weiter bis in den Süden Tibets, wo sie auf ansässige Bauern stießen (HERMANNS 1949).
Der Übergang von Wildziegen zu Hausziegen (Capra hircus) vollzog sich am Ende des 9. Jtsd. v. Chr. im Gebiet des fruchtbaren Halbmondes. Ausgangsgruppe waren wahr-scheinlich die in den Bergregionen Klein- und Vorderasiens verbreiteten Bezoaziegen, Capra aegagrus (BENECKE 1994, SCHOLZ 1995). Der heutige Yak oder Grunzochse

5. Der Untersuchungsraum – Ernährung 46
(Poephagus grunniens) ist aus dem ursprünglich in den Hochebenen Zentralasiens weit verbreiteten Wildyak (Poephagus mutus) hervorgegangen. Der Beginn der Domestikation liegt im Dunkeln, wird aber gelegentlich bereits für das Neolithikum vermutet. Ein Stoff-stück Yakwolle aus dem 5 Jtsd. v. Chr. belegt die frühzeitige Nutzung von Poephagus spec. (BENECKE 1994, SCHOLZ 1995).
Ihren Bedarf an Gerste handeln sich die Nomaden im Herbst in den Ackerbaugebieten ein. Sie nennen dieses Getreide Na. Die von den Chinesen gebrauchte Bezeichnung Tsing guo bedeutet wörtlich übersetzt Grünkorn, dem das Getreide eher gleicht als reifer Gerste. Die seßhaften Bauern lassen ihr Getreide an Gestellen aufgehängt nachreifen. Das traditionelle Grundnahrungsmittel Tzam pa stellen die Tibeter durch Mahlen der gerösteten Gerste her. Die hitzeschonende Zubereitung des Tzam pa mag zur Vitaminversorgung der Bevölkerung beitragen, ist doch der Anteil von Gemüse an der Ernährung ausgesprochen gering. Lediglich die Rhizome von Potentilla anserina werden im Frühjahr gegessen. Zudem nutzt man kleine gelbe Wildrüben und Wildzwiebeln. Im Herbst ergänzen Pilze das Nahrungsangebot (HERMANNS 1949). Rheum dient zwar den Mongolen zur Ernährung, den Tibetern aber nur zum Färben ihrer Stoffe. Die Mongolen verzehren zudem die Rhizome von Polygonum bistorta und P. viviparum. Aus den Zwiebeln von Lilium und Allium legen sie Wintervorräte an. Im Qaidam-Becken haben die süßsauren Beeren von Nitraria schoberi
große Bedeutung, zudem sammelt man die Beeren von Lonicera. Mehl gewinnen die Mongolen aus den Samen von Chenopodiaceen. Die Körner von Psamma villosa und Elymus giganteum ergeben eine Grütze. Andere Hirtenvölker (Tungusen, Burjaten, sibirische Tataren) nutzen die unterirdischen Pflanzenteile von Sanguisorba, Campanula und Hedysarum. Aus der inneren Rinde von Pinus, Picea und Abies wird ein Mehl gewonnen. (TAKAMIYA 1978).
5.6.2 Karakorum
Im Karakorum und im Hindukusch sorgen Acker- und Gartenbau für die Nahrungsgrundlage der Bevölkerung. Aufgrund der vernachlässigbar geringen Niederschläge der Tallagen findet der Pflanzenbau nur auf bewässerten Terrassen statt. Die Flüsse der Haupttäler sind meist zu tief eingeschnitten, um von ihnen Wasser auf die Felder leiten zu können. Daher wird in langen, störanfälligen Kanälen Wasser von höher gelegenen Seitentälern herangeführt. Bewässert wird vor der Saat und anschließend in etwa einwöchigen Abständen. In den warmen Talabschnitten baut man Weizen, Gerste, Hirse und Buchweizen an. Hülsenfrüchte, Lein und Kartoffeln sind von geringer Bedeutung. Wo es das Klima zuläßt, wird eine zweite Ernte im Jahr eingebracht. In warmen Lagen tritt Mais an die Stelle des Buchweizens als Zweitfrucht. In den ansteigenden Seitentälern und auf den Feldern in der Nähe der Hochweiden gedeiht nur Sommergerste.
Von den Früchten der angebauten Obstbäumen dienen besonders getrocknete Aprikosen als wichtige Zusatznahrung für den Winter. Ihre an Blausäure armen Steinkernen werden ebenso bevorratet wie auch Walnüsse.
Den größten Teil des Viehbestandes bilden Ziegen und Schafe. Sie liefern Milch, Wolle und Dung. Fleisch ist bei der Ernährung von geringer Rolle. Die wenigen Schlachtungen

5. Der Untersuchungsraum – Ernährung 47
erfolgen meist zu festlichen Anlässen vor Winterbeginn, da trotz der Zufütterung von Stroh, Heu und Laub die geringen winterlichen Weideflächen die Größe der Herden stark begrenzen. Dank der Hochweiden in 3000 bis 4000 m Höhe ist die Viehhaltung im Sommer problemlos. Die wenigen Rinder benötigt man als Zugtiere bei der Feldbestellung und zum Dreschen des Getreides (BRAUN 1996, JETTMAR 1993, MÜLLER-STELLRECHT 1979, SCHWEINFURTH 1957, 1983, SNOY 1975).

6. Ergebnisse und Diskussionen – Lingfeng im Qinling-Gebirge 48
6 Ergebnisse und Diskussionen
6.1 Lingfeng im Qinling-Gebirge
6.1.1 Das Untersuchungsgebiet
Lage und Geologie. Der Höhenzug des Qinling schließt sich östlich an das Tibetanische Hochland an. Sein Erscheinungsbild ist durch den steilen Nordabfall zum Graben des Wei He, durch hohe Gebirgsrücken und tiefe Flußtäler geprägt. Die welligen Bergrücken senken sich von 3000 m im Westen auf 2000 m nach Osten ab. Gemeinsam mit dem Dabie Shan und dem Huai-Fluß gliedert er den chinesischen Kernraum in den feucht-subtropischen Süden und den humid bis subhumiden, warmgemäßigten Norden. Hier verlaufen wichtige Wasser-, Klima - und Kulturscheiden sowie tier- und pflanzengeographische Grenzen (BLUNDEN &
ELVIN 1992, ERKES 1956, HENDL et al. 1978, LIU KAMBIU 1988, LI WENYI 1987, WANG
HESHENG 1981, ZHANG JINGWEI et al. 1981, ZHANG SHANZHEN et al.1985, ZHAO SONGQIAO
1986). Bis zum Übergang vom Devon zur Karbonzeit waren der heute nördliche und südliche Teil des Qinling durch einen Ozean getrennt. Dieser schloß sich beim Zusammenstoß vom Nordchina-Block mit dem Südchina-Block. Seit dem mittleren Karbon kam es zu Granitintrusionen. Den Qinling durchzieht die östliche Fortsetzung der sich an Qilian Shan und Kunlun Shan zum Pamir entlang ziehenden 3500 km langen Kollisionslinie der Blöcke. Der Qinling ist durch einen Staffelbruch vom deutlich höheren Tibetanischen Hochland abgesetzt (CHANG CHENGFA 1996, MENG QINGREN et al. 1996). Nach HSÜ
JEN (1983) erreichte der Qinling vermutlich schon nach dem frühen Pleistozän seine heutige Höhe. Tektonische Bewegungen mit regional differenzierten Hebungsvorgängen sind aber bis in das späte Pleistozän nachweisbar (HOU JIANJUN et al. 1995, PORTER et al. 1992).
Seine höchste Erhebung ist der Taibai Shan mit 3767 m. Hier stellte ROST (1992) zwei höchstwahrscheinlich durch ein Interstadial getrennte jungpleistozäne Vergletscherungen fest. Die zugehörigen Seitenmoränenwälle sind zwischen 2850 und 3310 m ausgebildet. Die Schneegrenzdepression im LGM betrug vermutlich 1000 m. Relikte spät- oder neoglazialer Vereisungen fehlen (ROST 1992, 1994). Lößablagerungen sind im Qinling bis in 3000 m
Höhe vorhanden (FANG XIAOMIN 1995). Es treten verbraunte Böden auf (CHEN CUNGEN
1987, ZHAO SONGQIAO 1994).
Nördlich des Qinling erreicht die Mächtigkeit des Löß in weiten Bereichen des Löß-plateaus (440.000 km2) über 100 m. Nahe der Stadt Lanzhou lagerte sich im Laufe der letzten 2,5 Mio. Jahre Löß von bis zu 500 m Dicke ab. Die nach Südosten hin abnehmende Mächtigkeit des Löß und seine geringer werdende Korngröße weist auf einen äolischen Transport des Staubes aus den innerasiatischen Trockengebieten (nordwestlicher Gansu Korridor, Gobi, Takla Makan) hin. Weitere Ursprungsgebiete waren die Hochebenen Tibets sowie trocken gefallene Flüsse, Seen und Meeresbuchten. Für den Transport sorgten der Wintermonsun, die Westwinde und die vom Tibetanischen Plateau herabwehenden Winde (AN ZHISHENG et al. 1995, LEHMKUHL 1997b, LEI XIANGYI 1995, LI PEIYING & ZHOU LIPING
1993, LIU TUNGSHENG et al. 1986, WU ZIRONG & GAO FUQING 1985, ZHANG DEER 1985).

6. Ergebnisse und Diskussionen – Lingfeng im Qinling-Gebirge 49
Wo die Geschwindigkeit dieser Winde durch die absteigenden Luftmassen des Lanzhou Hochs gebremst wurde, fiel besonders viel vom mitgeführten Staub nieder, und es entstanden die mächtigsten Ablagerungen (FANG XIAOMIN 1995). Die Bildung des Löß erfolgte wohl überwiegend unter den trockenen Bedingungen der Kaltzeiten. Die in den Lößprofilen enthaltenen Paläoböden werden hingegen den feuchtwarmen Bedingungen der Interglaziale und Interstadiale zugeschrieben und zeugen vom Erstarken des Sommer-monsuns. Der daraus abzuleitende Klimaverlauf korreliert mit den Ergebnissen der marinen Sauerstoffisotopen-Stratigraphie. So wird der unter dem spätpleistozänen Malan-Löß anstehende Paläoboden (S1) dem Isotopenstadium 5 zugeordnet (AN ZHISHENG et al. 1991a, DING ZHONGLI 1996, FORMAN 1991, KEMP et al. 1996, KUKLA & AN ZHISHENG 1989, LIU
TUNGSHENG et al. 1986, LIU TUNGSHENG & DING ZHONGLI 1993, PORTER et al. 1992, ZHENG
HONGHAN 1985).
Abb. 3.: Blockbild der Landschaft bei Lingfeng und Lage des untersuchten Profils (aus PACHUR et al. 1994). Das Gelände erreicht Höhen zwischen etwa 2500 und 2900 m ü. M.
Das Gebiet um Lingfeng (34°17’N/104°08’E) liegt im westlichen Qinling in der Provinz Gansu und weist Höhen zwischen etwa 2500 bis 2900 m auf (Abb. 3). Über roten Sedimenten des Tertiärs liegen hier pleistozäne sandige Flußschotter und mehrere Meter mächtige schluffige Schichten mit eingeschalteten organischen Lagen. Die Bildung einer Deckschicht aus umgelagertem Löß begann am Ende des Pleistozäns (PACHUR et al. 1994).

6. Ergebnisse und Diskussionen – Lingfeng im Qinling-Gebirge 50
Nördlich schließt sich das Lößplateau, südlich das Rote Becken der Provinz Sichuan mit dem Hauptort Chengdu an (ZHAO SONGQIAO 1986). Nach Südwesten folgen auf die mehr als 4000 m hohen Gebirgszüge des Min Shan am Ostrand des Tibetanischen Plateaus das Becken von Zoige (Rouergai Plateau) und in etwa 300 km Entfernung das Unter-suchungsgebiet des Nianbaoyeze.
Klima und Vegetation. Die Ortschaft Lingfeng liegt im Übergangsbereich von maritim zu kontinental getöntem Klima. Aufgrund der im Jahresverlauf wechselnden Einflüsse von Ostasien- und Wintermonsun fällt der überwiegende Teil der Niederschläge im Sommer, allein 60 % von Juni bis August. Hingegen sind die Winter ausgesprochen kalt und trocken. Die JMT dürfte bei etwa 5 °C liegen. Der Niederschlag beträgt etwa 550 mm, die potentielle Evaporation ist vermutlich 2,5 mal so hoch (DOMRÖS & PENG GONGBING 1988, PACHUR et al. 1994, PÖRTGE 1994). An der Nordseite weist der Fuß des Gebirges (um 500 m NN) vergleichbare Niederschläge wie Lingfeng auf. Die Niederschläge nehmen mit der Höhe stark zu, so daß in 800 m Höhe gleiche Werte wie an der in den Tieflagen feuchteren Südseite auftreten. An der Nordseite liegt das Niederschlagsmaximum von 1100 mm in 1500 m Höhe (ZHANG JIACHENG & LIN ZHIGUANG 1992).
Die im Norden an den Qinling angrenzenden Gebiete sind warm temperiert mit einer ausgeprägten Trockenzeit im Winter. Die Niederschläge nehmen zum Landesinneren erheblich ab. Entsprechend wird die Zone der sommergrünen Laubwälder auf dem Löß-plateau im großen Bogen des Huang He von Waldsteppe, Steppe und schließlich Wüste abgelöst. Die Steppen reichen bis an den westlichen Qinling heran. Die intensive Landwirt-schaft hat im Laufe von Jahrtausenden die Vegetation und das Landschaftsbild mitgeprägt. Der Ackerbau führte trotz der Anlage von Terrassen zu mehrere hundert Meter tiefen Erosionsgräben. Soweit vorhanden stehen die Wälder auf zimtfarbenen Böden und Braunerden. Sie ähneln denen des mitteleuropäischen Buchenwaldgebietes, doch spielen unter dem kontinental getönten Klima Arten der Gattung Fagus nur eine geringe Rolle. Statt dessen sind Eichen wie z. B. Quercus liotungensis vorherrschend. Im Süden sind Q. acutissima und Q. variabilis häufig. Daneben findet man weitere Arten von Quercus, Betula und Populus sowie Tilia mongolica, T. mandshurica, Pinus tabulaeformis und Juniperus chinensis. Zudem spielt Thuja (Platycladus) orientalis eine wichtige Rolle. In den Hochlagen des Berglandes sind Picea wilsonii, P. meyerii, Larix principis-rupprechtii und Pinus tabulaeformis verbreitet.
Entlang des Waldgebietes zieht sich im Westen ein schmaler Streifen mit Steppen-wäldern. Der Jahresniederschlag beträgt etwa 400 mm bei einer JMT von 1 bis 6 °C. Die sommergrünen Laubwaldbestände lösen sich hier zum Landesinneren hin auf, bis von ihnen nur noch kleine Bauminseln an Schattenhängen bleiben. Die von Westen eindringende Stipa bungeana-Steppe nimmt die baumfreien Flächen ein. Sie gedeiht noch bei 250 mm Nieder-schlag und reicht bis an den westlichen Qinling heran. Neben Stipa und weiteren Poaceen kommen in der deckungsarmen Vegetation u. a. dornbewehrte Sträucher der Gattungen Zizyphus und Caragana vor (CHEN CUNGEN 1987, HOU HSIOHYU 1983, LI BO et al. 1993, WALTER 1974, ZOU HOUYUAN et al. 1980).

6. Ergebnisse und Diskussionen – Lingfeng im Qinling-Gebirge 51
Die subtropischen Wälder südlich des Gebirges werden wegen der kühlen Winter besonders von sommergrünen Laubbäumen aufgebaut und leiten mit ihren immergrünen Elementen zu den Wäldern im Süden über. Die obere Baumschicht wird oft wie auf der Nordseite von sommergrünen Eichen (vor allem Quercus acutissima und Q. variabilis), Castanea sequinii, Liquidambar formosa, Platycarya strobilacea sowie verschiedenen Arten von Tilia und Acer aufgebaut. Außerdem tritt an der Südseite Pterocarya auf. Zu den überwiegend kälte- und trockenheitsresistenten Arten der zweiten Baumschicht und der Strauchschicht zählen die immergrünen und halbimmergrünen Castanopsis sclerophylla, Cyclobalanopsis glauca, Quercus engleriana, Q. acrodonta, Ilex, Ligustrum, Photinia und Phoebe. Die Hochlagen werden großflächig von Nadelwäldern mit Pinus henryi, P. bungeana, P. armandii, P. tabulaeformis, Picea wilsonii, P. brachytyla, Abies fargesii, A. chensiensis und Tsuga chinensis eingenommen.
In den Tälern beiderseits des Gebirges wird Ackerbau betrieben. Die an Baumarten reichen Waldbestände werden hier von Thuja orientalis (Nordseite) und Eichen dominiert. Zwischen 800 und 1300 m Höhe herrscht Quercus variabilis neben weiteren Arten von Eichen, Castanea und Populus vor, die oft unter niederwaldartiger Nutzung stehen. Die höher liegenden Wälder enthalten mehr als 40 Baumarten. Am häufigsten sind Quercus-Arten, Pinus tabulaeformis, P. armandii, Populus, Betula platyphylla var. szechuanica, B. luminifera, Tsuga chinensis, Carpinus cordata und C. fargesii. Tsuga chinensis hat im Qinling-Gebirge ihre nördliche Verbreitungsgrenze und steigt bis 2400 m. Zwischen 2200
bis 2600 m dominiert die Rotbirke Betula albo-sinensis. Ihr beigemischt sind Populus und Pinus armandii. Gruppen von Picea und Abies weisen den Birkengürtel als Ersatzgesellschaft aus, die in China häufig anstelle des unteren Nadelwaldgürtels zu finden ist. In Schlußgesellschaften tritt Pinus armandii nur bis in 2000 m Höhe auf. In höheren Lagen wird sie durch die Fichten verdrängt. Häufigste Fichte ist die bis 1600 m in den Laubwald hinabsteigende Picea wilsonii. Neben ihr treten P. asperata, P. neoveitchii und P. brachytyla auf. Aufgrund von Rodungen sind Fichtenwälder nur noch kleinflächig vorhanden. Unter ungestörten Bedingungen würden sie wohl die Höhen zwischen etwa 2000
bis 2500 m einnehmen.
Wie in fast allen chinesischen Gebirgen lösen mit zunehmender Höhe Tannen die Fich-ten ab. Am Taibai Shan haben Tannenwälder ihren Schwerpunkt von 2600 bis 3000 m. Sie werden von Abies fargesii und A. chensiensis aufgebaut. Beide Arten stoßen im Qinling an ihre nördliche Verbreitungsgrenze. Abies chensiensis bildet zwischen 2300 und 2700 m
Reinbestände aus. Darüber folgt Abies fargesii in Reinbeständen oder in tieferen Lagen gemeinsam mit Betula bzw. mit Larix chinensis in den höheren Lagen. In feucht-schattigen Wäldern bilden Rhododendren eine üppige Strauchschicht aus. Beigemischt sind Gehölze von Spiraea alpina, Lonicera tibetica und Sorbus koehneana. An trockenen Standorten fehlen die Rhododendren, und es entwickelt sich eine üppige Kraut- und Moosschicht.
Abies fargesii ist zwar bis in Höhen von 3300 m zu finden, doch wird der extrem kalte, windige und schneereiche subalpine Waldgürtel von 3000 bis 3350 m Höhe durch die Qinling-Lärche, Larix chinensis, einer geographischen Vikarianten der eng verwandten

6. Ergebnisse und Diskussionen – Lingfeng im Qinling-Gebirge 52
L. potaninii, beherrscht. An die Lärchenwälder schließt sich bis in 3500 m ein Zwergstrauch-gürtel mit Rhododendron capitatum, Salix cupularis, S. luctuosa und Spiraea alpina an. Stellenweise reichen die Lärche mit niedrigwüchsigen Exemplaren und Rhododendron, Juniperus, Potentilla, Salix, Spiraea und Lonicera in der Strauchschicht bis in Höhen von 3500 m. Wo die Gehölze fehlen, nehmen alpine Matten die Hänge bis zum Gipfel in 3767 m Höhe ein. Sie enthalten Poaceen (Roegneria, Poa, Trisetum, Deschampsia), Cyperaceen (Carex, Kobresia), Liliaceen (Allium, Fritellaria), Asteraceen (Anaphalis, Aster, Ligularia), Anemone, Sedum, Arenaria, Polygonum sphaerostachyum sowie zahlreiche Moose und Flechten. Eine nivale Stufe ist nicht ausgebildet (CHEN CUNGEN 1987, ROST 1992, ROST et al. 1996, WANG CHIWU 1961).
Zum Tibetanischen Hochland hin steigt die Höhenlage der Nadelwälder an. Im nördlich der Ortschaft Lingfeng verlaufenden Tal des Tao He liegt die Obergrenze des Gürtels von Fichten und Tannen um etwa 300 m über der des Taibai Shan. Picea asperata ist die dominierende Fichtenart. Daneben sind P. wilsonii, P. brachytyla und P. purpurea anzutreffen. Die Tannen sind durch Abies fargesii, A. faxoniana, A. recurva, A. ernestii und die Lärchen durch L. potaninii vertreten.
Besiedlung. Nach FENG SHENGWU (1985) nahm der Trockenfeldbau Chinas mit Beginn der Dadiwan-Kultur vor mehr als 7500 BP seinen Anfang auf dem Lößplateau. Siedlungen dieser frühen Zeit lagen unmittelbar nördlich des Qinling am Wei He und in dessen Einzugsgebiet. Hier war auch das Zentrum der ebenfalls neolithischen Yangshao-Kultur und der nachfolgenden bronzezeitlichen Kulturen. Auch der erste Kaiser Chinas (221 - 206 v. Chr.) hatte seinen Sitz am Wei He und ließ unweit des heutigen Xian bei Lintong seine Begräbnisstätte mit über 7000 Tonfiguren errichten (BLUNDEN & ELVIN 1992, GERNET
1994, ZHAO SONGQIAO 1986). Entsprechend lang und intensiv ist der anthropogene Einfluß auf die Natur des Lößplateaus einzuschätzen. Die Gebirgswälder unterliegen spätestens seit der Yuan-Dynastie (1279 - 1386) einer merklichen Abholzung, die in den nachfolgenden Jahrhunderten noch erheblich zunahm (XING JIAMING 1988, TANG BANGXING et al. 1994). Das Gebiet um Lingfeng ist heute so waldarm, daß Holzreste aus Sedimenten wie dem des untersuchten Profils von der Bevölkerung verfeuert werden (frdl. persönliche Mitteilung von Dr. B. WÜNNEMANN).
6.1.2 Das Profil Lingfeng
Das Profil „Lingfeng“ besteht zu einem großen Teil aus schluffigem Material, der obere Meter aus umgelagerten Löß. Torfige Lagen treten zwischen 2,2 - 4 m und, wechselnd mit sandigen Lagen, von 10,8 bis 17,5 m auf (Abb. 4., in der Beilage). Besonders im Bereich der torfigen Schichten gibt es größere Holzreste, die wegen der gegenwärtigen Waldarmut von der Bevölkerung ausgegraben und als Feuerholz genutzt werden. In situ eingebettete Wurzeln liegen u. a. in 11 und 16 m Tiefe. Nach den U/Th-Datierungen begann die Ablagerung des Profis in der letzten Warmzeit oder zu Beginn des Frühglazials (PACHUR
et al. 1994).

6. Ergebnisse und Diskussionen – Lingfeng im Qinling-Gebirge 53
Für die Pollenanalysen standen Proben zur Verfügung, die nach dem Schweretrennungs-Verfahren aufbereitet waren (vergl. Kap. 4). Bei der Berechnung wurde die Summe des GhP gleich 100 % gesetzt und die Prozentwerte aller Pollentypen und der Sporen hierauf bezogen. Das in das Pollendiagramm eingefügte Blockdiagramm enthält alle Pollentaxa des GhP und NGhP ausschließlich der Cyperaceen. Die Abfolge der Stratigraphie folgt PACHUR et al. (1994). Das bei PACHUR et al. (1994) dargestellte Pollendiagramm (dortige Fig. 8) stammt wie das hier vorgestellte Diagramm „Lingfeng“ vom Profil LIN 1 (dortige Fig. 4), berücksichtigt aber u. a. nicht die zur Interpretation wichtigen Sippen Juniperus, Hippophaë, Cyperaceen und Poaceen. Auf eine vergleichende Diskussion wird daher verzichtet.
Ergebnisse. In der Zunahme des GhP-Anteils am Gesamtpollen auf über 60 % und im gleichzeitigen Rückgang der Werte von Hippophaë, Salix und Betula sowie von Cyperaceen und Poaceen zeigt sich zu Beginn des DA 1 ein fortgeschrittenes Vorrücken der Wälder auf zuvor von lichten Pioniergesellschaften besiedelte Flächen. Pionierstandorte dürften angesichts der Gerölllagen im Profil und den vergleichsweise geringen organischen Beimengungen in den Sedimenten zunächst relativ häufig gewesen sein (PACHUR
et al. 1994).
Weit mehr als die Hälfte des gesamten GhP stammt aus der Gattung Picea (Picea p. p., Picea omorika-Typ), deren Arten in den vor Ort herrschenden Nadelwäldern dominierten. Die Werte von Abies und Larix erreichen nur wenige Prozente. Trotzdem ist mit dem Vorkommen von Abies zu rechnen. Larix, deren Pollen im Vergleich zu den übrigen Nadelhölzern zweifellos unterrepräsentiert ist, könnte auf konkurrenzarmen Standorten bestandsbildend aufgetreten sein.
Die geringen Werte von Pterocarya und Ulmus (Ulmus-Typ) sowie die PK von Tilia, Alnus, Corylus und des Carpinus cordata-Typs deuten vermutlich auf ein Vorkommen von Laubbäumen entlang der Wasserläufe hin. Den Unterwuchs der Wälder beherrschten Farne (Filicatae trilet p. p., Filicatae monolet p. p.) und Schachtelhalme (Equisetum). Ihre hohe Werte deuten auf feucht-schattige Standorte hin.
Nasse Standorte und offenes Wasser lassen sich von den Nachweisen der Algengat-tungen Botryococcus und Pediastrum ableiten. Nicht zuletzt kann angesichts der insgesamt geringen Nachweise der trockenheitsresistenten Gattung Ephedra (Ephedra distachya-Typ und E. fragilis-Typ) mit einem humiden Klima gerechnet werden.
In Analogie zu den heutigen Vorkommen von Picea und Abies zwischen 1800 und 3500 m Höhe im Tao He-Tal (WANG CHIWU 1961) dürfte sich die Umgebung von Lingfeng (ca. 2500 - 2900 m NN) zur Zeit des DA 1 in einer Stufe mit Picea(-Abies)-Wäldern befunden haben. Die NGhP-Anteile können dabei durchaus allein aus dem Unterwuchs der Wälder und von offenen Standorten des Flußtales stammen; sie sind daher keine sicheren Hinweise auf gehölzarme Hänge. Nach WISSMANN (1961) liegt Lingfeng heute zwar an der östlichen Grenze des Gebietes, in dem Vorkommen von Picea bzw. Abies an Schatthänge

6. Ergebnisse und Diskussionen – Lingfeng im Qinling-Gebirge 54
gebunden sind. Heute spielt hier allerdings der anthropogene Einfluß eine entscheidende Rolle auf die geringe Waldverteilung.
PK von anspruchsvollen Baumgattungen wie z. B. Pinus und Tsuga, ab dem DA 2 auch von Quercus, die heute im unteren Bereich der Picea-Wälder oder in den tiefer gelegenen Laubwäldern vorkommen (CHEN CUNGEN 1987), dürften fast ausschließlich aus dem Fernflug stammen. Die Landschaft um Lingfeng bot den genannten Gattungen mit Höhen von zumeist über 2500 m offenbar keine geeigneten Standorte.
Am Ende des DA 1 geht der Anteil des GhP deutlich zurück. Bestände mit den weniger anspruchsvollen Gehölzgattungen Betula und Juniperus nahmen im Vergleich zu denen der Fichten zu. Der Anstieg der Werte von Cyperaceen und Poaceen weist auf Auflichtungen hin. Angesichts der hohen Prozentwerte dürften im wesentlichen Cyperaceen und Poaceen die Vegetation (Matten, Wiesen) aufgebaut haben, wo Baumbestände verschwunden waren. Eine geringe Zunahme von Artemisia und Chenopodiaceen sowie Nachweise von Thalictrum und Brassicaceen sprechen für eine Ausweitung gestörter Standorte.
Am Beginn des DA 2 weist der steile Abfall der Kurven der Farnsporen und der Rückgang der Picea-Werte auf eine vollständige Verdrängung der feucht-schattigen Picea (-Abies)-Wälder im Umfeld des Untersuchungspunktes hin. Entsprechend nimmt die Häufigkeit der aus tieferen Lagen heraufgewehten PK von Tsuga und Quercus zu. Das Aufkommen von Gebüschen mit Hippophaë und Weiden (Salix, Salix oritrepha-Typ) sowie die Bildung schluffig-sandiger Sedimente (PACHUR et al. 1994) zeigen eine Pioniergehölz-besiedlung offener, durch verstärkte fluviale Dynamik entstandener Flächen an. Auf diese Pionierphase folgte in der Mitte des DA 2 ein Gebüschstadium mit Juniperus, Rhodo-
dendron und Potentilla. Auf das Vorkommen von Rhododendron weisen neben den Pollenfunden auch Hölzer aus 17,1 m und 16,5 m Tiefe hin, die nach den Abbildungen bei CUTLER et al. (1987) zu Rhododendron gestellt werden können. Diese heute besonders in der oberen subalpinen Stufe anzutreffenden Gebüsche bildeten vermutlich eine an verschlechterte klimatische Verhältnisse angepaßte Klimaxgesellschaft aus, die am Ende des DA 2 wiederum durch Pioniergehölze abgelöst wurde. Neben Hippophaë und Salix beteiligten sich nun auch Lärchen (Larix) an der Wiederbesiedlung der offenen Flächen, die u. a. durch erhöhte fluviale Aktivität (stark minerogene Sedimente) entstanden.
Im Unterwuchs der Gebüsche und in der übrigen, wohl in weiten Bereichen gehölz-armen Vegetation des DA 2 herrschten Cyperaceen und Poaceen vor. Hinweise auf Änderungen des Arteninventars deuten sich im etwas stärkeren Auftreten des Anemone/Ranunculus-Typs, von Brassicaceen und von Liliaceen sowie in den ersten Pollenfunden des Swertia- und des Bupleurum-Typs an. Die Sporenfunde von Botrychium und vom Riccia-Typ verweisen auf Lücken in der Pflanzendecke.
Nachweise von Myriophyllum und Hippuris zeigen Stillgewässer an. Daher können die hohen Werte des Potamogeton/Triglochin-Typs wohl auf ein reichliches Vorkommen von Potamogeton zurückgeführt werden.
Im DA 3 kam es unter Verdrängung der Pioniergehölze zur Wiederansiedlung von Nadelwäldern die in Flächenausdehnung, Artenzusammensetzung und Unterwuchs den im

6. Ergebnisse und Diskussionen – Lingfeng im Qinling-Gebirge 55
DA 1 existierenden Picea-(Abies)-Wäldern glichen. Ein Teil der Nadelbäume war von dem Parasiten Arceuthobium (Loranthaceae, Mistelgewächse) befallen. Von den Pinus-PK konnten trotz schlechter Erhaltung manche als PK der Untergattung Diploxylon oder Haploxylon angesprochen werden (vergl. Kap. 4.4.). Die PK des Diploxylon-Typs mögen aus Mischwäldern mit Pinus tabulaeformis stammen, die wie heute unterhalb der Picea-Abies-Stufe angesiedelt waren. Der im Diagramm allgemein etwas häufigere Haploxylon-Typ hat in Pinus armandii einen klimatisch weniger anspruchsvollen Vertreter, der heute bis in den unteren Bereich der Picea-Wälder hinein in Reinbeständen oder mit Picea wilsonii vorkommt. Das gehäufte Auftreten des Diploxylon-Typs im DA 3 mag zusammen mit den Nachweisen von Arceuthobium sowie den Funden von Fagus und Hedera auf etwas günstigere klimatische Bedingungen als im DA 1 hindeuten.
In der Mitte des DA 3 treten vereinzelt etwas geringere Picea-Werte auf. Die in diesem Bereich des Profils eher stärker minerogenen Sedimente und die leicht erhöhten Werte von Hippophaë, Salix und Lonicera sprechen für eine fluviale Beeinträchtigung der flußnahen Nadelwälder. Neben den Pioniergehölzen profitierten hiervon auch Ulmus und Carpinus (C.
cordata-Typ). Hinweise auf Verlandungsgesellschaften an ruhigen Gewässern sind allein die wenigen Funde vom Sparganium-Typ.
Mit dem Übergang zum DA 4 wandelten sich die dichten Nadelwälder in lichte Gehölze mit Juniperus und Rhododendron um die in einer von Cyperaceen und Poaceen beherrschten Landschaft standen. Die Bedingungen ähneln zunächst denen, wie sie für die Mitte des DA 2 abgeleitet wurden. Zwar kamen Juniperus-Gehölze und Bestände von Nadelwald wohl auch bis zum Ende des DA 4 vor, doch weisen das Aussetzen der Rhododendron-Kurve und die zunehmenden Anteile von Hippophaë sowie das zunehmend minerogene Sediment auf einen deutlichen Wandel des Landschaftsbildes und der geodynamischen Prozesse hin, der auch zu einem stärkeren Auftreten von Ranunculaceen (Ranunculus/Anemone-Typ), Asteraceen (Aster-Typ, Saussurea-Typ, Cichorioideae) und des Polygonum bistorta-Typs führte.
Die Pollenführung der zwei Proben des DA 5 ist gering. Die Proben enthalten fast ausschließlich PK von Hippophaë, Potentilla, Artemisia und vom Aster-Typ, deren Erhaltung z. T. schlecht ist. Vermutlich hat die Trockenheit des Sedimentes eine Rolle gespielt. Die Pollenspektren sind für eine Rekonstruktion der damaligen Vegetation wenig aussagekräftig. Ein Vergleich zwischen den Prozentwerten des DA 4 und des DA 6 zeigt immerhin, wie sehr sich während des DA 5 die Vegetation außerhalb der Hippophaë-Gebüsche gewandelt hat.
Im DA 6 macht der GhP weit weniger als 5 % des gesamten Polleneintrages (inkl. NGhP und Cyperaceen) aus. In mehreren Proben sind allein die PK der Cyperaceen mehr als 25 mal häufiger als solche von Bäumen und Sträuchern. Bei Ausschluß dieser vermutlich durch lokale Vorkommen überhöhten Cyperaceen-Anteile ergibt sich für den GhP ein Abfall von 60 % im DA 4 auf nun mehr etwa 20 % im DA 6. Bis auf einzelne Bestände von Hippophaë, Juniperus und Salix wird die Landschaft im DA 6 fast frei von Gehölzen gewesen sein. Zwar liegt ein Fund von Picea-Holz aus 4,75 m Tiefe vor, doch könnte Umlagerung eine Rolle spielen. Die Annahme kleiner Fichtenrestbestände an günstig exponierten Hängen erscheint eher unwahrscheinlich. PK der Gattungen Quercus und Tilia stammen wohl sicher aus

6. Ergebnisse und Diskussionen – Lingfeng im Qinling-Gebirge 56
tieferen Lagen oder aus älteren Sedimenten. Nachweise der heute im Gebiet des Qinling an ihre nördliche Verbreitungsgrenze stoßenden Gattungen Tsuga und Pterocarya fehlen ganz. Auch der Ulmus-Typ wurde nicht nachgewiesen. Die vergleichsweise hohen Werte von Pinus und Betula könnten u. a. von gestörten Standorten stammen, an denen Arten beider Gattungen als Pioniere aufgetreten sein dürften.
Die hohen Werte des Anemone/Ranunculus-, Trollius- und des Caltha-Typs sowie von Thalictrum belegen ein artenreiches Vorkommen der Familie der Ranunculaceen. Zahlreich vertreten waren auch die Asteraceen (besonders Artemisia, Aster-Typ, Saussurea-Typ). Die hohen Werte des Polygonum alpinum-Typs, der Gentianaceae p. p. und des Swertia-Typs kennzeichnen den alpinen Charakter der damals allgemein vorherrschenden und von Cypera-ceen und Poaceen aufgebauten Matten- und Rasengesellschaften.
Zwei an Pflanzenresten reiche Sedimentlagen zwischen 5 und 6 m Tiefe sprechen am Untersuchungspunkt für eine zeitweise dichte Vegetationsdecke. Diese wurde zunächst von Vertretern von Cyperaceen, Artemisia und Trollius sowie des Anemone/Ranunculus-Typs aufgebaut. Anschließend deuten die hohen Werte von Sanguisorba filiformis, vom Caltha-, Aster- und Saussurea-Typ sowie von Gentianaceen und Apiaceen auf die Entstehung von Sumpfwiesen hin, wie sie aus dem Becken von Zoige (um 3500 m) im östlichen Tibet mit Sanguisorba filiformis und Arten von Caltha, Aster, Cremanthodium, Saussurea, Gentiana und Chamaesium beschrieben worden sind (BJÖRK 1993, THELAUS 1992). Angesichts der sich abzeichnenden Klimaverhältnisse könnte einsetzender Permafrost saisonal zu staunassen Verhältnissen geführt haben, doch gibt es in der Sedimentstratigraphie keine Belege für diese Annahme (frdl. persönliche Mitteilung Dr. B. WÜNNEMANN).
Im DA 7 dürfte es in günstigen Lagen zu einer Wiederbewaldung mit Arten der Gat-tungen Picea, Abies und Larix gekommen sein. Selbst Pterocarya und Ulmus scheinen ebenso wie Arceuthobium in das Gebiet zurückgekehrt zu sein. Die hierin erkennbare Klimagunst zeigt sich auch in den höheren organischen Anteilen des Sediments. Der GhP-Anteil bleibt aber unter dem vom DA 2 und DA 4. Am Ende des DA 7 erfolgte ein rapider Rückgang der geringen Waldbestände, und alpine Matten und Rasen mit Cyperaceen und Poaceen breiteten sich aus.
Die in den DA 2, DA 4 und DA 7 relativ geringen Picea-Werte sprechen für eine gerin-gere Bedeutung der Nadelwälder als in den DA 1 und DA 3. Inwieweit die Obergrenze der Wälder dabei allgemein tiefer lag oder Baumbestände auch in hohen Lagen, beschränkt auf zusagende Standorte in günstiger Hangexposition, vorkamen, kann nicht sicher beurteilt werden.
Das Landschaftsbild im DA 8 dürfte dem des DA 6 ähnlich gewesen sein. Jedoch sind beispielsweise der im DA 6 häufige Sanguisorba filiformis und der Polygonum alpinum-Typ nicht vertreten. Aufgrund der zunehmenden Lößführung wurden die oberen 40 cm

6. Ergebnisse und Diskussionen – Lingfeng im Qinling-Gebirge 57
des Profils nicht untersucht und die Bedeutung des hohen Anteils des Polygonum bistorta-Typs bleibt offen.
Diskussion. Zeitstellung. Anhand der U/Th-Datierungen ist die Basis des Profils in das letzte Interglazial oder den Beginn des Frühglazials zu stellen (PACHUR et al. 1994). Zu der im folgenden zugrunde gelegten Lößstratigraphie sei auf AN ZHISHENG et al. (1991a,b), FORMAN (1991), KUKLA & AN ZHISHENG (1989), LIU TUNGSHENG et al. (1986) und ZHANG
XIAOYE et al. (1994) verwiesen. Überträgt man die aus Profilen des Lößplateaus bekannte Gliederung des Paläobodenkomplexes S1 in bis zu drei durch Lößlagen getrennte Bodenhorizonte und deren Korrelation mit den Abschnitten des Sauerstoffisotopenstadiums 5 auf das Pollendiagramm von Lingfeng, so läßt sich der jüngere der beiden klimatisch besonders günstigen vegetationsgeschichtlichen Diagrammabschnitte, der DA 3, mit dem jüngsten Boden des S1, dem S1SS1, und damit zum Isotopenstadium 5a in Bezug setzen. Der DA 1 entspräche damit dem mittleren Boden S1SS2 des Stadiums 5c und der klimatisch ungünstige DA 2 dem jüngeren Lößhorizont S1LL2 in 5b. Die beiden Waldzeiten dürften dem Odderade (DA 1) bzw. dem Brörup (DA 3) des mitteleuropäischen Raumes entsprechen (GRÜGER 1989, 1995, BEHRE & LADE 1986). Der DA 4 bis 8 entsprächen somit dem Zeitraum der letzglazialen Bildung des Malan-Löß (L1).
Es darf jedoch nicht übersehen werden, daß einerseits die Korrelation der Lößstrati-graphie mit den Isotopenstadien der Tiefseesedimente noch gewisse Unsicherheiten birgt und andererseits die Ergebnisse der U/Th-Datierungen des Profils für eine Zuordnung der DA 1 bis 3 zu den Abschnitten des Stadiums 5 ungeeignet sind.
Klima. Während des DA 1 und DA 3 lag das Gebiet um Lingfeng in einer Stufe mit Picea-(Abies)-Wäldern. Aus dem nahen Tao He-Tal gibt WANG CHIWU (1961) Picea-Abies-Wälder heute aus Höhen zwischen 1800 m und 3500 m an. Bei Lingfeng dürfte damit im DA 1 und DA 3 ein in etwa ähnliches Klima wie in der Gegenwart geherrscht haben: JMT um 4,5 °C, Jahresniederschlag über 500 mm (frdl. persönliche Mitteilung Dr. J. BÖHNER). Für einen differenzierteren Vergleich fehlt es an palynologischen Untersuchungen zur holo-zänen Vegetationsgeschichte des Gebietes.
Geht man für eine Strauchformation, wie sie zur Mitte des DA 2 und zu Beginn des DA 4 vorlag, von einer JMT von weniger als 2 °C aus (LI WENHUA 1993), so dürfte die JMT der subalpinen Phasen mindestens 2,5 °C unter der heutigen gelegen haben.
Während des waldfreien DA 6 muß die Obergrenze der Wälder mehr als 1000 m tiefer als heute gelegen haben. Bei einem Temperaturgradienten zwischen 0,5 °C und 0,6 °C pro 100 m Höhendifferenz (CHEN CUNGEN 1987, LEHMKUHL & LIU SHIJIAN 1994b, LI WENHUA
1993, WARDLE 1981, YU XIAOGAN & SUN SHANGZHI 1981, ZHAO SONGQIAO 1986)
entspricht dies einer um mindestens 5 bis 6 °C geringeren JMT. Nach ROST et al. (1996) treten alpine Matten wie im DA 6 heute am Taibai Shan im östlichen Qinling etwa ab einer JMT von weniger als -1 °C auf. Aus beiden Ableitungen ergibt sich eine Differenz von mindestens etwa 5,5 °C zur heutigen Temperatur. Die fortwährende Existenz
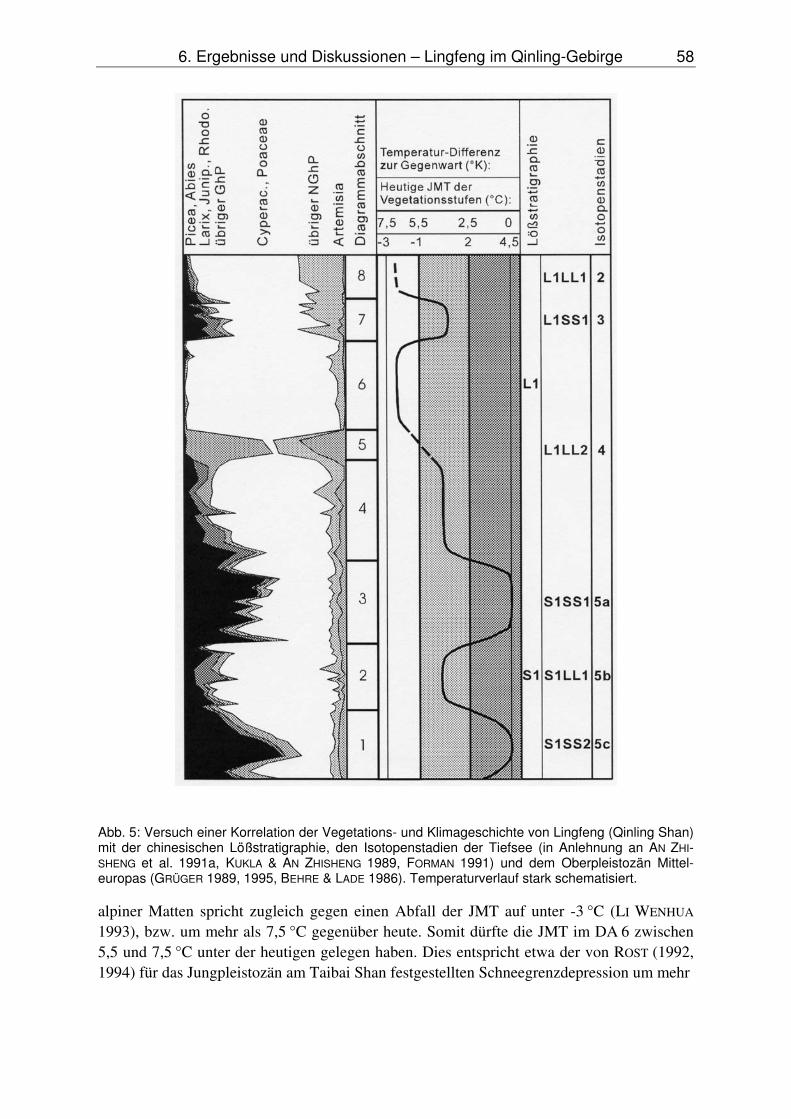
6. Ergebnisse und Diskussionen – Lingfeng im Qinling-Gebirge 58
Abb. 5: Versuch einer Korrelation der Vegetations- und Klimageschichte von Lingfeng (Qinling Shan) mit der chinesischen Lößstratigraphie, den Isotopenstadien der Tiefsee (in Anlehnung an AN ZHI-SHENG et al. 1991a, KUKLA & AN ZHISHENG 1989, FORMAN 1991) und dem Oberpleistozän Mittel-europas (GRÜGER 1989, 1995, BEHRE & LADE 1986). Temperaturverlauf stark schematisiert. alpiner Matten spricht zugleich gegen einen Abfall der JMT auf unter -3 °C (LI WENHUA
1993), bzw. um mehr als 7,5 °C gegenüber heute. Somit dürfte die JMT im DA 6 zwischen 5,5 und 7,5 °C unter der heutigen gelegen haben. Dies entspricht etwa der von ROST (1992, 1994) für das Jungpleistozän am Taibai Shan festgestellten Schneegrenzdepression um mehr

6. Ergebnisse und Diskussionen – Lingfeng im Qinling-Gebirge 59
als 1000 m. Dabei spiegelt die Entwicklung der Vegetation vorrangig die Temperaturverhältnisse der Vegetationsperiode wider. Sollte die Abkühlungstendenz der Sommermonate stärker als die des Winters gewesen sein (HÖVERMANN & SÜSSENBERGER
1986), dann müßte die tatsächliche mittlere Temperatur des Jahres höher, im umgekehrten Fall niedriger gewesen sein (KONG ZHAOCHEN et al. 1990, WANG PINXIAN & SUN XIANGJUN
1994, ZHANG LANSHENG 1984). Stand die damalige Sumpfvegetation mit Sanguisorba filiformis etc. tatsächlich auf Permafrostboden, so würde dies für einen sehr kalten Abschnitt im jüngsten Teil des DA 6 sprechen. Nach ROST (1998) sowie ZHOU YOUWU et al. (1991) und HÖVERMANN & LEHMKUHL (1994) kann im Qinling und anderen Gebirgen mit Permafrost ab einer JMT um -2 °C gerechnet werden. Die Differenz zur heutigen JMT (4,5 °C) von 6,5 °C läge noch im Rahmen des aus der übrigen Vegetationsentwicklung abgeleiteten Temperaturrückganges.
Ist die angenommene zeitliche Einordnung des DA 4 bis DA 6 (s. o., Abb. 5) richtig, so ergibt sich für das Isotopenstadium 4 die Gliederung in einen wärmeren (DA 4) und einen kälteren (DA 6) Abschnitt.
Den 14C-Datierungen zufolge ist der DA 7 älter als 38800 BP (Hv 21337 - 21340). Die geringe Wiederbewaldung dieser Zeit fällt somit in den Beginn eines aus den Lößprofilen bekannten Interstadials (AN ZHISHENG et al. 1995, PORTER et al. 1992, ROST 1994, 1998, ZHANG XIAOYE et al. 1994) mit einer als L1SS1 bezeichneten schwachen Bodenbildung – manche Profile zudem mit einem L1SS2 – (AN ZHISHENG et al. 1991a, FORMAN 1991, KUKLA & AN ZHISHENG 1989) und erhöhten Seeständen (LEHMKUHL & HASELEIN 1999). Vermutlich ist der jüngere Teil dieses bis etwa 25000 BP währenden Interstadials im Pollenprofil nicht archiviert. Nach den Pollen- und Makrorestanalysen am Profil von Bei-zhuangcun (530 m ü. M.) nahe Xian, das etwa 31000 Jahre zurückreicht, bestanden zumindest in den tieferen Lagen des Qinling noch bis etwa 26000 BP Wälder mit Picea wilsonii und Abies (HEAD et al. 1989, LIU KAMBIU 1988, ROST 1994, 1998, SUN JIANZHONG
et al. 1991). Die JMT im DA 7 wird ähnlich dem DA 2 und DA 4 um 2,5 - 5,5 °C unter der heutigen gelegen haben. Für den Zeitraum 31000 - 26000 BP rekonstruiert ROST (1998) eine ganz ähnliche Temperaturabsenkung von 3 - 6 °C.
Für den DA 8, dessen Vegetationsverhältnisse den alpinen Matten des DA 6 am nächsten stehen, ergaben Proben aus 107 und 135 cm Tiefe Alter von 17.500 und 27.205 BP, die in die Zeit des L1LL1 fallen. Die unzureichende sedimentologische Archivierung des DA 8 läßt genauere klimatische Angaben nicht zu, doch ist ein Vergleich mit dem DA 6 am nahe liegendsten. Hierfür sprechen auch die Hinweise auf eine im entsprechenden Zeitraum um ca. 6 °C geringere Temperatur in Ost-Tibet (LEHMKUHL 1995a). Die Annahme einiger Autoren von Temperaturabsenkungen während des LGM zwischen 10° und 13 °C (z. B. AN
ZHISHENG et al. 1991C, LIU KAMBIU 1988, ZHANG LANSHENG 1984) kann bei Beachtung der zugrundeliegenden Befunde nicht geteilt werden (ROST 1998). Die Klimaänderungen im China südlich des Qinling dürften allgemein deutlich geringer gewesen sein als bei Lingfeng und im Norden Chinas (LI WENYI 1987, LIU JINLING et al. 1986, ZHANG LANSHENG 1984).

6. Ergebnisse und Diskussionen – Lingfeng im Qinling-Gebirge 60
Die nachgewiesenen Vegetationstypen erfordern nach heutigen Bedingungen zumeist Niederschläge von 500 mm oder mehr im Jahr (CHANG D. H. S. 1981, LI WENHUA 1993, MOU XINDAI et al. 1992). Das zeitweise häufige Auftreten von Pioniergehölzen deutet auf ein saisonal stark schwankendes Abflußregime hin. Das mag in klimatischen Übergangs-phasen (Beginn DA 1, Grenze DA 1/DA 2, Grenze DA 2/DA 3, DA 5) durch eine im Umbruch begriffene, der Erosion weniger Widerstand entgegensetzende Vegetationsdecke mitbedingt sein. Im DA 3 werden zudem hohe sommerliche Monsunniederschläge von Bedeutung gewesen sein.

