
Immunohistochemical lozalization of estrogen receptor in the normal canine female genital tract
15
ELSEVIER DOMESTIC ANIMAL ENDOCRINOLOGY Vol. 14(3):133-147, 1997 IMMUNOHISTOCHEMICAL LOCALIZATION OF ESTROGEN RECEPTOR IN THE NORMAL CANINE FEMALE GENITAL TRACT H. De Cock, R. Ducatelle, and J.P. Logghe Department of Veterinary Pathology University of Ghent (RUG) Ghent, Belgium Received October 31, 1995 Accepted December 13, 1996 The distribution of estrogen receptor (ER) in the ovaries and uterus was.studied throughout the estrous cycle in the bitch. The stage in the estrous cycle of 25 dogs was assessed by gross and histologic appearance of the uterus and ovaries and blood steroid hormone values. Demonstration of ER was performed by an indirect immunohistochemical technique with monoclonal antibodies. ER was found as a red nuclear staining in the surface, crypt, and glandular epithelium; in endometrial stroma cells; and in smooth muscle cells of the tunica muscularis of the uterus. Total scores of ER-positive cells varied during the estrous cycle, with the highest scores in the early proliferative stage and the lowest scores in the early secretory stage of the estrous cycle. These results are in concordance with the observations in other animals and the human uterus. In the canine ovaries, positive staining for ER was present in surface epithelium and subsurface epithelial ingrowths, which is in contrast with negative staining of human ovaries. This study confirms the earlier hypothesis that the epithelial ingrowths in canine ovaries may be responsive to blood steroid hormones. © Elsevier Science Inc. 1997 INTRODUCTION A key to the diagnosis and therapy of hormonal imbalances and related diseases is the understanding of normal biochemical pathways and physiologic responses. In the human, the relationship between cyclic changes in histomorphology and the distribution of estrogen receptor (ER) in the female genital tract has been studied extensively. ER has been found in the nuclei of epithelial, stromal, and smooth muscle cells of the reproductive tract in both postmenopausal woman and women of reproductive age (1-3). During the menstrual cycle, the epithelial and stromal cells of the endometrium functionales show substantial changes in the pattern of staining for ER (4,5). Such ER studies have proved to be of great value in the investigation ot~ endocrine disturbances, tumor pathology, and therapy (6-11). The expression of ER in tumors seems to have a predictive value of the malignancy both in endometfial and in breast carcinomas (8,9,11-14). The estrous cycle in the bitch is monestral. The canine interestrus (estrus to estrus) period is about 7 mo. The estrous cycle is divided into proestrus, estrus, metestrus, and anestrus. The bitch differs from other species in that the corpora lutea already develop before ovulation and remain functional for approximately the same length of time (50- 80 d), whether pregnancy occurs or not. Moreover, in the bitch, a long period of anestrus (60-240 d) follows the demise of the corpus luteum (15,16). Taking into account these peculiarities of the estrous cycle in the dog, it was considered © ElsevierScience Inc. 1997 0739-7240/97/$17.00 655 Avenue of the Americas, New York, NY 10010 PII S0739-7240(97)00001-5
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Immunohistochemical lozalization of estrogen receptor in the normal canine female genital tract

ELSEVIER
DOMESTIC ANIMAL ENDOCRINOLOGY Vol. 14(3):133-147, 1997
IMMUNOHISTOCHEMICAL LOCALIZATION OF ESTROGEN RECEPTOR IN THE NORMAL CANINE FEMALE
GENITAL TRACT
H. De Cock, R. Ducatelle, and J.P. Logghe
Department of Veterinary Pathology University of Ghent (RUG)
Ghent, Belgium
Received October 31, 1995 Accepted December 13, 1996
The distribution of estrogen receptor (ER) in the ovaries and uterus was.studied throughout the estrous cycle in the bitch. The stage in the estrous cycle of 25 dogs was assessed by gross and histologic appearance of the uterus and ovaries and blood steroid hormone values. Demonstration of ER was performed by an indirect immunohistochemical technique with monoclonal antibodies. ER was found as a red nuclear staining in the surface, crypt, and glandular epithelium; in endometrial stroma cells; and in smooth muscle cells of the tunica muscularis of the uterus. Total scores of ER-positive cells varied during the estrous cycle, with the highest scores in the early proliferative stage and the lowest scores in the early secretory stage of the estrous cycle. These results are in concordance with the observations in other animals and the human uterus. In the canine ovaries, positive staining for ER was present in surface epithelium and subsurface epithelial ingrowths, which is in contrast with negative staining of human ovaries. This study confirms the earlier hypothesis that the epithelial ingrowths in canine ovaries may be responsive to blood steroid hormones. © Elsevier Science Inc. 1997
I N T R O D U C T I O N
A key to the diagnosis and therapy of hormonal imbalances and related diseases is the understanding of normal biochemical pathways and physiologic responses. In the human, the relationship between cyclic changes in his tomorphology and the distribution of estrogen receptor (ER) in the female genital tract has been studied extensively. ER has been found in the nuclei of epithelial, stromal, and smooth muscle cells of the reproductive tract in both postmenopausal woman and women of reproductive age (1-3). During the menstrual cycle, the epithelial and stromal cells of the endometrium functionales show substantial changes in the pattern of staining for ER (4,5).
