
Regulation of calcium balance in the sturgeon Acipenser naccarii: a role for PTHrP
11
doi: 10.1152/ajpregu.00203.2007 293:R884-R893, 2007. First published 9 May 2007; Am J Physiol Regul Integr Comp Physiol and Adelino V. M. Canário Juan Fuentes, Christophe Haond, Pedro M. Guerreiro, Nádia Silva, Deborah M. Power : a role for PTHrP naccarii Acipenser Regulation of calcium balance in the sturgeon You might find this additional info useful... 45 articles, 16 of which you can access for free at: This article cites http://ajpregu.physiology.org/content/293/2/R884.full#ref-list-1 2 other HighWire-hosted articles: This article has been cited by http://ajpregu.physiology.org/content/293/2/R884#cited-by including high resolution figures, can be found at: Updated information and services http://ajpregu.physiology.org/content/293/2/R884.full can be found at: and Comparative Physiology American Journal of Physiology - Regulatory, Integrative about Additional material and information http://www.the-aps.org/publications/ajpregu This information is current as of November 7, 2012. http://www.the-aps.org/. Copyright © 2007 the American Physiological Society. ISSN: 0363-6119, ESSN: 1522-1490. Visit our website at 12 times a year (monthly) by the American Physiological Society, 9650 Rockville Pike, Bethesda MD 20814-3991. levels of biological organization, ranging from molecules to humans, including clinical investigations. It is published investigations that illuminate normal or abnormal regulation and integration of physiological mechanisms at all publishes original American Journal of Physiology - Regulatory, Integrative and Comparative Physiology by guest on November 7, 2012 http://ajpregu.physiology.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Regulation of calcium balance in the sturgeon Acipenser naccarii: a role for PTHrP

doi: 10.1152/ajpregu.00203.2007293:R884-R893, 2007. First published 9 May 2007;Am J Physiol Regul Integr Comp Physiol
and Adelino V. M. CanárioJuan Fuentes, Christophe Haond, Pedro M. Guerreiro, Nádia Silva, Deborah M. Power
: a role for PTHrPnaccariiAcipenserRegulation of calcium balance in the sturgeon
You might find this additional info useful...
45 articles, 16 of which you can access for free at: This article citeshttp://ajpregu.physiology.org/content/293/2/R884.full#ref-list-1
2 other HighWire-hosted articles: This article has been cited by http://ajpregu.physiology.org/content/293/2/R884#cited-by
including high resolution figures, can be found at: Updated information and serviceshttp://ajpregu.physiology.org/content/293/2/R884.full
can be found at: and Comparative PhysiologyAmerican Journal of Physiology - Regulatory, Integrative about Additional material and information
http://www.the-aps.org/publications/ajpregu
This information is current as of November 7, 2012.
http://www.the-aps.org/. Copyright © 2007 the American Physiological Society. ISSN: 0363-6119, ESSN: 1522-1490. Visit our website at12 times a year (monthly) by the American Physiological Society, 9650 Rockville Pike, Bethesda MD 20814-3991. levels of biological organization, ranging from molecules to humans, including clinical investigations. It is publishedinvestigations that illuminate normal or abnormal regulation and integration of physiological mechanisms at all
publishes originalAmerican Journal of Physiology - Regulatory, Integrative and Comparative Physiology
by guest on Novem
ber 7, 2012http://ajpregu.physiology.org/
Dow
nloaded from

Regulation of calcium balance in the sturgeon Acipenser naccarii: a rolefor PTHrP
Juan Fuentes, Christophe Haond, Pedro M. Guerreiro, Nadia Silva, Deborah M. Power,and Adelino V. M. CanarioCentro de Ciencias do Mar, CIMAR-Laboratorio Associado, Universidade do Algarve, Campus de Gambelas, Faro, Portugal
Submitted 22 March 2007; accepted in final form 8 May 2007
Fuentes J, Haond C, Guerreiro PM, Silva N, Power DM,Canario AV. Regulation of calcium balance in the sturgeon Aci-penser naccarii: a role for PTHrP. Am J Physiol Regul Integr CompPhysiol 293: R884–R893, 2007. First published May 9, 2007;doi:10.1152/ajpregu.00203.2007.—Calcium regulation in sturgeon isof special interest because they are a representative of the ancientfishes possessing mainly cartilaginous skeletons and a supposedly lowcalcium demand. The present study aimed to characterize the effect ofa chronic absence of dietary calcium and the effect of parathyroidhormone-related protein (PTHrPA) (1-34) (7) on calcium balance injuvenile sturgeon (Acipenser naccarii). At rest, sturgeon juveniles arein net positive calcium balance, since whole body calcium uptake issignificantly higher than efflux and calcium accumulates in the body.To study the importance of dietary calcium, the sturgeon were kept ona calcium-free diet for 8 wk. This manipulation impaired growth asmeasured by failure to gain weight or increase in length and indicatesthat dietary calcium is important for growth in sturgeon. An increasedwhole body calcium uptake partially compensated dietary calciumdeficiency and was associated with increased gill chloride cell numberin lamellae and filaments in parallel with increased gill Na�K�-ATPase activity. In addition, a single injection of piscinePTHrP(1-34) significantly increased whole body calcium uptake anddecreased whole body calcium efflux. Administration of PTHrP sig-nificantly increased circulating plasma calcium 4–24 h postinjection.The increase in net calcium transport and increased plasma levels ofcalcium is consistent with the actions of a hypercalcemic factor. Itwould appear that the sturgeon rely on calcium for growth and tightlyregulate calcium transport. The action in calcium balance is consistentwith PTHrP acting as a hypercalcemic factor in sturgeon.
chloride cells; fish; growth
UNLIKE TERRESTRIAL VERTEBRATES that rely on dietary calcium asa source of calcium, fish have access to a vast calcium pool inwater even in the case of calcium-poor fresh water (15). Theworking model for teleost fish calcium balance entails thecooperation and cross-compensation (16, 19) of the gills andintestine to make enough calcium available for body func-tions. In freshwater teleosts, the gills are assumed to be theprimary entry point for nondietary calcium (12) and in bothfreshwater and seawater species, calcium uptake from thewater by the gill is mediated by the chloride cells. This celltype is a specialized ion-transporting cell characterized bythe presence of numerous mitochondria and a high expres-sion of the Na�-K�-ATPase (12, 35, 37). This cell typecontains all the machinery for transcellular calcium move-ment, such as the Na�/Ca2� exchanger and the Na�-K�-ATPase (16, 19, 29). Extrabranchial chloride cells present in
the fish skin may also play a role in calcium uptake (30, 32).In seawater, fish have high drinking rates, and the intestinebecomes an additional route for calcium uptake from thewater (19, 21).
In recent years, there have been significant advances in theunderstanding of the endocrine control of calcium balance inteleost fish. Although stanniocalcin secreted from the corpus-cles of Stannius is hypocalcemic (or antihypercalcemic factor)(46), there is strong evidence that parathyroid hormone-relatedprotein (PTHrP) has strong short-term hypercalcemic actionson calcium balance (7, 21, 24). A variety of other endocrinefactors described as having long-term calcitropic actions in fishinclude prolactin (18), vitamin D (42), estrogen (22, 34), andcortisol (17).
A recent review (24) provides an up-to-date account of thestructure and physiology of PTHrP in Teleosts. PTHrP, inparticular the PTHrPA(1-34) form has been shown to have arole in calcium regulation in teleost, in particular in sea breameither at whole body level (24) and in in vitro studies in theintestine (21). The mature peptides of the PTHrPA in teleostssuch as the puffer fish, (Takifugu rubripes), sea bream (Sparusauratus), and European flounder (Platichthys flesus) are verysimilar, and identity is over 90% in the first 34 amino acids(24). In contrast, in cartilaginous fish, the PTHrP mRNA hasnot been isolated. However, PTHrP and PTHrP receptor im-munoreactivity have been identified using human antisera inelasmobranchs (3, 26, 43, 44).
