
Novel adenoviral vectors coding for GFP-tagged wtCFTR and ΔF508-CFTR: characterization of...
10
ION CHANNELS, TRANSPORTERS Horia Vais Guang-Ping Gao Michael Yang Phoi Tran Jean-Pierre Louboutin Suryanarayan Somanathan James M. Wilson William W. Reenstra Novel adenoviral vectors coding for GFP-tagged wtCFTR and DF508-CFTR: characterization of expression and electrophysiological properties in A549 cells Received: 24 June 2004 / Accepted: 29 July 2004 / Published online: 28 September 2004 Ó Springer-Verlag 2004 Abstract E1/E3-deleted adenoviral vectors expressing an N-terminal green fluorescent protein (GFP) reporter gene fused to either wtCFTR (H5.040CMVEGFP-wtCFTR) or DF508-CFTR (H5.040CMVEGFP-DF508CFTR) were generated. To characterize the expression and activity, A549 cells were infected with vectors expressing GFP-tagged and non-tagged forms of CFTR and DF508CFTR. CFTR activity was assayed in cell-attached and excised patches. For H5.040CMVEGFP-wtCFTR, forskolin-dependent outward current was observed in cell-attached patches from 56 of 67 GFP-positive cells. Single-channel conductances, open probability, mean open and mean closed time values for GFP-CFTR and CFTR were not significantly different. After excision, GFP-CFTR activity required ATP and exhibited a linear I-V relationship. For H5.040CMVEGFP-DF508CFTR, media were supplemented with 5 mM butyrate 16 h after infection. Forskolin-dependent outward current was observed in cell-attached patches from 21 of 30 butyrate- treated GFP-positive cells and 0 of 8 GFP-positive cells without butyrate. Single-channel conductances, open probability, mean open and mean closed time values for GFP-DF508CFTR and DF508CFTR were not signifi- cantly different. However, the increase in open proba- bility with genistein was significantly smaller for GFP- DF508CFTR than for DF508CFTR. In excised patches, GFP-DF508CFTR activity required ATP and exhibited a linear I-V relationship. Despite the consistent detection of GFP-CFTR and GFP-DF508CFTR channels in the plasma membrane by patch clamping, GFP fluorescence was observed only in intracellular regions and was not altered by butyrate. The data show that high levels of functional GFP-tagged CFTR channels can be expressed with these adenoviral vector constructs. Keywords A549 cells Channel conductance Channel kinetics Genistein Viral transfection Introduction Recently, a large number of studies on CFTR function and membrane trafficking have made use of CFTR constructs with an N-terminal green fluorescent pro- tein (GFP) tag (GFP-CFTR). In general, investigators have not reported measurements of channel activity for GFP-CFTR or have demonstrated channel activity by whole-cell or short-circuit current measurements [3, 10, 12, 22, 27]. In no case has it been established that GFP-tagged forms of CFTR have single-channel properties like those of the untagged forms, apart from the pore properties [20, 25]. The choice of an N- terminal tag is reasonable, because it is less likely to interfere with membrane-targeting information thought to be encoded in the C terminal [24, 26]. However, as interactions between the N terminal and the R domain have been shown to alter CFTR activity [6, 7, 28], there is concern that the presence of an N-terminal GFP might alter channel kinetics. This may be even greater concern with the most common disease-asso- ciated mutant of CFTR, DF508, where GFP-tagged constructs have been used to evaluate procedures that alter the processing and functional activity of DF508CFTR [13, 22, 27, 34]. One of the critical factors in studying the electro- physiology of the CFTR gene product is to achieve efficient gene transduction in the target cells. We chose recombinant adenovirus as the gene transfer vehicle. Adenovirus-based vectors are well known for their H. Vais G.-P. Gao M. Yang P. Tran J.-P. Louboutin S. Somanathan J. M. Wilson W. W. Reenstra (&) Division of Medical Genetics, School of Medicine, University of Pennsylvania, Philadelphia, PA 19104, USA E-mail: [email protected] Fax: +1-215-5737414 Pflugers Arch – Eur J Physiol (2004) 449: 278–287 DOI 10.1007/s00424-004-1331-0
Transcript of Novel adenoviral vectors coding for GFP-tagged wtCFTR and ΔF508-CFTR: characterization of...

ION CHANNELS, TRANSPORTERS
Horia Vais Æ Guang-Ping Gao Æ Michael Yang
Phoi Tran Æ Jean-Pierre Louboutin
Suryanarayan Somanathan Æ James M. Wilson
William W. Reenstra
Novel adenoviral vectors coding for GFP-tagged wtCFTRand DF508-CFTR: characterization of expressionand electrophysiological properties in A549 cells
Received: 24 June 2004 / Accepted: 29 July 2004 / Published online: 28 September 2004� Springer-Verlag 2004
Abstract E1/E3-deleted adenoviral vectors expressing anN-terminal green fluorescent protein (GFP) reporter genefused to either wtCFTR (H5.040CMVEGFP-wtCFTR)or DF508-CFTR (H5.040CMVEGFP-DF508CFTR)were generated. To characterize the expression andactivity, A549 cells were infected with vectors expressingGFP-tagged and non-tagged forms of CFTR andDF508CFTR. CFTRactivity was assayed in cell-attachedand excised patches. For H5.040CMVEGFP-wtCFTR,forskolin-dependent outward current was observed incell-attached patches from 56 of 67 GFP-positive cells.Single-channel conductances, open probability, meanopen and mean closed time values for GFP-CFTR andCFTR were not significantly different. After excision,GFP-CFTR activity required ATP and exhibited a linearI-V relationship. For H5.040CMVEGFP-DF508CFTR,media were supplemented with 5 mM butyrate 16 h afterinfection. Forskolin-dependent outward current wasobserved in cell-attached patches from 21 of 30 butyrate-treated GFP-positive cells and 0 of 8 GFP-positive cellswithout butyrate. Single-channel conductances, openprobability, mean open and mean closed time values forGFP-DF508CFTR and DF508CFTR were not signifi-cantly different. However, the increase in open proba-bility with genistein was significantly smaller for GFP-DF508CFTR than for DF508CFTR. In excised patches,GFP-DF508CFTR activity required ATP and exhibited alinear I-V relationship. Despite the consistent detectionof GFP-CFTR and GFP-DF508CFTR channels in theplasma membrane by patch clamping, GFP fluorescencewas observed only in intracellular regions and was not
altered by butyrate. The data show that high levels offunctional GFP-tagged CFTR channels can be expressedwith these adenoviral vector constructs.
