
Molecular Properties of Structure and Regulation of the Calcium Channel
16
Molecular Properties of Structure and Regulation of the Calcium Channel“ MICHEL LAZDUNSKI,~ JACQUES BARHANIN,~ MARC BORSOTTO,~ CHRISTIAN COGNARD: CHRISTIE COOPER? THIERRY COPPOLA,~ MICHEL FOSSET,~ JEAN-PIERRE GALIZZI,~ MARLENE M. HOSEY,~ CHRISTIANE MOURRE,~, JEAN-FRANCOIS RENAUD,~ GEORGES ROMEY,~ ANNIE SCHMID,~ AND SYLVIE VANDAELE~ bBiochernistry Center National Centerfor Scientijc Research (CNRS) Pare Valrose 06034 Nice Cedex, France ‘General Physiology Laboratory 40 Avenue du Recteur Pineau 86021 Poitiers Cedex, France dUniversity of Health Sciences The Chicago Medical School 3333 Green Bay Road North Chicago, Illinois 60064 Voltage-dependent calcium channels are known to play important roles in excitation- contraction coupling in cardiac and smooth muscles and also in a number of neurosecretory processes. They have recently become accessible to biochemical study as a result of the availability of a number of tritiated calcium channel inhibitors belonging to the dihydropyridine and the phenylalkylamine series.’ Dihydropyridine derivatives such as nitrendipine and other calcium inhibitors such as verapamil, bepridil, and diltiazem are very important therapeutic agents in the treatment of cardiovascular disorders.’ One of the difficulties associated with the biochemical characterization of macro- molecules that confer electrical excitability to biological membranes, such as the voltage-sensitive calcium channel, is that they are almost always present in very low amounts in membranes. For most membrane preparations in which the voltage- sensitive calcium channel has been characterized, the dihydropyridine receptor is only present at a density in the range of 0.1 to 1 pmol/mg protein.’ Therefore, the transverse tubule (T-tubule) membrane preparation isolated from rabbit skeletal muscle appears to be exceptional in that it contains more than 50 pmol [3H]dihydropyridine-binding sites/mg p r ~ t e i n . ~ Voltage-clamp analyses of skeletal muscle have shown that essen- tially all specific Ca’+ conductances are localized in the transverse tubular system and that the channel responsible for these conductances is inhibited by low concentrations ‘This work was supported by the Association des Myopathes, the Centre National de la Recherche Scientifique, the Fondation sur les Maladies Vasculaires, the Fondation pour la Recherche Mkdicale, and the Ministere de I’Industrie et de la Recherche (grant no. 83.C.0696). 134
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Molecular Properties of Structure and Regulation of the Calcium Channel

Molecular Properties of Structure and Regulation of the Calcium Channel“
MICHEL LAZDUNSKI,~ JACQUES BARHANIN,~ MARC BORSOTTO,~ CHRISTIAN COGNARD:
CHRISTIE COOPER? THIERRY COPPOLA,~ MICHEL FOSSET,~ JEAN-PIERRE GALIZZI ,~
MARLENE M. HOSEY,~ CHRISTIANE MOURRE,~, JEAN-FRANCOIS RENAUD,~ GEORGES ROMEY,~
ANNIE SCHMID,~ AND SYLVIE VANDAELE~ bBiochernistry Center
National Center for Scientijc Research (CNRS) Pare Valrose
06034 Nice Cedex, France ‘General Physiology Laboratory
40 Avenue du Recteur Pineau 86021 Poitiers Cedex, France
dUniversity of Health Sciences The Chicago Medical School
3333 Green Bay Road North Chicago, Illinois 60064
Voltage-dependent calcium channels are known to play important roles in excitation- contraction coupling in cardiac and smooth muscles and also in a number of neurosecretory processes. They have recently become accessible to biochemical study as a result of the availability of a number of tritiated calcium channel inhibitors belonging to the dihydropyridine and the phenylalkylamine series.’ Dihydropyridine derivatives such as nitrendipine and other calcium inhibitors such as verapamil, bepridil, and diltiazem are very important therapeutic agents in the treatment of cardiovascular disorders.’
One of the difficulties associated with the biochemical characterization of macro- molecules that confer electrical excitability to biological membranes, such as the voltage-sensitive calcium channel, is that they are almost always present in very low amounts in membranes. For most membrane preparations in which the voltage- sensitive calcium channel has been characterized, the dihydropyridine receptor is only present at a density in the range of 0.1 to 1 pmol/mg protein.’ Therefore, the transverse tubule (T-tubule) membrane preparation isolated from rabbit skeletal muscle appears to be exceptional in that it contains more than 50 pmol [3H]dihydropyridine-binding sites/mg p r ~ t e i n . ~ Voltage-clamp analyses of skeletal muscle have shown that essen- tially all specific Ca’+ conductances are localized in the transverse tubular system and that the channel responsible for these conductances is inhibited by low concentrations
‘This work was supported by the Association des Myopathes, the Centre National de la Recherche Scientifique, the Fondation sur les Maladies Vasculaires, the Fondation pour la Recherche Mkdicale, and the Ministere de I’Industrie et de la Recherche (grant no. 83.C.0696).
134
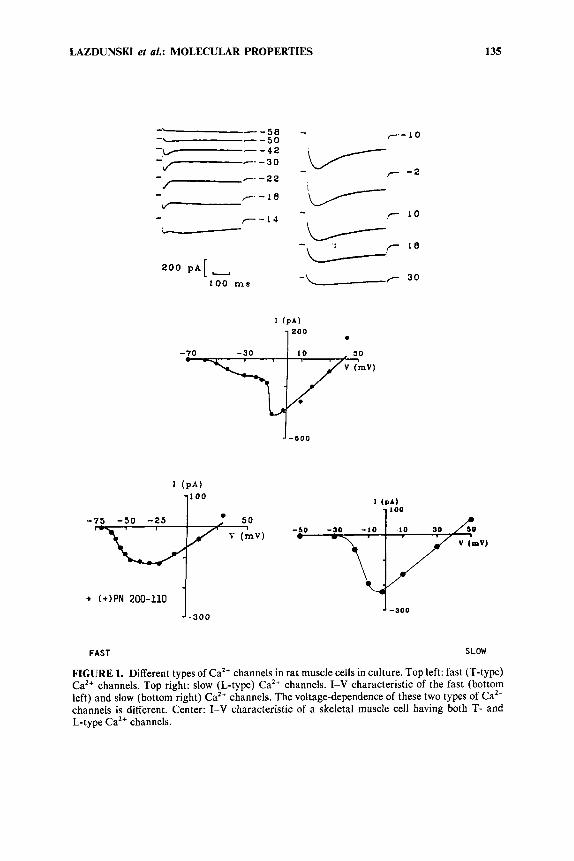
LAZDUNSKI et al.: MOLECULAR PROPERTIES 135
+ (+)PN 200-110 Loo
J - 6 0 0
' - 3 0 0
FAST SLOW
FIGURE 1. Different types of Ca2+ channels in rat muscle cells in culture. Top left: fast (T-type) Ca2+ channels. Top right: slow (L-type) Ca2+ channels. I-V characteristic of the fast (bottom left) and slow (bottom right) Ca2+ channels. The voltage-dependence of these two types of Ca2+ channels is different. Center: I-V characteristic of a skeletal muscle cell having both T- and L-type Ca2+ channels.