Such ER studies have proved to be of great value in the investigation ot ~ endocrine disturbances, tumor pathology, and therapy (6-11) . The expression of ER in tumors seems to have a predictive value of the malignancy both in endometfial and in breast carcinomas (8,9,11-14).
The estrous cycle in the bitch is monestral. The canine interestrus (estrus to estrus) period is about 7 mo. The estrous cycle is divided into proestrus, estrus, metestrus, and anestrus. The bitch differs from other species in that the corpora lutea already develop before ovulation and remain functional for approximately the same length of t ime ( 5 0 - 80 d), whether pregnancy occurs or not. Moreover, in the bitch, a long period of anestrus (60 -240 d) follows the demise of the corpus luteum (15,16).
Taking into account these peculiarit ies of the estrous cycle in the dog, it was considered
© Elsevier Science Inc. 1997 0739-7240/97/$17.00 655 Avenue of the Americas, New York, NY 10010 PII S0739-7240(97)00001-5

134 DE COCK ET AL.
worthwhile to study the ER distribution in the uterus and ovaries in this species. In 1977, Greene et al. (17) proved by a radioimmunoassay technique that the ER from various mammalian species have similar antigenic sites. This was confirmed by Traish et al. in 1990 (18).
The technique for the immunohistochemical localization of ER in tissue has been optimized in rodent laboratory animals (1,2,19). Examination of the cellular localization of ER in male and female primates was done by Brenner et al. (20) and by McLellan et al. (21). In domestic animals, studies of ER distribution and changes in the endometrium during different stages of the estrous cycle were performed in the mare (22), the ewe (23), the pig (24), and the cat (25). ER have been investigated immunohistochemically in the male dog in normal and hyperplastic prostatic tissue (26). In the bitch, changes of cytosolic ER in the uterus during the reproductive cycle and in response to estrogen and progesterone treatment were only examined with a receptor-binding assay (27,28). The purpose of this study was to investigate the ER distribution in the canine ovary and uterus throughout the estrous cycle by use of an immunohistochemical technique.
MATERIALS AND METHODS
Tissue and Blood Samples. Samples from 25 healthy adult female dogs presented for ovariohysterectomy were collected at the time of surgery. Surgery was accomplished after sedation with Thalamonal (Janssens Pharmaceutica, Beerse, Belgium) and premedication with atropine, followed by an intravenous injection of pentothal sodium (Abbott Labo- ratories, Chicago, IL). Anesthesia was maintained during surgery with fluothane (Zeneca, Destelbergen, Belgium). Samples from the following parts of the female genital tract were taken for further investigation: both ovaries, left and right uterine horns and corpus uteri. A blood sample was taken before sedation for the measurement of sex steroid hormone levels. The stage in the estrous cycle was determined according to the histologic aspect of the ovaries and uterus together with the blood serum steroid hormone levels.
Sample treatment. The tissue samples were fixed immediately in 10% formalin for 48 hr and thereafter routinely processed in a tissue processor (Shandon Elliott duplex processor, Type SCE-0545, Shandon LTD, Camberley, U.K.). In brief, this consisted of dehydration in ethanol, clearing in xylene, and embedding in paraffin. Sections of 5-pm thickness were cut, mounted on 3-aminopropyltriethoxysilane (Sigma, St Louis, MO)- coated slides, and dried overnight at 60 ° C.
Serum Steroid Hormone Measurement. For estradiol, the IMx Estradiol Micropar- ticle Enzyme Immunoassay technique (Abbott Laboratories, Abbott Park, IL, USA) was used. Testosterone and progesterone were measured, by a solid-phase radioimmunoassay, based on, respectively, a testosterone and progesterone-specific antibody immobilized to the wall of a polypropylene tube (Diagnostic Products Corporation, Los Angeles) (15).
Immunohistochemical Procedure. After deparaffinizing in xylene and rehydration (four steps: 100% alcohol, 95% alcohol, 50% alcohol to distilled water), an antigen retrieval technique was performed. In this procedure, slides were placed in Antigen Retrieval Citra Solution (BioGenex, San Ramon, CA), microwaved at high power (750 W) for 4 min, and then kept boiling lbr l 0 rain at 250 W. The tissues were kept in the antigen retrieval solution for 30 min at 4 ° C. Distilled water was added if necessary to prevent the slides from drying out. Hereafter, the slides were rinsed in phosphate-buffered saline (PBS) at pH 7.6 for 15 min. A mouse monoclonal antibody to human ER (clone 1D5; BioGenex), diluted 1/10 in buffered glycine phosphate, pH 7.1, with 6% bovine serum albumin (BioGenex), was applied overnight at 20 ° C.
After rinsing for 15 min with PBS, pH 7.6, the slides were incubated for 30 min with biotinylated goat anti-mouse antibodies (Link, supersensitive kit; BioGenex). After an-

ER IN CANINE FEMALE GENITAL TRACT 135
TABLE 1. MEAN BLOOD STEROID HORMONE LEVELS IN THE DIFFERENT STAGES OF THE ESTROUS CYCLE.
