
Normal development of the outflow tract in the rat
10
Normal Development of the Outflow Tract in the Rat Jing Ya, Maurice J.B. van den Hoff, Piet A.J. de Boer, Sabina Tesink-Taekema, Diego Franco, Antoon F.M. Moorman, Wouter H. Lamers Abstract—The outflow tract (OFT) provides the structural components forming the ventriculoarterial connection. The prevailing concept that this junction “rotates” to acquire its definitive topography also requires a concept of “counterrotation” and is difficult to reconcile with cell-marking studies. Rats between 10 embryonic days (EDs) and 2 postnatal days were stained immunohistochemically and by in situ hybridization. DNA replication was determined by incorporation of bromodeoxyuridine and apoptosis by the annexin V binding and terminal deoxynucleotidyl transferase– mediated dUTP-X nick end labeling (TUNEL) assays. Starting at ED12, cardiomyocytes in the distal (truncal) part of the OFT begin to shed their myocardial phenotype without proceeding into apoptosis, suggesting transdifferentiation. Myocardial regression is most pronounced on the dextroposterior side and continues until after birth, as revealed by the disappearance of the myocardial cuff surrounding the coronary roots and semilunar sinuses and by the establishment of fibrous continuity between mitral and aortic semilunar valves. Fusion of the endocardial ridges of the truncus on late ED13 is accompanied by the organization of a-smooth muscle actin–and nonmuscle myosin heavy chain–positive myofibroblasts into a central whorl and the appearance of the semilunar valve anlagen at their definitive topographical position within the proximal portion of the truncus. After fusion of the proximal (conal) portion of the endocardial ridges, many of the resident myofibroblasts undergo apoptosis and are replaced by cardiomyocytes. The distal myocardial boundary of the OFT is not a stable landmark but moves proximally over the spiraling course of the aortic and pulmonary routes, so that the semilunar valves develop at their definitive topographic position. After septation, the distal boundary of the OFT continues to regress, particularly in its subaortic portion. The myocardializing conus septum, on the other hand, becomes largely incorporated into the right ventricle. These opposite developments account for the pronounced asymmetry of the subaortic and subpulmonary outlets in the formed heart. (Circ Res. 1998;82:464-472.) Key Words: development n outflow tract n semilunar valves n rat n transdifferentiation T he OFT of the embryonic heart is that portion of the heart tube that lies downstream from the developing ventricular segments 1 (for nomenclature, see “Materials and Methods”). As long as the embryonic heart has no properly functioning one-way valves, the OFT functions as a sphincter to prevent regurgitation of the blood by virtue of its slow conduction of the depolarizing impulse in conjunction with a long-lasting contraction and the abundant presence of endocardial jelly. 1–4 The subsequent remodeling of the cardiac jelly of the OFT into distinct endocardial ridges is accompanied by an extensive population with extracardiac mesenchymal cells that can be traced to the branchial arches and that originate largely from the neural crest. 5,6 As a result, the neural crest– derived mes- enchyme between the fourth and sixth arches is laterally continuous with that in the endocardial ridges. 7,8 It is generally agreed that apposition and fusion of the spiraling endocardial ridges from distal to proximal sites leads to the formation of the aorticopulmonary septum. 9 –11 However, considerable contro- versy continues to exist regarding the mechanism by which the OFT is septated. According to the concept of Thompson (discussed in References 7, 8, and 12 to 14), the condensed mesenchyme of the aorticopulmonary septum and of the endocardial ridges forms, together with the distal myocardial boundary of the OFT, a “septation complex,” which translo- cates toward the ventricle. This concept tacitly assumes that the distal (myocardial) rim of the OFT, once formed, can serve as a stable landmark on the cardiac tube. Because (1) the semilunar valves develop near the distal rim of the OFT and (2) the fourth branchial arch lies anterior to the sixth, whereas the pulmonary semilunar valve lies anterior to the aortic semilunar valve, this assumption implies that the junction of the OFT with great arteries rotates over ’150°. 15–19 To accommodate this rotation of the valve anlagen, “absorption” or “retraction” of the myocardium of the OFT is said to account for the compensatory counterrotation. 9,15,17,19,20 However, this hypoth- esis is difficult to reconcile with the finding that in vivo labeling of the OFT myocardium does not reveal such rota- tion. 21,22 Therefore, we investigated the alternative hypothesis that the distal (myocardial) boundary of the OFT is not a stable landmark but moves proximally over the spiraling course of the aortic and pulmonary routes before the semilunar valves develop, so that these valves can develop at their definitive Received June 5, 1997; accepted November 20, 1997. From the Department of Anatomy and Embryology, Academic Medical Center, University of Amsterdam, Amsterdam, the Netherlands. Correspondence to Wouter H. Lamers, MD, PhD, Department of Anatomy and Embryology, Academic Medical Center, University of Amsterdam, Meibergdreef 15, 1105 AZ, Amsterdam, Netherlands. E-mail [email protected] © 1998 American Heart Association, Inc. 464 by guest on January 22, 2015 http://circres.ahajournals.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Normal development of the outflow tract in the rat

Normal Development of the Outflow Tract in the RatJing Ya, Maurice J.B. van den Hoff, Piet A.J. de Boer, Sabina Tesink-Taekema, Diego Franco,
Antoon F.M. Moorman, Wouter H. Lamers
Abstract—The outflow tract (OFT) provides the structural components forming the ventriculoarterial connection. Theprevailing concept that this junction “rotates” to acquire its definitive topography also requires a concept of“counterrotation” and is difficult to reconcile with cell-marking studies. Rats between 10 embryonic days (EDs) and 2postnatal days were stained immunohistochemically and by in situ hybridization. DNA replication was determined byincorporation of bromodeoxyuridine and apoptosis by the annexin V binding and terminal deoxynucleotidyl transferase–mediated dUTP-X nick end labeling (TUNEL) assays. Starting at ED12, cardiomyocytes in the distal (truncal) part of theOFT begin to shed their myocardial phenotype without proceeding into apoptosis, suggesting transdifferentiation.Myocardial regression is most pronounced on the dextroposterior side and continues until after birth, as revealed by thedisappearance of the myocardial cuff surrounding the coronary roots and semilunar sinuses and by the establishment offibrous continuity between mitral and aortic semilunar valves. Fusion of the endocardial ridges of the truncus on late ED13is accompanied by the organization of a-smooth muscle actin–and nonmuscle myosin heavy chain–positive myofibroblastsinto a central whorl and the appearance of the semilunar valve anlagen at their definitive topographical position within theproximal portion of the truncus. After fusion of the proximal (conal) portion of the endocardial ridges, many of the residentmyofibroblasts undergo apoptosis and are replaced by cardiomyocytes. The distal myocardial boundary of the OFT is nota stable landmark but moves proximally over the spiraling course of the aortic and pulmonary routes, so that the semilunarvalves develop at their definitive topographic position. After septation, the distal boundary of the OFT continues to regress,particularly in its subaortic portion. The myocardializing conus septum, on the other hand, becomes largely incorporatedinto the right ventricle. These opposite developments account for the pronounced asymmetry of the subaortic andsubpulmonary outlets in the formed heart. (Circ Res. 1998;82:464-472.)