6. Ergebnisse und Diskussionen – Der Gaxun Nur in der Inneren Mongolei 61
6.2 Der Gaxun Nur in der Inneren Mongolei
6.2.1 Das Untersuchungsgebiet
Der Gaxun Nur, auch Gaschun-Nuur oder Kashunnuer (42°06’N/100°51’E), ist ein ehemaliger Endsee im zentralasiatischen Wüstengebiet des innermongolischen Hochlands nahe der Grenze zur Äußeren Mongolei. Sein Seeboden liegt in etwa 900 m Höhe auf dem im weiteren Umfeld durchschnittlich 1000 bis 1500 m hohen Alaschan Plateau. Im Norden steigt das Noyan Bogda-Gebirge über 2500 m Höhe an.
Zur Äußeren Mongolei hin beherrschen die Steinwüsten der Gobi das Landschaftsbild. Der größte Teil des Alaschan Gebietes sind Sandwüsten wie die Badain Jaran Shamo und Tengger Shamo. Die aus den Gebirgen kommenden Flüsse enden oft in salzigen Seen und Sümpfen.
Die beiden im Qilian Shan entspringenden wasserreichen Flüsse Hei He und Beida He vereinigen sich zum Edsin Gol (Ruo Shui), der früher über den Mören Gol in den Gaxun Nur floß, heute aber nach menschlichen Eingriffen im östlich des Gaxun Nur gelegenen Sogo Nur endet. Oftmals ziehen von Schmelzwässern gespeiste Grundwasserströme aus den Gebirgen entlang alter Flußläufe in die Wüsten hinein. Die von ihnen gespeisten Wasser-becken erreichen ein halbes Jahr nach der sommerlichen Schneeschmelze ihren Höchststand. Vom Noyan Bogda-Gebirge im Norden führen zahlreiche trockene Flußbetten zur weiträumigen Beckenlandschaft des Gaxun Nur und Sogo Nur.
Über die Jahre gemittelt fallen im Gebiet des Gaxun Nur weniger als 50 mm Nieder-schlag, und zwar größtenteils während des Sommermonsuns von Juni bis August/ Sep-tember. Die jährlichen Mengen schwanken sehr stark. In manchen Jahren bleibt der Regen fast ganz aus. Die geringe Menge und starke Variabilität werden durch die von Gebirgen abgeschirmte kontinentale Lage bestimmt. Die JMT beträgt etwa 8,4 °C, die mittlere Temperatur des Januar -12 °C und die des Juli 26,3 °C (BÖHNER 1996, DOMRÖS & PENG
GONGBING 1988, FEZER & HALFAR 1994, NORLINDH 1949, WALTER 1974, YANG XIAOPING
1991, ZHANG JIACHENG & LIN ZHIGUANG 1992, ZHAO SONGQIAO 1986, 1994).
In den Steinwüsten der Gobi ist über hunderte von Kilometern fast keine Vegetation zu finden. Dort, wo Pflanzen gedeihen, bedecken sie oft nicht mehr als 5 % der Bodenober-fläche. Auf unbeweglichen Sanden der Shamo kann die Deckung 40 - 60 % betragen. Häufig sind es Sträucher und Halbsträucher aus den Familien der Chenopodiaceen, Zygophyllaceen und Asteraceen. In sandigen Senken, oft zwischen Dünen, wachsen Haloxylon und Nitraria. In Übergangsbereichen von Salzwasserstellen zu Sanddünen und bei hohem Grundwasserstand trifft man auf Nitraria und Kalidium. Die Asteraceen sind u. a. durch die Gattungen Artemisia und Aster vertreten. Phragmites communis steht im Röhricht der Seen und auf halophilen Sumpfwiesen.

6. Ergebnisse und Diskussionen – Der Gaxun Nur in der Inneren Mongolei 62
Die häufigste Baumart ist Ulmus pumila. Sie tritt in einzelnen Exemplaren und offenen Gruppen auf grundwassernahen Standorten auf. Alte Bäume weisen auf frühere Auenstandorte hin. Die westliche Grenze ihrer Verbreitung verläuft am Gaxun Nur. Populus diversifolia begleitet Flußläufe und Bewässerungskanäle der Oasen, ist zuweilen aber auch in Dünentälern zu finden.
Mit der Höhe sinken die Temperaturen und die Niederschläge nehmen zu, so daß man in manchen Bereichen der die Wüsten umrandenden und in sie vorstoßenden Gebirge auf Nadelwälder stößt. Im Mongolischen Altai sind es zwischen Höhen von etwa 1900 und 2300 m Wälder von Larix sibirica. Nach Nordosten nehmen mit den Niederschlägen die Bestände von Abies sibirica, Picea obovata und Pinus cembra ssp. sibirica erheblich zu. Pinus sylvestris steht auf trockenen, sandigen Böden der Gebirgsränder.
Im Tien Shan geht die spärlich bewachsene Steppe der tieferen Lagen in 1100 m Höhe in eine Steppe mit Artemisia terrealbe, Festuca ovina und Stipa-Arten über. Von 1800 bis 2900 m treten auf den Schattenhängen der Nordseite Picea schrenkiana und Larix sibirica auf. Daneben ist auch Abies semenovii anzutreffen. Zumeist handelt es sich um Waldsteppen, selten um flächendeckende Wälder. Auf der Südseite liegt die Waldstufe um 500 m tiefer.
Der Gebirgszug des Helan Shan, unmittelbar westlich des Huang He-Bogens trägt in Höhen zwischen 2500 und 3000 m Wälder mit Picea asperata und Pinus tabulaeformis. An der Nordseite des Tibetanischen Plateaus im Qilian Shan liegt zwischen etwa 2000 und 3500 m eine Stufe mit Waldbeständen an schattigen, feuchten Hängen aus Picea crassifolia. Zudem kommen auch Picea wilsonii und Pinus tabulaeformis vor (BARTHEL 1990, CHEN
CUNGEN 1987, CHEN GUICHEN et al. 1994, HILBIG & KNAPP 1983, HUANG RONGFU 1987, HILBIG 1995, PLAETSCHKE 1939, SCHMIDT-VOGT 1987, SUCCOW 1989, WALTER 1974, WANG CHIWU 1961, YUAN GUOYING 1986).
6.2.2 Das Profil Gaxun Nur – Erste Voruntersuchungen
In der Inneren Mongolei wurde im seit Jahrzehnten ausgetrockneten Seebecken des Gaxun Nur (42°06’N/100°51’E) ein 235 m langer Kern erbohrt, in dem die Brunhes/Matuyama-Grenze in 191 m Tiefe liegt (frdl. persönliche Mitteilung Prof. Dr. H.-J. PACHUR und Dr. B. WÜNNEMANN). Es wurden 16 Proben auf ihren Pollengehalt hin untersucht werden, um die Brauchbarkeit des Profils für vegetationsgeschichtliche Zwecke zu testen. Nach dem derzeitigen Bearbeitungsstand ist das Profil besonders in den Bereichen zwischen 100 und 235 m, um 50 m sowie in einer Tiefe von 20 m für pollenanalytische Untersuchungen geeignet. Wenn auch die bisherigen Ergebnisse noch spärlich sind, sollen sie wegen der ungenügenden Datenlage des zur Diskussion stehenden Untersuchungs- und Zeitraumes doch dargestellt werden.
Den in Abb. 6 dargestellten Proben liegt eine durchschnittliche Zählsumme von lediglich 100 PK zugrunde. In den übrigen 5 Proben (17, 34, 65, 77 und 96 m) konnten nur weniger als 10 PK ausgezählt werden. Die an jeder zweiten Probe vorgenommene Bestimmung der Pollendichte ergab Werte zwischen 8 und 6100 PK pro cm3.
fschluetz
Highlight
fschluetz
Highlight

6. Ergebnisse und Diskussionen – Der Gaxun Nur in der Inneren Mongolei 63
Abb. 6: Pollendiagramm des Profils „Gaxun Nur“, Innere Mongolei, ca. 950 m. ü. M. Brunhes/-Matuyama-Grenze in 191 m Tiefe. (GS = GhP + NGhP, vereinfachte Wiedergabe).
Ergebnisse. Die Proben des DA 1 zeichnen sich durch GhP-Anteile von über 60 % aus. Die Nachweise von Koniferen überwiegen. Zu Beginn nehmen die Werte von Picea und Abies zu und deuten wie der Rückgang von Juniperus auf eine Klimabesserung zur Brunhes/Matuyama-Grenze hin an. Später weisen die zurückgehenden Werte von Picea/Abies und die Zunahme von Juniperus auf eine Klimaverschlechterung hin. Zudem treten Pinus und Larix verstärkt auf.
Häufige Laubgehölze sind Betula und Vertreter des Ulmus-Typs. Der hohe Anteil tetraporater PK des Ulmus-Typs könnte auf Ulmus pumila hinweisen, die relativ hohen Werte von Hippophaë und Pterocarya auf die Nähe von Flußläufen. Daneben konnten durch wenige Pollenfunde noch Tilia, Alnus, Rosaceen, Weigela und Elaeagnus nachgewiesen werden.
Die Vegetation trockener und vermutlich teilweise versalzter Standorte ist durch Ephedra, Nitraria, Chenopodiaceen und den Armeria-Typ belegt. Die Nachweise des Sparganium/Typha- und des Typha latifolia-Typs weisen auf Röhrichte an Seeufern oder in Sumpfwiesen hin. Auf feuchte Standorte verweisen auch die Funde von Farnsporen. Im Rückgang der trileten Farnsporen (Filicatae trilet) und in der Zunahme monoleter Sporen (Filicatae monolet) spiegelt sich vermutlich eine Änderungen der lokalen Wuchsbedingungen wider. Nach den niedrigen Werten von Poaceen und Artemisia zu urteilen können Steppen kaum von Bedeutung gewesen sein.

6. Ergebnisse und Diskussionen – Der Gaxun Nur in der Inneren Mongolei 64
Auf das pollenarme Material des DA 2 folgt der DA 3 mit zwei Proben, deren Werte von Picea/Abies, Pinus und Larix eine ähnliche Vegetation wie am Ende des DA 1 vermuten lassen. Die Anteile des Ulmus-Typs sind verhältnismäßig hoch, ebenso die Hinweise auf ausgedehnte Röhricht- und Farnbestände.
Von den drei Proben des DA 4 hat nur die mittlere Probe (20 m) einen ausreichenden Pollengehalt. Diese Probe zeigt aber erhebliche Unterschiede zu den übrigen Pollenspektren. So fehlen Picea/Abies, Larix und Pinus. Mit den Nachweisen von Juniperus, Betula und Hippophaë sind vermutlich Hinweise auf Gehölze mit Sträuchern und kleinen Bäumen gegeben. Eine erheblich größere Verbreitung scheinen nun Steppen mit Poaceen und Artemisia gehabt zu haben.
Diskussion. Zur Zeit des DA 1 gab es sowohl Nadelwälder mit Picea, Abies, Larix und Pinus als auch Bereiche mit einer vermutlich wüstenartigen Vegetation mit Ephedra, Nitraria, Chenopodiaceen und Plumbaginaceen (Armeria-Typ).
Nach dem hohen Anteil des GhP zu urteilen, kann der weitere Bereich um den Gaxun Nur nicht völlig waldfrei gewesen sein, wie es TONG GUOBANG & ZHANG JUNPAI (1991) bzw. TONG GUOBANG et al. (1992) bei der Rekonstruktion der Vegetationsverhältnisse verschiedener Zeitscheiben für die Innere Mongolei annehmen. Vielmehr scheinen boreale Nadelwälder von Picea und Abies von Norden her erheblich näher zum Gaxun Nur vorgedrungen zu sein als in der Gegenwart. Sie wurden am Ende des DA 1 etwa von lichter Kieferntaiga und heller Lärchentaiga abgelöst. Die Nadelwälder waren vermutlich auf niederschlagsgünstige Hochlagen von Gebirgen beschränkt, und das weite Tiefland wird von einer halbwüstenartigen Vegetation eingenommen worden sein. Soweit es Steppen gab, werden sie auf eine schmale Höhenstufe beschränkt gewesen sein.
Betula-Arten könnten kleine Laubwaldbestände gebildet haben. Bestände von Betula platyphylla stehen heute in der Äußeren Mongolei am Übergang vom Wald zur Steppe (HILBIG 1995). Von den Vertretern des Ulmus-Typs reicht Ulmus pumila heute in Grund-wassernähe weit in die Wüstengebiete Innerasiens hinein. Hier ist U. pumila vielerorts der einzige Laubbaum und kennzeichnet ehemalige Auenstandorte (WALTER 1974). Die Ulme stößt heute am Gaxun Nur an ihre westliche Verbreitungsgrenze (HILBIG & KNAPP 1983), scheint aber im DA 1 und DA 3 weiter verbreitet gewesen zu sein. Auch Hippophaë und Pterocarya weisen auf eine günstige Wasserversorgung durch Flüsse und zudem auf saisonale Hochwässer hin. Während Hippophaë auf armen Böden an Pionierstandorten gedeiht, steht Pterocarya auf Schwemmböden der Auenbereiche. Die im jüngeren Teil des DA 1 zunehmenden Werte von Nitraria und Hippophaë sowie das Aussetzen von Pterocarya-Funden mögen auf eine voranschreitende Versalzung wassernaher Standorte hinweisen, die sich auch auf die Farnbestände auswirkte. Heute ist das Vorkommen von Pterocarya in China auf den Osten des Landes südlich vom Huang He beschränkt (SCHAAR-SCHMIDT 1988), und die nächsten Vorkommen von Hippophaë liegen im Tibetanischen Hochland bzw. mehrere 100 km entfernt in der Äußeren Mongolei (BAT et al. 1988, LU
RONGSEN 1992, MEUSEL et al. 1978). Angesichts der Verbreitungsmechanismen von Pterocarya (Wind, Fließgewässer) und Hippophaë (Vögel) sowie des Pioniercharakters von

6. Ergebnisse und Diskussionen – Der Gaxun Nur in der Inneren Mongolei 65
Hippophaë kann man das heutige Fehlen beider Gattungen am Gaxun Nur wohl besonders auf einen Mangel geeigneter Standorte mit hoher Flußaktivität zurückführen.
Insgesamt scheint das Vorkommen von Laubgehölzen auf Standorte an Fließgewässern beschränkt gewesen zu sein. Hinweise auf großflächige Laubwälder, vergleichbar mit Quercus-Wäldern wie im heutigen östlichen China, fehlen.
Nach AN ZHISHENG et al. (1988) lagen zumindest während der Bildung des Paläoboden-komplexes S5 (500.000 - 460.000 BP) die Niederschläge im Bereich des Lößplateaus um 250 mm höher als in der Gegenwart. Bei höheren Niederschlägen werden Fremdlingsflüsse stärker in die Wüstengebiete vorgedrungen sein. Zudem lassen die nach JIANG DEXIN (1988) häufigen palynologischen Nachweise von Cedrus im Qaidam-Becken während der sog. Sebei-, der darauf folgenden Senie- (500.000 - 292.000 BP) und selbst noch in der Dabuxun-Palynoflora (292.000 - 15.000 BP) eine Nährung der tibetischen Gletscher durch hohe Niederschläge im Winter/Frühjahr und einen daraus resultierenden höheren sommerlichen Abfluß vermuten. Zugleich spricht aber das Fehlen von Cedrus selbst im klimatisch günstigen DA 1 gegen einen bedeutenden Anteil von Winter- oder Frühjahrs-Niederschlägen im Untersuchungsgebiet am Gaxun Nur.
Das Fehlen von Pterocarya im jüngeren Teil des DA 1 und die zunehmenden Nachweise von Hippophaë, Nitraria, Pinus und Larix können – vorbehaltlich der geringen Zählsummen – mit der von KUKLA & AN ZHISHENG (1989) vom Lößplateau beschriebenen deutlichen Verschiebung zu einem trockeneren und kälteren Klima zusammenhängen. So deutet die Zunahme von Nitraria auf zunehmend sulfatreicheres Wasser hin, und salztolerante Hippo-
phaë-Arten mögen im Verlauf der allmählichen Klimaverschlechterung Pterocarya verdrängt haben (CHEN KEZAO & XU ZHIQIANG 1991, KINZEL 1982, LERCH 1991). Neben klimatischen Änderungen ist bei der Länge der betrachteten Zeiträume auch an neo-tektonische Einflüsse auf Klima- und Wasserhaushalt zu denken.
Der vom DA 1 erfaßte Zeitraum beträgt etwa 500.000 Jahre und dürfte vor etwa 1 Mio. Jahren begonnen haben. In dieser Zeit traten auf dem Lößplateau Zentralchinas mindestens drei ausgeprägte Kaltzeiten auf, die zusammen mindestens die halbe Dauer des DA 1 umfassen (LIU TUNGSHENG et al. 1986). Keines der Pollenspektren zeigt aber ausgeprägt kaltzeitliche Merkmale. Vielmehr entsteht der Eindruck eines allmählichen Überganges von klimatisch weniger günstigen Bedingungen hin zu einem Klimaoptimum und einer erneuten Klimaverschlechterung. Die Klärung dieses scheinbar widersprüchlichen Sachverhaltes bedarf der weiteren Untersuchung mit einer ausreichenden zeitlichen Auflösung.
Während des DA 3 scheinen boreale Nadelwälder bestanden zu haben. Die Hinweise auf Ulmus und auf nasse Bereiche mit Röhrichten sowie das Fehlen von Nitraria deuten auf ein reiches Wasserangebot hin.
Die pollenführende Probe des DA 4 zeigt mit ihren Hinweisen auf eine Steppenvege-tation (hohe Werte von Artemisia, Poaceen) und durch das Vorkommen der heute im

6. Ergebnisse und Diskussionen – Der Gaxun Nur in der Inneren Mongolei 66
Gebiet ausgestorbenen Gattung Hippophaë auf erhebliche Unterschiede zur vergangenen und heutigen Vegetation hin.
Inwieweit der DA 3 mit dem letzten Interglazial und der DA 1 mit dem von PACHUR et al. (1995) vom Gaxun Nur und aus der südwestlich angrenzenden Tengger Shamo beschriebenen Feuchtphase vor dem LGM (s. a. YANG XIAOPING 1991, FANG JINQI 1991) übereinstimmt, muß zunächst offen bleiben.
Insgesamt lassen die einmalige geographische und zeitliche Stellung des Materials (zur Lückenhaftigkeit der bisherigen Untersuchungsdichte siehe in TONG GUOBANG & ZHANG
JUNPAI (1991), Fig. 11.1) und die sich schon jetzt abzeichnenden Hinweise zur Floren-, Vegetations- und Klimageschichte eine detaillierte Untersuchung des Profils lohnend erscheinen.

6. Ergebnisse und Diskussionen – Das Hochland von Tibet 67
6.3 Das Hochland von Tibet
Das Hochland von Tibet umfaßt die Autonome Region Tibet (Xizang) und die Provinz Qinghai. Entsprechend wird von chinesischen Autoren die Bezeichnung Qinghai-Xizang Plateau benutzt. Außerdem gehören Teile des Autonomen Gebietes Xinjiang sowie der Provinzen Gansu, Sichuan und Yunnan zum Hochland. Bei einer Fläche von 2,5 Mio. km2 – etwa 25 % der Fläche Chinas – besitzt Tibet nur 0,8 % der Einwohner und 0,8 % des Ackerlandes Chinas. Die Indus-Yarlung-Zangbo-Sutur (vergl. Kap. 5.1.) trennt Tibet vom Gebirgszug des Himalaja. Zur Abgrenzung gegen die tiefer liegenden Gebiete mag die 3000 m Isohypse dienen. Die durchschnittliche Höhe des Plateaus beträgt über 4000 m und steigt zum abflußlosen, trocken-kalten Qiangtang Plateau auf 4500 - 5000 m an. In den höher gelegenen Bereichen tritt Permafrost bis in 90 m Tiefe auf. Etwa 80 % (47000 km2) der Gletscher Chinas liegen in Tibet (SHI YAFENG et al. 1986, WANG JINTING 1981, ZHAO SONG-QIAO 1986, 1994). Die folgende geographische Gliederung (südöstliches Tibet, südliches Tibet) lehnt sich an ZHAO SONGQIAO an.
6.3.1 Südöstliches Tibet: Der Nianbaoyeze
6.3.1.1 Das Untersuchungsgebiet
Lage und Geologie. Den größten Teil des südöstlichen Plateaus nehmen die meridionalen Stromfurchen oder i. w. S. das Gebiet des Hengduan Shan ein. Der Nianbaoyeze hat mit seiner südlichen Hälfte Anteil an dieser Landschaft. Er liegt im Osten des Bayan Har Shan bei etwa 33°N/101°E an der Provinzgrenze zwischen Qinghai und Sichuan. Die Entwässerung erfolgt im Süden durch tief eingeschnittene V-Täler zum Yangzi und nach Norden durch die weiten flachen Täler einer ehemaligen Einebnungsfläche zum Huang He. Das Gebirge geht auf eine jura- bis kreidezeitliche Granitintrusion zurück und erhebt sich mit seinem höchsten Gipfel von 5369 m Höhe einige hundert Meter über die triaszeitlichen Gesteine des Songpan-Garzè Terrain (CHANG CHENGFA 1996, LEHMKUHL 1995a, LEHMKUHL
& LIU SHIJIAN 1994b, LEHMKUHL & SPÖNEMANN 1994).
Ein rezenter Gletscher (5,1 km2) bedeckt den Gipfel und reicht auf der Nordseite bis 4600 m herab. Die Schneegrenze liegt unterhalb 5000 m. Die Frostschuttstufe ist durch Solifluktion, Gesteinssprengung, Schutt sowie steile Felshänge gekennzeichnet und reicht bis 4400 m herab. Von der pleistozänen Gletschertätigkeit zeugen Trogtäler, erratische Granitblöcke, Moränen und Seen. Die ältesten dieser Spuren sind mehr als 200.000 Jahre alt. Die Schneegrenzdepression des Ximenzuo-Standes zur Zeit des LGM betrug etwa 700 m. Moränen des jüngeren Spätglazials oder des Neoglazials treten um 4100 m und Spuren der Kleinen Eiszeit oberhalb von 4200 m auf (LEHMKUHL 1995a).
Die Böden unterhalb der Frostschuttzone lassen sich entsprechend der vorherrschenden Vegetation als alpine Wiesen- oder Mattenböden bezeichnen (CHEN CUNGEN 1987, ZHAO
SONGQIAO 1994). Nach ZHAO SONGQIAO (1994) und GELLERT (1987) sind für das südöstliche Tibet verlehmte und verbraunte sowie lessivierte Böden mit hoher Basensätti-

6. Ergebnisse und Diskussionen – Das Hochland von Tibet 68
gung anzunehmen. Unter Wald kann es zur Podsolierung kommen (LERCH 1991, LI
WENHUA 1993, WANG LIANGJIAN et al. 1995). Die Untersuchungen von LEHMKUHL (1995a) erbrachten für den Nianbaoyeze dunkle Braunerden mit A-C-Profilen auf 40 - 60 cm mächtigen Sandlössen. Diese sind aus entkalkten und durchschlämmten schwarzerde-ähnlichen Böden hervorgegangen. Ihr C/N-Verhältnis von 10 - 12 entspricht dem von Steppenböden. Gleye, An- und Niedermoore trifft man in Wasserstaulagen an. Oberhalb von 4400 m ist die Bodenbildung nicht über Gesteinsrohböden hinausgegangen (LEHMKUHL 1995a).
Klima. Das Klima an der Ostabdachung Tibets wird vom Monsun beherrscht. Im Winter bringt er die trocken-kalten Winde und Stürme aus dem Inneren Asiens und Sibiriens, im Sommer die feuchten Luftmassen vom Pazifik und teilweise vom Indischen Ozean (DOMRÖS
& PENG GONGBING 1988, WINKLER & WANG PAO K. 1993, ZHANG JIACHENG & LIN
ZHIGUANG 1992). Entsprechend fallen im Nianbaoyeze die Niederschläge überwiegend in den Monaten Mai bis Oktober und sind im Süden des Gebirges höher als im Norden (LEHMKUHL & LIU SHIJIAN 1994b, PÖRTGE 1994). Aus Modellberechnungen ergibt sich ein Anstieg von durchschnittlichen 900 mm in 4000 m auf etwa 1000 mm in 4250 m Höhe (frdl. persönliche Mitteilung Dr. J. BÖHNER, vergl. BÖHNER 1996). Die Mitteltemperaturen des Sommers sind gering. Nordöstlich des Nianbaoyeze in Jiuzhi (3629 m) beträgt die Temperatur im Juli 9,9 °C (JMT 0,1 °C). Durch seine Beckenlage begünstigt, verzeichnet das südöstlich gelegene Aba (3275 m) im Juli 12,5 °C (JMT 3,2 °C). Im Nianbaoyeze dürfte die Temperatur in 4000 m Höhe im Juli bei 8,4 °C und im Januar bei -11,7 °C liegen (JMT -0,7 °C). Für die Obergrenze des Baumwuchses in 4250 m ist in der Vegetationsperiode im Juli eine Temperatur von 7,1 °C bei einer JMT von -2 °C und einer Januar-Temperatur von -13 °C zu erwarten (frdl. persönliche Mitteilung Dr. J. BÖHNER). Die kalten Winter (Januar: Jiuzhi -11,2 °C; Aba -7,9 °C) sind bei monatlichen Niederschlägen von weniger als 10 mm trocken und schneearm. Zudem treten starke Winde aus westlichen und nordwestlichen Richtungen auf (LEHMKUHL 1995a, LEHMKUHL & LIU SHIJIAN 1994b, ZHENG YUANG-CHANG 1994).
Vegetation. Teile von Qinghai und Sichuan gehören der Zone der subtropischen Laub- und Nadelwälder der chinesischen Hochgebirge an (Abb. 7). Unterhalb von 2400 m treten am Westrand des Beckens von Sichuan Laubwälder mit immergrünen Arten auf. In höheren Lagen bis etwa 2800 m sind Wälder aus sommergrünen Laubbäumen und Nadelhölzern vorhanden. Unter den Laubbäumen sind die Gattungen Acer, Tilia, Fraxinus, Zelkova, Quercus und Betula zu nennen.
An feuchten und nebelreichen Talhängen wachsen in der montanen Stufe bis in Höhen um 3000 m Tsuga chinensis und Tsuga dumosa. Kiefern sind mit mehreren Arten der Unter-gattung Diploxylon vertreten. In den subtropischen Tieflagen unterhalb 1600 m wächst Pinus
massoniana, darüber bis in etwa 2800 m P. tabulaeformis und P. yunnanensis. P. densata ist eine Art relativ kalter und trockener Lagen. Sie tritt in großflächigen Reinbeständen in den westlichen Hochgebirgen von Sichuan bis etwa 3500 m auf. Ihre Obergrenze hat sie hier bei 3800 m. Pinus armandii, ein Vertreter der Untergattung Haploxylon, wächst kleinflächig und verstreut in feuchten Lagen bis in 3400 m Höhe.
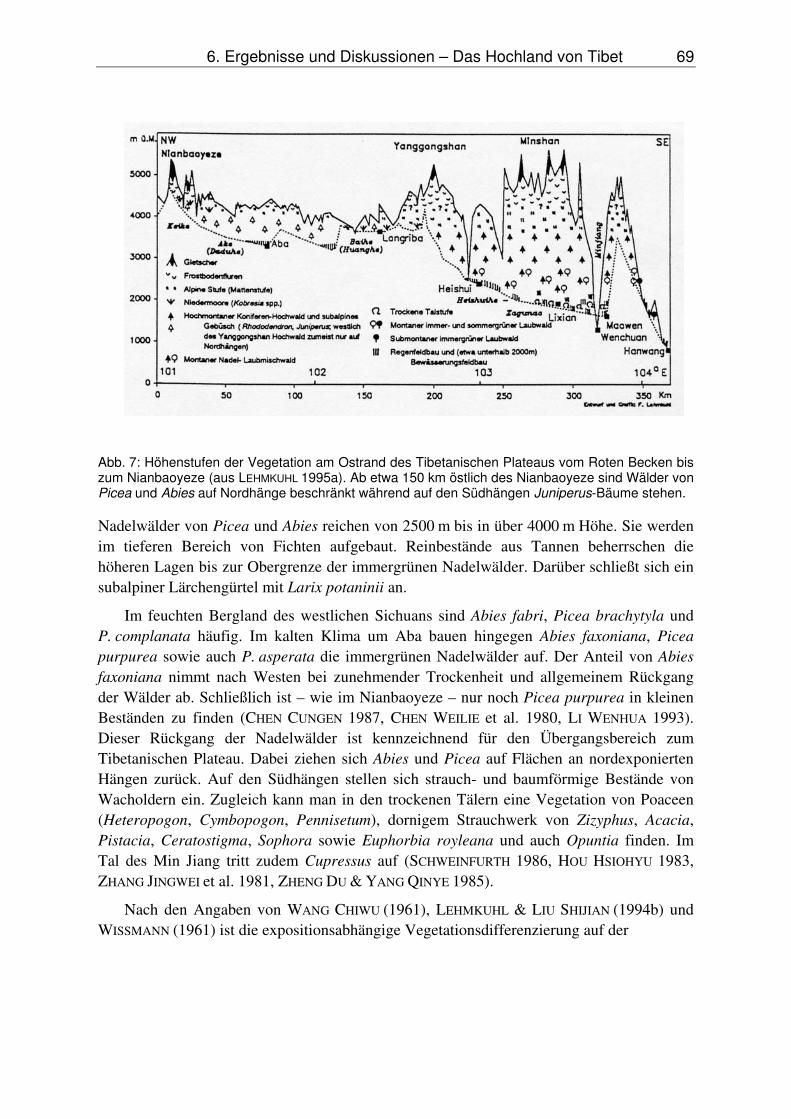
6. Ergebnisse und Diskussionen – Das Hochland von Tibet 69
Abb. 7: Höhenstufen der Vegetation am Ostrand des Tibetanischen Plateaus vom Roten Becken bis zum Nianbaoyeze (aus LEHMKUHL 1995a). Ab etwa 150 km östlich des Nianbaoyeze sind Wälder von Picea und Abies auf Nordhänge beschränkt während auf den Südhängen Juniperus-Bäume stehen. Nadelwälder von Picea und Abies reichen von 2500 m bis in über 4000 m Höhe. Sie werden im tieferen Bereich von Fichten aufgebaut. Reinbestände aus Tannen beherrschen die höheren Lagen bis zur Obergrenze der immergrünen Nadelwälder. Darüber schließt sich ein subalpiner Lärchengürtel mit Larix potaninii an.
Im feuchten Bergland des westlichen Sichuans sind Abies fabri, Picea brachytyla und P. complanata häufig. Im kalten Klima um Aba bauen hingegen Abies faxoniana, Picea
purpurea sowie auch P. asperata die immergrünen Nadelwälder auf. Der Anteil von Abies
faxoniana nimmt nach Westen bei zunehmender Trockenheit und allgemeinem Rückgang der Wälder ab. Schließlich ist – wie im Nianbaoyeze – nur noch Picea purpurea in kleinen Beständen zu finden (CHEN CUNGEN 1987, CHEN WEILIE et al. 1980, LI WENHUA 1993). Dieser Rückgang der Nadelwälder ist kennzeichnend für den Übergangsbereich zum Tibetanischen Plateau. Dabei ziehen sich Abies und Picea auf Flächen an nordexponierten Hängen zurück. Auf den Südhängen stellen sich strauch- und baumförmige Bestände von Wacholdern ein. Zugleich kann man in den trockenen Tälern eine Vegetation von Poaceen (Heteropogon, Cymbopogon, Pennisetum), dornigem Strauchwerk von Zizyphus, Acacia, Pistacia, Ceratostigma, Sophora sowie Euphorbia royleana und auch Opuntia finden. Im Tal des Min Jiang tritt zudem Cupressus auf (SCHWEINFURTH 1986, HOU HSIOHYU 1983, ZHANG JINGWEI et al. 1981, ZHENG DU & YANG QINYE 1985).
Nach den Angaben von WANG CHIWU (1961), LEHMKUHL & LIU SHIJIAN (1994b) und WISSMANN (1961) ist die expositionsabhängige Vegetationsdifferenzierung auf der

6. Ergebnisse und Diskussionen – Das Hochland von Tibet 70
Breitenlage des Nianbaoyeze bereits in den Nebentälern des Min Jiang und östlich des Beckens von Zoige zu beobachten; eine Karte der nordexponierten Wälder (Picea, Abies) gibt WISSMANN (1961). Neben der Trockenheit wirken sich nach TAFEL (1914) auch die winterlichen Weststürme begrenzend auf den Baumwuchs aus.
Der Nianbaoyeze liegt in einem Gebiet, dessen Vegetation überwiegend von alpinen Matten geprägt wird. Cyperaceen, Poaceen, Asteraceen, Ranunculaceen und Fabaceen sind artenreich vertreten. Vorherrschend sind Kobresia pygmaea, K. tibetica, K. royleana, K. humilis, Polygonum viviparum, Thalictrum alpinum, Pedicularis und Poaceen. In nassen Bereichen wachsen Carex, Caltha scaposa, Glyceria aquatica, Utricularia vulgaris und
Potamogeton. Größere Gebüschgruppen treten an Schattenhängen auf. Hier sind Salix
oritrepha, Salix cupularis und Arten der Gattungen Caragana, Potentilla, Sabina, Salix, Spiraea, Sibiraea, Rosa, Sorbus, Rhododendron und Lonicera zu nennen. Hippophaë wächst besonders an immer wieder gestörten Pionierstandorten, wie sie u. a. die Schotterbänke der Flüsse bieten. Die Vegetation der alpinen Matten und Gebüsche kann bis 3500 m
hinunterreichen (HOU HSIOHYU 1983, CHEN CUNGEN 1987, WANG MANHUA 1987).
Bäume sind im Nianbaoyeze selten. Picea purpurea bildet nur kleine Bestände an Nordhängen nahe dem Magenacuo-See (östlich der Gebirgsmitte) und im Kekehe-Tal (Südseite des Gebirges). Diese Vorkommen stellen vermutlich die westliche Verbrei-tungsgrenze von P. purpurea dar. In Südexposition wachsen baumförmige Wacholder. Die Baumgrenze liegt bei etwa 4250 m. Dendrochronologische Untersuchungen an P. purpurea aus 4000 m und an Juniperus aus 4100 m erbrachten Zeitreihen von 264 Jahren bzw. 521 Jahren (BRÄUNING 1994).
Sowohl Fichten als auch die Wacholder bevorzugen im Untersuchungsgebiet Standorte, an denen die ansonsten dichtfilzigen Kobresia-Matten schlecht ausgebildet (Blockhalden, Felshänge) oder aufgerissen (Viehtritt, Erosion) sind (LEHMKUHL 1995a), ganz ähnlich wie dies von MIEHE et al. (1997) bezüglich Juniperus in ganz Hochasien beobachtet und schon von WALTER (1968) für Baumwuchs bei dichter Grasnarbe aus den Steppengebieten berichtet wurde.
Für die offene Vegetation der Frostschuttzone (ab etwa 4400 m) sind Polsterpflanzen charakteristisch. Ihre Arten stammen oft aus den Gattungen Saussurea, Saxifraga, Aconitum, Androsace, Potentilla und den Familien der Caryophyllaceen, Fabaceen, Cyperaceen und Poaceen (HUANG RONGFU 1987, LEHMKUHL & LIU SHIJIAN 1994b, HOU HSIOHYU 1983).
Besiedlung. Nach den prähistorischen Befunden wird für das Tibetanische Hochland eine Besiedlungsperiode im späten Pleistozän angenommen. Im mittleren Holozän weitete sich die menschliche Tätigkeit aus (HUANG WEIWEN 1994). Im Becken von Sichuan ist mit Ackerbau und Beweidung bis in den Bereich der alpinen Matten hinein seit über 2000 Jahren zu rechnen. Um 1650 n. Chr. wanderten die Golog aus Zentraltibet in das Gebiet am Oberlauf des Huang He zwischen dem Anyemaqen Shan und der Provinz Sichuan ein. Die am Anyemaqen Shan ansässigen A rig-Stämme wurden um 1850 von den Golog in das

6. Ergebnisse und Diskussionen – Das Hochland von Tibet 71
Gebiet nördlich des Qinghai Hu verdrängt. Schließlich besiedelten die Wang thag die reichen Weidegründe am Anyemaqen Shan. Sie waren nach eigener Überlieferung vom Osten her eingewandert, von wo sie durch Chinesen verdrängt wurden. Die Umgebung des Nianbaoyeze ist spätestens seit Ende des vorigen Jahrhunderts von starker Abholzung betroffen. Seit den 50er Jahren des 19. Jh. wurde die Entwaldung erheblich beschleunigt (HERMANNS 1949, LEHMKUHL 1995a, b, LEHMKUHL & SHIJIAN 1994b).
Die heutige Bevölkerungsdichte beträgt im Verwaltungsbereich von Aba etwa 3,8 Einwohner pro km2 und in Jiuzhi 1,7 Einwohner pro km2. Im Becken von Aba betreibt man bis in 3300 m Höhe Rotationsbrache mit Gerstenanbau. Die Halbnomaden von Aba nutzen den südlichen Teil des Nianbaoyeze im Sommer als Weidegrund. In der Nordhälfte sind das ganze Jahr über Nomaden mit ihren Viehherden anzutreffen (DAMM 1993, LEHMKUHL
1995a, LEHMKUHL & SHIJIAN 1994b).
6.3.1.2 Rezenter Pollenniederschlag
Oberflächenproben. Aus dem Nianbaoyeze-Gebirge außer den Niedermoorprofilen auch Oberflächenproben zur Verfügung. Für die Ermittlung der heutigen Verteilung des Pollenniederschlages wurden die sechs außerhalb von Vermoorungen gesammelte Moos-polster sowie ein Polster von Arenaria kansuensis MAXIM. (Caryophyllaceae; det. Dr. B. DICKORÉ, Göttingen) untersucht. Die Grundsummen (ohne Caryophyllaceen und Cypera-ceen) der Pollenspektren liegen zwischen 250 und 425 PK (Abb. 8).
Die Nordseite des Nianbaoyeze unterliegt heute einer relativ starken Weidenutzung durch Herden von Nomaden. Die Probe 1 stammt aus einer Höhe von 4300 m von einem Rücken zwischen den Tälern des Ke`erzha und Duomeizhalong. Der Anteil des Gehölzpol-lens (GhP) der Probe hat mit fast 35 % den höchsten Wert aller Oberflächenproben. Neben Juniperus mit 9 % und Betula mit 8 % ist der aus dem Fernflug stammende Pollen von Quercus mit 6 % beteiligt. Picea erreicht wie in den meisten übrigen Oberflächenproben kaum 1 %.
Die Probe 2 wurde am nördlichen Ende eines oberhalb vom Ximencuo-See gelegenen kleineren Sees in 4100 m Höhe genommen. Die Probe 3 stammt aus dem selben Gebirgsteil und gleicher Höhe vom Gletschervorfeld des Nianbaoyeze-Gletschers. Der Anteil des GhP beträgt in beiden Proben weniger als 30 %. In der Probe 2 tritt mit 2 % der geringste Oberflächenwert von Juniperus auf. PK von Rhododendron (3 %) gehen auf nahe gelegene Rhododendron-Gebüsche zurück. In Probe 3 erreicht der sonst gering vertretene Pollen von Picea knapp 4%. Der Anteil der Cyperaceen liegt bei 45 % und 75 %. Poaceen sind mit 20 % vertreten. Artemisia erreicht bis zu 15 %, und der Anemone/Ranunculus-Typ bis zu 18 %.
Drei weitere Proben stammen von der Westseite des Nianbaoyeze aus einem Gebiet, das einer kurzen Beweidung im Sommer unterliegt. Die Proben 4, 5 und 6 wurden südlich des Nianbaoyeze-Gipfels gesammelt. Es stammt die Probe 4 (4380 m) aus der Umgebung des Ma`erdangcuo-Sees, die Probe 5 (4300 m) aus der Nachbarschaft des Emucuogongma-Sees

6. Ergebnisse und Diskussionen – Das Hochland von Tibet 72
und die Probe 6, ein Polster von Arenaria kansuensis, aus der Nähe von Profil 2 (4200 m). Der Anteil des GhP beträgt in 4200 m 21 % bzw. 32 % in 4300 m und 4380 m Höhe. Die Cyperaceen-Anteile nehmen mit steigender Geländehöhe von 95 % auf 120 % zu. Die Prozentwerte der Poaceen gehen hingegen von 25 % auf 13 % zurück. Artemisia erreicht etwa 6 %, der Anemone/Ranunculus-Typ 2,5 %. Der Polygonum bistorta-Typ hat in 4380 m mit 14 % seinen Höchstwert. Trollius tritt in der Probe 6 mit einem hohen Anteil auf (15 %). Der ungewöhnlich starke Caryophyllaceen-Anteil dieser Probe – bezogen auf die Grundsumme ohne Caryophyllaceen 1150 % – stammt sicher in erster Linie aus dem untersuchten Arenaria-Polster selbst. Abgesehen von diesem Extremwert haben aber auch die beiden anderen Proben von der Westseite des Gebirges etwas höhere Caryophyllaceen-Werte als die übrigen Oberflächenproben.
Abb. 8: Pollendiagramm der Oberflächenproben aus dem Nianbaoyeze. Moospolster, Probe 6 Caryophyllaceen-Polster. (GS = GhP + NGhP ohne Caryophyllaceen).
An der Südseite des Nianbaoyeze wurde nördlich von Profil 16 in etwa 4000 m Höhe die Probe 7 entnommen. Das Gebiet wird von Halbnomaden genutzt. Der GhP-Anteil liegt unter 25 %. Cyperaceen und Poaceen sind mit 40 bzw. 15 % nur gering vertreten, was durch den hohen Artemisia-Anteil von 45 % bedingt ist.

6. Ergebnisse und Diskussionen – Das Hochland von Tibet 73
Diskussion. Die überwiegende Menge des Pollens der Oberflächenproben stammt von Cyperaceen und Poaceen. An nicht vermoorten Standorten, wie sie bei der Probenentnahme gewählt wurden, können im Nianbaoyeze offenbar Cyperaceen-Werten von 40 bis 120 % (bezogen auf die GS) erwartet werden. Der GhP erreicht in der GS Anteile von 20 bis 35 %. Die bei Annäherung an die Mattenobergrenze höheren Prozentwerte des GhP sind durch die mit der Meereshöhe abnehmende Pollenproduktion der lokalen Vegetation zu verstehen. Der prozentuale Anteil des vom Wind aus den tieferen Lagen und fernen Gebieten herangewehten GhP steigt dadurch notwendigerweise an.
Während die Werte der Cyperaceen und des Polygonum bistorta-Typs an der Westseite des Nianbaoyeze mit Annäherung an die Frostschuttstufe zunehmen, gehen die Anteile der Poaceen und von Trollius zurück. Der Anemone/Ranunculus-Typ tritt in den Proben von der stärker beweideten Nordseite deutlicher hervor als an der Westseite. Die unterschiedliche Häufigkeit der Pollentypen läßt sich also vermutlich sowohl durch verschiedene klimatische Verhältnisse (Meereshöhe, Geländeausrichtung) als auch durch den Grad der anthropo-zoogenen Beeinflussung zu erklären. Die uneinheitlichen Artemisia-Werte der Nordseite (6 - 15 %) und der hohe Wert der Südseite (45 %) läßt zudem lokale Einflüsse vermuten.
6.3.1.3 Die Profile
Aus dem Nianbaoyeze-Gebirge und seinem Vorland standen 16 Niedermoorprofile für die palynologischen Untersuchungen zur Verfügung. Nach palynologischen Vorunter-suchungen und ersten 14C-Datierungen konzentrierte sich die vegetationsgeschichtliche Bearbeitung im wesentlichen auf fünf Profile. Genauere Angaben zur Herkunft (zumeist frdl. persönliche Mitteilungen von PD Dr. F. LEHMKUHL) und eine Liste der 14C-Datierungen sind dem Anhang zu entnehmen. Profil 3 und Profil 9 aus dem Norden des Gebirges (Abb. 9, in der Beilage) umfassen den größten Teil des Holozäns, zeigen aber bei einer Distanz von nur etwa 7 km deutliche Unterschiede einer kleinräumig differenzierten Vegetationsentwicklung. Noch deutlichere Belege für die Rolle lokaler Einflüsse erbrachte die Bearbeitung von Profil 15 und Profil 16 von der Südseite des Gebirges. Das Profil 14 repräsentiert die Vegetationsentwicklung in dem versumpften Ji’ea-Becken östlich des Nianbaoyeze. Weitere Profile sind im Anschluß kurz angeführt.
Profil 3
Das Profil 3 wurde im Nordwesten des Nianbaoyeze im weiten Flußtal des Lerzha außerhalb des Moränendoppelwalls des Ximentzuo-Standes (LGM) in einem Niedermoor in 4170 m Höhe abgeteuft (Abb. 9, vergl. auch LEHMKUHL 1995a, dort Abb. 18). Der Fluß Lerzha ist etwa 35 m entfernt. Die relativ feuchten Standortbedingungen bieten gute Weide-möglichkeiten, die insbesondere im Winter genutzt werden. Bei einer Gesamtlänge von 3,30 m besteht das Profil überwiegend aus Cyperaceen-Torf. An der Basis tritt eine tonig-schluffige Fein- bis Mitteldetritusmudde auf, deren Bildung etwa 9500 BP begann.
Wegen der Bedeutung des Betula-Pollens im Diagramm wurde eine größenstatistische Artansprache versucht (Abb.10). Der größte Teil der PK ist kleiner als 30 µm und kann keiner der in Frage kommenden Arten Betula utilis, B. platyphylla oder B. albo-sinensis zu

6. Ergebnisse und Diskussionen – Das Hochland von Tibet 74
geordnet werden, deren PK Durchmesser von über 30 µm aufweisen. Möglicherweise geht vom Sediment ein Einfluß auf die PK-Größe aus oder es ist mit anderen als den oben angeführten Arten im Nianbaoyeze zu rechnen. Immerhin deutet eine leichte Zwei-gipfeligkeit der Größenverteilung auf mindestens zwei Betula-Arten hin.
Abb. 10: Größenverteilung der Betula-PK im Profil 3 aus dem Nianbaoyeze. Die PK ab einer Größe von 30 µm dürften von Betula utilis stammen oder aus größerer Distanz angeweht sein (B. platyphylla, B. albo-sinensis). PK unter 30 µm lassen sich, soweit bekannt, keiner Art zuordnen, und ihre geringe Größe ist möglicherweise auf Sedimenteinflüsse zurückzuführen.
Im DA 1 (330 - 295 cm) ist wie im gesamten Diagramm der Anteil des GhP geringer als die Anteile des NGhP und der Cyperaceen (Abb. 11, in der Beilage). Auch wenn die Werte von Picea und Abies unter 5 % liegen, lassen sie doch eine, wenn auch sehr geringe, frühholozäne Verbreitung beider Gattungen im Nianbaoyeze vermuten. In gleicher Weise – im Vergleich mit den Oberflächenproben und den Kurvenverläufen auch der übrigen Diagramme – sind die geringen Prozentwerte von Juniperus als Hinweis auf Vorkommen

6. Ergebnisse und Diskussionen – Das Hochland von Tibet 75
des Wacholders im Nianbaoyeze zu verstehen. Keine der drei Gattungen kann aber besonders häufig gewesen sein. Vermutlich handelte es sich um ähnlich kleine Baumbestände, wie sie stellenweise heute noch in Teilen des Gebirges zu finden sind. Die PK von Pinus, Quercus, Pterocarya, Corylus, Tsuga und Ulmus lassen sich auf Fernflug zurückführen.
Der Anstieg der Hippophaë-Kurve von 5 auf 15 % sowie die vergleichsweise hohen Werte von Salix p. p., des Potentilla-Typs und der Rosaceen p. p. deuten auf Pioniergehölz-bestände hin, wie man sie etwa auf Schotterflächen entlang von Flußläufen erwarten darf. Von dort stammt vermutlich auch ein Teil der Betula-Nachweise. Ursache solcher Standorte wird vermutlich eine hohe Flußdynamik gewesen sein, hervorgerufen durch die zu Beginn des Holozäns schwache Vegetationsdecke und eine Stärkung der Sommermonsuns mit der eine Zunahme der Niederschläge einherging.
Im gesamten Diagramm überwiegt der Polleneintrag der Cyperaceen den aller übrigen Pflanzen. Dies wird durch die vorherrschende Stellung der Gattungen Kobresia und Carex in den das Landschaftsbild bestimmenden Cyperaceen-Matten und in der feuchten Umgebung der Bohrstelle bedingt sein. Von den Kräutern und Stauden des NGhP treten besonders die Familie der Poaceen (über 30 %), die Gattung Artemisia (über 15 %) sowie der Aster-Typ und die Gattung Thalictrum (beide um 5 %) hervor. Die im DA 1 und im DA 2 gefunden Poaceen-PK von über 37 µm Durchmesser sind nicht mit Ackerbau in Verbindung zu bringen, sondern belegen die Existenz wildwachsender Poaceenarten mit PK vom Getreide-Typ (vergl. Kap. 4.4.). Soweit bestimmbar, gehören sie dem Hordeum-Typ an.
Die Pollennachweise vom Myriophyllum spicatum-Typ, von Hippuris und vom Potamogeton/Triglochin-Typ sowie die Funde von Potamogeton-Früchten und die Detritusmudde belegen die Existenz eines frühholozänen Gewässers.
Am Ende des DA 1 fällt die Hippophaë-Kurve stark ab. Ebenso gehen die Nachweise von Salix p. p., des Potentilla-Typs und der Rosaceen p. p. zurück. Die Cyperaceen-Werte nehmen zu. Diese Hinweise auf einen Rückgang der Pioniergehölze und die Ausbreitung von Cyperaceen sprechen für ein erhebliches Nachlassen der Flußdynamik gegen 8900 BP und Verlandung des Untersuchungspunktes.
Der DA 2 (295 - 228 cm) stellt sich mit geringen Hippophaë-Werten als Zeitraum relativ geringer Dynamik dar. Der Rückgang von Salix p. p., des Potentilla-Typs und der überwiegend organogene Sedimentanteil deuten auf eine weitgehend geschlossene Vege-tationsdecke hin. Arten aus den Familien der Cyperaceen und Poaceen dürften die Pionier-gehölze abgelöst haben. Die etwas erhöhten Anteile von Betula mögen z. T. aus Beständen ehemaliger Pionierstandorte stammen. Außer in vorübergehend höheren Trollius-Werten zeigen sich aber kaum Hinweise auf Änderungen in der floristischen Zusammensetzung.
Gegen 7400 BP deuten der Rückgang der Poaceen-Werte und der Anstieg der GhP-Anteile eine Klimabesserung an, die während des DA 3 (228 - 150 cm) in tendenziell

6. Ergebnisse und Diskussionen – Das Hochland von Tibet 76
höheren Werten von Picea und Betula zum Ausdruck kommt. Um 6600 BP hat der GhP mit Werten um 40 % seine größte Häufigkeit. Ein nicht geringer Teil stammt von Pionier-gehölzen (Salix p. p., Potentilla-Typ, Rosaceae p. p.) und aus dem Fernflug (Pinus). Im erneuten Aufkommen von Pioniergehölzen und in dem Rückgang der Cyperaceen und Poa-ceen machen sich Störungen der Vegetationsdecke bemerkbar. Unmittelbar vor Beginn dieser erneuten Pionierholzphase deuten Höchstwerte insektenbestäubter Sippen (Swertia/Lomatogonium-Typ 25%, Liliaceen 15%) auf Änderungen der lokalen Vegetations-verhältnisse hin. Mögliche Ursache könnte eine Erosion der Vegetationsdecke gewesen sein, auf die dann die Pioniergehölze folgten. Die Kurven von Picea, Betula und Juniperus weisen in der selben Probe geringe Werte auf, erreichen aber anschließend wieder ähnliche Anteile wie zuvor, so daß ihr Rückgang überwiegend rechnerisch bedingt ist. Die Werte von Abies bleiben hingegen gering; ihr Rückgang deutet auf eine Abnahme der Tannenbestände hin.
Aus den hohen Picea-Werte um 6600 BP lassen sich relativ günstige Temperatur-verhältnisse ableiten. Die Bildung von Pioniergehölzen spricht für wieder stärker saisonal schwankende Abflußverhältnisse. Beides weist auf die erneute Stärkung des Sommer-monsuns hin. Durch die um einen Meter angewachsenen Torfe mögen aber viele Standorte für Pioniergehölze (Hippophaë, Salix) verloren gegangen sein.
Das Auftreten von Selaginella weist auf jahreszeitlich trockene Standorte hin. Vertreter des Caragana- und des Thymelaea-Typs sowie von Pistacia und Tribulus könnten auf den trockenen Schottern und Sanden des Flußtales gestanden haben. Die Zunahme des Potamo-
geton/Triglochin-Typs spricht vermutlich für die Ausbreitung von Triglochin.
Der Rückgang von Picea, Betula und von Pioniergehölzen deutet bei Zunahme von Cyperaceen und Poaceen auf wieder stabilere Vegetationsverhältnisse gegen Ende des DA 3 unter kühlerem und weniger saisonal geprägten Klima hin. Die Abnahme der zuvor weitgehend unveränderten Werte des Salix oritrepha-Typs spricht für einen Rückgang der subalpinen Gebüsche. Der Polygonum bistorta-Typ und der Typ N1 waren nun stärker vertreten. Die Zunahme des Polygonum bistorta-Typs deutet vermutlich auf kühlere Bedingungen hin (vergl. Oberflächenproben). Auch Vertreter des Caltha-Typs wurden häufiger. Die Bedeutung von Artemisia ging hingegen allmählich zurück.
Im DA 4 (150 - 65 cm) sinkt der Anteil des GhP unter 20 %. Die Anteile von Picea und Abies liegen deutlich unter 5 %. Betula liegt noch bei über 5%. Der Rückgang von Betula sowie die geringe Zunahme von Hippophaë und des Potentilla-Typs gegen etwa 3500 BP lassen eine erneute fluvial bedingte Öffnung der Vegetationsdecke vermuten. Dies könnte auf ein kühleres Klima hinweisen. Die Anteile von Asteraceen (Aster-Typ, Anthemis-Typ, Saussurea-Typ) und Gentianaceen p. p. nehmen ab etwa 3500 BP zu. Vermutlich deuten die Asteraceen ebenfalls auf kühlere Verhältnisse hin (vergl. Diagramm „Lingfeng“ DA 6, Kap. 6.1.2.).
Nach dem Beginn des DA 5 (65 - 0 cm) um 2700 BP bleibt Betula deutlich unter 5 %. Hingegen nimmt der Anteil von Juniperus auf etwa 5 % zu und der Potamogeton/Tri-
glochin-

6. Ergebnisse und Diskussionen – Das Hochland von Tibet 77
Typ steigt auf 10 % an. Der Potamogeton/Triglochin-Typ könnte für eine gewisse Versalzung sprechen.
In den letzten etwa 800 Jahren (interpoliert) geht der Anteil des GhP auf 5 % zurück. Besonders deutlich sind die Abnahme von Hippophaë und das Aussetzen der Picea-Kurve. Die Anteile der NGhP-Taxa verhalten sich in dieser Zeit recht unterschiedlich. Eine Abnahme zeigen Artemisia und der Potamogeton/Triglochin-Typ. Die Anteile des Anemone/Ranunculus-Typs und des Typs N1 nehmen deutlich zu. Die Werte des Saussurea-Typs sowie die Nachweise der bisher seltenen PK von Rheum und vom Rumex-Typ steigen am Ende des DA 5 deutlich an. Nach einer teilweise deutlichen Zunahme gehen die Werte der Poaceen, des Polygonum bistorta-Typs und des Polygonum alpinum-Typs wieder zurück. Die Cyperaceen verhalten sich ähnlich. Die Zunahme der Cyperaceen und Poaceen deuten auf eine Klimaphase mit guten Wuchsbedingungen hin, an die sich bis zur Gegenwart eine Zeit deutlicher Beweidung mit Zunahme von Rheum- und Rumex-Typ anschließt.
Profil 9
Das Profil 9 wurde etwa 7 km nordöstlich vom Profil 3 im Ximencuo-Tal in 4050 m Höhe genommen. Der Bohrpunkt liegt in unmittelbarer Nähe eines kleinen Sees (etwas nördlich des Ximencuo-Sees) an der Innenseite der Haupteisrandlage des letztglazialen Ximencuo-Standes (vergl. LEHMKUHL 1995a, dort Abb. 21). Das Gebiet wird besonders als Sommerweide genutzt. Eine Sommersiedlung befindet sich in der Nähe.
Das Profil ist 143 cm lang und besteht überwiegend aus Cyperaceen-Torfen. Zwischen 95 bis 105 cm sowie unterhalb von 133 cm enthalten sie hohe Schluffanteile. Nach den Datierungen ist die Profilbasis älter als 8000 BP (extrapoliert). Das Pollendiagramm (Abb. 12) setzt somit im Zeitraum des DA 2 von Profil 3 ein.
Im DA 1 (140 - 95 cm) erreicht der GhP einen Anteil von etwas mehr als 10 %. Die Betula-Werte schwanken stark. Picea und Juniperus sind schwach vertreten.
Der Anteil der Poaceen liegt um 40 %. Ab der Mitte des DA 1 sind Poaceen-PK mit einem Durchmesser von 37 µm und mehr vorhanden. Weder ihre frühen Funde im DA 1 noch die späteren im DA 3 können aber mit Ackerbau in Verbindung gebracht werden (vergl. auch Profil 3). Getreideanbau (Hordeum) gibt es heute erst im Becken von Aba in einer Entfernung von über 70 km und nur unterhalb von 3400 m. Die Prozentwerte der Cyperaceen steigen von 90 % auf 200 - 300 % an. Zugleich gehen die Nachweise der Algengattungen Pediastrum und Botryococcus sowie die minerogenen Sedimentanteile stark zurück und die Pollenführung wird besser. Anschließend sind die Nachweise von Pediastrum gering und setzen am Ende des DA 2 aus. Diese Entwicklung deutet auf Ver-landungsvorgänge hin. Die im Diagramm teilweise stark vertretene Algengattung Botryococcus (6 - 900 %) läßt relativ nasse Standortbedingungen vermuten. Die im DA 1 und auch noch im DA 2 und DA 3 gefundenen PK der windblütigen Gattung Myriophyllum dürften aus den benachbarten Seen stammen. Gegenüber den Cyperaceen und den Poaceen

6. Ergebnisse und Diskussionen – Das Hochland von Tibet 78
ist der Polleneintrag anderer Sippen gering. Nur Artemisia ist mehrfach mit Anteilen von mehr als 10 % vertreten.
Abb. 12: Pollendiagramm des Profils 9 aus dem nördlichen Nianbaoyeze, Ximencuo-Tal, 4050 m. ü. M. Stratigraphie: an der Basis Schluffmudde, darüber Cyperaceen-Torf, bei 1 m stark schluffig. (GS = GhP + NGhP).
Insgesamt weist der DA 1 nur geringe Unterschiede zum DA 3 und den Oberflächen-proben auf. Alpine Matten und feuchte Standorte wurden auch im frühen Holozän von Cyperaceen und Poaceen beherrscht. Nur Betula war häufiger als in der Gegenwart.
Im DA 2 (95 - 55,5 cm) erreicht der GhP Werte von über 30 %. Stärker vertreten als im DA 1 sind Salix (18 %), der Potentilla-Typ (11 %) und Hippophaë (3,5 %). Ihre Zunahme deutet die Ausdehnung von Pioniergehölzen an, in denen weitere Rosaceen (Rosaceen p. p., Sibiraea-Typ) vorkamen. Außer Gebüschen scheinen auch Bäume (Picea) häufiger gewesen zu sein. Die etwas erhöhten Brassicaceen-Werte und das Einsetzen der Kurve des Bupleurum-Typs zählen zu den wenigen Hinweisen auf Veränderungen der Kraut- und Staudenschicht.
Gegen Ende des DA 2 nimmt der Anteil des GhP ab. Während der Rückgang von Picea und Betula am ehesten durch rückläufige Temperaturen zu erklären ist, kann die Abnahme der auf Störungen der Vegetationsdecke angewiesenen Pioniergehölze auf schwächere sommerliche Abflußereignisse zurückgeführt werden. Beides dürfte mit dem Nachlassen des Sommermonsuns zusammenhängen.