The physiological and endocrine model for calcium regula-tion in fish has been developed from studies on teleosts. Yet,considering the evolutionary history of fishes and the manyadaptations to different environments, it is unlikely that anysingle model is representative of the diversity of fish species.The sturgeons (chondrosteans) offer alternative models tostudy the evolution of calcium regulation, as they are repre-sentatives of an ancient branch of the fish phylogenetic tree(10). Calcium regulation in chondrosteans is of particularinterest because they have a cartilaginous skeleton, which mayimply a reduction in calcium needs and thus representing adifferent model of calcium homeostasis.
No account on the calcium balance of sturgeon exists in theliterature, and the potential function of PTHrP has not beenstudied in this group of fish. Thus, the present study aimed tocharacterize the mechanisms by which calcium balance isattained in the sturgeon (Acipenser naccarii) and to examinethe potential action of piscine PTHrPA(1-34) (7) on calciumbalance in this species.
Address for reprint requests and other correspondence: J. Fuentes, Centro deCiencias do Mar, CIMAR-Laboratorio Associado, Universidade do Algarve,Campus de Gambelas, 8005-139 Faro, Portugal (e-mail: [email protected]).
The costs of publication of this article were defrayed in part by the paymentof page charges. The article must therefore be hereby marked “advertisement”in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Am J Physiol Regul Integr Comp Physiol 293: R884–R893, 2007.First published May 9, 2007; doi:10.1152/ajpregu.00203.2007.
0363-6119/07 $8.00 Copyright © 2007 the American Physiological Society http://www.ajpregu.orgR884
by guest on Novem
ber 7, 2012http://ajpregu.physiology.org/
Dow
nloaded from

MATERIALS AND METHODS
Fish
Immature juvenile sturgeon (Acipenser nacarii, Bonaparte 1836)were obtained from commercial dealers and transported to RamalheteMarine Station (University of Algarve, Faro, Portugal). Fish (43 � 2 gbody mass; 25 � 0.45 cm fork length) were maintained in 60-literglass freshwater aquaria (four fish per tank) in a closed circuit with abiological filter at 22°C for 2 wk before experiments. Sea breamjuveniles (Sparus auratus, 26 � 1 g body mass) were obtained froma stock raised at Ramalhete Marine Station and kept in 500-literrunning seawater tanks (36 ppt, 22°C) for at least 2 wk before death.Tilapia (Oreochromis mossambicus, 40 � 3 g body mass) of mixedsexes were obtained from a stock raised at the University of Algarveand maintained in 60-liter glass freshwater aquaria (four fish per tank)in a closed circuit with a biological filter at 27°C for 2 wk beforedeath. All fish were fed daily at a rate of 2% of body weight oncommercial pellets (Durasoja, Provimi, Faro, Portugal). Food waswithheld 24 h prior to, and on the day of experiments and sampling.All experimental work was carried out in compliance with nationalregulations for the use of laboratory animals.
Comparative Analysis of Plasma and Whole Body Calcium
For comparison of whole body calcium and plasma parametersbetween teleosts and chondrosteans, tilapia (n � 14; 7 males and 7females), sea bream juveniles (n � 18; immature), and sturgeon (n �10; immature) were used. Fish were killed with an overdose ofanesthetic with 2-phenoxyethanol (1:1,000 vol/vol, Sigma-Aldrich,Madrid, Spain), weighed (to the nearest gram) and measured (to thenearest millimeter). Before blood collection, the puncture area wasdried with tissue paper to avoid contamination of the samples withwater. A 0.3-ml blood sample was collected by caudal puncture into1-ml heparinized syringes (ammonium heparin, 30 units/ml; Sigma-Aldrich), and plasma was separated by centrifugation of whole blood(10,000 rpm for 3 min.), aliquoted into 1.5-ml vials, snap-frozen, andstored in liquid nitrogen until analysis. The plasma parameters mea-sured were PTHrP, total calcium, and the filtered plasma calciumfraction.
For measurement of whole body calcium content in sturgeon andsea bream, carcasses were frozen, homogenized for 10 min untilreduced to a paste. Subsequently, a subsample was transferred topreweighed glass vials and digested with nitric acid (1:1 weight:volume) at 30°C for 1 wk. After digestion, samples were neutralizedwith equal volumes of 2 M NaOH, and calcium content was deter-mined in triplicate using a colorimetric assay (Sigma-Aldrich proce-dure no. 587).
Determination of Whole Body 45Ca Fluxes in Sturgeon
Whole body calcium uptake. For determinations of whole bodycalcium uptake, three fish per tank in duplicate (six fish per treatment)were transferred to freshwater flux tanks (1.5 l/fish) set up in closedcircuit with aeration. After a 30-min recovery period, the tank waterwas spiked with 37 kBq/l 45CaCl2 (specific activity 14.8 MBq/�mol;NEN, Life Sciences Products, Boston, MA) and allowed to mix. Thetanks were partially darkened with a black lid, and fish were left toswim undisturbed. Exactly 4 h after the addition of the tracers,triplicate 2-ml water samples were collected from each tank forcounting, and the tank water was replaced with tracer-free water. Thefish were left to swim for 15 min to remove any radioactivity,adhering to the body surfaces and then killed with an overdose of2-phenoxyethanol (1:250, vol/vol), blotted dry, quickly weighed to thenearest gram, measured to the nearest millimeter, and frozen at�20°C. The whole frozen intestine, from esophagus to rectum, fromindividual fish was dissected out, and the carcass was transferredseparately to 2 volumes of double-distilled water and homogenized
for 10 min until reduced to a paste. A subsample was transferred topreweighed glass vials and digested with nitric acid (1:1 weight:volume) at 30°C for 1 wk. After digestion, samples were neutralizedwith equal volumes of 2 M NaOH and transferred to scintillation vials(2 ml per vial to count the whole subsample). All samples, includingtank water and digested carcasses, were bleached with 2 ml 35%hydrogen peroxide (Fluka; Sigma-Aldrich) to prevent color quench-ing, incubated overnight at room temperature, and then counted for 15min in a Beckman LS6000IC scintillation counter after the addition of20 ml scintillation cocktail (OptiPhase HiSafe II, Wallac; AmershamPharmacia Biotech, Piscataway, NJ).
Whole body calcium uptake (CU) was calculated according to thefollowing equation: CU � (FC/WCa)/(t �w), where FC is the total45Ca activity in the fish carcass (i.e., the whole intestine excluded),WCa is the 45Ca specific activity in the water (cpm/nmol), t is theduration of exposure (h), and w is fish wet weight (g). Whole bodycalcium influx rate is expressed as nanomoles per hour per gram.
Whole body calcium efflux. Fish were loaded with 45Ca by intra-peritoneal injection of 37 kBq 45CaCl2 dissolved in 300 �l 0.9% NaCland returned to holding tanks for 36 h for recovery and to achieve aconstant specific activity of the readily available calcium pool in thewhole fish. For determinations of whole body calcium efflux, fishfrom experiment 1, or control and PTHrP injected from experiment 2(see below) were lightly anesthetized (2-phenoxyethanol, 1:20,000vol/vol) and transferred to 0.5 liter freshwater flux tanks (fish to tankratio 1:10) with aeration. Five-milliliter water samples were takenafter 30 min and every 20 min thereafter for 2 h. Fish were then killedwith an overdose of anesthetic (1:250); fish were then weighed, and ablood sample was collected and plasma was separated. Water andplasma samples (200 �l) were measured for 45Ca radioactive decay aspreviously described. Calcium efflux rate (ER) was calculated asfollows: ER � (Aw �Ct)/(At � t �W), where Aw is the final specificactivity in water, Ct is the total plasma calcium (in millimoles perliter), At is the specific activity of 45Ca in the plasma, t is time (h), andW is fish wet weight (g). Results are expressed as nanomoles per hourper gram. Calculations of calcium efflux from individual fish weretaken to show initial velocity if individual timed collections compliedwith linearity (R2 � 0.98), to rule out back flux.