Key Words: development n outflow tract n semilunar valves n rat n transdifferentiation
The OFT of the embryonic heart is that portion of the hearttube that lies downstream from the developing ventricular
segments1 (for nomenclature, see “Materials and Methods”).As long as the embryonic heart has no properly functioningone-way valves, the OFT functions as a sphincter to preventregurgitation of the blood by virtue of its slow conduction ofthe depolarizing impulse in conjunction with a long-lastingcontraction and the abundant presence of endocardial jelly.1–4
The subsequent remodeling of the cardiac jelly of the OFTinto distinct endocardial ridges is accompanied by an extensivepopulation with extracardiac mesenchymal cells that can betraced to the branchial arches and that originate largely fromthe neural crest.5,6 As a result, the neural crest–derived mes-enchyme between the fourth and sixth arches is laterallycontinuous with that in the endocardial ridges.7,8 It is generallyagreed that apposition and fusion of the spiraling endocardialridges from distal to proximal sites leads to the formation of theaorticopulmonary septum.9–11 However, considerable contro-versy continues to exist regarding the mechanism by which theOFT is septated. According to the concept of Thompson(discussed in References 7, 8, and 12 to 14), the condensed
mesenchyme of the aorticopulmonary septum and of theendocardial ridges forms, together with the distal myocardialboundary of the OFT, a “septation complex,” which translo-cates toward the ventricle. This concept tacitly assumes that thedistal (myocardial) rim of the OFT, once formed, can serve asa stable landmark on the cardiac tube. Because (1) thesemilunar valves develop near the distal rim of the OFT and (2)the fourth branchial arch lies anterior to the sixth, whereas thepulmonary semilunar valve lies anterior to the aortic semilunarvalve, this assumption implies that the junction of the OFTwith great arteries rotates over '150°.15–19 To accommodatethis rotation of the valve anlagen, “absorption” or “retraction”of the myocardium of the OFT is said to account for thecompensatory counterrotation.9,15,17,19,20 However, this hypoth-esis is difficult to reconcile with the finding that in vivolabeling of the OFT myocardium does not reveal such rota-tion.21,22 Therefore, we investigated the alternative hypothesisthat the distal (myocardial) boundary of the OFT is not a stablelandmark but moves proximally over the spiraling course of theaortic and pulmonary routes before the semilunar valvesdevelop, so that these valves can develop at their definitive
Received June 5, 1997; accepted November 20, 1997.From the Department of Anatomy and Embryology, Academic Medical Center, University of Amsterdam, Amsterdam, the Netherlands.Correspondence to Wouter H. Lamers, MD, PhD, Department of Anatomy and Embryology, Academic Medical Center, University of Amsterdam,
Meibergdreef 15, 1105 AZ, Amsterdam, Netherlands.E-mail [email protected]© 1998 American Heart Association, Inc.
464 by guest on January 22, 2015http://circres.ahajournals.org/Downloaded from

topographic position. We will argue that transdifferentiation,rather than differential growth or apoptosis, is primarily re-sponsible for the proximal movement of the distal myocardialboundary during septation. After septation, the distal boundaryof the OFT continues to regress, particularly in its subaorticportion. The conus septum, on the other hand, becomes awell-developed mainly right ventricular structure as a result ofingrowth of myocytes from the surrounding part of the OFTmyocardium into the conus septum (“myocardialization”23).These opposite developments account for the pronouncedasymmetry of the subaortic and subpulmonary outlets in theformed heart. Finally, we show that a-SMA–positive neuralcrest–derived myofibroblasts that populate the walls of thebranchial arches and the endocardial ridges7,8,24,25 differ froma-SMA–positive cardiomyocytes by their exclusive expressionof nonmuscle MHCs.
Materials and Methods
AnimalsWistar rats, obtained from Broekman, Someren, the Netherlands,were kept in a controlled dark-light cycle (light, 11:00 AM to 11:00PM). Females in estrus were identified26 and mated. The time of matingwas regarded as the beginning of gestation (ED0). Rats were decap-itated under CO2 anesthesia, and embryos were harvested. To identifyproliferating cells, pregnant animals were equipped with an intraperi-toneal Alzet miniosmotic pump (model 2001D) supplying 80 mgBrdU/h, primed with 50 mg/kg BrdU (Sigma Chemical Co) intra-peritoneally, and killed 24 hours later. To identify early apoptotic cells,embryos between ED14 and ED16 were isolated without disturbingthe fetal membrane and placental circulation, injected with 2.75 mg(50 mL) recombinant human fluorescein-labeled annexin V, and killed15 minutes later. Animal welfare was in accordance with institutionalguidelines of the University of Amsterdam.
Embryos were fixed overnight in ice-cold methanol/acetone/water(2:2:1 [vol/vol]) for immunohistochemistry in 4% (wt/vol) formalde-hyde in PBS (150 mmol/L NaCl and 10 mmol/L sodium phosphate,pH 7.4) for in situ hybridization or in 4% (wt/vol) formaldehyde,1 mmol/L MgCl2, and 1 mmol/L CaCl2 for 4 hours at 4°C forapoptosis studies.27 Embedding, storage, and staining procedures wereas detailed elsewhere.28
A total of 50 immunohistochemically stained embryos (ED10, 3embryos; ED11, 5 embryos; ED12, 4 embryos; ED13, 9 embryos;ED14, 8 embryos; ED15, 3 embryos; ED16, 7 embryos; ED17, 1embryo; ED18, 2 embryos; ED19, 3 embryos; ED20, 3 embryos; andpostnatal day 2, 2 embryos), 20 BrdU-stained embryos (4 each forED13 to ED17), 8 TUNEL-stained embryos (2 each for ED14 toED17), and 16 annexin V–stained embryos (4 each for ED14 toED17) were investigated. In addition, our hematoxylin-azophloxine–
stained reference series of rat embryos (45 specimens between ED10and birth) was examined when appropriate.
ImmunohistochemistrySerial sections were incubated overnight with appropriately dilutedmonoclonal antibodies against either a-MHC (1:10, code 249 to5-A-4),28 b-MHC (1:10, 219–1D1),29 a-SMA (1:1000, SigmaIMMH-2), desmin (1:50, Monosan, Mon 3001, clone 33), a cyto-plasmic antigen of macrophages (1:200, Instruchemie, IC25–896,clone ED1),30 BrdU (1:50, Becton Dickinson), or a polyclonalantibody against fibronectin (1:6000, AB 1942, Brunschwig Chemie).To retrieve the BrdU epitope, sections were submerged in boiling10 mmol/L sodium citrate buffer for 1 minute and quenched inice-cold PBS before being exposed to antibody. Early apoptotic cellswere identified by their binding of annexin V,31 whereas late apoptoticcells were identified by the TUNEL assay32 according to the manu-facturer’s (Boehringer-Mannheim) instructions. Annexin V bindingand labeled DNA nicks were demonstrated with a peroxidase-coupledmonoclonal antibody against fluorescein (converter-POD, 1:15,Boehringer-Mannheim). Antibody binding was visualized with dia-minobenzidine and H2O2 as described.28
In Situ HybridizationIn situ hybridization was performed as described in detail previously.33
Antisense RNA probes were made from human NMMHCA cDNA(934 bp),34 human NMMHCB cDNA (4650 bp),34 mouse SMMHCcDNA (350 bp),35 and rat MLC2A cDNA (550 bp).36 NMMHCA andNMMHCB cRNAs were labeled with two nucleotides; the others,with one.
Three-Dimensional ReconstructionSelected hearts of immunostained serial sections of rat embryos werereconstructed three-dimensionally as described37 and rendered withthe help of a medical artist.
NomenclatureThe OFT is defined as that part of the embryonic heart that connectsthe embryonic right ventricle with the aortic sac. The aortic sac, inturn, is the mesenchymal structure that connects the OFT with thebranchial arch arteries. Our demarcation of the boundaries of the OFTfollows that provided by Pexieder.38 Its distal arterial boundary is takento coincide with the distal boundary of the myocardium. Externally,its proximal ventricular boundary can be identified by a furrow on themyocardial surface, whereas its internal boundary corresponds withthat of the endocardial ridges and with the boundary between thedeveloping trabeculae of the right ventricle and the nontrabeculatedwall of the OFT. The septal endocardial ridge takes origin on theventricular septum and covers the left lateral and posterior wall of theOFT, whereas the parietal ridge takes origin on the right lateral part ofthe junction between the OFT and right ventricle and covers the rightlateral and anterior wall of the OFT (see Reference 39). A character-istic bend halfway down the OFT corresponds internally with a fairlyacute change in direction of the endocardial ridges; in accordance withPexieder, we will designate the portion of the OFT that is proximal(upstream) to this bend as the conus and the portion distal (down-stream) to it as the truncus.