136 ANNALS NEW YORK ACADEMY OF SCIENCES
of dihydr~pyridines.'~. T-tubule membranes were used to analyze the molecular pharmacology of the CaZC channel as starting material for the solubilization and purification of this channeI.
DIFFERENT TYPES OF Ca" CHANNELS IN SKELETAL MUSCLE
There are a t least two different types of Ca2+ channels in skeletal muscle (FIGURE l).6v7 One class of these Ca2+ channels activates a t very low membrane potentials and opens and closes rapidly; another type of Ca2+ channels is much slower and only activates near -40 mV. This latter type of CaZ+ channel (L-type) is theonly one that is sensitive to 1,4-dihydropyridine~.~
The L-type Ca2+ channel is blocked by compounds such as (+)PN 200-1 10 and activated by Caz+ channel activators such as Bay K 8644: Blockade by (+)PN 200-1 10 is voltage dependent and is more potent when the channel is in the inactivated state (in a situation where the drug interacts with the depolarized membrane). There is an excellent correlation between binding constants determined with tritiated ligands on isolated membranes and binding constants measured by electrophysiology (voltage clamp) on depolarized cells in which the L-type Caz+ channel is inactivated.' L-type
TABLE 1. Affinities of Different 1,4-Dihydropyridines for Binding to Rabbit Microsome Membranes from Different Tissues
Microsome ( + ) [3H]PN 200-1 10 ['HINitrendipine Membranes KdnMY KdnMY Skeletal muscle 0.2 1 .a Heart muscle 0.06 0.3 Smooth muscle 0.4 0.4 Brain 0.04 0.1
'K , is the equilibrium dissociation constant.
Ca2+ channels in skeletal muscle are slower than in cardiac cells and have a different voltage dependence of inactivation.
PHARMACOLOGICAL PROPERTIES FOR CALCIUM CHANNEL BLOCKER RECEPTORS
TABLE 1 indicates some of the affinities found for the binding of different 1,4-dihydropyridines to the Ca2+ channel protein in skeletal muscle as compared to other tissues. The most active 1,4-dihydropyridine is (+)PN 200-1 10. The binding of 1,4-dihydropyridines to the voltage-dependent Ca2+ channel in cardiac and skeletal muscle cells7** is voltage-dependent, the highest affinity being found when the membrane is depolarized. The affinity of (+)PN 200-1 10 found from binding experiments to purified membranes is the same as that found by electrophysiology on cells having a membrane potential near 0 mV.
Verapamil, bepridil, and diltiazem are known to allosterically regulate 1,4- dihydropyridine binding sites in brain, smooth, skeletal, and cardiac muscles.' T- tubule membranes also have high-affinity binding sites for ( -) ['H]desmethoxyvera- pamil [( -)D888], d-cis-['Idiltiazem, and (*)['H]bepridil. Binding a t these sites is

LAZDUNSKI et al.: MOLECULAR PROPERTIES 137
reversible and saturable with the following equilibrium dissociation constants ( K D ) and maximum binding capacities (Bmax): ( -) [3H]desmethoxyverapamil (KD = 1.5 nM, B,,, = 70 f 10 pmol/mg protein), d-cis-['H]diltiazern ( K D = 50 nM, B,,, = 50 * 8 pmol/mg protein), and (r)['H]bepridil (KD = 20 nM, B,,, = 80 15 pmol/mg protein). Desmethoxyverapamil, diltiazem, and bepridil are competitive for binding to skeletal muscle T-tubule membranes." A high-affinity Ca2+-binding site (KCaz+ = 5 pM) modulates the affinity of binding of desmethoxyverapamil, diltiazem, and bepridil a t their presumably common binding site."
AFFINITY LABELING EXPERIMENTS OF BEPRIDIL, OF DILTIAZEM, AND OF 194-DIHYDROPYRIDINE RECEPTORS IN SKELETAL MUSCLE
Affinity labeling experiments with (k) ['Hlbepridil, d-~is-[~H]diltiazem, and ( +)[3H]PN 200-1 10 have shown that a polypeptide of 170 c 10 kDa is specifically labeled." Affinity labeling results suggest that the Ca2+ channel structure is a large protein of 170 kDa carrying receptors for 1,4-dihydropyridines and for verapamil, D888, diltiazem, and bepridil.
PURIFICATION OF THE 1,4-DIHYDROPYRIDINE RECEPTOR COMPONENT OF THE Ca2+ CHANNEL
Among 14 detergents of varied chemical natures, only one, the 3-[(3-cholamido- propy1)-dimethyl-ammoniol- 1 propane sulfonate (CHAPS), presented the required qualities for solubilization of the dihydropyridine receptor from T-tubule mem- branes.12 Different unlabeled calcium inhibitors have been tested for their ability to interfere with the ['Hlnitrendipine binding on the solubilized receptor.12 These molecules not only include dihydropyridine derivatives [( +)PN 200-1 10, nitrendipine, and nifedipine] but also benzothiazepine (1-cis and d-cis diltiazem), phenylalkylamine [( +) and (-)verapamil], and bepridil compounds. Rank order of potencies to inhibit 50% of binding (Ko.5) are: (+)PN 200-110 (Ko.5 = 0.7 nM) > nifedipine (K,,5 7 nM) > nitrendipine = 9 nM) for 1,4-dihydropyridine and (+)verapamil
= 50 000 nM) >> 1-cis-diltiazem for the other class of drugs. d-cis-Diltiazem has a potentiating effect on (+)['H]PN 200-110 binding to T-tubule membranes with a = 500 nM. Evidence that the solubilization procedure used in this work is useful to purify the voltage-sensitive calcium channel arises from the observations that ligand specificity and stereoselectivity are very close to those known for the membrane-bound receptor.