Experiment 1: Effect of Dietary Calcium on Growthand Calcium Balance
After the acclimation period, 20 juvenile sturgeons were randomlyallocated in four 65-liter tanks (5 fish/tank; 2 tanks per treatment), andthe diet was gradually (over 4 days) changed from the control pellets(calcium-sufficient) to the test pellets [calcium-deficient; provided byProf. G. Flik, Radboud University Nijmegen and previously tested inthe sea bream (1)]. Fish were fed twice daily to a final ration of exactly1.5% of the total body mass per day. The calcium-deficient and-sufficient diets were identical in appearance (shape and color), and noreduction in feeding was observed after switching to test diet pellets,and all food offered was consumed by the animals. The fish were kepton the experimental regime for 8 wk and were fasted for 24 h beforeeach sampling. The sturgeon were sampled immediately before(time 0) and 2, 4, 6, and 8 wk after the start of the experiments.
At each sampling, the sturgeon were weighed (to the nearest gram)and measured (to the nearest millimeter) under anesthesia. A bloodsample (0.3 ml) was collected and processed as described above. Fishwere returned to their holding tanks and resumed normal feeding theday after sampling. Food rations were adjusted after each samplingusing the recorded total wet weight of fish in the tanks and maintainedconstant until the next sampling. No mortalities occurred during theexperiment. The condition factor (K) was calculated using the equa-tion: K � (W/L3) �1,000, where W is body weight (g) and L isstandard length (cm). At the end of experiments (week 8), both controlfish and those maintained on a calcium-deficient diet were subject tothe same experimental procedure. All fish in one tank (n � 5) were
R885CALCIUM REGULATION IN STURGEON
AJP-Regul Integr Comp Physiol • VOL 293 • AUGUST 2007 • www.ajpregu.org
by guest on Novem
ber 7, 2012http://ajpregu.physiology.org/
Dow
nloaded from

netted and three (from each duplicate tank) were transferred to fluxboxes (see above). The remaining fish were anesthetized to recordweight and length and for a final collection of plasma as previouslydescribed. A biopsy of the first left gill filaments was collected fromindividual fish in 100 �l of ice-cold sucrose-EDTA-imidazole buffer(SEI; or 150 mM sucrose, 10 mM EDTA, 50 mM imidazole, pH 7.3)in 0.5-ml vials, snap-frozen and stored in liquid nitrogen or fixed inBouin’s fixative and used for quantification of branchial chloride cellnumbers (see Chloride Cell Detection and Quantification).
Experiment 2: Effect of Piscine PTHrP(1-34) on Calcium Balance
After the acclimation period of 2 wk, groups of four sturgeonjuveniles per tank (two tanks per treatment) were anesthetized,weighed to the nearest gram, and injected intraperitoneally with saline(0.9% NaCl, control fish) or freshly prepared piscine (1-34) Takifugurubripes PTHrPA (7, 23, 24) to give final doses of 10 ng/g or 50 ng/gwet weight. This process was achieved in less than 3 min with twooperators. At 4 and 24 h after hormone treatments, all fish in one tank(n � 4) were anesthetized and weighed, and the blood was collectedand processed as described above. Experiments were performed twiceas to give a final n � 8 per treatment.
For determinations of whole body calcium uptake, control andPTHrP-injected fish (n � 6 per treatment) were slightly anesthetized(2-phenoxyethanol, 1:20,000 vol/vol), injected to give final doses of 0,10, or 50 ng PTHrP(1-34)/g wet weight, transferred to freshwater fluxtanks, and processed as described above.
Blood Plasma Calcium and PTHrP
Calcium was measured in triplicate by colorimetric assay (Sigma-Aldrich procedure no. 587). The plasma total calcium fraction wasmeasured by ultrafiltration of plasma by centrifugation (12,000 g, 10min at 4°C) through a 10-kDa cutoff filter (Millipore) followed bycolorimetric assay (Sigma-Aldrich procedure no. 587). Values forcalcium are shown as millimoles per liter for plasma or milligram pergram for whole body measurements. Plasma levels of PTHrP weremeasured by radioimmunoassay (RIA) after assessing parallelism ofdiluted sturgeon plasma to the assay standard curve (39). Plasmavalues of PTHrP are shown as nanograms per milliliter.
Gill ATPase Activity
The activity of branchial Na�-K� ATPase is often used as a markerof chloride cells in the gill epithelium (12, 35, 37) Gill Na�-K�-ATPase activity was determined using a microassay (31). In brief, gillbiopsies were homogenized in 125 ml of SEI buffer with 0.1%deoxycholic acid and spun at 2,000 g for 30 s. Ten microliters ofhomogenate were added to a 200-�l assay mixture in duplicate in thepresence or absence of 0.5 mM ouabain in 96-well plates at 25°C andread at 340 nm for 15 min with intermittent mixing. Ouabain-sensitiveATPase activity was detected by enzymatic coupling of ATP dephos-phorylation to NADH oxidation. ATPase activity was expressed asmicromoles ADP per milligram protein per hour. Protein was assayedin triplicate in homogenates using the Bradford method with BSA(Sigma-Aldrich) as a standard (6). Both assays were run in microtiterplates, and absorption readings were carried out using a Benchmarkmicroplate reader (Bio-Rad, Hercules, CA).
Chloride Cell Detection and Quantification
For chloride cell detection by immunohistochemistry (IHC) for theNa�-K�-ATPase, gill samples were collected and fixed in Bouin’sfluid for 48 h and washed and stored in 70% ethanol. Whole mountIHC was carried out on individual gill filaments dissected from thefixed gill samples. Before IHC gill filaments were washed 24 h in 70%ethanol containing 1% H2O2 followed by 24 h in renewed baths ofTris-buffered saline/Triton X (TBS/TX) (50 mM TBS, 150 mM NaCl,with 0.1% TX-100, pH 7.6). Incubation with the primary antibody
(anti Na�-K�-ATPase, �5 mouse monoclonal from DevelopmentalStudies Hybridoma Bank, University of Iowa, Iowa City, IA) wasperformed over 48 h at a dilution of 1/400 dilution in TBS/TXcontaining 1% BSA, 5% normal goat serum, and 0.05% sodium azide.Samples were washed over 4 h with several changes of TBS/TX andincubated for 24 h with a goat-anti-mouse antibody (GAM IgG H�L,Nordic Immunology, Tilburg, The Netherlands) at a dilution of 1/400in TBS/TX. After washing in TBS/TX for 4 h, mounts were incubated24 h with a mouse peroxidase-antiperoxidase complex (M/PAP;Nordic Immunology, Tilburg, The Netherlands) at a dilution of 1/600in TBS/TX and finally washed several hours in TB/TX (50 mM TBSwith 0.01% TX-100, pH 7.6). All incubations were performed at roomtemperature in 2-ml tubes. Detection was carried out by incubatingwith the chromagen, DAB (0.005% wt/vol; Sigma-Aldrich) in TB/TXcontaining 0.002% H2O2 and 0.1% (wt/vol) NiNH3 to enhance thesignal. Stained filaments were washed and kept in 70% ethanol andobserved with a stereomicroscope (Olympus SZ60) equipped with adigital camera (Olympus DP11).
Samples from fish fed calcium-sufficient or calcium-deficient dietwere collected for fixation as described above, processed for standardparaffin histology, and cut at 7 �m to obtain longitudinal sections ofthe gill filaments. Sections were mounted on glass slides coated inpoly-L-lysine and dried. IHC was carried out on dewaxed sectionsfollowing the general methodology indicated above with the follow-ing differences: endogenous peroxidase inactivation with 0.35% H2O2
for 20 min in 100% methanol; incubation for 1 h with goat-anti-mouseIgG H�L, 1 h with the M/PAP. Sections were mounted in DePex(Merck, Boston, MA) and analyzed with a microscope (Leica DM2000) equipped with a digital camera (Leica DFX 400).