6. Ergebnisse und Diskussionen – Das Hochland von Tibet 79
Im DA 2 stellt sich der Zeitraum von etwa 5000 bis 2000 BP also als günstiger für Gehölzwuchs dar als der DA 1. Dies weicht von den am Profil 3 gewonnenen Ergebnissen ab (Diskussion in Kap. 6.3.1.4.).
Im DA 3 (55,5 - 0 cm) ist der GhP-Anteil mit weniger als 10 % geringer als im DA 1 und DA 2 vertreten. Das geht neben dem Rückgang verschiedener Gehölzsippen besonders auf die Abnahme der Betula-Werte auf unter 2 % zurück. Fernflug von Pinus ist zunächst mit ähnlichen Werten vorhanden wie zuvor. Die wenigen PK von Abies dürften auf Weitflug zurückgehen.
Gegen 1300 BP lassen die Zunahme der Cyperaceen und des Polygonum bistorta-Typs kühl-feuchte Verhältnisse vermuten. Bestätigung findet diese Annahme im Vorstoßen des Nianbaoyeze-Gletschers und der Datierung des dabei überfahrenen Bodens auf 1385 ± 170 (LEHMKUHL 1995a u. BRÄUNING & LEHMKUHL 1996: Hv 18029).
Um 900 BP gehen die Pinus-Anteile zurück, und die Cyperaceen sowie der Polygonum bistorta-Typ und Typ N1 erreichen ihre Höchstwerte. Am Ende des DA 3 sinken die Werte der Cyperaceen und Polygonaceen dann wieder ab. Hingegen steigen die Anteile von Juniperus, vom Potentilla-, Anemone/Ranunculus- und Riccia-Typ in den jüngsten Proben an. Auch Trollius und Koenigia zeigen höhere Werte. Während die Zunahme der Cypera-ceen und Polygonaceen durch geänderte Klimaverhältnisse zu erklären sind, dürfte bei ihrem Rückgang und gleichzeitiger Zunahme von Juniperus etc. ein zunehmender anthropo-zoogener Einfluß (Weide) von Bedeutung gewesen sein.
Profil 15
Das Profil 15 wurde an der Südseite des Nianbaoyeze in einem kleinen Trogtal im Einzugs-gebiet des Kekehe-Flusses innerhalb des maximalen Standes der letztglazialen Vereisung in 4130 m Höhe entnommen. Das Profil ist 60 cm lang und reicht nach den 14C- Datierungen vermutlich bis 5300 BP zurück. Heute treten am Untersuchungspunkt Sträucher aus den Gattungen Potentilla und Salix auf. Größere Gebüsche und Nadelbäume stehen erst in etwa 3 km Entfernung am Kekehe.
Im DA 1 (60 - 40,5 cm) erreicht der GhP Werte bis 40 % (Abb. 13). Während die Anteile von Juniperus und Picea auf über 5 bzw. 10 % ansteigen, gehen die zu Beginn noch hohen Werte von Salix p. p. (12 %) und vom Salix oritrepha- sowie Potentilla-Typ deutlich zurück. Die Anteile von Abies (2 %) und Betula (4 %) ändern sich nur wenig. Der Sibiraea-Typ, Hippophaë, Rosaceen p. p. und Rhododendron sind nur durch wenige PK vertreten. Die Werte der Cyperaceen liegen zu Beginn unter 100 %, steigen aber auf über 200 % an.
Abnehmende Anteile von Salix und Rosaceen (Potentilla-, Sibiraea-Typ) sowie die Zunahme der Cyperaceen deuten auf die Verdrängung subalpiner Gebüsche durch eine Sumpf- und Matten-Vegetation hin. Die Zunahme von Picea und Juniperus dürften auf eine Ausbreitung der Gattungen auf einem Teil der Gebüschstandorte zurückgehen.
Den Anfang dieser Vegetationsveränderung mögen Erosionsvorgänge vor der Profil-bildung gebildet haben, die die vorübergehende Ansiedlung der Sträucher ermöglichten.

6. Ergebnisse und Diskussionen – Das Hochland von Tibet 80
Abb. 13: Pollendiagramm des Profils 15 aus dem südlichen Nianbaoyeze, Einzugsbereich des Kekehe, 4130 m. ü. M. Stratigraphie: Cyperaceen-Torf. (GS = GhP + NGhP).
Während Picea und Juniperus, vermutlich auch Abies, in kleinen nahegelegenen Baum-beständen standen, stammen die Nachweise von Pinus, Quercus und Ulmus (später auch Pterocarya) aus dem Fernflug. Gegen 2700 BP kennzeichnet der Rückgang von Picea, Abies und Juniperus das Ende des mittelholozänen Klimaoptimums.
Im DA 2 (40,5 - 0 cm) liegt der Anteil des GhP deutlich unter 20 %. Die Werte des Anemone/Ranunculus-, des Polygonum bistorta-, des Aster-Typs und von Trollius, Gentianaceen und Apiaceen sind deutlich höher als im DA 1. Die Zunahme der Werte dieser insektenbestäubten Sippen weist auf lokale Veränderungen der Vegetation hin, die mit dem Rückgang der Sträucher und der Ausdehnung der Cyperaceen-Bestände im DA 1 begannen. PK von Saxifraga stellaris-, Caltha-, Swertia-, Pedicularis oederi- und Primula farinosa-Typ sowie von Crassulaceen stammen von nassen, quellflurartigen Standorten.

6. Ergebnisse und Diskussionen – Das Hochland von Tibet 81
Im jüngeren Teil des DA 1 können einige Veränderungen durch anthropo-zoogene Einflüsse erklärt werden. Hierzu gehören u. a. die ab etwa 800 BP (interpoliert) vergleichs-weise hohen Werte der Chichorioideen und Chenopodiaceen sowie der Anstieg von Juniperus und Apiaceen, der auf Weidetätigkeit hinweist. Juniperus-Arten sind durch Inhaltsstoffe und z. T. auch Nadeln vor Verbiß geschützt. Auch verschiedene Apiaceen schützen sich durch giftige Verbindungen. Einjährige Vertreter von Chenopodiaceen sowie von Koenigia weisen außerdem auf eine Öffnung der Vegetationsdecke hin. Vielleicht gehen die Nachweise von Chenopodiaceen und die hohen Werte von Botryococcus auf einen Nährstoffeintrag durch Vieh zurück.
Die abfallenden Werte von Cyperaceen und Poaceen deuten auf eine Überbeweidung hin, die auch den Rückgang von Artemisia und vom Polygonum bistorta-Typ verursacht haben mag. Die Bedeutung von Ranunculaceen (Anemone/Ranunculus-Typ, Trollius) scheint dagegen unter dem Weidedruck zugenommen zu haben. Das Auftreten von Rosaceen (Potentilla-, Sibiraea-Typ, Rosaceen p. p.) kann auf eine durch Störung der Mattennarbe (Chenopodiaceen, Koenigia) induzierte Verbuschung zurückgehen.
Alle diese Vegetationsänderungen sprechen für eine starke Weidenutzung ab etwa 800 BP (interpoliert). Vielleicht steht auch bereits der Rückgang der Picea-Kurve (gegen 1050 BP) mit menschlichen Aktivitäten in Verbindung.
Profil 16
Das Profil 16 wurde etwa 2,5 km südöstlich vom Profil 15 auf einem flachen Felsterras-senrest außerhalb des Ximencuo-Standes in 3960 m Höhe entnommen. Bei einer Torf-mächtigkeit von nur 80 cm reicht die Überlieferung bis weit in das mittlere Holozän zurück (52 - 67 cm: 5400 ± 95 BP). Gebüsche und Bäume finden sich heute erst in etwa 500 m Entfernung am Kekehe.
Der DA 1 (80 - 45,5 cm) weist GhP-Werte von weniger als 6,5 % auf (Abb. 14). Ge-büsche oder Bäume werden daher im mittleren Holozän wohl kaum hier gestanden haben. Anfangs hohe Poaceen-Werte (über 60 %) gehen bei der Zunahme der Cyperaceen-Anteile (von 65 % auf 200 %) zurück. Die Ausbreitung der Cyperaceen dürfte als Folge des feuchteren Klimas im mittleren Holozän zu verstehen sein.
Im Verlauf des DA 2 (45,5 - 0 cm) steigt der Anteil des GhP auf etwa 15 % an. Juniperus-Werte von 7 % und die Zunahme des Anemone/Ranunculus-Typs, der Apiaceen und von Trollius deuten auf Beweidung hin. Die nach 700 BP und um 2700 BP vorübergehend höheren Werte der Cyperaceen und Poaceen mögen klimatisch, etwa durch ein stärkeres Feuchteangebot, bedingt sein. Für den jüngsten Rückgang von Cyperaceen und Poaceen wird neben klimatischen Änderungen wohl auch Beweidung verantwortlich sein.
Profil 14
Das Profil 14 (Abb. 15) wurde östlich des Nianbaoyeze im Ji’ea-Becken in 3870 m Höhe genommen. Das Gebiet befindet sich außerhalb der eiszeitlichen Vereisungen des Nianbao-

6. Ergebnisse und Diskussionen – Das Hochland von Tibet 82
yeze. Im Frühjahr und Herbst wird das kleine Becken von Nomaden auf ihrem Weg von und zu den Weideplätzen des Nianbaoyeze für mehrere Wochen als Weidegrund benutzt.
Abb. 14: Pollendiagramm des Profils 16 aus dem südlichen Nianbaoyeze, Einzugsbereich des Kekehe, 3960 m. ü. M. Stratigraphie: Cyperaceen-Torf. (GS = GhP + NGhP).
Der GhP-Anteil erreicht im DA 1 (90 - 53,5 cm) kaum über 5 %. Regelmäßig vertreten sind nur Picea, Juniperus und Betula sowie Pinus. Angesichts der geringen Werte können Baumbestände im Bereich des Ji´ea-Becken nur sehr unbedeutend gewesen sein, sofern sie überhaupt auftraten. Die Poaceen erreichen zu Beginn mehr als 40 %, und die Cyperaceen sind mit über 100 % vertreten. Häufige Taxa des NGhP sind Thalictrum, der Anemone/Ranunculus- und der Polygonum alpinum-Typ. Sanguisorba filiformis, in keinem anderen Profil vom Nianbaoyeze gefunden, ist von Beginn an mit etwa 2 % vertreten. Eine Abnahme der Poaceen und ein Anstieg der Cyperaceen-Kurve sowie der Rückgang von Picea um etwa 2750 BP korrelieren gut mit den Ergebnissen aus Profil 3 und Profil 15 im Hinblick auf eine Klimaverschlechterung.
Am Übergang zum DA 2 ändert sich die Häufigkeit einer ganzen Reihe von Pollentypen. So gehen die Werte der Poaceen und die Anteile des Anemone/Ranunculus-, Polygonum alpinum- und Polygonum bistorta-Typs sowie die Anzahl der Farnsporenfunde zurück. Die Kurven der Valerianaceen und des Anthemis-Typ setzen aus. Eine Zunahme der Anteile ist

6. Ergebnisse und Diskussionen – Das Hochland von Tibet 83
bei Artemisia, Trollius, Liliaceen, Apiaceen p. p. sowie bei den Chenopodiaceen zu verzeichnen, und es setzen Nachweise des Bupleurum-Typs ein. Mögliche Ursache dieses Vegetationswandels könnte eine zunehmende Vernässung (Sanguisorba filiformis, Cypera-ceen) des Beckens sein, die bei einer Alterstellung von etwa 1850 BP vermutlich in einen größeren klimatischen Zusammenhang zu stellen ist (vergl. Kapitel 6.3.1.4.), ohne aber einen im Ji’ea-Becken durch Konzentration der Viehherden früh wirksamen anthropo-zoogenen Einfluß ganz ausschließen zu wollen. So weicht beispielsweise Sanguisorba filiformis dem Weidedruck durch seine Kleinwüchsigkeit aus.
Abb. 15: Pollendiagramm des Profils 14 aus dem Ji’ea-Becken östlich des Nianbaoyeze, 3870 m. ü. M. Stratigraphie: Cyperaceen-Torf, im unteren Teil schluffig-tonig (GS = GhP + NGhP).
Die im DA 2 ab etwa 450 BP stärkeren Anteile von Juniperus sind wohl als Folge einer Überbeweidung zu verstehen, zumal der Anemone/Ranunculus-Typ und Sanguisorba filiformis mit hohen Werten auftreten und Nachweise von Rheum einsetzen.

6. Ergebnisse und Diskussionen – Das Hochland von Tibet 84
Weitere Profile
Weitere Profile liegen aus dem mittleren und nördlichen Teil des Nianbaoyeze vor. Auch sie zeigten bei den palynologischen Voruntersuchungen ein Vorherrschen der Cyperaceen, geringe GhP-Werte und hohe Poaceen- und Artemisia-Anteile.
Im Profil 2, nahe des Ekuoduoma-Sees am Westrand des Nianbaoyeze, ist 290 cm lang. Über tonig-schluffigem Sediment folgt ab 130 cm Tiefe ein Cyperaceen-Torf. Dieser Wechsel erfolgte etwa 5000 BP und läßt sich als Anzeichen für ruhigere Abflußverhältnisse nach dem Klimaoptimum werten. Das Profil 8 wurde südlich des Ximencuo-Sees entnommen und besteht aus 91 cm Cyperaceen-Torf der in 50 bis 79 cm Tiefe von einer Gruslage unterbrochen ist. Der untere Torf (80 bis 89 cm) wurde auf 3055 BP datiert. Nimmt man bei der Interpretation der Datierung eine kurze Bildungsdauer des Gruses an, so ergibt sich für die Gruslage ein Alter von etwa 2750 BP. Die Einlagerung des Gruses fällt also zeitlich etwa mit dem vor allem in den Profilen 3 und 15 erkennbaren Beginn einer Klimaverschlechterung zusammen. Vom Oberlauf des Nigequ an der Nordseite des Nian-baoyeze stammt das Profil 11 (Tab. 1, im Anhang). Es ist 254 cm mächtig und unterhalb von 220 cm stark tonig-schluffig. Die Bildung des Torfes begann um etwa 4300 BP (Datierungen vergl. Kap. 11.2). Der Anteil des GhP in den vier untersuchten Proben ist anfangs mit 25 % und später mit 10 % etwa so hoch wie in den vergleichbaren Zeiten im nächstgelegenen Profil 9. Der Bildungsbeginn des 153 cm mächtigen Cyperaceen-Torfes des Profils 12 (Nordostseite des Nianbaoyeze nördlich des Ha’acuo-Sees) steht vermutlich im Zusammenhang mit der schon mehrfach angesprochenen Klimaänderung um 2700 BP. In der jüngsten Probe (3,5 cm) gibt es höhere Juniperus-Werte und geringe Poaceen- und Artemisia-Anteile, d. h. Anzeichen für Beweidung (Tab. 2, im Anhang). Vom Magenacuo-See an der Ostseite des Gebirges stammt das 95 cm lange Profil 13. Der Torf enthält zwischen 85 und 45 cm eine Gruslage, die nach 1380 BP gebildet wurde (Datierung in 85 bis 90 cm).
Holzkohle und Makroreste
Während der palynologischen Untersuchungen der Profile 3, 14, 15 und 16 wurde auch das Auftreten von Holzkohlenflittern erfaßt. Holzkohlenflitter sind im gesamten Profil 3 und Profil 15 vorhanden. Vom Profil 16 enthielten die vier jüngsten Proben (0 bis 16,5 cm) sowie jede zweite der älteren Proben Holzkohlepartikel. Im Profil 14 wurden sie im gesamten DA 2, nicht aber im DA 1 gefunden.
Aus dem Profil 3 wurden zusätzlich Proben von durchschnittlich 35 ml Volumen auf-geschlämmt. Der Bereich 300 - 309 cm besteht aus einer tonig-schluffigen Fein- bis Mittel-detritus-Mudde, in der Steinkerne von Potamogeton gefunden wurden. Die Bereiche 222 - 232, 201 - 209 und 70 - 76 cm bestehen aus Cyperaceen-Torf mit vereinzelten Laubmoos-resten. Es traten weder Funde von Potamogeton noch von Triglochin auf. Alle vier Proben enthielten kleine Holzkohlestückchen.

6. Ergebnisse und Diskussionen – Das Hochland von Tibet 85
6.3.1.4 Diskussion
Älteres Holozän (ca. 10000 - 7500 BP)
Die Überlieferung der holozänen Vegetationsgeschichte des Nianbaoyeze beginnt im Profil 3 um 9600 BP. Bis 8900 BP deutet die Existenz von Pioniergehölzen (Hippophaë etc.) auf saisonal stärkere Wasserführung der Flüsse hin. Als Ursache kann eine frühholozäne Stärkung des Sommermonsuns angenommen werden, die zu erhöhten Niederschlägen führte. Ein Einfluß durch Schmelzwasser des Nianbaoyeze-Gletschers ist wegen der abpuffernden Wirkung der eingeschalteten Seen unwahrscheinlich. In Westtibet herrschten ähnliche Verhältnisse. Dort ließ sich aus Seespiegeländerungen und der Vegetationsentwicklung eine frühholozäne Phase höherer monsunaler Niederschläge zwischen 9500 und 8700 BP ableiten (FAN HUI et al. 1996, GASSE et al. 1996). Die Versalzung einiger Seen West- und Zentral-Tibets weist auf ein Nachlassen des Wasserangebots ab etwa 9000 BP hin (CHEN KEZAO et al. 1981).
Der starke Rückgang der Hippophaë-Bestände um 8900 BP markiert im Nianbaoyeze das Ende der ersten holozänen Monsunzunahme. Neben Laubgehölzen kamen seit Beginn des Profils 3 auch Picea und Juniperus im Nianbaoyeze vor. Ab etwa 9200 BP kann ebenso mit dem Vorkommen von Abies gerechnet werden.
Von 8900 bis 7400 BP folgt dann ein Zeitraum, in dem die aus der Pionierphase verblie-benen Sträucher der Gattung Salix und der Familie der Rosaceen von Cyperaceen und Poa-ceen verdrängt wurden. Diese Entwicklung, die auf abnehmende Niederschläge und tiefere Temperaturen hinweist, spricht für einen schwächeren Einfluß des Sommermonsuns. Auch FAN HUI et al. (1996) und GASSE et al. (1996) führen den zwischen 8600 bis 7500 BP niedrigen Seespiegel des Bangong Co (Westtibet) auf verringerte monsunale Niederschläge zurück. Der Rückgang des Torfwachstums im Becken von Zoige ab etwa 8200 BP (THELAUS
1992) und die Unterbrechung von Bodenbildungen in Zentralchina gegen 8100 BP (LIU
TUNGSHENG et al. 1985 in ROST 1997, ZHOU S. Z. et al. 1991) mögen sowohl auf eine Temperaturabsenkung als auch auf geringere Niederschläge zurückzuführen sein.
Mittleres Holozän (ca. 7500 - 2700 BP)
Eine Zunahme von Picea, Abies, Juniperus und Betula ab 7400 BP im Profil 3 weist auf den Beginn einer mittelholozänen Klimabesserung hin. Hohe Picea-Werte und das Auftreten von Pioniergehölzen um 6600 BP markieren das Klimamaximum mit seinen höheren Temperaturen und Niederschlägen. Der Rückgang von Picea und Betula weist dann auf eine Klimaverschlechterung um 5300 BP hin, nach der es in der Umgebung von Profil 3 offenbar keine wesentlichen Gehölzbestände mehr gab.
Das Profil 9 weist zwischen 95 bis 105 cm eine tonig-schluffige Torflage mit geringen Anteilen von Picea und Betula auf. Ihre Bildung dürfte mit dem im Profil 3 erkennbaren Klimarückgang zusammenhängen. Etwa zur gleichen Zeit setzt das Profil 15 ein. Die Dia-gramme beider Profile zeigen, daß es auch nach 5300 BP stellenweise noch Baum- und Strauchbestände gab und daß es vorübergehend zu einer Zunahme von Picea, Juniperus und

6. Ergebnisse und Diskussionen – Das Hochland von Tibet 86
Betula kam. Während aber nach den Ergebnissen von Profil 15 – wie auch von Profil 3 und Profil 14 – die Klimagunst gegen 2700 BP endete, hielten sich Sträucher (Salix, Potentilla-Typ) in der Umgebung von Profil 9 noch bis 2000 BP. Die mit der Klimaverschlechterung einhergehende Bildung der Gruslage von Profil 8 deutet auf Umlagerungsvorgänge hin, die mit offenen Standorten vermutlich eine Voraussetzung für die lange Strauchphase von Profil 9 schufen.
Die Interpolation des Beginns des holozänen Klimaoptimums auf 7400 BP (Profil 3) wird durch die Angaben anderer Autoren bestärkt. So wird für den Beginn des auch als Yali-Periode oder Qilongduo-Intervall bekannten Klimaoptimums oft ein Alter von 7500 BP genannt (HUANG CIXUAN & LIANG YULIAN 1981, LI BINGYUAN et al. 1985, LI TIANCHI 1988, LIN ZHENYAO & WU XIANGDING 1987, SUN XIANGJUN & CHEN YINSHUO 1991b, WANG
FUBAO & FAN 1987, WINKLER & WANG PAO K. 1993). Vom Qinghai Hu (Nordost Tibet) geben KONG ZHAOCHEN et al. (1990) und LISTER et al. (1991) aufgrund von Pollen- bzw. Ostracoden-Untersuchungen den Beginn des Klimaoptimums mit 8000/7500 und 7000 BP an. Palynologische Ergebnisse vom Bangong Co sprechen ab 7200 BP für erhöhte Feuchtigkeit (VAN CAMPO et al. 1996) und Cyclotella-Algenblüten für eine erhöhte Produktivität des Sees (FAN HUI et al. 1996). Der nahe Sumxi Co und andere tibetische Seen erreichten ab etwa 7500 BP ihre größte Ausdehnung (AVOUAC et al. 1996, VAN CAMPO &
GASSE 1993, ZHOU S. Z. et al. 1991). Auch in der Inneren Mongolei und auf dem Löß-Plateau gab es nach 7500 BP ein für Vegetation und Bodenbildung günstigeres Klima (HOFMANN 1993, RHODES et al. 1996, ROST 1997, ZHOU S. Z. et al. 1991).
Der frühe mittelholozäne Klimarückgang um 5300 BP im Profil 3 stimmt mit den An-gaben von WANG FUBAO & FAN (1987), LI TIANCHI (1988) und anderen Autoren überein, wonach die um 7500 BP begonnenen feucht-warmen Klimaverhältnisse gegen 5500 BP nachließen. Dabei ist zu bedenken, daß die GhP-Anteile der Diagramme bei Klimaverschlechterungen wegen des Überdauerungsvermögens der Bäume (HOLTMEIER
1985, WALTER 1974) z. T. erst „verspätet“ reagieren. Einige Seen weisen bereits während oder kurz nach dem Klimaoptimum des Nianbaoyeze um 6600 BP auf einen deutlich nachlassenden Einfluß des Sommermonsuns hin. So stiegt im Baijian Hu (Innere Mongolei) die Salinität bereits ab 6600 BP an (PACHUR et al. 1995), und der Bangong Co im westlichen Tibet begann 6300 BP zu versumpfen (FAN HUI et al. 1996, GASSE et al. 1996). Das Bildungsende der in den Kakitu-Bergen (Qilian Shan) von BEUG (1987) untersuchten Torfe um 6200 BP mag ebenfalls mit abnehmender Feuchtigkeit sowie Wärmemangel zusammenhängen.
Im Profil 3 finden sich während des Klimaoptimums vereinzelte Hinweise auf trockene Standorte (Caragana-Typ, Pistacia, Thymelaea-Typ, Selaginella). Die Zunahme des Potamogeton/Triglochin-Typs könnte auf leichte Versalzung hindeuten. In dem mehr im Einflußbereich des Sommermonsuns gelegenen Zoige-Becken nehmen Nachweise des Potamogeton-Typs ab etwa 5200 BP erheblich zu (FRENZEL 1994).

6. Ergebnisse und Diskussionen – Das Hochland von Tibet 87
In Tibet folgt auf die Yali-Zeit das sogenannte Neoglazial, dessen Beginn sich mit dem Vorstoß des Arza-Gletschers im südöstlichen Tibet auf 3000 BP datieren läßt (HUANG
CIXUAN & LIANG YULIAN 1981, LEHMKUHL 1997a, LISTER et al. 1991, OSMASTON 1989, SHI
YAFENG et al. 1986, WANG FUBAO & FAN 1987, WINKLER & WANG PAO K. 1993, ZHENG
BENXING & SHI YAFENG 1985).
Die im Profil 3 und im Profil 15 mit 2680 bzw. 2660 BP datierten Vegetationsän-derungen sind jünger und mit dem Klimawechsel vom Subboreal zum Subatlantikum zwischen 2750 und 2450 BP zu korrelieren. VAN GEEL et al. (1996) führen zahlreiche europäische und einige weltweite Belege von Klimaänderungen aus diesem nur 90 Kalenderjahre umfassenden Zeitraum an. Weitere Beispiele aus Asien bieten die Unter-suchungen von HAHNE & MELLES (1997) in Sibirien und die eigenen Ergebnisse aus N-Pakistan (Kap. 6.4.).
Jüngeres Holozän (ab ca. 2700 BP)
Innerhalb der jüngsten Diagrammabschnitte zeigen sich Vegetationsänderungen, die sowohl klimatisch als auch anthropo-zoogen bedingt sein können.
Der deutliche Vegetationswandel um 1850 BP im Profil 14 mag im wesentlichen auf eine zunehmende Versumpfung des Ji’ea-Beckens zurückzuführen sein. Jedenfalls lassen Gletschervorstöße in den Jahrhunderten nach 2000 BP ein höheres Feuchteangebot vermuten (LI BINGYUAN et al. 1985, SHI YAFENG et al. 1986, ZHENG BENXING & SHI YAFENG 1985, ZHOU S. Z. et al. 1991). Bei Manzhang, nur 50 km nördlich des Nianbaoyeze beginnt zu dieser Zeit eine Phase mit Solifluktionsaktivitäten (LEHMKUHL 1995a). Da das Becken auf dem Weg zum Nianbaoyeze aber einen wichtigen Weidegrund darstellt, kann die Ausbreitung von Sanguisorba filiformis und anderer Sippen (vergl. Profil 14) auch durch eine vergleichsweise hohe Viehdichte im Becken gefördert worden sein.
Vermehrte Hinweise auf einen anthropo-zoogenen Einfluß finden sich erst ab etwa 800 BP. Die in den Diagrammen zu beobachtende Zunahme der Cyperaceen wie auch gelegentlich der Poaceen mag durch feucht-kühle Klimaverhältnisse bedingt sein. Für die Abnahme der Gräser und einiger weiterer Sippen (Artemisia, Polygonum bistorta-, Potamogeton/Triglochin-Typ) in jüngster Zeit können sowohl klimatische Ursachen als auch ein stärkerer Weidedruck verantwortlich sein.
Sowohl die Klima- als auch Siedlungsgeschichte werden daher in den letzten Jahrhun-derten Einfluß auf die Vegetationsentwicklung genommen haben. Unter den mongolischen Eroberungen des 13. Jahrhunderts und der fortschreitenden Urbarmachung von Ackerland in den Bergregionen während der südlichen Song Dynastie (1129 - 1279 n. Chr.) mögen die Nomaden zunehmend in den Nianbaoyeze gedrängt worden ein. Dies wird sich während der Ming Dynastie (1368 - 1644 n. Chr.) noch verstärkt haben, als chinesische Militärkolonien die Züge der Nomaden am Ostrand Tibets begrenzten. HERMANNS (1949) berichtet von der Einwanderung der Golog an den Oberlauf des Huang He zwischen dem Anyemaqen Shan und der Provinz Sichuan um 1650 n. Chr. Dies war zugleich eine ausgesprochen kühle Zeit (LIN ZHENYAO & WU XIANGDING 1987). In der folgenden Tsing Dynastie (1644 - 1911 n. Chr.) engten immer mehr chinesische Ackerbauern die verfügbaren Weiden in den

6. Ergebnisse und Diskussionen – Das Hochland von Tibet 88
Grenzlanden zu Tibet ein. Um 1850 n. Chr. wurden die A rig-Stämme aus dem Anyemaqen Shan verdrängt und schließlich besiedelten die vor den Chinesen zurückweichenden Wang thag die reichen Weidegründe am Anyemaqen Shan (HERMANNS 1949, ZHAO SONG-QIAO 1994).
Waldverteilung
Die Diagramme belegen kleinräumige Unterschiede der vegetationsgeschichtlichen Ent-wicklung im Nianbaoyeze. Picea und Juniperus (bis 7100 BP wohl auch Abies) wuchsen seit dem frühen Holozän im Nordwesten des Nianbaoyeze (Profil 3). Offenbar fehlte es hier aber bereits nach dem frühen mittelholozänen Klimarückgang um 5300 BP an ausreichend günstigen Standorten, an denen Fichten in größerem Umfang noch hätten gedeihen können. Das schmalere Ximencuo-Tal (Profil 9) bot offenbar bessere Voraussetzungen für Baum- und Strauchwuchs. Die für Baumwuchs besonders geeigneten, nordexponierten Hänge sind im flachen, nach Nordwesten weit offenen Lerzha-Tal kaum vorhanden und zudem den trocken-kalten Winden des Wintermonsuns unmittelbar ausgesetzt. Die vermutlich geringen winterlichen Niederschläge dürften auch keine schützende Schneedecke gebildet haben, wodurch die Wirkung der starken Winde noch verstärkt wurde (Schneegebläse). Vermutlich verschlechterten sich die winterlichen Niederschlagsverhältnisse nach dem mittelholozänen Klimaoptimum erheblich (vergl. die Ergebnisse zur Klimageschichte am Nanga Parbat, Kap. 6.4.).
Die Südseite des Nianbaoyeze scheint trotz ihrer Leelage zum Wintermonsun und dem stärkeren Einfluß des Sommermonsuns keine wesentlich besseren Voraussetzungen für Wälder geboten zu haben als die Nordseite. Das Profil 16 zeigt weder im frühen noch im mittleren Holozän Hinweise auf Baum- oder Strauchbestände auf dem flachen Südhang in seiner Umgebung.
Aus dem nahen Tal von Profil 15 sind hingegen Vorkommen von Bäumen und Sträuchern bis zum Ende des mittleren Holozäns belegt. Hier mögen die geschützte Tallage und Schneeanreicherungen in Leelagen die Wirkung der kalt-trockenen Winter mit ihren Stürmen abgeschwächt haben.
In den Tälern von Profil 9 und Profil 15 wird zudem der Gehölzwuchs gefördert worden sein, da infolge der hohen Reliefenergie der Hänge die Vegetationsdecke stärker einer Erosion ausgesetzt war und immer wieder geöffnet werden konnte. Da Profil 9 und Profil 15 außerhalb, Profil 3 wie Profil 16 innerhalb des im letzten Glazial vergletscherten Bereiches liegen, könnte auch die Qualität der Böden eine gewisse Rolle spielen.
Juniperus kommt im Nianbaoyeze seit dem frühen Holozän vor. Angesichts der Vege-tationsgeschichte des Nianbaoyeze handelt es sich bei den Wacholderbäumen nicht um Ersatzgesellschaften nach Rodungen von Picea- oder Abies-Wäldern, sondern vielmehr um die natürliche Baumvegetation der Südhänge. Die expositionsabhängige Verteilung der Nadelhölzer dürfte primär durch die unterschiedliche solare Strahlungsbilanz der Hänge und die daraus resultierenden kleinklimatischen Unterschiede bedingt sein (TRETER 1996). Besonders ins Gewicht fällt dabei vermutlich die Schneearmut der Winter, die Vegetation der Südhänge nicht schützt und im Frühjahr kaum mit Schmelzwasser versorgt. In der

6. Ergebnisse und Diskussionen – Das Hochland von Tibet 89
wolken- und niederschlagsreichen Vegetationsperiode dürften die Wuchsbedingungen der Hänge nur wenig differenziert sein (MIEHE 1984).
Der Rückgang der Bäume und Sträucher im mittleren Holozän (5300 BP) und am Ende des mittleren Holozäns (2700 BP) korreliert mit Klimaänderungen, so daß ein anthropo-zoogener Einfluß nicht angenommen werden muß. Insgesamt kamen Bäume und Sträucher während des frühen und mittleren Holozän wie in der Gegenwart nur kleinflächig und lokal vor (vergl. Abb. 9). Ausgedehnte Wälder hat es nicht gegeben, wohl aber Baumgruppen und Gebüsche an heute gehölzfreien Standorten.
Neben dem Klima könnte auch Feuer einer der Gründe für die geringen Gehölzvor-kommen im Nianbaoyeze sein. Die Holzkohlenfunde sprechen für immer wieder auftretende Brände. An Waldgrenzstandorten, wie dem Nianbaoyeze, kann selbst Feuer in Abständen von Jahrzehnten oder Jahrhunderten die Situation nachhaltig beeinflussen (HOLTMEIER
1985). Solche Brände mögen durch Funkenflug bei Steinschlägen besonders nach der Frostperiode im trockenen Frühjahr ausgelöst werden. So berichtet CHEN ZHIMING (1996) von Bränden nach Erdbeben infolge von Steinschlag. Die vegetationsarmen nivalen Hochlagen, der anstehende Granit und die tektonischen Aktivitäten (TENG JIWEN & LIN
BANZUO 1984) bieten im Nianbaoyeze die entsprechenden Voraussetzungen. Unter dem kontinental beeinflußtem Klima dürften auch Blitze von Bedeutung sein. GOLDAMMER
(1993) sieht in den montan-borealen Wäldern Nordost-Chinas (Provinz Heilongjiang) natürliche, durch Blitzschläge induzierte Feuerökosysteme, deren Ursprung bis weit vor die erste menschliche Besiedlung zurückreicht.
Nach den palynologischen Daten sind aber wohl die dichtfilzigen Kobresia-Matten der Hauptgrund für die Gehölzarmut des Nianbaoyeze. Ihre Rohhumusböden können von Baum- und Strauchkeimlingen kaum oder gar nicht durchwurzelt werden. So erzwingen sie ein Ausweichen der Gehölze auf Sonderstandorte, etwa auf von Natur aus gestörte Bereiche an Flüssen und auf felsige Hänge, wo die Gehölze mit ihrem tiefreichenden Wurzelwerk den Gräsern überlegen sind und sich keine dichten Kobresia-Matten bilden können. Heute sind die wenigen Vorkommen von Picea und Juniperus im Nianbaoyeze auf solche Hanglagen beschränkt. Gebüsche finden sich entlang der Flüsse und auf offenen Moränenschottern. Wenn Viehtritt die Matten aufreißt werden Sträucher wie z. B. Potentilla fruticosa zu „Weideunkräutern“.
Auf die hemmende Wirkung der rohhumusreichen Kobresia-Matten Hochasiens und die Bedeutung natürlicher und anthropo-zoogener Öffnung für das Auftreten von Juniperus-Bäumen und -Sträuchern weisen MIEHE et al. (1997) hin. Vergleichbare Einflüsse der Wurzelkonkurrenz auf die Gehölzverteilung sind auch anderswo in Zentralasien zu beobachten (WALTER 1968, HILBIG 1995).
Nach den palynologischen Befunden sind der Nianbaoyeze und vermutlich auch weitere Gebiete im Osten Tibets ein von Natur aus durch Kobresia-Matten beherrschter Raum mit

6. Ergebnisse und Diskussionen – Das Hochland von Tibet 90
kleinen Waldbeständen. Ähnliche, nach HOLTMEIER ohne menschlichen Einfluß entstandene Waldgrenzökotone, finden sich auch in anderen Hochgebirgen Asiens und der Welt (HOLTMEIER 1985, 1989).
Nicht zu übersehen ist aber der verheerende Raubbau in den letzten Jahrzehnten an den schon von Natur aus nur geringen Waldbeständen des Nianbaoyeze und anderer Gebiete Ost-Tibets (LEHMKUHL 1995b, WINKLER 1994, 1996).
In einer Landschaft, in der die Waldgrenze nicht eine geschlossene Linie darstellt, sondern in Form von Kobresia-Matten mit Baumgruppen und kleinen Wäldern ausgebildet ist, ist der Rückgang des GhP in Diagrammen unterhalb der heutigen Waldgrenze kein zwingender Beweis für eine anthropo-zoogene Beeinflussung. Bei entsprechendem zeitlichen Kontext zur Klimaentwicklung, wie oben darzustellen versucht wurde, ist von einer klimagesteuerten Reduzierung der waldfähigen Flächen – sei es durch für Gehölze ungünstige Kimaparameter oder dichteren Schluß der Kobresia-Matten – auszugehen.
Pollentypen, die in geschlossenen Waldgebieten bei anthropo-zoogenen Eingriffen als Störungszeiger auftreten, müssen hier äußerst kritisch gewertet werden. Die in Frage kommenden Pflanzensippen finden in einem Waldgrenzökoton wie im Osten Tibets zu jeder Zeit und nicht zuletzt während des Kimaoptimums (höhere Flußaktivität) natürliche Standorte vor. Eine anthropo-zoogene Mitwirkung bei der Vegetationsänderungen im Nian-baoyeze während des mittleren Holozäns ist nicht zu belegen, auch wenn THELAUS (1992) und FRENZEL (1994) bezüglich des Beckens von Zoige zu anderen Annahmen kommen. Hinreichende Hinweise auf eine anthropo-zoogene Beeinflussung treten im Nianbaoyeze erst seit weniger als 1000 Jahren auf.
Während des gesamten Holozäns scheint die Vegetation im Nianbaoyeze nur relativ geringen Änderungen unterlegen gewesen zu sein. Da es nicht zur Ausbildung einer Waldstufe mit Vertretern der Gattungen Pinus oder gar Quercus kam, erscheinen Angaben zur mittelholozänen Temperaturzunahme um mehr 3 °C gegenüber heute für den Nianbao-yeze als zu hoch (LI TIANCHI 1988, SHI YAFENG et al. 1993, WANG FUBAO & FAN 1987, WANG MANHUA 1987, WU XIANGDING & LIN ZHENYAO 1981, ZHENG BENXING & SHI
YAFENG 1985). Eher ist wohl an Beträge von 2 °C oder weniger zu denken (FRENZEL 1994, LIN ZHENYAO & WU XIANGDING 1987, SHI YAFENG & ZHANG XIANGSONG 1995).

6. Ergebnisse und Diskussionen – Das Hochland von Tibet 91
6.3.2 Südliches Tibet: Im Stadtbereich von Lhasa
6.3.2.1 Das Untersuchungsgebiet
Lage und Geologie. Das südliche Tibet wird im Norden durch die westöstlich verlaufenden Höhenzüge des Gangdise Shan und Nyainqentanglha Shan sowie im Süden vom Himalaja begrenzt (Abb. 1). Teile des Gebietes bezeichnet man nach ihrer Lage auch als Tibetischen Himalaja oder Transhimalaja sowie nach der Entstehungsgeschichte als Tethys-Himalaja.
In der oberen Trias zerbrach der Gondwana-Kontinent in die Indische Platte und die Gangdise-Platte. Zwischen beiden entstand die Neo-Thetys. Beim späteren beim Zusam-menschluß entstand entlang der Plattenränder ein Ophiolith-Gürtel. Der Yarlung Zangbo und weiter im Westen der Indus folgen dem Verlauf der dabei entstandenen Grabenstruktur und geben der Indus-Yarlung-Zangbo-Sutur ihren Namen (CHANG CHENGFA 1996, LI TINGDONG
et al. 1986, SHEN XIANJIE 1996, SINHA & UPADHYAY 1995). Lhasa (29°42’N/91°08’E) liegt in etwa 3650 m Höhe am Lhasa He, einem Nebenfluß des Yarlung Zangbo. Das Gelände steigt im Umfeld der Stadt auf über 5400 m an (LIU YANHUA 1992). Hier stehen Schiefer und Sandsteine der Trias und des Jura sowie Granite der späten Kreide und des Tertiärs an. In den Flußtälern lagern quartäre Lockergesteine (CHANG CHENGFA 1996, SHEN XIANJIE 1996, ZHANG RONGZU 1989). Im Gebiet um Lhasa wie im Tibetischen Himalaja treten verbraunte und verlehmte Böden auf, in der Hochgebirgshalbwüste herrscht Frostschuttbedeckung vor (MIEHE 1991, ZHAO SONGQIAO 1986).
Klima. Für Lhasa wird eine JMT von 7,5 °C angegeben. Fast 90 % des Jahresnieder-schlags von nur 88 mm fallen von Mai bis September. Die Monatsmittel liegen in dieser Zeit deutlich über 12 °C. Wärmster Monat ist der Juni mit 15,5 °C. Die relative Feuchte beträgt von Juni bis Oktober zwischen 50 % und 70 %. Nur der Dezember (-1,9 °C) und Januar (-2,3 °C) haben Temperaturmittelwerte unter dem Gefrierpunkt (DOMRÖS & PENG GONGBING
1988).
Vegetation. Im unteren Yarlung Zangbo-Längstal reichen die subalpinen Nadelwälder an feuchten Hängen bis in 4100 m Höhe hinauf. In der mittleren Nebelwaldstufe zwischen 2500 und 3200 m hat Picea likiangensis ihren Verbreitungsschwerpunkt. In ganzjährig feuchten Lagen treten Tsuga dumosa und Tsuga lijiang hinzu. Die Wälder der oberen Nebelwaldstufe werden von Abies delavayi und A. georgei aufgebaut. Im darüber liegenden Krummholzgürtel trifft man verschiedene Rhododendron-Arten sowie Betula utilis an. Zwergsträucher von Rhododendron anthopogon und R. nivale kennzeichnen die untere Mattenstufe (ab 4400 m). Die Vegetation der oberen Mattenstufe wird von ausgedehnten Cyperaceen-Rasen aus Kobresia pygmaea und K. angusta beherrscht. In 5300 m beginnt die nivale Stufe mit ihren Felsspalten und Frostbodenfluren, in der u. a. Arten der Gattungen Saussurea und der Familie der Brassicaceen wachsen.