Mean Blood Mean Blood Endometrial Dating Progesterone Levels Estradiol Levels
(n) Ovaries (ng/ml) (_+SE) (pg/ml) (-+SE)
Anestrus (4) PF, SF 0.13 (-+0.03) 51 (-+3) Early proliferative (4) TF 0.36 (-+ 0.21) 60 (-+ 13) Late proliferative (4) young CL 9,55 (-+2.98) 61 (-+24) Early secretory (3) young CL 27.58 (29.88) 138 (-+51) Late secretory (8) old CL 0.34 (-+0.34) 57 (_+ 12) Pregnant (2) young CL 25.15 (-+7.99) 63 (-+3)
Progesterone and estradiol levels in blood during the different stages of the estrous cycle. The values are expressed as means -+ SE. The following abbreviations are used: n, number of dogs observed; PF, primary follicle; SF, secondary follicle; TF, tertiary follicle; CL, corpus luteum.
other rinse of 10 min, an alkaline phosphatase-conjugated streptavidin (Label, supersen- sitive kit; BioGenex) was applied as indicated by the manufacturer. After a final wash, substrate solution (5 mg of Fast Red in 5 ml of naphthol-phosphate/Tris-HC1 buffer; Fast red substrate pack; Biogenex) was added for 10 min. Nuclear counterstain was achieved by incubation with Mayer ' s hematoxylin for 10 sec,
Posit ive control tissue consisted of human breast tissue known to possess ER. More- over, samples of expected positive (uterus) and negative canine tissue (mesenteric lymph nodes) were incubated with the selected antibody (MU272-UC; BioGenex). Addit ional controls included incubation with irrelevant antibodies, the antibody diluent (glycine phosphate buffered at pH 7.1 with 6% bovine serum albumin; Biogenex), and PBS in the absence of primary antibodies.
Scor ing of I m m u n o s t a i n i n g . The expression levels for grading the immunostaining of ER were calculated by the following formula:
TS = PS + IS
In which PS stands for the proportion score, IS is the intensity score, and TS stands for the total score.
The PS numbers the cells that have been stained. Zero means no positive cells, 1 is between 0 and 1 posit ive cells for 100 negative cells, and 2 is given for a proportion of posit ive to negative cells between 1 in 100 and 1 in 10. In the same way, 3 ranges from 1 in 10 to 1 in 3, 4 ranges from 1 in 3 to 2 in 3, and finally, 5 is between 2 in 3 and all cells posit ive (1 in 1). IS indicates the degree of intensity of staining and is given as follows: O is negative, 1 is a weak staining, 2 is intermediate staining, and 3 is strong staining. The sum of these two numbers is the TS, ranging from 0 to a maximum level of 8 (note that 1 cannot be given).
S ta t i s t ica l Analys is . Statistical analysis of the differences in total score between the different stages in the estrous cycle was performed by the Kruskal-Wall is , nonparametric test.
R E S U L T S
Es t ima t i on of Stage in Es t rous Cycle. The results of staging of the estrous cycle are summarized in Table 1. Dogs are classified from anestrus to early proliferative phase, proliferative phase, and secretory phase. Four dogs were in the anestrus stage of the estrous cycle. Endometr ia consistent with the proliferative phase were found in eight dogs. Eleven dogs were in the secretory phase of the estrous cycle. Two dogs were pregnant.

136 DE COCK ET AL.
The results of blood progesterone and estradiol measurements are also given in Table 1. Blood testosterone levels were in all cases lower than 5 ng/dl, except in one pregnant dog with a value of 5 ng/dl.
Immunohistochemical Localization of ER in the Canine Female Reproductive Tract. Positive staining for ER was observed as a dark-red nuclear staining in epithelial, stromal (Figure la), and smooth muscle cells (Figure lb). The intensity of specific staining and the proportion of positive to negative cells varied between the different layers of the uterus wall and between the dogs. Lymphocytes, endothelial cells (Figure lb), and bands of connective tissue in the myometrium and lining the epithelium of the serosa were always negative. In the negative control tissue, incubated with PBS, pH 7.6, nonsense antiserum, or buffered glycine phosphate, pH 7.1, with 6% bovine serum albumin (BioGenex), instead of the primary antiserum, no red nuclear staining was observed (Figure 2). In the positive control tissue, nuclear staining was always present. Nonspecific background staining was limited to the serum in the blood vessels and glycogen-contain- ing cytoplasmic granules present in the uterine epithelium during metestrus (see Figure 9). This aspecific staining did not interfere with the specific staining.
Ovary. The ovaries of all dogs under investigation were lined by a low cuboidal surface epithelium. This epithelium became multilayered with prominent subsurface epithelium ingrowths (SKI) during anestrus, early proliferative phase, and late secretory phase. In anestrus, several small primary and secondary follicles were seen in the ovarian stroma. In the early proliferative phase, the follicles were large, tertiary follicles with preovulatory luteinization. In the late proliferative phase, in the early secretory phase, and in the pregnant dogs, the ovaries contained several large corpora lutea. In the late secretory phase, the corpora lutea were atrophic. Histologic and immunohistochemical observations were identical in the left and right ovary. A summary of the nuclear staining scores in the ovaries is listed in Table 2. ER was found in the surface epithelium and in the SKI (Figure 3). No positive staining could be found in any other part of the ovarian tissue, independent of the stage in the estrous cycle.
Significant differences (P < 0.05) were found between the different stages of the estrous cycle in the lining epithelium and SKI. The highest total scores for ER in the ovary were found in the epithelium of dogs with early proliferative and late secretory phase endometrium (Figure 3). Positive and negative cells were present in the same tissue. Many of the positive cells were situated in the stratum basale and particularly at the edge of the SKI.