In the sectioned tissue, two different regions of the gill filamentwere considered for chloride cell distribution/density analysis accord-ing to the results of the whole mount IHC: region 1, the afferent sideof the filament devoid of lamellae; and region 2, the median part of thefilament-bearing lamellae. For each fish, five images were recordedfrom each of the defined regions of the filament. In region 1, thelength of the epithelium lining the filament was measured and immu-nopositive cells counted. In region 2, the length of the epitheliumlining the lamellae (4 to 6 lamellae per fish) was measured, and theimmunopositive cells in this epithelium were counted. In addition,the length of the interlamellar filament epithelium was measuredand the immunopositive cells in this epithelium were counted. Thedensity of immunopositive cells was expressed as the number ofchloride cells per linear millimeter of lining epithelium. The dataobtained from the different images were averaged for each fish, andeight fish per group (one control group and one calcium-deficient dietgroup) were used. Three sets of data were obtained for each sample,the density of chloride cells in the afferent filament epithelium (AFE),the interlamellar filament epithelium (IFE), and the lamellar epithe-lium (LE). All measurements were performed using the open sourcesoftware Image J (2).
Detection and Identification of Calcified Tissue
A preliminary analysis to assess the presence of calcified tissue wasconducted. Sturgeon snout, vertebrae, and ganoid scales were fixed forhistology in sublimated Bouin-Hollande and embedded in paraffin(Histosec, Merck). Serial sections (5 �m) were mounted on poly-L-lysine-coated glass slides. Sections were stained with Masson’sTrichrome to distinguish mineralized tissue and nonmineralized con-nective tissue (11). Briefly, to stain nuclei, sections were immersed inMayer’s hematoxylin, followed by staining of mineralized tissue byan acid fuchsin/xylidine Ponceau solution. Differentiation was per-formed with phosphomolybdic acid followed by staining of thenon-mineralized tissue with fast green. Alcian blue staining of muco-polysaccharides in connective tissue allowed the identification ofcartilage and was carried out with dewaxed and rehydrated sections byimmersion for 30 min in Alcian blue (1% Alcian blue in 3% acetic
R886 CALCIUM REGULATION IN STURGEON
AJP-Regul Integr Comp Physiol • VOL 293 • AUGUST 2007 • www.ajpregu.org
by guest on Novem
ber 7, 2012http://ajpregu.physiology.org/
Dow
nloaded from

acid); sections were then counterstained with Harris hematoxylin.Vertebrae and ganoid scales were fixed in ethanol and the presence ofcalcium detected with alizarin red S whole-mount staining, using anadaptation of a previously described method (13). Stained sectionsand whole mount preparations were analyzed using a microscope(Olympus BH2) and a stereomicroscope (Olympus SZ-60) coupled toa digital camera (Olympus DP11) linked to a computer for digitalimage analysis.
Statistics
Values are shown as means � SE, unless otherwise stated. Afterassessing normality and homogeneity of variances, differences be-tween groups were established as appropriate by Student’s t-test, orone-way ANOVA followed when necessary by the a posteriori Bon-ferroni multicomparison test to identify groups different from con-trols. For chloride cell distribution/proliferation data were analyzedusing a two-way ANOVA considering treatment (food calcium level)and histological location (lamellae, interlamellae, and filament epithe-lia) as main factors, followed by the Bonferroni multicomparison testto identify significantly different groups. Differences between groupswere considered significant at P � 0.05.
RESULTS
Basal Plasma and Body Calcium
Table 1 shows the summary of comparative values of wholebody calcium and plasma calcium parameters in juvenile seabream, tilapia, and sturgeon. Plasma calcium in sturgeon juve-niles ranged between 1.2 and 1.3 mM, roughly half that ofteleost fish. While the ratio of filtered (i.e., ionic) to totalcalcium in the sea bream and tilapia ranged between 0.4 and0.8, in sturgeon it was 0.99, which means that virtually all ofthe calcium circulating in sturgeon plasma is ionic.
Whole body 45Ca fluxes in juvenile sturgeon indicate thatthere is a net calcium uptake of 12.6 nmol �h�1 �g�1 (Fig. 1).The overall positive net influx is a consequence of the highwhole body calcium uptake and the significantly lower calciumefflux (P � 0.05, Student’s t-test).
Whole body calcium in sturgeon juveniles (5.67 � 0.19mg/g) was slightly lower than that of the sea bream (6.36 �0.14 mg/g) and tilapia (14) (12 mg/g). The high bodycalcium content and the significant net calcium uptakeobserved in sturgeon juveniles raise questions about thepotential localization of this calcium. The distribution ofcalcium in the endoskeleton compared with the exoskeleton
of sturgeon is in agreement with the generally acceptedabsence of calcification of the endoskeleton (Fig. 2), asexemplified by the vertebrae (Fig. 2E). In contrast, thesturgeon ganoid scales contain abundant calcium (Fig. 2, Cand D) as does the snout area (Figs. 2, A and B), indicatingthat the exoskeleton most likely accounts for the high valuesof whole body calcium detected.
Dilutions of sturgeon blood plasma show good cross-reac-tion and parallelism to the standard curve in the PTHrP RIAand measurements of blood plasma in normal untreated juve-nile sturgeon were 0.7 � 0.07 ng/ml (n � 10), roughlyone-third of the plasma levels measured in the normal un-treated juvenile sea bream and tilapia (Table 1).
Effect of Calcium-Deficient Diet on Calcium Balance
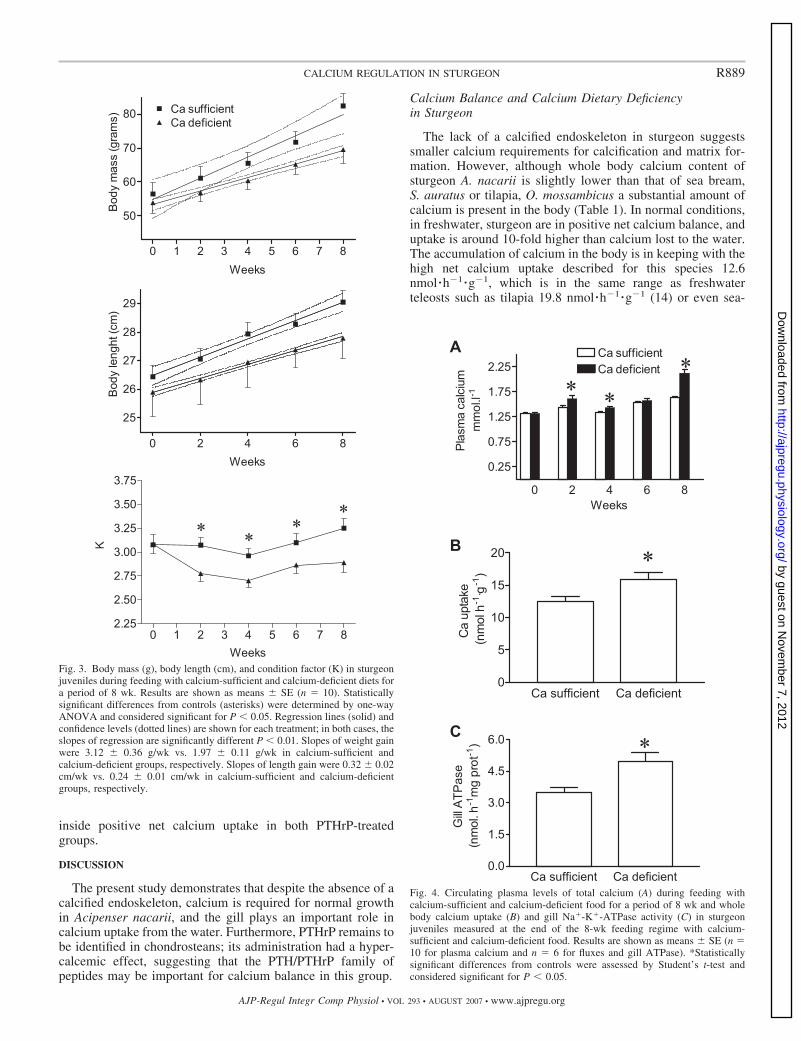
The effect of feeding calcium-deficient and calcium-suffi-cient diets on body mass, length, and condition factor (K) insturgeon juveniles is shown in Fig. 3, A–C. In both treatments,there was a positive mass gain during the 8 wk, although lackof dietary calcium significantly impairs growth in weight (Fig.3A, F � 8.92, P � 0.024) and length (Fig. 3B, F � 27.2, P �0.001). The value of K in fish maintained on a calcium-deficient diet was consistently and significantly lower than inthe calcium-sufficient control group throughout the experimen-tal period (Fig. 3C, P � 0.05, one-way ANOVA).