ResultsChanges in the Configuration of the DistalMyocardial BoundarySoon after the heart tube becomes identifiable at Theiler’s stage 12(ED9),40 the OFT develops into a prominent feature of theascending loop of the heart tube. As demonstrated by the stainingpattern of a- and b-MHC, the distal boundary of the myocardi-um of the OFT initially does not reach the aortic sac.41,42 In thespecimen shown in Fig 1A, the myocardium reaches up to thedeveloping bend between the conal and the truncal part of the
Selected Abbreviations and Acronyms
BrdU 5 bromodeoxyuridineED (with number) 5 embryonic day
MHC 5 myosin heavy chainMLC 5 myosin light chain
NMMHCA 5 nonmuscle MHC ANMMHCB 5 nonmuscle MHC B
OFT 5 outflow tractSMA 5 smooth muscle actin
SMMHC 5 smooth-muscle MHCTUNEL 5 terminal deoxynucleotidyl
transferase–mediated dUTP-X nick endlabeling
Ya et al 465
by guest on January 22, 2015http://circres.ahajournals.org/Downloaded from

OFT. The myocardial expansion toward the arterial pole iscompleted in the course of ED10 (Theiler’s stages 13 and 14).During ED11 (Theiler’s stages 15 and 16), the distal myocardialrim remained circular in configuration and apposed to the aorticsac (Fig 1B). However, at ED12 (Theiler’s stage 17), we observedthat the staining intensity of both a-MHC (not shown) andb-MHC (Fig 1C to 1F) started to decline on the right side of thedistal boundary. Although the cells in the area showing thisheterogeneous weak MHC staining property on ED12 no longerexpressed MHCs on early ED13 (Theiler’s stage 18), the processwas seen to have spread to adjacent myocardial cells (Figs 2 and 3).Meanwhile, a similar process had also started on the left side of theOFT myocardium (Fig 3), so that the originally flat distalmyocardial boundary of the OFT had transformed into a saddle-shaped structure by late ED13 (Theiler’s stage 19). At this stage,the “seat” of the saddle was deeper on the right than on the leftside and extended halfway through the truncus (Fig 3F and 3G).Meanwhile, growth of the right ventricle caused the bend in theOFT to begin straightening. In the course of ED14, MHC-positive cells continued to shed their myocardial phenotype, butnow the anterior and posterior “tongues” had also becomeinvolved. As a result, the MHC-positive dextroposterior bound-
ary of the myocardium was located more proximal than the rest ofthe distal rim of the myocardium (Figs 3F, 3G, and 4).
We measured the length of the myocardial OFT, ie, thedistance between the distal rim and the ventricular base of theendocardial ridges, between ED13 and ED18. The distancedeclined '60% between ED13 and ED14 to remain virtuallyconstant thereafter until ED18. This regression could not beaccounted for by local apoptosis, which was absent from theheart at this stage (Fig 5E). Furthermore, the OFT myocardi-um had a very low DNA-synthesizing activity, as demon-strated by a very low incorporation of BrdU between ED12and ED15 (Fig 5A and 5C). In fact, the difference in BrdUincorporation between OFT and ventricular myocardium wasso pronounced in this period that it could have been used todemarcate both cardiac segments. In sharp contrast, however,the areas that were in the process of shedding their myocardial
Figure 1. Right lateral views of reconstructions of the OFT inED10, ED11, and ED12 rat embryos (panels A, B, and C,respectively) and sections of the ED12 embryo (panel C) show-ing the expression of b-MHC (panel D), a-SMA (panel E), anddesmin (panel F) in the heart. Panels A to C show the distalboundary of the OFT as identified by the expression of b-MHC(interrupted line), the myocardium losing its myocardial pheno-type (panel C, asterisk in shaded area), and the reflection of theepicardium to the pericardium (arrow pointing at stippled line).Note the development of the bend between the truncal and theconal portions of the OFT. Note also that the area showingregression of b-MHC staining (panel D, arrows) cannot yet berecognized in the a-SMA–stained and desmin-stained sections(panels E and F, respectively). AS indicates aortic sac; 4th, partof aortic sac giving access to fourth branchial arch arteries; 6thand 314th, sixth and combined third and fourth branchial arter-ies, respectively; and RV, right ventricle. Bar550 mm.
Figure 2. Fate of the distal boundary of the OFT myocardium inearly (panels A to D) and mid-ED13 (panels E to J) rat embryos.The embryo in panels A to D was sectioned transversely, andthat in panels E to J was sectioned frontally. Panels A, B, H,and I were stained for the presence of b-MHC; panels C and Eto G, for the presence of a-SMA; and panels D and J, for thepresence of desmin. Panel B is a higher magnification of thebox appearing in panel A. Panels F, I, and J are serial sections,and panels E and H are neighboring sections. Note the regres-sive staining for b-MHC (panels A, B, H, and I), leaving the ante-rior and posterior “tongues” of remaining myocardium (panel H).Panels C and E to G show the population of endocardial ridgesby a-SMA–positive myofibroblasts; the aorticopulmonary sep-tum (APS, panel E) is laterally continuous with the septal andparietal ridges (S and P, respectively; panels F and G). Theintercalated ridges (I) contain many desmin-positive cells (panelJ) but virtually no a-SMA–positive cells (panel F). A indicatesaorta; PT, pulmonary trunk; RV, right ventricle; and 6th, 6th archarteries. Bar5100 mm (panels A, C, and D) and 50 mm (panels Band E to J).
466 Development of Outflow Tract in Rat
by guest on January 22, 2015http://circres.ahajournals.org/Downloaded from

phenotype were characterized by a rather high level ofincorporation of BrdU on both ED12 (not shown) and ED13(Fig 5A and 5C). The loss of myofibrillar staining thereforeoccurs in healthy nonreplicating cells and is followed by amitotic wave. From ED15, DNA synthetic activity graduallyincreased in the remaining myocardium of the OFT, so thatfrom ED17 onward, it could no longer be distinguished by thiscriterion from the ventricular myocardium (not shown).
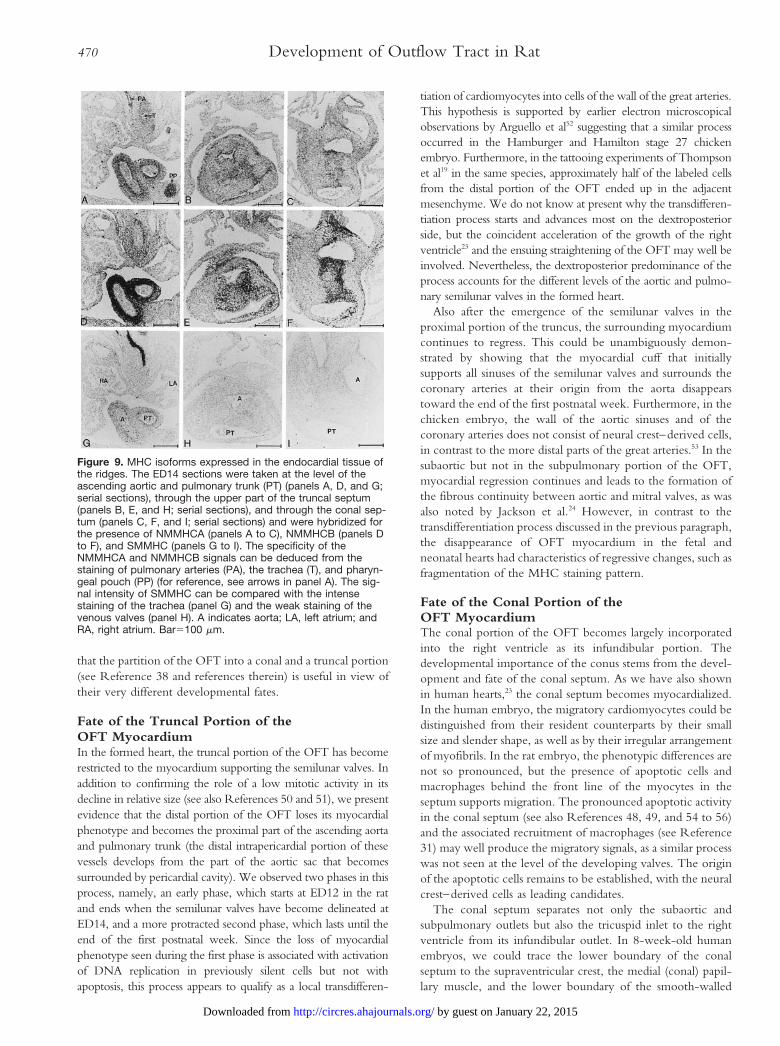
The coronary arteries were found to be useful structures forfollowing the subsequent fate of the distal boundary of theOFT myocardium. Coronary arteries could first be identifiedunambiguously near the distal rim of the OFT at ED16 (Fig6A), although suggestive structures were already seen at ED15(Fig 4G to 4I). More important, Fig 6A clearly shows that the
aortic root of the coronary arteries was localized within theconfines of the myocardium of the OFT. This cuff of OFTmyocardium still supported the semilunar valves and, hence,still surrounded the root of the coronary arteries 2 days afterbirth (Fig 6B to 6D), but the fragmented staining pattern ofb-MHC suggested regressive changes. Indeed, 1 week later,the myocardial cuff was no longer present, and a fibrouscontinuity had developed between the mitral valve and thesemilunar valves of the aorta, whereas the semilunar valves ofthe pulmonary trunk were supported by a free-standingpulmonary infundibulum (not shown; see Reference 24).