During the late proliferative and early secretory phases, the total score of ER-responsive cells in the epithelium decreased (Figure 4). SKI were often not present, and if they were, they stained only weakly. One of the pregnant dogs had small amounts of ER in the epithelium and SKI; the other dog stayed completely negative.
Uterus. The endometrium is divided in three zones, in accordance with McEntee (29): the zone of crypts (CZ), the intermediate zone (IZ), and the basal zone (BZ). The CZ included the single lining epithelium with short, epithelial-lined recesses, the crypts. The IZ was composed predominantly of connective tissue. The BZ consisted mainly of glands that follow cyclic changes. From the three different places of the uterus observed, the staining was always more pronounced in the corpus than in the uterus horns.
In anestrus to early proliferative phase endometria, positive reaction increased gradually from faint nuclear staining (Figure 5) to strong and homogeneous coloring of the nuclei of the cells from the lining and glandular epithelium in the early proliferative stage (Figure 6). In anestrus, not all surface epithelial cells contain ER and the positive cells were more or less grouped together. In the endometrial glands, ER reactivity was always more
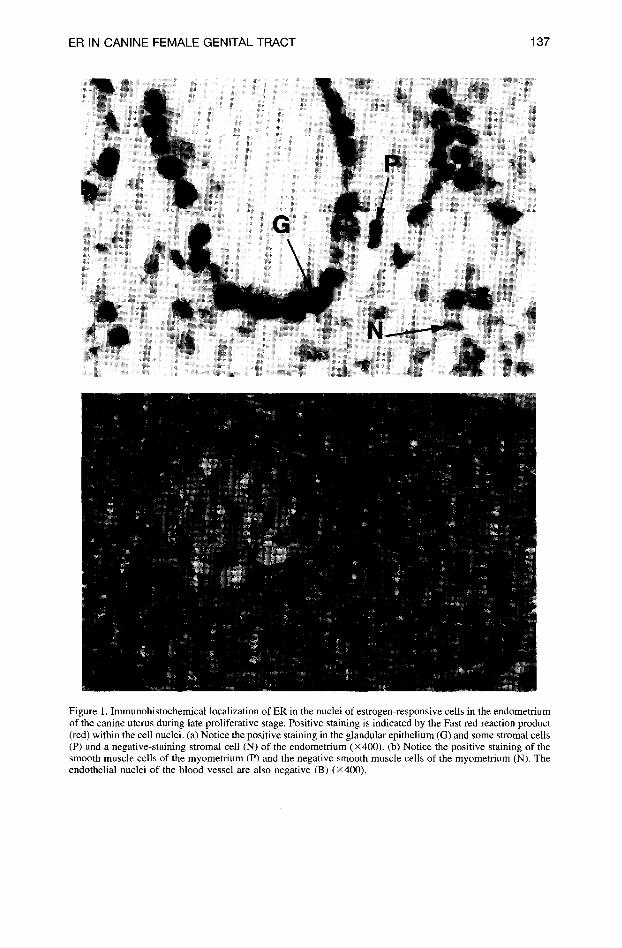
ER IN CANINE FEMALE GENITAL TRACT 137
Figure 1. Immunohistochemical localization of ER in the nuclei of estrogen-responsive cells in the endometrium of the canine uterus during late proliferative stage. Positive staining is indicated by the Fast red reaction product (red) within the cell nuclei. (a) Notice the positive staining in the glandular epithelium (G) and some strornal cells (P) and a negative-staining stromal cell (N) of the endometrium (x400). (b) Notice the positive staining of the smooth muscle cells of the myometrium (P) and the negative smooth muscle cells of the myometrium (N). The endothelial nuclei of the blood vessel are also negative (B) (x400).

138 DE COCK ET AL.
Figure 2. Negative control. Canine uterine tissue incubated with buffered glycine phosphate, pH 7.1, with 6% bovine serum albumin instead of the primary antiserum. No positive nuclear staining is observed (X 160).
TABLE 2. MEAN TOTAL SCORE FOR ER-SPECIFIC STAINING OF OVARIES AND UTERUS IN THE DIFFERENT STAGES OF THE ESTROUS CYCLE,
Ovaries Uterus
OE SKI CZ IZ BZ TM Endometrial ( -+ SE) ( -+ SE) ( -+ SE) ( -+ SE) ( -+ SE) ( -+ SE)
Dating I/T I/T I/T I/T I/T I/T
Anestrus 5.25 5.67 4.25 3.00 3.50 3.75 to early (-+0.48) (-+0.33) (-+ 1.32) (+0.58) (-+0.65) (-+0.48) proliferative 0.33 0.35 0.31 0.33 0.29 0.40
Early 4.40 4.75 5.40 4.60 5.20 5.20 proliferative (-+0.51) (-+0.75) (-+0.68) (-+0.51) (-+0.58) (-+0.66)
0.41 0.32 0.26 0.39 0.27 0.35 Late 2.67 2.00 2.00 0.67 3.33 1.67
proliferative (-+0.33) (-+0.00) (-+0.84) (+0.47) (-+1.33) (-+0.62) 0.37 0.50 0.50 0.49 0.40 0.40
Early secretory 2.33 0.00 2.66 2.33 5.00 2.33 (-+0.33) (-+0.00) (-+1.25) (-+0.33) (-+1.15) (-+0.33) 0.43 0.00 0.25 0.22 0.28 0.24
Late secretory 4.29 4.80 5.75 4.50 4.75 4.25 (-+0.52) (-+0.58) (-+0.49) (-+0.42) (+0.25) (-+0.31) 0.27 0.29 0.30 0.36 0.32 0.29
Pregnant 1.50 3 (a) 0.00 1.00 3.00 1.00 (-+2.12) (+0.00) (-+0.00) (+-0.00) (-+1.41) 0.33 0.33 0.00 0.50 0.33 0.50
OE, the values are expressed as means of the total score, -+SE observed in the different stages of the estrous cycle; I/T, the proportion of the intensity score to the total score 'a) score based on measurements in one dog; in the other dog, SKI were not present. The following abbreviations are used: OE, ovarian epithelium; TM, tunica muscularis.

