
Clinically Driven Smart Implantable Electronic Devices Moving ...
Upload
independentCategory
view
2download
0
PRIMARY RESEARCH PAPER
A fully implantable multi-channel biotelemetry systemfor measurement of blood flow and temperature: a firstevaluation in the green sturgeon
A. Grans Æ M. Axelsson Æ K. Pitsillides ÆC. Olsson Æ J. Hojesjo Æ R. C. Kaufman ÆJ. J. Cech Jr.
Received: 26 March 2008 / Revised: 17 August 2008 / Accepted: 24 August 2008
� Springer Science+Business Media B.V. 2008
Abstract The objective of this study was to evaluate
a novel fully implantable radio-based blood flow
biotelemetry system which allows simultaneously
measurement of blood flow on two channels and
temperature on one channel, in fish. These are the first
recordings of blood flow from free-swimming fish,
showing that the system is capable of recording blood
flow in the ventral aorta (cardiac output) and celia-
comesenteric artery (gastrointestinal blood flow) in
green sturgeon Acipenser medirostris exposed to a
series of different stimuli for up to 7 days after
implantation. The results showed stable base line
recordings and blood flow was used to calculated heart
rate (fH) and stroke volume (Vs). It was possible to
reproduce the same type of responses as has previously
been reported during exposure to hypoxia, tempera-
ture, stress and feeding. The mass of our implant was
less than 2% of the body mass which is well within the
recommended sizes for surgically implanted telem-
etry transmitters and it fitted easily within the abdom-
inal cavity of the sturgeon. A fully implantable system
minimizes the risk of infection/expulsion and maxi-
mizes the likelihood that the studied fish will behave
naturally and be treated normally by surrounding
fish. The use of biotelemetry in basic comparative
physiology and applied animal ecology could help
scientists to collect information that has previously
been challenging to obtain and to open the possibility
for new types of physiological and ecophysiological
studies.
Keywords Biotelemetry � Blood flow �Temperature � Doppler flow � Chronic measurements
Introduction
Biotelemetry systems are rapidly becoming important
tools for in vivo studies in medical and comparative
physiology. The major benefits of biotelemetry record-
ing systems, compared to traditional methods where
the experimental animals are physically connected to
the acquisition equipment through wires, are reduced
confinement stress, reduced risk of infections and the
possibility of long-term measurements in semi-natural
or natural environments. Biotelemetry makes it pos-
sible to record, for example, the effects of social
interactions and also to correlate normal behaviours
with physiological variables, something which is
Handling editor: K. Martens
A. Grans (&) � M. Axelsson � C. Olsson � J. Hojesjo
Department of Zoology, University of Gothenburg,
P.O. Box 463, 405 30 Goteborg, Sweden
e-mail: [email protected]
K. Pitsillides
EndoSomatic Technologies LLC, Sacramento, CA, USA
R. C. Kaufman � J. J. Cech Jr.
Department of Wildlife, Fish and Conservation Biology,
University of California, Davis, Davis, CA, USA
123
Hydrobiologia
DOI 10.1007/s10750-008-9578-7

practically impossible with ‘‘hardwired’’, cage-con-
fined animals. Many previous interpretations of
physiological data may be heavily affected or even
incorrect due to the stress induced by surgery,
confinement or human handling. The fact that the
animals studied using biotelemetry are no longer
physically connected to the recording equipment and
have the freedom to move around in a more natural
way may thus lead to higher quality of the data
obtained.
Biotelemetry is not a new idea. The first successful
transmission of biological information from a living
animal was performed in 1869 (Marey, 1896).
Winters (1921) was probably the first who success-
fully used radiotelemetry when he introduced the use
of a radiolink for sailors to get emergency advice
from doctors onshore when accidents happened at
sea. More than 20 years later, Fuller & Gordon
(1948) were the first to describe the use of biotelem-
etry to measure physiological activities from
unrestrained animals. When the transistor was intro-
duced in 1952 it became possible to design smaller
and more efficient biotelemetry equipment and this
led to a rapid increase in the use of telemetric
devices.
Blood flow is a central variable in cardiovascular
research and by measuring the blood flow of several
separate blood vessels (e.g. cardiac output and
gastrointestinal blood flow) information is obtained
about both the distribution of blood between tissues
and heart rate. Over the last 30 years there have been
several attempts to design an implantable blood
flowmeter, although none has been aimed for use in
fish. The new system is developed with the aim to
solve two central problems in fish biology: to
quantitatively describe the processing of food and
to evaluate the effects of stress on physiological and
behavioural mechanisms. These problems can be
addressed by measuring changes in distribution
between systemic blood flow and gastrointestinal
blood flow together with heart rate and temperature.