Remodeling of the Endocardial Tissue ofthe OFTUp to ED12, the endocardial tissue of the OFT lined the inside ofthe myocardial tube as an almost uniformly thick cuff. a-SMA–positive cells could be seen to populate the endocardial ridge tissue
Figure 3. Fate of the distal boundary of the OFT myocardium inlate ED13 embryos (panels A to E), dextroanterior view of OFTon early ED13 revealing area with regressing myocardial pheno-type (asterisk in shaded area in panel F; reconstruction is basedon the embryo shown in panels A to D of Fig 2), and right lateralview of OFT late on ED13 (panel G; reconstruction is based onthe embryo shown in panels A to E). Panels A to E show trans-verse sections of the embryo stained for the presence ofb-MHC (panels A and C), a-SMA (panels B and E), and desmin(panel D). The sections show that the distal boundary of theOFT is further distal on the left than on the right side, that theportion of the wall of the OFT that has lost its b-MHC–positivestaining has become intensely desmin-positive (see panels Cand D), and that the a-SMA–positive myofibroblasts in the aorti-copulmonary septum (APS, panel E) are anteriorly and posteri-orly (panel B) continuous with the same cells in the septal andparietal endocardial ridges (S and P, respectively). Note theregression of the bend between truncal and conal portions ofthe OFT (panels F and G) and the anterior course of the ascend-ing aortic trunk above the OFT compared with the straightascending course of the pulmonary trunk (PT, panel G). A indi-cates aorta; RV, right ventricle; and 6th, 6th arch arteries.Bar5100 mm.
Figure 4. Fate of the distalboundary of the OFT myocar-dium in ED14 (panels A to F)and ED15 (panels G to I)embryos and right lateralview of the OFT of the ED14embryo (panel J) showingthat fusion is complete in thetruncus portion and is pro-gressing in the conus portion.The panels show frontal sec-tions stained for the presencea-SMA (panels A, D, and G),b-MHC (panels B, E, and H),and desmin (panel I). PanelsC and F are stained with
hematoxylin and azoflochsin to show the transition of endotheli-um (thick layer) into endocardium (thin layer) at the level of thevalve swellings. Panels B, E, H, and J show that the myocardialrim supporting the aortic semilunar valve occupies a more prox-imal position than that supporting the pulmonary valve. PanelsA, D, and G show that the accumulation of the a-SMA–positivemyofibroblasts into a central whorl-shaped structure leaves thedeveloping valve swellings in the septal and parietal ridges virtu-ally free of a-SMA–positive cells. The valve swelling of the inter-calated ridge does not express a-SMA either but is positive fordesmin (panels G and I). Panels G and H show a structure thatis suggestive for a developing coronary artery (arrows). A indi-cates aorta; APS, aorticopulmonary septum; and PT, pulmonarytrunk. Dotted lines in panels D, G, and I indicate luminal bound-ary of endocardial tissue. The interrupted line in panel J indi-cates the distal rim of myocardium. Bar5100 mm.
Ya et al 467
by guest on January 22, 2015http://circres.ahajournals.org/Downloaded from
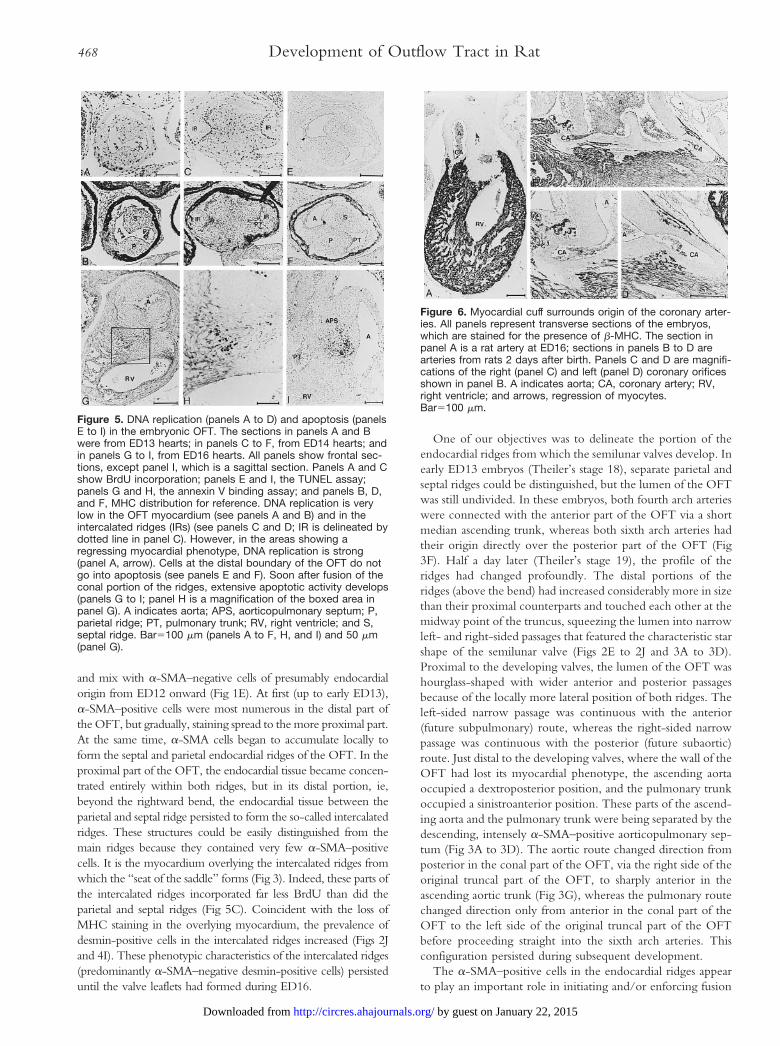
and mix with a-SMA–negative cells of presumably endocardialorigin from ED12 onward (Fig 1E). At first (up to early ED13),a-SMA–positive cells were most numerous in the distal part ofthe OFT, but gradually, staining spread to the more proximal part.At the same time, a-SMA cells began to accumulate locally toform the septal and parietal endocardial ridges of the OFT. In theproximal part of the OFT, the endocardial tissue became concen-trated entirely within both ridges, but in its distal portion, ie,beyond the rightward bend, the endocardial tissue between theparietal and septal ridge persisted to form the so-called intercalatedridges. These structures could be easily distinguished from themain ridges because they contained very few a-SMA–positivecells. It is the myocardium overlying the intercalated ridges fromwhich the “seat of the saddle” forms (Fig 3). Indeed, these parts ofthe intercalated ridges incorporated far less BrdU than did theparietal and septal ridges (Fig 5C). Coincident with the loss ofMHC staining in the overlying myocardium, the prevalence ofdesmin-positive cells in the intercalated ridges increased (Figs 2Jand 4I). These phenotypic characteristics of the intercalated ridges(predominantly a-SMA–negative desmin-positive cells) persisteduntil the valve leaflets had formed during ED16.