The CHAPS-solubifized extract from rabbit skeletal muscle microsomes was used for purification studies. The dihydropyridine receptor was substantially purified by a combination of ion exchange chromatography (DEAE), lectin affinity chromatogra- phy on wheat germ agglutinin (WGA), and gel filtration on Sephacryl Sdm. After solubilization, a trace amount of (+)[3H]PN 200-1 10 was added to the solubilized receptor. In this way a small part of the solubilized receptor was specifically labeled (less than 5% of the total receptor complex). This small proportion of the ['Hldihydro- pyridine receptor complex was used to monitor the purification steps by counting aliquots of each fraction, while the total active binding site capacity can still be measured by equilibrium binding experiments.'' Sodium dodecyl sulfate (SDS) gel electrophoresis analysis was carried out after each purification step. The protein
= 500 nM) > (t)bepridil = 4000 nM) > (-)verapamil
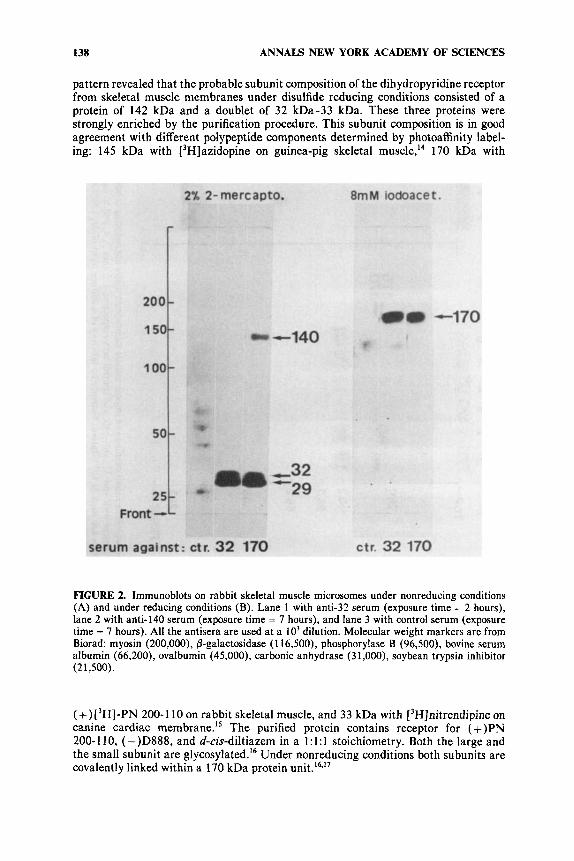
138 ANNALS NEW YORK ACADEMY OF SCIENCES
pattern revealed that the probable subunit composition of the dihydropyridine receptor from skeletal muscle membranes under disulfide reducing conditions consisted of a protein of 142 kDa and a doublet of 32 kDa-33 kDa. These three proteins were strongly enriched by the purification procedure. This subunit composition is in good agreement with different polypeptide components determined by photoaffinity label- ing: 145 kDa with [3H]azidopine on guinea-pig skeletal muscle,I4 170 kDa with
FIGURE 2. Immunoblots on rabbit skeletal muscle microsomes under nonreducing conditions (A) and under reducing conditions (B). Lane 1 with anti-32 serum (exposure time = 2 hours), lane 2 with anti-140 serum (exposure time = 7 hours), and lane 3 with control serum (exposure time = 7 hours). All the antisera are used at a lo3 dilution. Molecular weight markers are from Biorad: myosin (200,000), 0-galactosidase (1 16,500), phosphorylase B (96,500), bovine serum albumin (66,200), ovalbumin (45,000), carbonic anhydrase (3 1 ,OOO), soybean trypsin inhibitor (2 1,500).
(+)[3H]-PN 200-1 10 on rabbit skeletal muscle, and 33 kDa with [3H]nitrendipine on canine cardiac membrane.15 The purified protein contains receptor for (+)PN 200-110, (-)D888, and d-cis-diltiazem in a 1:l:l stoichiometry. Both the large and the small subunit are glycosylated.'6 Under nonreducing conditions both subunits are covalently linked within a 170 kDa protein
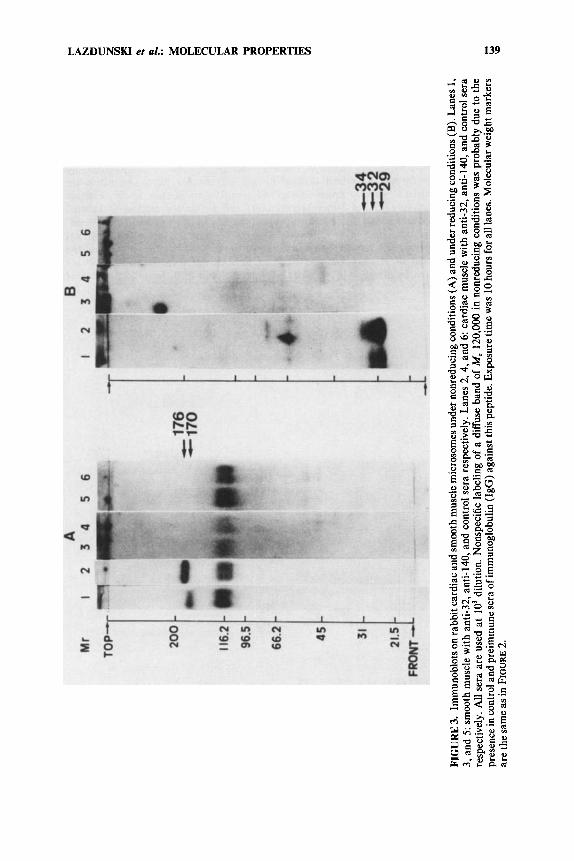
LAZDUNSKI et al.: MOLECULAR PROPERTIES 139
140 ANNALS NEW YORK ACADEMY OF SCIENCES
IMMUNOCHEMICAL ANALYSIS OF SUBUNIT STRUCTURES OF
VOLTAGE-DEPENDENT Ca2’ CHANNELS IN SKELETAL, CARDIAC, AND SMOOTH MUSCLES
1,4-DIHYDROPYRIDINE RECEPTORS ASSOCIATED WITH
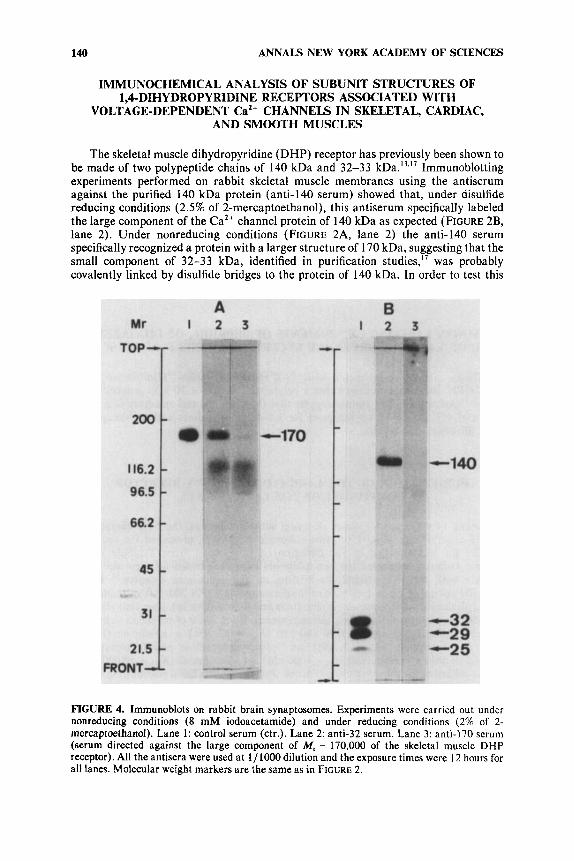
The skeletal muscle dihydropyridine (DHP) receptor has previously been shown to be made of two polypeptide chains of 140 kDa and 32-33 kDa.I3.l7 Immunoblotting experiments performed on rabbit skeletal muscle membranes using the antiserum against the purified 140 kDa protein (anti-140 serum) showed that, under disulfide reducing conditions (2.5% of 2-mercaptoethanol), this antiserum specifically labeled the large component of the CaZ+ channel protein of 140 kDa as expected (FIGURE 2B, lane 2). Under nonreducing conditions (FIGURE 2A, lane 2) the anti-I40 serum specifically recognized a protein with a larger structure of 170 kDa, suggesting that the small component of 32-33 kDa, identified in purification studies,” was probably covalently linked by disulfide bridges to the protein of 140 kDa. In order to test this