ER IN CANINE FEMALE GENITAL TRACT 139
Figure 3. Immunohistochemical localization of ER in the ovary during late proliferative stage. Positive staining is present in surface epithelium (E) and SKI (X400).
pronounced at the base of the glands. Crypts of the CZ had the tendency to stain stronger than the basal glands.
Endometrial stroma cells and smooth muscle cells stained less intensely than the surface and the glandular epithelium, but the proportion of positive cells was high. Positive and negative endometrial stromal cells and smooth muscle cells were randomly distributed. In the tunica muscularis, there was a tendency for the inner layers to stain more intensely than the subserosal layers.
From the early proliferative phase to the late proliferative phase, there was a decrease in nuclear staining in all layers of the endometrium as well as in the myometrium (Figures 7 and 8). The most prominent staining in late proliferative phase was found in the glands of the BZ; this is in contrast to the situation described in the early proliferative stage. In late proliferative stage, the proportion of the intensity score to total score of ER was remarkably higher in all layers of the uterus than in all other stages under investigation. During early secretory phase, the progesterone levels in the blood were highest (Table 1).
With the onset of the late secretory phase and back to anestrus, the staining again appeared more intense and uniform throughout the uterine wall (Figure 9). The stromal
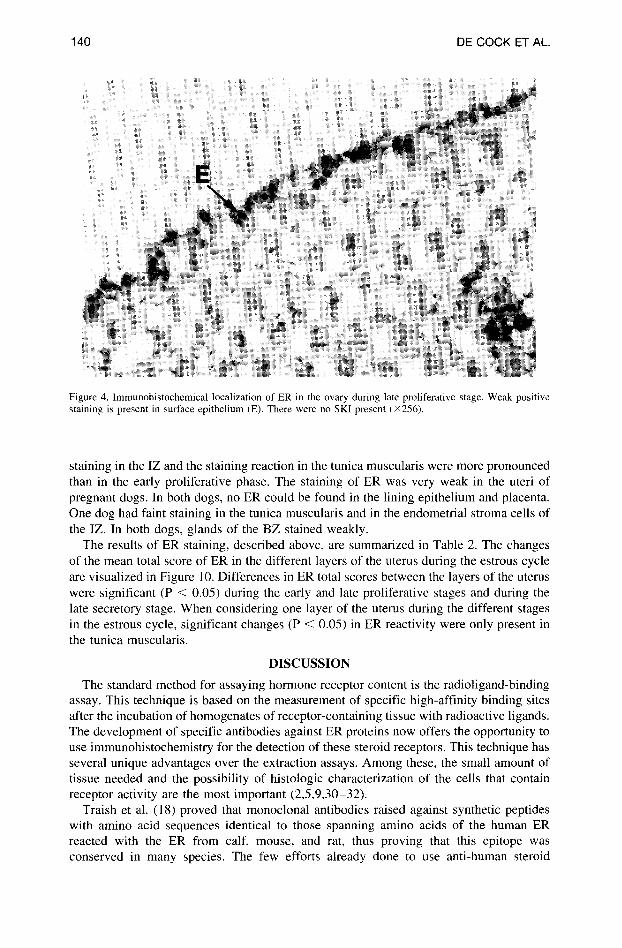
140 DE COCK ET AL.
Figure 4. lmmunohistochemical localization of ER in the ovary during late proliferative stage. Weak positive staining is present in surface epithelium (E). There were no SKI present (×256).
staining in the IZ and the staining reaction in the tunica muscularis were more pronounced than in the early proliferative phase. The staining of ER was very weak in the uteri of pregnant dogs. In both dogs, no ER could be found in the lining epithelium and placenta. One dog had faint staining in the tunica muscularis and in the endometrial stroma cells of the IZ. In both dogs, glands of the BZ stained weakly.
The results of ER staining, described above, are summarized in Table 2. The changes of the mean total score of ER in the different layers of the uterus during the estrous cycle are visualized in Figure 10. Differences in ER total scores between the layers of the uterus were significant (P < 0.05) during the early and late proliferative stages and during the late secretory stage. When considering one layer of the uterus during the different stages in the estrous cycle, significant changes (P < 0.05) in ER reactivity were only present in the tunica muscularis.
D I S C U S S I O N
The standard method for assaying hormone receptor content is the radioligand-binding assay. This technique is based on the measurement of specific high-affinity binding sites after the incubation of homogenates of receptor-containing tissue with radioactive ligands. The development of specific antibodies against ER proteins now offers the opportunity to use immunohistochemistry for the detection of these steroid receptors. This technique has several unique advantages over the extraction assays. Among these, the small amount of tissue needed and the possibility of histologic characterization of the cells that contain receptor activity are the most important (2,5,9,30-32).