Keywords A549 cells Æ Channel conductance Æ Channelkinetics Æ Genistein Æ Viral transfection
Introduction
Recently, a large number of studies on CFTR functionand membrane trafficking have made use of CFTRconstructs with an N-terminal green fluorescent pro-tein (GFP) tag (GFP-CFTR). In general, investigatorshave not reported measurements of channel activityfor GFP-CFTR or have demonstrated channel activityby whole-cell or short-circuit current measurements [3,10, 12, 22, 27]. In no case has it been established thatGFP-tagged forms of CFTR have single-channelproperties like those of the untagged forms, apartfrom the pore properties [20, 25]. The choice of an N-terminal tag is reasonable, because it is less likely tointerfere with membrane-targeting information thoughtto be encoded in the C terminal [24, 26]. However, asinteractions between the N terminal and the R domainhave been shown to alter CFTR activity [6, 7, 28],there is concern that the presence of an N-terminalGFP might alter channel kinetics. This may be evengreater concern with the most common disease-asso-ciated mutant of CFTR, DF508, where GFP-taggedconstructs have been used to evaluate procedures thatalter the processing and functional activity ofDF508CFTR [13, 22, 27, 34].
One of the critical factors in studying the electro-physiology of the CFTR gene product is to achieveefficient gene transduction in the target cells. We choserecombinant adenovirus as the gene transfer vehicle.Adenovirus-based vectors are well known for their
H. Vais Æ G.-P. Gao Æ M. Yang Æ P. Tran Æ J.-P. LouboutinS. Somanathan Æ J. M. Wilson Æ W. W. Reenstra (&)Division of Medical Genetics, School of Medicine,University of Pennsylvania,Philadelphia, PA 19104, USAE-mail: [email protected]: +1-215-5737414
Pflugers Arch – Eur J Physiol (2004) 449: 278–287DOI 10.1007/s00424-004-1331-0

broad tropism and ability to express high levels oftransgene products in infected cells [1, 11, 18, 30, 32].Adenoviral vectors have become a frequently usedmethod for CFTR expression in cell lines and inanimal studies, resulting in high levels of CFTRexpression. In this study, minigene cassettes expressingGFP-tagged CFTR wild-type and DF508-CFTR fusionproteins were cloned into E1/E3-deleted adenovirus.A549 cells, a small-cell lung carcinoma cell line, weretransfected with vector and the single-channel activityof CFTR constructs assayed in cell-attached patches.Our studies suggest that the single-channel propertiesof GFP-tagged CFTRs are not appreciably altered bythe presence of an N-terminal GFP.
Materials and methods
Construction AdCMVCFTR.GFP andAdCMVCFTRD508.GFP
The construction of adenovirus vectors expressingEGFP-CFTR fusion proteins was accomplished inthree steps. First, pShuttle-EGFP was generated byisolating the EGFP gene in pIRES2-EGFP (Clontech),by digestion with BstXI and Not1 and by inserting itinto pShuttle (Clontech) between the Bsa1 and Nhelsites by blunt-end ligation. Two fragments were thengenerated from pAd-CB-CFTR [33]. The first requiredPCR amplification of a region that contained a BsrG1site 24 bp upstream from the CFTR start codon andthe only XbaI site in the CFTR gene, followed bydigestion with BsrG1 and XbaI. The second requiredisolating a fragment of the CFTR gene from the XbaIsite to an EcoRV site 83 bp beyond the CFTR stopcodon. These two fragments were then stitched intopShuttle-EGFP between a BsrGI site at the N-terminalof EGFP and the PmeI upstream of the BGHpoly(A)+ site to produce pShuttle-EGFP-CFTR. In ananalogous way, pShuttle-EGFP-F508CFTR was cre-ated with an XbaI-to-EcoRV fragment from pAd-CB-F508CFTR. The resulting constructs contain an ORFthat codes for EGFP lacking the N-terminal K, aneight-amino acid spacer REPSARET, and the completeCFTR (or DF508CFTR) protein. The EGFP-CFTRand EGFP-F508CFTR ORFs were cleaved from thepShuttle vector with I-CeuI and PI-SceI and insertedinto pAdenoX backbones at these sites. As the pAde-noX backbone contained a GFP construct between theCeuI and PI-SceI sites, transfectans with the CFTRinsert were selected with a green/white selection [9].These were then used to transfect 293 cells and gen-erate the E1/E3-deleted adenoviral vectors Ad-CMVCFTR.GFP and AdCMVCFTRDF508.GFP.Vectors were expanded in 293 cells and purified byCsCl density gradient centrifugation. The infectivity ofviral vectors was determined on 293 cells by plaqueassay as described previously [8].
Cell culture and transformation
A549 cells were grown in F-12 K nutrient mixture(Kaighn’s medium, Cellgro-Mediatech, Herndon, Va.,USA,), supplemented with 10% fetal bovine serum(Atlanta Biologicals, Norcross, Ga., USA) and 1%penicillin/streptomycin (10,000 IU/ml, Cellgro-Media-tech). Cells were maintained in 150-mm plates. Mediawere changed every 2 days. Cells were passaged at 90%to 95% confluence and split 1 to 10. One day prior toinfection, cells were plated onto glass coverslips (8 mm ·8 mm) in a 60-mm plate. Approximately 107 cells wereinfected with AdCMVCFTR.GFP, AdCBCFTR, Ad-CMVCFTRD508.GFP, or AdCBCFTRD508 [9] at anmultiplicity of infection [(MOI) taken as tenfold thenumber of viral particles per cell] of 100 in 4 ml ofmedium. Cells were used for patch clamping 1–3 dayspost-infection, with 2 days being optimal. To increaseplasma membrane expression of DF508CFTR andDF508CFTR-GFP, 16-h-postinfection media were re-placed with fresh media, supplemented with 5 mM bu-tyric acid [15]. After 2 days, approximately 60% of cellsinfected with AdCMVCFTR.GFP and 50% of cells in-fected with AdCMVCFTRD508.GFP expressed EGFPas assessed by fluorescence. While 5 mM butyrate hadno effect on the fluorescence of cells infected with Ad-CMVCFTRD508.GFP, it was essential for the detectionof DF508CFTR-GFP and DF508CFTR channel activity.
Electrophysiology
Pipettes were pulled from PG52151–4 glass capillaries(World Precision Instruments, Sarasota, Fla., USA),using a vertical puller (PP-830, Narishige InstrumentsLaboratory, Tokyo). They had resistances of 4–6 MWwhen filled with pipette solution: 140 mM NMDG-Cl,5 mM CaCl2, 2 mM MgCl2, and 10 mM HEPES (pH7.4 adjusted with NMDG). For cell-attached measure-ments, the bath solution was 145 mM NaCl, 5 mMKCl,2 mM CaCl2, 2 mM MgCl2, 5 mM glucose, and 5 mMHEPES (pH 7.4, adjusted with NaOH). For inside-outpatches, the bath solution was 145 mM NaCl, 2 mMMgCl2, 5 mM EGTA, and 5 mM HEPES (pH 7.4, ad-justed with NaOH).