6. Ergebnisse und Diskussionen – Das Hochland von Tibet 92
Die Vegetation der weniger feuchten Lagen, insbesondere der Sonnenhänge, enthält andere Baum- und Straucharten. Bis in 3400 m Höhe stehen trockene Offenwälder von Pinus
densata. Darüber folgt ein etwa 200 m mächtiger Bereich mit Quercus apuifolioides, an den sich ein Krummholzgürtel mit Juniperus indica anschließt. Zu den Zwergsträuchern der unteren Mattenstufe gehören neben J. indica auch J. squamata und Potentilla arbuscula (LI
WENHUA 1993, MIEHE 1991, MOU XINDAI et al. 1992, ZHANG JINGWEI et al. 1988).
Westlich der Wälder ist der Talgrund des Yarlung Zangbo ausgesprochen trocken und wird von Strauchsteppen mit Sophora moorcroftii, S. viciifolia, Caragana spinifera, Ceratostigma plumbaginoides und C. griffithii sowie von Poaceen aus den Gattungen Aristida, Pennisetum und Orinus beherrscht. Vereinzelt kommen auch Sträucher von Juniperus squamata vor. (LI WENHUA 1993, MIEHE 1991, MOU XINDAI et al. 1992, ZHANG
JINGWEI et al. 1988). Die endemische Cupressus gigantea bildet am Mittellauf des Yarlung Zangbo offene Wäldchen von 10 m Höhe (CHEN WEILIE 1981, LI WENHUA 1993). Entlang der Flußläufe stehen auch Bäume von Populus spp. und Salix spp. Diese sind wohl fast alle angepflanzt und bis auf eine endemische Weidenart vom Menschen eingeführt. (ZHANG
JINGWEI et al. 1988, WARDLE 1981). Bis 4 m hohe Dickichte von Hippophaë rhamnoides sind auf den Überflutungsebenen noch in 4450 m Höhe zu finden (WARDLE 1981).
In etwa 3700 m Höhe beginnt eine Stipa bungeana-Steppe mit Sträuchern von Salix spp., Potentilla fruticosa und Spiraea mollifolia. Die alpinen Matten aus Kobresia pygmaea beginnen in 4400 m und gehen oberhalb von 5100 m in alpine Wüsten mit Arenaria, Androsace, Saussurea und Rhodiola über. Niedrige Büsche von Potentilla fruticosa sind noch in 4900 m Höhe zu finden (ZHANG JINGWEI et al. 1988, WARDLE 1981, MOU XINDAI et al. 1992, LIU YANHUA 1992).
Sumpfige Bereiche werden von Phragmites communis, Acorus calamus, Scirpus sp., Triglochin maritimum, Blysmus sinocompressus, Pedicularis longiflora var. tubiformis, Hippuris vulgaris, Ranunculus (Batrachium) bungei, Potamogeton spp. und Typha latifolia
eingenommen. Im offenen Wassers wachen Potamogeton pectinatus, P. natans, P. crispus, Najas minor, Ranunculus (Batrachium) trichophyllum sowie Utricularia aurea. In den Bulten-Schlenken-Komplexen werden die feuchten Mulden von Triglochin palustris, Ranunculus nephelogenes und Halerpestes sarmentosa eingenommen, während die Bulte von Blysmus sinocompressus und Kobresia littledalei gebildet werden (ZHANG JINGWEI et al. 1988).
Besiedlung. In der Umgebung von Lhasa gehen die Hinweise auf menschliche Sied-lungen (Gebäudereste, Töpferwaren und Tierknochen) bis in das Neolithikum zurück (HUANG WEIWEN 1989). Nach schriftlichen Quellen gibt es Ackerbau in den Hochlagen Tibets seit dem 2. oder 3. Jh. n. Chr. Vom Beginn des 6. Jh. ist überliefert, daß der Pflug von zwei Yaks gezogen wurde und ein Bewässerungsystem bestand (ZHANG RONGZU 1989). 607 n. Chr. kam es zur ersten Einigung der tibetischen Stämme. Als Sitz der Monarchie wurde Lhasa gewählt. Der Hauptausbau des in dieser Zeit begonnenen Potala Palastes

6. Ergebnisse und Diskussionen – Das Hochland von Tibet 93
erfolgte in der Regierungszeit des Fünften Dalai Lama zwischen 1645 und 1693 n. Chr. (GERNET 1994, HAMBLY 1995, PEWE et al. 1995).
Auf den Feldern um Lhasa werden heute bis in über 4000 m Höhe Gerste, Weizen, Raps und Hülsenfrüchte angebaut. Dabei machte die Gerste 1958 etwa 50 % und der Weizen weniger als 10 % der gesamten Erntemenge aus, wobei Gemüse keine nennenswerte Rolle spielte. 1984 lagen die Anteile von Gerste und Weizen bei etwa 20 % und 30 %, die übrige Ernte bestand nun zum größten Teil aus Gemüse (LIU YANHUA 1992). Der höchstgelegene Gersteanbau findet heute in 4750 m Höhe statt (YU XIAOGAN & SUN SHANGZHI 1981). Neben den zahlreichen Beständen von Populus und Salix sind bei Lhasa auch Aprikosen- und Apfelbäume, Juglans cathayensis, Ulmus parvifolia, Cupressus torulosa sowie Thuja orientalis gepflanzt worden (WARDLE 1981). Die häufigsten Herdentiere sind Schafe und Ziegen. Yaks hatten 1981 nur einen zahlenmäßigen Anteil von 16 % (NI ZUBIN 1981, SCHOLZ 1995).
6.3.2.2 Das Profil Lhasa Stadtbereich
Östlich von Lhasa konnte in 3650 m Höhe ein 150 cm langes Profil genommen werden. Die unteren 60 cm bestehen aus einem minerogenem Sediment mit einer geringen Pollendichte, das von organogene Lagen überlagert wird.
Im gesamten Diagramm überwiegt der NGhP (Abb. 16, in der Beilage). Im DA 1 ist der GhP mit kaum 10 % vertreten. Dabei erreicht Hippophaë Werte bis 5 % und Salix sowie Betula etwa 2 %. Die Basisprobe jedoch weist einen hohen Wert des Juniperus-Typs von 12 % auf. Bis in die Gegenwart ist Fernflug von Pinus, Tsuga und Quercus sowie vereinzelt auch von Abies und Picea zu beobachten. Abgesehen von Sanddorn, Weiden und Birken dürfte das Gebiet weitgehend gehölzfrei gewesen sein. Die Poaceen sind mit Werten von 20 bis 50 % stark vertreten. Weitere häufige Vertreter des NGhP sind Artemisia, Cichorioideae, Brassicaceen und der Aster-Typ. Nachweise von Ceratostigma und das niedrige Verhältnis von Artemisia zu Chenopodiaceen (Artemisia/Chenopodiaceen) lassen trockene Verhältnisse annehmen.
Farne (Filicatae monolet/trilet, Polypodium), Farnverwandte (Equisetum, Selaginella) und Lebermoose (Riccia-Typ, Riella, Hepaticeae p. p.) sind durch eine ganze Reihe von Sporenfunden belegt. Dabei weist Selaginella auf saisonal höhere Niederschläge hin, die zur Bildung annueller Wasserkörper ausreichte und in denen sich Rasen von Riella ansiedelten. Die Zunahme des Polygonum persicaria- und des Polygonum sibirica-Typs deuten auf eine Ausdehnung nasser Bereiche hin. Riella und P. sibiricum weisen dabei auf salzige Bedingungen hin. Angesichts der Pollenfunde vom Myriophyllum spicatum- und vom Potamogeton/Triglochin-Typ scheint darüber hinaus auch ein dauerhaftes Gewässer bestanden zu haben.
Einige der Pollentypen weisen auf die Tätigkeit des Menschen hin, so z. B. der Triticum- und der Hordeum-Typ auf den Anbau von Weizen bzw. Gerste. Ein Teil des Hordeum-Typs kann dabei aber auch von den reichlich am Untersuchungspunkt wild wachsenden Poaceen stammen (Kap. 4.5.). Der Polygonum aviculare- und Plantago major/media-Typ sowie

6. Ergebnisse und Diskussionen – Das Hochland von Tibet 94
Koenigia weisen auf eine Trittbelastung der Vegetation und damit wohl auf ein Abweiden der Grasbestände hin.
Die im DA 2 organogenen Sedimente entstanden in einem vegetationsreichen Wasser-körper mit Arten von Myriophyllum, Eupatomogeton und Batrachium sowie mit Nymphaea
tetragona. Auch Hippuris und Utricularia wurden nachgewiesen. Die hohen prozentualen Werte der Poaceen von über 80 % und die hohe Pollendichte von über 25000 PK/cm3 sprechen für einen dichten Grasgürtel am Gewässerrand, in dem vermutlich auch Vertreter des Polygonum bistorta-, Eurumex-, Sparganium- und Urtica dioica-Typs sowie von Valeriana standen. Offenbar entwickelte sich im DA 2 ein teichartiges Gewässer mit einer reichen Wasser- und Ufervegetation. Arten der Gattung Tribulus können als Weidezeiger angesehen werden, deren Trittkletten-Früchte vom dem Vieh verbreitet wurden, das hier nicht nur weidete sondern den kleinen Teich als Tränke genutzt haben dürfte.
Soweit die im Verhältnis zu den Chenopodiaceen deutlich häufiger gewordenen Artemisia-Nachweise nicht allein aus der Nähe des Gewässers stammen, deuten sie auf feuchtere Verhältnisse in der Steppenvegetation hin (VAN CAMPO & GASSE 1993). Wesentlich verändert haben können sich die klimatischen Bedingungen allerdings nicht, liegen doch keine Anzeichen für eine Ansiedlung von Picea- oder Abies-Wäldern vor. Der niedrige Anteil des GhP von kaum 5 % darf aber angesichts der hohen Pollenproduktion der Poaceen nicht über den Fortbestand der bereits im DA 1 bestehenden wenigen Gehölze hinwegtäuschen. Die PK von Juglans mögen auf Pflanzungen von Walnußbäumen zurückgehen.
Am Übergang zum DA 3 sinken die Werte der Poaceen ab, während die Anteile von Chenopodiaceen und Cyperaceen zunehmen. Die Standortbedingungen näherten sich damit wieder denen des DA 1 an. Nymphaea tetragona und Myriophyllum belegen aber zunächst noch den Fortbestand des Teiches, in dessen Verlandungsbereich sich nun Igel- und/oder Rohrkolben (Sparganium-Typ) ansiedelten. Den DA 4 kennzeichnen hohe Salix-Werte (über 35 %) sowie PK von Populus.
6.3.2.3 Diskussion
Eine Extrapolation der 14C-Datierung (575 ± 120 BP für 81 - 86,5 cm) auf das Alter der Profilbasis in 150 cm ist wegen des Sedimentwechsels schwierig. Vermutlich sind weniger als 1000 Jahre der Vegetationsgeschichte überliefert. Für diesen Zeitraum konnten Viehhaltung und der Anbau von Getreide pollenanalytisch nachgewiesen werden. Belege für Nadelwälder wurden nicht gefunden, ob aber der hohe Wert des Juniperus-Typs der Basisprobe auf ehemalige Wacholderwälder zurückgeht, bleibt offen.
An die Teichphase des DA 3 von 1300 bis 1650 n. Chr. schließt sich im DA 4 eine Verlandung des Gewässers an. Die Dauer des DA 3 deckt sich mit der Zeit höherer See-spiegelstände Chinas, die von Fang Jinqi (1993) mit 1250 bis 1650 n. Chr. angegeben wird. Nach Untersuchungen im Qilian Shan am Eis des Dunde-Gletschers sollen auch die Temperaturen von 1250 n. Chr. bis 1360 n. Chr. angestiegen sein, um dann aber bis

6. Ergebnisse und Diskussionen – Das Hochland von Tibet 95
1650 n. Chr. auf ein Minimum zu fallen (YAO TANDONG & THOMPSON 1992). Somit dürfte das üppige Pflanzenwachstum während der Teichphase sowohl durch höhere Feuchtigkeit und anfangs auch durch erhöhte Temperaturen gefördert worden sein. Eine sich darin andeutende Stärkung des Sommermonsuns kann aber insgesamt nur gering gewesen sein, finden sich doch keine deutlichen Hinweise auf eine Änderung der zonalen Vegetation.
Nach Wardle (1981) soll es sich bei den Salix- und Populus-Bäumen weiter Gebiete Südtibets wie bei Lhasa fast vollständig um Anpflanzungen eingeführter Arten handeln. Diese Annahme wird durch die palynologischen Ergebnisse unterstützt, die für eine Anpflanzung von Salix und Populus östlich von Lhasa vor vielleicht 50 Jahren sprechen (DA 4).
6.3.3 Weitere Untersuchungen in Tibet
Aus dem südöstlichen und südlichen Tibet liegen eine ganze Anzahl von Rohhumus-Soden und Niedermoorprofilen vor, von denen bislang erst 2 Sodenprofile untersucht werden konnten
6.3.3.1 Sodenprofile
Die Untersuchung von Sodenprofilen aus den dichtfilzigen Kobresia pygmaea-Matten verspricht Aussagen über ihre Entstehungsgeschichte sowie den Beginn ihres bei Einfluß des Himalaja-Föhn zu beobachtenden Rückgang (MIEHE 1993). Leider hat sich gezeigt, daß eine zeitliche Einordnung mit konventionellen 14C-Datierungen nicht möglich ist (vergl. Anhang).
Soden 3
Die Unterschiede der drei Pollenspektren von Soden 3 aus 5040 m Höhe sind gering (Abb. 17). Der GhP wird von Juniperus (5 - 13 %) und Betula (6 %) dominiert. Die Werte des Potentilla-Typs liegen unter 5 %. GhP-Fernflug von Koniferen und Quercus ist mit etwa 10 % vertreten, was auf die relativ geringe Pollenproduktion nahe der Mattenobergrenze hinweist. Der Rückgang der Prozentwerte und der Pollendichte der Cyperaceen deutet auf eine nachlassende Wüchsigkeit der Kobresia-Matten hin. Auch der Anstieg der Crassulaceen auf 15 % und der Fabaceen auf 5 % sowie PK von Koenigia sprechen sogar für zunehmende Lücken in der Mattennarbe und ein Näherrücken der hochalpinen Steppe und somit für einen Rückgang der Kobresia-Matten.
Soden 18
Das Sodenprofil 18 stammt aus den alpinen Kobresia-Matten aus 4260 m Höhe. Die Werte der Cyperaceen sinken von 70 % auf 45 %, die Werte der Poaceen von 35 % auf 15 % (Abb. 17). Der Potentilla-Typ nimmt hingegen auf 30 % zu, während die Pollendichte absinkt. In der jüngsten Probe ist der Potentilla-Typ dann mit 25 % vertreten und die Pollendichte wieder etwas höher. PK von Koniferen und Quercus stammen aus dem Fernflug und erreichen zusammen etwa 5 %.

6. Ergebnisse und Diskussionen – Das Hochland von Tibet 96
Abb. 17: Pollendiagramme der Soden 3 und 18, südliches Tibet, 5040 und 4260 m. ü. M. Beide Profile stammen aus dichtfilzigen Kobresia-Matten. (GS = GhP + NGhP).
Der Rückgang der Cyperaceen- und Poaceen-Werte bei Zunahme des Potentilla-Typs sowie die geringe Pollendichte deuten auf eine verstärkte Weidenutzung hin, die durch Öffnung der Mattennarbe die Ausbreitung von Potentilla fruticosa-Sträuchern förderte. Der geringe Fernflug spricht für eine flächendeckende Vegetation mit recht hoher Pollen-produktion im Vergleich zu dem 800 m höher gelegenen Soden 3.
6.3.3.2 Weitere Profile
Im Südosten des Tibetanischen Plateaus wurde an der westlichen Grenze der Picea-Abies-Wälder das Profil Lager 16 (3730 m) genommen. Stichprobenartige Pollenanalysen ergaben eine erhebliche Zunahme von Juniperus etwa in den letzten 600 Jahren. Das Profil Ganzi (3550 m) stammt aus einem Gebiet mit Kobresia-Matten unweit der Nadelwälder. Die bisherigen Ergebnisse deuten darauf hin, daß am Untersuchungspunkt Nadelwälder bestanden, die ab ca. 900 BP durch Kobresia-Matten abgelöst wurden.
In den älteren Proben des Profils Malinganko (4050 m) westlich von Ganzi ist die Pollenüberlieferung schlecht. Ab etwa 4000 BP ist die Pollenführung besser, und die Werte der Cyperaceen erreichen 200 %. Die Oberflächenprobe wird zu gleichen Teilen von PK der Cyperaceen und von Sanguisorba filiformis bestimmt. Das Material von Nagar tse 3 am Yamco Yumco-See erwies sich für Pollenanalysen wenig geeignet. Im Profil Nagqu 3, aus der Stufe der alpinen Kobresia-Matten nördlich von Lhasa, beherrschen Cyperaceen mit weit über 500 % die Pollenspektren. Eine eingehende Bearbeitung der Profile könnte sicherlich zur Klärung der vegetations- und siedlungsgeschichtlichen Vorgänge im heutigen Grenzbereich zwischen montanen Nadelwäldern und alpinen Kobresia-Matten beitragen.

6. Ergebnisse und Diskussionen – Nanga Parbat und Karakorum 97
6.4 Nanga Parbat und Karakorum
6.4.1 Das Untersuchungsgebiet
Die Talschaft des Yasin, der Nanga Parbat und die übrigen Untersuchungspunkte sind Teil des „Kulturraum Karakorum“ (Culture Area Karakoram, CAK) und werden seit Jahren u. a. im Rahmen von DFG-Projekten untersucht.
Lage. Das Gebirge des Karakorum erstreckt sich nördlich des Himalaja von 35° bis 36° N und von 72° bis etwa 79° E. Im Westen ist es vom Hindukusch sowie vom Pamir, im Norden vom Kunlun und im Osten von Tibet umgeben (vergl. Abb. 1 und Abb. 18). TAHIRKHELI & JAN (1984) ziehen die Grenze zwischen Karakorum und Hindukusch im Westen der Talschaft Yasin. Den Südosten der etwa 650 km langen Gebirgskette des Karakorum nimmt die Deosai-Ebene ein.
Die von Nordwest nach Südost streichenden Ketten des Karakorum enthalten die weltweit größte Häufung hoher Gipfel. Sechs Berge erreichen annähernd 8000 m Höhe oder sind sogar höher. Die Entwässerung der das Gebirge tief zerfurchenden Täler erfolgt nach Süden über das Indus-Tal. Zwischen dem Boden des Hunza-Tales und dem Gipfel des Rakaposhi (7793 m) beträgt die Höhendifferenz auf einer Entfernung von nur 11 km etwa 5938 m. Dies ist nach PAFFEN et al. (1956) der Ort der größten Reliefenergie der Erde (s. a. GOUDIE et al. 1984).
Mit einer Vielzahl von Gletschern, die gemeinsam eine Fläche von ca. 18000 km2 be-decken, stellt der Karakorum einen der am stärksten vergletscherten Räume außerhalb der Polargebiete dar (LI JIJUN & XU SHUYING 1984, OWEN & DERBYSHIRE 1989, WANG
ZONGTAI & YANG HUIAN 1992, WISSMANN 1960). Für das Pleistozän wurden 3 große Vereisungsphasen nachgewiesen, von denen die mittlere die größte Ausdehnung besaß. Dabei haben nach DERBYSHIRE (1996) die tief reichenden Abtragungen die geomorpho-logischen Verhältnisse soweit verändert, daß für die späteren Vereisungen nur noch kleinere Flächen zur Verfügung standen (s. a. FORT 1996). Im Holozän erfolgten 3 bis 5 Gletschervorstöße, u. a. während des mittleren Holozäns und z. T. in der aus Europa bekannten „Kleinen Eiszeit“. Dabei kann die Reaktion einzelner Gletscher auf Klima-änderungen durch die lokalen Gegebenheiten dem der übrigen Gletscher entgegenlaufen. Zudem ist durch die weit verbreitete Schuttbedeckung der Gletscher ein Abschmelzen oftmals nicht unmittelbar mit einem Rückzug, sondern mit dem Einsinken der Oberfläche unter annäherndem Beibehalten der Gletscherlänge verbunden (DERBYSHIRE et al. 1984, HASERODT 1989, KALVODA 1992, OWEN 1989, RÖTHLISBERGER 1986).
Geologie. Der Südwesten des Karakorum besteht aus dem Karakorum-Block und dem Kohistan-Bogen. Ersterer wird parallel zu seiner Längserstreckung aus drei Gürteln unterschiedlicher Gesteinskomplexe aufgebaut. Auf perm- bis kreidezeitliche Sedimente mit wenigen spätmesozoischen Intrusionen folgen nach Süden Batholithe (Kreide bis Miozän) sowie ein Komplex metamorph überprägter Sedimente, Vulkanite und Plutonite (BELYAEVSKY 1976, LEMENNICIER et al. 1996, ZANCHI & GRITTI 1996). Während für die

6. Ergebnisse und Diskussionen – Nanga Parbat und Karakorum 98
letzten 25 Mio. Jahre im Schnitt eine Hebungsrate von weniger als 0,8 mm/a vermutet wird, nimmt OWEN (1989) für den Karakorum eine gegenwärtige Hebung um 2 mm pro Jahr an.
Die Talschaft des Yasin grenzt an den Hindukusch. Die höchsten Erhebungen liegen über 6500 m. Bei einer klimatischen Schneegrenzlage um 4900 m ist das Gebiet stark vergletschert. Das Tal zeigt mit seiner U-Form, den hängenden Seitentälern und Moränen, die typischen Merkmale eines Gletschertales. Die älteste noch erkennbare Vergletscherung soll vor 50.000 Jahren stattgefunden haben (HUGHES 1984). Die Gletscher reichten bis unter die Ortschaft Gupis herab. Eine vermutlich nacheiszeitliche Haupttalvergletscherung kam bis in 2350 m Höhe in die Nähe des Hauptortes Yasin (JACOBSEN & SCHICKHOFF 1995).
Bei Alambar stehen schwach metamorphe paläozoische Sedimentgesteine an, darunter Kalk, Marmor und auch Dolomit. Ähnliche Gesteine, aber stärker metamorphisiert, sind auch bei Batyatum Jut verbreitet. Zwischen diesen beiden Gebieten treten sowohl Granite als auch Granodiorite auf (BUCHROITHNER 1978, EBBLIN 1988, HUGHES 1984, TAHIRKHELI &
JAN 1984). Soweit ersichtlich, liegt das Nazbar-Tal und mit ihm das Tal des Shukan an der Nahtlinie von Karakorum-Block und Kohistan-Bogen. An dieser Südgrenze der Asiatischen Platte findet man neben den bereits erwähnten Gesteinen auch solche der Yasin-Gruppe (kreidezeitliche Sedimente, Vulkanite) und des Grünstein-Komplexes (grünfarbene, metamorphisiert Wechsellagerung von Vulkaniten mit Sedimenten der Trias) sowie Ophiolithe (HUGHES 1984, RAZA & BENDER 1995, SEARLE 1991, TAHIRKHELI & JAN 1984).
Mit dem Nanga Parbat-Haramosh-Massiv schiebt sich die Indische Platte spornartig von Süden her in den Kohistan-Bogen hinein (BENDER 1995a, DESIO 1976, MILLER 1982). Von Osten kommend fließt der Indus zwischen den mächtigen Gebirgsmassiven des Nanga Parbat und des Haramosh hindurch, um sich unmittelbar westlich am Nanga Parbat entlang nach Süden zu wenden. Da die Himalaja-Hauptkette westlich des Indus kaum noch über 5000 m hinausragt, stellt der Gipfel des Nanga Parbat (8126 m) gleichsam den westlichen Pfeiler der Gebirgskette dar. Im Vergleich zu dieser ist er in relativ schneller Hebung begriffen. Während aber ZEITLER et al. (1982) noch von 7 mm pro Jahr ausgingen, ergaben die neueren Untersuchungen von WHITTINGTON (1996) nur 3 - 4 mm.
Die Märchenwiese liegt nördlich des Gipfels des Nanga Parbat im Rakhiot-Tal, das sich ebenso wie das weiter östlich gelegene Astor-Tal (Profil Sir le Bala) in die Granite und Gneise des Massivs einschneidet. Östlich der Ortschaft Astor verläuft das Astor-Tal streckenweise in den Gesteinen des Kohistan-Bogens (SEARLE 1991, SHAMS 1995).
Eine Aufzählung der zahlreichen Gesteinsarten des Karakorum und des Nanga Parbat würde zu weit führen, zumal sie angesichts der starken tektonischen Einflüsse auf kleinstem Raume wechseln und die dominierenden Vegetationstypen kaum eine Bindung an Substratgrenzen zeigen, sondern vielmehr vom Klima abhängen (SCHICKHOFF 1993).

6. Ergebnisse und Diskussionen – Nanga Parbat und Karakorum 99
Abb. 18: Klimaeinflüsse im Raum Karakorum/Nanga Parbat und den umliegenden Gebieten. Schraffur: waagerecht = subtropisch-mediterraner Klimaeinfluß von Westen, Niederschläge im Frühjahr, senkrecht = Übergangsgebiet, Frühjahrs-Niederschläge höher als Sommer-Niederschläge, diagonal (im Süden) = randtropisch-monsunaler Klimaeinfluß von Südosten, Sommer-Niederschläge mindestens so hoch wie die Frühjahrs-Niederschläge, diagonal (im Nordosten) = zentralasiatisch-kontinentaler Klimaeinfluß von Nordosten, Wintertrockenheit und gradueller Sommer-Niederschlag (nach REIMERS 1992).

6. Ergebnisse und Diskussionen – Nanga Parbat und Karakorum 100
Eine Bodenentwicklung wird durch das steile Relief, die damit verbundenen Umlage-
rungsprozesse und das Klima erschwert. Auf den ebenen Flußterrassen greift oftmals der Mensch mit seinem Kulturland in die natürlichen Prozesse ein (FURRER 1986, GRÜNEBERG
1995).
Klima. Der Gang der Niederschläge wird im nordwestlichen Himalaja, im Karakorum und den angrenzenden Gebirgsräumen durch drei Klimaregime bestimmt (Abb. 18). Lage und Verlauf der Westwinde, des Sommermonsuns und der Strahlströme der höheren Tropo-sphäre werden vom Tibetanischen Hochland beeinflußt. Das Hochland stellt ein mechanisches Hindernis für die Luftmassen dar und trägt durch seine „Heizfläche“ zur Bildung eines den Monsun verstärkenden sommerlichen Hitzetiefs bei.
Im Winter wirkt sich das über dem Tarim-Becken, dem Kunlun Shan und Tibet herr-schende trocken-kalte, zentralasiatisch-kontinentale Klima nach Südwesten bis zum Quertal des Indus aus. Während des Frühjahrs sorgen westliche Tiefdruckausläufer aus dem sub-tropisch-mediterranen Klimabereich über das nördliche Afghanistan hinweg bis in den Pamir, Hindukusch, Karakorum und Himalaja hinein für Niederschläge. Diese bringen bis in die nordindischen Landesteile Jammu und Kaschmir etwa 40 % der jährlichen Nieder-schläge. Die Stadt Kargil, östlich der Deosai-Ebene, liegt von November bis April unter einer dichten Schneedecke.
Im Sommer erhält die Südseite des Himalaja hohe Regenfälle durch den Südost-Monsun. Dieser randtropisch-monsunale Einfluß wirkt bis in den Himalaja und Karakorum, wo er nach dem frühjährlichen Maximum der Niederschläge einen sekundären sommer-lichen Anstieg bringt. Dabei kann das monsunale Wettergeschehen vor der Gebirgshaupt-kette aufgrund seiner geringen vertikalen Erstreckung von etwa 2000 m unmittelbar nur entlang der Täler in das Gebirge vordringen. Jedoch steigen im Vorland des Himalaja feuchtlabile Luftmassen auf, die mit einer Südströmung der mittleren Troposphäre über den Himalaja-Hauptkamm hinweg gelangen, wo ihre Konvektion im Luv der Karakorum-Ketten zu Niederschlägen führt. Diese werden bei Zusammentreffen mit gelegentlichen mediterranen Störungen noch verstärkt. Insgesamt nimmt der Anteil der sommerlichen Niederschläge an der Jahressumme von Südwesten nach Nordosten ab. Mit der in gleicher Richtung ansteigenden Höhenlage der Talböden gehen auch die Temperaturen zurück. So fallen südlich der Hauptkette im Indus-Tal bei Chilas (1260 m) bei einer JMT von 20,2 °C 187 mm Regen. Bunji (1372 m), nördlich des Nanga Parbat, weist 156 mm und 17,8 °C auf. Im weiter nördlicheren Gilgit (1460 m) wurden 132 mm und 15,8 °C registriert. Gupis (2156 m) in der Talschaft Yasin hat einen jährlichen Niederschlag von 126 mm und eine JMT von 12,9 °C. Der Temperaturgradient beträgt im Jahresdurchschnitt 0,77 °C/100 m, im Januar 0,68 °C/100 m und im Juli 0,87 °C/100 m (zumeist über 30-jährige Meßreihen; vergl. REIMERS 1992 sowie WEIERS 1995).
Angesichts der Lage im altweltlichen Trockengürtel erstaunt die geringe Niederschlags-menge an den Meßstationen der halbwüstenhaften Talabschnitte nicht. An den Talflanken steigt die Ergiebigkeit der Niederschläge aber erheblich an. Eine wichtige Rolle bei dieser Verteilung der Feuchte spielen Hangwinde („Troll-Effekt“). Sie setzen mit der nach

6. Ergebnisse und Diskussionen – Nanga Parbat und Karakorum 101
Sonnenaufgang zunehmenden Thermik gegen 10 bis 11 Uhr ein und wehen vom heißen Talboden die kühleren Hänge hinauf. Im Tagesverlauf führt die einsetzende Kondensation zu Wolkenbändern entlang der Talflanken zwischen 3100 und 3500 m. Die aus ihnen fallenden Niederschläge sind eher gering, doch bieten sie der Vegetation einen Schutz vor Sonnenstrahlung und Verdunstung. Zur Talmitte hin verdunstet der Regen in der heißen Luft oft schon vor dem Erreichen des Bodens. Selbst Schlechtwetterereignisse der Hochlagen geben sich oft nur als Spurenniederschläge zu erkennen. An den Hängen des Nanga Parbat fallen bereits ab etwa 3200 m Höhe mehr als 500 mm Niederschlag. Diese Menge wird in semiariden Tälern wie dem des Yasin erst in etwa 3800 m und in noch trockeneren Seitentälern in etwa 4200 m Höhe erreicht. Oberhalb der Schneegrenze fallen am Batura-Gletscher im nördlichen Karakorum über 2000 mm Niederschlag. In den Einzugsgebieten der Flüsse Gilgit, Hunza und Astor kann ab etwa 6000 m Höhe mit einer solchen Nieder-schlagsmenge gerechnet werden.
Die großen vereisten Flächen führen wie im Yasin-Tal und in anderen Tälern zu kata-batischen Gletscherabwinden, die Getreideaussaat behindern und Obstbäume schädigen können (BÖHNER 1996, DING YIHUI 1994, FLOHN 1981, KACHROO et al. 1977, NEGI 1990, 1995, REIMERS 1992, SCHWEINFURTH 1956, WEIERS 1995, u. a.).
Vegetation. Wo an den Hängen mit der Höhe die Feuchtigkeit zunimmt, beginnt ein Vegetationsgürtel mit feucht-temperierten Nadelwäldern. Diese finden sich am Nanga Parbat großflächig zwischen etwa 2800 m und 3700 m Höhe. Sie werden von Pinus wallichiana und Picea smithiana gebildet. Südlich des Nanga Parbat und im Astor-Tal tritt Abies spectabilis hinzu. Auch Juniperus excelsa ist im Bereich der feuchten Nadelwälder anzutreffen. Die in niederschlagsreichen Gebieten wie dem Becken von Kaschmir vertretenen Gattungen Acer, Aesculus, Juglans und Corylus spielen in den Wäldern des Untersuchungsgebietes keine Rolle, sind aber z. T. in dem dem Sommermonsun zugewendeten Kaghan-Tal noch vertreten.
In 3800 - 3900 m Höhe, lokal bei 4250 m, bildet Betula utilis die obere Baumgrenze. Arten von Salix wachsen in diesen subalpinen Wäldern wie auch in der darüberliegenden alpinen Stufe, wo der gelbblühende Rhododendron hypenanthum hinzutritt. Lawinengassen können tief in die feucht-temperierten Nadelwälder hineinschneiden und schaffen dann durch mechanische Belastung und spät ausapernde Schneemassen quasi alpine bis subalpine Verhältnisse. Entsprechend besiedeln u. a. Betula utilis und Weiden diese Schneisen.
Oberhalb des Birkengürtels bildet Kobresia capillifolia geschlossene Matten, in denen Polygonum affine, Gentiana, Pedicularis und Swertia wachsen. Zwischen 4200 und 4300 m
Höhe löst sich die Vegetationsdecke in Flecken auf. Die Schneegrenze liegt bei 4600 m.
Wo an den trockenen Südhängen die Artemisia-Steppe bis zur alpinen Stufe reicht, fehlen Birken, Weiden und Rhododendren. Statt dessen findet man Juniperus excelsa an der Südseite des Nanga Parbat bis in 4200 m Höhe zusammen mit krüppelwüchsigen Bäumen von J. turkestanica sowie niederliegenden Sträuchern von J. nana und J. squamata.

6. Ergebnisse und Diskussionen – Nanga Parbat und Karakorum 102
Unterhalb der feucht-temperierten Nadelwälder schließt eine Artemisia maritima-Steppe an. Ihr Deckungsgrad kann über 70 % betragen. Die allmähliche Umwandlung der Steppe in eine Wüstensteppe der trocken-heißen Talstufe erfolgt bei etwa 2000 m Höhe, wobei die Deckung auf 5-35 % absinkt. Bei den wenigen Gehölzen der Wüstensteppe handelt es sich um Bäumchen von Pistacia integerriana. Daneben kommen Olea ferruginea und Sträucher von Capparis spinosa vor. Die Baumbestände in den Artemisia-Steppen können hingegen das Erscheinungsbild einer Baumsteppe bis hin zu Steppenwald besitzen. In den Tälern des Rakhiot und Astor werden diese Trockenwälder von Pinus gerardiana und Juniperus excelsa gebildet, daneben kommt J. macropoda vor.
Die untersten Vorkommen von J. excelsa beginnen am Nanga Parbat in 2200 m Höhe. Ab 2600 m Höhe erst wird der Wacholder häufiger. Unterhalb der Rakhiot-Mündung tritt in den Trockenwäldern jenseits des Indus Quercus baloot auf. Diese früher mit Q. ilex gleichgesetzte Art dringt im Karakorum bis zur Südabdachung des Rakaposhi vor. Pinus gerardiana reicht in kleinen Beständen bis in das Chaprot-Tal westlich von Chalt. Von hier an ist Juniperus excelsa der einzige Baum der Steppe. Die in nicht zu feuchten Lagen an der Südseite des westlichen Himalaja oberhalb der Trockenwälder waldbildende Cedrus
deodara hat ihre nördlichsten Vorkommen am Indus etwa 20 km flußabwärts des Rakhiot.
Entsprechend dem klimatischen Übergangscharakter des Gebietes zieht sich mit zunehmender Trockenheit der feucht-temperierte Nadelwald beim Vordringen in das Gebirge auf die Nord- und Nordwest-Hänge zurück. An seiner Stelle breitet sich die Artemisia-Steppe aus. Diese expositionsabhängige Differenzierung der Vegetation ist kleinflächig bereits im mittleren Kaghan-Tal südwestlich des Nanga Parbat zu beobachten. Im Hunza-Tal erreichen die nördlichsten Vorkommen von Pinus wallichiana und Picea smithiana das Rakaposhi-Massiv (Abb. 19). Auf der gegenüberliegenden Talseite markieren Bestände von Juniperus excelsa in 3500 bis 3800 m Höhe die äußerste Grenze der feuchten Nadelwälder. Folgt man dem Lauf des Gilgit flußaufwärts in Richtung Yasin, so reicht die Verbreitung von Picea smithiana und Pinus wallichiana südlich bis Singal bzw. zum Einmündungsbereich des Ishkoman bei Gakuch. Pinus wallichiana kommt zudem in kleinen Beständen im Bereich des Ishkomans, teilweise mit Picea smithiana, sowie südwestlich von Gupis vor. Darüber hinaus gibt es im übrigen Kara
korum weitere reliktartige Einzelvorkommen von Pinus, Picea und von Abies, die auf eine vormals größere Verbreitung der feuchten Nadelwälder hindeuten (BRAUN 1996, PAFFEN et al. 1956, SCHICKHOFF 1993, 1995, SCHWEINFURTH 1957, TROLL 1939, 1967).
Im trockenen Haupttal des Yasin sind der Artemisia-Steppe bei nördlicher Exposition oberhalb von 2800 m geschlossene Baumbestände von Juniperus macropoda und J. turkestanica beigemischt. Subalpine Vorkommen von Betula utilis sind auf strenge Nordexpositionen beschränkt. In ihrem Unterwuchs und der alpinen Stufe kommen
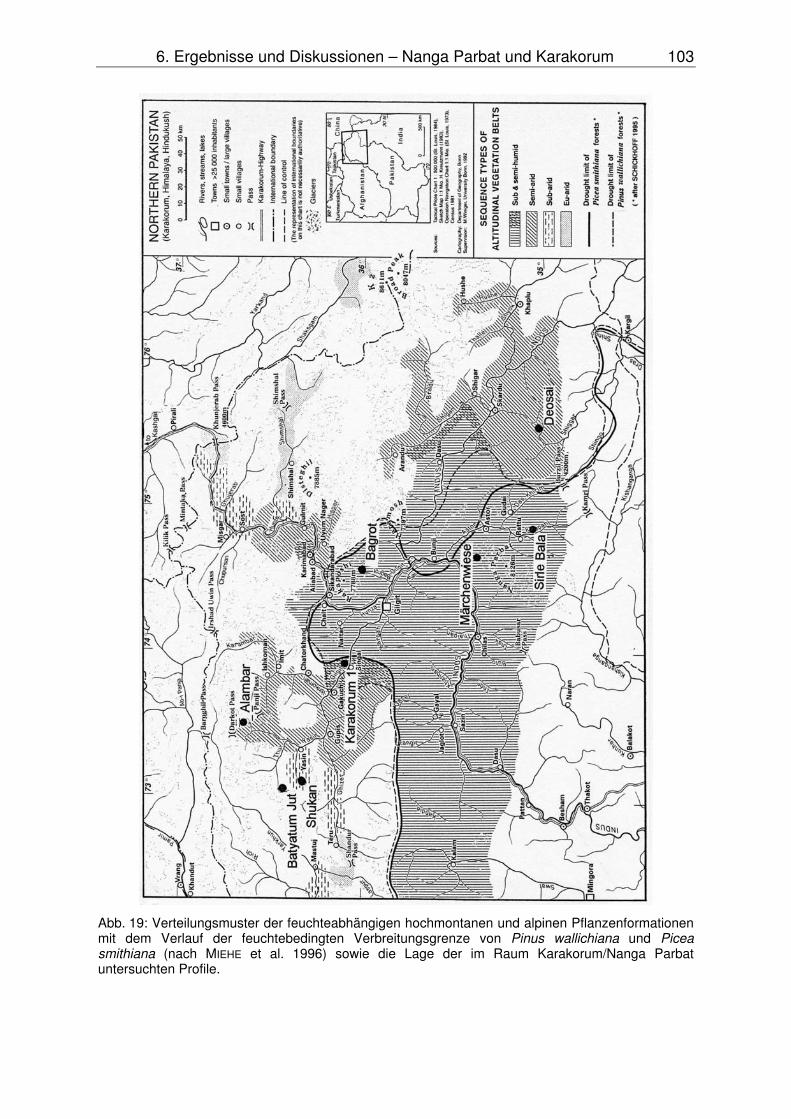
6. Ergebnisse und Diskussionen – Nanga Parbat und Karakorum 103
Abb. 19: Verteilungsmuster der feuchteabhängigen hochmontanen und alpinen Pflanzenformationen mit dem Verlauf der feuchtebedingten Verbreitungsgrenze von Pinus wallichiana und Picea smithiana (nach MIEHE et al. 1996) sowie die Lage der im Raum Karakorum/Nanga Parbat untersuchten Profile.

6. Ergebnisse und Diskussionen – Nanga Parbat und Karakorum 104
niedrige Sträucher von Salix karelinii und Juniperus communis vor. Die geschlossenen Matten von Kobresia capillifolia gehen an den Südhängen in mehr oder weniger offene krautreiche Steppen mit Horstgräsern und Hemikryptophyten über.
In Seitentälern des Yasin, so im Tal des Nazbar und Shukan, stehen die Wacholder-bäume überwiegend in offenen Beständen an steilen Felsklippen. Wälder mit Kronenschluß fehlen. Aufgrund der größeren Trockenheit sind Gehölze der subalpinen Stufe auf Juniperus communis in Nordexposition beschränkt. Betula und Salix sind nur azonal an Standorten mit Wasserzufluß anzutreffen. Vom Talgrund in etwa 2800 m bis in 3800 m Höhe erstreckt sich eine trockene Artemisia-Steppe, die durch häufiges Auftreten von Ephedra und Acantholimon gekennzeichnet ist. Während die Polster von Acantholimon durch ihre Dornen und die vorhandenen Eremurus-Arten durch ihre Inhaltsstoffe gegen Viehverbiß geschützt sind, unterliegen Vertreter der Poaceen und selbst Artemisia maritima der Beweidung. Die alpine Steppe ist krautreich. An den Schattenhängen wird sie von tussockförmigen Poaceen und Cyperaceen sowie Hemikryptophyten aufgebaut. Es wachsen hier Potentilla, Nepeta, Oxytropis und Geranium sowie sukkulente Crassulaceen. An den Sonnenhängen bedeckt die Vegetation oft weniger als 50 % des Bodens. Geschlossene Kobresia-Matten treten nur azonal an feuchten Standorten auf (frdl. persönliche Mitteilung von Dr. S. MIEHE u. Prof. Dr. G. MIEHE, MIEHE et al. 1996).
Besiedlung. Hinweise auf Bewässerungsfeldbau sind im Yasin-Tal auf 2380 ‒ 185 BP datiert (JACOBSEN & SCHICKHOFF 1995). Im etwa 100 km nördlich gelegenen Wakhan-Tal deuten palynologische Untersuchungen auf Ackerbau mindestens seit dem 1. Jh. n. Chr. hin (BORTENSCHLAGER & PATZELT 1978). Das Alter einer ehemaligen Bewässerungseinrichtung im Wakhan wird mit 1350 ± 60 BP, das einer Feuerstelle mit 3750 ± 60 BP angegeben (RAUNIG 1978).
Besonders im 4. und 6. Jh. n. Chr. stand das nördlich des Pamir gelegene Sogdien mit den Handelswegen am oberen Indus über Routen, die durch den Wakhan und die Täler des Karakorum (Hunza, Gilgit, Yasin) führten, in Verbindung (RAUNIG 1983, JETTMAR 1980b, JETTMAR 1993). Diese Täler gehörten auch zu einer südlich von der „Seidenstraße“ zum Indischen Ozean führenden Route, deren Verlauf heute der Karakoram Highway nach-zeichnet. Das im Karakorum gelegene Königreich Bolor teilte sich im 8. Jh. Die Räume Astor und Chilas gehörten zu Groß-Bolor, Gilgit und Yasin zu Klein-Bolor (JETTMAR 1977, 1980a). Im 10. Jh. wurde in der Siedlung Yasin ein buddhistisches Kloster erbaut. Zwischen 1120 und 1160 n. Chr. kam es zur Islamisierung. Im 14. und 15. Jh. stellte Yasin ein selbständiges Königreich dar. Wegen der häufigen kriegerischen Auseinandersetzungen besaß noch im 19. Jh. jede Siedlung ihren Wehrturm (HUGHES 1984). Heute befindet sich in der Yasin-Siedlung das Verwaltungszentrum des Gebietes (KREUTZMANN 1996).
Die Ernährung der Bevölkerung sichern Ackerbau, Viehzucht und Obstanbau. Die zahlreichen Schaf- und Ziegenherden dienen im wesentlichen der Herstellung von Milch-produkten. Schlachtungen erfolgen oft nur in Verbindung mit festlichen Anlässen. Dauer-siedlungen und deren Kulturland liegen in der Artemisia-Steppe. Im Sommer bezieht ein Teil der Bevölkerung höher gelegene Sommerfeldsiedlungen in den Seitentälern, um die dort

6. Ergebnisse und Diskussionen – Nanga Parbat und Karakorum 105
gelegenen Äcker zu bestellen und der Hitze der unteren Talstufen zu entgehen. Die Viehherden werden dabei mitgeführt und im Jahresverlauf weiter zu den Almen aufgetrie-ben, die einen guten Weidegrund bieten. Die Größe der Viehbestände wird durch die winterliche Futterknappheit in den Dauersiedlungen bestimmt. Heute werden auf dem Sommerweiden zusätzliche Herdenzuwächse für den Fernhandel (Karakorum Highway) erwirtschaftet (SNOY 1975, 1993, MÜLLER-STELLRECHT 1979, PAFFEN et al. 1956).
Für den Bau von Häusern werden die vor Ort vorhandenen Materialien Steine und Erde verwendet. Ein weiteres wichtiges Baumaterial ist Holz. Die tragenden Pfeiler und Balken im Inneren der Häuser fertigt man aus dem hochgeschätzten Walnußholz. Dank seiner langen Haltbarkeit kann es mehrfach wiederverwendet werden. Die Wände sind durch den horizontalen Einbau von Kiefern- und Weidenhölzern relativ erdbebenfest (HUGHES 1984).
In weiten Bereichen des Karakorum ist Wacholder die einzige Quelle zur Brennholz-gewinnung. Im Hunza-Tal beschränkt sich die Nutzung aber weitgehend auf das Astwerk. Da Bäume erst im abgestorbenen Zustand geschlagen werden, können sie Alter von über 1200 Jahren erreichen (BILHAM et al. 1984). Juniperus dient zudem der rituellen Beräucherung von Ziegen und Ställen (SNOY 1975).
Die Untersuchungen konzentrierten sich auf den semihumiden Raum des Nanga Parbat im NW-Himalaja und auf subaride Abschnitte der Talschaft Yasin im Westen des Karakorum (Abb. 19)
6.4.2 Die Profile
6.4.2.1 Der Nanga Parbat
In den Tälern am Nanga Parbat liegen die Dauersiedlungen und ihr Kulturland zumeist in der Stufe der Artemisia-Steppe (2000 bis 3000 m Höhe). Das Rakhiot-Tal besitzt mit Tato eine Dauersiedlung, zu der zwei Sommerfeldsiedlungen gehören. Von ihnen befindet sich Tatoniril unweit des Zungenendes des Rakhiot-Gletschers. Die Rodungsfluren reichen bis in 3400 m Höhe in den feuchten Nadelwald hinein. Die Lichtungen der sogenannten Märchenwiese (TROLL 1939) nutzt man als Triftweide. Durch Lawinengefahr und spät ausapernde Schneedecken ist die Nutzung der Sommerfeldsiedlungen auf wenige Monate beschränkt. Im südlichen Bereich der Märchenwiese liegt in etwa 3300 m Höhe eine Lichtung von ungefähr 300 m Länge und 200 m Breite an. Eine ehemalige Seitenmoräne des Rakhiot-Gletschers staut hier zwei Bäche auf, was zu einer teilweisen Vermoorung führte. Diese auf der Karte von TROLL als „Rampe“ vermerkte Moräne ist heute teilweise von Pinus wallichiana und Juniperus–Bäumen bestanden (frdl. persönliche Mitteilung Prof. Dr. M. WINIGER und Dr. U. SCHICKHOFF). Von den entnommenen Profilen wurden die Profile „Märchenwiese Rampe“ und „Märchenwiese Hang“ palynologisch untersucht. Die angegebenen Prozentwerte beziehen sich auf die Summe des Gehölzpollens (GhP = 100%). Im Summendiagramm – zwischen den Kurven des GhP und den Cyperaceen – sind wie bei

6. Ergebnisse und Diskussionen – Nanga Parbat und Karakorum 106
den übrigen holozänen Pollendiagrammen die Anteile des GhP denen des NGhP gruppenweise gegenübergestellt (GhP + NGhP = 100%).
Märchenwiese, Profil Rampe
Im nordöstlichen Bereich der teilweise vermoorten Lichtung wurde ein in seinem unteren Teil holzreicher 3,30 m mächtiger Cyperaceen-Torf mit einem Alter von fast 8000 BP angetroffen.
Der GhP im DA 1 stammt fast ausschließlich von Pinus (Abb. 20, in der Beilage). Von den übrigen Gehölzsippen ist Betula mit Werten bis 4 % am häufigsten. Juniperus, Quercus, Salix und Fraxinus sind gering vertreten. Zudem liegen Einzelfunde von Arceuthobium, Rosaceen, Hippophaë, Ulmus und Ephedra vor. Der Anteil des NGhP ist sehr gering und liegt im Summendiagramm unter 3%. Am häufigsten sind Poaceen, Artemisia und Chenopodiaceen, doch bleiben alle Werte unter 2 % der GhP-Summe. Weitere Sippen wie Ranunculaceen, Crassulaceen, Liliaceen und Koenigia sind nur durch wenige PK nachgewiesen. Auch die Cyperaceen-Werte sind gering. Die Umgebung des Bohrpunktes trägt also seit mindestens 7900 BP einen dichten Wald von Pinus wallichiana, in dem zunächst noch keine Lichtung bestand. Gegen etwa 7200 BP deutet sich im leichten Rückgang von Pinus und in der Zunahme von Juniperus und Artemisia eine Öffnung des Kiefernwaldes an. Die Entstehung der Lichtung dürfte auf eine verstärkte Versumpfung zurückgehen, die zur Zunahme der Cyperaceen-Bestände führte.
Im DA 2 sind die Pinus-Werte geringer und die Cyperaceen-Werte höher als im DA 1. Die aus einigen Kilometern Entfernung stammenden PK von Quercus und Cedrus konnten vermehrt eingeweht werden. Ein PK von Pistacia dürfte aus der trocken-heißen Wüstensteppe stammen. Auch der Pollen von Betula, Salix, Juniperus und Artemisia wird z. T. aus mehr oder weniger entfernten Vegetationseinheiten wie den subalpinen Gebüschen und der Steppe kommen. Vertreter von Betula und Salix werden aber auch im feuchten Bereich der Lichtung, Juniperus sowie Artemisia in ihrem trockenen Randbereich gestanden haben. Die PK der insektenbestäubten Sippen Asteraceen, Apiaceen, Gentianaceen und vom Polygonum bistorta-Typ dürften unmittelbar aus der Pflanzendecke der Lichtung stammen. Die zunehmenden Farnsporen-Anteile (Filicatae monolet) deuten auf erhöhte Feuchtigkeit hin.
Zu Beginn des DA 3 um 6300 BP geht der Pinus-Anteil stark zurück. Die Zunahme von Betula und Salix ist als Hinweis auf Lawinenbahnen zu werten. Lawinen können bis tief in die Stufe der feuchten Nadelwälder hinabreichen, und dort durch mechanische Belastung und spät ausapernde Schneemassen quasi „subalpine“ Verhältnisse schaffen. Im DA 3 scheinen derartige Lawinenbahnen bis zur Lichtung vorgedrungen zu sein. Zu den Erstbesiedlern der Schneisen zählen Betula utilis sowie Weiden, während Rhododendron nicht in die Stufe der Nadelwälder herabsteigt und im Diagramm entsprechend rar ist. Die Gattung Lonicera, mit ihren an Schneebedeckung angepaßten, lichtbedürftigen Sträuchern (SCHICKHOFF 1993) ist hingegen mit einigen PK vertreten. Sehr schlecht erhaltene Betula-

6. Ergebnisse und Diskussionen – Nanga Parbat und Karakorum 107
PK (im Diagramm: Betula korrodiert) wurden vermutlich mit abfließendem Schmelzwasser umgelagert.
Ein Anstieg der Picea-Werte deutet auf eine Sukzession, die von den Birken- und Weidengebüschen hin zu fichtenreichen Waldbeständen führte. Die einzige im Unter-suchungsraum vorkommende Fichtenart Picea smithiana ist feuchteliebend und dürfte von der Öffnung des Kiefernwaldes ebenso profitiert haben wie von der bis in den Sommer hinein vom Lawinenschnee ausgehenden Feuchtigkeit. Nachweise von Abies-PK fehlen, so daß die anspruchsvolle Abies spectabilis im Bereich der Märchenwiese wohl keine Rolle spielte. Im Schutz der Fichten scheinen sich die bei der Birkenausbreitung zurückgegangenen Farnbestände erholt zuhaben. Zu Beginn der Fichtenphase sind auch die Nachweise von Arceuthobium häufiger. Diese auf Koniferen schmarotzende Gattung dürfte an die klimatisch günstigsten Zeiten gebunden gewesen sein (vergl. S. 14). Die Abnahme von Arceuthobium und der Rückgang der Farne zur Mitte des DA 3 deuten vermutlich auf ein verringertes Feuchteangebot hin.
Anschließend spricht der Rückgang von Betula und Salix für ein Nachlassen der Lawi-nenaktivität. Ein Anstieg der Pinus-Werte zeigt, daß die entsprechenden Standorte wieder von Kiefern eingenommen wurden. Die Fichten, langlebiger als Birken und Weiden, wurden etwas später, gegen 2900 BP, von den Kiefern zurückgedrängt, behielten aber fortan einen gewissen Anteil am Waldaufbau.
Am Übergang zum DA 4 endete die Kiefernzunahme. Der starke Anstieg der Cypera-ceen weist auf eine Vergrößerung der Lichtung zu dieser Zeit hin. Mit der weiteren Öffnung gelangten nun vermehrt PK aus dem Weit- und Fernflug ins Profil, so daß aus einem Anstieg der Quercus- und Cedrus-Werte keine sicheren Annahmen zur Entwicklung ihrer Wälder abzuleiten sind. Die zunehmenden Nachweise von Poaceen und Artemisia werden aus der Steppe sowie z. T. auch von der Lichtung stammen. Die Zunahme von Juniperus könnte mit der Ausbreitung auf der an die Lichtung angrenzenden Seitenmoräne zusammenhängen. Etwas erhöhte Werte von Picea und Salix in der zweiten Hälfte des DA 4 weisen möglicherweise wieder auf etwas feuchtere Bedingungen hin.
Ein erneuter Anstieg des NGhP leitet dann zum DA 5 über. Innerhalb dieses vielleicht 500 Jahre umfassenden Abschnittes weisen zeitweise erhöhte Werte des Rumex-Typs, der Gentianaceen, des Aster-Typs und der Apiaceen sowie PK von Plantago lanceolata und ein Rückgang der Cyperaceen auf eine Beweidung der Lichtung hin. Höhere Werte von Salix und Betula können als Folge anthropo-zoogener Störungen gesehen werden, doch ist zudem auch mit dem Einfluß klimatischer Änderungen zu rechnen (HASERODT 1989, RÖTHLISBERGER 1986). Ein deutlich früheres Einsetzen der Weidenutzung, etwa schon nach dem Rückgang der Lawinengefahr des DA 3, kann nicht ausgeschlossen werden, doch sind Hinweise dafür (z. B. durch Pollenfunde von Eremurus) sehr gering.