Several interesting systems have been described
that use different flowmeter techniques, such as,
electromagnetic flowmeters (Fryer et al., 1975),
ultrasonic Doppler shift techniques (Cathignol et al.,
1976; Allen et al., 1978, 1979) and interferometric
ultrasonic techniques (Rader et al., 1975). However,
most system, have parts of the devices externally
mounted and are consequently limited for use in
larger animals (Franklin et al., 1964; Yonezawa
et al., 1989, 1992; Spelman et al., 1991; Kong et al.,
2007). Other problems with these devices are the high
power consumption (Fryer et al., 1975; Cathignol
et al., 1976; Allen et al., 1978, 1979) and the bulk
flow probes (Rader et al., 1975).
Recently the first paper describing a multi-channel
biotelemetry system capable of recording both blood
pressure and blood flow was presented (Axelsson
et al., 2007). This multi-channel flow and pressure
biotelemetry system was designed for larger animals
and has so far been used and tested in pigs and
alligators (Axelsson et al., 2007). The system has
capacity to simultaneously measure blood flow at
four channels and blood pressure at three channels
together with ECG and temperature. This resulted in
a fairly large system (around 130 g, 60 cc volume)
(Axelsson et al., 2007), precluding application for
fish. However, based on this larger multi-channel
system a smaller device has been developed and the
present study includes the first evaluation of this
system. In the smaller system two of the blood flow
channels are retained together with one channel
allowing measurement of temperature. Due to the
difficulty to accurately measure blood pressure when
the external pressure fluctuates, e.g. when the freely
swimming fish changes depths, pressure was not
recorded in the smaller version. Our dual-channel
system consists of two parts: a component that is
fully implanted in the fish and a base station. Similar
to the larger system, the dual-channel system uses a
bidirectional communication protocol between the
base station and the implant. This allows not only
data to be received and processed from the implant,
but also commands to be sent to the implant.
Two important aspects during the development of
either system were the requirements for a fairly long
transmission range and a relatively long battery
lifetime. Long range transmission means that the
base station/computer and implant can communicate
with each other which allows the fish under study
more freedom compared to hardwired animals that
are usually restricted to relatively small experimental
chambers.
The evaluation study was conducted on green
sturgeon, Acipenser medirostris (Ayres), which is
native to North America. The choice of species
allowed us to compare our new data with older
studies using traditional methods, looking at the same
Hydrobiologia
123

cardiovascular variables in sturgeon, conducted in the
same laboratory (Crocker et al., 2000). The sturgeon
were available at different size ranges facilitating
determination of the appropriate size of animals for
the implant.
The aim of this study was to test the newly
developed dual blood-flow and temperature biote-
lemetry system in free-swimming fish. Data obtained
from telemetrically instrumented animals exposed to
temperature challenges, hypoxia, chasing and feeding
in the laboratory were compared with data obtained
from earlier studies using traditional techniques.
Material and methods
Animals used
The green sturgeon (n = 3) used in this study were
3 years old and had the masses 3.3 (sturgeon #1), 3.6
(sturgeon #2) and 4.2 (sturgeon #3) kg. They were
descendants of wild-caught Klamath River stur-
geon that were artificially spawned in 2004 (Van
Eenennaam et al., 2001). The eggs were incubated at
the University of California, Davis, at the Center for
Aquatic Biology and Aquaculture (CABA) and the
juveniles were kept in air-equilibrated water at tem-
peratures similar to the Klamath River (11–15�C)
during late spring. The fish were fed commercial
Silvercup trout pellets at 3–5% body weight ration per
day based on a feeding table for white sturgeon
Acipenser transmontanus (Richardson) that also has
been successfully used for green sturgeon (Kaufman
et al., 2006). At 31 days post-hatching, the fish were
placed in round 284-l fibreglass holding tanks and kept
there for their first 2 years, and then transferred to
12,800-l tanks until needed for experiments. The tanks
received a continuous flow of air-equilibrated, 19�C
well water.
Short description of the newly developed system
A simplified block diagram of the system is presented
in Fig. 1. This fully implantable biotelemetry system
consists of two channels for Doppler-based flow
measurements and one channel for temperature
measurement. The Doppler flowmeter is designed to
reduce power consumption through micro-power
analog and radio-frequency integrated circuits, as
well as advanced power management techniques. For
a further description see Axelsson et al. (2007). The
power reducing design results in a system which can
run on a single AA-size lithium battery with all three
channels operating continuously for approximately
14 days. Table 1 summarizes the technical specifica-
tions of the biotelemetry system. The implantable
flowmeter is also capable of timed-acquisition oper-
ation where the average power consumption is further
reduced. The timed-acquisition operation controls the
turning on and off of the implant in repeated, timed
intervals, thus substantially extending battery life.