FIGURE 4. Immunoblots on rabbit brain synaptosomes. Experiments were carried out under nonreducing conditions (8 mM iodoacetamide) and under reducing conditions (2% of 2- mercaptoethanol). Lane 1: control serum (ctr.). Lane 2: anti-32 serum. Lane 3: anti-I70 serum (serum directed against the large component of M, = 170,000 of the skeletal muscle DHP receptor). All the antisera were used at 1/ 1000 dilution and the exposure times were 12 hours for all lanes. Molecular weight markers are the same as in FIGURE 2.

LAZDUNSKI et al.: MOLECULAR PROPERTIES 141
TABLE 2. Binding of (+)[3H]PN 200-110 and Anti-32 Serum as Respectively Determined by Autoradiographic and Indirect Immunofluorescence Methods"
Specific Binding Anti- of (+)13HlPN 200-110 32 Serum
Hippocampus Ammon's horn Dentate gyrus
Cerebral cortex Striatum Thalamus Hypothalamus Substantia nigra Colliculi Cerebellum cortex
Granular layer Molecular layer
+ +++ ++ + + + + +
+++ ++
+ +++ ++ + + + + +
+++ +
'+ + +, very high label; + +, high label; +, moderate label; +, low label.
hypothesis, an antiserum directed against the purified 32-33 kDa component (anti-32 serum) was prepared and its ability to react with the DHP receptor of skeletal muscle was studied in immunoblots. FIGURE 2B, lane 1, shows that one band revealed by anti-32 serum under disulfide reducing conditions corresponded effectively to a low component of 32 kDa, but it also shows the presence of bands a t 29 kDa and 25 kDa. Under nonreducing conditions the only peptide revealed by anti-32 serum had a structure of 170 kDa (FIGURE 2A, lane 1). Nonreducing conditions caused not only the complete disappearance of the band of 32 kDa but also the disappearance of the bands a t 29 kDa and 25 kDa. This observation suggests that the two smaller polypeptides of 25 kDa and 29 kDa are degradation products of the 32 kDa component. Other structural studies carried out in this laboratory which did not involve the use of antibodies confirmed this view.I6 All these data taken together obtained both with the anti-140 serum and the anti-32 serum indicate that the DHP receptor is indeed a protein of about 170 kDa made of two chains of 140 kDa and 32 kDa covalently linked by disulfide bridge(s).
One important question was to know to what extent the molecular structure of the D H P receptor Ca2+ channel protein in nerve cells and in cardiac and smooth rabbit muscles was similar to that found in skeletal muscle. In order to answer this question, immunoblotting experiments were carried out using microsomal fractions from cardiac and smooth muscles and brain synaptosomes. FIGURE 3A (lanes 1 and 2) shows that proteins of 170 kDa (smooth muscle) and 176 kDa (heart) were specifically labeled by the anti-32 serum under nonreducing conditions. Under disulfide reducing conditions, after 2-mercaptoethanol treatment, the anti-32 serum revealed smaller components of 32 kDa and 29 kDa in the case of smooth muscle (FIGURE 3B, lane 1) and of 34,32, and 29 kDa in the case of cardiac muscle (FIGURE 3B, lane 2). It is probable that the 29 kDa component in smooth muscle membranes is a degradation product of the 32 kDa peptide and that peptides a t 32 kDa and 29 kDa are degradation products of the 34 kDa peptide in cardiac muscle membranes. No specific labeling of peptide bands was observed with the anti-140 serum either in the presence or in the absence of the reducing agent (FIGURE 3A and B, lanes 3 and 4). The presence in cardiac and smooth muscle membranes of the 170-176 kDa protein under nonreducing conditions and of the 32-34 kDa protein under reducing conditions clearly indicates, however, that a component of about 140 kDa exists in these tissues as in skeletal muscle. The lack of
142 ANNALS NEW YORK ACADEMY OF SCIENCES
c
X 15
5 U
cn F z 3 0
W l-
Y
010 n
a t E
a 2 s A
T O *
0 . 5 -
r
0
N z 0.