Data acquisition
Cell-attached and excised inside-out patch currentswere recorded at room temperature (22�C), using anEPC-9 patch-clamp amplifier controlled by the associ-ated PULSE software (Heka Elektronik, Lambrecht,Germany). For cell-attached patches, the apparentmembrane potential (bath minus pipette potential) was50 mV. For I-V plots, single current data were obtainedfrom inside-out patches clamped at membrane potentialsbetween ±50 mV. Separate records were obtained ateach value of membrane potential and single-channel
279

currents measured off-line. Data were sampled at 200 Hz,filtered at 100 Hz with a built-in three-pole Bessel filter,and stored directly onto a PC hard disk for analysis.
Data analysis
Data were digitally filtered at 10 Hz and analyzed usingIGOR Pro (WaveMetrics, Lake Oswego, Ore., USA)software. Open probability, Po, was determined from thevariance in mean current, r2, with the following rela-tionship:
Po ¼ 1� r2=I � i� �
ð1Þ
where I is the mean current and i is the single-channelcurrent [5, 31]. I and r2 were determined in records of15 min or longer, where I in the presence of forskolinwas constant. i was determined from amplitude histo-grams. The mean open time, so, was calculated from theratio of mean current to single-channel current, I/i, andthe number of channel openings per unit time, n/T, withthe formula:
so ¼ ðI=iÞ=ðn=T Þ ð2Þ
where T is the length of the record (>600 s) and n is thetotal number of openings in the record [23]. Mean closedtime, sc, was determined from so and Po.
Immunocytochemistry
Cells were fixed in acetone:methanol (50:50) for 5 min.Blocking was performed by a 1-h incubation with 10%goat serum. Sections were incubated for 1 h at roomtemperature with the primary antibody, mouse mono-clonal anti-CFTR (C terminus) (Research Diagnostics,Flanders, N.J., USA) at a 1:20 dilution, then washedthree times with PBS, and incubated with 1:100 dilutedrhodamine-labeled goat anti-mouse IgG (Sigma, StLouis, Mo., USA). Mounting medium containing DAPIwas used to visualize the nuclei. Specimens were exam-ined with a fluorescence microscope, Nikon Microphot-FXA, using FITC, rhodamine, and triple filters tovisualize GFP, anti-CFTR immunostaining, and the twoprevious ones plus DAPI staining of the nuclei, respec-tively. Negative controls, consisting in the omission ofprimary antibody and substitution of the primary anti-body by an isotype-matched, non-immune controlantibody were performed.
Reagents
Forskolin (Calbiochem, La Jolla, Calif., USA) andgenistein (Alexis, San Diego, Calif., USA) were kept asstock solutions (10 mM and 50 mM, respectively) inDMSO at �20�C. All other chemicals were from Sigma(St.Louis, MO).
Statistics
Calculated values are given as means±SEM. Compari-sons among data were made using the Student’s t-testwith P<0.05 considered significant.
Results
To compare the kinetic properties of GFP-CFTR withCFTR as well as GFP-DF508CFTR with DF508CFTR,A549 cells were infected with one of four geneticallyengineered adenoviruses, AdCMVCFTR.GFP, Ad-CBCFTR, AdCMVCFTRD508.GFP, or AdCBCFTRD508, at an MOI of 100. Under the conditions describedin ‘‘Materials and methods,’’ patches with CFTR chan-nels were observed from 1 to 3 days post-infection,with 2 days being optimal. In cell-attached patchesfrom non-infected A549 cells, forskolin-dependent chlo-ride channels were not observed. Infection withAdCMVCFTR.GFP and AdCMVCFTRD508.GFP re-sulted in GFP fluorescence in 50% to 70% (GFP-CFTR)and 40% to 60% (GFP-DF508CFTR) of cells. As pre-sented in Fig. 1a–c for AdCMVCFTR.GFP (GFP-CFTR) and in Fig. 1d–i for AdCMVCFTRD508.GFP (GFP-DF508CFTR), transduced cells mainlyshowed a diffuse intracellular localization of the ex-pressed proteins. As described in the ‘‘Materials andmethods,’’ functional GFP-DF508CFTR channels re-quired the presence of butyrate in the culture medium.However, as shown in Fig. 1d, e, g, h, there was no dis-cernable change in GFP-DF508CFTR distribution whenassayed by GFP fluorescence (Fig. 1e, h) or immunore-activity (Fig. 1d, g). As immunoreactivity for CFTR andGFP fluorescence co-localize (Fig. 1c, f, i), the intracel-lular GFP fluorescence is not generated during the deg-radation of CFTR or DF508-CFTR. The predominantintracellular localization was not due to the GFP tag,because antibodies to the C terminal of CFTR generatedthe same fluorescence pattern in cells infected with vec-tors for CFTR, AdCBCFTR, and DF508CFTR, Ad-CBCFTRD508 (data not shown). No immunostainingfor CFTR was seen when cells were incubated withoutthe primary antibody or when the primary antibody wassubstituted by an isotype-matched non-immune controlantibody (not shown).
Comparison of GFP-CFTR and CFTR channel kinetics
Cell-attached patches were formed in the absence ofagonist. Once the absence of agonist-independentchannel activity was confirmed, cells were perfused witha bathing solution containing 1 lM forskolin. For Ad-CMVCFTR.GFP-infected cells, an outward current wasseen in cell-attached patches from 56 of 67 GFP-positivecells. For AdCBCFTR-infected cells, channel activitywas observed in 24 of 28 cells examined. Since
280

morphological clues were used to identify AdCBCFTR-infected cells, the high level of patches with CFTR forAdCBCFTR-infected cells is not inconsistent with thelevel of expression and number of successful patchesobtained with cells expressing GFP-CFTR. Channelactivity for both GFP-CFTR and CFTR was forskolindependent (data not shown). At a pipette potential of�50 mV, the mean current was 13±4 pA (n=11) butranged between 0.1 pA and 200 pA for GFP-CFTR andwas similar (up to 200 pA) for CFTR. Figure 2a shows arecord from a cell-attached patch in which up to sevenGFP-CFTR channels were active. From records like theone in Fig. 2a, with less than 20 active channels, andrecordings of 10 min or longer without significantchanges in I (measured in 2-min windows), Po wasestimated from the variance in I (Eq. 1). As shown inTable 1, for GFP-CFTR, a Po value of 0.21±0.03(n=6) was obtained; the same analysis for cellsexpressing CFTR gave a Po value of 0.20±0.02 (n=5).