One of our objectives was to delineate the portion of theendocardial ridges from which the semilunar valves develop. Inearly ED13 embryos (Theiler’s stage 18), separate parietal andseptal ridges could be distinguished, but the lumen of the OFTwas still undivided. In these embryos, both fourth arch arterieswere connected with the anterior part of the OFT via a shortmedian ascending trunk, whereas both sixth arch arteries hadtheir origin directly over the posterior part of the OFT (Fig3F). Half a day later (Theiler’s stage 19), the profile of theridges had changed profoundly. The distal portions of theridges (above the bend) had increased considerably more in sizethan their proximal counterparts and touched each other at themidway point of the truncus, squeezing the lumen into narrowleft- and right-sided passages that featured the characteristic starshape of the semilunar valve (Figs 2E to 2J and 3A to 3D).Proximal to the developing valves, the lumen of the OFT washourglass-shaped with wider anterior and posterior passagesbecause of the locally more lateral position of both ridges. Theleft-sided narrow passage was continuous with the anterior(future subpulmonary) route, whereas the right-sided narrowpassage was continuous with the posterior (future subaortic)route. Just distal to the developing valves, where the wall of theOFT had lost its myocardial phenotype, the ascending aortaoccupied a dextroposterior position, and the pulmonary trunkoccupied a sinistroanterior position. These parts of the ascend-ing aorta and the pulmonary trunk were being separated by thedescending, intensely a-SMA–positive aorticopulmonary sep-tum (Fig 3A to 3D). The aortic route changed direction fromposterior in the conal part of the OFT, via the right side of theoriginal truncal part of the OFT, to sharply anterior in theascending aortic trunk (Fig 3G), whereas the pulmonary routechanged direction only from anterior in the conal part of theOFT to the left side of the original truncal part of the OFTbefore proceeding straight into the sixth arch arteries. Thisconfiguration persisted during subsequent development.
The a-SMA–positive cells in the endocardial ridges appearto play an important role in initiating and/or enforcing fusion
Figure 5. DNA replication (panels A to D) and apoptosis (panelsE to I) in the embryonic OFT. The sections in panels A and Bwere from ED13 hearts; in panels C to F, from ED14 hearts; andin panels G to I, from ED16 hearts. All panels show frontal sec-tions, except panel I, which is a sagittal section. Panels A and Cshow BrdU incorporation; panels E and I, the TUNEL assay;panels G and H, the annexin V binding assay; and panels B, D,and F, MHC distribution for reference. DNA replication is verylow in the OFT myocardium (see panels A and B) and in theintercalated ridges (IRs) (see panels C and D; IR is delineated bydotted line in panel C). However, in the areas showing aregressing myocardial phenotype, DNA replication is strong(panel A, arrow). Cells at the distal boundary of the OFT do notgo into apoptosis (see panels E and F). Soon after fusion of theconal portion of the ridges, extensive apoptotic activity develops(panels G to I; panel H is a magnification of the boxed area inpanel G). A indicates aorta; APS, aorticopulmonary septum; P,parietal ridge; PT, pulmonary trunk; RV, right ventricle; and S,septal ridge. Bar5100 mm (panels A to F, H, and I) and 50 mm(panel G).
Figure 6. Myocardial cuff surrounds origin of the coronary arter-ies. All panels represent transverse sections of the embryos,which are stained for the presence of b-MHC. The section inpanel A is a rat artery at ED16; sections in panels B to D arearteries from rats 2 days after birth. Panels C and D are magnifi-cations of the right (panel C) and left (panel D) coronary orificesshown in panel B. A indicates aorta; CA, coronary artery; RV,right ventricle; and arrows, regression of myocytes.Bar5100 mm.
468 Development of Outflow Tract in Rat
by guest on January 22, 2015http://circres.ahajournals.org/Downloaded from

of the ridges. In late ED13 hearts, the a-SMA–positive cellscould be seen to align longitudinally between the attachmentof the septal and parietal ridges on the lateral wall of the OFTon the one hand and their place of origin in the mesenchymesurrounding the aortic sac on the other hand (Fig 3A to 3D).Fusion of the ridges is accompanied by accumulation of thea-SMA–positive cells into a highly characteristic whorllikestructure at the site of fusion (Fig 4), depleting the moreperipheral parts of the newly formed septum of a-SMA–positive cells. These a-SMA–depleted parts of the septumsubsequently formed, together with the a-SMA–sparse inter-calated ridges, the leaflets of the semilunar valves (Fig 7).
Below the valvular swellings, the laterally positioned ridgeswere still separate at ED14, even though their volume hadmarkedly increased (Fig 4). On their fusion on ED15 (Theiler’sstage 21), the parietal and septal endocardial ridges form theconal (or infundibular) septum between the subpulmonary andsubaortic outlets of the ventricles. Soon after fusion, many ofthe nonmyocardial cells of the septum were seen to undergoapoptosis (Fig 5G to 5I). Apoptotic cells were identified bothby binding of annexin V to their membranes (early apoptoticcells31) and by terminal-transferase labeling of nicked nuclearDNA (late apoptotic cells32). Annexin V binding was first seenon ED15, whereas both parameters were positive on ED16 andED17. Apoptotic involution did not include the surroundingmyocardium. The apoptotic cells were found to attract fetalmacrophages to execute their removal (Fig 8B and 8C).Meanwhile, myocardial cells at the margin of the fused ridgesstarted to migrate into the ridges to replace the regressingnonmyocardial cells (Fig 8A). Active migration is suggested bythe slender shape of the myocytes and by the presence ofapoptotic cells and macrophages behind the myocardial frontline (Fig 8B and 8C). The frequency of BrdU accumulation inthese migratory myocardial cells was not higher than in theadjacent OFT myocardium (not shown), suggesting that mi-gratory cells were not primarily recruited by cell multiplica-tion. Myocardialization was completed first at the ventricularend of the conal septum on ED16 and last at the distal end (Figs7 and 8). On myocardialization, the conal septum forms the
supraventricular crest, which separates the right ventricularinlet from the outlet, and the outlet portion of the muscularventricular septum, which separates the subaortic from thesubpulmonary outlet. As a result of the ongoing regression inthe myocardium in the subaortic outlet (see previous section),the distal portion of the outlet septum becomes the posteriorpart of the freestanding subpulmonary infundibulum.
MHCs of the SMA-Positive Cells in the RidgesThe a-SMA–positive cells in the endocardial ridges of the OFTappear to have an important morphogenetic function. To estab-lish their contractile properties, we determined the type of MHCwith which the actin was associated (Fig 9). Staining for MLC2Awas used to delineate the myocardium (not shown). In situhybridization analysis showed that cells in the endocardial ridgeswere characterized by a strong expression of NMMHCB and toa much lesser extent by the expression of NMMHCA, whereasno expression of SMMHC was seen. Further distal, the walls ofthe branchial arch arteries stained very intensely for NMMHCBand, to a lesser extent, NMMHCA and weakly for SMMHC.The wall of the dorsal (descending) aorta was negative forNMMHCA and NMMHCB but strongly positive for SMMHC.The differential staining of the periesophageal tissue and thepharyngeal pouch confirmed that the NMMHCA signal did notresult from cross hybridization with NMMHCB. The intensity ofthe SMMHC signal could be related to the very strong signal thatwas seen in the smooth muscle cells in the developing lung.
DiscussionThe OFT provides the structural components to form thedefinitive architecture of the ventriculoarterial connection. Itsnormal development has been the subject of numerous stud-ies.7–11,13–19,43–49 The formation of the OFT portion of themyocardial tube follows a proximodistal direction and iscompleted at ED12 in the rat (Theiler’s stage 17, Carnegiestage 14 in human embryos [authors’ unpublished data, 1997],and Hamburger and Hamilton stage 22 in the chick21).Subsequently, its relative size declines.15,20 The present studyaddresses this second phase in OFT development and shows
Figure 7. Phenotypic characteristics of the cells forming thesemilunar valves. The panels represent frontal sections of anED17 embryo stained for the presence of b-MHC (panel A),a-SMA (panel B), and fibronectin (panel C). Note that theattachment of the valve leaflets to the myocardium is formed bya-SMA–positive and fibronectin-positive tissue, whereas thevalve leaflets themselves are a-SMA–negative and fibronectin-negative. A indicates aorta; PT, pulmonary trunk. Bar5100 mm.