6. Ergebnisse und Diskussionen – Nanga Parbat und Karakorum 108
Abb. 21: Pollendiagramm des Profils „Märchenwiese Hang“, Nanga Parbat, 3300 m. ü. M. Strati-graphie: schluffig-toniger Sand mit humosen Lagen, oben Cyperaceen-Torf. (GS = GhP).
Märchenwiese, Profil Hang
Das Profil Märchenwiese Hang wurde etwas hangaufwärts außerhalb des eigentlichen vermoorten Bereiches entnommen. Schluffige, teilweise tonige Sande (74 cm) werden hier von einer 10 cm starken organischen Deckschicht überlagert. Pollenanalysen und 14C-Datierung waren im unteren Bereich nur an eingeschalteten humosen Lagen möglich.
Die Überlieferung setzt im Zeitraum des DA 4 des Profils Märchenwiese Rampe ein. Die Anteile des GhP, insbesondere von Pinus, sind entsprechend hoch (Abb. 21). Doch um 1500 BP zeichnet sich in einem starken Abfall der Pinus-Werte ein einschneidender Rückgang der Kiefernbestände ab. An die Stelle des Waldes traten Cyperaceen, Poaceen und Artemisia sowie einer Reihe von Kräutern und Stauden. Auch Schachtelhalm (Equisetum) scheint relativ häufig gewesen zu sein. Über ein Birkengebüsch (Betula 90 %) führte die Sukzession zurück zum Kiefernwald. Dieser Vorgang zeichnet sich im Diagramm Märchenwiese Rampe nicht ab, was für einen sehr lokalen Einfluß auf einen nur kleinen Teil des Kiefernwaldes am Rande der Lichtung spricht. Die Sukzessionsdauer ist wegen der

6. Ergebnisse und Diskussionen – Nanga Parbat und Karakorum 109
Sedimentationsverhältnisse nicht abschätzbar. Der Auslöser des Waldrückganges wird in Kap. 6.4.3. diskutiert.
6.4.2.2 Der Karakorum
Im trockenen Karakorum stellte die Beschaffung von Material für die Pollenanalysen besondere Ansprüche an die Gebietskenntnis der im Gelände tätigen Wissenschaftler. Mit dem Profil Sir le Bala liegt ein relativ junges vegetationsgeschichtliches Archiv aus dem Grenzbereich zwischen dem Nanga Parbat und dem östlichen Teil des Karakorum vor. Einen weiter zurückreichenden Aufschluß zur Vegetationsgeschichte des subariden westlichen Karakorum geben drei Profile aus der Talschaft Yasin. Hier reicht die Überlieferung im Profil Shukan 2 bis in den jüngeren Teil des mittleren Holozäns, in den Profilen Batyatum Jut und Alambar bis an den Beginn des jüngeren Holozäns zurück. Auf weitere Profile wird kurz eingegangen. Die angegebenen Prozentwerte beziehen sich auf die Grundsumme aller Landblütenpflanzen (GhP + NGhP) ohne Cyperaceen.
Profil Sirle Bala
Im oberen Astor-Tal lagert in einer von Quell- und Schmelzwasser gespeisten Hang-verflachung in 3010 m Höhe über einer hellen Mudde schwarzes anmooriges Material und darüber helles, gleyartiges Material sowie 30 cm Cyperaceen-Torf. Der GhP und NGhP sind etwa gleich stark vertreten (Abb. 22). Besonders häufig sind dabei Pinus, Betula, Poaceen und Artemisia. Zu Beginn deuten hohe Werte der Filicatae monolet noch auf Farnbestände hin, die durch eine Ausdehnung der Cyperaceen-Bestände im DA 1 zurückgedrängt wurden. Auf die Cyperaceen-Zunahme folgen um 1900 BP zu Beginn des DA 2 mit PK von Juniperus und Eremurus erste Hinweise auf eine Beweidung. Später treten mit dem Poly-
gonum aviculare- und Plantago major/media-Typ sowie mit Plantago lanceolata weitere Weidezeiger auf. Der Rückgang von Poaceen und die Zunahme von Apiaceen und Rumex(-Typ) können als Reaktion auf den zunehmenden Beweidungsdruck gewertet werden. Damit zeigt die Abnahme der Betula-Werte in den obersten Proben wohl einen anthropo-zoogen bedingten Birkenrückgang an.
Im anmoorigen Sediment in der Mitte des Profils (etwa 1600 BP) gibt es einen sehr hohen Wert von Betula und eine Lage von Cyperaceen-Blattscheiden. Es ist vielleicht durch die schnelle Sedimentation gößerer Mengen eingespülten Materials entstanden. Die Ursache dieses Ereignisses scheint auf die weitere Vegetationsgeschichte keinen Einfluß gehabt zu haben. Möglicherweise handelte es sich um die Folge eines kurzzeitigen Wettergeschehens (Diskussion in Kap. 6.4.3.).
Profil Shukan 2
Das Profil Shukan 2 konnte in einem bachnahen Niedermoor in 3360 m Höhe in einem Seitental des Nazbar-Tales genommen werden. Die sandig-kiesige Basis datiert auf 5000 BP. In nächster Nähe liegen die Bewässerungsfelder (Hordeum), Pferche und Hütten einer von der Ortschaft Shukan aus betriebenen Sommerfeldsiedlung. Nach chinesischen Quellen

6. Ergebnisse und Diskussionen – Nanga Parbat und Karakorum 110
besteht die Ortschaft Shukan seit mindestens 700 AD. Auf den Hängen in der Umgebung der Sommerfeldsiedlung weiden Schaf- und Ziegenherden. Entlang der natürlichen Wasserläufe und der Bewässerungskanäle grasen Rinder. Laubgehölze mit Birken und Weiden sind auf gewässernahe Gunststandorte beschränkt. Die zonale Vegetation des Untersuchungsgebietes ist eine Artemisia-Steppe mit Beständen der Wacholder-Steppenwälder. Die Vorkommen der Wacholderbäume beschränken sich dabei überwiegend auf offene Gruppen an Felshängen ohne Kronenschluß (frdl. persönliche Mitteilung Prof. Dr. G. MIEHE).
Abb. 22: Pollendiagramm des Profils „Sirle Bala“ aus dem Karakorum, oberes Astor-Tal, 3010 m. ü. M. Stratigraphie: unterer Teil helle Mudde, darüber anmooriger Bereich mit Pflanzenstreu, darüber gleyartiges Material, obere 30 cm Cyperaceen-Torf. (GS = GhP + NGhP).
Zu Anfang des DA 1 weisen hohe Pinus-Werte (anfangs 45 %) auf Kiefernwälder hin (Abb. 23). Die Niederschläge müssen damals höher gewesen sein als in der Gegenwart. Makroreste der Kiefern wurden nicht gefunden. Zusagende Standortverhältnisse für Pinus wallichiana dürfte zumindest das nahe Nazbar-Tal mit seinen Nordhängen geboten haben. Juniperus-Werte von unter 5 % sprechen nicht für größere Vorkommen von Steppenwäldern. Bei relativ hohen Werten von Betula und Salix werden ausgedehnte gewässerbegleitende Gehölze bestanden haben, zu denen vermutlich auch Vertreter der

6. Ergebnisse und Diskussionen – Nanga Parbat und Karakorum 111
Rosaceen (Rosaceen p. p.) gehörten. Hohe Einzelwerte von Ephedra und Cichorioideen dürften auf kleinflächige lokale Vorkommen zurückgehen. PK von Picea, Cedrus und Quercus zählen zur Fernflug-Komponente der Pollenspektren. Der folgende Abfall der Pinus-Kurve weist auf einen Rückgang der Niederschläge um etwa 5000 BP hin. Der gleichzeitige Rückgang der Cyperaceen-Werte könnte auf einen trockenheitsbedingten Abbau alpiner Kobresia-Matten hindeuten.
Abb. 23: Pollendiagramm des Profils „Shukan 2“ aus dem Karakorum, Talschaft Yasin, Seitental des Naz Bar-Tales, 3360 m. ü. M. Stratigraphie: an der Basis sandig-humoser Kies, darüber Cyperaceen-Torf. (GS = GhP + NGhP).
Am Übergang zum DA 2 steigen die Artemisia-Werte deutlich an und belegen die mit Zunahme der Aridität anwachsende Bedeutung der Artemisia-Steppe. Die Nachweise von Ephedra und Acantholimon lassen bereits eine subaride Steppe ähnlich jener der Gegenwart annehmen. Bei Pinus-Werten von fortan nur mehr 5 bis 10 % wird sich auch bereits die Kiefernwaldverteilung der heutigen angenähert haben. Kaum veränderte Betula- und Salix-Werte sind als Beweis für den ungestörten Fortbestand gewässernaher Gehölze aufzufassen. Eine Ausdehnung der Cyperaceen-Bestände im nassen Umfeld des Untersuchungspunktes zeigt sich in der Zunahme der Cyperaceen-Werte und der Bildung des Cyperaceen-Torfes.

6. Ergebnisse und Diskussionen – Nanga Parbat und Karakorum 112
Mit Dornpolstern von Acantholimon tritt zu Beginn des DA 2 eine Wuchsform auf, die nicht nur an relative Trockenheit, sondern auch an Abweidung angepaßt ist. Auf Weide-gänger weisen auch erhöhte Werte vom Polygonum aviculare-Typ, vom Plantago
major/media-Typ und von Rheum hin. Arten der genannten Sippen stellen allerdings unter den naturräumlichen Gegebenheiten des Karakorum mit seinem vegetationsfeindlichen Klima und den (einst) vorkommenden Wildherden bis zu einem gewissen Grade wohl auch natürliche Bestandteile der Vegetation dar. In der Mitte des DA 2 weisen erhöhte Werte des Potentilla-Typs, der Apiaceen und der Fabaceen auf eine Störung der lokalen Vegetation hin. Hier tritt Eremurus als ein weiteres Weideunkraut auf. Wahrscheinlich geht dieser Befund tatsächlich auf anthropo-zoogene Einflüsse zurück (siehe Kap. 6.4.3.).
Im DA 3 deutet dann die Zunahme der Anteile vom Plantago major/media-Typ und die PK von Urtica auf einen starken Viehbesatz hin. Der Rückgang der Poaceen- und Cypera-ceen-Werte mag z. T. auf eine Abweidung der Gräser (durch Viehherden) zurückzuführen zu sein, wodurch Gattungen mit weideresistenten Vertretern wie z. B. Juniperus und Artemisia häufiger wurden. In den rückläufigen Anteilen von Betula und Salix zeigt sich nun eine Abnahme der gewässerbegleitenden Gehölze. Mögliche Ursachen könnten die Viehherden, der Feuerholzbedarf der Hirten sowie die Verwendung von Birkenrinde zur Verpackung von Milchprodukten gewesen sein. Andererseits deuten die PK des Hordeum-Typs auch auf Bewässerungsfelder hin, die vermutlich wie heute in Gewässernähe angelegt wurden. Die heutige Sommerfeldsiedlung dürfte also wohl am Übergang vom DA 2 zum DA 3 (ca. 1400 BP) gegründet worden sein. Ein hoher Juniperus-Wert sowie der Nachweis von Thesium sprechen dann für einen ansteigenden Beweidungsdruck in jüngster Zeit.
Profil Batyatum Jut
Getrennt von einem über 5000 m hohen Gebirgszug fließt nördlich des Nazbar-Tales der Khaimetbar-Fluß. In seinem oberen Talabschnitt hat sich in einer staunassen Hohlform auf einer Gletscherschliffläche in 3550 m Höhe ein Niedermoor gebildet, aus dem das 93 cm lange Profil stammt. Von einer Alm aus wird das Gebiet als Schafweide genutzt. Baumstümpfe zeugen von ehemals stärkeren Beständen von Juniperus-Bäumen. Jungwuchs von Baumwacholdern ist in unzugänglichen Lagen zu beobachten (frdl. persönliche Mitteilung Dr. J.-P. JACOBSEN).
Die Überlieferung der Vegetationsgeschichte setzt im DA 1 um 2700 BP zu Beginn des jüngeren Holozäns ein (Abb. 24). Der NGhP erreicht Werte um 80 % und stammt zum großen Teil von einer Steppenvegetation aus Poaceen und Artemisia. Juniperus-Pollen bildet den Hauptteil des GhP. Die Werte von Juniperus schwanken zwar, liegen aber insgesamt auf einem relativ hohen Niveau und sprechen damit für die Existenz von Steppenwäldern. Die zu Anfang hohen Cyperaceen-Werte weisen auf eine gute lokale Wasserversorgung hin. Die PK von Salix, Betula, Hippophaë und Rosaceen werden wohl aus dem feuchten Umfeld des Niedermoores sowie vom Nahflug aus subalpinen und gewässernahen Gebüschen stammen. Mit etwa 5 % ist der Fernflug von Pinus, Cedrus und Quercus recht gering. Die ansteigenden Werte des Plantago major/media- und des Polygonum aviculare-Typs deuten auf einen erhöhten Weidedruck ab etwa 2250 BP hin. Wohl als Folge davon bahnen sich

6. Ergebnisse und Diskussionen – Nanga Parbat und Karakorum 113
gegen Ende des DA 1 deutliche Änderungen der Steppenvegetation an. So nahm die Häufigkeit von Artemisia im Vegetationsbild offenbar zu, während die der Poaceen abnahm. Zudem treten mit dem Centaurea/Cousinia-Typ Hinweise auf das Vorkommen bewehrter Weideunkräuter auf.
Abb. 24: Pollendiagramm des Profils „Batyatum Jut“ aus dem Karakorum, Talschaft Yasin, Khaimetbar-Tal, 3550 m. ü. M. Stratigraphie: Cyperaceen-Torf. (GS = GhP + NGhP).
Der DA 2 zeichnet sich zu Anfang durch besonders hohe Werte des Plantago major/-media- und des Anemone/Ranunculus-Typs sowie durch eine geringe Pollenkonzentration aus, so daß der Weidedruck etwa gegen 1700 BP noch zugenommen haben muß. Der Rückgang der Juniperus-Werte geht wohl auf ein Abholzen von Wacholderbäumen zurück. Die um 800 BP erhöhten Cyperaceen-Werte sprechen für eine verbesserte Wasserversorgung des Niedermoores. Anschließend gehen die Cyperaceen-Werte zurück und die Konzentration des Pollens der Grundsumme steigt an, so daß eine Verlangsamung des Torfzuwachstums anzunehmen ist. Als Ursachen kommen Beweidung wie auch klimatische Änderungen in Frage (siehe Kap. 6.4.3.).

6. Ergebnisse und Diskussionen – Nanga Parbat und Karakorum 114
Profil Alambar
Das Alambar-Tal liegt im Nordosten der Talschaft Yasin. Unter dem subariden Klima erfolgt der Übergang von der Artemisia-Steppe zu einer offenen, krautreichen alpinen Steppe in 3600 bis 3800 m Höhe (S. MIEHE et al. 1996). Die Vegetationsgeschichte der letzten 2500 Jahre ist in einem 80 cm mächtigen Niedermoor archiviert. Dieses bildete sich in 3700 m Höhe an einem quelligen Schutthang unterhalb einer Gletscherzunge. Hier weiden heute durchziehende Herden; eine 3 km talabwärts gelegene Almhütte wird nicht mehr betrieben (frdl. persönliche Mitteilung Dr. H.-J. JACOBSEN).
Abb. 25: Pollendiagramm des Profils „Alambar“ aus dem Karakorum, Talschaft Yasin, Alambar-Tal, 3700 m. ü. M. Stratigraphie: Cyperaceen-Torf. (GS = GhP + NGhP).
Der Anteil des GhP beträgt im DA 1 etwa 20 % (Abb. 25). Etwa die Hälfte davon stammt aus dem Fernflug, was für eine relativ spärliche Vegetation mit geringer Pollen-produktion am Untersuchungspunkt spricht und durch die Nähe der alpinen Steppe bedingt gewesen sein dürfte. Die wenigen Bäume und Büsche des Tales waren Arten der Gattungen Juniperus, Ephedra, Salix, Hippophaë, Betula und vermutlich Fraxinus. Hohe Werte von Artemisia, Poaceen und Chenopodiaceen (Chenopodiaceen p. p., Haloxylon-Typ) deuten auf das Vorherrschen einer relativ trockene Steppe hin. Der Plantago major/media- und der Polygonum aviculare-Typ lassen von Beginn des DA 1 an einen gewissen Weidedruck vermuten, der am Übergang zum DA 2 vor ca. 2000 Jahren noch zunahm. Dafür sprechen neben den erhöhten Werten des Plantago major/media- und des Polygonum aviculare-Typs die Nachweise von Sippen, die durch Dornen, Stacheln oder bestimmte Inhaltsstoffe gegen

6. Ergebnisse und Diskussionen – Nanga Parbat und Karakorum 115
Fraß geschützt sind: Acantholimon, Centaurea/Cousinia-Typ, Rheum und Eremurus. Dem entsprechend steht hier die Zunahme der Juniperus-Werte für eine Ausbreitung strauchförmiger Wacholder wie z. B. J. communis. Ein auf 1115 ± 190 BP datiertes Stück Juniperus-Holz aus dem Profil belegt Wacholder-Vorkommen vor Ort (vergl. Anhang). Erhöhte Cichorioideen-Werte treten gleichzeitig mit höheren Anteilen des Plantago
major/media- sowie des Polygonum aviculare-Typs auf und lassen an die Ausbreitung von Weideunkräutern wie z. B. Taraxacum und Leontopodium denken.
Im Verlauf des DA 2 gehen die Hinweise auf eine Beweidung der Vegetation zunächst zurück. Es treten aber höhere Cyperaceen-Werte auf und diese mögen Ausdruck feuchterer Verhältnisse sein, die eine Ausdehnung der Weidemöglichkeiten erlaubten und so den lokalen Weidedruck verringert haben mögen. In jüngerer Zeit sprechen dann erhöhte Juniperus- und Artemisia-Werte wieder für die selektive Wirkung einer verstärkten Bewei-dung auf die Zusammensetzung der Vegetation.
Über den gesamten Zeitraum von etwa 2500 Jahren betrachtet, zeigen die Poaceen eine leichte Abnahme, während die Werte von Artemisia ansteigen. Der sich darin abzeichnende allmähliche Wandel der Steppenvegetation hin zu einem größeren Reichtum an Artemisia ist wohl durch den langen Weidedruck bei allgemein trockenem Klima zu erklären.
Weitere Profile
Im wenige Kilometer vom Profil Shukan 2 gelegenen Profil Shukan 1 (Tab. 3, diese und folgende im Anhang) liegt über einer klastischen Schüttung ein Cyperaceen-Torf noch unbekannten Alters. Die palynologischen Daten spiegeln das Ersetzen einer an Poaceen und Lamiaceen reichen Pioniervegetation durch Cyperaceen ebenso wie die oben bereits dargestellte allgemeine Gehölzarmut des Gebietes wider. Neben Poaceen und Lamiaceen gehörten u. a. auch Caryophyllaceen, Campanulaceen (Campanula-Typ) sowie Acantholi-
mon zu den Erstbesiedlern des sandig-kiesigen Sediments der Profilbasis. Die Untersuch-ungen am Profil Karakorum 2 (Tab. 4) nahe von Shukan bestätigen die Waldarmut des Nazbar-Tales seit dem jüngeren Mittelholozän. Das Profil Karakorum 1 (Tab. 5) aus dem Gilgit-Tal weist entsprechend seiner Lage im Verbreitungsgebiet von Pinus wallichiana (Abb. 19) einen relativ hohen Pinus-Anteil auf. Ein Rückgang der Juniperus-Werte deutet auf eine Abnahme der Wacholder in den letzten 1000 Jahren hin. Im Bagrot-Tal stehen oberhalb einer trockenen Talstufe ab etwa 3000 m Höhe ausgedehnte Kiefernwälder. Im Diagramm Bagrot (Tab. 6) spricht der deutliche Abfall der Pinus-Werte und die Zunahme der Cyperaceen-Anteile für eine vermutlich sehr junge Abnahme der Kiefernwälder. Zur Klärung von Alter, Ausmaß und Ursachen sind weitere Untersuchungen notwendig. Das Profil Deosai Plateau (Tab. 7) stammt von der gleichnamigen Hochebene im Osten des Karakorum. Die hier auf Löß stehenden Wiesensteppen unterliegen der Korrasion durch die über den Himalaja-Hauptkamm einfallenden Winde. Die Ausblasung der feinen

6. Ergebnisse und Diskussionen – Nanga Parbat und Karakorum 116
Bodenteilchen hinterläßt Steinpflaster (frdl. persönliche Mitteilung Prof. Dr. G. MIEHE). Es bleibt unsicher, inwieweit die Zunahme der Pinus-Werte (Fernflug) auf einem Rückgang der Wiesensteppe oder der Zunahme von Kiefern auf der Südseite des Himalaja beruhen (SHARMA & CHAUHAN 1988, SHARMA 1991). Die Klärung dieses Problems steht noch aus.
6.4.3 Diskussion
Älteres Holozän (bis ca. 7200 BP)
Die Vegetationsgeschichte des Nanga Parbat ist im Profil Märchenwiese Rampe ab etwa 7900 BP überliefert. Seitdem stehen hier bereits feuchte Kiefernwälder von Pinus wal-
lichiana ähnlich denen der Gegenwart.
Mittleres Holozän (ca. 7200 - 2700 BP)
Die Bildung der versumpften Lichtung am Nanga Parbat (Märchenwiese Rampe) deutet auf zunehmende Nässe um 7200 BP hin. Diese dürfte auf eine Klimaänderung zurückzuführen sein, begann doch im westlichen Tibet um 7200 BP durch eine Zunahme des Sommermonsuns der feuchteste Abschnitt des Holozäns (GASSE et al. 1996, VAN CAMPO et al. 1996). Aufgrund der örtlichen geomorphologischen Situation führten die erhöhten Niederschläge zu einer so starken Vernässung, daß sich der Kiefernwald infolge einer Versumpfung lokal lichtete. Der im Umfeld stehende Kiefernwald änderte sich kaum; zur Ausbreitung feuchteliebender Baumarten wie Picea smithiana und Abies spectabilis kam es im Bereich der Märchenwiese nicht.
Erst gegen 6300 BP, d. h. bereits nach dem Maximum des monsunalen Einflusses (vergl. Nianbaoyeze, Kap. 6.3.1), änderte sich das Waldbild. Lawinenbahnen zerschnitten den bis dahin dichten Kiefernwald und Birken sowie Weiden konnten sich ausbreiteten. Ein Teil dieser Gebüsche wurde anschließend durch Fichtenbestände ersetzt. Picea smithiana vermochte demnach erst nach massiven Störungen der Kiefernwaldstruktur und zudem gefördert durch die Feuchtigkeit des Lawinenschnees lokal mit Pinus wallichiana erfolgreich zu konkurrieren. Angesichts der nur geringen vertikalen Reichweite der monsunalen Niederschläge geht die erhöhte Lawinentätigkeit vermutlich auf einen stärkeren Einfluß zyklonaler Störungen mit höheren Niederschlägen im Winter und Frühjahr zurück. Diese Annahme wird durch palynologische Befunde aus den Durchzugsgebieten der mediterranen Störungen unterstützt. So führten im Zagros-Gebirge im Iran ab 6000 BP diese verstärkten Winterniederschläge innerhalb weniger Jahrhunderte über eine Verdichtung der Waldsteppe zu einer durchgängigen Waldbedeckung (ZEIST & BOTTEMA 1988) und beendeten die bereits aus archäologischen Befunden erkannte postglaziale Trockenperiode (MEDER 1979). Die hohe Fichtenhäufigkeit an der Märchenwiese korreliert mit der relaltiv jungen mittelholozänen Niederschlagszunahme der Winterregengebiete und nicht mit einer Stärkung des Sommermonsuns. Möglicherweise stehen auch die nach 6000 BP erhöhten Seespiegelstände in der Rajasthan-Wüste Indiens mit einer zyklonalen Niederschlags-zunahme in Verbindung (AGRAWAL & GUPTA 1988, BHATIA & SINGH 1988). Schließlich bezeugt der Rückgang von Birken, Weiden und Fichten das Nachlassen der

6. Ergebnisse und Diskussionen – Nanga Parbat und Karakorum 117
Winterniederschläge und damit die Abnahme der zyklonalen Störungen. Das Ende hoher Picea-Werte um 2900 BP dürfte aufgrund der Lebensdauer der Fichten gegenüber dem Niederschlagsrückgang verzögert sein. Ein markanter Gletschervorstoß im nahen Bagrot-Tal endete bereits um 3200 BP (RÖTHLISBERGER 1986), und die Trockenheit der Wüsten Indiens wird durch ein Absinken der winterlichen und sommerlichen Niederschläge seit etwa 4000 bis 3000 BP erklärt (VISHNU-MITTRE 1979, AGRAWAL 1992, BRYSON & SWAIN 1981). Der Rückgang der Kiefernbestände in der trockenen Talschaft Yasin (Shukan 2) belegt ein Nachlassen der seit 7200 BP erhöhten monsunalen Sommerniederschläge (Märchenwiese Rampe) bereits um 5000 BP. Am viel feuchteren Nanga Parbat deutet hingegen nur die Abnahme der Farne und der Gattung Arceuthobium auf eine verringerte sommerliche Feuchtigkeit an.
Die palynologischen Untersuchungen am Nanga Parbat und im Karakorum betreffen einen Raum, dessen Niederschlagsverhältnisse durch seine Übergangslage zwischen dem vorderasiatisch-subtropischen (Winter-Frühjahrs-Niederschläge) und dem südasiatisch-randmonsunalen Klimasystem (Sommerregen) bestimmt werden (Abb. 18; REIMERS 1992). Das Paläoklima wurde zum einen durch die Zunahme des sommermonsunalen Niederschlagseinflusses um 7200 BP und dessen Nachlassen um 5000 und 2700 BP bestimmt, sowie durch erhöhte Winterniederschläge infolge einer verstärkten Einwirkung des vorderasiatisch-subtropischen Klimasystems etwa von 6300 bis 3000 BP.
Jüngers Holozän (ab ca. 2700 BP)
Zu Beginn des jüngeren Holozäns dürften sich die Wuchsbedingungen für Cyperaceen-Niedermoore lokal verbessert haben. Die Lichtung an der Märchenwiese weitete sich aus, und in der Talschaft Yasin bildete sich der Niedermoortorf bei Batyatum Jut, und wenig später auch der von Alambar. Die bisherigen Untersuchungen ergaben keine Anhaltspunkte für eine über die stellenweise vorkommenden Juniperus-Steppenwälder (Batyatum Jut) hinausgehende Bewaldung der Talschaft Yasin im jüngeren Holozän. Das entspricht auch Ergebnissen von BORTENSCHLAGER & PATZELT (1978) aus dem unmittelbar nördlich gelegenen Wakhan.
Das Diagramm von Batyatum Jut enthält von Beginn der vegetationsgeschichtlichen Überlieferung (ca. 2700 BP) an Hinweise auf eine Trittbelastung der Vegetation, die allerdings ebenso von Wildherden wie von einer frühen anthropo-zoogenen Nutzung ausgegangen sein kann. Eine Zunahme von Weideunkräutern um etwa 2250 BP könnte dagegen den Herden der im Yasin-Haupttal ansässigen Bevölkerung zugeschrieben werden (JACOBSEN & SCHICKHOFF 1995). Auch die kurzzeitige Vegetationsstörung im Nazbar-Tal (Shukan 2: Mitte DA 2) fällt vermutlich in diese Zeit und dürfte ebenfalls anthropo-zoogenen Ursprungs sein. Es bleibt noch zu klären, welcher Einfluß dabei von den Skythen/Saken ausging, die seit dem 7. oder 5. Jh. v. Chr. in den Karakorum vordrangen und dabei die Infrastruktur für die aus dem Ostpamir nachrückenden Truppen schufen, die ab dem 2. Jh. v. Chr. nach Indien einfielen, indem sie u. a. durch das östlich des Yasin gelegene Ishkoman-Tal zogen (JETTMAR 1993, JETTMAR & THEWALT 1985). Zu fragen wäre auch,

6. Ergebnisse und Diskussionen – Nanga Parbat und Karakorum 118
welche Auswirkungen die Eroberungszüge Alexander des Großen durch die dem Karakorum benachbarten Gebiete von Sogdien und NW-Indien für den Karakorum hatten.
Auch bei Alambar treten von Beginn an (2500 BP) Beweidungszeiger auf. Ab etwa 1950 BP nahm die Weideeinflußstärke zu. Die sich darin andeutende Nutzung des Tales mag i. w. S. mit der sog. Seidenstraße zusammenhängen. Von dem an den Handelstätigkeiten beteiligten Sogdien führte ein Weg zum Indus-Tal über den nur etwa 10 km von Alambar entfernten Dakrot-Pass durch die Talschaft Yasin (MODE 1991, s. a. Kap. 5.5.). Bei Batyatum Jut verstärkte sich die Weidenutzung gegen 1700 BP. Vielleicht ist auch der Beginn des Bewässerungsfeldbaus im nahen Nazbar-Tal bei Shukan (Shukan 2, Übergang DA 2/DA 3) an den Beginn des 2. Jahrtausend zu stellen.
Am Untersuchungspunkt im oberen Astor-Tal bei Sirle Bala schuf die lokale Zunahme der Cyperaceen-Bestände eine Weidemöglichkeit, die seit etwa 1900 BP genutzt wird. Die um 1600 BP gebildete anmoorige Lage im Profil Sirle Bala und die im Profil Märchen-wiese Hang dokumentierte lokale Öffnung des Kiefernwaldes am Nanga Parbat um 1500 BP gehen auf kurzzeitige Störungen zurück. Es könnte sich dabei um extreme Nieder-schlagsereignisse gehandelt haben, wie sie in einzelnen Jahren immer wieder zu beobachten sind (REIMERS 1994). Ein Beispiel für solche Ereignisse im Untersuchungszeitraum ist möglicherweise eine Überschwemmungskatastrophe in Südmesopotamien im Jahr 627 n. Chr. (OSTEN 1956).
Höhere Werte der Cyperaceen (Batyatum Jut, Alambar) sowie von Picea und Salix (Märchenwiese Rampe) lassen für eine Phase von wenigen Jahrhunderten nach Beginn unseres Jahrtausends etwas bessere Feuchtigkeitsverhältnisse annehmen.
Im Diagramm Märchenwiese Rampe zeichnet sich ein anthropo-zoogene Einfluß erst seit etwa 500 Jahren ab. Auch im Yasin scheint die Nutzung in den letzten Jahrhunderten noch zugenommen zu haben. Dies deckt sich im wesentlichen mit Ergebnissen aus benach-barten Räumen. So weist RAUNIG (1983) auf einen erhöhten Bevölkerungsdruck im Wakhan ab der 2. Hälfte des 14. Jh. hin, und pollenanalytische Arbeiten in Indien (SHARMA 1991) und Nepal (BEUG & MIEHE 1997, 1999) beschreiben vergleichbar junge Vorgänge in den Tälern an der Südseite des Himalaja.

7. Zusammenfassung – Pleistozän 119
7 Zusammenfassung
7.1 Pleistozän
7.1.1 Gaxun Nur
An einem 1 Mio. Jahre zurückreichendem Kern aus einem Endsee in den Wüstengebieten der Inneren Mongolei, dem Gaxun Nur, wurden erste palynologische Untersuchungen durchgeführt. Es ließen sich im älteren Teil der Ablagerungen von Picea, Abies und Pinus dominierte Nadelwälder mit weiteren holarktischen Vertretern (Larix, Juniperus) und subtropischen Elementen (Tsuga) nachweisen. Unter den Laubbäumen war Ulmus häufiger als in der Gegenwart. Zudem scheinen Pterocarya und Hippophaë einst im Untersuchungsgebiet vorgekommen zu sein. Eine Versalzung der Böden wird durch Nitraria angezeigt.
7.1.2 Lingfeng
Im Pollendiagramm Lingfeng (Qinling Shan, östliches China) zeichnen sich zu Beginn des letzten Glazials zwei Waldzeiten ab, die den mitteleuropäischen Interstadialen Brörup und Odderade sowie Bodenbildungsphasen der chinesischen Lößstratigraphie entsprechen dürften. Das Landschaftsbild wurde von Picea-(Abies)-Wäldern beherrscht. Die klimatischen Verhältnisse waren ähnlich denen der Gegenwart (JMT 4,5 °C, >500 mm Niederschlag). Im Klimaoptimum der zweiten Waldzeit wurden Pioniergehölze (Hippophaë, Salix) und Laubbäume (Ulmus, Carpinus) durch fluviale Störungen der Nadelwälder gefördert.
In dem Stadial zwischen beiden Waldzeiten herrschten subalpine Gebüsche mit Juniperus und Rhododendron. Auf eine vergleichbare subalpine Phase nach der zweiten Waldzeit folgten mit fortschreitender Klimaverschlechterung alpine Bedingungen mit einer von Cyperaceen beherrschten krautreichen Vegetationsdecke. Danach trat eine geringe Wiederbewaldung ein. Im anschließenden Abschnitt, in den das LGM fällt, herrschten erneut alpine Verhältnisse. Für die subalpinen Phasen ist eine um mindestens 2,5 °C geringere Jahresmitteltemperatur als in der Gegenwart anzunehmen. Zur Zeit der alpinen Verhältnisse betrug die Absenkung vermutlich 5,5 bis 7,5 °C.
7.2 Holozän
7.2.1 Nianbaoyeze
Der Nianbaoyeze, ein Granitmassiv im Osten Tibets, besaß im gesamten Holozän nur kleinflächig begrenzte Baumvorkommen. Kobresia-Matten beherrschten das Vegetationsbild und behinderten die Ansiedlung von Bäumen und Sträuchern. Eine gewisse Rolle für die Waldarmut mögen auch die seit dem älteren Holozän anhand von Holzkohlen

7. Zusammenfassung – Holozän 120
nachgewiesenen Brände gespielt haben. Gehölze konnten besonders an felsigen Hängen und bei mechanische Störungen der Mattendecke aufkommen. Vor allem zu Beginn des Holozäns und durch erhöhte sommermonsunale Niederschläge zur Zeit des holozänen Klimaoptimums um 6600 BP konnten sich Pioniergehölze wie Hippophaë und Salix in Flußnähe ausbreiten. Eine leichte Zunahme der Nadelbaumbestände zu Beginn des mittleren Holozäns (7500 BP) weist auf eine Klimabesserung hin. Während des Klimaoptimums dürfte die Jahresmitteltemperatur um weniger als 2 °C über der heutigen gelegen haben. Es folgten zwei deutliche Klimarückgänge, ein erster im mittleren Holozän um 5300 BP und ein zweiter am Übergang zum jüngeren Holozän um 2700 BP.
Da schon zu Beginn des Holozäns Picea, Abies und Juniperus auftraten, handelt es sich bei den rezent in Südexposition auftretenden Baumbeständen von Juniperus weniger um Ersatzgesellschaften nach einem Abholzen ehemaliger Picea-Abies-Wälder, als vielmehr um einen natürlichen Vegetationstyp. Dies ist bis zu einem gewissen Grade auch für andere Gebiete des östlichen Tibets anzunehmen. Eine anthropo-zoogene Einflußnahme zeichnet sich in den letzten 800 Jahren durch Änderungen in der Zusammensetzung der Mattenvegetation ab. Das heutige Auftreten der strauchförmigen Potentilla fruticosa als Weideunkraut kann auf die Öffnung der Mattennarbe durch Viehtritt zurückgeführt werden. Feldbeobachtungen zeigen, daß die von Natur aus geringen Waldbestände seit einigen Jahrzehnten durch verstärkte Nutzung aufgebraucht werden.
7.2.2 Lhasa
Im östlichen Stadtbereich von Lhasa (3650 m) ist der heutige Getreideanbau von Weizen und Gerste pollenanalytisch über mehr als 600 Jahre zurückzuverfolgen. Zwischen 1300 und 1650 n. Chr. kam es in einer Feuchtphase zur Entstehung eines pflanzenreichen Teiches. Wie die Pollenanalysen nahelegen, werden Anpflanzungen von Salix und Populus seit dem 20. Jahrhundert vorgenommen.
7.2.3 Nanga Parbat
Im Bereich der sog. Märchenwiese (3300 m) am Nanga Parbat (NW-Himalaja) bildete sich in dichtem Kiefernwald (Pinus wallichiana) während des älteren Holozäns ein ver-sumpfter Bereich, der sich mit Zunahme der sommermonsunalen Niederschläge ab 7200 BP ausdehnte. Gegen 6300 BP führte der verstärkte Einfluß zyklonaler Störungen aus dem mediterranen Raum zu einer Erhöhung der winterlichen bis frühjahrszeitlichen Niederschläge. Vermutlich öffneten vermehrte Lawinenabgänge den Kiefernwald soweit, daß sich Birken, Weiden und Fichten ausbreiten konnten.
Während der Rückgang der sommermonsunalen Niederschläge gegen 5000 BP kaum Auswirkungen auf die Vegetation hatte, zeigt sich ein Ausbleiben der erhöhten winterlichen Niederschläge gegen 3000 BP im Rückgang von Birken, Weiden und Fichten. Der Beginn des jüngeren Holozäns um 2700 BP ist durch einen vermehrten Eintrag von Pollen mehr oder minder trockenresistenter Sippen aus der näheren (Juniperus, Artemisia, Poaceen) und weiteren Umgebung (Quercus, Cedrus) gekennzeichnet. Erst etwa in den letzten 500 Jahren liegen Hinweise auf einen anthropo-zoogenen Einfluß vor.

7. Zusammenfassung – Holozän 121
7.2.4 Karakorum
Die Verbreitung der Kiefernwälder war im mittleren Holozän etwas größer als heute. Spätestens seit der Abschwächung des Sommermonsuns gegen 5000 BP herrscht in der Talschaft Yasin im westlichen Karakorum (N-Pakistan) jedoch eine trockene Artemisia-Steppe, gebietsweise mit Juniperus-Steppenwäldern, vor. Im jüngeren Holozän (ab 2700 BP) kam es lokal zur Bildung vermutlich schmelzwassergespeister Niedermoore, die von Beginn an einer geringen Beweidung unterlagen. Zur Zeit der Zunahme der Weideunkräuter um 2300 BP war das Yasin-Tal bereits besiedelt. Die weitere Zunahme des Weidedrucks gegen 2000 BP könnte in Bezug mit der Seidenstraße stehen, von der eine Route durch das Yasin-Tal zum Indus führte. In den letzten Jahrhunderten nahm der anthropo-zoogene Einfluß weiter zu.
7.2.5 Klimageschichtliche Ergebnisse: Ein überregionaler Vergleich
In der Vegetationsgeschichte des Nianbaoyeze und des Nanga Parbat zeichnet sich eine Stärkung des Sommermonsuns bei Beginn des mittleren Holozäns um 7500 bzw. 7200 BP ab. Zu einer Abschwächung des Sommermonsuns kam es um 5300 BP (Nianbaoyeze) bzw. 5000 BP (Nanga Parbat, Karakorum). Eine weitere Klimaverschlechterung erfolgte am Übergang zum jüngeren Holozän um 2700 BP. Abgesehen von der Zunahme der winterlichen/frühjahrszeitlichen Niederschläge durch eine Stärkung zyklonaler Störungen zwischen etwa 6300 bis 3000 BP (Nanga Parbat) entspricht die Gliederung der Klimageschichte im Holozän also weitgehend der von Mitteleuropa (FIRBAS 1949, LANG
1994). Das ältere Holozän umfaßt in der vorliegenden Arbeit etwa das Präboreal und Boreal. Das mittlere Holozän entspricht der Zeit des Atlantikums und des Subboreals, mit dem mittelholozänen Klimarückgang um 5300/5000 BP. Das Einsetzen des jüngeren Holozäns ist zeitgleich mit der weltweit nachzuweisenden Klimaänderung am Übergang vom Subboreal zum Subatlantikum (VAN GEEL et al. 1996).
Daneben zeichnen sich in Gruslagen (Nianbaoyeze) und mit kurzzeitigen Vegetations-änderungen (Nanga Parbat, Karakorum) wiederkehrende Ereignisse ab, wie sie etwa durch Starkniederschläge bei Schlechtwetterlagen auftreten können. Ein Einfluß eines Hebungsvorganges des Tibetanischen Hochlandes auf die holozäne Klimaentwicklung, wie von einigen Autoren angenommen, hat sich nicht gezeigt.

8. Summary – Pleistocene 122
8 Summary
Palynological investigations on the holocene history of vegetation, climate and human influence in High Asia (Nanga Parbat, Karkoram, Nianbaoyeze, Lhasa) and on
the Pleistocene of China (Qinling Shan, Gaxun Nur)
8.1 Pleistocene
8.1.1 Gaxun Nur (Inner Mongolia)
Pollen analysis was done on sediments, 1 million years old, from Gaxun Nur, an endoreic lake in the desert near the borderline to Outer Mongolia. The analyse shows that coniferous forest dominated by Picea, Abies, Pinus with subtropical species (Tsuga) and other holarctic elements (Larix, Juniperus) existed in that area until about 500.000 years ago. Ulmus was more frequent than today. Pterocarya and Hippophaë seem to have occurred there, too. The existence of salty soils is indicated by Nitraria.
8.1.2 Lingfeng (E-China)
At Lingfeng (2550 m, Qinling Shan) two interstadial phases with Picea-(Abies)-forests occurred at the beginning of the last glacial period. The interstadials can be correlated with phases of soil formation in the Chinese Loess Plateau and also with the Brörup and Odderade interstadials in Central Europe. Climate was presumably similar to the modern one (4.5 °C, >500 mm). Fluvial erosion disturbed the coniferous forests and favoured the spreading of pioneer species like Hippophaë and Salix and of some deciduous tree species (Ulmus, Carpinus) during the second interstadial phase. Between the two forest-phases, under stadial conditions, subalpine shrubs (Juniperus, Rhododendron) developed. A comparable subalpine phase after the end of the second interstadial was followed by a phase with alpine vegetation dominated by Cyperaceae and later by a time of a slight reforestation. Finally once more a phase with alpine conditions appeared, including the time of the so-called Last Glacial Maximum. Temperatures were at least 2.5 °C lower during the subalpine phases and 5.5 to 7.5 °C lower during the alpine phases.
8.2 Holocene
8.2.1 Nianbaoyeze (E-Tibet)
In the Nianbaoyeze mountains, a granite massif with an elevation of about 3900 to 5369 m, only small groves occurred during the Holocene. Dense alpine Kobresia-matts dominated and prevented the expansion of forests and shrubbery. Also fires, indicated by charcoal found since the older Holocene, may have contributed to the rarity of woods. Particularly at the beginning of the Holocene and during the period of the climatic optimum around 6600 BP, when the summer monsoon caused more precipitation, pioneer woods like

8. Summary – Holocene 123
Hippophaë and Salix could spread along the rivers. A slight increase of forests at the beginning of the middle Holocene (7500 BP) hints to higher temperatures. However, during the climatic optimum the temperature was less than 2 °C higher than today. Two phases of climatic deterioration are clearly indicated by pollen data, the first at about 5300 BP, the other at about 2700 BP.
Today forests of Picea are found on north facing slopes, while Juniperus trees are re-stricted to south facing slopes. The occurrence of Juniperus since the beginning of the Holocene most likely indicates, that such a vegetation pattern, depending on the exposition, is natural in the Nianbaoyeze. Therefore the open forests of Juniperus cannot be considered a kind of secondary vegetation following the cutting of Picea and Abies, but are a natural type of vegetation. This is, to a certain extent, probably also true for other areas in eastern Tibet with the same recent vegetation pattern.
An anthropo-zoogenous influence is indicated by a change of composition of plant communities during the last 800 years. The occurrence of Potentilla fruticosa as a pasture weed can be seen as a result of the opening of the plant cover by the footprints of livestock. Recent observations indicate a progressive consumption of the small natural tree reserves of the Nianbaoyeze by man.
8.2.2 Lhasa (S-Tibet)
In the eastern area of the city of Lhasa (3650 m) the recent cultivation of wheat and bar-ley were traced back over more than 600 years. During a moist phase between 1300 and 1650 AD a pond with a rich vegetation developed. The planting of Salix and Populus takes place since the 20th century.
8.2.3 Nanga Parbat (7126 m, NW-Himalaya)
In the pine forest (Pinus wallichiana) near the so called „Märchenwiese“ (3300 m) a swampy area developed during the older Holocene and expanded with the increase of summer monsoon precipitation at about 7200 BP. About 6300 BP, precipitation increased by the strengthened influence of low pressure areas from the region of the Mediterranean Sea in winter- and springtime. It seems, that avalanches opened the pine forest, so that Betula, Salix and Picea got more frequent. Whereas a decrease of the summer monsoon precipitation at about 5000 BP had only a little influence on the vegetation, the reduction of precipitation in winter- and springtime at about 3000 BP made Betula, Salix and Picea decrease. With the beginning of the younger Holocene (2700 BP) pollen of drought resistant taxa (Juniperus, Artemisia, Poaceae) and from farther away (Quercus, Cedrus) got more common. Human influence is indicated since 500 years ago only.
8.2.4 Karakoram (N-Pakistan)
Pine forests were more common in the middle Holocene than today. At the latest since the lowering of the summer monsoon precipitation around 5000 BP a dry Artemisia-steppe with some open Juniperus-forests is dominating the landscape. Locally swamps fed by

8. Summary – Holocene 124
meltwater developed in younger Holocene (2700 BP). They are grazed since that time. At 2300 BP, when more pasture weeds occurred, settlements already existed in the Yasin-valley. The increase of grazing pressure at about 2000 BP may be caused by an indirect influence from the Silk Road, a branch of which led through the Yasin-valley to the Indus. Anthropo-zoogenous influence increased further during the last centuries.
8.2.5 Conclusions
The vegetation history of the Nianbaoyeze and the Nanga Parbat proves the strengthen-ing of summer monsoon at the beginning of the middle Holocene at 7500 � P and 7200 BP respectively. A decrease of the influence of the summer monsoon took place about 5300 BP (Nianbaoyeze) and 5000 BP (Nanga Parbat, Kararkoram). A further climatic deterioration started at the beginning of the younger Holocene (2700 BP). Apart from the increase of precipitation in winter/springtime between 6300 and 3000 BP (Nanga Parbat), the climatic development during the Holocene seems to be similar to that of Central Europe (FIRBAS 1949, LANG 1994). The older Holocene of the present paper covers the Pre-Boreal and Boreal. The middle Holocene includes the Atlantic and Sub-Boreal. The transition between Atlantic and Sub-Boreal is marked by the climatic decline at 5300/5000 BP. The start of the younger Holocene at 2700 BP agrees with the world-wide occurring climatic change at the transition from the Sub-Boreal period to the Sub-Atlantic period (VAN GEEL et al. 1996).
In addition, sandy layers (Nianbaoyeze) and short lasting disturbances of the vegetation (Nanga Parbat, Karakoram) indicate the effect of temporary climatic influences like catastrophic precipitation etc. No indications were found of an influence of a tectonic rise on the climate, as assumed by some authors.

9. Nachwort und Danksagung 125
9 Nachwor t und Danksagung
„Wir können nicht bestimmt angeben, wie groß die Vermehrung oder Verminderung sein muß, damit etwas das Eine oder das Andere werde.“
(CICERO: Lehre der Akademie. Zitiert aus ST. READ (1997): Philosophie der Logik.- Reinbeck b. Hamburg. 312S.)
Bereitete es zu Anfang Probleme, ausreichend Literatur zu den verschiedenen Untersuch-ungsräumen der vorliegenden Arbeit zu beschaffen, so wandelte sich dies mit dem Fort-schreiten der Bemühungen in das Gegenteil, und es kristallisierten sich bald neue Schwierig-keiten heraus. In früherer Zeit wurde in China fast ausschließlich in der Landessprache publiziert. Die Inhalte dieser Artikel waren somit nur in den gelegentlich zu findenden englischsprachigen Übersichtsartikeln, also aus zweiter Hand, zugänglich. Schon seit einiger Zeit aber erscheinen immer mehr Aufsätze in chinesischen Zeitschriften mit englischsprachigen Zusammenfassungen und Abbildungsunterschriften. Doch es fällt oft schwer den Weg der Datenerhebung und Interpretation ganz nachzuvollziehen. Inzwischen veröffentlichen die chinesischen Wissenschaftlern vermehrt in englischer Sprache. Auf der anderen Seite hat das weltweite Interesse an der Erforschung Hochasiens – dem „Dach der Welt“ – zu Unter-suchungen von Wissenschaftlern verschiedener Nationalitäten und Schulen geführt und damit z. T. sehr unterschiedliche Interpretationsansätze der gewonnen Daten nach sich gezogen. So kostet es schon einige Zeit, sich einen Überblick zu verschaffen – entsprechend zahlreich sind die Literaturzitate.
Glücklicherweise lag der Schwerpunkt der eigenen Arbeit in der Datengewinnung an einem sehr umfangreichen Materialbestand, und die dabei gewonnenen Ergebnisse stehen – soweit Vergleiche möglich sind – im großen und ganzen mit bereits Bekanntem im Einklang. Abweichungen in einzelnen Punkten erscheinen ausreichend abgesichert.
Die Art der vorliegenden Arbeit erforderte die Festlegung von Grenzen zwischen Zeit-abschnitten unterschiedlicher paläoökologischer Ausprägung – Grenzen wie sie die Natur so nicht kennt. Notwendigerweise stößt man dabei immer wieder auf das seit Jahrtausenden bekannte im obigen Zitat angesprochene Problem des sog. Sorites. Bei der für die Erfassung und für eine sinnvolle, zur Verständigung wohl unerläßliche Gliederung der zeitlichen Abläufe standen dann auch weniger die einzelnen Prozentwerte, als vielmehr der sich in ihren Änderungen abzeichnende Wandel der Umwelt im Vordergrund.
Eine so weit gespannte Arbeit wie die vorliegende konnte natürlich nicht ohne die Unter-stützung Dritter durchgeführt werden. Allen im folgenden genannten Personen und Institutionen gebührt für ihre Hilfe mein herzlicher Dank.
An erster Stelle seien die Herren Professoren Dr. H.-J. BEUG (Göttingen), Dr. G. MIEHE
(Marburg) und Dr. H.-J. PACHUR (Berlin) genannt, deren gemeinsamer Antrag zur

9. Nachwort und Danksagung 126
dankenswerten finanziellen Unterstützung durch die DFG führte und den thematischen Rahmen der Arbeit absteckte.
Durch Herrn Prof. Dr. H.-J. BEUG erfuhr ich während des gesamten Zeitraumes meiner Promotion fortwährende Unterstützung durch Anregung und Diskussion.
Herrn Prof. Dr. G. MIEHE und Frau Dr. S. MIEHE legten durch die Beschaffung von Profilen und das Sammeln von Pflanzenbelegen während ihrer zahlreichen Geländeaufenthalte in den verschiedenen Gebieten Hochasien einen wichtigen Grundstein für die vorliegende Arbeit. Beide schufen zudem durch konstruktive Gespräche und einen gemeinsamen mehrwöchigen Geländeaufenthalt mit dem Autor ein besseres Verständnis der Zusammenhänge.
Diskussion und reger Informationsaustausch mit Herrn Prof. Dr. H.-J. PACHUR und Herrn Dr. B. WÜNNEMANN (Berlin) trugen entscheidend zur vorliegenden Interpretation der von ihnen gewonnenen pleistozänen Profile bei.
Das umfangreiche Material von Herrn PD Dr. F. LEHMKUHL (Göttingen) aus dem Nianbao-yeze und zahlreiche Gespräche schufen die Grundlage zum Erkennen der kleinräumig differenzierten Vegetationsgeschichte im östlichen Tibet.
Die von Herrn Prof. Dr. M. WINIGER (Bonn), Herrn Dr. U SCHICKHOFF (Bonn) und Herrn Dr. J.-P. JACOBSEN (Bederkesa) beigesteuerten Profile, aus dem bisher palynologisch völlig unzureichend erschlossenen nördlichen Pakistan, ergänzten die Ergebnisse am MIEHE’schen Material um ausgesprochen wichtige Resultate.
Herr Prof. Dr. M. GEYH (Hannover) trug durch zahlreiche physikalische Altersbestim-mungen zum Gelingen der Arbeit bei.
Ohne den offenen Zugang zu dem unter der Bearbeitung von Herrn Dr. B. DICKORÉ (Göttingen) stehenden „Hochasien-Herbarium“, zu dessen umfangreichen Bestand zahlreiche Personen beitragen, wäre so manches pollenanalytische Problem ungelöst geblieben und manche Interpretation ohne die Gespräche mit Herrn Dr. B. DICKORÉ nicht möglich gewesen.
Den Zugang zu Werken chinesischer Autoren eröffneten Frau Prof. SUN XIANGJUN
(Peking) und Herr Dr. SHI NING (Göttingen). An dieser Stelle soll auch die Göttinger Staats- und Universitätsbibliothek mit ihrem umfangreichen Bestand an Zeitschriftenserien und Büchern positiv hervorgehoben werden.
Herr PD Dr. T. ROST und Herr Dr. J. BÖHNER halfen mit wichtigen Informationen und Diskussionen weiter.
Zum Rückhalt während der Arbeit am Institut trugen besonders Herr Prof. Dr. H.-J. BEUG, Herr Prof. Dr. E. GRÜGER, Frau Dr. R. VOIGT, Frau Dr. L. DUPONT, Herr. Dr. F. BITTMANN, Herr Dr. SHI NING und Frau U. GROTHMANN bei.
Tatkräftige Unterstützung wurde dem Verfasser besonders durch Frau U. GROTHMANN, Herrn Dr. B. SCHÖNE, Herrn Dr. W. KRIBBE und Herrn M. KOCH zuteil.
Nicht zuletzt möchte ich meinen Eltern und Frau S. BÄTZING für Verständnis und Rücksichtnahme danken.