From cell-attached patch records that showed <10active channels, n was determined and mean so wascalculated (Eq. 2). As shown in Fig. 2 and Table 1,mean so values were 1.5±0.3 s (n=4) for GFP-CFTRand 1.5±0.3 s (n=5) for CFTR. Mean sc values werecalculated from values of Po and mean so. For GFP-CFTR and CFTR, mean sc values were 4.9±1.5 s (n=3)and 6.2±1.3 s (n=5), respectively. Neither the open northe sc values were significantly different for the twoforms of CFTR.
To demonstrate the ATP-dependence of GFP-CFTR,patches containing GFP-CFTR channels were excised inATP-free solutions. A rapid loss of channel activity wasobserved (Fig. 3a). However, channel activity was re-stored when 1 mM ATP was added to the bath. Thesubsequent removal and readdition of ATP causedchannel activity to cease and to recover, respectively(Fig. 3b). Our results suggest that, in the presence ofATP, channel rundown in A549 cells is relatively slow.This allowed us to measure single-channel conductancewithout having to add protein kinase A to the bath.Figure 4 shows an I-V plot for GFP-CFTR and CFTR.Like CFTR, GFP-CFTR currents were ohmic betweenmembrane potentials ±50 mV, and single-channelconductances, g, for GFP-CFTR and CFTR were8.2±0.2 pA and 8.4±0.3 pA, respectively.
Comparison of GFP-DF508CFTR and DF508CFTRchannel kinetics
A459 cells were infected with AdCMVCFTRD508.GFPand AdCBCFTRD508. To increase GFP-DF508CFTRand DF508CFTR expression in the plasma membrane,at 16 h post-infection, the vector was removed, and themedia were supplemented with 5 mM Na-butyrate.After cell-attached patch formation and upon noticingthe absence of agonist-independent channel activity,channel activation was induced by adding 10 lM fors-
Fig. 1 Localization of green fluorescent protein (GFP)-CFTRand GFP-DF508CFTR. A549 cells were infected with Ad-CMVCFTR.GFP (a–c) or AdCMVCFTRD508.GFP (d–i). Cellswere fixed and treated with an anti-CFTR (C terminal) antibodyfollowed by a tetramethy-rhodamine-conjugated secondary anti-body. Cells shown in g–i were cultured in the presence of 5 mM
butyrate for 24 h prior to processing. a, d, and g show CFTRimmunostaining (red) while b, e, and h show the correspondingGFP fluorescence (green). In c, f, and i, a triple overlay of CFTR,GFP, and DAPI staining (blue) of the nucleus is shown. Scale bar(bottom right) is 15 lm
281

kolin into the bath. For A549 cells infected with Ad-CMVCFTRD508.GFP and exposed to media supple-mented with butyrate, forskolin-dependent channelactivity was observed in 32 of 55 patches from GFP-positive cells. In cells that were not kept in media sup-plemented with butyrate, channels were observed in
patches from 0 of 8 GFP-positive cells. For A549 cellsinfected with AdCBCFTRD508, where media weresupplemented with butyrate, channel activity was ob-served in 13 of 25 patches.
Upon activation with 10 lM forskolin, cell-attachedpatches expressing GFP-DF508-CFTR showed infre-quent channel openings (Fig. 5a). The addition of 50 lMgenistein increased channel activity. Due to the largeerrors in values of Po obtained from variance analysiswhen Po is very small, the Po for GFP-DF508CFTR was
Table 1 Gating and conductance properties of green fluorescent protein (GFP)-tagged CFTR constructs
GFP-CFTR CFTR GFP-DF508 DF508
Patches with CFTRa 84% (56/67)b 86% (24/28) 56% (32/55)b,c 52% (13/25)c
Ic, pA (n) 0.27±0.03 (7) 0.29±0.03 (5) 0.27±0.06 (4) 0.24±0.01 (4)Po (n) 0.21±0.03 (6) 0.20±0.02 (5) 0.011±0.004 (5) 0.005±0.002 (4)Po (+ genistein) (n) - - 0.25±0.07e (5) 0.60±0.07e,f (4)so, s (n) 1.5±0.3 (4) 1.5±0.3 (5) 0.46±0.14 (5) 0.41±0.09 (4)so, s (+ genistein) (n) - - 2.8±0.8e (5) 6.8±1.0e,f (5)sc, s (n) 4.9±1.5 (3) 6.2±1.3 (5) 67±19 (5) 154±58 (4)sc, s (+ genistein) (n) - - 11±4f (5) 6.0±3.0f (5)g, pS (n) 8.2±0.2 (7) 8.4±0.3 (7) 8.2±0.2 (7) 8.2±0.3 (6)
Fig. 2 Cell-attached recordings from A549 cells expressed GFP-CFTR (a) and CFTR (b). Cells were infected with Ad-CMVCFTR.GFP (a) or AdCBCFTR (b) and patched after 48 hto 72 h. Channels were activated with 1 lM forskolin prior to thestart of the traces shown (pipette potential = �50 mV). Traces arefiltered at 10 Hz. Insets are at a tenfold expanded time scale. Closedstate is indicated by the dashed line
Fig. 3 ATP-dependent GFP-CFTR activity in excised patches. Inboth traces, cell-attached patches were formed from A549 cells thathad been infected with AdCMVCFTR.GFP. Channel activity wasstimulated with 1 lM forskolin prior to the start of the trace andrecorded at a holding pipette potential of �50 mV. Prior toexcision, bathing media were changed to a Ca2+-free solution with5 mM EGTA. In a, inside-out patches were excised into ATP freebuffer at the arrow. The inset is an expansion of the indicated partof the trace. In b, excision was made into bathing solutioncontaining 1.0 mM ATP. ATP was removed and readmitted to thebath, as indicated by the bars
aI Mean current, Po open probability, so mean open time, sc, meanclosed time,g single-channel conductancebFor the GFP-tagged constructs, only green fluorescent cells werepatched
cCells treated with 5 mM butyrate for 24 f to 48 hdCell-attached patches with Vmem=50 mVeSignificantly different (P £ 0.05) than value without genisteinfSignificantly different (P £ 0.05) than value for GFP-DF508CFTR
282

first calculated from r2 in the presence of genistein:Po=0.25±0.07 (n=5). Based on the assumption thatgenistein does not increase the number of channels, Po
with forskolin alone, 0.011±0.004, was calculated fromthe change in I. A similar procedure was used to analyzeDF508CFTR channels (Fig. 5b); Po was 0.005±0.002(n=5) with forskolin and 0.60±0.07 (n=5) with fors-kolin plus genistein. Our assumption that genistein doesnot increase the number of active DF508CFTR channelsis consistent with previous studies [16, 17]. However, theobserved increases in Po, 37±13-fold for GFP-DF508CFTR and 170±50-fold for DF508CFTR, aregreater that those we have previously observed in stablytransfected NIH-3T3 cells [16]. As (1) the data with asingle active channel in the patch (Fig. 5B) had a geni-stein-dependent increase in mean current of 56-fold, and(2) an examination of all data suggested that the foldincreases were multiples of this value, we reanalyzed thedata, allowing the number of active channels in the ab-sence of genistein to differ by integral values less than themaximum number of channels observed in the presenceof genistein. This value was selected so that the variancein the fold increase in I by genistein was minimized.This procedure gave Po values for GFP-DF508CFTRand DF508CFTR, in the absence of genistein, of0.013±0.003 and 0.011±0.001, respectively. Fold in-creases in Po with genistein were calculated to be 13±0.6for GFP-DF508CFTR and 54±6 for DF508CFTR.Irrespective of the procedure used, the forskolin-depen-dent Po of GFP-DF508CFTR and DF508CFTR were notsignificantly different, and only in the presence of geni-stein and forskolin was the Po of DF508CFTR signifi-cantly greater than that for GFP-DF508CFTR.