- A
-
0
1 J m- 0 0 0-0-0
Q-8 1 1 I
1 6 5 1 0-4 1 6 3 1@
--4
A- I 1 F - I 2k4-4 ASCITES 1 o - ~ FLUID 10-4 DILUTION * IN 10-3 THE ASSAY 1 ( r 2
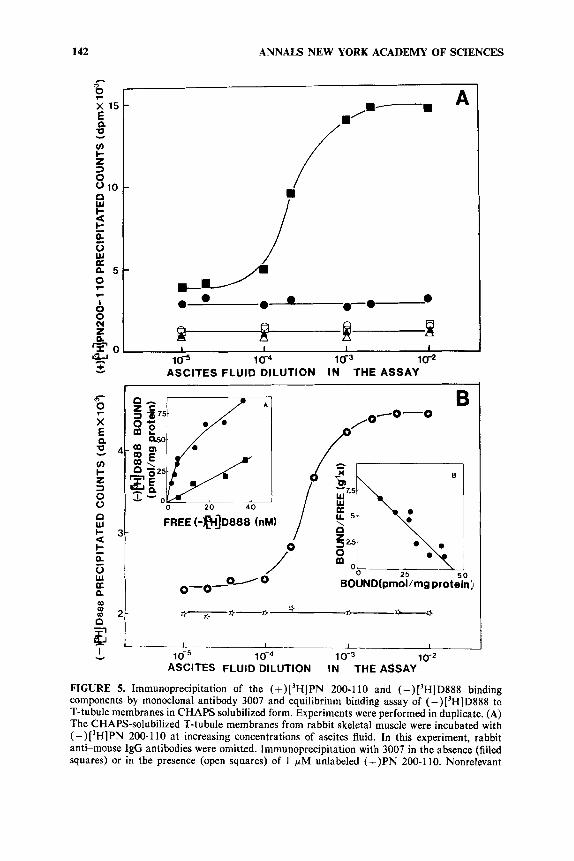
FIGURE 5. Immunoprecipitation of the (+)['H]PN 200-1 10 and (-)['HID888 binding components by monoclonal antibody 3007 and equilibrium binding assay of (-)['HID888 to T-tubule membranes in CHAPS solubilized form. Experiments were performed in duplicate. (A) The CHAPS-solubilized T-tubule membranes from rabbit skeletal muscle were incubated with (+)['H]PN 200-1 10 at increasing concentrations of ascites fluid. In this experiment, rabbit anti-mouse IgG antibodies were omitted. Immunoprecipitation with 3007 in the absence (filled squares) or in the presence (open squares) of 1 pM unlabeled (+)PN 200-1 10. Nonrelevant

LAZDUNSKI et al.: MOLECULAR PROPERTIES 143
labeling of such a polypeptide component by the anti-140 serum could be due to the fact that antigenic determinants are different in skeletal muscle and in cardiac and smooth muscle DHP receptors. It could also be due to an insufficient titer of the anti-140 serum used. This second explanation seems more likely since the titer of the anti-140 serum was lower than the titer of the anti-32 serum and since the amount of DHP receptors measured with (+)[3H]-PN 200-110 in heart and in smooth muscle was considerably lower than in skeletal muscle, making these receptors more difficult to detect in these tissues. Attempts to obtain more potent anti-140 sera were unsuccessful.
FIGURE 4 shows that the same type of subunit structure exists for neuronal Ca2+ channels.'8 In conclusion, this work provides evidence that the DHP receptor, which presumably corresponds to the Ca2+ channel protein of skeletal, cardiac, and smooth muscle as well as of nerve cells, has a very similar subunit composition in all these tissues comprising a large polypeptide of about 140 kDa associated with a smaller polypeptide of 32-34 kDa by disulfide bridge(s). The small subunit could be the same polypeptide that was previously photolabeled with [3H]nitrendipine in canine cardiac membrane^.'^
The series of antibodies described in this report can of course be very useful tools for future purifications of the nerve and smooth muscle Caz+ channel proteins, for the analysis of the distribution of these channels, as previously done for N a + channel^.'^ Immunofluorescence studies of brain sections have already indicated a distribution of the Ca2+ channel protein similar to that of 1,4-dihydropyridine-binding sites with (+)['H]PN 200-1 10 by autoradiographic techniques (TABLE 2)."
MONOCLONAL ANTIBODIES AGAINST THE DIHYDROPYRIDINE RECEPTOR
Monoclonal hybridoma cell lines secreting antibodies against the ( + ) P N 200-1 10 and the (- )desmethoxyverapamil-binding components of the voltage-dependent cal- cium channel from rabbit T-tubule membranes have been isolated.".21 The specificity of these monoclonal antibodies was established by their ability to coimmunoprecipitate (+)['H]PN 200-1 10 and ( -)[3H]desmethoxyverapamil receptors (FIGURE 5). Mono- clonal antibodies described in this work cross-reacted with rat, mouse, chicken, and frog skeletal muscle Ca2+ channels, but not with crayfish muscle CaZ+ channels. Cross-reactivity was also detected with membranes prepared from rabbit heart, brain, and intestinal smooth muscle.
These antibodies were used in immunoprecipitation experiments with i251-labeled detergent (CHAPS and digitonin) solubilized membranes. They revealed a unique
ascites fluid in the absence (filled circles) or in the presence (open circles) of 1 pM (+)PN 200-1 10. Antibody 3007 without CHAPS-soluhilized membranes in the absence (filled triangles) or in the presence (open triangles) of 1 pM (+)PN 200-1 10. (B) Inset A: T-tubule membranes at 20 pg/ml in 20 mM Hepes/NaOH buffer (pH 7.5 and OOC) were incubated for 60 minutes (time sufficient to attain equilibrium) with increasing concentrations of (-)[3H]D888 in the absence (circles) or in the presence (squares) of 1 pM (-)D888. Each sample was then CHAPS solubilized.*' Bound (-)['HID888 was separated from the free ligand by filtration (200 pl aliquots) through 2.8 ml Sephadex G-50 columns. Inset B: Scatchard plot for the specific (-) ['HID888 binding component. Main panel: T-tubule membranes from rabbit skeletal muscle labeled with (-) ['HID888 and CHAPS solubilized were incubated with increasing concentra- tions of ascites fluid." Immunoprecipitation with 3007 in the absence (circled stars) or in the presence (stars) of 1 pM (-)D888.