10. Literaturverzeichnis 127
10 Literaturverzeichnis
AGARKAR, D. S., AGARKER, M. S. & S. BANERJEE (1984): Desmids of Jabalpur, Madhya Pradesh,
India.- Bibliotheca Phycologica 66: 333-370.
AGARWAL, A. (1983): Studies in the pollen morphology of some cultivated Angiosperms.-
Advances in Pollen Spore Res. 11: 67.
AGRAWAL, D. P. (1991): Last 20,000 years of climatic change.- The Palaeobotanist 40: 393.
AGRAWAL, D. P. (1992): Man and environment in India through ages.- New Delhi. 293.
AGRAWAL, D. P. & S. K. GUPTA (1988): Climatic changes in India during the last 700,000 years.-
IN: WHYTE, P. (ed.): The palaeoenvironment of East Asia from the Mid-Tertiary. Geology,
tectonics, orogeny, geomorphology. Centre of Asian Studies Occas. Pap. a. Monogr. 77,1:
488-499.
AN ZHISHENG, LIU TUNGSHENG, ZHOU YISHI, SUN FUQING & DING ZHONGLI (1988): A prominent
climatic optimum during last 1.2 million years: records in China Loess Plateau.- IN: WHYTE,
P. (ed.): The palaeoenvironment of East Asia from the Mid-Tertiary. Geology, tectonics,
orogeny, geomorphology. Centre of Asian Studies Occas. Pap. a. Monogr. 77,1: 414-420.
AN ZHISHENG, KUKLA G. J., PORTER S. C. & XIAO JULE (1991a): Magnetic susceptibility evidence
of monsoon variation on the Loess Plateau of Central China during the last 130,000 years.-
Quat. Res. 36: 29-36.
AN ZHISHENG, WU XIHAO, WANG PINXIAN, WANG SUMIN, SUN XIANGJUN & LU YANCHOU (1991b):
An evolution model for paleomonsoon of China during the last 130000 years.- IN: LIU
TUNGSHENG (ch. ed.): Quaternary Geology and Environment in China. Science Press Beijing.
237-244.
AN ZHISHENG, WU XIHAO, LU YANCHOU, ZHANG DEER, SUN XIANGJUN, DONG GUANGRONG &
WANG SUMIN (1991c): Paleoenvironmental changes of China during the last 18000 years.-
IN: LIU TUNGSHENG (ch. ed.): Quaternary Geology and Environment in China. Science Press
Beijing. 228-236.
AN ZHISHENG, SUN DONGHUAI, ZHANG XIAOYE & ZHOU WEIJIAN (1995): Accumulation sequence
of chinese loess and climatic records of Greenland Ice Core during the last 130 ka.- Chinese
Scie. Bull. 40: 1272-1276.
ANDERSEN, S. T. & F. BERTELSEN (1972): Scanning electron microscope studies of pollen of
cereals and other grasses.- Grana 12: 79-86.
ANIYA M. & K. YOKOYAMA (1991): Glacial limits, climate and uplift in the Mount Meylixueshan
area of Yunnan Province, China: a simulation modelling approach.- The Holocene 1: 273-
280.

10. Literaturverzeichnis 128
AUTORENGRUPPE (1982): Angiosperm pollen flora of tropic and subtropic China.- Institute of
Botany and South Chinas Institute of Botany. Academia Sinica. 453.
AVOUAC, J.-P., DOBREMEZ, J.-F. & L. BOURJOT (1996): Palaeoclimatic interpretation of a
topographic profile across middle Holocene regressive shorelines of Longmu Co (Western
Tibet).- Palaeogeogr. Palaeoclimat. Palaeoecol. 120: 93-104.
BARRIENTOS, O. O. P. (1979): Revision der Gattung Pediastrum MEYEN (Chlorophyta).-
Bibliotheca Phycologica 48: 183.
BARTENS, H. (1990): Untersuchungen über die Vegetationsgeschichte des Bruchberges im
Oberharz.- Unveröff. Dipl.-Arbeit, Göttingen. 64.
BARTHEL, H. (1990): Mongolei - Land zwischen Taiga und Steppe.- Geogr. Bausteine 8: 220.
BAT, S., OTSCHIRBAT, G. & U. LIGAA (1988): Natürliche Vorkommen von Sanddorn in der
Mongolischen Volksrepublik.- Gartenbau 35: 85-86.
BEAZLEY, M. (1985): The world atlas of Archaeology.- Malesherbs, France. 423.
BELYAEVSKY, N. A. (1976): Tectonic of the nothern part of the Pamirs-Himalayan Syntaxis.- Atti
dei convegni lincei 2: 29-42.
BENDER, F. K. (1995a): Geological framework. Introduction.- IN: BENDER, F. K. & H. A. RAZA
(eds.): Geology of Pakistan.- Beitr. z. Regio. Geol. d. Erde 25: 11-17.
BENDER, F. K. (1995b): Palaeogeography and geodynamic evolution.- IN: BENDER, F. K. & H. A.
RAZA (eds.): Geology of Pakistan.- Beitr. z. Regio. Geol. d. Erde 25: 162-181.
BENDER, H. (1995c): Water.- IN: BENDER, F. K. & H. A. RAZA (eds.): Geology of Pakistan. Beitr.
z. Regio. Geol. d. Erde 25: 291-328.
BENECKE, N. (1994): Der Mensch und seine Haustiere. Die Geschichte einer jahrtausende alten
Beziehung.- Stuttgart. 470.
BENNINGHOFF, W. S. (1962): Calculation of pollen and spores density in sediments by addition of
exotic pollen in known quantities.- Pollen et Spores 4: 332-333.
BERTSCH, A. (1961): Untersuchungen an rezenten und fossilen Pollen von Juniperus.- Flora 150:
503-513.
BEUG, H.-J. (1957): Untersuchungen zur spätglazialen und frühpostglazialen Floren- und Vege-
tationsgeschichte einiger Mittelgebirge (Fichtelgebirge, Harz und Rhön).- Flora 145: 167-
211.
BEUG, H.-J. (1961): Leitfaden der Pollenbestimmung für Mitteleuropa und angrenzende Gebiete.-
Lieferung 1. Stuttgart 63.

10. Literaturverzeichnis 129
BEUG, H.-J. (1987): Palynological studies on a peat layer in Kakitu Mountain, northeastern
Qinghai-Xizang Plateau.- IN: HÖVERMANN & WENYING WANG (ch. eds.): Reports on the
northeastern part of the Qinghai-Xizang (Tibet) Plateau by Sino-W. German Scientific
Expedition. Science Press Beijing. 496-501.
BEUG, H.-J. & G. MIEHE (1998): Vegetationsgeschichtliche Untersuchungen in Hochasien. 1.
Anthropogene Vegetationsveränderungen im Langtang-Tal, Himalaya, Nepal. Petermanns
Geogr. Mitt. 142: 141-148.
BEUG, H.-J. & G. MIEHE (1999): Vegetation history and human impact in the Eastern Cenral
Himalaya.- Diss. Bot. in Vorbereitung.
BHATIA, S. B. & N. SINGH (1988): Middle holocene palaeoclimatic and palaeoenvironmental
events in southern Haryana.- IN: JAIN, D. V. S., DHARA, M. & J. SAKETHARAMAN (eds.):
Palaeoclimatic and palaeoenvironmental changes in Asia during the last 4 million years. New
Delhi. 236-246.
BHATT, D. K. (1989): The lithostratigraphy of Karewa Group, Kashmir Valley, India and a critical
review of its fossil record.- Geol. Survey of India 122: 85.
BHATTACHARYYA, A. (1989): Vegetation and climate during the last 30,000 years in Ladakh.-
Palaeogeogr. Palaeoclimat. Palaeoecol. 73: 25-38.
BILHAM, R., PANT, C. B. & G. C. JACOBY (1984): A preliminary study of ancient trees in the
Hunza Valley and their dendroclimatic potential.- IN: MILLER, K. J. (ed.): The international
Karakoram Project. Cambridge. 2: 599-606.
BJÖRK, S. (1993): The Hongyuan wetland research project. An ecological and technical feasibility
study of peat mining in Hongyuan, Sichuan, China.- Departement of Limnology University
of Lund. 100
BLACKMORE, S. (1982): Palynology of subtribe Scorzonerinae (Compositeae: Lactuceae) and its
taxonomic significance.- Grana 21: 149-160.
BLACKMORE, S. (1984): Compositae - Lactuceae.- IN: PUNT, W. & G. C. S. CLARKE (edts.): The
northwest european pollen flora 4: 45-85.
BLUNDEN, C. B. & M. ELVIN (1992): Weltatlas der Alten Kulturen. China.- 6. Aufl. München. 239.
BLÜTHGEN, J. & W. WEISCHET (1980): Allgemeine Klimageographie.- 3. Aufl. Berlin. 887.
BÖHNER, J. (1996): Säkulare Klimaschwankungen und rezente Klimatrends Zentral- und
Hochasiens.- Göttinger Geogr. Abh. 101: 166.
BONNEFILLE, R. & G. RIOLLET (1980): Pollens des savanes D'Afrique orientale.- Editions du
Centre National de la Recherche Scientifique. Paris. 140.

10. Literaturverzeichnis 130
BORTENSCHLAGER, S. (1967): Vorläufige Mitteilungen zur Pollenmorphologie in der Familie der
Geraniaceen und ihre systematische Bedeutung.- Grana 7: 400-468.
BORTENSCHLAGER, S. (1990): Aspects of pollen morphology in the Cupressaceae.- Grana 29: 129-
137.
BORTENSCHLAGER, S. & G. PATZELT (1978): Das Pollendiagramm vom Kol-e Ptukh (3272 m) im
östlichen Wakhan.- IN: GRANCY, R. S. DE & R. KOSTKA (Hrsg.): Großer Pamir. Graz/Austria.
193-200.
BOS, J. A. A. & W. PUNT (1991): Juglandaceae.- IN: PUNT, W. & S. BLACKMORE (edts.): The
northwest european pollen flora 6: 79-95.
BOSINSKI, G. (1992): Die ersten Menschen in Eurasien.- Jahrb. d. Röm.-Germ. Zentralmuseums
Mainz. 39: 131-181.
BRAUN, G. (1996): Vegetationsgeographische Untersuchungen im NW-Karakorum (Pakistan).-
Bonner Geogr. Abh. 93: 156.
BRÄUNING, A. (1994): Dendrochronology for the last 1400 years in eastern Tibet.- IN: LEHMKUHL,
F. & LIU SHIJIAN (eds.): Landscape and quaternary climatic changes in eastern Tibet and
surroundings. GeoJour. 34: 75-95.
BRÄUNING, A. & F. LEHMKUHL (1996): Glazialmorphologische und dendrochronologische
Untersuchungen neuzeitlicher Eisrandlagen Ost- und Südtibets.- Erdkunde 50: 341-359.
BRENTJES, B. (1984): Die "Baktrischen Bronzen" - Ein Brückenpfeiler zwischen Ost- und
Westasien?- IN: BRENTJES, B. & H.-J. PEUKE (Hrsg.): Kulturhistorische Probleme Südasiens
und Zentralasiens.- Wissenschaft. Beitr. 1984/25: 51-71.
BRENTJES, B. (1991): Klimaschwankungen, Umweltveränderungen und Geschichte in
Zentralasien.- Hallesche Beiträge zur Orientwissenschaft. Wissenschaft. Beitr. 1991,16: 5-
31.
BRONGER, A. & T. HEINKELE (1989): Paleosol sequences as witnesses of pleistocene climatic
history.- Catena, Suppl. 16: 163-186.
BRYSON, R. A. & A. M. SWAIN (1981): Holocene variations of monsoon rainfall in Rajasthan.-
Quat. Res. 16: 135-145.
BUCHROITHNER, M. F. (1978): Zur Geologie des Afghanischen Pamir.- IN: GRANCY, R. S. DE & R.
KOSTKA (Hrsg.): Großer Pamir. Graz/Austria. 85-118.
BURKI, S. J. (1991): Historical dictionary of Pakistan.- Asian Historical Dictionaries Bd. 3.
Metuchen & London. 251.
CHANG CHENGFA (1996): Geology and tectonics of Qinghai-Xizang Plateau.- A Series of Solid
Earth Scie. Res. in China. 153.

10. Literaturverzeichnis 131
CHANG, D. H. S. (1981): The vegetation zonation of the Tibetan Plateau.- Mount. Res. a. Develop.
1: 29-48.
CHANG, DAVID H. S. (1983): The Tibetan Plateau in relation to the vegetation of China.- Annals
of the Missouri Bot. Gard. 70: 564-570.
CHEN CUNGEN (1987): Standörtliche, vegetationskundliche und waldbauliche Analyse
chinesischer Gebirgsnadelwälder und Anwendung alpiner Gebirgswaldbau-Methoden im
chinesischen fichtenreichen Gebirgsnadelwald.- Diss. Univ. f. Bodenkultur, Wien, VWGÖ,
30: 316.
CHEN FENGHUAI & HU QIMING (1981): Distribution of the genus Primula L. in Xizang.- IN: LIU
DONGSHENG (ch. ed.): Geological and ecological studies of Qinghai-Xizang Plateau. Science
Press Beijing. 2: 1299-1303.
CHEN GUICHEN, PENG MIN, HUANG RONGFU & LU XUEFENG (1994): Vegatation characteristics and
its distribution of Qilian Mountain region.- Acta Bot. Sinica 36: 63-72.
CHEN KEZAO & XU ZHIQIANG (1991): Salt lakes in China.- IN: LIU TUNGSHENG (ch. ed.):
Quaternary Geology and Environment in China. Science Press Beijing. 434-441.
CHEN KEZAO, YANG SHAOXIU & ZHEN XIYU (1981): The saline lakes on the Qinhgai-Xizang
Plateau.- IN: LIU DONGSHENG (ch. ed.): Geological and ecological studies of Qinghai-Xizang
Plateau. Science Press Beijing. 2: 1719-1724.
CHEN WEILIE (1981): The main types of forest vegetation and their distribution in Xizang.- IN:
LIU DONGSHENG (ch. ed.): Geological and ecological studies of Qinghai-Xizang Plateau.
Science Press Beijing. 2: 1947-1952.
CHEN WEILIE, & ZHANG JINGWEI, WANG JINTING & LI BOSHENG (1980): The pines and pine forests
of Xizang.- Acta Bot. Sinica 22: 170-176.
CHEN ZHIMING (1996): Sequential analysis of China's hazards in geoscience.- GeoJour. 38: 259-
263.
CLARKE, G. (1978): Pollen morphology and generic relationships in the Valerianaceae.- Grana 17:
61-75.
CLARKE, G. C. S. & M. R. JONES (1980a): Plantaginaceae.- IN: PUNT, W. & G. C. S. CLARKE
(edts.): The northwest european pollen flora 2: 129-154.
CLARKE, G. C. S. & M. R. JONES (1980b): Valerianaceae.- IN: PUNT, W. & G. C. S. CLARKE
(edts.): The northwest european pollen flora 2: 155-179.
CLARKE, G. C. S. & M. R. JONES (1981): Dipsacaceae.- IN: PUNT, W. & G. C. S. CLARKE (edts.):
The northwest european pollen flora 3: 1-25.

10. Literaturverzeichnis 132
CLARKE, G. C. S., PUNT, W. & P. P. HOEN (1991): Ranunculaceae.- IN: PUNT, W. & S.
BLACKMORE (edts.): The northwest european pollen flora 6: 117-271.
CUI ZHIJIU, WU YONGQIU & GENGNIAN LIU (1995): Discovery and environmental significance of
neolithic trace in Kunlun Mountains Pass area.- Chinese Scie. Bull. 40: 1451-1455.
CUTLER, D. F., RUDALL, P. J., GASSON, P. E. & R. M. O. GALE (1987): Root identification manual
of trees and shrubs.- London. 245.
DAMM, B. (1993): Weidewirtschaft, Faktoren und Ausmaß von Degradationserscheinungen in
Hochweidegebieten Osttibets (Sichuan/Qinghai), China.- Geoökodynamik 14: 117-139.
DANI, A. H. (1986): The historic city of Taxila.- Japan. Hinode Printing. 190.
DANI, A. H. & P. BERNARD (1994): Alexander and his successors in Central Asia.- IN: HARMATTA,
J., PURI, B. N. & G. F. ETMADI (eds.): The development of sedentary and nomadic
civilizations: 700 B. C. to A. D. 250. History of civilizations of Central Asia. UNESCO
Publ. 2: 67-97.
DERBYSHIRE, E. (1996): Quaternary glacial sediments, glaciation style, climate and uplift in the
Karakoram and Northwest Himalaya: review and speculations.- Palaeogeogr. Palaeoclimat.
Palaeoecol. 120: 147-157.
DERBYSHIRE, E., LI JIJUN, PERROTT, F. A., XU SHUYING & R. S. WATERS (1984): Quaternary
glacial history of the Hunza Valley, Karakoram Mountains, Pakistan.- IN: MILLER, K. J.
(ed.): The international Karakoram Project. Cambridge. 2: 456-495.
DESIO, A. (1976): Some geotectonic problems of the Kashmir Himalaya-Karakorum-Hindu Kush
and Pamir area.- Atti dei convegni lincei 2: 115-129.
DEXEL, T. (1973): Frühe Keramik in China: Die Entwicklung der Hauptformen vom Neolithikum
bis in die T'ang-Zeit.- Braunschweig. 212.
DICKORÉ, W. B. (1995): Systematische Revision und chorologische Analyse der Monocotyledonae
des Karakorum (Zentralasien, West-Tibet) = Flora Karakorumensis. I. Angiospermae,
Monocotyledoneae.- Stapfia 39: 298.
DING YIHUI (1994): Monsoons over China.- Atmospheric Scie. Library 16: 419.
DING ZHONGLI (1996): Correlation of climatic periodicities between loess and deep sea record in
the past 2.5 Ma.- IN: PANG ZHONGHE, ZHANG JINDONG & SUN JIANHONG (eds.): Advances in
solid earth sciences. Science Press Beijing. 59-63.
DODIA, R., AGRAWAL, D. P. & A. B. VORA (1984): New pollen data from the Kashmir bogs.- IN:
WHYTE, R. O., CHIU TZENANG, LEUNG CHIKEUNG & SO CHAKLAM (eds.): The evolution of
the East Asian environment: palaeobotany, palaeozoology and palaeoanthropology. Centre of
Asian Studies Occas. Pap. a. Monogr. 59,2: 569-578.

10. Literaturverzeichnis 133
DOMRÖS, M. & PENG GONGBING (1988): The climate of China.- Berlin, Heidelberg. 360.
DRESCHER-SCHNEIDER, R. (1993): Funde nordafrikanischer Pollen in spät- und postglazialen
Sedimenten am Südrand der Alpen.- IN: BROMBACHER, C., JACOMET, S. & J. N. HAAS
(Hrsg.): Festschrift Zoller. Diss. Bot. 196: 415-425.
DUHOUX, E. (1982): Mechanism of exine rupture in hydrated taxoid type of pollen.- Grana 21: 1-
7.
EBBLIN, C. (1988): Geological structure of the Yasin-Ishkuman area, Karakorum, Pakistan.- Geol.
Survey of Pakistan Rec. 43: 15.
EBERHARD, W. (1948): Chinas Geschichte.- Bern. 404.
EIDE, F. (1981): Key for northwest european Rosaceae pollen.- Grana 20: 101-118.
EIKELAND, F. E. (1979): Species distinction in the "Potentilla pollen type" (Rosaceae) of NW
Europe.- Grana 18: 189-192.
EISELE, G., HAAS, K. & S. LINER (1994): Methode zur Aufbereitung fossilen Pollens aus
minerogenen Sedimenten.- Göttinger Geogr. Abh. 95: 165-166.
EL GHAZALI, G. E. B. (1993): A study on the pollen flora of Sudan.- Rev. of Palaeobot. a.
Palynol. 76: 99-345.
ENGEL, M. S. (1980a): Haloragaceae.- IN: PUNT, W. & G. C. S. CLARKE (edts.): The northwest
european pollen flora 2: 199-208.
ENGEL, M. S. (1980b): Hippuridaceae.- IN: PUNT, W. & G. C. S. CLARKE (edts.): The northwest
european pollen flora 2: 195-198
ENGLER, A. (1954): Syllabus der Pflanzenfamilien. I. Allgemeiner Teil, Bakterien,
Gymnospermen.- Berlin-Nikolassee. 367.
ENGLER, A. (1964): Syllabus der Pflanzenfamilien. II. Angiospermen und Übersicht über die
Florengebiete der Erde.- Berlin-Nikolassee. 666.
ENOKI, K., KOSHELENKO, G. A. & Z. HAIDARY (1994): The Yüeh-chih and their migrations.- IN:
HARMATTA, J., PURI, B. N. & G. F. ETMADI (eds.) (1994): The development of sedentary and
nomadic civilizations: 700 B. C. to A. D. 250. History of civilizations of Central Asia.
UNESCO Publ. 2: 171-189.
ERDTMAN, G. (1952): Pollen morphology and plant taxonomy - Angiosperms.- An introduction to
Palynology. I. Stockholm. 539.
ERDTMAN, G. (1957): Pollen and spore morphology/plant taxonomy. Gymnospermae,
Pteridophyta, Bryophyta (Illustrations).- An introduction to Palynology. II. Stockholm. 151.

10. Literaturverzeichnis 134
ERDTMAN, G. & G. NORDBORG (1961): Über Möglichkeiten die Geschichte verschiedener
Chromosomenzahlenrassen von Sanguisorba officinalis und S. minor pollenanalytisch zu
beleuchten.- Botaniska Notiser 114: 19-21.
ERKES, E. (1956): Geschichte Chinas von den Anfängen bis zum Eindringen des ausländischen
Kapitals.- Berlin. 155.
FAEGRI, K. & J. IVERSEN (1989): Textbook of pollen analysis.- Chichester, New York. 4th. ed.
328.
FAN HUI, GASSE F., HUC A., LI YUANFANG, SIFEDDINE, A. & I. SOULIÉ-MÄRSCHE (1996): Holocene
environmental changes in Bangong Co basin (Western Tibet). Part 3: Biogenic remains.-
Palaeogeogr. Palaeoclimat. Palaeoecol. 120: 65-78.
FANG JINQI (1991): Lake evolution during the past 30,000 years in China, and its implications for
environmental change.- Quat. Res. 36: 37-60.
FANG JINQI (1993): Lake evolution during the past 3000 years in China and its implications for
environmental change.- Quat. Res. 39: 175-185.
FANG RUIZHENG & MING TIANLU (1981): Differentiation and development of genus Rhododendron
during the process of the uplift of Qinghai-Xizang Plateau.- IN: LIU DONGSHENG (ch. ed.):
Geological and ecological studies of Qinghai-Xizang Plateau. Science Press Beijing. 2:
1289-1298.
FANG XIAOMIN (1995): The origin and provenance of Malan loess along the eastern margin of the
Qinghai-Xizang (Tibetan) Plateau and its adjacent area.- Science in China (B) 38: 876-887.
FASSL, K. (1996): Die Bewertung von Zeigerarten in europäischen Pollendiagrammen für die
Rekonstruktion des Klimas im Holozän.- Paläoklimaforschung 22: 371.
FENG SHENGWU (1985): Origin of chinese agriculture as viewed from Dadiwan Cultural relicts.-
Acta Geogr. Sinica 40: 207-214.
FENG ZHAODONG, THOMSON L. G., MOSLEY-THOMPSON E. & YAO TANGDONG (1993): Temporal
and spatial variations of climate in China during the last 10000 years.- The Holocene 3: 174-
180.
FEZER, F. & R. HALFAR (1994): Oasen im winterkalten Zentralasien in den Provinzen Gansu und
Qinghai in Westchina.- IN: DOMRÖS, M. & W. KLAER (1994): Festschrift für Erdtmann
Gormsen zum 65. Geburtstag. Mainzer Geogr. Schr. 40: 643-652.
FIELDING, E. J. (1996): Tibet uplift and erosion.- Tectonophysics 260: 55-84.
FIRBAS, F. (1937): Der pollenanalytische Nachweis des Getreidebaus.- Zeitschrift f. Botanik 31:
447-478.

10. Literaturverzeichnis 135
FIRBAS, F. (1949): Spät- und nacheiszeitliche Waldgeschichte Mitteleuropas nördlich der Alpen.
1. Band: Allgemeine Waldgeschichte.- Jena. 480.
FIRBAS, F. (1952): Spät- und nacheiszeitliche Waldgeschichte Mitteleuropas nördlich der Alpen.
2. Band: Waldgeschichte der einzelnen Landschaften.- Jena. 256.
FITSCHEN, J. (1987): Gehölzflora. 8. Aufl.- Bearbeitet von MEYER, F. H., HECKER, U., HÖSTER,
H. R. & F.-G. SCHRÖDER. Heidelberg, Wiesbaden. ohne Seitenzählung.
FLOHN, H. (1968): Contributions to a meteorology of the Tibetan Highlands.- Dept. Atmospheric
Scie., Colorado State Univ., Fort Collins, Colorado, Atmospheric Scie. Pap. 130: 120.
FLOHN, H. (1981): The elevated heat source of the Tibetan Highlands and its role for the large-
scale atmospheric circulation.- IN: LIU DONGSHENG (ch. ed.): Geological and ecological
studies of Qinghai-Xizang Plateau. Science Press Beijing. 2: 1463-1470.
FONTES, J.-C., GASSE, F. & E. GIBERT (1996): Holocene environmental changes in Lake Bangong
basin (Western Tibet). Part 1: Chronology and stable isotopes of carbonates of a holocene
lacustrine core,- Palaeogeogr. Palaeoclimat. Palaeoecol. 120: 25-47.
FORMAN, S. L. (1991): Late pleistocene chronology of loess deposition near Luochuan, China.-
Quat. Res. 36: 19-28.
FORT, M. (1996): Late cenozoic environmental changes and uplift on the nothern side of the
central Himalaya: A reappraisal from field data.- Palaeogeogr. Palaeoclimat. Palaeoecol.
120: 123-145.
FRENZEL, B. (1993): Klimageschichte Hochtibets während des jüngeren Quartärs;
Methodologische Arbeiten zur Palynologie.- Akad. d. Wiss. u. d. Lit. Mainz, Jahresb. 1993:
165-171.
FRENZEL, B. (1994): Über Probleme der holozänen Vegetationsgeschichte Osttibets.- Göttinger
Geogr. Abh. 95: 143-166.
FREY, W., FRAHM, J.-P., FISCHER, E. & W. LOBIN (1995): Die Moos- und Farnpflanzen Europas. 6.
Auflage.- IN: Kleine Kryptogamenflora, begründet von H. GAMS. 4: 426.
FUKAREK (1993): Blütenpflanzen 1.- IN: Urania Pflanzenreich. 590.
FUKAREK (1994): Blütenpflanzen 2.- IN: Urania Pflanzenreich. 609.
FUKAREK, F., SCHULTZE-MOTEL, J. & M. SIEGEL (1992): Moose, Farne, Nacktsamer.- IN: Urania
Pflanzenreich. 390.
FURRER, G. (1986): Solifluidale Bodenformen und Gliederung der subnivalen Höhenstufe im
Braldo- und Biafotal (Karakorum).- IN: KUHLE, M. (Hrsg): Internationales Symposium über
Tibet und Hochasien vom 8. - 11. Oktober 1985 im Geographischen Institut der Universität
Göttingen. Göttinger Geogr. Abh. 81: 143-144.

10. Literaturverzeichnis 136
GARCKE, A. (1972): Illustrierte Flora. Deutschland und angrenzende Gebiete.- Berlin, Hamburg.
1607.
GASSE, F., FONTES, J. C., VAN CAMPO, E. & K. WEI (1996): Holocene environmental changes in
Bangong Co basin (Western Tibet). Part 4: Discussion and conclusions.- Palaeogeogr.
Palaeoclimat. Palaeoecol. 120: 79-92.
GERNET, J. (1994): Die chinesische Welt.- Suhrkamp Verlag. 714.
GLASENAPP, H. VON (1996): Die fünf Weltreligionen: Hinduismus, Buddhismus, chinesischer
Universismus, Christentum, Islam.- München. 480.
GOLDAMMER, J. G. (1993): Feuer in Waldökosystemen der Tropen und Subtropen.- Basel, Boston,
Berlin. 251.
GOUDIE, A. S., BRUNSDEN, D., COLLINS, D. N., DERBYSHIRE, E., FERGUSON, R. I., HASHMET, Z.,
JONES, D. K. C., PERROTT, F. A., SAID, M., WATERS, R. S. & W. B. WHALLEY (1984): The
geomorphology of the Hunza Valley, Karakoram Mountains, Pakistan.- IN: MILLER, K. J.
(ed.): The international Karakoram Project. Cambridge. 2: 359-410.
GRÜGER, E. (1989): Palynostratigraphy of the last Interglacial/Glacial cycle in Germany.- Quat.
Intern. 3/4: 69-79.
GRÜGER, E (1995): Correlation of middle-european late-pleistocene pollen sequences of the
Pfefferbichl and Zeifen types.- Mededelingen Rijks Geologische Dienst 52 :97-104.
GRÜNEBERG, F. H. (1995): Soils. IN: BENDER, F. K. & H. A. RAZA (eds.): Geology of Pakistan.
Beitr. z. Regio. Geol. d. Erde 25: 329-355.
GRUSCHKE, A. (1997): Demographie und Ethnographie im Hochland von Tibet.- Geogr. Rundsch.
49: 279-286.
GULLVÅG, B. M. (1966): The fine structure of some Gymnosperm pollen walls.- Grana 6: 435-
475.
GUO SHUANGXING (1981): On the elevation and climatic changes of the Qinghai-Xizang Plateau
based on fossil Angiosperms.- IN: LIU DONGSHENG (ch. ed.): Geological and ecological
studies of Qinghai-Xizang Plateau. Science Press Beijing. 1: 201-206.
GUO SHUANGXING (1985): Preliminary interpretation of tertiary climate by using megafossil floras
in China.- Palaeontologia Cathayana 2: 169-176.
GUO SHUANGXING (1988): The palaeoenvironment in the Hengduan Mountain area since the
Eocene.- IN: WHYTE, P. (ed.): The palaeoenvironment of East Asia from the Mid-Tertiary.
Geology, tectonics, orogeny, geomorphology. Centre of Asian Studies Occas. Pap. a.
Monogr. 77,1: 523-527.

10. Literaturverzeichnis 137
GUPTA, H. P. (1991): Changing pattern of vegetation in the intermontane basin of Kashmir since 4
Ma: a palynological approach.- The Palaeobotanist 4: 354-373.
GUPTA, H. P. & C. SHARMA (1986): Pollen flora of North-West Himalaya.- Lucknow. 181.
GUPTA, H. P. & S. K. BERA (1995): Silent Valley: A correlation between pollen spectra and
vegetation.- The Palaeobotanist 43: 139-144.
HAFFNER, W. (1997): Hochasien: Der Effekt großer Massenerhebungen.- Geogr. Rundsch. 49:
307-314.
HAHNE, J. & M. MELLES (1997): Late- and post-glacial vegetation and climate history of the
south-western Taymyr Peninsula, central Siberia, as revealed by pollen analysis of a core
from Lake Lama.- Veget. Hist. a. Archaeobot. 6: 1-8.
HAMBLY, G. (Hrsg.) (1995): Zentralasien.- Fischer Weltgeschichte. Frankf. a. M. 16: 365.
HAMILTON, A. C. & R. A. PERROTT (1980): Modern pollen deposition on a tropical african
mountain.- Pollen et Spores 22: 437-468.
HANDEL-MAZZETTI, H. (1936): VII. Teil: Anthophyta.- IN: HANDEL-MAZZETTI, H. (Hrsg.):
Botanische Ergebnisse der Expedition der Akademie der Wissenschaften in Wien nach
Südwest-China 1914/1918. Symbolae Sinicae 7: 1450.
HARALDSON, K. (1978): Anatomy and taxonomy in Polygonaceae Subfam. Polygonoideae MEISN.
EMEND. JARETZKY.- Acta Univ. Upsaliensis Symb. Bot. Upsalienses 22,2: 96.
HARRISON, T. M., COPELAND, P., KIDD, W. S. F. & Y. AN (1992): Raising Tibet.- Science 255:
1663-1670.
HARTMANN, H. (1987): Pflanzengesellschaften trockener Standorte aus der subalpinen und
alpinen Stufe im südlichen und östlichen Ladakh.- Candollea 42: 277-326.
HASERODT, K. (1989): Zur pleistozänen und postglazialen Vereisung zwischen Hindukusch,
Karakorum und Westhimalaya.- IN: HASERODT, K. (Hrsg.): Hochgebirgsräume Nordpakistans
im Hindukusch, Karakorum und Westhimalaya. Beitr. u. Material. z. Region. Geogr. 2: 181-
233.
HE DAZHANG & HE DONG (1988): On the nothern boundary of tropical climate in China.- Acta
Geogr. Sinica 43: 176-183.
HE TINGNONG (1981): The floristic analysis of Gentianaceae in Xizang and its geographical
relationships.- IN: LIU DONGSHENG (ch. ed.): Geological and ecological studies of Qinghai-
Xizang Plateau. Science Press Beijing. 2: 1305-1312.
HEAD, M. J., ZHOU WEIJIAN & ZHOU MINGFU (1989): Evaluation of 14C ages of organic fractions
of paleosols from loess-paleosol sequences near Xian, China.- 31: 680-696.

10. Literaturverzeichnis 138
HEDBERG, O. (1946): Pollen morphology in the genus Polygonum L. s.lat. and its taxonomical
significance.- Svensk Botanisk Tidskrift 40: 371-404.
HEGDE, K. T. M. (1977): Late quaternary environments in Gujarat and Rajasthan.- IN: AGRAWAL,
D. P. & B. M. PANDE (eds.): Ecology and Archaeology of Western India. Delhi. 169-180.
HENDL, M., MARCINEK, J. & E. J. JÄGER (1978): Allgemeine Klima-, Hydro- und
Vegtationsgeographie.- Gotha/Leipzig. 166.
HERMANNS, M. (1949): Die Nomaden von Tibet.- Wien. 325.
HEYWOOD, V. H. (Hrsg.) (1982): Blütenpflanzen der Welt.- Basel. 356.
HILBIG, W. (1995): The vegetation of Mongolia.- Amsterdam. 258.
HILBIG W. & H. D. KNAPP (1983): Vegetationsmosaik und Florenelemente an der Wald-Steppen-
Grenze im Chentej-Gebirge (Mongolei).- Flora 174: 1-89.
HILBIG W., STUBBE M., DAWAA, N., SCHAMSRAN, Z., DORN, M., HELMECKE, K., BUMZAA, D. & K.
ULYPKAN (1984): Vergleichend biologisch-ökologische Untersuchungen in Hochgebirgen der
Nordwest- und Südostmongolei. Allgemeiner Bericht. IN: Biologische Ressourcen der
Mongolischen Volksrepublik 4. Wissenschaft. Beitr. 1984,59: 5-49.
HILDEBRAND, J. (1990): China. Erlangen. 148.
HOCHULI, P. A. & U. GRONER (1985): Heerisporites, a new spore-genus of possible Riellacean
affinity.- Rev. of Palaeobot. a. Palynol. 44: 261-275.
HOFMANN, J. (1993): Geomorphologische Untersuchungen zur jungquartären Klimaentwicklung
des Helan Shan und seines westlichen Vorlandes (Autonomes Gebiet Innere Mongolei/VR
China).- Berliner Geogr. Abh. 57: 1-187.
HOLMES, J. A. & A. STREET-PERROTT (1989): The quaternary glacial history of Kashmir, North-
West Himalaya: A Revision of de Terra and Paterson's sequenze.- Zeitsch. f. Geomorphol. N.
F. Supp. 76: 195-212.
HOLTMEIER, F.-K. (1985): Die klimatische Waldgrenze – Linie oder Übergangsraum (Ökoton)?
Ein Diskussionsbeitrag unter besonderer Berücksichtigung der Waldgrenzen in den mittleren
und hohen Breiten der Nordhalbkugel.- Erdkunde 39: 271-285.
HONG BIGONG & AN SHUQING (1993): Preliminary studies on the geographic distribution of Fagus
in China.- Acta Bot. Sinica 35: 229-233.
HONG SUKPYO (1991): Taxonomy of Aconogonon s. lat. (Polygonaceae).- Acta Univ. Upsaliensis
Compreh. Summar. 351: 30.
HONG SUKPYO (1992): Taxonomy of the genus Aconogonon (Polygonaceae) in Himalaya and
adjacent regions.- Acta Univ. Upsaliensis Symb. Bot. Upsalienses 30,2: 118.

10. Literaturverzeichnis 139
HONG SUKPYO & O. HEDBERG (1990): Parallel evolution of aperture numbers and arrangement in
the genera Koenigia, Persicaria and Aconogonon (Polygonaceae).- Grana 29: 177-184.
HOROWITZ, A. (1992): Palynology of arid lands.- Elsevier Science Publishers B. V. 546.
HOU HSIOHYU (1983): Vegetation of China with reference to its geographical distribution.- Annals
of the Missouri Bot. Gard. 70: 509-549.
HOU JIANJUN, HAN MUKANG, ZHANG BAOZENG, CHAI BAOLONG & HAN HENGYUE (1995):
Geomorphic expressions of the activity along north Qinling piedmont fault zone in the late
quaternary period.- Acta Geogr. Sinica 50: 138-146.
HÖVERMANN, J. & F. LEHMKUHL (1994): Vorzeitliche und rezente geomorphologische Höhen-
stufen in Ost- und Zentraltibet. - Eine Bestandsaufnahme -.- IN: PÖRTGE, K.-H. & LI JIAN
(Hrsg.): Beiträge zur Tibetforschung. Göttinger Geogr. Abh. 95: 15-69.
HÖVERMANN, J. & H. SÜSSENBERGER (1986): Zur Klimageschichte Hoch- und Ostasiens.- Berliner
Geogr. Studien 20: 173-186.
HÖVERMANN, J. & WANG WENYING (ch. edts.) (1987): Reports on the northeastern part of the
Qinghai-Xizang (Tibet) Plateau by Sino - W. German Scientific Expedition.- Science Press
Beijing. 510.
HOW FOONCHEW, WU TELIN, KO WANCHEUNG & CHEN TECHOA (1982): A dictionary of the
families and genera of chinese seed plants (chin.).- 2nd. ed. 632.
HSÜ JEN (1983): Late cretaceous and cenozoic vegetation in China, emphasizing their connections
with North America.- Annals of the Missouri Bot. Gard. 70: 490-508.
HUANG CIXUAN & LIANG YULIAN (1981): Based upon palynological study to discuss the natural
environment of the central and southern Qinghai-Xizang Plateau of Holocene.- IN: LIU
DONGSHENG (ch. ed.): Geological and ecological studies of Qinghai-Xizang Plateau. Science
Press Beijing. 1: 215-224.
HUANG RONGFU (1987): Vegetation in the northeastern part of the Qinghai-Xizang Plateau.- IN:
HÖVERMANN & WENYING WANG (ch. eds.): Reports on the northeastern part of the Qinghai-
Xizang (Tibet) Plateau by Sino-W. German Scientific Expedition. Science Press Beijing.
438-495.
HUANG TSENGCHIENG (1972): Pollen flora of Taiwan.- National Taiwan Univ. Bot. Depart. Press.
297.
HUANG TSENGCHIENG (1981): Spore flora of Taiwan (Pteridophyta).- National Taiwan Univ. 111.
HUANG WEIWEN (1989): The early Paleolithic of China.- The Quat. Res. 28: 237-242.

10. Literaturverzeichnis 140
HUANG WEIWEN (1990): The Late Paleolithic in North China.- IN: SHAKHMATOVA, N. M. (ed.):
Chronostratigraphy of the Paleolithic in North, Central, East Asia and America (Pap. for the
Intern. Symp.). Novosibirsk. 35-45.
HUANG WEIWEN (1994): The prehistoric human occupation of the Qinghai-Xizang Plateau.-
Göttinger Geogr. Abh. 95: 201-219.
HUGHES, R. E. (1984): Yasin Valley: The analysis of geomorphology and building types.- IN:
MILLER, K. J. (ed.): The international Karakoram Project. Cambridge. 2: 253-288.
JACOBSEN, J.-P. & U. SCHICKHOFF (1995): Untersuchungen zur Besiedlungsgeschichte und
gegenwärtigen Waldnutzung im Hindukush/Karakorum.- Erdkunde 49: 49-59.
JAHN, R. & P. SCHÖNFELDER (1995): Exkursionsflora für Kreta.- Stuttgart (Hohenheim). 446.
JARVIS, D. I. (1993): Pollen evidence of changing holocene monsoon climate in Sichuan Province,
China.- Quat. Res. 39: 325-337.
JARVIS, D. I. & S. T. CLAY-POOLE (1992): A comparison of modern pollen rain and vegetation in
southwestern Sichuan Province, China.- Rev. of Palaeobot. a. Palynol. 75: 239-258.
JETTMAR, K. (1977): Bolor - a contribution to the political and ethnic geography of North
Pakistan.- Zentarlasiat. Stud. 11: 412-448.
JETTMAR, K. (1980a): Bolor - zum Stand des Problems.- Zentarlasiat. Stud. 14: 115-132.
JETTMAR, K. (1980b): Neuentdeckte Felsbilder und -inschriften in den Nordgebieten Pakistans.-
Allg. u. Vergl. Archäologie - Beitr. 2: 151-199.
JETTMAR, K. (1984): Tierstil am Indus.- IN: BRENTJES & PEUKE (Hrsg.): Kulturhistorische
Probleme Südasiens und Zentralasiens. Wissenschaft. Beitr. 1984,25: 73-93.
JETTMAR, K. (1993): Voraussetzungen, Verlauf und Erfolg menschlicher Anpassung im
nordwestlichen Himalaya mit Karakorum.- IN: SCHWEINFURTH, U.: Neue Forschungen im
Himalaya.- Erdkundliches Wissen 112: 31-47.
JETTMAR, K. & V. THEWALT (1985): Zwischen Gandhara und den Seidenstraßen. Felsbilder am
Karakorum Highway.- Mainz. 60.
JI JINJUN (1996): Climatic change in arid areas of China and monsoon fluctuations during the past
10 kyears.- Jour. of Arid Environ. 32: 1-7.
JIA LANPO & HUANG WEIWEN (1991): The palaeolithic culture of China.- Quat. Scie. Revi. 10:
519-521.
JIANG DEXIN (1988): Quaternary palynofloras and palaeoclimate of Qaidam Basin, Qinghai,
China.- IN: WHYTE, P. (ed.): The palaeoenvironment of East Asia from the Mid-Tertiary.
Geology, tectonics, orogeny, geomorphology. Centre of Asian Studies Occas. Pap. a.
Monogr. 77,1: 571-578.

10. Literaturverzeichnis 141
JIANG FUCHU & WU XIHAO (1993): Fundamental charcteristics of the stepped landform in China
Continent.- Marine Geol. & Quat. Geol. 13: 15-24.
JOCHIMSEN, M. (1986): Zum Problem des Pollenfluges in den Hochalpen.- Diss. Bot. 90: 249.
JOHNSON, G. (1995): Weltatlas der Alten Kulturen. Indien, Pakistan, Nepal, Bhutan, Bangladesch,
Sri Lanka.- München. 240.
JONES, M. R. & G. C. S. CLARKE (1981): Nymphaeaceae.- IN: PUNT, W. & G. C. S. CLARKE (edts.):
The northwest european pollen flora 3: 57-67.
JONSSON, L. (1973): Pollen morphology in african species of Swertia L. (Gentianaceae).- Grana
13: 119-128.
KACHROO, P., SAPRU, B. L. & U. DHAR (1977): Flora of Ladakh. An ecological and taxonomical
appraisal.- Dehra Dun, India. 172.
KALVODA, J. (1992): Geomorphological record of the quaternary orogeny in the Himalaya and the
Karakoram.- Developm. in Earth Surface Proces. 3: 315.
KAZMI, A. H. (1989): A brief overview of the geology and metallogenic provinces of Pakistan.-
IN: KAZMI, A. H. & L. W. SNEE (eds.): Emeralds of Pakistan: geology, gemology and genesis.
Pakistan. 1-11.
KE MANHONG (1994): A method of pollen-spore analysis in loess.- Acta Bot. Sinica 36: 144-147.
KEMP, R. A., DERBYSHIRE, E., CHEN FAHU & MA HAIZHOU (1996): Pedosedimentary development
and palaeoenvirenmental significance of the S1 palaeosol on the northeastern margin of the
Qinghai-Xizang (Tibetan) Plateau.- Jour. of Quat. Scie. 11: 95-106.
KEZINA, T. V. (1988): Morphology of some Salicaceae pollen grains.- Palynology in the USSR.
Novosibirsk. 61-64.
KINZEL, H. (1982): Pflanzenökologie und Mineralstoffwechsel.- Stuttgart. 534.
KLOTZ, G. (1989): Der Himalaja.- IN: KLOTZ, G. (Hrsg.): Hochgebirge der Erde und ihre Pflanzen-
und Tierwelt. Leipzig, Jena, Berlin. 210-230.
KLÖTZLI, F. A. (1993): Dornpolster und Kissenpolster - zwei divergierende Adaptionen.- IN:
BROMBACHER, C., JACOMET, S. & J. N. HAAS (Hrsg.): Festschrift ZOLLER. Diss. Bot. 196:
155-162.
KÖHLER, E. & E. LANGE (1979): A contribution to distinguishing cereal from wild grass pollen by
LM and SEM.- Grana 18: 133-140.
KOMAREK, J. & B. FOTT (1983): Chlorophyceae (Grünalgen). Ordnung: Chlorococcales.- IN:
HUBER-PESTALOZZI, G. (Hrsg.): Das Phytoplankton des Süßwassers, Systematik und
Biologie. 7.1: Stuttgart. 1044.

10. Literaturverzeichnis 142
KONG ZHAOCHEN & DU NAIQIU (1981): Preliminary study on the vegetation of the Qinghai-Xizang
Plateau during Neogene and Quaternary periods.- LIU DONGSHENG (ch. ed.): Geological and
ecological studies of Qinghai-Xizang Plateau. 1: 239-246.
KONG ZHAOCHEN, DU NAIQIU, SHAN FASHOU, TONG GUOBANG, LUO SHAOJIE & FAN SHUXIAN
(1990): Vegetational and climatic changes in the last 11,000 years in Qinghai Lake -
numerical analysis based on palynology in Core QH85-14C.- Marine Geol. & Quat. Geol. 10:
79-90.
KREMP, G. O. W. & T. KAWASAKI (1972): The spores of Pterodophytes.- Tokyo. 398.
KREUTZMANN, H. (1996): Siedlungsprozesse und territoriale Aneignung im zentralen Hunza-Tal.-
Erdkunde 50: 173-189.
KRÜSSMANN, G. (1983): Handbuch der Nadelgehölze. 2. Aufl.- Berlin, Hamburg. 396.
KUHLE, M. (1987): Subtropical mountain- and highland-glaciation as Ice Age triggers and the
waning of the glacial periods in the Pleistocene.- GeoJour. 14: 393-421.
KUHLE, M. (1993): Eine Autozyklentheorie zur Entstehung und Abfolge der quartären Kalt- und
Warmzeiten auf der Grundlage epirogener und glazialisostatischer Bewegungsinterferenzen
im Bereich des tibetischen Hochlands.- Petermanns Geogr. Mitt. 137: 133-152.
KUKLA, G. & AN ZHISHENG (1989): Loess stratigraphy in Central China.- Palaeogeogr.
Palaeoclimat. Palaeoecol. 72: 203-225.
KVAVADZE, E. V. (1996): Redeposited pollen in recent and holocene sediments of the Caucasus
Mountains.- Grana 35: 33-37.
KWANTEN, L. (1979): Imperial Nomads. A history of Central Asia, 500 - 1500.- University of
Pennsylvania Press. 352.
LAMB, H. H. (1995): Climate, history and the modern world.- London, New York. 2nd. ed. 433.
LANG, G. (1993): Holozäne Veränderungen der Waldgrenze in den Schweizer Alpen -
Methodische Ansätze und gegenwärtiger Kenntnisstand.- IN: BROMBACHER, C., JACOMET, S.
& J. N. HAAS (Hrsg.): Festschrift Zoller. Diss. Bot. 196: 317-327.
LANG, G. (1994): Quartäre Vegetationsgeschichte Europas: Methoden und Ergebnisse.- Jena,
Stuttgart, New York. 462.
LAUER, W. (1993): Klimatologie.- IN: LESER, H. & K. ROTHER (Hrsg.): Das Geographische
Seminar. Braunschweig. 267.
LAUER, W., RAFIQPOOR, M. D. & P. FRANKENBERG (1996): Die Klimate der Erde.- Erdkunde 50:
275-300.
LEEUWEN, P. VAN, PUNT, W. & P. P. HOEN (1988): Polygonaceae.- IN: PUNT, W., BLACKMORE, S.
& G. C. S. CLARKE (edts.): The northwest european pollen flora 5: 81-151.

10. Literaturverzeichnis 143
LEHMKUHL, F. (1995a): Geomorphologische Untersuchungen zum Klima des Holozäns und
Jungpleistozäns Osttibets.- Göttinger Geogr. Abh. 102: 184.
LEHMKUHL, F. (1995b): Aktuelle Inwertsetzung und Gefährdung im Hochgebirge am Ost- und
Nordrand des tibetischen Plateaus.- Regensburger Geogr. Schr. 25: 75-93.
LEHMKUHL, F. (1997a): Late pleistocene, late-glacial and holocene glacier advances on the
Tibetan Plateau.- Quat. Intern. 38/39: 77-83.
LEHMKUHL, F. (1997b): The spatial distribution of loess-like sediments in the mountain areas of
Central and High Asia.- Zeitsch. f. Geomorphol. N. F. Suppl. 111: 97-116.
LEHMKUHL, F. & F. HASELEIN (1999): Pleistocene paleoenvironmental changes on the Tibetan
Plateau and adjacent areas.- Quat. Intern. submitted.
LEHMKUHL, F. & LIU SHIJIAN (1994a): Landscape and quaternary climatic changes in eastern Tibet
and surroundings.- IN: LEHMKUHL, F. & LIU SHIJIAN (eds.): Landscape and quaternary
climatic changes in eastern Tibet and surroundings. GeoJour. 34: 5-6.
LEHMKUHL, F. & LIU SHIJIAN (1994b): An outline of physycal geography including pleistocene
glacial landforms of eastern Tibet (Provinces Sichuan and Qinghai).- IN: LEHMKUHL, F. &
LIU SHIJIAN (eds.): Landscape and quaternary climatic changes in eastern Tibet and
surroundings. GeoJour. 34: 7-30.
LEHMKUHL, F. & T. ROST (1993): Zur pleistozänen Vergletscherung Ostchinas und Nordosttibets.-
Petermanns Geogr. Mitt. 137: 67-78.
LEHMKUHL, F. & J. SPÖNEMANN (1994): Morphogenetic problems of the upper Huang He.-
GeoJour. 31: 31-40.
LEI XIANGYI (1995): Formation age and microfabric characteristics of loess in Chaoxian,
Jingyuan, Gansu, China.- Acta Geogr. Sinica 50: 521-533.
LEMENNICIER, Y., FORT, P. LE, LOMBARDO, B., PECHER, A. & F. ROLFO (1996):
Tectonometamorphic evolution of the central Karakorum (Baltistan, nothern Pakistan).-
Tectonophysics 260: 119-143.
LERCH, G. (1991): Pflanzenökologie.- Berlin. 535.
LI BINGYUAN, YANG YICHOU, ZHANG QINGSONG & WANG FUBAO (1985): On the environmental
evolution of Xizang (Tibet) in Holocene.- IN: LIU TUNGSHENG (ch. ed.): Quaternary Geology
and Environment of China. China Ocean Press Beijing. 234-240.
LI BO, YONG SHIPENG, LIU ZHUNGLING, SUN HONGLIANG & ZENG SIDI (1993): The eco-
geographical rule of stipeta steppe on the Sunglia Plain.- Acta Bot. Sinica 22: 270-279.