For DF508CFTR and GFP-DF508CFTR, channelkinetics in the presence and absence of genistein were
also investigated. Mean so values were determined asdescribed for the wild-type CFTR and therefore areindependent of the value of Po. As shown in Table 1, inthe absence of genistein, mean so values of 0.41±0.09 s(n=4) and 0.46±0.14 s (n=5) were observed forDF508CFTR and GFP-DF508CFTR, respectively. sovalues were increased by genistein to 6.8±1.0 s (n=5)and 2.8±0.8 s (n=5) for DF508CFTR and GFP-DF508CFTR, respectively. Mean sc values were calcu-lated from Po. When mean sc values were calculated withthe assumption that the number of channels in thepresence and absence of genistein was the same, mean scvalues in the absence of genistein of 154±58 s (n=4) and67±19 s (n=5) were observed for DF508CFTR andGFP-DF508CFTR, respectively; these values are notsignificantly different (P=0.16). sc values in the presenceof genistein were 6±3 s (n=5) and 11±4 s (n=5) forDF508CFTR and GFP-DF508CFTR, respectively. Thedata demonstrate that the single-channel kinetics ofGFP-DF508CFTR and DF508CFTR are similar withrespect to activation by forskolin and potentiation bygenistein. The only significant difference is the increasedPo in the presence of genistein, reflecting changes inboth mean so and mean sc, for DF508CFTR. In theabsence of genistein, single-channel kinetics of DF508-CFTR and GFP-DF508CFTR showed no significantdifferences. Our data also show that genistein increasesthe forskolin-dependent Po of DF508CFTR (and
Fig. 4 Single-channel I-V plots from A549 cells infected withAdCMVCFTR.GFP or AdCBCFTR. Channels were activated inthe initial cell-attached configuration with 1 lM forskolin. Inside-out patches, containing GFP-CFTR (d) and CFTR (s) channels,were generated, and separate records were obtained at eachmembrane potential indicated in the graph. The insets showrepresentative data from which single-channel currents weremeasured. Closed state is indicated by the dashed line
Fig. 5 Cell-attached recordings from A549 cells expressed GFP-DF508CFTR (a) and DF508CFTR (b). Cells were infected withAdCMVCFTRD508.GFP (a) or AdCBCFTRD508 (b) and patchedafter 48 h to 72 h. Growth media were supplemented with 5 mMbutyrate 16 h after infection. Channels were activated with 10 lMforskolin prior to the start of the traces and with 50 lM genisteinwhere indicated. Patches were held at a pipette potential of�50 mV throughout the record
283
GFP-DF508CFTR) by increasing the mean so anddecreasing the mean sc.
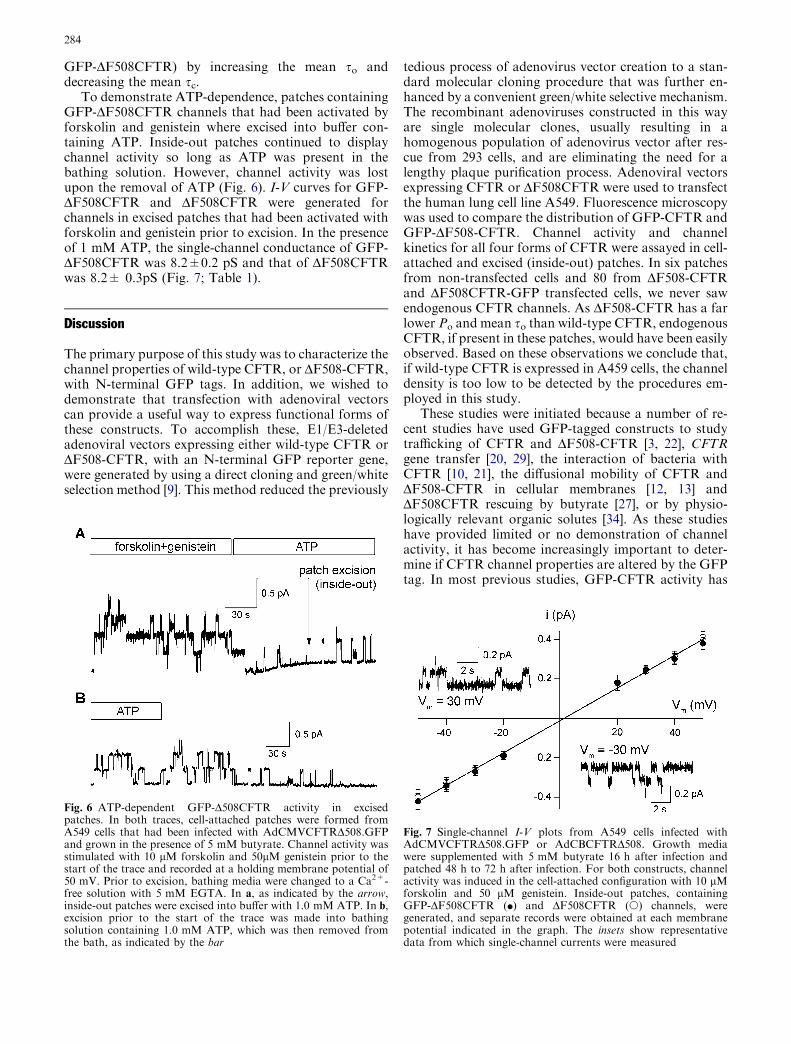
To demonstrate ATP-dependence, patches containingGFP-DF508CFTR channels that had been activated byforskolin and genistein where excised into buffer con-taining ATP. Inside-out patches continued to displaychannel activity so long as ATP was present in thebathing solution. However, channel activity was lostupon the removal of ATP (Fig. 6). I-V curves for GFP-DF508CFTR and DF508CFTR were generated forchannels in excised patches that had been activated withforskolin and genistein prior to excision. In the presenceof 1 mM ATP, the single-channel conductance of GFP-DF508CFTR was 8.2±0.2 pS and that of DF508CFTRwas 8.2± 0.3pS (Fig. 7; Table 1).