The implant stores in internal memory its calibra-
tions for the flow and temperature channels. These
calibrations are easily accessed from the base station
user interface by sending a calibration command to the
implant. The implant responds with a 3-step calibra-
tion signal for the two flow channels and a 2-step
calibration signal for the temperature. The bidirec-
tional transceiver range is approximately 2–6 m
depending on enclosure, water depth and surrounding
radio frequency (RF) interference.
Implant description
The implant is controlled by a low-power microcon-
troller that generates all power control and timing
signals to the Doppler flowmeter and temperature
subsystems. The microcontroller also encodes and
formats the data for transmission through the RF link,
and processes any received commands. The Doppler
Flow 1Flow 2Temp
Trigger
RS232
RFLink Digital to
AnalogConverters
Controller
Battery
Flow 1Flow 2
Temp TempSensor
RFLinkController
DopplerFlowmeter
Module
A
B
Fig. 1 Block diagram of (A) base station and (B) implant
Hydrobiologia
123

flowmeter module operates at 20 MHz. Each channel
has independent adjustment controls for range-gate
positioning, signal direction and channel on/off.
Temperature is measured using an internal, digital
sensor.
Base station description
There are three basic functions that are performed in
the base station decoder/controller unit:
1. Reception and decoding of data from implant.
2. Transmission of user-selected commands and
configuration parameters to the implant.
3. Conversion of decoded implant data to analog
signals (using the digital-to-analog section). The
control of the implant/base station is mediated
via the computer using a custom-made program
and a serial communication protocol between the
computer and the base station.
The analog output from the base station was
connected to a commercial data acquisition system
(ML865 PowerLab 4/25T Data acquisition System,
AD Instruments Pty Ltd, Castle Hill, Australia)
running the software Chart 5 for Windows on a
PC-computer (Dell Latitude 820). The sampling rate
was set to 100 Hz for all channels.
Preoperative care
The sturgeon were anaesthetized by placing them in a
40-l well water tank containing 0.2 g l-1 3-amino-
benzoic acid ethyl ester (MS-222), 10 g l-1 NaCl and
buffered to pH 7.0 with 4.2 g l-1 NaHCO3. The
sturgeon were kept in the anaesthetics until ventila-
tory movements ceased and then weighed and
measured before being transferred to an operating
table covered with wet sponges. The fish were
positioned with the ventral side facing up and the
eyes shielded from the bright light using wet paper
towels. Anaesthesia was maintained by pumping
oxygenated, buffered MS-222 (0.075 g l-1) over
the gills during the surgery. A sterile surgical drape
(1051 Incise Drape, 3 M, USA) was used to cover
the fish and to keep the surgical surface sterile and
sterile gloves (Sensi-Touch Latex Powdered Sterile
Surgeons Glove, Ansell Healthcare Products Inc.,
Canada) were used throughout the surgery. Before
the surgery, the fish were injected with 2.5 mg/kg
[1 M] Enrofloxacin (Baytrilo�, Bayer, USA) antibi-
otics and all surgical instruments, the implant and
associated electrical leads were sterilized by the use
of a cold sterilant (Cidex, Johnson & Johnson
Company, USA). The surgical precautions taken
have been stressed in several reviews on fish surgery
(Butcher & Wildgoose, 2001; Fontenot & Neiffer,
2004) and were decided together with veterinary staff
at the University of California (UC), Davis. All
animal experiments were performed in accordance
with national and local ethical guidelines (EH&S
protocol: 07-12677).
Gastrointestinal blood flow and cardiac output
Green sturgeon have a single large celiacomesenteric
artery (CoMe) that supplies the entire gastrointestinal
tract with blood. To expose the celiacomesenteric
artery, a 5-cm-long midventral incision was made
posterior to the pectoral girdle and the liver was
carefully retracted. Gastrointestinal blood flow
(qCoMe) was recorded using silicon cuff transducer
probe with two integral canals for insertion of sutures
Table 1 A summary of the specifications for the system
Dimensions and battery life
Implant dimensions 60 9 30 9 10 mm
Battery pack dimensions 50 mm, Ø15 mm
Total weight 45 g
Battery life 340 h continuously ON
Doppler flowmeter
Number of channels 2
Ultrasound frequency 20 MHz
Pulse repetition frequency 64 kHz
Minimum range gate adjustments 1 mm
Maximum range gate adjustments 6 mm
Doppler frequency shift measured [18 kHz
Blood velocity range [100 cm/s
Transducer diameter 0.7 mm
Temperature
Number of channels 1
Temperature range 0–50�C
Resolution 0.0625�C
Radio frequency link section
Frequency 433 MHz
Power out 0 dBm
Hydrobiologia
123

(ES-2.5, Iowa Doppler products, Iowa City, USA)
that were placed around the CoMe. The probe cuffs
were closed using the integrated silk sutures.