Plasma levels of calcium in response to feeding with calcium-deficient and calcium-sufficient diets in sturgeon juveniles areshown in Fig. 4A. Fish fed the calcium-deficient diet had higherplasma levels of calcium at all sampling points, with theexception of the data point on week 6. After 8 wk of treatment,plasma calcium in fish fed with the calcium-sufficient dietranged from 1.3 to 1.45 mM, while fish fed the on calcium-deficient diet ranged from 2.1 to 2.2 mM.
The effect of dietary calcium on whole body calciumuptake is shown in Fig. 4B. In fish fed with calcium-sufficient diets, calcium uptake averaged 12.4 � 0.8nmol �h�1 �g�1, and an increase of whole body calciumuptake of 28% was observed in the calcium-deficient dietgroup (P � 0.05, Student’s t-test).
Gill Na�-K�-ATPase activity
The effect of dietary calcium on gill Na�-K�-ATPase ac-tivity is shown in Fig. 4C. In the calcium-sufficient diet group,the activity was 3.4 � 0.3 nmol �h�1 �mg protein�1 and in-
Table 1. Whole body and plasma parameters in sturgeon,tilapia, and sea bream
Sturgeon Tilapia Sea Bream
Body calcium, mg/g 5.67�0.19 12* 6.36�0.14Plasma total calcium,
mmol/l1.32�0.02 5.68�1.21 (female) 2.56�0.08
2.71�0.07 (male)Plasma free calcium,
mmol/l1.30�0.03 1.96�0.05 1.65�0.04
Ratio free/total calcium 0.99�0.02 0.40�0.08 (female) 0.64�0.010.74�0.02 (male)
Plasma PTHrP, ng/ml 0.71�0.07 2.11�0.11 2.43�0.08
Results are shown as means � SE; n � 10 for sturgeon, n � 18 for seabream, n � 7 for tilapia female, and n � 7 for tilapia male. Tilapia results notseparated by sex were not significantly different (Student’s t-test) and werepooled. PTHrP, parathyroid hormone-related protein. *Data for male tilapiafrom Ref. 14.
Fig. 1. Whole body calcium uptake, efflux, and resulting net flux in juvenilesturgeon. Results are shown as average � SE (n � 6). Differences betweenuptake and efflux were assessed by Student’s t-test and considered significantfor P � 0.05. Sturgeon juveniles are in positive whole body calcium uptake.
R887CALCIUM REGULATION IN STURGEON
AJP-Regul Integr Comp Physiol • VOL 293 • AUGUST 2007 • www.ajpregu.org
by guest on Novem
ber 7, 2012http://ajpregu.physiology.org/
Dow
nloaded from

creased 40% in the calcium-deficient diet group (P � 0.05,Student’s t-test).
Numerous putative chloride cells, immunopositive for Na�-K�-ATPase, were observed in the sturgeon gill filaments (Fig.6A). Chloride cells were mainly distributed in the epitheliumlining the blood afferent (afferent or efferent refer to bloodflow hereafter) side of the filament (AFE), the epitheliumlining the lamellae (LE), and the epithelium lining the interla-mellar spaces (IFE, Fig. 5D). Few or no chloride cells wereobserved on the efferent side of the filament, either on thefilament epithelium or the lamellar epithelium close to thisedge of the filament (Fig. 5, A–D). In the calcium-sufficientdiet group, the density of chloride cells was significantlydifferent in the three distinct areas (AFE, IFE and LE). Thehighest density of chloride cells was observed on the IFE andwas approximately one-half in the AFE, whereas the LE hadthe lowest chloride cell density. In the calcium-deficient dietgroup, the distribution of chloride cells was similar to that ofthe control group, but the density of chloride cells increasedsignificantly (Fig. 5E). There was no significant interactionbetween the main factors (location in epithelia food calciumlevel; two-way ANOVA F � 2.05, P � 0.14), indicating thatthe effect of dietary calcium is homogeneous in the epithelialareas under study. Overall, there was a 20% increase of
chloride cell density in the three distinct epithelia of the gillfilament (Fig. 5E).
Effect of PTHrP on Calcium Balance
Alterations of circulating levels of PTHrP in response to theinjection of piscine PTHrP(1-34) are shown in Fig. 6A. Therewere no differences between control and PTHrP-treated groupsat 4 h postinjection. However, by 24 h, circulating PTHrPlevels were significantly higher in fish injected with 10 and 50ng/g PTHrP(1-34) (P � 0.05, one-way ANOVA).
PTHrP(1-34) induced a significant increase in circulatingplasma calcium levels (Fig. 6A). This increase was effective asearly as 4 h after injection with 50 ng/g and becomes signifi-cantly higher than controls at 24 h at doses of 10 and 50 ng/g(Fig. 6B). In addition, the ratio of filtered to total plasmacalcium shows a significant decrease of 10% at doses of 10 and50 ng/g 24 h after the injection of PTHrP (Fig. 6C).
PTHrP(1-34) had two main actions related to whole bodycalcium balance. It evoked a significant reduction of calciumefflux at 50 ng/g and not at a dose of 10 ng/g (Fig. 7A) andresulted in an increase in calcium uptake at both doses testedwith no evident dose-response (Fig. 7B). The combination ofboth effects resulted in a substantial increase in the already
Fig. 2. Masson’s trichrome (A) and Alcian blue (B)staining of a section through the snout. Alizarin redwhole mount staining of skin with ganoid scales (C andD) and vertebra (E). A: mineralized structures are iden-tified by the pink stain and correspond to the rostralbones of the snout; these bones are located beneath theskin (s) and are supported by nonmineralized connec-tive tissue (ct) and cartilage (c), which is stained lightgreen. B: Alcian blue staining clearly identifies thecartilaginous (c) structure in the snout characterized bytheir intense blue staining; the rostral bones (rb) do notstain using this method. C: lateral view of a ganoidscale, which as indicated by its affinity for Alizarinred is calcified. D: top view of an alizarin red-stainedganoid scale; calcified thorn-shaped spines are ob-served scattered in the skin (arrowheads) around thescale. E: vertebra subject to alizarin red staining has noaffinity for the stain, suggesting it is not calcified. ct,Connective tissue; c, cartilage; rb, rostral bone; s, skin;v, vertebra; n, notochord.
R888 CALCIUM REGULATION IN STURGEON
AJP-Regul Integr Comp Physiol • VOL 293 • AUGUST 2007 • www.ajpregu.org
by guest on Novem
ber 7, 2012http://ajpregu.physiology.org/
Dow
nloaded from
inside positive net calcium uptake in both PTHrP-treatedgroups.
DISCUSSION
The present study demonstrates that despite the absence of acalcified endoskeleton, calcium is required for normal growthin Acipenser nacarii, and the gill plays an important role incalcium uptake from the water. Furthermore, PTHrP remains tobe identified in chondrosteans; its administration had a hyper-calcemic effect, suggesting that the PTH/PTHrP family ofpeptides may be important for calcium balance in this group.