Discussion
The primary purpose of this study was to characterize thechannel properties of wild-type CFTR, or DF508-CFTR,with N-terminal GFP tags. In addition, we wished todemonstrate that transfection with adenoviral vectorscan provide a useful way to express functional forms ofthese constructs. To accomplish these, E1/E3-deletedadenoviral vectors expressing either wild-type CFTR orDF508-CFTR, with an N-terminal GFP reporter gene,were generated by using a direct cloning and green/whiteselection method [9]. This method reduced the previously
tedious process of adenovirus vector creation to a stan-dard molecular cloning procedure that was further en-hanced by a convenient green/white selective mechanism.The recombinant adenoviruses constructed in this wayare single molecular clones, usually resulting in ahomogenous population of adenovirus vector after res-cue from 293 cells, and are eliminating the need for alengthy plaque purification process. Adenoviral vectorsexpressing CFTR or DF508CFTR were used to transfectthe human lung cell line A549. Fluorescence microscopywas used to compare the distribution of GFP-CFTR andGFP-DF508-CFTR. Channel activity and channelkinetics for all four forms of CFTR were assayed in cell-attached and excised (inside-out) patches. In six patchesfrom non-transfected cells and 80 from DF508-CFTRand DF508CFTR-GFP transfected cells, we never sawendogenous CFTR channels. As DF508-CFTR has a farlower Po and mean so than wild-type CFTR, endogenousCFTR, if present in these patches, would have been easilyobserved. Based on these observations we conclude that,if wild-type CFTR is expressed in A459 cells, the channeldensity is too low to be detected by the procedures em-ployed in this study.
These studies were initiated because a number of re-cent studies have used GFP-tagged constructs to studytrafficking of CFTR and DF508-CFTR [3, 22], CFTRgene transfer [20, 29], the interaction of bacteria withCFTR [10, 21], the diffusional mobility of CFTR andDF508-CFTR in cellular membranes [12, 13] andDF508CFTR rescuing by butyrate [27], or by physio-logically relevant organic solutes [34]. As these studieshave provided limited or no demonstration of channelactivity, it has become increasingly important to deter-mine if CFTR channel properties are altered by the GFPtag. In most previous studies, GFP-CFTR activity has
Fig. 6 ATP-dependent GFP-D508CFTR activity in excisedpatches. In both traces, cell-attached patches were formed fromA549 cells that had been infected with AdCMVCFTRD508.GFPand grown in the presence of 5 mM butyrate. Channel activity wasstimulated with 10 lM forskolin and 50lM genistein prior to thestart of the trace and recorded at a holding membrane potential of50 mV. Prior to excision, bathing media were changed to a Ca2+-free solution with 5 mM EGTA. In a, as indicated by the arrow,inside-out patches were excised into buffer with 1.0 mM ATP. In b,excision prior to the start of the trace was made into bathingsolution containing 1.0 mM ATP, which was then removed fromthe bath, as indicated by the bar
Fig. 7 Single-channel I-V plots from A549 cells infected withAdCMVCFTRD508.GFP or AdCBCFTRD508. Growth mediawere supplemented with 5 mM butyrate 16 h after infection andpatched 48 h to 72 h after infection. For both constructs, channelactivity was induced in the cell-attached configuration with 10 lMforskolin and 50 lM genistein. Inside-out patches, containingGFP-DF508CFTR (d) and DF508CFTR (s) channels, weregenerated, and separate records were obtained at each membranepotential indicated in the graph. The insets show representativedata from which single-channel currents were measured
284

either not been measured, or measurements have beenlimited to short circuit currents [22, 27, 29] or whole-cellcurrents and their inhibition or activation by pharma-cological agents [25]. In one study [25], the channelconductance of GFP-CFTR was determined. In nocases, however, have Po values or mean so and sc valuesof GFP-tagged constructs been determined. One reasonwhy the channel kinetics of GFP-tagged constructsmight be altered from those of the non-tagged forms isthe observation that the binding of the N terminus ofCFTR to the R domain regulates CFTR activity [6, 7,28]. In addition, as adenoviral vectors have become amore widely used method for in vitro gene delivery,studies are needed to demonstrate that the properties ofCFTR in virally transformed cells are comparable tothose observed with other means of transformation or incells that naturally express CFTR.
We must point out that there are several potentialproblems with the kinetic parameters obtained in thisstudy, which place limits on the quantitative and com-parative conclusions that we have drawn. Values of Po
were calculated from I, r2, and i. This method requiresthat i can be clearly identified in the traces and that I beconstant over the time course of the experiment. Apractical consequence of this is that our analysis is re-stricted to patches having no more than 15 channelsactive at any time. In addition, since Po is calculatedfrom the difference between 1 and the experimentalparameter, r2/I*i, values of Po that are close to 0 or 1 arelikely to be inaccurate. For this reason, Po values forDF508CFTR and GFP-DF508CFTR were first calcu-lated in the presence of genistein. Po values in the ab-sence of genistein were then calculated from the ratio ofthe mean currents in the presence and absence of geni-stein. This requires the experimentally verifiable fact thatgenistein does not change i and the assumption that thenumber of active channels is not altered by genistein.This assumption has been supported by several previousstudies [16, 17]. In this study however, the large andvariable increases in mean current with genistein sug-gested that, in the presence of genistein, there were moreactive channels than before genistein addition. Accord-ingly, values for Po and mean sc values were also cal-culated without the assumption that the number ofchannels is unaffected by genistein.
Values of mean so were determined from I, i, and thenumber of openings per time unit. This required theanalysis of current records where all openings wereclearly discernable which meant, in practice, that nomore than six channels were simultaneously open at anytime. The procedure does not permit open-time histo-grams to be generated and is therefore unable to deter-mine if there is more than one so constant; nevertheless,values of so determined in this way are not dependent onPo. However, as values of mean sc were calculated fromPo and so, these values are dependent on Po and, forDF508CFTR constructs in the absence of genistein,values for mean sc values were calculated with bothvalues of Po. Independent of any errors that may have
been introduced by our methods for calculating kineticparameters, since the same procedures were applied toboth CFTR and GFP-CFTR (and DF508CFTR andGFP-DF508CFTR), our conclusions about the relativeeffects of an N-terminal GFP tag on channel kinetics arelikely to be correct.