Figure 8. Myocardialization of the conal septum (CS) at ED16.The panels show frontal sections of the embryo that are stainedfor expression of b-MHC (panels A and B) and for the presenceof fetal macrophages (panel C). Panel A is distal to panels Band C, which are serial sections. The boundary of the advancingmyocardium as seen in panel B is superimposed on panel C(dotted line) to show that it has moved ahead of the recruitedmacrophages (box). A indicates aorta; PT, pulmonary trunk.Bar5100 mm.
Ya et al 469
by guest on January 22, 2015http://circres.ahajournals.org/Downloaded from
that the partition of the OFT into a conal and a truncal portion(see Reference 38 and references therein) is useful in view oftheir very different developmental fates.
Fate of the Truncal Portion of theOFT MyocardiumIn the formed heart, the truncal portion of the OFT has becomerestricted to the myocardium supporting the semilunar valves. Inaddition to confirming the role of a low mitotic activity in itsdecline in relative size (see also References 50 and 51), we presentevidence that the distal portion of the OFT loses its myocardialphenotype and becomes the proximal part of the ascending aortaand pulmonary trunk (the distal intrapericardial portion of thesevessels develops from the part of the aortic sac that becomessurrounded by pericardial cavity). We observed two phases in thisprocess, namely, an early phase, which starts at ED12 in the ratand ends when the semilunar valves have become delineated atED14, and a more protracted second phase, which lasts until theend of the first postnatal week. Since the loss of myocardialphenotype seen during the first phase is associated with activationof DNA replication in previously silent cells but not withapoptosis, this process appears to qualify as a local transdifferen-
tiation of cardiomyocytes into cells of the wall of the great arteries.This hypothesis is supported by earlier electron microscopicalobservations by Arguello et al52 suggesting that a similar processoccurred in the Hamburger and Hamilton stage 27 chickenembryo. Furthermore, in the tattooing experiments of Thompsonet al19 in the same species, approximately half of the labeled cellsfrom the distal portion of the OFT ended up in the adjacentmesenchyme. We do not know at present why the transdifferen-tiation process starts and advances most on the dextroposteriorside, but the coincident acceleration of the growth of the rightventricle23 and the ensuing straightening of the OFT may well beinvolved. Nevertheless, the dextroposterior predominance of theprocess accounts for the different levels of the aortic and pulmo-nary semilunar valves in the formed heart.
Also after the emergence of the semilunar valves in theproximal portion of the truncus, the surrounding myocardiumcontinues to regress. This could be unambiguously demon-strated by showing that the myocardial cuff that initiallysupports all sinuses of the semilunar valves and surrounds thecoronary arteries at their origin from the aorta disappearstoward the end of the first postnatal week. Furthermore, in thechicken embryo, the wall of the aortic sinuses and of thecoronary arteries does not consist of neural crest–derived cells,in contrast to the more distal parts of the great arteries.53 In thesubaortic but not in the subpulmonary portion of the OFT,myocardial regression continues and leads to the formation ofthe fibrous continuity between aortic and mitral valves, as wasalso noted by Jackson et al.24 However, in contrast to thetransdifferentiation process discussed in the previous paragraph,the disappearance of OFT myocardium in the fetal andneonatal hearts had characteristics of regressive changes, such asfragmentation of the MHC staining pattern.
Fate of the Conal Portion of theOFT MyocardiumThe conal portion of the OFT becomes largely incorporatedinto the right ventricle as its infundibular portion. Thedevelopmental importance of the conus stems from the devel-opment and fate of the conal septum. As we have also shownin human hearts,23 the conal septum becomes myocardialized.In the human embryo, the migratory cardiomyocytes could bedistinguished from their resident counterparts by their smallsize and slender shape, as well as by their irregular arrangementof myofibrils. In the rat embryo, the phenotypic differences arenot so pronounced, but the presence of apoptotic cells andmacrophages behind the front line of the myocytes in theseptum supports migration. The pronounced apoptotic activityin the conal septum (see also References 48, 49, and 54 to 56)and the associated recruitment of macrophages (see Reference31) may well produce the migratory signals, as a similar processwas not seen at the level of the developing valves. The originof the apoptotic cells remains to be established, with the neuralcrest–derived cells as leading candidates.
The conal septum separates not only the subaortic andsubpulmonary outlets but also the tricuspid inlet to the rightventricle from its infundibular outlet. In 8-week-old humanembryos, we could trace the lower boundary of the conalseptum to the supraventricular crest, the medial (conal) papil-lary muscle, and the lower boundary of the smooth-walled
Figure 9. MHC isoforms expressed in the endocardial tissue ofthe ridges. The ED14 sections were taken at the level of theascending aortic and pulmonary trunk (PT) (panels A, D, and G;serial sections), through the upper part of the truncal septum(panels B, E, and H; serial sections), and through the conal sep-tum (panels C, F, and I; serial sections) and were hybridized forthe presence of NMMHCA (panels A to C), NMMHCB (panels Dto F), and SMMHC (panels G to I). The specificity of theNMMHCA and NMMHCB signals can be deduced from thestaining of pulmonary arteries (PA), the trachea (T), and pharyn-geal pouch (PP) (for reference, see arrows in panel A). The sig-nal intensity of SMMHC can be compared with the intensestaining of the trachea (panel G) and the weak staining of thevenous valves (panel H). A indicates aorta; LA, left atrium; andRA, right atrium. Bar5100 mm.
470 Development of Outflow Tract in Rat
by guest on January 22, 2015http://circres.ahajournals.org/Downloaded from

conus of the right ventricle.23 The distal end of the conalseptum may not be myocardialized and persist as the ligamentof the conus, at the base of the aortic and pulmonary roots. Asa result of the ongoing regression of the myocardium in thesubaortic outlet, the distal portion of the myocardialized outletseptum becomes the posterior part of the freestanding subpul-monary infundibulum.
Myofibroblasts of the Endocardial TissueThe very strong expression of a-SMA in neural crest–derivedcells that populate the walls of the branchial arches and theendocardial ridges of the OFT is well known in both mammalsand birds.7,8,24,25 Although several studies have shown that thesmooth muscles cells in the wall of the aorta and pulmonaryarteries of embryonic and fetal mammals express the SM1 andSMemb isoforms of SMMHC and the B form of nonmuscleMHC (NMMHCB),35,57–59 data on the differential distributionof SMMHC and nonmuscle MHC in the wall of arteries ofneural crest and mesodermal origin were not yet available. Weshow that the expression of NMMHCB and, to a lesser extent,NMMHCA are confined to the branchial arch arteries, ie,arteries with a contribution from the neural crest. Thisassociation of NMMHCB with the neural crest is alsostrengthened by its presence in neural cells.60 On the otherhand, the descending aorta, which does not receive a contri-bution of the neural crest, expresses only SMMHC. Inside theendocardial tissues of the OFT, SMMHC was not expressed(see also Reference 35), whereas its expression in the ascendingaorta and pulmonary trunk was initially weak.
Although we studied only phenotypic markers, the coex-pression of a-SMA and nonmuscle MHCs by a subpopulationof cells in the endocardial ridges and in the wall of the branchialarch arteries certainly suggests that this phenotype identifiescells with a neural crest origin. In this respect, it is remarkablethat such cells were virtually absent in the intercalated ridgesand subsequently seemed to migrate away from the part of theparietal and septal ridges that form the semilunar valve leafletsto generate a very dense “whorl” at the site of fusion of theridges. This description implies that the neural crest does notplay a prominent role in semilunar valve development. Thishypothesis is supported by the finding that mice deficient forthe transcription factors Sox461 and Pax362 or retinoic acidreceptors63,64 all suffer from a common trunk, but only adeficient function of the endocardial ridge tissue (Sox4 defi-ciency) is associated with a deficient morphogenesis of thesemilunar valves.