10. Literaturverzeichnis 144
LI BOSHENG (1981): A preliminary study on the subnival vegetation in Xizang.- IN: LIU
DONGSHENG (ch. ed.): Geological and ecological studies of Qinghai-Xizang Plateau. Science
Press Beijing. 2: 1987-1990.
LI BOSHENG, ZHANG JINGWEI, WANG JINTING & CHEN WEILIE (1985): The alpine cushion
vegetation of Xizang.- Acta Bot. Sinica 27: 311-317.
LI JIJUN (1991a): The environmental effects of the uplift of the Qinghai-Xizang Plateau.- Quat.
Scie. Revi. 10: 479-483.
LI JIJUN (1991b): The uplift of the Qinghai-Xizang Plateau and its effect on environment.- IN: LIU
TUNGSHENG: Quaternary Geology and Environment in China. Science Press Beijing. 265-
272.
LI JIJUN & XU SHUYING (1984): The distribution of glaciers on the Qinghai-Xizang plateau and its
relationship to atmospheric circulation.- IN: MILLER, K. J. (ed.) The international Karakoram
Project. Cambridge. 1: 84-93.
LI JIJUN, WEN SHIXUAN, ZHANG QINGSONG, WANG FUBAO, ZHENG BENXING & LI BINGYUAN
(1979): A discussion on the period, amplitude and type of the uplift of the Qinghai-Xizang
Plateau.- Scientia Sinica 22: 1314-1328.
LI JIJUN et al. (1995): Uplift of Qinghai-Xizang (Tibet) Plateau and Global Change.- Lanzhou
University Press. 207.
LI MINGSEN (1981): Soil formation and distribution on Qiangtang Plateau.- IN: LIU DONGSHENG
(ch. ed.): Geological and ecological studies of Qinghai-Xizang Plateau. Science Press
Beijing. 2: 1877-1887.
LI PEIQIONG & NI ZHICHENG (1981): The formation and differentiaton of the Leguminosae flora in
Xizang.- IN: LIU DONGSHENG (ch. ed.): Geological and ecological studies of Qinghai-Xizang
Plateau. Science Press Beijing. 2: 1277-1282.
LI PEIYING & ZHOU LIPING (1993): Occurence and palaeoenvironmental implications of the late
pleistocene loess along the eastern coasts of the Bohai Sea, China.- Geol. Soc. (London).
Spec. Publ. 72: 293-309.
LI TIANCHI (1988): A preliminary study on the climatic and environmental changes at the turn
from Pleistocene to Holocene in East Asia.- GeoJour. 17: 649-657.
LI Tianchi (1990): Landslide management in the mountain areas of China.- Internat. Centre f.
Integr. Mount. Develop. Occas. Paper 15: 50.
LI TINGDONG (1996): The process and mechanism of the rise of the Qinghai-Tibet Plateau.-
Tectonophysics 260: 45-53.
LI TINGDONG, XIAO XUCHANG, LI GUANGCEN, GAO YANLIN & ZHOU WEIQIN (1986): The crustal
evolution and uplift mechanism of the Qinghai-Tibet Plateau.- Tectonophysics 127: 279-289.

10. Literaturverzeichnis 145
LI WENHUA (1993): Forests of the Himalayan-Hengduan Mountains of China and strategies for
their sustainable development.- Kathmandu. 175.
LI WENHUA, HAN YUFENG & LI YUJIU (1981): Fundamental characteristics of the forests in Xizang
in relation to the uplift of the Plateau.- IN: LIU DONGSHENG (ch. ed.): Geological and
ecological studies of Qinghai-Xizang Plateau. Science Press Beijing. 2: 1937-1945.
LI WENYI (1987): Vegetation and environment of East China in quaternary cold stage.- Acta
Geogr. Sinica 42: 299-307.
LI WENYI & YAO ZUJU (1990): A study on the quantitative relationship between Pinus pollen in
samples and Pinus vegetation.- Acta Bot. Sinica 32: 943-950.
LI XU & LIU JINLING (1988): Holocene vegetational and environmental changes at Mt. Luoji,
Sichuan.- Acta Geogr. Sinica 43: 44-51.
LI YUANFANG, ZHANG QINGSONG, LI BINGYUAN & F. GASSE (1994): Ostracod fauna and
environmental changes during the past 17000 years in the western Tibet.- Acta Geogr. Sinica
49: 46-54.
LIN ZHENYAO & WU XIANGDING (1981): Climatic classification of Qinghai-Xizang Plateau.- IN:
LIU DONGSHENG (ch. ed.): Geological and ecological studies of Qinghai-Xizang Plateau.
Science Press Beijing. 2: 1575-1580.
LIN ZHENYAO & WU XIANDING (1987): Some features on climatic fluctuation over the Qinghai-
Xizang Plateau.- IN: YE DUZHENG, FU CONGBIN, CHAO JIPING & M. YOSHINO (eds.): The
climate of China and global climate. China Ocean Press Beijing. 116-123.
LISTER, G. S., KELTS, K., CHEN KE ZAO, YU JUNQING & F. NIESSEN (1991): Lake Qinghai, China:
closed-basin lake levels and the oxygen isotope record for ostracoda since the latest
Pleistocene.- Palaeogeogr. Palaeoclimat. Palaeoecol. 84: 141-162.
LIU KAMBIU (1988): Quaternary history of the temperate forests of China.- Quat. Scie. Revi. 7: 1-
20.
LIU JINLING, TANG LINGYU, QIAO YULOU, HEAD, M. J. & D. WALKER (1986): Late Quaternary
vegetation history at Menghai, Yunnan Province, southwest China.- Jour. of Biogeogr. 13:
399-418.
LIU TUNGSHENG & DING ZHONGLI (1993): Stepwise coupling of monsoon circulations to global ice
volume variations during the late Cenocoic.- Global a. Planet. Change 7: 119-130.
LIU TUNGSHENG, ZHANG SHOUXIN & HAN JIAOMAO (1986): Stratigraphy and paleoenvironmental
changes in the loess of Central China.- Quat. Scie. Revi. 5: 489-495.
LIU TUNGSHENG, DING MENGLIN & E. DERBYSHIRE (1996): Gravel deposits on the margins of the
Qinghai-Xizang Plateau and their environmental significance.- Palaeogeogr. Palaeoclimat.
Palaeoecol. 120: 159-170.

10. Literaturverzeichnis 146
LIU YANHUA (1992): Dynamics of highland agriculture in Lhasa, Tibet. A case study.- Internat.
Centre f. Integr. Mount. Develop. Occas. Paper 22: 1-43.
LIU ZECHUN (1990) Study of the geomorphology and stratigraphy of Homo erectus pekinensis site
at Zhoukoudian, Beijing.- IN: SHAKHMATOVA, N. M. (ed.): Chronostratigraphy of the
Paleolithic in North, Central, East Asia and America (Pap. for the Intern. Symp.).
Novosibirsk. 25-34.
LLOYD, P. S. & S. LLOYD (1968): A study of the autecology of Polygonum affine D. DON in the
Karakoram Mountains.- Jour. of Ecol. 56: 723-738.
LU RONGSEN (1992): Seabuckthorn: a multipurpose plant species for fragile mountains.- Internat.
Centre f. Integr. Mount. Develop. Occas. Paper 20: 62.
MÄKELÄ, E. M. (1996): Size distinctions between Betula pollen types - a review.- Grana 35: 248-
256.
MANI, M. S. (1978): Ecology and phytogeography of high altitude plants of the Northwest
Himalaya.- New Delhi, Bombay, Calcutta. 205.
MARKGRAF, V. (1980): Pollen dispersal in a mountain area.- Grana 19: 127-146.
MARTENSEN, H. O. & W. PROBST (1990): Farn- und Samenpflanzen in Europa.- Stuttgart, New
York. 525.
MEDER, O. G. (1979): Klimaökologie und Siedlungsgang auf dem Hochland von Iran in vor- und
frühgeschichtlicher Zeit.- Marburger Geogr. Schr. 80: 221.
MEHER-HOMJI (1973): A phytoclimatic approach to the problem of mediterraneity in the Indo-
Pakistan Sub-continent.- Feddes Repertorium 83: 757-788.
MENG QINGREN, ZHANG GUOWEI, YU ZAIPING & MEI ZHICHAO (1996): Orogenic processes of the
Qinling.- IN: PANG ZHONGHE, ZHANG JINDONG & SUN JIANHONG (eds.): Advances in solid
earth sciences. Science Press Beijing. 8-15.
MEUSEL, H. & R. SCHUBERT (1971a): Beiträge zur Pflanzengeographie des Westhimalajas. 1. Teil:
Die Arealtypen.- Flora 160: 137-194.
MEUSEL, H. & R. SCHUBERT (1971b): Beiträge zur Pflanzengeographie des Westhimalajas. 2. Teil:
Die Waldgesellschaften.- Flora 160: 373-432.
MEUSEL, H. & R. SCHUBERT (1971c): Beiträge zur Pflanzengeographie des Westhimalajas. 3. Teil:
Die pflanzengeographische Stellung und Gliederung des Westhimalayas.- Flora 160: 573-
606.
MEUSEL, H., JÄGER, E., RAUSCHERT, S. & E. WEINERT (1978): Vergleichende Chorologie der
zentraleuropäischen Flora - Karten. Bd. 2. Jena. 421.

10. Literaturverzeichnis 147
MIEHE, G. (1984): Vegetationsgrenzen im extremen und multizonalen Hochgebirge (Zentraler
Himalaya).- Erdkunde 38: 268-277.
MIEHE, G. (1988): Geoecological reconnaissance in the alpine belt of southern Tibet.- GeoJour.
17: 635-638.
MIEHE, G. (1991): Der Himalaya, eine multizonale Gebirgsregion.- IN: WALTER & BRECKLE:
Ökologie der Erde. Band 4. Gemäßigte und arktische Zonen außerhalb Euro-Nordasiens.
Stuttgart. 181-230.
MIEHE, G. (1993): Vegetationskundliche Beiträge zur Klimageographie im Hochgebirge am
Beispiel des Langtang Himal (Nepal).- IN: SCHWEINFURTH, U.: Neue Forschungen im
Himalaya. Erdkundliches Wissen 112: 155-190.
MIEHE, G., MIEHE, S., HUANG JIAN & OTSU TSEWANG (1997): Forschungsdefizite und -
perspektiven zur Frage der potentiellen natürlichen Bewaldung in Tibet.- Petermanns Geogr.
Mitt. (im Druck).
MIEHE, S., CRAMER, T., JACOBSEN, J.-P. & M. WINIGER (1996): Humidity conditions in the
western Karakorum as indicated by climatic data and corresponding distribution patterns of
the montane and alpine vegetation.- Erdkunde 50: 190-204.
MILLER, K. (1982): Continents in collision. The International Karakoram Project.- London. 212.
MODE, M. (1991): Einige Bemerkungen zur sogdischen Handelsroute über das obere Industal.-
Hallesche Beiträge zur Orientwissenschaft. Wissenschaft. Beitr. 16: 45-52.
MOORE, P. D., WEBB, J. A. & M. E. COLLINSON (1991): Pollen analysis.- Oxford. 2nd. ed. 216.
MOU XINDAI, DENG LIYON, GU ANLIN, LIU FENGXIAN, QIAO BINGJUN, BAO CHUNZHI & ZHANG
PUJIN (1992): Rangelands of Xizang (Tibet).- The series of the scientific expedition of the
Qinghai-Xizang Plateau. Science Press House Beijing. Map 1:2,5 Mio. 158.
MÜLLER-STELLRECHT, I. (1979) Materialien zur Ethnographie von Dardistan (Pakistan). Teil I:
Hunza.- IN: JETTMAR, K. (Hrsg.): Bergvölker im Hindukusch und Karakorum. Graz/Austria.
3,1: 383.
MÜLLER-STELLRECHT, I. (1980): Materialien zur Ethnographie von Dardistan (Pakistan). Teil II:
Gilgit und Teil III: Chitral und Yasin.- IN: JETTMAR, K. (Hrsg.): Bergvölker im Hindukusch
und Karakorum. Graz/Austria. 3/2,3: 261.
NAIR, P. K. K: (1965): Pollen grains of western himalayan plants.- Asian Monogr. 5: 102.
NAKAGAWA, G., GARFI, G., REILLE, M. & R. VERLAQUE (1998): Pollen morphology of Zelkova
sicula (Ulmaceae), a recently discovered relic species of the European Tertiary flora: des-
cription, chromosomal relevance, and palaeobotanical significance.- Palaeogeo.
Palaeoclimat. Palaeoeco. 100 1/2 :27-37.

10. Literaturverzeichnis 148
NAKAMURA, J. (1980a): Diagnostic characters of pollen grains of Japan. Part I.- Spec. Publ. from
the Osaka Museum of Natural History 13: 91.
NAKAMURA, J. (1980b): Diagnostic characters of pollen grains of Japan. Part II.- Spec. Publ. from
the Osaka Museum of Natural History 12: 157.
NAYAR, B. K. (1963): Spore morphology of Loxogramme.- Grana 4: 388-392.
NAYAR, B. K. & S. DEVI (1964a): Spore morphology of indian ferns. I. Aspidiaceae.- Grana 5: 80-
120.
NAYAR, B. K. & S. DEVI (1964b): Spore morphology of indian ferns. II. Aspleniaceae and
Blechnaceae.- Grana 5: 222-246.
NAYAR, B. K. & S. DEVI (1964c): Spore morphology of indian ferns. III. Polypodiaceae- Grana 5:
342-395.
NAYAR, B. K. & S. DEVI (1965): Spore morphology of indian ferns. IV. Grammitidaceae.- Grana
6: 121-127.
NAYAR, B. K. & S. DEVI (1966): Spore morphology of the Pteridaceae. I. The pteridoid ferns.-
Grana 6: 476-502.
NAYAR, B. K. & S. DEVI (1967): Spore morphology of the Pteridaceae. II. The gymnogrammoid
ferns.- Grana 7: 468-600.
NAYAR, B. K. & S. DEVI (1968a): Spore morphology of the Pteridaceae. III. The Dicksonioid,
Dennstaedtioid and Lindsayoid ferns.- Grana 8: 185-203.
NAYAR, B. K. & S. DEVI (1968b): Spore morphology of the Pteridaceae. IV. Taxonomic and
phyletic considerations.- Grana 8: 517-535.
NEGI, S. S. (1990): A handbook of the Himalaya.- New Delhi. 350.
NEGI, S. S. (1991): Forest types of India, Nepal and Bhutan.- New Delhi. 322.
NEGI, S. S. (1995): Cold deserts of India.- New Delhi. 248.
NI ZUBIN (1981): Zonation of animal husbandry of Qinghai-Xizang Plateau.- IN: LIU DONGSHENG
(ch. ed.): Geological and ecological studies of Qinghai-Xizang Plateau. Science Press
Beijing. 2: 2091-2102.
NILSSON, S. (1964): On the pollen morphology in Lomatogonium A. BR.- Grana 5: 298-329.
NILSSON, S. (1967): Pollen morphological studies in the Gentianaceae-Gentianinae.- Grana 7: 46-
145.
NILSSON, S. & J. PRAGLOWSKI (1992): Erdtman's Handbook of Palynology.- Copenhagen. 2nd. ed.
580.

10. Literaturverzeichnis 149
NORLINDH, T. (1949): Flora of the mongolian steppe and desert areas.- IN: Reports from the
scientific expedition to the north-western provinces of China under the leadership of Dr.
Sven Hedin. The Sino-Swedish Expedition- Publication 31, XI Botany 4. Stockholm. 155.
NOWICKE, J. W. & J. J. SKVARLA (1977): Pollen morphology and the relationship of the
Plumbaginaceae, Polygonaceae and Primulaceae to the Order Centrospermae.- Smithsonian
Contrib. to Bot. 73: 64.
OBERDORFER, E. (1990): Pflanzensoziologische Exkursionsflora. 6. Aufl.- Stuttgart. 1050.
OKA, H. I. (1988): Origin of cultivated rice.- Develop. in Crop Scie. 14: 254.
OSMASTON, H. (1989): Problems of the quaternary geomorphology of the Xixabangma region in
South Tibet and Nepal.- Zeitsch. f. Geomorphol. N. F. Suppl. 76: 147-180.
OSTEN, H. H. v.d. (1956): Die Welt der Perser.- IN: BOSSERT, T. (Hrsg.): Große Kulturen der
Frühzeit. Stuttgart. 299.
OWEN, L. K. (1989): Terraces, uplift and climate in the Karakorum Mountains, nothern Pakistan:
Karakoram intermontane basin evolution.- Zeitsch. f. Geomorphol. N. F. Suppl. 76: 117-146.
OWEN, L. A. & E. DERBYSHIRE (1989): The Karakorum glacial depositional system.- Zeitsch. f.
Geomorphol. N. F. Suppl. 76: 33-73.
PACHUR, H.-J., WÜNNEMANN B., ZHANG LINYUAN, ZHANG HUCAI & MA YUZHENG (1994):
Floodplain sediments on the divide between Huang He and Chang Jiang, Gansu Province,
NW China.- IN: LEHMKUHL, F. & LIU SHIJIAN (eds.): Landscape and quaternary climatic
changes in eastern Tibet and surroundings. GeoJour. 34: 97-106.
PACHUR, H.-J., WÜNNEMANN, B. & HUCAI ZHANG (1995): Lake evolution in the Tengger Desert,
northwestern China, during the last 40,000 years.- Quat. Res. 44: 171-180.
PAFFEN, K., PILLEWITZER, W. & H.-J. SCHNEIDER (1956): Forschungen im Hunza-Karakorum.-
Erdkunde 10: 1-33.
PANT, G. B. & K. RUPA KUMAR (1997): Climates of South Asia.- Belhaven Studies in
Climatology. Chichester, New York, Weinheim. 320.
PETZSCH, H. & R. PIECHOCKI (1986): Säugetiere.- IN: Urania Tierreich. Leipzig. 604.
PEWE, T. L., LIU TUNGSHENG, SLATT, R. M. & LI BINGYUAN (1995): Origin and character of
loesslike silt in the southern Qinghai-Xizang (Tibet) Plateau, China.- U.S. Geol. Survey
Profess. Pap. 1549: 55.
PLAETSCHKE, B. (1939): Landschaftskundliche Wesenszüge der östlichen Gobi.- Wissenschaft.
Veröffent. d. Dt. Museums f. Länderkunde zu Leipzig. N. F. 7: 103-148.
POHLE, P. (1997): Felsbilder in Zentralasien.- Geogr. Rundsch. 49: 287-292.

10. Literaturverzeichnis 150
POLUNIN, O. & A. STAINTON (1990): Concise flowers of the Himalaya.- Delhi, Bombay, Calcutta,
Madras. 283.
PORTER, C. S., AN ZHISHENG & ZHENG HONGBO (1992): Cyclic quaternary alluviation and
terracing in a nonglaciated drainage basin on the north flank of the Qinling Shan, Central
China.- Quat. Res. 38: 157-169.
PÖRTGE, K.-H. (1994): Hydrologische Untersuchungen an Flüssen und Seen im tibetischen
Plateau.- IN: PÖRTGE, K.-H. & LI JIAN (Hrsg.): Beiträge zur Tibetforschung. Göttinger Geogr.
Abh. 95: 273-282.
PRACH, K. (1994): Vegetation succession on river gravel bars across the Northwestern Himalayas,
India.- Arctic a. Alpine Res. 26: 349-353.
PRAGLOWSKI, J. (1970): The pollen morphology of the Haloragaceae with references to
taxonomy.- Grana 10: 159-239.
PRAGLOWSKI, J. (1982): Fagaceae L., Fagoideae.- World Pollen a. Spore Flora 11: 28.
PRAGLOWSKI, J. (1987): Pollen morphology of Tribulaceae.- Grana 26: 193-211.
PRELL, W. L. & J. E. KUTZBACH (1992): Sensitivity of the Indian Monsoon to forcing parameters
and implications for its evolution. Nature 360: 647-652.
PUNT, W. (1976): Sparganiaceae and Typhaceae.- IN: PUNT, W., JANSSEN, C. R., REITSMA, T. & G.
C. S. CLARKE (eds.): The northwest european pollen flora 1: 75-88.
PUNT, W. (1984): Umbelliferae.- IN: PUNT, W. & G. C. S. CLARKE (edts.): The northwest european
pollen flora 4: 155-369.
PUNT, W. & S. BLACKMORE (Edts.) (1991): The northwest european pollen flora, VI. Parts 44-51.-
Reprint from Review of Palaeobotany and Palynologie 69. Amsterdam. 275.
PUNT, W. & P. DEN BREEJEN (1981): Linaceae.- IN: PUNT, W. & G. C. S. CLARKE (edts.): The
northwest european pollen flora 3: 75-115.
PUNT, W. & G. C. S. CLARKE (edts.) (1980): The northwest european pollen flora, II. Parts 8-20.-
Reprint from Review of Palaeobotany and Palynologie 23, 24, 26, 28. Amsterdam. 265.
PUNT, W. & G. C. S. CLARKE (edts.) (1981): The northwest european pollen flora, III. Parts 21-
28.- Reprint from Review of Palaeobotany and Palynologie 33. Amsterdam. 138.
PUNT, W. & G. C. S. CLARKE (edts.) (1984): The northwest european pollen flora, IV. Parts 29-
37.- Reprint from Review of Palaeobotany and Palynologie 42. Amsterdam. 369.
PUNT, W. & M. MALOTAUX (1984): Cannabaceae, Moraceae and Urticaceae.- IN: PUNT, W. &
G. C. S. CLARKE (edts.): The northwest european pollen flora 4: 23-44.

10. Literaturverzeichnis 151
PUNT, W., JANSSEN, C. R., REITSMA, T. & G. C. S. CLARKE (edts.) (1976): The northwest european
pollen flora, I. Parts 1-7.- Reprint from Review of Palaeobotany and Palynologie 17, 19 and
21. Amsterdam. 145.
PUNT, W., BLACKMORE, S. & G. C. S. CLARKE (edts.) (1988): The northwest european pollen flora,
V. Parts 38-43.- Reprint from Review of Palaeobotany and Palynologie, 57. Amsterdam. 154.
PUNT, W., BOS, A. A. & P. P. HOEN (1991): Oleaceae.- IN: PUNT, W. & S. BLACKMORE (edts.): The
northwest european pollen flora 6: 23-47.
PUNT, W., BLACKMORE, S., NILSSON, S. & A. LE THOMAS (1994): Glossary of pollen and spore
terminology.- Laborat. of Palaeob. & Palynol. Contr. Series. 1: 71.
PURI, B. N. (1994): The Kushans.- IN: HARMATTA, J., PURI, B. N. & G. F. ETMADI (eds.) (1994):
The development of sedentary and nomadic civilizations: 700 B. C. to A. D. 250. History of
civilizations of Central Asia. UNESCO Publ. 2: 246-263.
QI GUOQIN (1989): Pleistocene human environment in North China.- The Quat. Res. 28: 327-336.
QI HUANG, CAI BIQIN & YU JUNQING (1981): The C14 age and cycle of sedimentation of some
saline lakes on the Qinghai-Xizang Plateau.- Kexue Tongbao 26: 66-70.
QIN ZHIYE & XIE WENZHONG (1980): Obsevations on the morphology and ecology of the cushion
plants in Togme Region of northern Xizang.- Acta Bot. Sinica 22: 177-181.
RAM, J. (1992): Effects of clipping on aboveground plant biomass and total herbage yields in a
grassland above treeline in Central Himalaya, India.- Arctic a. Alpine Res. 24: 78-81.
RAO, A. R. & P. SHUKLA (1975): Pollen flora of Upper Gangetic Plain.- Indian Pollen-Spore
Floras 1: 130.
RAUNIG, W. (1978): Zur materiellen Kultur der Bewohner des Wakhan.- IN: GRANCY, R. S. DE &
R. KOSTKA (Hrsg.): Großer Pamir. Graz/Austria. 263-271.
RAUNIG, W. (1983): Spuren alter Besiedlung auf den Talterrassen des östlichen Wakhan
(Nordostafghanistan).- IN: SNOY, P. (Hrsg): Ethnologie und Geschichte. Festschrift für Karl
Jettmar. Beitr. z. Südasienforsch. 86: 496-508.
RAZA, H. A. & F. K. BENDER (1995): General introduction.- BENDER, F. K. & H. A. RAZA (eds.):
Geology of Pakistan. Beitr. z. Regio. Geol. d. Erde 25: 1-10.
REILLE, M. (1992): Pollen et Spores D'Europe et D'Afrique du nord.- Laboratoire de Botanique
historique et Palynologie. Marseille. 520.
REILLE, M. (1995): Pollen et Spores D'Europe et D'Afrique du nord.- Supplement 1. Laboratoire
de Botanique historique et Palynologie. Marseille. 327.
REIMERS, F. (1992): Untersuchungen zur Variabilität der Niederschläge in den Hochgebirgen
Nordpakistans und angrenzender Gebiete.- Beitr. u. Material. z. Region. Geogr. 6: 219.

10. Literaturverzeichnis 152
REIMERS, F. (1994): Die Niederschlagssituation in den Hochgebirgen Nordpakistans während der
Flutkatastrophe vom September 1993.- Beitr. u. Material. z. Region. Geogr. 7: 1-19.
REITSMA, T. (1966): Pollen morphology of some european Rosaceae.- Acta Bot. Neerlandica 15:
290-307.
RHODES, T. E., GASSE, F., LIN RUIFEN, FONTES, J.-C., WEI KEQIN, BERTRAND, P., GIBERT, E.,
MÉLIÈRES, F., TUCHOLKA, P., WANG ZHIXIANG & CHENG ZHIYUAN (1996): A late
pleistocene-holocene lacustrine record from Lake Manas, Zunggar (nothern Xinjiang,
western China).- Palaeogeogr. Palaeoclimat. Palaeoecol. 120: 105-121.
RIKHARI, H. C., NEGI, G. C. S., PANT, G. B., RANA, B. S. & S. P. SINGH (1992): Phytomass and
primary productivity in several communities of a Central Himalaya alpine meadow, India.-
Arctic a. Alpine Res. 24: 344-351.
RIKHARI, H. C., NEGI G. C. S., RAM, J. & S. P. SINGH (1993): Human-induced secondary
succession in an alpine meadow of Central Himalaya, India.- Arctic a. Alpine Res. 25 8-14.
ROLPH, T. C., SHAW JOHN, DEBRYSHIRE E. & WANG JINGTAI (1993): The magnetic mineralogy of
a loess section near Lanzhou, China.- Geol. Soc. (London). Spec. Publ. 72: 311-323.
ROST, K. T. (1992): Geomorphologische Höhenstufen im Qinling Shan (VR China) unter
besonderer Berücksichtigung der jungpleistozänen Vergletscherungen.- Göttinger Geogr.
Abh. 97: 89.
ROST, K. T. (1994): Paleoclimatic field studies in and along the Qinling Shan (Central China).-
IN: LEHMKUHL, F. & LIU SHIJIAN (eds.): Landscape and quaternary climatic changes in
eastern Tibet and surroundings. GeoJour. 34: 107-120.
ROST, K. T. (1997): Observations on distribution and age of loess-like sediments in the high-
mountain ranges of Central China.- Suppl. 111: 117-129.
ROST, K. T. (1998): Geomorphologische und paläoklimatsiche Untersuchungen in
zentralchinesischen Gebirgen und Gebirgsvorländern.- Göttinger Geogr. Abh. 105:1-168.
ROST, K T., BÖHNER, J. & MA ZHIMIN (1996): Untersuchungen zum klimatischen, geomorpho-
logischen und geoökologischen Stockwerkbau des zentralen Qinling Shan (VR China,
Provinz Shaanxi).- Petermanns Geogr. Mitt. 140: 177-191.
RÖTHLISBERGER, F. (1986): 10000 Jahre Gletschergeschichte der Erde. Aarau, Frankfurt am Main,
Salzburg. 416.
ROUSI, A. (1971): The genus Hippophaë L. A taxonomic study.- Annales Bot. Fennici 8: 117-227.
ROWLEY, J. B. (1960): The exine structure of "cereal" and "wild" type grass pollen.- Grana 2: 9-
15.

10. Literaturverzeichnis 153
SARASWAT, K. S. (1991): Archaeobotanical remains in ancient cultural and socioeconomical
dynamics of the Indian Subcontinent.- The Palaeobotanist 40: 514-545.
SCHAARSCHMIDT, H. (1988): Die Walnussgewächse.- Neue Brehm-Bücherei 591: 116.
SCHICKHOFF, U. (1993): Das Kaghan-Tal im Westhimalaya (Pakistan).- Bonner Geogr. Abh. 87:
268.
SCHICKHOFF, U. (1995): Verbreitung, Nutzung und Zerstörung der Höhenwälder im Karakorum
und in den angrenzenden Hochgebirgsräumen Nordpakistans.- Petermanns Geogr. Mitt. 139:
67-85.
SCHLÜTZ, F. (1995): Zur holozänen Vegetationsgeschichte im Gebiet des Nianbaoyeze an der
Grenze der Provinzen Qinghai und Sichuan.- Göttinger Geogr. Abh. 102: 185-200.
SCHLÜTZ, F. (1997): Beiträge zur Vegetations- und Siedlungsgeschichte im Wesertal bei Höxter-
Corvey.- Ausgrab. u. Funde i. Westf.-Lippe 9/A: 55-72.
SCHLÜTZ, F. (1998): Vegetationsgeschichtliche Untersuchungen in Hochasien. 2. Zum Holozän
des Karakorum und zum letzten Glazial im Qinling Shan.- Petermanns Geogr. Mitt. 142:
149-154.
SCHMIDT-VOGT, H. (1987): Die Fichte, Band 1: Taxonomie, Verbreitung, Morphologie, Ökologie,
Waldgesellschaften.- Hamburg, Berlin. 2. Aufl. 647.
SCHOLZ, F. (1995): Nomadismus. Theorie und Wandel einer sozio-ökologischen Kulturweise.-
Erdkundliches Wissen 118: 300.
SCHUBERT, R., HANDKE, H. & H. PANKOW (1983): Niedere Pflanzen - Grundband.- IN:
ROTHMALER, W.: Exkursionsflora für die Gebiete der DDR und BRD. 1: 811.
SCHÜTT, P., SCHUCK, H. J. & B. STIMM (1992): Lexikon der Forstbotanik.- Landsberg/Lech. 581.
SCHWEINFURTH, U. (1956): Über klimatische Trockentäler im Himalaya.- Erdkunde 10: 297-302.
SCHWEINFURTH, U. (1957): Die horizontale und vertikale Verbreitung der Vegetation im
Himalaya.- Bonner Geogr. Abh. 20: 370.
SCHWEINFURTH, U. (1982): Der innere Himalaya.- IN: MEYEN, E. & E. PLEWE: Beiträge zur
Hochgebirgsforschung und zur Allgemeinen Geographie. Erdkundliches Wissen 59: 15-24.
SCHWEINFURTH, U. (1983): Mensch und Umwelt im Indus-Durchbruch am Nanga Parbat (NW-
Himalaya).- IN: SNOY, P. (Hrsg.) Beitr. z. Südasienforsch. 86: 536-559.
SCHWEINFURTH, U. (1986): Zur Landschaftsgliederung im Chinesisch-Tibetischen
Übergangsraum.- Berliner Geogr. Studien 20: 237-249.
SCHWEINFURTH, U. (1993): Vegetation und Himalaya-Forschung.- IN: SCHWEINFURTH, U.:Neue
Forschungen im Himalaya.- Erdkundliches Wissen 112: 11-29.

10. Literaturverzeichnis 154
SCHWEINFURTH, U. & CHEN WEILIE (1984): Vegetation und Landesnatur im südlichen Gaoligong
Shan (West Yünnan).- Erdkunde 38: 278-288.
SEARLE, M. P. (1991): Geology and tectonics of the Karakoram Mountains.- Cichester, New York.
358.
SEKINA, M. A. & P. D. MOORE (1995): Morphological studies of the pollen grains of the semi-arid
region of Egypt.- Flora 190: 115-133.
SENGÖR, A. M. C. (1986): The dual nature of the alpine-himalayan system: progress, problems and
prospects.- Tectonophysics 127: 177-195.
SHAMS, F. A. (1995): The Tethyan Belt.- IN: BENDER, F. K. & H. A. RAZA (eds.): Geology of
Pakistan. Beitr. z. Regio. Geol. d. Erde 25: 31-62.
SHAN RENHUA & SHE MENGLAN (1981): Xizang Umbellifers: their geographical patterns and
floristic characteristics.- IN: LIU DONGSHENG (ch. ed.): Geological and ecological studies of
Qinghai-Xizang Plateau. Science Press Beijing. 2: 1283-1287.
SHARMA, C. (1991): Palaeoclimatic oscillations since last deglaciation in western Himalaya: a
palynological assay.- The Palaeobotanist 40: 374-382.
SHARMA, C. & M. S. CHAUHAN (1988): Palaeoenvironmental interferences from the quaternary
palynostratigraphy of Himachal Pradesh and Kumaon, India.- IN: JAIN, D. V. S., DHARA, M.
& J. SAKETHARAMAN (eds.): Palaeoclimatic and palaeoenvironmental changes in Asia during
the last 4 million years. New Delhi. 178-191.
SHEN CAIMING & TANG LINGYU (1996): A preliminary study of vegetation history since Pliocene
in Xizang.- IN: PANG ZHONGHE, ZHANG JINDONG & SUN JIANHONG (eds.): Advances in solid
earth sciences. Science Press Beijing. 74-79.
SHEN XIANJIE (1996): Crust-mantle thermal structure and tectonothermal evolution of the Tibetan
Plateau.- A Series of Solid Earth Scie. Res. in China. 224.
SHEN ZHENSHU, WANG QIANG, CHENG GUO & QI GUOZHU (1992): The lower quarternary boundary
in the Chaidamu Basin of Qinghai Province.- Marine Geol. & Quat. Geol. 12: 97-107.
SHI YAFENG & ZHANG XIANGSON (1995): Impact of climate change on surface water resource and
tendency in the future in the arid zone of northwestern China.- Science in China (B) 38: 11,
1395-1408.
SHI YAFENG, REN BINGHUI, WANG JINGTAI & E. DERBYSHIRE (1986): Quaternary glaciation in
china.- Quat. Scie. Revi. 5: 503-507.
SHI YAFENG, KONG ZHAOZHENG, WANG SUMIN, TANG LINGYU, WANG FUBAO, YAO TANDONG,
ZHAO XITAO, ZHANG PEIYUAN & SHI SHAOHUA (1993): Mid-holocene climates and
environments in China.- Global a. Planet. Change 7: 219-233.

10. Literaturverzeichnis 155
SHI ZHU & CHEN YILIN (1981): The Compositae of Xizang.- IN: LIU DONGSHENG (ch. ed.):
Geological and ecological studies of Qinghai-Xizang Plateau. Science Press Beijing. 2:
1313-1319.
SHILOVA, G. N. (1984): To the method of the interpretation of spore-pollen spectra in mountain
districts of the Mongolia.- Problems of modern Palynology. Novosibirsk. 61-65.
SINGH, G. & R. D. JOSHI (1969): Pollen morphology of some eurasian species of Artemisia.- Grana
9: 50-62.
SINGH, G., WASSON R. J. & D. P. AGRAWAL (1990): Vegetational and seasonal climatic changes
since the last full glacial in the Thar Desert, northwestern India.- Rev. of Palaeobot. a.
Palynol. 64: 351-358.
SINHA, A. K. & R. UPADHYAY (1995): Himalya: geological aspects.- The Palaeobotanist 44: 9-27.
SNOY, P. (1975): Bagrot. Eine Dardische Talschaft im Kararkorum.- IN: JETTMAR, K. (Hrsg.):
Bergvölker im Hindukusch und Karakorum. Graz/Austria. 2: 235.
SONG ZHICHEN & LIU GENGWU (1981): Tertiary palynological assemblages from Xizang with
references to their paleogeographical significance.- IN: LIU DONGSHENG (ch. ed.): Geological
and ecological studies of Qinghai-Xizang Plateau. Science Press Beijing. 1: 207-214.
SORSA, P. (1971): Pollen morphological study of the genus Hippophaë L., including the new taxa
recognized by A. ROUSI.- Annales Bot. Fennici 8: 228-236.
STAFFORD, P. J. & S. BLACKMORE (1991): Geraniaceae.- IN: PUNT, W. & S. BLACKMORE (edts.):
The northwest european pollen flora 6: 49-78.
STAHLBERG, S. (1996): Der Gansu-Korridor: Barbarenland diesseits und jenseits der Großen
Chinesischen Mauer.- Hamburg. 243.
STEFFAN, H. & L. DUPONT (1988): Note on a program for plotting of pollen diagrams with an
Atari ST microcomputer.- Pollen et Spores 30: 125-130.
STOCKMARR, J. (1971): Tablets with spores used in absolute pollen analysis.- Pollen et Spores 13:
615-621.
STONE, D. E. & C. R. BROOME (1975): Juglandaceae A. RICH ex KUNTH.- World Pollen a. Spore
Flora 4: 35.
STORRS, A. & J. STORRS (1990): Trees and shrubs of Nepal and the Himalaya.- Bangkok. 367.
SUCCOW, M. (1989): Die Mittelasiatischen Hochgebirge. Der Altai.- IN: KLOTZ, G. (Hrsg.):
Hochgebirge der Erde und ihre Pflanzen- und Tierwelt. Leipzig, Jena, Berlin. 187-209.
SUN DONGHUAI, JIANG FUCHU & S. C. PORTER (1996): Preliminary study on the summer monsoon
climate of Chinese Loess Plateau during the last interglacial period.- Chinese Scie. Bull. 41:
321-325.

10. Literaturverzeichnis 156
SUN JIANZHONG, ZHAO JINGBO et al. (eds.) (1991): Quaternary of Loess Plateau in China.- Science
Press Beijing (chin.).
SUN XIANGJUN & CHEN YINSHUO (1991a): Palynological records of the last 11,000 years in
China.- Quat. Scie. Revi. 10: 537-544.
SUN XIANGJUN & CHEN YINSHUO (1991b): Holocene palynological research in China.- IN: LIU
TUNGSHENG (ch. ed.): Quaternary Geology and Environment in China. Science Press Beijing.
214-227.
SUN XIANGJUN & WU YUSHU (1988): The distribution of pollen and algae in surface sediments of
Dianchi, Yunnan Province, China.- Rev. of Palaeobot. a. Palynol. 55: 193-206.
SUN XIANGJUN, DU NAIQIU & CHEN MINGHONG (1981): The paleovegetation and paleoclimate
during time of Homudu people.- Acta Bot. Sinica 23: 145-151.
TAFEL, A. (1914): Meine Tibetreise. Eine Studienfahrt durch das nordwestliche China und durch
die innere Mongolei in das östliche Tibet.- Stuttgart, Berlin, Leipzig. 1 & 2: 698.
TAHIRKHELI, R. A. K. & Q. M. JAN (1984): The geographical and geological domains of the
Karakoram.- IN: MILLER, K. J. (ed.): The international Karakoram Project.- Cambridge. 2:
57-70.
TAKAMIYA, T. (1978) Beiträge zur Geschichte der Nahrung und der Nahrungsbereitung bei den
Hirtenvölkern Mittel- und Innerasiens.- Diss. Ludwig-Maximilians-Univ. München. 376.
TANG BANGXING, LIU SUQING & LIU SHIJIAN (1994): Mountain disaster formation in northwest
Sichuan.- IN: LEHMKUHL, F. & LIU SHIJIAN (eds.): Landscape and quaternary climatic
changes in eastern Tibet and surroundings. GeoJour. 34: 41-46.
TAO JUNRONG (1981): Succession of the floras in Xizang during Upper Cretaceous-Paleogene and
Neogene.- Acta Bot. Sinica 23: 140-145.
TAO JUNRONG (1988): The paleocene flora and palaeoclimate of Liuqu Formation in Xizang
(Tibet).- IN: WHYTE, P. (ed.): The palaeoenvironment of East Asia from the Mid-Tertiary.
Geology, tectonics, orogeny, geomorphology. Centre of Asian Studies Occas. Pap. a.
Monogr. 77,1: 520-522.
TENG JIWEN & LIN BANZUO (1984): Earthquake activity and tectonics of the Himalaya and its
surrounding regions.- IN: MILLER, K. J. (ed.) The international Karakoram Project.
Cambridge. 1: 221-235.
THELAUS, M. (1992): Some characteristics of the mire development in Hongyuan County, eastern
Tibetan Plateau.- Proc. of the 9th Intern. Peat Congr. 1992. 1: 334-351.
THOMAS, A. (1994): Raum-zeitliche Variation der Temperaturgradienten und thermischen
Raumgliederung in der Provinz Yunnan, VR China.- IN: DOMRÖS, M. & W. KLAER (Hrsg.):
Festschrift für Erdtmann Gormsen zum 65. Geburtstag. Mainzer Geogr. Schr. 40: 625-642.

10. Literaturverzeichnis 157
TOMB, A. S. (1975): Pollen Morphology in Tribe Lactuceae (Compositae).- Grana 15: 79-89.
TONG GUOBANG & ZHANG JUNPAI (1991): Quaternary palynoflora of China.- IN: ZHANG ZONGHU
& SHAO SHIXIONG (eds.): The Quaternary of China. China Ocean Press. 375-391.
TONG GUOBANG, ZHANG JUNPAI & FAN SHUXIAN (1992): Distribution of quaternary palynoflora in
China.- Marine Geol. & Quat. Geol. 12: 45-56.
TRETER, U. (1996): Gebirgs-Waldsteppe in der Mongolei.- Geogr. Rundschau 11: 655-661.
TRINKLER, E. (1932): Geographische Forschungen im westlichen Zentralasien und Karakorum-
Himalaya.- IN: TRINKLER, E. & H. DE TERRA (Bearb.): Wissenschaftliche Ergebnisse der Dr.
TRINKLERschen Zentralasien-Expedition. Berlin. 1: 133.
TROLL, C. (1939): Das Pflanzenkleid des Nanga Parbat.- Wissenschaft. Veröffent. d. Dt. Museums
f. Länderkunde zu Leipzig. N. F. 7: 149-193.
TROLL, C. (1967): Die klimatische und vegetationsgeographische Gliederung des Himalaya-
Systems.- Ergebn. Forsch. - Unternehmen Nepal Himalaya 1: 353-388.
TROLL, C. (1978): Klima und Pflanzenkleid der Erde in dreidimensionaler Sicht.- Wege d.
Forschung 130: 355-400.
TUBEUF, K. FREIHERR VON (1923): Monographie der Mistel.- München, Berlin. 832.
TURNER, S. & S. BLACKMORE (1984): Plumbaginaceae.- IN: PUNT, W. & G. C. S. CLARKE (edts.):
The northwest european pollen flora 4: 133-154.
UBERA, J. L., GALAN, C. & F. H. GUERRERO (1988): Palynological study of the genus Plantago in
the Iberian Peninsula.- Grana 27: 1-15.
VAN CAMPO. E. & F. GASSE (1993): Pollen- and diatom-inferred climatic and hydrological
changes in Sumxi Co Basin (Western Tibet) since 13,000 yr B.P.- Quat. Res. 39: 300-313.
VAN CAMPO, E., COUR, P. & HANG SIXUAN (1996): Holocene environmental changes in Bangong
Co Basin (Western Tibet). Part 2: The pollen record.- Palaeogeogr. Palaeoclimat. Palaeoecol.
120: 49-63.
VAN GEEL, B., BUURMANN, J. & H. T. WATERBOLK (1996): Archaeological and palaeoecological
indications of an abrupt climate change in The Netherlands, and evidence for climatological
teleconnections around 2650 BP.- Jour. of Quat. Scie. 11: 451-460.
VERBEEK-REUVERS, A. A. M. L. (1980): Saxifragaceae.- IN: PUNT, W. & G. C. S. CLARKE (edts.):
The northwest european pollen flora 2: 31-58.
VISHNU-MITTRE (1979): Palaebotanical evidences of the environment of early man in
northwestern and western India.- Grana 18: 167-181.
VISHNU-MITTRE (1981): Possible significance of pre-neolithic cereal type in South Asia.- Proc.
IV. Int. Palynol. Conf., Lucknow (1976-77). 3: 291-294.

10. Literaturverzeichnis 158
VISHNU-MITTRE & B. D. SHARMA (1963): Pollen morphology of the indian species of Alnus.-
Grana 4: 302-305.
WAGENITZ, G. (1956): Pollenmorphologie der mitteleuropäischen Valerianaceen.- Flora 143: 473-
485.
WAGNER, D. B. (1993): Iron and steel in ancient China.- IN: B. SPULER (Hrsg.): Handbuch der
Orientalistik. Leiden. 4 Abt. 9: 573.
WALTER, H. (1968): Die Vegetation der Erde in öko-physiologischer Betrachtung. Bd. II: Die
gemäßigten und arktischen Zonen.- Stuttgart. 1001.
WALTER, H. (1974): Die Vegetation Osteuropas, Nord- und Zentralasiens.- Stuttgart. 452.
WALTER, H. & H. STRAKA (1970): Arealkunde: Floristisch-historische Geobotanik.- IN: WALTER,
H. (Hrsg.): Einführung in die Phytologie. 3,2: 478.
WANG CHIWU (1961): The forests of China with a survey of grassland and desert vegetation.-
Maria Moors Cabot Foundation Publ. 5: 313.
WANG FUBAO & FAN C. Y. (1987): Climatic changes in the Qinghai-Xizang (Tibetan) Region of
China during the Holocene.- Quat. Res. 28: 50-60.
WANG FUHSIUNG, CHIEN NANFEN, ZHANG YULONG & YANG HUIQIU (1995): Pollen flora of China
(chin.). 2nd. ed. 461.
WANG HESHENG (1981): A study on the distribution and origin of endemic genera of
Spermatophytes in China.- IN: LIU DONGSHENG (ch. ed.): Geological and ecological studies
of Qinghai-Xizang Plateau. Science Press Beijing. 1: 605-620.
WANG JINGQUAN (1985): Studies on the pollen morphology of family Salicaceae in Gansu.- Acta
Bot. Sinica 27: 594-598.
WANG JINTING (1981): On the fundamental characteristics of the steppe vegetation in Xizang
Plateau.- IN: LIU DONGSHENG (ch. ed.): Geological and ecological studies of Qinghai-Xizang
Plateau. Science Press Beijing. 2: 1929-1936.
WANG LIANGJIAN, LI XIANMING & LIN ZHIYUAN (1995): A discussion on the soil below dark
coniferous forests of the high mountains in Southwest China.- Acta Geogr. Sinica 50: 542-
551.
WANG MANHUA (1987): The spore-pollen groups of peatland on Ruoergai Plateau and
paleobotany and paleoclimate.- Acta Geogr. Sinica 7: 147-155.
WANG PINXIAN & SUN XIANGJUN (1994): Last glazial maximum in China: comparison between
land and sea.- Catena 23: 341-353.
WANG SHUJI & SUN YAN (1987): Cenozoic evolution of geographical environment in north and
south sides of the Tianshan Mountains.- Acta Geogr. Sinica 42: 211-220.

10. Literaturverzeichnis 159
WANG XIANZENG (1988): The miocene palynofloras of China and their palaeoenvironmental
significance.- IN: WHYTE, P. (ed.): The palaeoenvironment of East Asia from the Mid-
Tertiary. Geology, tectonics, orogeny, geomorphology. Centre of Asian Studies Occas. Pap.
a. Monogr. 77,1: 564-570.
WANG ZONGTAI & YANG HUIAN (1992): Characteristics of the distribution of glaciers in China.-
Annals of Glaciology 16: 17-20.
WARDLE, P. (1981): The upper limits of tree growth in oceanic and continental environments.- IN:
LIU DONGSHENG (ch. ed.): Geological and ecological studies of Qinghai-Xizang Plateau.
Science Press Beijing. 2: 1953-1961.
WEIERS, S. (1995): Zur Klimatologie des NW-Karakorum und angrenzender Gebiete.- Bonner
Geogr. Abh. 92: 168.
WENG CHENGYU, SUN XIANGJUN & CHEN YINSHUO (1993): Numerical characteristics of pollen
assemblages of surface samples from the West Kunlun Mountains. Acta Bot. Sinica 35: 69-
79.
WHITEHOUSE, D. & R. WHITEHOUSE (1975): Archaeological atlas of the world.- London. 272.
WHITTINGTON, A. G. (1996): Exhumation overrated at Nanga Parbat, nothern Pakistan.-
Tectonophysics 260: 215-226.
WINKLER, D. (1994): Die Waldvegetation in der Ostabdachung des Tibetischen Hochlandes.-
Berliner Geogr. Abh. Beih. 2: 129.
WINKLER, D. (1996): Forests, forest economy and deforestation in the Tibetan Prefectures of West
Sichuan.- Commonwealth Forestry Rev. 75: 296-301.
WINKLER, M. G. & WANG PAO K. (1993): The late-quaternary vegetation and climate of China.-
IN: WRIGHT, H. E, KUTZBACH, J. E., WEBB, T., RUDDIMAN, W. F., STREET-PERROTT, F. A. &
P. J. BARTLEIN (eds.): Global climates since the Last Glacial Maximum.- Minneapolis,
London. 221-264.
WISSMANN, H. VON (1960): Stufen und Gürtel der Vegetation und des Klimas in Hochasien und
seinen Randgebieten. A: Hygrische Raumgliederung und Exposition.- Erdkunde 14: 249-272.
WISSMANN, H. VON (1961): Stufen und Gürtel der Vegetation und des Klimas in Hochasien und
seinen Randgebieten. B: Thermische Raumgliederung (1. Teil) und Frostboden.- Erdkunde
15: 19-44.
WU SHUHUI (1995): Die Eroberung von Qinghai unter Berücksichtigung von Tibet und Khams
1717 - 1727 anhand der Throneingaben des Großfeldherrn Nian Gengyao.- Tunguso Sibirica
2: 365.
WU SHUHUI (1996): How the Qing army entered Tibet after the tibetan civil war.-
Zentarlasiatische Studien 26: 122-138.

10. Literaturverzeichnis 160
WU WEITANG (1985): Origin and spread of paddy rice cultivation in China.- Acta Geogr. Sinica
40: 29-36.
WU XIANGDING & LIN ZHENYAO (1981): A preliminary analysis of climatic change during the
historical time of Qinghai-Xizang Plateau.- IN: LIU DONGSHENG (ch. ed.): Geological and
ecological studies of Qinghai-Xizang Plateau. Science Press Beijing. 2: 1581-1587.
WU XIANGDING & LIN ZHENYAO (1988): Climatic fluctuation in Tibet since the Late Tertiary.- IN:
WHYTE, P. (ed.): The palaeoenvironment of East Asia from the Mid-Tertiary. Geology,
tectonics, orogeny, geomorphology. Centre of Asian Studies Occas. Pap. a. Monogr. 77,1:
421-432.
WU XINZHI (1988): China report - palaeoanthropology.- WHYTE, P. (ed.): The palaeoenvironment
of East Asia from the Mid-Tertiary. Invertebrate palaeontology and oceanography.- Centre of
Asian Studies Occas. Pap. a. Monogr. 77,2: 971-980.
WU ZHENGYI, TANG YANCHENG, LI XIWEN, WU SUGONG & LI HENG (1981): Dissertations upon the
origin, development and regionalization of Xizang flora through the floristic analysis.- IN:
LIU DONGSHENG (ch. ed.): Geological and ecological studies of Qinghai-Xizang Plateau.
Science Press Beijing. 2: 1219-1244.
WU ZIRONG & GAO FUQING (1985): The formation of loess in China.- IN: LIU TUNGSHENG (ch.
ed.): Quaternary Geology and Environment of China. China Ocean Press Beijing. 137-138.
XI YIZHEN (1993): Investigation on pollen morphology of Aconitum L.- Acta Bot. Sinica 35: 674-
686.
XIN YIQUN (1993): Studies on the pollen morphology of the genus Ulmus L. in China and its
taxonomic significance.- Acta Bot. Sinica 35: 91-95.
XING JIAMING (1988): The relationship between environment changes and human activities since
late Quaternary in North China.- IN: WHYTE, P. (ed.): The palaeoenvironment of East Asia
from the Mid-Tertiary. Invertebrate palaeontology and oceanography.- Centre of Asian
Studies Occas. Pap. a. Monogr. 77,2: 1076-1083.
XU QINGHAI, YANG XIAOLAN, WU CHEN, MENG LINGYAO & WANG ZIHUI (1996): Alluvial pollen
on the China Plain.- Quat. Res. 46: 270-280.
XU REN (1981): Vegetational changes in the past and the uplift of Qinghai-Xizang Plateau.- IN:
LIU DONGSHENG (ch. ed.): Geological and ecological studies of Qinghai-Xizang Plateau.
Science Press Beijing. 1: 139-144.
YALDIZ, M. (1987): Archäologie und Kunstgeschichte Chinesisch-Zentralasiens (Xinjiang).- IN:
B. SPULER (Hrsg.): Handbuch der Orientalistik. Leiden. 7. Abt. 7, Bd. 3, Abs. 2: 237.