Calcium Balance and Calcium Dietary Deficiencyin Sturgeon
The lack of a calcified endoskeleton in sturgeon suggestssmaller calcium requirements for calcification and matrix for-mation. However, although whole body calcium content ofsturgeon A. nacarii is slightly lower than that of sea bream,S. auratus or tilapia, O. mossambicus a substantial amount ofcalcium is present in the body (Table 1). In normal conditions,in freshwater, sturgeon are in positive net calcium balance, anduptake is around 10-fold higher than calcium lost to the water.The accumulation of calcium in the body is in keeping with thehigh net calcium uptake described for this species 12.6nmol �h�1 �g�1, which is in the same range as freshwaterteleosts such as tilapia 19.8 nmol �h�1 �g�1 (14) or even sea-
Fig. 4. Circulating plasma levels of total calcium (A) during feeding withcalcium-sufficient and calcium-deficient food for a period of 8 wk and wholebody calcium uptake (B) and gill Na�-K�-ATPase activity (C) in sturgeonjuveniles measured at the end of the 8-wk feeding regime with calcium-sufficient and calcium-deficient food. Results are shown as means � SE (n �10 for plasma calcium and n � 6 for fluxes and gill ATPase). *Statisticallysignificant differences from controls were assessed by Student’s t-test andconsidered significant for P � 0.05.
Fig. 3. Body mass (g), body length (cm), and condition factor (K) in sturgeonjuveniles during feeding with calcium-sufficient and calcium-deficient diets fora period of 8 wk. Results are shown as means � SE (n � 10). Statisticallysignificant differences from controls (asterisks) were determined by one-wayANOVA and considered significant for P � 0.05. Regression lines (solid) andconfidence levels (dotted lines) are shown for each treatment; in both cases, theslopes of regression are significantly different P � 0.01. Slopes of weight gainwere 3.12 � 0.36 g/wk vs. 1.97 � 0.11 g/wk in calcium-sufficient andcalcium-deficient groups, respectively. Slopes of length gain were 0.32 � 0.02cm/wk vs. 0.24 � 0.01 cm/wk in calcium-sufficient and calcium-deficientgroups, respectively.
R889CALCIUM REGULATION IN STURGEON
AJP-Regul Integr Comp Physiol • VOL 293 • AUGUST 2007 • www.ajpregu.org
by guest on Novem
ber 7, 2012http://ajpregu.physiology.org/
Dow
nloaded from

water fish such as the sea bream 18 nmol �h�1 �g�1 (22). Theidentification of all areas in the body in which calcium accu-mulates still requires a detailed study, but the histology resultssuggest significant amounts of calcium are deposited in theexoskeleton, that is, the ganoid scale system and cranium.
Total calcium circulates at much lower levels in A. nacariiand Acipenseridae (38) compared with sea bream or tilapia(see Table 1 for comparison). We show also that, strikingly, inthe sturgeon A. nacarii nearly all (99%) of the calcium circu-lates in the plasma unbound to protein (i.e., in ionic form),which is significantly higher than in sea bream (around 65%) ortilapia (between 45 and 75% depending on sex, Table 1). Thisfeature is, as far as we know, unique to this group of verte-brates. Thus, the high calcium uptake observed in our study insturgeon, together with a low efflux and the limited amount ofcalcium circulating in the plasma reinforces the idea of calciumaccumulation in the exoskeleton.
In teleosts, the lack of dietary calcium combined withreduced water calcium availability results in reduced or evenarrested (with limiting water calcium) growth, as recentlyshown for the sea bream (1). Although lack of dietary calciumimpaired growth in A. nacarii, the fish were still able tomaintain positive growth. Thus, if calcium is important tomaintain positive growth, compensatory mechanisms to in-crease calcium uptake may have been activated. The cross-regulation of intestinal (dietary) and branchial calcium uptake
was first demonstrated in goldfish (25), where increasedbranchial calcium uptake was confirmed in the absence ofdietary calcium. Furthermore, it has been recently shown thatincreased dietary calcium results in lower gill uptake in rain-bow trout (5). Indeed, the measurements of whole body cal-cium uptake in juveniles of A. nacarii show that lack of dietarycalcium results in increased epithelial calcium uptake (Fig.4B). Thus sturgeon is regulating calcium, and because thepotential sources of calcium can be the surrounding water andthe diet, the available routes of calcium entry into the body arethe gill and the intestine. The low freshwater calcium content(0.7 mM) and the expected low drinking rate for all juvenilesof freshwater fish (20), such as the sturgeon used in this study,suggest the intestine may have only a minor role in thecompensatory response to lack of dietary calcium, and the gillis the likely target of increased epithelial calcium uptake.
In fish, the branchial epithelium contains specialized ion-transporting cells, the chloride cells (also called mitochondrion-rich cells), which are believed to be the main site of active iontransport (12, 35, 37) and nearly all of the calcium uptake infreshwater in teleosts (19). In this study, we applied for the firsttime the immunolocalization of the Na�-K�-ATPase in thegills of A. naccarii, an immunostaining procedure widely usedin fish to specifically visualize the chloride cells. In controlfish, the labeling procedure revealed the presence of numerouschloride cells located mostly on the afferent region of the gill
Fig. 5. A–D: distribution of the chloride cells (CCs) in the gillfilament of sturgeon. CCs were visualized by immunohisto-chemistry for the Na�-K�-ATPase (dark staining). A andB: General distribution of the CCs, whole mount immunohis-tochemistry in A and paraffin section in B. CCs are mostlylocated on the epithelium lining the afferent region of thefilament (Af) and on the region of the lamellae (Lam) close tothe Af. The efferent region of the filament (Ef) (A and B) isdevoid of CCs. C: transverse section through the afferent regionof the filament. On both sides of the filament, the afferentfilament epithelium (AFE) hosts numerous CCs. D: trans-verse section through the filament showing lamellae close tothe afferent region of the filament. The lamellar epithelium(LE) and the interlamellar filament epithelium (IFE) displaynumerous CCs. The AFE, the IFE, and the LE illustrated in Band C were chosen for CC quantification. Scale bars: A, 500�m; B, 200 �m; C and D, 50 �m. E: chloride cell (CC) densityin different regions of the gill epithelium in juvenile sturgeonfed on calcium-sufficient and calcium-deficient diet for a periodof 8 wk. Results are shown as means � SE (n � 8 pertreatment). *Significant (Students t-test P � 0.05) differencesbetween treatments in discrete gill regions.
R890 CALCIUM REGULATION IN STURGEON
AJP-Regul Integr Comp Physiol • VOL 293 • AUGUST 2007 • www.ajpregu.org
by guest on Novem
ber 7, 2012http://ajpregu.physiology.org/
Dow
nloaded from

filament; on the filament epithelium covering the trailing edgeand the interlamellar spaces but also on the lamellae. Chloridecells have already been described in the gill of A. naccariiusing transmission electron microscopy (8) or the zinc iodide-osmium staining procedure (9, 33). According to the authors, infreshwater acclimated fish, the chloride cells are present in thelamellar and interlamellar epithelia, but the filament epitheliumwas not studied (8, 33) or did not show chloride cells (9). Chloridecells were also observed all over the filament epithelium in thesturgeon Acipenser oxyrinchus (4).
A whole mounted staining of gill filament was also per-formed after immunostaining for the Na�-K�-ATPase. Thistechnique clearly illustrated the distribution of chloride cellsand particularly showed that the presence of lamellar chloridecells is a characteristic feature of the A. naccarii in normal
conditions and confirms our observations made on paraffinsections.
In our study, we observed an increase in chloride cell densityof 20% in the calcium-deficient diet group compared withcontrols. This increase in chloride cell numbers in the gillcorrelates with the increase of gill Na�-K�-ATPase activity.The values of gill Na�-K�-ATPase activity measured inA. nacarii are similar to values published for other freshwaterspecies or euryhaline species adapted to freshwater such asstingray (36), rainbow trout (28), or killifish (40). Both theincrease in chloride cell numbers and the increase in theactivity of the Na�-K�-ATPase in response to deficient dietarycalcium suggests the activation of compensatory mechanismsleading to an increase in calcium uptake from the water (18), inwhich apical epithelial calcium channels in gill nonchloridecells could also be involved (41). However, the impairedgrowth of the sturgeon on reduced calcium diets indicates thatbranchial epithelial transport only partially compensates fordietary deficiency.