ConclusionA striking feature of the aortic and pulmonary blood channelsis that major changes in the direction of both routes are realizedat different locations. The aortic route is characterized by apronounced and long forward bend in its extracardiac trajec-tory to connect the posterior subaortic outlet with the fourtharch arteries (aortic arc). The pulmonary route, on the otherhand, takes a forward turn inside the right ventricle, windsaround the supraventricular crest and the muscular outletseptum, but, from there on, ascends straight into the pulmo-nary trunk. The layout of this pattern becomes visible and iscompleted in the rat embryo in a single day (ED13, Carnegie
stage 16 in the human embryo) and basically does not changethereafter. It coincides temporally with the appearance of thesemilunar valve anlagen. Although it is generally accepted thatthe spiraling course of the septal and parietal ridges plays amajor role in realizing these connections between ventriclesand arteries, the transformation of these ridges into structures(the semilunar valves and the outlet septum) without such aspiraling topographical relation has remained an issue. We havenow shown that this issue can be resolved if the transfer of thecells of the distal portion of the OFT to the root of the greatarteries is taken into account. Nevertheless, the fate of thesecells will need to be established more definitely by lineagemarking.
AcknowledgmentsWe thank I.E.M. Oosterling for skillfully preparing the drawings ofthe three-dimensional reconstructions and C. Gravemeijer forhis photographs.
References1. Moorman AFM, Lamers WH. Molecular anatomy of the developing heart.
Trends Cardiovasc Med. 1994;4:257–264.2. Arguello C, Alanis J, Pantoja O, Valenzuela B. Electrophysiological and
ultrastructural study of the atrioventricular canal during the development ofthe chick embryo. J Mol Cell Cardiol. 1986;18:499–510.
3. de Jong F, Opthof T, Wilde AAM, Janse MJ, Charles R, Lamers WH,Moorman AFM. Persisting zones of slow impulse conduction indeveloping chicken hearts. Circ Res. 1992;71:240–250.
4. Lamers WH, de Jong F, de Groot IJM, Moorman AFM. The developmentof the avian conduction system. Eur J Morphol. 1991;29:233–253.
5. Kirby ML, Gale TF, Stewart DE. Neural crest cells contribute to normalaorticopulmonary septation. Science. 1983;220:1059–1061.
6. Kirby ML, Waldo K. Neural crest and cardiovascular patterning. Circ Res.1995;77:211–215.
7. Rosenquist GC, Fray-Gavalas C, Waldo K, Beall AC. Development of themusculoelastic septation complex in the avian truncus arteriosus. Am JAnat. 1990;189:339–356.
8. Beall AC, Rosenquist TH. Smooth muscle cells of neural crest origin formthe aorticopulmonary septum in the avian embryo. Anat Rec. 1990;226:360–366.
9. Kramer TC. The partitioning of the truncus and conus and the formationof the membranous portion of the interventricular septum in the humanheart. Am J Anat. 1942;71:343–370.
10. de Vries PA, de CM Saunders JB. Development of the ventricles and spiraloutflow tract in the human heart. Contrib Embryol. 1962;37:87–114.
11. van Mierop LHS, Alley RD, Kausel HW, Stranahan A. Pathogenesis oftransposition complexes, I: embryology of the ventricles and great arteries.Am J Cardiol. 1963;12:216–225.
12. Thompson RP, Fitzharris TP. Morphogenesis of the truncus arteriosus ofthe chick embryo heart: tissue reorganization during septation. Am J Anat.1979;156:251–264.
13. Thompson RP, Wong M, Fitzharris TP. A computer graphic study ofcardiac truncal septation. Anat Rec. 1983;206:207–214.
14. Rosenquist TH, McRoy JR, Waldo K, Kirby ML. Origin and propagationof elastogenesis in the developing cardiovascular system. Anat Rec. 1988;221:860–871.
15. Goor DA, Dische R, Lillehei CW. The conotruncus, I: its normalinversion and conus absorption. Circulation. 1972;46:375–384.
16. Asami I. Partitioning of the arterial end of the embryonic heart. In: VanPraagh R, Takao A, eds. Etiology and Morphogenesis of Congenital HeartDisease. 4th ed. Mt Kisco, NY: Futura Publishing Co; 1992:51–61.
17. Asami I. Beitrag zur Entwicklung des Kammerseptums in menschlichenHerzen mit besonderer Berucksichtigung der sogenannten Bulbusdrehung.Z Anat Entw Gesch. 1969;128:1–17.
18. Lomonico MP, Moore GW, Hutchins GM. Rotation of the junction ofthe outflow tract and great arteries in the embryonic human heart. AnatRec. 1986;216:544–549.
19. Thompson RP, Abercrombie V, Wong M. Morphogenesis of the truncusarteriosus of the chick embryo heart: movements of autoradiographictattoos during septation. Anat Rec. 1987;218:434–440.
Ya et al 471
by guest on January 22, 2015http://circres.ahajournals.org/Downloaded from

20. Thurkow EW, Wenink ACG. Development of the ventriculoarterialsegment of the human embryonic heart: a morphometric study. Anat Rec.1993;236:664–670.
21. de la Cruz MV, Sanchez Gomez C, Arteaga MM, Arguello C. Experi-mental study of the development of the truncus and the conus in the chickembryo. J Anat. 1977;123:661–686.
22. Seichert V. Study of the tissue and organ anlage shifts by the method ofplastic linear marking. Folia Morphol (Praha). 1965;13:228–238.
23. Lamers WH, Viragh S, Wessels A, Moorman AFM, Anderson RH. For-mation of the tricuspid valve in the human heart. Circulation. 1995;91:111–121.
24. Jackson M, Connell MG, Smith A, Anderson RH. Immunohistochemicalevaluation of the developing outflow tract in the rat: achieving aortic tomitral fibrous continuity. Cardiovasc Res. 1995;30:262–269.
25. Noden DM, Poelmann RE, Gittenberger-de Groot AC. Cell origins andtissue boundaries during outflow tract development. Trends Cardiovasc Med.1995;5:69–75.
26. Lamers WH, te Kortschot A, Moorman AFM, Los JA. Acetylcholinesterasein prenatal rat heart: a marker for the early development of the cardiacconductive tissue? Anat Rec. 1987;217:361–370.
27. Vermes I, Haanen C, Steffens-Nakken H, Reuteligsperger C. A novelessay for apoptosis flow cytometric detection of phosphatidylserineexpression on early apoptotic cells using fluorescein labelled annexin V.J Immunol Methods. 1995;184:39–51.
28. de Groot IJM, Lamers WH, Moorman AFM. Isomyosin expression patternduring rat heart morphogenesis: an immunohistochemical study. Anat Rec.1989;224:365–373.
29. Sant’Ana Pereira JAA, Wessels A, Nytmans L, Moorman AFM, SargeantAJ. New method for the accurate characterization of single human skeletalmuscle fibres demonstrates a relation between mATPase and MyHCexpression in pure and hybrid fiber types. J Muscle Res Cell Motil. 1995;16:21–34.
30. Dijkstra CD, Dopp EA, Joling P, Kraal G. The heterogeneity of mono-nuclear phagocytes in lymphoid organs: distinct macrophage subpopu-lations in the rat recognized by monoclonal antibodies ED1, ED2 and ED3.Immunology. 1985;54:589–599.
31. Fadok VA, Voelker DR, Campbell PA, Cohen JJ, Bratton DL, HensonPM. Exposure of phosphatidylserine on the surface of apoptotic lym-phocytes triggers specific recognition and removal by macrophages.J Immunol. 1992;148:2207–2216.
32. Arends MJ, Morris RG, Wyllie AH. Apoptosis: the role of the endonu-clease. Am J Pathol. 1990;136:593–608.
33. Moorman AFM, Vermeulen JLM, Koban MU, Schwartz K, Lamers WH,Boheler KR. Patterns of expression of sarcoplasmic reticulum Ca21-ATPase and phospholamban mRNAs during rat heart development. CircRes. 1995;76:616–625.
34. Simons M, Wang M, McBride OW, Kawamoto S, Yamakawa K, Gdula D,Adelstein RS, Weir L. Human nonmuscle myosin heavy chains areencoded by two genes located on different chromosomes. Circ Res. 1991;69:530–539.