Traish et al. (18) proved that monoclonal antibodies raised against synthetic peptides with amino acid sequences identical to those spanning amino acids of the human ER reacted with the ER from calf, mouse, and rat, thus proving that this epitope was conserved in many species. The few efforts already done to use anti-human steroid

ER IN CANINE FEMALE GENITAL TRACT 141
Figure 5. Anestrus endometria with a weak nuclear staining in the glandular epithelium (G) and stromal cells (F) (x400).
receptor antibodies on animal tissue also show that the estradiol receptor is a highly conserved protein (22-26). In this study, we demonstrated the cross-reactivity of com- mercially available anti-human ER antibodies with the canine ER, thus confirming again the conservative nature of this protein.
Literature data on ER analysis of animal uterus hitherto give little detailed information on the changes during normal cyclicity (22-25). Most studies are done during exogenous hormone therapy (23,25). Therefore, it is difficult to compare our observations with the results from earlier investigations. Most detailed descriptions of ER changes in the uterus during the estrous cycle are found in humans (4,5) and primates (20,21).
The intracellular localization of ER in the nuclei is in concordance with the observa- tions in humans (2,3,5) and other animals (22-25). ER-containing cells in the uterus are endometrial surface and glandular epithelium, endometrial stroma cells of the propria, and smooth muscle cells of the tunica muscularis.
The intensity and distribution of the ER varied according to the dating of the endome- trium. During the early proliferative phase, a strong and ubiquitous staining for ER was observed in surface and glandular epithelium, as well as in the endometrial stromal and

142 DE COCK ET AL.
Figure 6. Early proliferative stage endometrium. Note homogeneous coloring of the nuclei of the cells from the lining (E) and glandular epithelium (G) and stromal cells (F) (×400).
myometr ial smooth muscle cells. During the late proliferative phase, after the rising blood progesterone concentrations, positive staining was reduced in both endometrium and myometrium. In the glandular epithelium of the BZ, staining intensity decreased less than in the CZ and IZ. All of these observations are in accordance with the findings in humans (4,5) primates (20,21), and sheep (23).
The rising progesterone concentrations are important for the sudden change in the staining pattern of ER in the endometrium and myometr ium (21,23,25,27). Estradiol seems to be less important for this phenomenon because it was seen with or without raised estradiol serum concentrations. These findings are in agreement with those of Lessey et al. (27), who demonstrated a decrease in ER during progesterone treatment with or without concomitant estrogen administration.
The onset of the decline in ER in the canine uterus starts before ovulation. The latter observation in the uterus from bitches is never demonstrated in the human (4,5), primate (20,21), mare (22), ewe (23), pig (24), or cat (25). This phenomenon can be explained by the differences in blood steroid hormone values during estrus in the dog. Tertiary follicles in the ovary of the bitch already show luteinization and secrete progesterone during estrus.

ER IN CANINE FEMALE GENITAL TRACT 143
Figure 7. Immunohistochemical localization of ER in the endometrium of the uterus during early secretory stage. Notice the absence of ER in the proliferated glands (G) in the BZ of the endometrium (×400).
Consequently, detectable progesterone values are already present before ovulation (to- gether with high estradiol levels); this is in contrast to other species.
In late secretory and anestrus phase, our observations in the canine differ slightly from those in other species. Strong staining is observed in all layers of endometrium and tunica muscularis, whereas in humans and female primates, only weak staining is observed (4,5,20,21). These differences may be related to differences in the estrous cycle between human and canine species. In the human, the superficial layer of the endometrium is lost during the menses; in the female dog, this tissue remains intact and goes into a long period of anestrus. In this view, the similarity between ER distribution during the anestrus of the dog and ER appearance in postmenopausal woman (4,5,33) is not surprising.
Primate and human ovaries are completely negative for ER (20,34). Nevertheless, ER was demonstrated in human ovarian carcinomas (34). In this study, ER was demonstrated in the ovarian SKI and surface epithelium in dogs. The staining score followed the involution (less ER) and hypertrophy (high score of ER) of these SKI during the estrous cycle. Until now, little was known about the function of these structures. The fact that ER

144 DE COCK ET AL.
Figure 8. Detail from the basal glands shown in Figure 7. Notice the slight positive coloring of the nuclei in the basal glands• The red granules in the cytoplasm are nonspecific staining, also observed in the control slides with irrelevant antibody (X400).
is present supports the hypothesis of an endocrine involvement, as proposed by McEntee (29).
In conclusion, in this study, the ER distribution in the canine female genital tract is described throughout the estrous cycle in the adult bitch. Differences were shown in ER distribution during the late secretory phase as compared with the human uterus. Further investigation of ER distribution in tissues from bitches with endometrial disturbances such as hyperplasia and pyometra may lead to a better understanding of the pathogenesis of these clinically important entities in the bitch. Furthermore, exploring ER changes in the anestrus stage of the bitch may lead to a better understanding of tissue changes in the postmenopausal endometrium of women.