10. Literaturverzeichnis 161
YANG XIAOPING (1991): Geomorphologische Untersuchungen in Trockenräumen NW-Chinas
unter besonderer Berücksichtigung von Badanjilin und Takelamagan.- Göttinger Geogr. Abh.
96: 124.
YANG YICHOU, LI BINGYUAN, YI ZESHENG & ZHANG QINGSONG (1981): Basic characteristics of
landforms on the Xizang Plateau.- IN: LIU DONGSHENG (ch. ed.): Geological and ecological
studies of Qinghai-Xizang Plateau. Science Press Beijing. 2: 1745-1757.
YANG YICHOU, LI BINGYUAN, YI ZESHENG & ZHANG QINGSONG (1982): The formation and
evolution of landforms in the Xizang Plateau.- Acta Geogr. Sinica 37: 76-87.
YAO TANDONG & L. G. THOMPSON (1992): Trends and features of climatic changes in the past
5000 years recorded by the Dunde Ice Core.- Annals of Glaciology 16: 21-24.
YOU YUZHU (1985): Preliminary study on Paleolithic bone engraving in Shiyu Shanxi Province.-
IN: LIU TUNGSHENG (ch. ed.): Quaternary Geology and Environment of China. China Ocean
Press Beijing. 269-272.
YU XIAOGAN & SUN SHANGZHI (1981): The upper limit of agriculture in Xizang and its factors
analysis.- IN: LIU DONGSHENG (ch. ed.): Geological and ecological studies of Qinghai-
Xizang Plateau. Science Press Beijing. 2: 2033-2044.
YUAN GUOYING (1986): The vertical zonation of the western Altay Mts.- Acta Geogr. Sinica 41:
32-40.
YUAN SHAOMIN & SUN XIANGJUN (1990): The vegetational and environmental history at the west
foot of Changbei Mountain, Northeast China during the last 10,000 years.- Acta Bot. Sinica
32: 558-567.
YUNUS, D. & P. K. K. NAIR (1989): Pollen morphology of indian Geraniales: a research
monograph.- Advances in Pollen Spore Res. 15/16 168.
ZANCHI, A. & D. GRITTI (1996): Multistage structural evolution of nothern Karakorum (Hunza
region, Pakistan).- Tectonophysics 260: 145-165.
ZAVADA, M. (1983): Pollen morphology of Ulmaceae.- Grana 22: 23-30.
ZEIST, W. VAN & S. BOTTEMA (1977): Palynological investigations in western Iran.-
Palaeohistoria 19: 19-85.
ZEIST, W. VAN. & S. BOTTEMA (1988): Late quaternary vegetational and climatic history of
Southwest Asia.- IN: JAIN, D. V. S., DHARA, M. & J. SAKETHARAMAN (eds.): Palaeoclimatic
and palaeoenvironmental changes in Asia during the last 4 million years. New Delhi. 129-
148.
ZEITLER, P. K., JOHNSON, N. M., NAESER, C. M. & R. A. K. TAHIRKHELI (1982): Fisson-track
evidence for quaternary uplift of the Nanga Parbat region, Pakistan. Nature 298: 255-257.

10. Literaturverzeichnis 162
ZHANG DEER (1985): Meteorological characteristics of dust fall in China since the historic times.-
IN: LIU TUNGSHENG (ch. ed.): Quaternary Geology and Environment of China. China Ocean
Press Beijing. 101-106.
ZHANG JIACHENG & LIN ZHIGUANG (1992): Climate of China.- New York. 376.
ZHANG JINGWEI, WANG JINTING, CHEN WEILIE & LI BOSHENG (1981): On the vegetation zoning in
the Qinghai-Xizang Plateau.- IN: LIU DONGSHENG (ch. ed.): Geological and ecological
studies of Qinghai-Xizang Plateau. Science Press Beijing. 2: 1919-1928.
ZHANG JINGWEI, LI BOSHENG, WANG JINTING, CHEN WEILIE & ZHAO KUIYI (1988): Vegetation of
Xizang (Tibet) (chin.).- The series of the scientific expedition of the Qinghai-Xizang
Plateau. Beijing. Map 1:3 Mio. 589.
ZHANG LANSHENG (1984): Reconstruction of the climate of the Late Pleistocene ice age in East
China.- IN: WHYTE, R. O., CHIU TZENANG, LEUNG CHIKEUNG & SO CHAKLAM (eds.): The
evolution of the East Asian environment: geology and palaeoclimate.- Centre of Asian
Studies Occas. Pap. a. Monogr. 59,1: 252-270.
ZHANG LANSHENG (1988): The trend towards dryness in north and western China since the Mid-
Pleistocene.- Centre of Asian Studies Occas. Pap. a. Monogr. 77,1: 445-452.
ZHANG PINGZHONG, WANG XIANBIN, CHEN JIANFA, LI CHUNYUAN & WANG SUMIN (1995): Delta 13C values and hydrogen index records in sediment organic matter of RH core of Zoige
Basin, eastern Qing-Zang (Tibet) Plateau and their environmental significance.- Science in
China (B) 38: 1015-1024.
ZHANG QINGSONG, LI BINGYUAN & ZHU LIPING (1994): New recognitions of quaternary
environment in the northwest Tibetan Plateau.- Acta Geogr. Sinica 49: 289-297
ZHANG RONGZU (1989): Case study on mountain environmental management: Nyemo County
(Tibet).- Internat. Centre f. Integr. Mount. Develop. Occas. Paper 13: 1-75
ZHANG SHANZHEN, HE YAUANLIANG & HE YUANLIANG (1985): Late palaeozoic
palaeophytogeographic provinces in China and their relationships with plate tectonics.-
Palaeontologia Cathayana 2: 77-86.
ZHANG XIAOYE, AN ZHISHENG, CHEN TUO, ZHANG GUANGYU, ARIMOTO, R. & B. J. RAY (1994):
Late Quaternary records of the atmospheric input of eolian dust to the center of the Chinese
Loess Plateau.- Quat. Res. 41: 35-43.
ZHANG YANQING & J. F. WELKER (1996): Tibetan alpine tundra responses to simulated changes in
climate: aboveground biomass and community responses.- Arctic a. Alpine Res. 28: 203-209.
ZHANG YONGZU & YANG QINYE (1981): Physico-geographical differentiation of the Qinghai-
Xizang Plateau.- Arctic a. Alpine Res. 1: 1851-1860.

10. Literaturverzeichnis 163
ZHANG YULONG & WU PANCHENG (1990): Studies on spore morphology of some chinese
Sphagnum L.- Acta Bot. Sinica 32: 85-90.
ZHANG YULONG, XI YIZHEN, ZHANG JINTAN, GAO GUIZHEN, DU NAIQIU, SUN XIANGJUN & KONG
ZHAOCHEN (1990): Spore morphology of chinese Pteridophytes.- Science Press Beijing. 592.
ZHAO KUIYI, WANG DEBIN & SONG HAIYUAN (1981): The swamps of the Xizang Plateau.- IN: LIU
DONGSHENG (ch. ed.): Geological and ecological studies of Qinghai-Xizang Plateau. Science
Press Beijing. 2: 1991-1998.
ZHAO SONGQIAO (1986): Physical geography of China.- Science Press Beijing. 209.
ZHAO SONGQIAO (1994): Geography of China.- New York. 322.
ZHAO YONGTAO (1994): The shrinking of Quaternary lakes and environmental changes on the
Tibetan Plateau.- IN: PÖRTGE, K.-H. & LI JIAN (Hrsg.): Beiträge zur Tibetforschung.
Göttinger Geogr. Abh. 95: 193-200.
ZHENG BENXING (1988): Studies of the glacial characteristics and environment of the Last Glaci-
ation in West China.- Centre of Asian Studies Occas. Pap. a. Monogr. 77,1: 453-463.
ZHENG BENXING (1989): The influence of himalayan uplift on the development of quaternary
glaciers.- IN: DERBYSHIRE E. & L. A. OWEN (eds.): Quaternary of the Karakoram and
Himalaya. Zeitsch. f. Geomorphol. N. F. Suppl. 76: 89-115.
ZHENG BENXING & SHI YAFENG (1985): Glacial variation since Late Pleistocene on the Qinghai-
Xizang (Tibet) Plateau of China.- IN: LIU TUNGSHENG (ch. ed.): Quaternary Geology and
Environment of China. China Ocean Press Beijing. 218-225.
ZHENG DU (1983): Untersuchungen zur floristisch-pflanzengeographischen Differenzierung des
Xizang-Plateaus (Tibet), China.- Erdkunde 37: 34-47.
ZHENG DU (1985): Study on the floristic phytogeographical differentiation of Xizang (Tibet).-
Acta Bot. Sinica 27: 84-93.
ZHENG DU & YANG QINYE (1985): Some problems on the altitudinal belts in southeastern
Qinghai-Xizang (Tibetan) Plateau.- Acta Geogr. Sinica 40: 60-69.
ZHENG DU, ZHANG YONGZU & YANG QINYE (1981): Physico-geographical differentiation of the
Qinghai-Xizang Plateau.- IN: LIU DONGSHENG (ch. ed.): Geological and ecological studies of
Qinghai-Xizang Plateau. Science Press Beijing. 2: 1851-1860.
ZHENG HONGHAN (1985): Paleoclimate events recorded in clay minerals in loess of China.- IN:
LIU TUNGSHENG (ch. ed.): Quaternary Geology and Environment of China. China Ocean
Press Beijing. 83-92.

10. Literaturverzeichnis 164
ZHENG YUANGCHANG (1994): Main problems of the ecologic environment in the north-eastern
part of the Qinghai-Xizang Plateau.- IN: PÖRTGE, K.-H. & LI JIAN (Hrsg.): Beiträge zur
Tibetforschung. Göttinger Geogr. Abh. 95: 263-271.
ZHOU GUOXING (1988): The first man in China: The date and environment of Yuanmou Man.- IN:
WHYTE, P. (ed.): The palaeoenvironment of East Asia from the Mid-Tertiary. Invertebrate
palaeontology and oceanography.- Centre of Asian Studies Occas. Pap. a. Monogr. 77,2:
1062-1068.
ZHOU JINSHANG (1987): The influence of Qinghai-Xizang on the climate over China.- IN: YE
DUZHENG, FU CONGBIN, CHAO JIPING & M. YOSHINO (eds.): The climate of China and global
climate. China Ocean Press Beijing. 345-346.
ZHOU S. Z., CHEN F. H., PAN B. T., CAO J. X., LI J. J. & E. DERBYSHIRE (1991): Environmental
change during the Holocene in western China on a millennial timescale.- The Holocene 1:
151-156.
ZHOU YOUWU, QIU GUOQING & GUO DONGXIN (1991): Quaternary permafrost in China.- Quat.
Scie. Revi. 10: 511-517.
ZHU CHENG (1995): The new recognitions to genesis quaternary sediments and neotectonics in the
east piedmont of the Lushan Mountain.- Acta Geogr. Sinica 50: 41-49.
ZOU HOUYUAN, LIANG YIMIN & SUN GIANTING (1980): A preliminary study on vegetation
regionalism in Loess Plateau of the North Shanxi Sheng.- Acta Bot. Sinica 22: 399-401.
ZOU JINSHANG (1987): The influence of Qinghai-Xizang Plateau on the climate over China.- IN:
YE DUZHENG, FU CONGBIN, CHAO JIPING & M. YOSHINO (eds.): The climate of China and
global climate. China Ocean Press Beijing. 345-346.

11. Anhang – Index der Pollentypen in Kap. 4.5 165
11 Anhang
11.1 Index der Pollentypen in Kap. 4.5
A
Abies........................................................................11
Acantholimon .........................................................20
Acer.........................................................................24
Aconitum ................................................................17
Alnus.......................................................................14
Androsace ...............................................................20
Anemone/Ranunculus-Typ ....................................17
Anthemis-Typ.........................................................23
Apiaceae p. p..........................................................20
Arceuthobium .........................................................14
Artemisia.................................................................23
Asteroideae .............................................................23
Aster-Typ................................................................23
B
Batrachium-Typ.....................................................17
Betula......................................................................13
Boraginaceae..........................................................24
Botrychium .............................................................26
Botryococcus ..........................................................24
Brassicaceae...........................................................17
Bupleurum-Typ ......................................................20
Buxus ......................................................................24
C
Caltha-Typ ..............................................................17
Campanula-Typ .....................................................24
Cannabis-Typ .........................................................14
Caragana-Typ.........................................................18
Carpinus cordata-Typ.............................................14
Cedrus.....................................................................12
Centaurea/Cousinia-Typ .......................................23
Cerastium-Typ .......................................................16
Ceratostigma ...........................................................20
Chenopodiaceae .....................................................16
Cichorioideae ..........................................................22
Corylus ....................................................................14
Crassulaceae...........................................................17
Cyperaceae.............................................................24
D
Diphasiastrum-Typ .................................................25
Diploxylon-Typ.......................................................11
E
Ephedra distachya-Typ...........................................13
Ephedra fragilis-Typ...............................................13
Equisetum ...............................................................26
Eremurus ................................................................23
Erodium ..................................................................19
Euphorbia ...............................................................24
Eurumex-Typ ..........................................................15
F
Fabaceae p. p. ........................................................18
Fagus ......................................................................14
Filicatae monolet p. p. ...........................................26
Filicatae tr ilet p. p. ................................................26
Fraxinus excelsior-Typ ..........................................20
Fraxinus floribunda-Typ .......................................21
G
Galium-Typ ............................................................21
Gentianaceae p. p. .................................................21
Geranium ................................................................19

11. Anhang – Index der Pollentypen in Kap. 4.5 166
H
Haloxylon-Typ........................................................16
Haploxylon-Typ .....................................................11
Hedera ....................................................................24
Hedysarum-Typ ......................................................18
Hepaticae p. p. .......................................................25
Hippophaë ..............................................................21
Hippuris ..................................................................19
Hordeum-Typ.........................................................24
Huperzia .................................................................25
Hypericum-Typ ......................................................24
J
Juglans ...................................................................13
Juniperus ................................................................12
Juniperus-Typ.........................................................12
K
Koenigia..................................................................16
L
Larix .......................................................................12
Lemna .....................................................................24
Liliaceae .................................................................23
Linum .....................................................................24
Lonicera..................................................................22
M
Malvaceae ..............................................................24
Melandrium-Typ ....................................................16
Mentha-Typ............................................................21
Myriophyllum alternifolium-Typs .........................19
Myriophyllum spicatum-Typ .................................19
Myriophyllum verticillatum-Typ ...........................19
N
Nitraria....................................................................19
Nymphaea tetragona...............................................17
O
Onagraceae ............................................................19
P
Pediastrum ..............................................................24
Pedicularis oederi-Typ ...........................................22
Pedicularis palustris-Typ .......................................22
Picea........................................................................11
Picea omorica/Abies-Typ........................................11
Picea omorica-Typ .................................................11
Pinaceae...................................................................11
Pinus .......................................................................11
Pistacia....................................................................21
Plagiogyria..............................................................26
Plantago alpina-Typ...............................................22
Plantago lanceolata ................................................22
Plantago major/media-Typ ....................................22
Poaceae ...................................................................24
Poaceae > 37 µm ....................................................24
Polygonum alpinum-Typ .......................................16
Polygonum amphibium-Typ ..................................16
Polygonum aviculare-Typ ......................................15
Polygonum bistorta-Typ.........................................15
Polygonum glaciale ................................................16
Polygonum persicaria-Typ.....................................16
Polygonum sibiricum ..............................................15
Polypodium-Typ .....................................................26
Populus ...................................................................13
Potamogeton/Triglochin-Typ.................................23
Potentilla-Typ.........................................................18
Primula farinosa-Typ.............................................20
Primula veris-Typ ..................................................20
Prunella-Typ ..........................................................21
Pteridium-Typ ........................................................27
Pteris-Typ ...............................................................27

11. Anhang – Index der Pollentypen in Kap. 4.5 167
Pterocarya...............................................................13
Q
Quercus...................................................................14
R
Rheum.....................................................................15
Rhinanthus-Typ .....................................................22
Rhododendron ........................................................20
Riccia-Typ ..............................................................25
Riella.......................................................................25
Rosaceae p. p. ........................................................18
Rumex-Typ .............................................................15
S
Salix ........................................................................13
Salix oritrepha-Typ ................................................13
Sanguisorba filiformis............................................18
Saussurea-Typ .......................................................23
Saxifraga granulata-Typ .......................................18
Saxifraga oppositifolia-Typ ...................................18
Saxifraga stellaris-Typ...........................................18
Schizaea ..................................................................26
Scrophular iaceae p. p............................................22
Selaginella ..............................................................25
Sibiraea-Typ...........................................................18
Sparganium-Typ ....................................................24
Sphagnum ...............................................................25
Stachys-Typ ............................................................22
Staurastrum ............................................................24
Swertia-Typ ............................................................21
T
Targionia ................................................................25
Thalictrum ..............................................................17
Thesium ..................................................................14
Thymelaea-Typ.......................................................19
Tilia .........................................................................24
Tribulus...................................................................19
Tripterospermum ....................................................21
Triticum-Typ ..........................................................24
Trollius....................................................................17
Tsuga.......................................................................12
Typ ..........................................................................16
U
Ulmus-Typ ..............................................................14
Urtica ......................................................................14
Utricularia ..............................................................22
V
Valer ianaceae.........................................................22
Viola palustris-Typ.................................................22
X
Xanthium ................................................................23

11. Anhang – Liste der Profile 168
11.2 Liste der Profile
Kapitel 6.1.: Lingfeng im Qinling-Gebirge
-Profil Lingfeng (Pollendiagramm: Abb. 4, in der Beilage) Prof. Dr. H.-J. Pachur und B. Wünnemann, 34°17’N/104°08’E Höhe: 2550 m Länge: 21 m (Straßenaufschluß und Grabung) Datierungen: 107 cm, 17500 +1240/-1075 BP (Hv 21342) 135 cm, 27205 +1275/-1100 BP (Hv 21341) 215 cm, >43770 BP (Hv 21340) 240 cm, 34230 +2480/-1890 BP durch rezente Verunreinigung (Hv 21339) 322 cm, >44000 BP (Hv 21338) 348 cm, >38800 BP (Hv 21337) aus Pachur et al. (1994): 1410 cm, 113000 +48000/-33000 (Uh 768) 1610 cm, 73000 +26000/-21000 (Uh 771) 2026 cm, 105000 ± 13900 (Uh 770)
Kapitel 6.2.: Der Gaxun Nur in der Inneren Mongolei
-Profil Gaxun Nur (100) (Pollendiagramm: Abb. 6) Prof. Dr. H.-J. Pachur und B. Wünnemann, Innere Mongolei bei etwa 41°30’N/101°30’E Höhe: ca. 950 m Länge: 235 m Datierung: in 191 m Tiefe Brunhes/Matuyama-Grenze
Kapitel 6.3.1.: Südöstliches Tibet: Der Nianbaoyeze
Die Profile wurden von Herrn PD Dr. F. LEHMKUHL (Göttingen) genommen, sofern keine anderen Angaben erfolgen handelt es sich um Cyperaceen-Torfe. -Profil 2, im Tal des Ekuoduoma-Sees 2.8.1991 bei 33°13’N/100°59’E Höhe: 4100 m Länge: 290 cm Datierung: 96 - 101 cm, 3635 ± 115 BP, 2140 - 1785 BC (Hv 21316) Die oberen 130 cm des Profils bestehen aus Cyperaceen-Torf, dessen Bildung etwa vor 5000 BP begann. Dieser Zeitraum ist in mehreren Diagrammen aus dem Nianbaoyeze erfaßt, so daß hier auf eine nähere Untersuchung verzichtet wurde, zumal unterhalb von 130 cm ein für Pollenanalysen unergiebiges tonig-schluffiges Material ansteht. Der Eintrag dieses Materials erfolgte während des mittelholozänen Klimaoptimums bis vor etwa 5000 BP.

11. Anhang – Liste der Profile 169
- Profil 3, im Tal des Lerzha-Flusses (Pollendiagramm: Abb.11, in der Beilage) 5.8.1991 bei 33°21’N/101°02’E Höhe: 4170 m Länge: 330 cm Datierungen: 62 - 69 cm, 2680 ± 100 BP, 910 - 790 BC (Hv 21318) 187 - 195 cm, 6555 ± 140 BP, 5580 - 5330 BC (Hv 21317) 300 - 320 cm, 9185 ± 145 BP, 8390 - 8040 BC (Hv 18411) -Profil 8, im Tal des Ximencuo-Sees 12.8.1991, 33°21’N/101°06’E Höhe: 4065 m Länge: 91 cm Datierung: 80 - 89 cm, 3055 ± 190 BP, 1515 - 1005 BC (Hv 18415) Der im Profil überlieferte Zeitraum ist in mehreren Diagrammen erfaßt. Die Voruntersuchungen der Proben gaben keine Hinweise auf abweichende vegetationsgeschichtliche Ergebnisse. In 50 bis 79 cm Tiefe ist in den Cyperaceen-Torf eine Granitgruslage eingeschaltet. - Profil 9, im Tal des Ximencuo-Sees (Pollendiagramm: Abb. 12) 13.8.1991 bei 33°24’N/101°06’E Höhe: 4050 m, Länge: 143 cm Datierungen: 52 - 54 cm, 1990 ± 110 BP, 100 BC - 130 AD (Hv 21320) 92 - 94 cm, 4900 ± 180 BP, 3940 - 3385 BC (Hv 21319) Parallelprofil: 60 - 100 cm, 4775 ± 55 BP, 3640 - 3385 BC (Hv 18416) -Profil 11, im Tal des Nigequ-Flusses (Tab. 1) 19.8.1991, 33°25’N/101°12’E Höhe: 4140 m Länge: 254 cm Datierung: 221 - 234 cm, 4295 ± 75 BP, 2925 - 2790 BC (Hv 20057) Die hohen Cyperaceen-Anteile und die wenigen Funde von Picea und Juniperus bestätigen die Baumarmut der Landschaft und die dominante Rolle der Kobresia-Matten in den letzten 5 Jahrtausenden. Die Flußaktivität am Oberlauf des Nigequ reichte offenbar nicht aus um das Aufkommen von Pioniergebüschen (Hippophaë, Salix, Potentilla) zu fördern. -Profil 12, im Tal des Ha’acuo-Sees (Tab. 12) 24.8.1991, 33°18’N/101°14’E Höhe: 4010 m Länge: 153 cm Datierung: 90 - 100 cm, 1630 ± 70 BP, 380 - 540 AD (Hv 21321) Das Profil reicht über 2000 Jahre zurück und belegt die Baumarmut des Gebietes. Im oberen Abschnitt (jünger 700 BP) lassen der Rückgang von Poaceen und die Zunahme von Juniperus eine anthropo-zoogene Beeinflussung vermuten.

11. Anhang – Liste der Profile 170
-Profil 13, im Tal des Magenacuo-Sees 26.8.1991, 33°12’N/101°12’E Höhe: 3990 m Länge: 95 cm Datierung: 85 - 90 cm, 1380 ± 95 BP, 605 - 760 AD (Hv 20058) Die Datierung erfolgte unmittelbar unterhalb einer Gruslage in 45 bis 85 cm Tiefe. -Profil 14, Becken des Ji’ea (Pollendiagramm: Abb. 15) 31.8.91, 33°16’N/101°24’E Höhe: 3870 m Länge: 91 cm Datierungen: 45 - 50 cm, 1285 ± 65 BP, 670 - 790 AD (Hv 21323) 75 - 86 cm, 4320 ± 150 BP, 3255 - 2700 BC (Hv 21322) -Profil 15, Tal im Einzugsgebiet des Kekehe (Pollendiagramm: Abb. 13) 9.9.1991, 33°08’N/101°01’E Höhe: 4130 m Länge: 60 cm Datierungen: 38 - 44 cm, 2660 ± 65 BP, 840 - 795 BC (Hv 21324) 51,5 - 58,5 cm, 4655 ± 75 BP, 3610 - 3350 BC (Hv 20059) -Profil 16, Tal im Einzugsgebiet des Kekehe (Pollendiagramm: Abb. 14) 9.9.1991, 33°08’N/101°00’E Höhe: 3960 m Länge: 80 cm Datierungen: 32 - 37 cm, 1890 ± 95 BP, 20 - 240 AD (Hv 21325) 52 - 67 cm, 5400 ± 95 BP, 4345 - 4090 BC (Hv 20060)
Kapitel 6.3.2.: Südliches Tibet: Im Stadtbereich von Lhasa
-Profil Lhasa Stadtbereich (Pollendiagramm: Abb. 16,in der Beilage) PROF. DR. G. MIEHE, 5.9.1994, 29°40’N/91°08’E Höhe: 3650 m Länge: 150 cm Datierung: 81 - 86,5 cm, 575 ± 120 BP, 1295 - 1440 AD (Hv 21331)
Kapitel 6.3.3.: Weitere Untersuchungen in Tibet
Alle untersuchten Profile wurden von Herrn Prof. Dr. G. MIEHE beschafft. Für die meisten der Profile liegen noch keine eingehenden Untersuchungen vor, so daß in solchen Fällen auf eine Darstellung der pollenanalytischen Ergebnisse verzichtet werden konnte. -Profil Soden 3 (Pollendiagramm: Abb. 17) 1994, 28°32’N/92°25’E Höhe: 5040 m Länge: 12 cm; Datierungen: keine

11. Anhang – Liste der Profile 171
-Profil Soden 18 (Pollendiagramm: Abb. 17) 1994, 31°43’N/100°44’E Höhe: 4260 m Länge: 18 cm Datierung: 6 - 7 cm, 106,2 ± 2,8 BP (Hv 21347) Auch die konventionellen 14C-Datierungen an weiteren Soden in 12,5 bzw. 16 cm Tiefe ergaben Alter zwischen 100 - 106 BP. Für die Rekonstruktion der Entwicklungsgeschichte der in weiten Teilen des östlichen Tibets dominierenden Kobresia-Matten wären vermutlich Altersbestimmungen an kleinen Kohlenstoffmengen (etwa Kobresia-Nüsschen) vorzuziehen. -Profil Lager 16 26.8.1994, 29°36’N/94°41’E Höhe: 3730 m Länge: 130 cm Datierungen: 51 - 58 cm, 1235 ± 100 BP, 675 - 950 AD (Hv 21333) 102 - 108,5 cm, 2415 ± 85 BP, 760 BC - 395 AD (Hv 21332) -Profil Ganzi 29.9.1994, 31°39’N/99°06’E Höhe: 3550 m Länge: 140 cm Datierung: 75 - 77 cm, 480 ± 90 BP, 1405 - 1480 AD (Hv 21334) -Profil Malinganko 28.9.1994, 31°52’N/99°06’E Höhe: 4050 m Länge: 140 cm Datierungen: 70 - 75 cm, 5295 ± 125 BP, 4320 - 3975 BC (Hv 21336) 108 - 113 cm, 6055 ± 150 BP, 5210 - 4785 BC (Hv 21335) -Profil Nagartse 3 1.8.1994, ca. 30°N/89° am See Yamco Yumco Länge: 188 cm -Profil Nagqu 3 12.9.1994 Länge: 90 cm
Kapitel 6.4.2.1.: Der Nanga Parbat
-Profil Märchenwiese Rampe (Pollendiagramm: Abb. 20, in der Beilage) Prof. Dr. M. WINIGER und Dr. U. SCHICKHOFF 25.8.1993, 35°23’N/74°34’E Höhe: ca. 3300 m Länge: 326 cm Datierungen: 51 - 59 cm, 660 ± 210 BP, 1195 - 1445 AD (Hv 21315) 132 - 140 cm, 2900 ± 310 BP, 1495 - 795 BC (Hv 21314) 202 - 209 cm, 6350 ± 235 BP, 5475 - 5045 BC (Hv 21313) 311 - 314 cm, 7725 ± 185 BP, 6700 - 6380 BC (Hv 20062) 311,5 - 324 cm, 7785 ± 100 BP, 6645 - 6965 BC (Hv 20063)

11. Anhang – Liste der Profile 172
-Profil Märchenwiese Hang (Pollendiagramm: Abb. 21) Prof. Dr. M. WINIGER und Dr. U. SCHICKHOFF 25.8.1993, 35°23’N/74°34’E Höhe: ca. 3300 m Länge: 84 cm Datierung: 57 - 60 cm, 1480 ± 145 BP, 425 - 675 AD (Hv 21312)
Kapitel 6.4.2.2.: Der Karakorum
-Profil Sirle Bala (Pollendiagramm: Abb. 22) Prof. Dr. M. WINIGER und Dr. U. SCHICKHOFF, 1993, oberes Astor-Tal Höhe: 3010 m Länge: 83 cm Datierung: 48 - 53 cm, 1880 ± 60 BP, 75 - 230 AD (Hv 20061) -Profil Shukan 2 (Pollendiagramm: Abb. 23) Prof. Dr. G. MIEHE 7.9.1990, 36°20’N/73°10’E Höhe: 3360 m Länge: 60 cm Datierungen: 15 - 18 cm, 1095 ± 45 BP, 895 - 1005 AD (Hv 20064) 28 - 31 cm, 980 ± 55 BP, 1010 - 1160 AD (Hv 20065) 53 - 60 cm, 4995 ± 190 BP, 3980 - 3630 BC (Hv 17900) Aufgrund der Altersinversion blieben die zwei jüngeren Datierungen bei der Interpretation unberücksichtigt. -Profil Batyatum Jut (Pollendiagramm: Abb. 24) Dr. J.-P. JACOBSEN 17.7.1994, 36°30’N/73°02’E Höhe: 3550 m Länge: 93 cm Datierungen: 37 - 38,5 cm, 1830 ± 70 BP, 120 - 320 AD (Hv 21327) 82 - 84 cm, 2550 ± 75 BP, 800 - 535 BC (Hv 21326) -Profil Alambar (Pollendiagramm: Abb.25) Dr. J.-P. JACOBSEN 21.7.1994, 36°40’N/73°30’E Höhe: 3700 m Länge: 78 cm Datierungen: 32,5 - 34 cm, 1230 ± 75 BP, 685 - 890 AD (Hv 21329) Juniperus - Holz aus 75 cm, 1115 ± 190 BP, 685 - 1160 AD (Hv 21330) 73 - 74,5 cm, 2355 ± 80, 490 - 375 BC (Hv 21328) An dem Juniperus - Holz aus 75 cm Tiefe war eine Ansprache als Stamm- oder Wurzelholz nicht möglich, seinem Alter nach dürfte es von einer Wurzel stammen.

11. Anhang – Liste der Profile 173
Kapitel 6.4.2.2.: Weitere Profile
Die Profile stammen von Herrn Prof. Dr. G. MIEHE: -Profil Deosai Plateau (Tab. 7) 13.08.1990, 35°02’N/75°25’E Höhe: 3950 m Länge: 76 cm, gegraben Datierung: 52 - 57 cm, 1500 ± 115, 420 - 650 AD (Hv 17901) -Profil Bagrot (Tab.6) 27.9.1990, 36°03’N/74°38’E Höhe: 3270 m Länge: 10 cm Datierungen: keine -Profil Shukan 1 (Tab.3) 7.9.1990, 36°23’N/73°05’E Höhe: 3400 m Länge: 29 cm Datierungen: keine -Profil Karakorum 1 (Tab. 5) 4.8.1990, 36°02’N/74°08’E Höhe: 3750 m Länge: 61 cm Datierung: 50 - 61 cm, 1105 ± 175 BP, 685 - 1150 AD (Hv 17899) -Profil Karakorum 2 (Tab. 4) 6.9.1990, nahe Shukan Höhe: 3600 m Länge: 77 cm Datierung: 54 - 58 cm, 3320 ± 105 BP, 1745 - 1460 BC (Hv 17898)

11. Anhang – Liste der Abbildungen 174
11.3 Liste der Abbildungen
im Text:
Abb. 1: Naturräumliche Gliederung Chinas nach ZHAO SONGQIAO (1994) in die Regionen A = Ost-China, B = Nord-China und C = Tibetanisches Plateau (Qinghai-Xizang-Plateau) mit den im Text erwähnten Unterregionen 1 = Südöstliches-Tibet, 2 = Süd-Tibet, 3 = Qaidam-Becken, 4 = Qinling Gebirge und Dabie Shan, 5 = Rotes Becken, 6 = Ordos-Gebiet, 7 = Alaschan Plateau, 8 = Tarim-Becken sowie Lage der Untersuchungsräume (Karakorum/Nanga Parbat, Nianbaoyeze) und einiger Untersuchungspunkte (Lhasa, Lingfeng, Gaxun Nur)……………………………………………………………............................………3
Abb. 2: Heutige Politische Gliederung Chinas (nach ZHAO SONGQIAO 1994). Das Tibetanische Plateau wird im wesentlichen von der Autonomen Region Tibet, der Provinz Qinghai und dem westlichen Sichuan eingenommen…….….......…………..........................................................................….38
Abb. 3.: Blockbild der Landschaft bei Lingfeng und Lage des untersuchten Profils (aus PACHUR et al. 1994). Das Gelände erreicht Höhen zwischen etwa 2500 und 2900 m ü. M..............................................................................................................................49
Abb. 5: Versuch einer Korrelation der Vegetations- und Klimageschichte von Lingfeng (Qinling Shan) mit der chinesischen Lößstratigraphie, den Isotopenstadien der Tiefsee (in Anlehnung an AN ZHISHENG et al. 1991a, KUKLA & AN ZHISHENG 1989, FORMAN
1991) und dem Oberpleistozän Mitteleuropas (GRÜGER 1989, 1995, BEHRE & LADE 1986). Temperaturverlauf stark schematisiert………………………………………….58
Abb. 6: Pollendiagramm des Profils „Gaxun Nur“, Innere Mongolei, ca. 950 m. ü. M. Brunhes/Matuyama-Grenze in 191 m Tiefe. (GS = GhP + NGhP, vereinfachte Wiedergabe)…………………………………………………………………………….63
Abb. 7: Höhenstufen der Vegetation am Ostrand des Tibetanischen Plateaus vom Roten Becken bis zum Nianbaoyeze (aus LEHMKUHL 1995a). Ab etwa 150 km östlich des Nianbaoyeze sind Wälder von Picea und Abies auf Nordhänge beschränkt während auf den Südhängen Juniperus-Bäume stehen………………………………………………69
Abb. 8: Pollendiagramm der Oberflächenproben aus dem Nianbaoyeze. Moospolster, Probe 6 Caryophyllaceen-Polster. (GS = GhP + NGhP ohne Caryophyllaceen)………72
Abb. 10: Größenverteilung der Betula-PK im Profil 3 aus dem Nianbaoyeze. Die PK ab einer Größe von 30 µm dürften von Betula utilis stammen oder aus größerer Distanz angeweht sein (B. platyphylla, B. albo-sinensis). PK unter 30 µm lassen sich, soweit bekannt, keiner Art zuordnen, und ihre geringe Größe ist möglicherweise auf Sedimenteinflüsse zurückzuführen..................................................................................74
Abb. 12: Pollendiagramm des Profils 9 aus dem nördlichen Nianbaoyeze, Ximencuo-Tal, 4050 m. ü. M. Stratigraphie: an der Basis Schluffmudde, darüber Cyperaceen-Torf, bei 1 m stark schluffig. (GS = GhP + NGhP)………………………………………………78
Abb. 13: Pollendiagramm des Profils 15 aus dem südlichen Nianbaoyeze, Einzugsbereich des Kekehe, 4130 m. ü. M. Stratigraphie: Cyperaceen-Torf. (GS = GhP + NGhP)……80
Abb. 14: Pollendiagramm des Profils 16 aus dem südlichen Nianbaoyeze, Einzugsbereich des Kekehe, 3960 m. ü. M. Stratigraphie: Cyperaceen-Torf. (GS = GhP + NGhP)…....82
Abb. 15: Pollendiagramm des Profils 14 aus dem Ji’ea-Becken östlich des Nianbaoyeze, 3870 m. ü. M. Stratigraphie: Cyperaceen-Torf, im unteren Teil schluffig-tonig (GS = GhP + NGhP)..……………………………………………………………………….…83

11. Anhang – Liste der Abbildungen 175
Abb. 17: Pollendiagramme der Soden 3 und 18, südliches Tibet, 5040 und 4260 m. ü. M. Beide Profile stammen aus dichtfilzigen Kobresia-Matten. (GS = GhP + NGhP)…….96
Abb. 18: Klimaeinflüsse im Raum Karakorum/Nanga Parbat und den umliegenden Gebieten. Schraffur: waagerecht = subtropisch-mediterraner Klimaeinfluß von Westen, Niederschläge im Frühjahr, senkrecht = Übergangsgebiet, Frühjahrs-Niederschläge höher als Sommer-Niederschläge, diagonal (im Süden) = randtropisch-monsunaler Klimaeinfluß von Südosten, Sommer-Niederschläge mindestens so hoch wie die Frühjahrs-Niederschläge, diagonal (im Nordosten) = zentralasiatisch-kontinentaler Klimaeinfluß von Nordosten, Wintertrockenheit und gradueller Sommer-Niederschlag (nach REIMERS 1992)…………………………………………………………………...99
Abb. 19: Verteilungsmuster der feuchteabhängigen hochmontanen und alpinen Pflanzenfor-mationen mit dem Verlauf der feuchtebedingten Verbreitungsgrenze von Pinus wallichiana und Picea smithiana (nach MIEHE et al. 1996) sowie die Lage der im Raum Karakorum/Nanga Parbat untersuchten Profile……………………………………….103
Abb. 21: Pollendiagramm des Profils „Märchenwiese Hang“, Nanga Parbat, 3300 m. ü. M. Stratigraphie: schluffig-toniger Sand mit humosen Lagen, oben Cyperaceen-Torf. (GS = GhP)…………………………………………………………………………………...108
Abb. 22: Pollendiagramm des Profils „Sirle Bala“ aus dem Karakorum, oberes Astor-Tal, 3010 m. ü. M. Stratigraphie: unterer Teil helle Mudde, darüber anmooriger Bereich mit Pflanzenstreu, darüber gleyartiges Material, obere 30 cm Cyperaceen-Torf. (GS = GhP + NGhP)……………………………………………………………………………….110
Abb. 23: Pollendiagramm des Profils „Shukan 2“ aus dem Karakorum, Talschaft Yasin, Seitental des Naz Bar-Tales, 3360 m. ü. M. Stratigraphie: an der Basis sandig-humoser Kies, darüber Cyperaceen-Torf. (GS = GhP + NGhP)………………………………..111
Abb. 24: Pollendiagramm des Profils „Batyatum Jut“ aus dem Karakorum, Talschaft Yasin, Khaimetbar-Tal, 3550 m. ü. M. Stratigraphie: Cyperaceen-Torf. (GS = GhP + NGhP)…………………………………………………………………………………113
Abb. 25: Pollendiagramm des Profils „Alambar“ aus dem Karakorum, Talschaft Yasin, Alambar-Tal, 3700 m. ü. M. Stratigraphie: Cyperaceen-Torf. (GS = GhP + NGhP)............................................................................................................................114
In der Beilage:
Abb. 4: Pollendiagramm des Profils „Lingfeng“, Qinling-Gebirge, 2250 m. ü. M. Stratigraphie siehe Legende. (GS = GhP).
Abb. 9: Karte des Nianbaoyeze mit Lage der untersuchten Profile und Angaben zur Vegetation und Geologie (nach Lehmkuhl 1995a).
Abb. 11: Pollendiagramm des Profils 3 aus dem nordwestlichen Nianbaoyeze, Lerzha-Tal, 4170 m. ü. M. Stratigraphie: tonig-schluffige Detritusmudde, darüber Cyperaceen-Torf. (GS = GhP + NGhP).
Abb. 16: Pollendiagramm des Profils „Lhasa Stadtbereich“, östliches Lhasa, 3650 m. ü. M. Stratigraphie: untere 60 cm schluffig-sandiges Material, darüber Detritusmudde, obere 10 cm Cyperaceen-Torf (GS = GhP + NGhP).
Abb. 20: Pollendiagramm des Profils „Märchenwiese Rampe“, Nanga Parbat, 3300 m. ü. M. Stratigraphie: Cyperaceen-Torf, besonders im unteren Teil sehr reich an (Pinus-)Holz. (GS = GhP).

11. Anhang – Verwendete Kürzel und fremdsprachliche Begriffe 176
11.4 Verwendete Kürzel und fremdsprachliche Begriffe
Abkürzungen:
BP years before present – Radiokarbonjahre vor 1950
AD Anno Domini, calibrated 14C-years
n. Chr. nach Christus, kalibrierte 14C-Jahre
DA Diagrammabschnitt
GhP Gehölzpollen
GS Grundsumme
JMT Jahresmitteltemperatur
LGM Letztglaziales Maximum
NGhP Nichtgehölzpollen
PK Pollenkorn/Pollenkörner
p. p. pro parte
VP Vergleichspräparat/Vergleichspräparate
fremdsprachliche Begriffe:
gobi Steinwüste
he/ho Fluß (z. B. Huang He)
hu See (z. B. Qinghai Hu)
-ling Gebirgszug (z. B. Qinling)
nur See (z. B. Gaxun Nur)
shan Berg, Gebirge (z. B. Qilian Shan)
shamo Sandwüste
tso (cuo, co) See (z. B. Ximencuo)

11. Anhang – Tabellen 177
11.5 Tabellen

11. Anhang – Tabellen 178
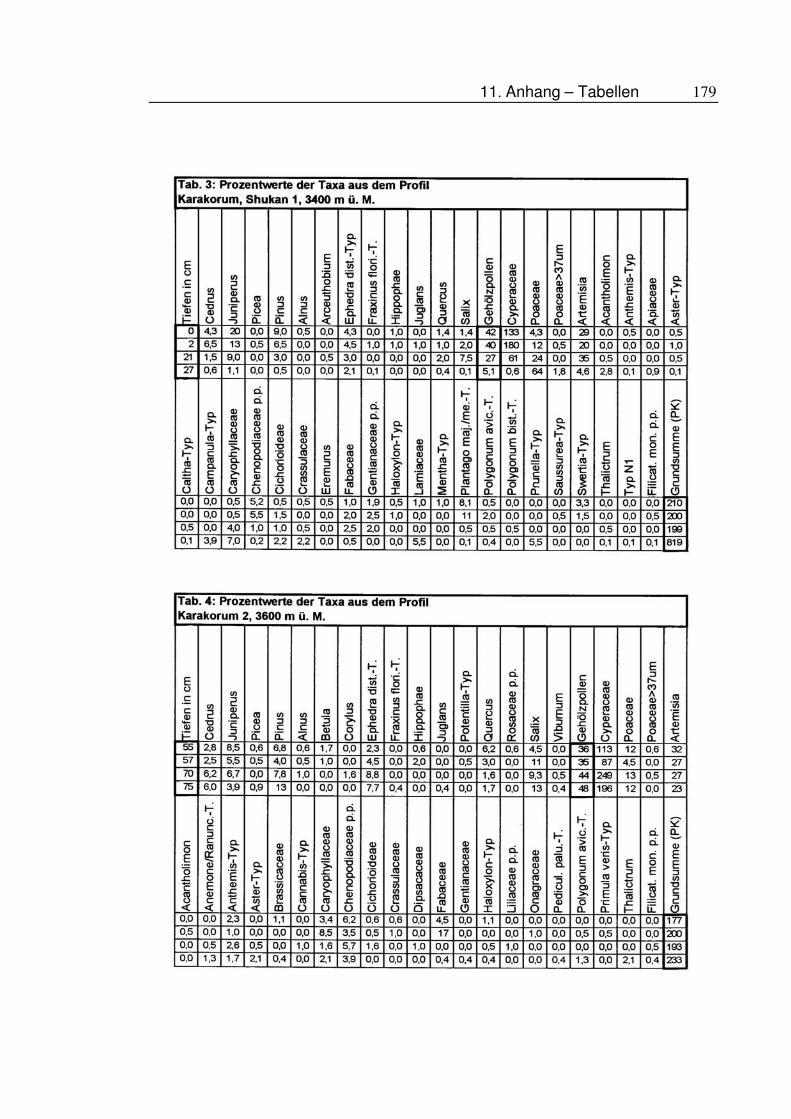
11. Anhang – Tabellen 179

11. Anhang – Tabellen 180
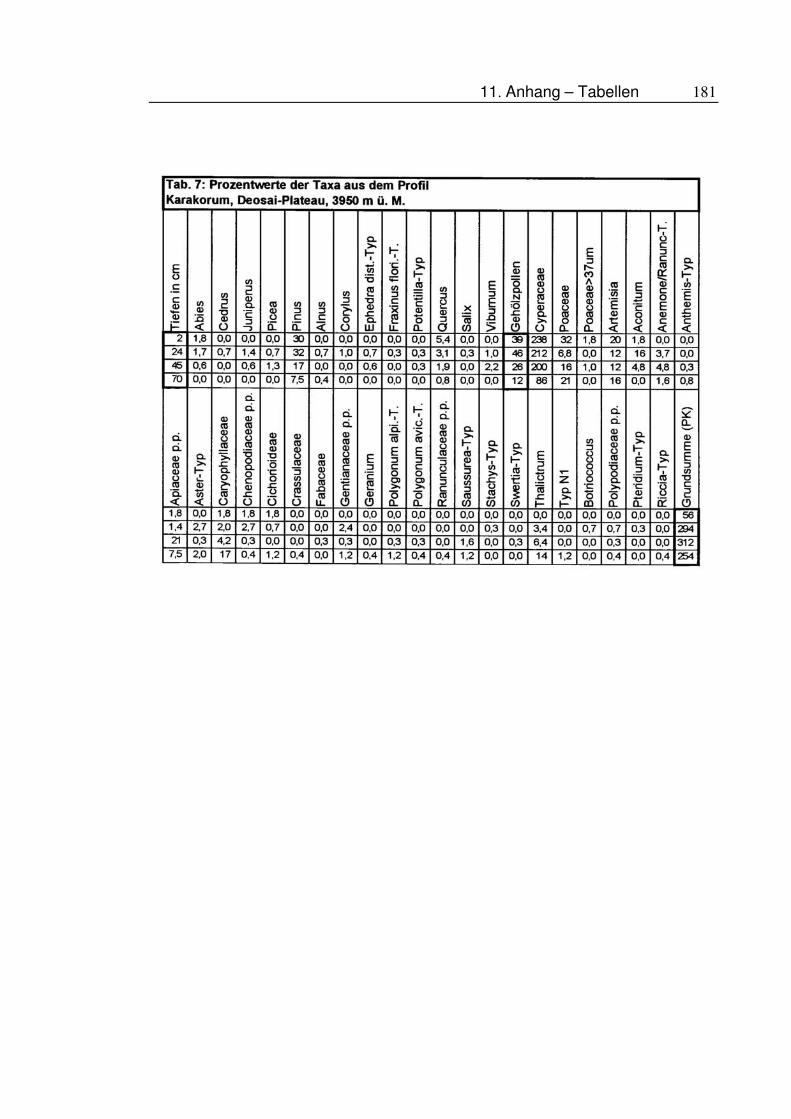
11. Anhang – Tabellen 181

11. Anhang – Fototafel 182
11.6 Fototafel
1,2: Haloxylon thomsonii (leg. SCHICKHOFF 1827/det. DICKORÉ; 1250fach):
Skulptur mikroechinat (1), Umriß des PK im optischen Schnitt polygonal (2).
3,4: Sanguisorba filiformis (Profil Malinganko, 1250fach):
abgebildete PK mit verhältnismäßig kleinen Opercula, (3) in äquatorialer und (4) in
polarer Ansicht.
5: Tribulus spec. (Profil Lhasa Stadtbereich, 1250fach):
PK sphäroidisch, jedes Lumen mit einer Pore.
6,7: Eremurus himalaicus (Alter Botanischer Garten Göttingen, 1250fach):
(6) distale Seite, (7) proximale Seite.
8,9: Riella cf. halophila (Profil Lhasa Stadtbereich, 500fach):
(8) Reticulum in Aufsicht (distale Seite?), (9) Reticulum im optischen Schnitt, die
Foveolae treten nur bei einzelnen Sporen auf und sind vermutlich erhaltungs-
bedingt.

11. Anhang – Fototafel 183

Beilage Abb. 4

Beilage Abb. 9
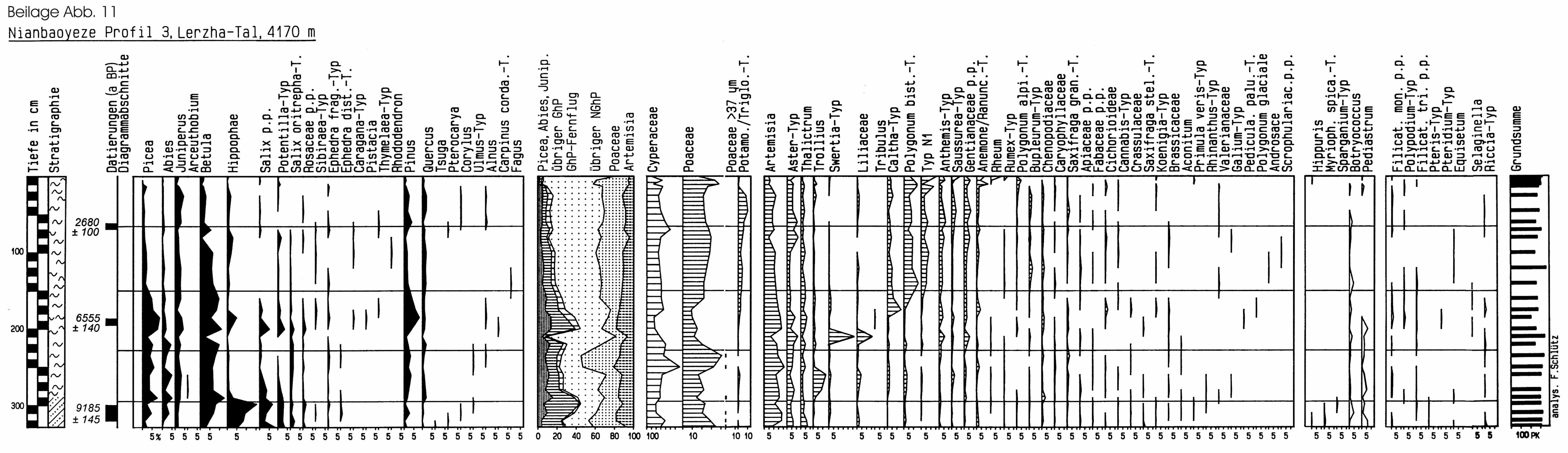
Beilage Abb. 11

Beilage Abb. 16
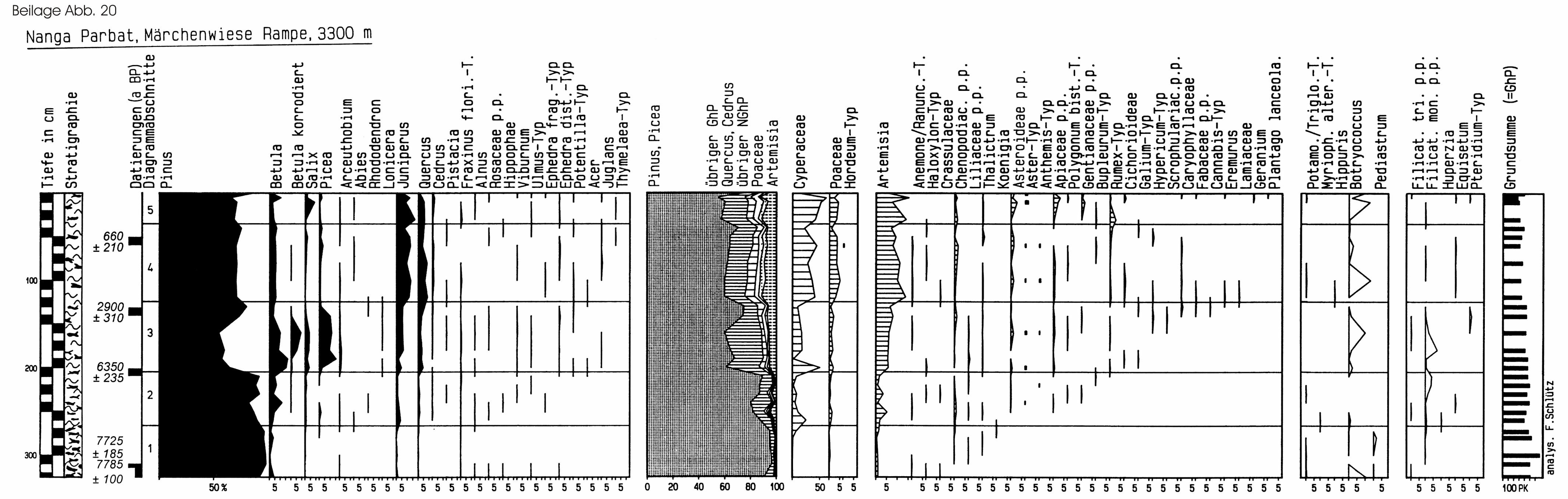
Beilage Abb. 20




