Our data confirm the observation of others [19, 20, 25]that GFP-CFTR has chloride channel activity. In cell-attached patches, this activity is only observed in thepresence of activators of PKA. Moreover, our studiesalso demonstrate that Po, so, sc, and i of GFP-CFTR andCFTR are not significantly altered from the corre-sponding values of CFTR. Our data demonstrate thatGFP-CFTR channel activity requires ATP and, based onthe similarity of our mean values for current in cell-at-tached patches, it appears that an N-terminus GFP tagdoes not alter CFTR trafficking in A549 cells. For GFP-DF508CFTR, our studies demonstrate that trafficking tothe plasma membrane does not occur in the absence ofbutyrate. As previous studies have demonstrated thatgenistein increases the channel activity of DF508CFTRby as much as 20-fold [16], genistein activation ofDF508CFTR and GFP-DF508CFTR was compared. Inthe presence of forskolin, infrequent channel open-ings—characteristic for the DF508 mutant [4, 14]—wereseen with both DF508CFTR andGFP-DF508CFTR, andno statistically significant difference in Po was observed.For both DF508CFTR and GFP-DF508CFTR, theaddition of genistein increased Po. However, in thepresence of genistein, the Po of DF508CFTR was signif-icantly greater than that of GFP-DF508CFTR; we haveno explanation for this effect. Analysis of so and sc re-vealed that genistein increases the Po of DF508CFTR andGFP-DF508CFTR by increasing so and decreasing sc.This requires that genistein alters the Po of DF508CFTRby binding to both the open and closed states of thechannel. As for the wild-type CFTR, the ATP require-ment for gating and the I-V relationship were unaffectedby the N-terminal GFP tag of DF508CFTR. Thus, in oursystem, the biophysical properties of CFTR andDF508CFTR are not altered by the GFP tag.
In this system, the majority of GFP-tagged CFTR andDF508CFTR were located in the cytoplasmic region ofthe cell. A similar distribution was seen with non-taggedCFTR and DF508CFTR. While the intracellular locali-zation might suggest a construct that was largely in theER or/and Golgi, our images are not of sufficient reso-lution for us to make this conclusion. To demonstratethat GFP fluorescence was due to GFP-CFTR and not tofree GFP that was generated during CFTR degradation,co-localization studies were performed. These demon-strated that CFTR and GFP were present in the samecompartments of the cell. The fluorescence studies alsodemonstrate that, for both GFP-CFTR and GFP-DF508CFTR, the vast majority of protein is in anintracellular location and not in the plasma membrane.While plasma membrane GFP-CFTR was undetectableby fluorescence, possibly because the high level ofintracellular construct would severely impede our ability
285

to detect plasma membrane CFTR, our electrophysio-logical studies clearly demonstrate the presence ofGFP-CFTR in the plasma membrane. They also dem-onstrate a butyrate-dependent redistribution of GFP-DF508CFTR that was not detectable by fluorescencemeasurements. These results emphasize that the detec-tion of plasma membrane CFTR by electrophysiologicalmeans is far more sensitive than by fluorescenttechniques.
Could a GFP-tag interfere with CFTR regulation? Ina number of studies, CFTR activity has been shown tobe decreased by co-expression with syntaxin-1A [2]. Ithas been proposed that an active state of CFTR requiresan interaction of the N terminal of CFTR with the Rdomain and that a less active state occurs when thisinteraction is disrupted. As syntaxin-1A has been shownto bind to an N-terminal peptide from CFTR, it hasbeen postulated that binding of the N terminal to syn-taxin-1A inhibits CFTR activity. If the GFP tag inter-feres with this process, the GFP tag could alter Po, so, orsc. If CFTR activity is regulated by binding of the Nterminal to the R domain, our results suggest that, forboth CFTR and DF508CFTR, this process is notblocked by an N-terminal GFP tag. However, thisprocess could potentially account for the differences ingenistein-dependent increases in so for DF508CFTR andthe GFP-tagged form.
In summary, our results suggest that the presence of aGFP tag on the N terminal of CFTR or DF508CFTRhas minimal effects on channel kinetic, single-channelconductance, or the expression of channels in the plasmamembrane. In contrast, the data clearly show that, whenhigh levels of CFTR are expressed, likely independent ofadenoviral infection, the vast majority of CFTR proteinaccumulates in intracellular vesicles.
Acknowledgements This work was supported by grants from theCystic Fibrosis Foundation (W.W.R.) and (J.M.W.) and R01HL70563 (W.W.R.). James M. Wilson is an inventor on patentslicensed to various commercial entities.