ACKNOWLEDGMENTS/FOOTNOTES
We thank Hilde De Bie from Klinipath, Belgium, for her technical advice. We are also grateful to the Department of Small Animal Medicine and the veterinarians who helped in collecting the tissues. We thank Christian Puttevils for photography.
Author of correspondence: Dr. H. De Cock, Department of Veterinary Pathology, University of Genth 24, 9000 Ghent, Belgium.
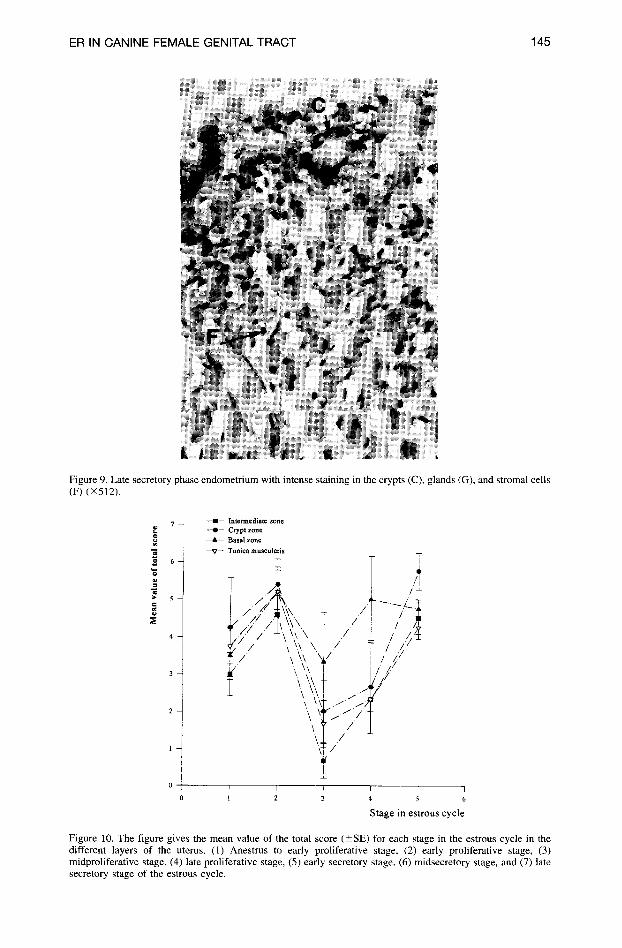
ER IN CANINE FEMALE GENITAL TRACT 145
Figure 9. Late secretory phase endometrium with intense staining in the crypts (C), glands (G), and stromal cells (F) (×512).
7 - -
6 -
--I1-- Intermediate zone --41-- Crypt zone
~ t . - - Basal zone
- -V- - Tuniea m u s c u l a r i s
:.i / ~ / / \:k\ ~- . /
\.~ /
I I
1 2 I I I
4 5 6
Stage in estrous cycle
Figure 10. The figure gives the mean value of the total score (-+SE) for each stage in the estrous cycle in the different layers of the uterus. (1) Anestrus to early proliferative stage, (2) early proliferative stage, (3) midproliferative stage, (4) late proliferative stage, (5) early secretory stage, (6) midsecretory stage, and (7) late secretory stage of the estrous cycle.

146 DE COCK ET AL.
REFERENCES
l. Greene GL, Jensen EV. Monoclonal antibodies as probes for estrogen receptor detection and characteriza- tion. J Steroid Biochem 16:353-359, 1982.
2. King W J, Greene GL. Monoclonal antibodies localize oestrogen receptor in the nuclei of target ceils. Nature 307:745-747, 1984.
3. Press MF, Greene GL. Methods in laboratory investigation. An immunohistochemical method for demon- strating estrogen receptor in human uterus using monoclonal antibodies to human estrophilin. Lab Invest 50:480-486, 1984.
4. Press MF, Nousek-Goebel N, King WJ, Herbst AL, Greene GL. Immunohistochemical assessment of estrogen receptor distribution in the human endometrium throughout the menstrual cycle. Lab Invest 51:495-503, 1984.
5. Snijders MPML. Oestrogen and progesterone receptors in the human uterus. PhD thesis, University of Limburg at Maastricht, The Netherlands, 1992.
6. Janne O, Kauppila A, Kontula K, Syrjala P, Vihko R. Female sex steroid receptors in normal, hyperplastic and carcinomatous endometrium. The relationship to serum steroid hormones and gonadotropins and changes during medroxyprogesterone acetate administration. Int J Cancer 24:545-554, 1979.
7. Ehrlich CE, Young CM, Cleary RE. Cytoplasmic progesterone and estradiol receptors in normal hyper- plastic, and carcinomatous endometria: Therapeutic implications. Am J Obstet Gynecol 141:539-546, 1981.
8. Kauppila AJI, Isotalo HE, Kivinen ST, Vihko RK. Prediction of clinical outcome with estrogen and progestin receptor concentrations and their relationships to clinical and histopathological variables in endometrial cancer. Cancer Res 46:5380 5384, 1986.
9. McCarty KS Jr, Szabo E, Flowers JL, Cox EB, Leight GS, Miller L, Konrath J, Soper JT, Budwit DA, Creasman WT, Seigler HF, McCarty KS Sr. Use of a monoclonal anti-estrogen receptor antibody in the immunohistochemical evaluation of human tumors. Cancer Res 46:4244s-4248s, 1986.