144 ANNALS NEW YORK ACADEMY OF SCIENCES

LAZDUNSKI ei al.: MOLECULAR PROPERTIES 145
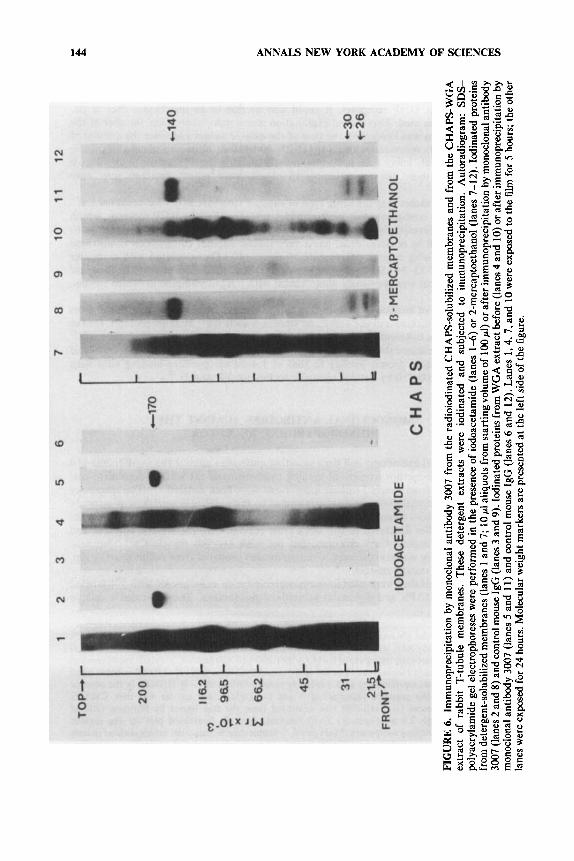
immunoprecipitating component of 170 kDa under nonreducing conditions. After disulfide bridge reduction the CHAPS-solubilized ( + ) P N 200- 1 l o / ( -)desmethoxy- verapamil binding component gave rise to a large peptide of 140 kDa and to smaller polypeptides of 30 kDa and 26 kDa (FIGURE 6) whereas the digitonin-solubilized receptor appeared with subunits of 170 kDa, 140 kDa, 30 kDa, and 26 kDa.
All these results taken together are interpreted as showing that both the 1,4- dihydropyridine and the phenylalkylamine receptors are part o f a single polypeptide chain of 170 kDa.
PURIFICATION OF THE CARDIAC Ca2+ CHANNEL PROTEIN
The dihydropyridine-sensitive voltage-dependent Ca2+ channel from chick heart has now been purified.20 It appears as a major component of 170 kDa linked by disulfide bridges to smaller polypeptides of 32-29 kDa. Both peptide maps of the 140 kDa peptide in cardiac and skeletal muscle preparations and studies with monoclonal antibodies suggest a high degree of homology for CaZ+ channels in the two tissues.
REGULATION OF Ca2+ CHANNEL ACTIVITY AND Ca2+ CHANNEL LEVEL BY ISOPROTERENOL AND cAMP
The effects of short-term stimulation of @-adrenergic receptors and elevations in intracellular cyclic adenosine monophosphate (CAMP) on nitrendipine-sensitive volt- age-dependent Ca2+ channels of skeletal muscle cells in vitro have been studied using both the 4sCa2+ flux technique and [3H] nitrendipine binding experiments.22 Isoprotere- no1 increased the nitrendipine-sensitive 4sCa2+ influx under depolarizing conditions. The effects of isoproterenol were additive to those of depolarization and were antagonized by alprenolol. Half-maximal inhibition of 4sCa2+ influx induced both by depolarization and by isoproterenol occurred a t a nitrendipine concentration of 1 nM. Treatments that resulted in an increased level of intracellular CAMP, such as treatment with 1-methyl 3-isobutylxanthine, theophylline, dibutyryl cAMP or 8- bromo cAMP also resulted in an activation of the nitrendipine-sensitive CaZ+ channel.
As for cardiac cells,23 phosphorylation of the Ca2+ channel in skeletal muscle via a CAMP-dependent kinase increases the probability of finding the Ca2+ channel in the open form.
In contrast, long-term treatment of myotubes in culture with isoproterenol and other compounds that increased intracellular cAMP led to a very large increase in the number of nitrendipine receptors. This increase was accompanied by a 4- to 10-fold decrease in the affinity of the receptors for nitrendipine.22 A similar increase has been observed in neuronal cells.24
Recent work from this laboratory2' has shown that the major subunit of the Ca2+ channel (140-145 kDa) is phosphorylated both by cAMP and Ca2'-calmodulin- dependent processes and that dephosphorylation occurs via calcineurin, a Ca2+- calmodulin-dependent phosphata~e.'~
A NEW EMERGING PHARMACOLOGY FOR Ca2+ CHANNELS
13H] Fluspirilene, a neuroleptic molecule of the diphenylbutylpiperidine series with anxiolytic properties, binds to skeletal muscle transverse tubule membranes with a
146
5 .
ANNALS NEW YORK ACADEMY OF SCIENCES
domperidona
9 .
a .
7 .
6 -
5 -
5 6 7 8 9 10 1 1
-log CKdl (M),C3Hlfluspirilene binding
r
sallopamil
, , , 8 , , ~ - cis-diltiazem
/ / 5 6 7 8 9 1 0
-log [Kd] (MI,(-) C3HlD888 binding
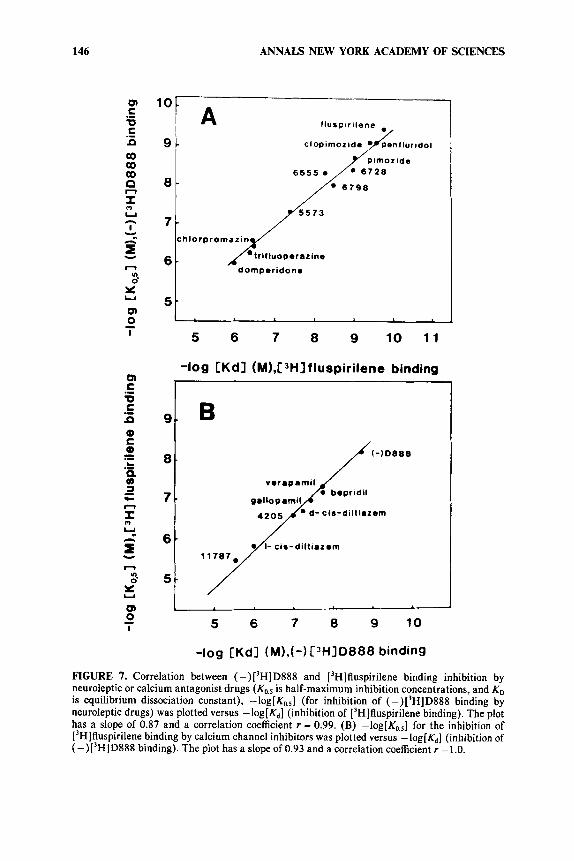
FIGURE 7. Correlation between (-)['HID888 and ['Hlfluspirilene binding inhibition by neuroleptic or calcium antagonist drugs is half-maximum inhibition concentrations, and KD is equilibrium dissociation constant), -log[Ko,,] (for inhibition of (-)['HID888 binding by neuroleptic drugs) was plotted versus -log[&] (inhibition of ['H]fluspirilene binding). The plot has a slope of 0.87 and a correlation coefficient r = 0.99. (B) -log[Ko.,] for the inhibition of ['H]fluspirilene binding by calcium channel inhibitors was plotted versus -log[&] (inhibition of (-)['HID888 binding). The plot has a slope of 0.93 and a correlation coefficient r =1.0.