35. Miano JM, Cserjesi P, Ligon KL, Periasamy M, Olson EN. Smooth musclemyosin heavy chain exclusively marks the smooth muscle lineage duringmouse embryogenesis. Circ Res. 1994;75:803–812.
36. Kubalak SW, Miller-Hance WC, O’Brien TX, Dyson E, Chien KR.Chamber specification of atrial myosin light chain-2 expression precedesseptation during murine cardiogenesis. J Biol Chem. 1994;269:16961–16970.
37. Verbeek FJ, Huysmans DP, Baeten RJAM, Schoutsen NJC, Lamers WH.Design and implementation of a database and program for3D-reconstruction from serial sections: a data-driven approach. Microsc ResTech. 1995;30:496–512.
38. Pexieder T. Conotruncus and its septation at the advent of the molecularbiology era. In: Clark EB, Markwald RR, Takao A, eds. DevelopmentalMechanisms of Heart Disease. Armonk, NY: Futura Publishing Co; 1995:227–247.
39. Los JA. Embryology. In: Watson H, ed. Paediatric Cardiology. London, UK:Lloyd Luke; 1968:1.
40. Theiler K. The House Mouse: Development and Normal Stages From Fertili-zation to 4 Weeks of Age. Berlin, Germany: Springer-Verlag; 1972.
41. de Jong F, Geerts WJC, Lamers WH, Los JA, Moorman AFM. Isomyosinexpression pattern during formation of the tubular chicken heart: a 3Dimmunohistochemical analysis. Anat Rec. 1990;226:213–227.
42. Jing Y, Markman MWM, Wagenaar GTM, Blommaart PJE, MoormanAFM, Lamers WH. Expression of the smooth-muscle proteins alphasmooth-muscle actin and calponin, and of the intermediate filamentprotein desmin are parameters of cardiomyocyte maturation in the prenatalrat heart. Anat Rec. 1997;249:495–505.
43. Bartelings MM, Gittenberger-de Groot AC. The arterial orifice level in theearly human embryo. Anat Embryol. 1988;177:537–542.
44. Thompson RP, Fitzharris TP. Morphogenesis of the truncus arteriosus ofthe chick embryo heart: the formation and migration of mesenchymaltissue. Am J Anat. 1979;154:545–556.
45. Fananapazir L, Kaufman MH. Observations on the development of theaortico-pulmonary spiral septum in the mouse. J Anat. 1988;158:157–172.
46. McBride RE, Moore GW, Hutchins GM. Development of the outflowtract and closure of the interventricular septum in the normal human heart.Am J Anat. 1981;160:309–331.
47. Orts Llorca F, Puerta Fonolla J, Sobrado J. The formation, septation andfate of the truncus arteriosus in man. J Anat. 1982;134:41–56.
48. Okamoto N, Akimoto N, Satow Y, Hidaka N, Miyabara S. Role of celldeath in conal ridges of developing human heart. Perspect Cardiovasc Res.1981;5:127–137.
49. Satow Y, Okamoto N, Akimoto N, Hidaka N, Miyabara S, Pexieder T.Comparative and morphometric study on genetically programmed celldeath in rat and chick embryonic heart. Cardiovasc Res. 1981;5:115–126.
50. Pexieder T. Development of the outflow tract of the embryonic heart. In:Rosenquist GC, Bergsma D, eds. Morphogenesis and Malformation of theCardiovascular System. New York, NY: AR Liss Inc; 1978:29–68.
51. Thompson RP, Lindroth JR, Wong YMM. Regional differences in DNA-synthetic activity in the preseptation myocardium of the chick. In: ClarkEB, Takao A, eds. Developmental Cardiology: Morphogenesis and Function. MtKisco, NY: Futura Publishing Co; 1990:219–234.
52. Arguello C, Delacruz MV, Sanchez C. Ultrastructural and experimentalevidence of myocardial cell differentiation into connective tissue cells inembryonic chick heart. J Mol Cell Cardiol. 1978;10:307–315.
53. Waldo K, Kumiski DH, Kirby ML. Association of the cardiac neural crestwith development of the coronary arteries in the chick embryo. Anat Rec.1994;239:315–331.
54. Takeda K, Yu ZX, Nishikawa T, Tanaka M, Hosoda S, Ferrans VJ,Kasajima T. Apoptosis and DNA fragmentation in the bulbus cordis of thedeveloping rat heart. J Mol Cell Cardiol. 1996;28:209–215.
55. Krstic R, Pexieder T. Ultrastructure of cell death in bulbar cushions ofchick embryo heart. Z Anat Entw Gesch. 1973;140:337–350.
56. Hurle JM, Ojeda JL. Cell death during the development of the truncus andconus of the chick embryo heart. J Anat. 1979;129:427–439.
57. Aikawa M, Sivam PN, Kuro-o M, Kimura K, Nakahara K, Takewaki S,Ueda M, Yamaguchi H, Yazaki Y, Periasamy M, Nagai R. Human smoothmuscle myosin heavy chain isoforms as molecular markers for vasculardevelopment and atherosclerosis. Circ Res. 1993;73:1000–1012.
58. Kuro-o M, Nagai R, Nakahara K, Katoh H, Tsai RC, Tsuchimochi H,Yazaki Y, Ohkubo A, Takaku F. cDNA cloning of a myosin heavy chainisoform in embryonic smooth muscle and its expression during vasculardevelopment and in arteriosclerosis. J Biol Chem. 1991;266:3768–3773.
59. Sakurai H, Matsuoka R, Furutani Y, Imamura S, Takao A, Momma K.Expression of four myosin heavy chain genes in developing blood vesselsand other smooth muscle organs in rabbits. Eur J Cell Biol. 1996;69:166–172.
60. Takahashi M, Kawamoto S, Adelstein RS. Evidence for inserted sequencesin the head region of nonmuscle myosin specific to the nervous system:cloning of the cDNA encoding the myosin heavy chain-B isoform ofvertebrate nonmuscle myosin. J Biol Chem. 1992;267:17864–17871.
61. Schilham MW, Oosterwegel MA, Moerer P, Jing Y, de Boer PAJ, VerbeekS, Lamers WH, Kruisbeek AM, Cumano A, Clevers H. Sox-4 gene isrequired for cardiac outflow tract formation and pro-B lymphocyteexpansion. Nature. 1996;380:711–714.
62. Franz T. Persistent truncus arteriosus in the Splotch mutant mouse. AnatEmbryol. 1989;180:457–464.
63. Mendelsohn C, Lohnes D, Decimo D, Lufkin T, LeMeur M, Chambon P,Mark M. Function of the retinoic acid receptors (RARs) during devel-opment, II: multiple abnormalities at various stages of organogenesis inRAR double mutants. Development. 1994;120:2749–2771.
64. Kastner P, Grondona JM, Mark M, Gansmuller A, LeMeur M, Decimo D,Vonesch P, Dolle P, Chambon P. Genetic analysis of RXRa develop-mental function: convergence of RXR and RAR signalling pathways inheart and eye morphogenesis. Cell. 1994;78:987–1003.
472 Development of Outflow Tract in Rat
by guest on January 22, 2015http://circres.ahajournals.org/Downloaded from

Antoon F. M. Moorman and Wouter H. LamersJing Ya, Maurice J. B. van den Hoff, Piet A. J. de Boer, Sabina Tesink-Taekema, Diego Franco,
Normal Development of the Outflow Tract in the Rat
Print ISSN: 0009-7330. Online ISSN: 1524-4571 Copyright © 1998 American Heart Association, Inc. All rights reserved.is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation Research
doi: 10.1161/01.RES.82.4.4641998;82:464-472Circ Res.
http://circres.ahajournals.org/content/82/4/464World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circres.ahajournals.org//subscriptions/
is online at: Circulation Research Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer about this process is available in the
located, click Request Permissions in the middle column of the Web page under Services. Further informationEditorial Office. Once the online version of the published article for which permission is being requested is
can be obtained via RightsLink, a service of the Copyright Clearance Center, not theCirculation Researchin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on January 22, 2015http://circres.ahajournals.org/Downloaded from




