10. Thowton JG, Wells M. Oestrogen receptor in glands and stroma of normal and neoplastic human endome- trium: A combined biochemical, immunohistochemical, and morphometric study. J Clin Pathol 40:1437- 1442, 1987.
11. Kuenen-Boumeester V, Van der Kwast TH, Van Putten WLJ, Claasen C, Van Ooijen B, Henzen-Logmans SC. Immunohistochemical determination of androgen receptors in relation to oestrogen and progesterone receptors in female breast cancer. Int J Cancer 52:581 584, 1992.
12. DeSombre ER, Greene GL, King W J, Jensen EV. Estrogen receptor antibodies and hormone dependent cancer. Hormones Cancer 5:1-2 l, 1984.
13. Ehrlich CE, Young CM, Stehman FB, Sutton GP, Alford WM. Steroid receptors and clinical outcome in patients with adenocarcinoma of the endometrium. Am J Obstet Gynecol 158:796-807, 1988.
14. Menendez-Bodet CJ, Schwartz MK. Estrogen and progesterone receptor proteins in patients with breast cancer. Adv Clin Chem 30:185-21, 1993.
15. Chastain CB, Ganjam VK. The Gonads. In: Clinical Endocrinology of Companion Animals, Chastain CB (ed). Lea & Febiger, Philadelphia, p. 439-534, 1986.
16. Pineda MH. Female reproductive system. In: Veterinary Endocrinology, McDonald LE and Pineda MH (eds). Lea & Febiger, Philadelphia, p. 303-355, 1989.
17. Greene GL, Closs LE, Fleming H, DeSombre ER, Jensen EV. Antibodies to estrogen receptor: Immuno- chemical similarity of estrophilin from various mammalian species. Proc Natl Acad Sci USA 74:3681-3685, 1977.
18. Traish AM, Ettinger R, Kim N, Marshak-Rothstein A, Wotiz HH. Development and characterization of monoclonal antibodies to specific domain of human estrogen receptor. Steroids 55:196-208, 1990.
19. Gee JMW, Nicholson RI, Jasani B, Newman GR, Amselgruber WM. An immunohistochemical method for the localization of estrogen receptors in rat tissues using a dinitropheny] (DNP)-labeled rat monoclonal primary antibody. J Histochem Cytochem 38:69-78, 1990.
20. Brenner RM, West NB, McClellan MC. Estrogen and progestin receptors in the reproductive tract of male and female primates. Biol Reprod 42:11-19, 1990.
21. McClellan M, West NB, Brenner RM. Immunohistochemical localization of estrogen receptors in the macaque endometrium during the luteal-follicular transition. Endocrinology 119:2467-2475, 1986.
22. Watson ED, Skolnik SB, Zanecosky HG. Progesterone and estrogen receptor distribution in the endome- trium of the mare. Theriogenology 38:575-580, 1992.
23. Cherny RA, Salamonsen LA, Findlay JK. Immunohistochemical localization of estrogen receptors in the endometrium of the ewe. Reprod Fertil Dev 3:321-331, 1991.
24. Stanchev P, Rodriguez-Martinez H, Edqvist LE, Eriksson H. Characterization of uterine sex steroid receptors in the pig and their variation during the oestrous cycle. J Steroid Biochem 35:689-699, 1990.

ER IN CANINE FEMALE GENITAL TRACT 147
25. Li W, Boomsma RA, Verhage HG. Immunohistochemical analysis of estrogen and progestin receptors in uteri of steroid-treated and pregnant cats. Biol Reprod 47:1073-1081, 1992.
26. Schulze H, Barrack R. Immunohistochemical localization of estrogen receptors spontaneous and experi- mentally induced canine benign prostatic hyperplasia. The Prostate 11:145-162, 1987.
27. Lessey BA, Wahawisan R, Gorell TA. Hormonal regulation of cytoplasmic estrogen and progesterone receptors in the beagle uterus and oviduct. Mol Cell Endocrinol 21:171-180, 1981.
28. Donnay I, Wouters-Ballman P, Vleeschonwer N, Leclercq G, Verstegen J. Changes in oestrogen, proges- terone and epidermal growth factor receptor concentrations and affinities during the oestrus cycle in the normal mammary gland and uterus of dogs. Vet Research Comm 19:101-113, 1995.
29. McEntee K. The uterus: Anatomy. In: Reproductive Pathology of Domestic Mammals, McEntee K (ed). Academic Press Inc., San Diego, p. 137-139, 1990.
30. Parl FF, Posey YF. Discrepancies of the biochemical and immunohistochemical estrogen receptor assays in breast cancer. Hum Pathol 19:960-966, 1988.
31. True LD. Atlas of Diagnostic Immunohistopathology, JB Lippincott Company, Philadelphia, p. 15.1-15.13, 1990.
32. Schutte HME, Scheres AFPM. Steroidreceptoren zichtbaar. Immunohistochemische en radiochemische bepaling van steroidreceptoren in borstkanker. Analyse 254-258, November 1993.
33. Press MF, Nousek-Goebel N, Bur M, Greene GL. Estrogen receptor localization in the female genital tract. Am J Pathol 123:280-292, 1986.
34. Press MF, Holt JA, Herbst AL, Greene GL. Immunohistochemical identification of estrogen receptor in ovarian carcinomas. Lab Invest 53:349-361, 1985.




