LAZDUNSKI et al.: MOLECULAR PROPERTIES 147
high affinity corresponding to a dissociation constant Kd = 0.1 1 f 0.04 nM.26 A 1:1 stoichiometry was found between ['HI fluspirilene binding and the binding of ( -) [3H]desmethoxyverapamil, one of the most potent Ca2+ channel inhibitors. Ca2+ channel inhibitors such as desmethoxyverapamil, verapamil, gallopamil, bepridil, or diltiazem antagonize ['HI fluspirilene binding besides antagonizing ['Hldesmethoxy- verapamil binding. Neuroleptics, especially those of the diphenylbutylpiperidine family, antagonize ( -) [3H]desmethoxyverapamil binding besides antagonizing [3H]fluspirilene binding. There is an excellent correlation between affinities found from ['HI fluspirilene and from (-) ['H]desmethoxyverapamil binding experiments (FIGURE 7).
Analysis of the properties of these cross-inhibitions indicates that [3H]fluspirilene binds to a site that is not the same as the site for phenylalkylamine derivatives (gallopamil, verapamil, diltiazem, bepridil).
Voltage-clamp experiments have shown that fluspirilene is a very efficient inhibitor of the voltage-dependent Ca2+ channel with a half-maximum effect near 0.1-0.2 nM and a nearly complete blocking effect a t a concentration of 1 nM. Fluspirilene blockade has very little voltage dependence.26
A PATHOLOGY INVOLVING THE Ca2+ CHANNEL
Binding experiments using tritiated dihydropyridines have shown that receptors for this drug were lacking in a mouse skeletal muscle di~ease.~ ' The mdg/mdg mouse has muscles that are unable to contract. This lack of contraction of course prevents birth. Muscles of mdg/mdg embryos present a lack of functional Ca2+ channels.28v29 This absence of Ca2' channels is only observed in muscle. Ca2+ channels are present on both nerve and cardiac cells. The absence of Ca2+ channels in muscle may be linked to the fact that well-formed triads are not observed in mdg/mdg muscie.
ACKNOWLEDGMENTS
We wish to thank Dr. Busch for the gift of (+)bepridil, Dr. Schramm from Bayer AG (FRG) for C G P 28932, and Dr. Hoffmeister and Dr. Garthoff from Bayer AG (FRG) for Bay K8644 and dihydropyridines. We also thank Bayer Pharma (France) for nitrendipine, Dr. Hollmann and Dr. Traut from Knoll AG (FRG) for (-)['H]des- methoxyverapamil, and CERM (Riom, France) and CEA (France) for [3H]bepridil. Thanks are due to M. T. Ravier, M. Valetti, and C. Roulinat-Bettelheim for expert technical assistance.
REFERENCES
1. JANIS, R. A. & D. J. TRIGGLE. 1984. 1.4-Dihydropyridine Ca2+ channel antagonists and activators: a comparison of binding characteristics with pharmacology. Drug Dev. Res. 4: 251-214.
FLECKENSTEIN, R. 1971. Specific pharmacology of calcium in myocardium, cardiac pace-maker and vascular smooth muscle. Annu. Rev. Pharmacol. Toxicol. 17: 149-166.
FOSSET, M., E. JAIMOVICH, E. DELPONT & M. LAZDUNSKI. 1983. ['HHInitrendipine receptors in skeletal muscles. Properties and preferential localization in transverse tubules. J. Biol. Chem. 258 6086-6092.
POTREAU, D. & G. RAYMOND. 1980. Calcium-dependent electrical activity and contraction of voltage-clamped frog single muscle fibers. J. Physiol. 307: 9-22.
2.
3.
4.

148 ANNALS NEW YORK ACADEMY OF SCIENCES
5.
6.
7.
8.
9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
20.
21.
22.
ALMERS, W., R. FINK & P. T. PALADE. 198 1. Calcium depletion in frog muscle tubules: the decline of calcium current under maintained depolarization. J. Physiol. 3 1 2 177-207.
COGNARD C., M. LAZDUNSKI & G. ROMEY. 1986. Different types of Ca2+ channels in mammalian skeletal muscle cells in culture. Proc. Nat. Acad. Sci. USA 8 3 517-521.
COGNARD C., G. ROMEY, J. P. GALIZZI, M. FOSSET & M. LAZDUNSKI. 1986. Dihydropyri- dine-sensitive Ca’+ channels in mammalian skeletal muscle cells in culture: electrophy- siological properties and interactions with Ca2+-channels activator (Bay K8644) and inhibitor (PN 200-110). Proc. Nat. Acad. Sci. USA 8 3 1518-1522.
BEAN, B. P. 1984. Nitrendipine block of cardiac channels: high affinity binding to the inactivated state. Proc. Nat. Acad. Sci. USA 81: 63884392.
TRIGGLE, D. J. & R. A. JANIS. 1984. Calcium channel antagonists: New perspectives from the radioligand binding assay. I n Modern Methods in Pharmacology 2 1-28. Alan R. Liss, Inc. New York, NY
GALIZZI, J. P., M. BORSOTTO, J. BARHANIN, M. FOSSET & M. LAZDUNSKI. 1986. Characterization and photoaffinity labelling of receptor sites for the Ca2+ channel inhibitors d-cis-diltiazem, (+)bepridil, desmethoxyverapamil and (+)PN 200-1 10 in skeletal muscle transverse tubule membranes. J. Biol. Chem. 261: 1393-1 397.