References
1. Berkner KL (1988) Development of adenovirus vectors for theexpression of heterologous genes. Biotechniques 6:616–629
2. Chang SY, Di A, Naren AP, Palfrey HC, Kirk KL, Nelson DJ(2002) Mechanisms of CFTR regulation by syntaxin 1A andPKA. J Cell Sci 115:783–791
3. Cheng J, Wang H, Guggino WB (2004) Modulation of maturecystic fibrosis transmembrane regulator protein by the PDZdomain protein CAL. J Biol Chem 279:1892–1898
4. Dalemans W, Barbry P, Champighny G, Jallat S, Dott K,Dreyer D, Crystal RG, Pavirani A, Lecocq J-P, Lazdunski M(1991) Altered chloride ion channel kinetics associated with theDF508 cystic fibrosis mutation. Nature 354:526–528
5. DeFelice L (1997) Electrical properties of cells—patch clampfor biologists. Plenum, New York, pp 135–136
6. Fu J, Kirk KL (2001) Cysteine substitutions reveal dual func-tions of the amino-terminal tail in cystic fibrosis transmem-brane conductance regulator channel gating. J Biol Chem276:35660–35668
7. Fu J, Ji H-L, Naren AP, Kirk KL (2001) A cluster of negativecharges at the amino terminal tail of CFTR regulates ATP-dependent channel gating. J Physiol (Lond) 536:459–470
8. Gao GP, Yang Y, Wilson JM (1996) Biology of adenovirusvectors with E1 and E4 deletions for liver-directed gene ther-apy. J Virol 70:8934–8943
9. Gao GP, Zhou X, Alvira MR, Tran P, Marsh J, Lynd K, XiaoW, Wilson JM (2003) High throughput creation of re-combinant adenovirus vectors by direct cloning, green-whiteselection and I-SceI-mediated rescue of circular adenovirusplasmids in 293 cells. Gene Ther 10:1926–1930
10. Gerceker AA, Zaidi T, Marks P, Golan DE, Pier GB (2000)Impact of heterogeneity within cultured cells on bacterialinvasion: analysis of Pseudomonas aeruginosa and Salmonellaenterica Serovar Thyphi entry into MDCK cells using a greenfluorescent protein-labelled cystic fibrosis transmembraneconductance regulator receptor. Infect Immun 68:861–870
11. Haddada H, Cordier L, Perricaudet M (1995) Gene therapyusing adenoviral vectors. Curr Top Microbiol Immunol199:297–306
12. Haggie PM, Stanton BA, Verkman AS (2002) Diffusionalmobility of the cystic fibrosis transmembrane conductanceregulator mutant, DF508-CFTR, in the endoplasmic reticulummeasured by photobleaching of GFP-CFTR chimeras. J BiolChem 277:16419–16425
13. Haggie PM, Stanton BA, Verkman AS (2004) Increased dif-fusional mobility of CFTR at the plasma membrane afterdeletion of its C-terminal PDZ binding motif. J Biol Chem279:5494–5500
14. Haws CM, Nepomuceno IB, Krouse ME, Wakelee H, Law T,Xia Y, Nguyen H, Wine JJ (1996) DF508-CFTR channels:kinetics activation by forskolin, and potentiation by xanthines.Am J Physiol 270:C1544–C1555
15. Heda GD, Marino CR (2000) Surface expression of the cysticfibrosis transmembrane conductance regulator mutant Del-taF508 is markedly upregulated by combination treatment withsodium butyrate and low temperature. Biochem Biophys ResCommun 271:659–664
16. Hwang T-C, Wang F, Yang IC, Reenstra WW (1997) Genisteinpotentiates wild-type and delta F508-CFTR channel activity.Am J Physiol 273:C988–C998
17. Illek B, Fisher H, Santos GF, Widdicombe JH, Machen TE,Reenstra WW (1995) cAMP-independent activation of CFTRCl channels by the tyrosine kinase inhibitor genistein. Am JPhysiol 268:C886–C893
18. Kay MA, Glorioso JC, Naldini L (2001) Viral vectors for genetherapy: the art of turning infectious agents into vehicles oftherapeutics. Nat Med 7:33–40
19. Kirk KL (2000) New paradigms of CFTR chloride channelregulation. Cell Mol Life Sci 57:623–634
20. Kollen WJW, Mulberg AE, Wei X, Sugita M, Raghuram V,Wang J, Foskett JK, Glick MC, Scanlin TF (1999) High-effi-ciency transfer of cystic fibrosis transmembrane conductanceregulator cDNA into cystic fibrosis airway cells in cultureusing lactosylated polylysine as a vector. Hum Gene Ther10:615–622
21. Kowalski MP, Pier GB (2004) Localization of cystic fibrosistransmembrane conductance regulator to lipid rafts of epithe-lial cells is required for Pseudomonas aeruginosa-induced cel-lular activation. J Immunol 172:418–425
22. Loffing-Cueni D, Loffing J, Shaw C, Taplin AM, Govindan M,Stanton CR, Stanton BA (2001) Trafficking of GFP-taggedDF508-CFTR to the plasma membrane in a polarized epithelialcell line. Am J Physiol 281:C1889–C1897
23. Mathews CJ, Tabcharani JA, Chang XB, Jensen TJ, RiordanJR, Hanrahan JW (1998) Dibasic protein kinase A sites regu-late bursting rate and nucleotide sensitivity of the transmem-brane conductance regulator chloride channel. J Physiol (Lond)508:365–377
24. Milewski MI, Mickle JE, Forrest JK, Stafford DM, Moyer BD,Cheng J, Guggino WB, Stanton BA, Cutting GR (2001) APDZ-binding motif is essential but not sufficient to localize the
286

C terminus of CFTR to the apical membrane. J Cell Sci114:719–726
25. Moyer BD, Loffing J, Schwiebert EM, Loffing-Cueni D, HalpinPA, Karlson KH, Ismailov II, Guggino WB, Langford GM,Stanton BA (1998) Membrane trafficking of the cystic fibrosisgene product, cystic fibrosis transmembrane conductance reg-ulator, tagged with green fluorescent protein in Madin-Darbycanine kidney cells. J Biol Chem 21:21759–21768
26. Moyer BD, Denton J, Karlson KH, Reynolds D, Wang S,Mickel JE, Milewski M, Cutting GR, Guggino WB, Li M,Stanton BA (1999) A PDZ-interacting domain in CFTR is anapical membrane polarization signal. J Clin Invest 104:1353–1361
27. Moyer BD, Loffing-Cueni D, Loffing J, Reynolds D, StantonBA (1999) Butyrate increases apical membrane CFTR but re-duces chloride secretion in MDCK cells. Am J Physiol277:F271–F276
28. Naren AP, Quick MW, Collawn JF, Nelson DJ, Kirk KL(1998) Syntaxin 1A inhibits CFTR by means of domain-specificprotein-protein interactions. Proc Natl Acad Sci USA95:10972–10977
29. Oceandy D, McMorran B, Schreiber R, Wainwright BJ,Kunzelmann K (2003) GFP-tagged CFTR transgene is func-
tional in the G551D cystic fibrosis mouse colon. J Membr Biol192:159–167
30. Trapnell BC, Gorziglia M (1994) Gene therapy using adeno-viral vectors. Curr Opin Biotechnol 5:617–625
31. Vais H, Zhang R, Reenstra WW (2004) Dibasic phosphoryla-tion sites in the R-domain of CFTR have stimulatory andinhibitory affects on channel activation. Am J Physiol287:C737–C745.DOI 10.1152/ajpcell.00504.2003
32. Wivel NA, Gao GP, Wilson JM (1999) Adenovirus vectors: thedevelopment of human gene therapy. In: The development ofhuman gene therapy. Cold Spring Harbor Laboratory Press,Cold Spring Harbor, pp 87–110
33. Yang Y, Nunes FA, Berencsi K, Gonczol E, Engelhardt JF,Wilson JM (1994) Inactivation of E2a in recombinant adeno-viruses improves the prospect for gene therapy in cystic fibrosis.Nat Genet 7:362–369
34. Zhang XM, Wang XT, Yue H, Leung SW, Thibodeau PH,Thomas PJ, Guggino SE (2003) Organic solutes rescue thefunctional defect in DF508 cystic fibrosis transmembrane con-ductance regulator. J Biol Chem 278:51232–51242
287




