PTHrP and Calcium Regulation in Sturgeon
Similar to what has been observed in teleosts (7, 21, 23),PTHrP(1-34) strongly modified calcium balance in the stur-geon A. nacarii. The action of PTHrP(1-34) was achievedthrough two processes: 1) stimulation of calcium uptake and2) inhibition of calcium efflux from the fish to water. Botheffects are consistent with its action as a hypercalcemic factorand are similar to PTHrP effects in sea bream larvae (23).Because in our study, the sturgeon were in freshwater wherelow drinking rates are expected, most of the effect of PTHrP(1-
Fig. 7. Effect of intraperitoneal single injection of 10 and 50 ng/g (1-34)PTHrP in whole body efflux (A) and uptake (B) in sturgeon juveniles.Results are shown as means � SE (n � 6). *Statistically significant differencesfrom controls were assessed by one-way ANOVA (P � 0.05, Bonferroni test).
Fig. 6. Effect of intraperitoneal single injection of saline (0), 10 or 50 ng/gPTHrP(1-34) on circulating plasma levels of PTHrP (A), total plasma calcium(B) in sturgeon juveniles sampled 4 or 24 h and the ratio of filtered/totalcirculating plasma levels of calcium (C) in sturgeon juveniles sampled 24 hafter a single injection of 10 and 50 ng/g (1-34)PTHrP. Results are shown asmeans � SE (n � 8). *Statistically significant differences from controls wereassessed by one-way ANOVA (P � 0.05, Bonferroni test).
R891CALCIUM REGULATION IN STURGEON
AJP-Regul Integr Comp Physiol • VOL 293 • AUGUST 2007 • www.ajpregu.org
by guest on Novem
ber 7, 2012http://ajpregu.physiology.org/
Dow
nloaded from

34) was likely to have been through the gills. Considering theapparent similarity in calcium handling between sturgeon andteleosts, it is likely that basolateral calcium movement in thegill is mediated by both the Na�/Ca2� exchanger and Ca2�-ATPase. Both transport systems respond to prolactin treatmentin teleosts (16) and could be likely targets for PTHrP. This issupported by the fact that in the parathyroidectomized ratkidney, PTH is able to restore the levels of protein expressionof Na�/Ca2� exchanger (NCX1, basolateral) and the epithelialcalcium channel (transient receptor potential cation channelsubfamily V member 5, apical) (45). This relationship may alsohold true in fish, and it is possible that one or more membersof the PTH/PTHrP family is involved in the regulation oftranscellular calcium transport in the epithelium.
The hypercalcemic action of PTHrP in sturgeon is reflectedby the direct increase in total plasma calcium in response tosingle injections. After hormonal treatment, the plasma ionicfraction remains unchanged and the fraction of calcium boundto protein is significantly increased. This fact suggests that theregulation of plasma calcium in chondrostean fish is very tight,keeping constant the concentration of ionic calcium in circu-lation. This observation suggests that calcium binding proteinscan act as temporary stores keeping ionic calcium constant andthat PTHrP may be involved in this process. Although thenature and identity of the calcium-binding proteins is un-known, in adult sturgeon total calcium increases in parallelwith vitellogenin (27).
In conclusion, the present study demonstrates that sturgeontightly regulate calcium balance and that PTHrP is a likelycandidate in the endocrine regulation of calcium in this groupof fish. The bioactivity of PTHrP(1-34) in sturgeon, furthersupports the suggestion for a generalized importance of thispeptide family in vertebrates and fish, in particular (7). Thetargets and mechanisms by which PTHrP exerts its hypercal-cemic require scrutiny.
ACKNOWLEDGMENTS
The authors thank J. Reis, M. Viegas, and A. Silva (Ramalhete MarineStation, Faro, Portugal) for fish care. The �5 monoclonal antibody developedby D. M. Fambrough was obtained from the Developmental Studies Hybrid-oma Bank developed under the auspices of the NICHD and maintained by theUniversity of Iowa, Department of Biological Sciences, Iowa City, IA.
GRANTS
This research has been carried out with the financial support of theCommission of the European Union, Quality of Life and Management ofLiving Resources specific Research Technology and Development program(Q5RS- Q5RS-2001-02904) and by European Social Fund and national fundsunder Portuguese National Science Foundation (FCT) project POCTI/CVT/48946/2002. P. Guerreiro and C. Haond were in receipt of FCT fellowshipsSFRH/BPD/9464/02 and SFRH/BPD/11527/02, respectively.
REFERENCES
1. Abbink W, Bevelander GS, Rotllant J, Canario AV, Flik G. Calciumhandling in Sparus auratus: effects of water and dietary calcium levels onmineral composition, cortisol and PTHrP levels. J Exp Biol 207: 4077–4084, 2004.
2. Abramoff MD, Magelhaes PJ, Ram SJ. Image processing with ImageJ.Biophotonics Int 11: 36–42, 2004.
3. Akino K, Ohtsuru A, Nakashima M, Ito M, Ting-Ting Y, Braiden V,Kawawaki T, Baba J, Yamashita S, Iwahori N. Distribution of theparathyroid hormone-related peptide and its receptor in the saccus vascu-losus and choroid plexus in the red stingray (Dasyatis akajei: Elasmo-branch). Cell Mol Neurobiol 18: 361–368, 1998.
4. Altinok I, Galli SM, Chapman FA. Ionic and osmotic regulation capa-bilities of juvenile Gulf of Mexico sturgeon, Acipenser oxyrinchus desotoi. Comp Biochem Physiol A 120: 609–616, 1998.
5. Baldisserotto B, Kamunde C, Matsuo A, Wood CM. A protective effectof dietary calcium against acute waterborne cadmium uptake in rainbowtrout. Aquatic Toxicol 67: 57–73, 2004.
6. Bradford MM. Rapid and sensitive method for quantitation of microgramquantities of protein utilizing principle of protein-dye binding. AnalBiochem 72: 248–254, 1976.
7. Canario AVM, Rotllant J, Fuentes J, Guerreiro PM, Teodosio HR,Power DM, Clark MS. Novel bioactive parathyroid hormone and relatedpeptides in teleost fish. FEBS Lett 580: 291–299, 2006.
8. Carmona R, Garcia-Gallego M, Sanz A, Domezain A, Ostos-GarridoMV. Chloride cells and pavement cells in gill epithelia of Acipensernaccarii: ultrastructural modifications in seawater-acclimated specimens. JFish Biol 64: 553–566, 2004.
9. Cataldi E, Ciccotti E, Dimarco P, Disanto O, Bronzi P, Cataudella S.Acclimation trials of juvenile ialian sturgeon to different salinities—morphophysiological descriptors. J Fish Biol 47: 609–618, 1995.
10. Choudhury A, Dick TA. The historical biogeography of sturgeons(Osteichthyes : Acipenseridae): a synthesis of phylogenetics, palaeontol-ogy and palaeogeography. J Biogeogr 25: 623–640, 1998.
11. Eckhard Witten P, Hall BK. Seasonal changes in the lower jaw skeletonin male Atlantic salmon (Salmo salar L.): remodelling and regression ofthe kype after spawning. J Anat 203: 435–450, 2003.
12. Evans DH, Piermarini PM, Choe KP. The multifunctional fish gill:dominant site of gas exchange, osmoregulation, acid-base regulation, andexcretion of nitrogenous waste. Physiol Rev 85: 97–177, 2005.
13. Faustino M, Power DM. Developmental of osteological structures in thesea bream: vertebral column and caudal Wn complex. J Fish Biol 52:11–22, 1998.
14. Flik G, Fenwick JC, Kolar Z, Mayergostan N, Bonga SEW. Whole-body calcium flux rates in cichlid teleost fish Oreochromis-mossambicusadapted to fresh-water. Am J Physiol Regul Integr Comp Physiol 249:R432–R437, 1985.
15. Flik G, Fenwick JC, Kolar Z, Mayergostan N, Wendelaar Bonga SE.Effects of low ambient calcium levels on whole-body Ca-2� flux rates andinternal calcium pools in the fresh-water cichlid teleost, Oreochromismossambicus. J Exp Biol 120: 249–264, 1986.