GALIZZI, J. P., M. FOSSET & M. LAZDUNSKI. 1985. Characterization of the Ca2+ coordination site regulating binding of Ca’+ channel inhibitors d-cis-diltiazem (+)be- pridil and (-)desmethoxyverapamil to their receptor site in skeletal muscle transverse tubule membranes. Biochem. Biophys. Res. Commun. 1 3 2 49-55.
BORSOITO, M., R. I. NORMAN, M. FOSSET & M. LAZDUNSKI. 1984. Solubilization of the nitrendipine receptor from skeletal muscle transverse tubule membranes. Interactions with specific inhibitors of the voltage-dependent Ca” channel. Eur. J. Biochem. 1 4 2 449-455.
BORSOTTO, M., J. BARHANIN, R. I. NORMAN & M. LAZDUNSKI. 1984. Purification of the dihydropyridine receptor of the voltage-dependent Ca’’ channel from skeletal muscle transverse tubules using (+)[3H]PN 200-1 10. Biochem. Biophys. Res. Commun. 1 2 2 1357-1366.
FERRY, D. R., M. ROMBLJSCH, A. GOLL & H. GLOSSMANN. 1984. Photoaffinity labelling of Ca2+ channels with [3H]azidopine. FEBS Lett. 1 6 9 112-1 18.
CAMPBELL, K. P., G. M. LIPSHUTZ & G. M. DENNEY. 1984. Direct photoaffinity labelling of the high affinity nitrendipine-binding site in subcellular membrane fractions isolated from canine myocardium. J. Biol. Chem. 2 5 9 5384-5387.
BARHANIN, J., T. COPPOLA, A. SCHMID, M. BORSOTTO & M. LAZDUNSKI. 1987. The calcium channel antagonists receptor from rabbit skeletal muscle. Reconstitution after purification and subunit characterization. Eur. J. Biochem. 164: 525-53 1.
BORSOITO, M., J. BARHANIN, M. FOSSET & M. LAZDUNSKI. 1985. The 1.4-dihydropyridine receptor associated with the skeletal muscle voltage-dependent Ca’+ channel: purification and subunit composition. J. Biol. Chem. 2 6 0 14255-14263.
SCHMID, A,, J. BARHANIN, C. MOURRE, T. COPPOLA, M. BORSOTTO & M. LAZDUNSKI. 1987. Antibodies reveal the cytolocalization and subunit structure of the 1,4 dihydropy- ridine component of the neuronal Ca’+ channel. Biochem. Biophys. Res. Commun.
ELLISMAN, M. H. & S. R. LEVINSON. 1982. Immunocytochemical localization of sodium channel distributions in the excitable membranes of Electrophorus electricus. Proc. Nat. Acad. Sci. USA 7 9 6707-67 11.
COOPER, C. L., S. VANDAELE, J. BARHANIN, M. FOSSET, M. LAZDUNSKI & M. M. HOSEY. 1987. Purification and characterization of the dihydropyridine-sensitive voltage-depen- dent Ca’+ channel from cardiac tissue. J. Biol. Chem. 2 6 2 509-512.
VANDAELE, S., M. FOSSET, J. P. GALIZZI & M. LAZDUNSKI. 1987. Monoclonal antibodies that co-immunoprecipitate the 1,4-dihydropyridine and phenylalkylamine binding com- ponents associated with the voltage-dependent Ca’+ channel from skeletal muscle. Biochemistry 26: 5-9.
SCHMID, A., J. F. RENAUD & M. LAZDUNSKI. 1985. Short-term and long-term effects of @-adrenergic effectors and cyclic AMP of nitrendipine-sensitive voltage-dependent Ca’+ channels of skeletal muscle. J. Biol. Chem. 260 13041-13046.
1 3 9 996-1002.

LAZDUNSKI et al.: MOLECULAR PROPERTIES 149
23. REUTER, H. 1983. Calcium channel modulation by neurotransmitters, enzymes and drugs. Nature 301: 569-574.
24. NIRENBERG, M., S. WILSON, H. HIGASHIDA, A. ROWER, K. KRUEGER, N. BASIS, R. RAY, S. KENINER & M. ADLER. 1983. Modulation of synapse formation by cyclic adenosine monophosphate. Science 222 794-799.
HOSEY, M., M. BORSOWO & M. LAZDUNSKI. 1986. Phosphorylation and dephosphoryla- tion of the major component of the voltage-dependent Ca2+ channel in skeletal muscle membranes by cyclic AMP and Ca2+-dependent processes. Proc. Nat. Acad. Sci. USA 83: 3733-3737.
GALIZZI, J. P., M. FOSSET, G. ROMEY, P. LADURON & M. LAZDUNSKI. 1986. Neuroleptics of the diphenylbutylpiperidine series are potent calcium channel inhibitors. Proc. Nat. Acad. Sci. USA 8 3 7513-7517.
27. PINCON-RAYMOND, M., F. RIEGER, M. FOSSET & M. LAZDUNSKI. 1985. Abnormal transverse tubule system and abnormal amount of receptors for Ca2+ channel inhibitors of the dihydropyridine family in skeletal muscle from mice with embryonic muscular dysgenesis. Dev. Biol. 112 458466.
ROMEY, G., F. RIEGER, J. F. RENAUD, M. PINCON-RAYMOND & M. LAZDUNSKI. 1986. The electrophysiology expression of Ca2+ channels and of apamin sensitive Ca2+ activated K+ channels is abolished in skeletal muscle cells from mice with muscular dysgenesis. Biochem. Biophys. Res. Commun. 136 935-940.
BEAM, K. G., C. M. KNUDSON & J. A. POWELL. 1986. A lethal mutation in mice eliminates the slow calcium current in skeletal muscle cells. Nature 320 168-170.
25.
26.
28.
29.














![Identification of Large Conductance Calcium Activated Potassium Channel Accessory [beta] 4 Subunit in Rat and Mouse Bladder Smooth Muscle](https://static.fdokumen.com/doc/165x107/633d9c4f199e0a385801d428/identification-of-large-conductance-calcium-activated-potassium-channel-accessory.jpg)