16. Flik G, Klaren PHM, Schoenmakers TJM, Bijvelds MJC, VerbostPM, Bonga SEW. Cellular calcium transport in fish: Unique and universalmechanisms. Physiol Zool 69: 403–417, 1996.
17. Flik G, Perry SF. Cortisol stimulates whole-body calcium-uptake and thebranchial calcium-pump in fresh-water rainbow trout. J Endocrinol 120:75–82, 1989.
18. Flik G, Rentierdelrue F, Bonga SEW. Calcitropic effects of recombinantprolactins in Oreochromis mossambicus. Am J Physiol Regul Integr CompPhysiol 266: R1302–R1308, 1994.
19. Flik G, Verbost PM. Calcium-transport in fish gills and intestine. J ExpBiol 184: 17–29, 1993.
20. Fuentes J, Eddy FB. Drinking in freshwater, euryhaline and marineteleosts. In: Ionic Regulation in Animals, edited by Hazon N, Eddy, FB,and Flik, G. New York: Springer-Verlag, pp. 135–149, 1997.
21. Fuentes J, Figueiredo J, Power DM, Canario AVM. Parathyroid hor-mone-related protein regulates intestinal calcium transport in sea bream(Sparus auratus). Am J Physiol Regul Integr Comp Physiol 291: R1499–R1506, 2006.
22. Guerreiro PM, Fuentes J, Canario AVM, Power DM. Calcium balancein sea bream (Sparus aurata): the effect of oestradiol-17 beta. J Endocri-nol 173: 377–385, 2002.
23. Guerreiro PM, Fuentes J, Power DM, Ingleton PM, Flik G, CanarioAVM. Parathyroid hormone-related protein: a calcium regulatory factor insea bream (Sparus aurata L.) larvae. Am J Physiol Regul Integr CompPhysiol 281: R855–R860, 2001.
24. Guerreiro PM, Renfro JL, Power DM, Canario AV. The parathyroidhormone family of peptides: structure, tissue distribution, regulation, andpotential functional roles in calcium and phosphate balance in fish. Am JPhysiol Regul Integr Comp Physiol 292: R679–R696, 2007.
25. Ichii T, Mugiya Y. Effects of a dietary deficiency in calcium on growthand calcium uptake from the aquatic environment in the goldfish, Caras-sius auratus. Comp Biochem Physiol A 74: 259–262, 1983.
26. Ingleton PM, Hazon N, Ho PM, Martin TJ, Danks JA. Immunodetec-tion of parathyroid hormone-related protein in plasma and tissues of an
R892 CALCIUM REGULATION IN STURGEON
AJP-Regul Integr Comp Physiol • VOL 293 • AUGUST 2007 • www.ajpregu.org
by guest on Novem
ber 7, 2012http://ajpregu.physiology.org/
Dow
nloaded from

elasmobranch (Scyliorhinus canicula). Gen Comp Endocrinol 98: 211–218, 1995.
27. Linares-Casenave J, Kroll KJ, Van Eenennaam JP, Doroshov SI.Effect of ovarian stage on plasma vitellogenin and calcium in culturedwhite sturgeon. Aquaculture 221: 645–656, 2003.
28. Lingwood D, Harauz G, Ballantyne JS. Regulation of fish gill Na�-K�-ATPase by selective sulfatide-enriched raft partitioning during seawateradaptation. J Biol Chem 280: 36545–36550, 2005.
29. Marshall WS, Bryson SE. Transport mechanisms of seawater teleostchloride cells: An inclusive model of a multifunctional cell. Comp Bio-chem Physiol A 119: 97–106, 1998.
30. Marshall WS, Bryson SE, Burghardt JS, Verbost PM. Ca2� transportby opercular epithelium of the fresh-water-adapted euryhaline teleost,Fundulus heteroclitus. J Comp Physiol [B] 165: 268–277, 1995.
31. McCormick SD. Methods for Nonlethal Gill Biopsy and Measurement ofNa�, K� -ATPase Activity. Can J Fish Aquat Sci 50: 656–658, 1993.
32. McCormick SD, Hasegawa S, Hirano T. Calcium-uptake in the skin ofa fresh-water teleost. Proc Natl Acad Sci USA 89: 3635–3638, 1992.
33. McKenzie DJ, Cataldi E, Di Marco P, Mandich A, Romano P,Ansferri S, Bronzi P, Cataudella S. Some aspects of osmotic and ionicregulation in Adriatic sturgeon Acipenser naccarii. II. Morpho-physiolog-ical adjustments to hyperosmotic environments. J Appl Ichthyol 15:61–66, 1999.
34. Pang PKT, Balbontin F. Effects of sex steroids on plasma calcium levelsin male killifish, Fundulus heteroclitus. Gen Comp Endocrinol 36: 317–320, 1978.
35. Perry SF. The chloride cell: Structure and function in the gills offreshwater fishes. Annu Rev Physiol 59: 325–347, 1997.
36. Piermarini PM, Evans DH. Effects of environmental salinity on Na�/K�-ATPase in the gills and rectal gland of a euryhaline elasmobranch(Dasyatis sabina). J Exp Biol 203: 2957–2966, 2000.
37. Pisam M, Rambourg A. Mitochondria-rich cells in the gill epithelium ofteleost fishes—an ultrastructural approach. Int Rev Cytol 130: 191–232,1991.
38. Rodriguez A, Gallardo MA, Gisbert E, Santilari S, Ibarz A, SanchezJ, Castello-Orvay F. Osmoregulation in juvenile Siberian sturgeon (Aci-penser baerii). Fish Physiol Biochem 26: 345–354, 2002.
39. Rotllant J, Worthington GP, Fuentes J, Guerreiro PM, Teitsma CA,Ingleton PM, Balment RJ, Canario AVM, Power DM. Determinationof tissue and plasma concentrations of PTHrP in fish: development andvalidation of a radioimmunoassay using a teleost 1-34 N-terminal peptide.Gen Comp Endocrinol 133: 146–153, 2003.
40. Scott GR, Rogers JT, Richards JG, Wood CA, Schulte PM. Intraspe-cific divergence of ionoregulatory physiology in the euryhaline teleostFundulus heteroclitus: possible mechanisms of freshwater adaptation. JExp Biol 207: 3399–3410, 2004.
41. Shahsavarani A, McNeill B, Galvez F, Wood CM, Goss GG, HwangPP, Perry S. Characterization of a branchial epithelial calcium channel(ECaC) in freshwater rainbow trout (Oncorhynchus mykiss). J Exp Biol209: 1928–1943, 2006.
42. Sundell K, Norman AW, Bjornsson BT. 1,25(Oh)2 Vitamin-D3 in-creases ionized plasma calcium concentrations in the immature Atlanticcod Gadus morhua. Gen Comp Endocrinol 91: 344–351, 1993.
43. Trivett MK, Walker TI, Clement JG, Ho PM, Martin TJ, Danks JA.Effects of water temperature and salinity on parathyroid hormone-relatedprotein in the circulation and tissues of elasmobranchs. Comp BiochemPhysiol B 129: 327–336, 2001.
44. Trivett MK, Walker TI, Macmillan DL, Clement JG, Martin TJ,Danks JA. Parathyroid hormone-related protein (PTHrP) production sitesin elasmobranchs. J Anat 201: 41–52, 2002.
45. van Abel M, Hoenderop JGJ, van der Kemp A, Friedlaender MM,van Leeuwen J, Bindels RJM. Coordinated control of renal Ca2�
transport proteins by parathyroid hormone. Kidney Int 68: 1708–1721,2005.
46. Wagner GF, DeNiu P, Jaworski E, Radman D, Chiarot C. Develop-ment of a dose-response bioassay for stanniocalcin in fish. Mol CellEndocrinol 128: 19–28, 1997.
R893CALCIUM REGULATION IN STURGEON
AJP-Regul Integr Comp Physiol • VOL 293 • AUGUST 2007 • www.ajpregu.org
by guest on Novem
ber 7, 2012http://ajpregu.physiology.org/
Dow
nloaded from




