In order to measure cardiac output (Q), a 2-cm
ventral incision was made posterior to the gill
juncture and carefully, without disrupting the peri-
cardium or damaging any vessels, the dermal and
sub-dermal musculature and connective tissue were
separated using blunt dissection tools to expose the
ventral aorta. A silicon, cuff-type Doppler blood flow
transducer with two integral canals for insertion of
sutures (ES-4.0, Iowa Doppler products, Iowa City,
USA) was placed around the ventral aorta. To
internalize the lead from the ventral aortic flow
probe, the probe was tunnelled under the skin from
the ventral incision to the midventral incision. The
probe was then placed around the ventral aorta and
closed using the integrated silk suture. The wires
from the probes were anchored with a single stitch of
3/0 silk suture in the sub-dermal muscle tissue. The
implant and battery were then carefully placed in the
abdominal cavity and the retracted organs restored to
their places of origin. The two incisions were closed
using sterile 3/0 nylon monofilament suture. The
locations of the probes and the implant are illustrated
in Fig. 2.
The surgical procedure including anaesthesia and
awakening took between 60 and 90 min.
Postoperative care
Post-operatively the sturgeon were given a subcuta-
neous injection of 0.1 mg/kg Torbugesic (butorphanol,
Fort Dodge, Iowa, USA) for post-operative pain relief,
in accordance with veterinarian instructions. The fish
were moved to 1.3-m-diameter fibreglass tanks
(750 l), where they could move freely, for recovery.
The tank received a continuous flow of air-equili-
brated, 19�C well water. Fish were allowed at least
24 h to recover from surgery before experimental
exposures started.
Experimental protocol
Cardiac output (Q), gastrointestinal blood flow (qCoMe)
and temperature (T) were continuously recorded
during the tests. Heart rate (fH) was calculated from
the phasic cardiac output signal using the cyclic
Q
qCoMe
Implant
battery
Fig. 2 Arrangement of the
implant, battery and the two
Doppler flow probes that
recorded cardiac output (Q)
and gastrointestinal blood
flow (qCoMe) inside the
sturgeon. The implant and
battery were positioned in
the abdominal cavity and
the flow probes measuring
Q and qCoMe were placed on
the ventral and
celiacomesenteric arteries,
respectively
Hydrobiologia
123

measurement feature in the Chart 5 software, while
stroke volume (Vs) was calculated from Q and fH.
During the experiments, all sturgeon were exposed
to three separate stimuli (hypoxia, increased water
temperature and chasing) once. Before each stimulus
all variables were recorded for at least 1 h to ensure
stable baseline values. The order of the stimuli was
randomized and the fishes were allowed at least 24 h
recovery between exposures. Precautions were made
to minimize long-term stress for the sturgeon in the
experiment. Similarly, the chasing exposure was
designed to give short-term effects. Chasing is a
comparably mild stimulus compared with other
previously used methods such as netting and water
reduction (Lankford et al., 2005), minimizing the
recovery time needed, and therefore reducing the risk
of carry-over effects between trials.
Lankford et al. (2005) showed in a study on
juvenile green sturgeon that plasma levels of two
indicators of stress, cortisol and glucose, were never
elevated for more than 180 min after a single stressor.
However, chronic stress (2–3 stressors randomly each
day) caused elevated levels of plasma glucose. More
than three stressors a day were not possible since it
causes increased mortality rate (Cech, unpublished).
In our study we wanted to measure blood flow
responses caused by several stimuli, with minimal
long-term effects to minimize the carry-over effects
between the different days and exposures. It has been
shown that green sturgeon stressed during nights
show higher levels of plasma cortisol and glucose
than do sturgeon stressed during the day (Lankford
et al., 2003). Consequently, all exposures were con-
ducted between 06:00 and 18:00 to minimize the
recovery times. It has also been shown that temper-
ature may affect recovery after stressful events.
Green sturgeon kept at 19�C recover from a 1-min
air emersion stressor faster (based on plasma cortisol
levels) compared to animals exposed to the same
stressor but acclimated to a lower temperature of
11�C (Lankford et al., 2003). Hence, the fish were
kept at 19�C, except during the temperature
challenge.
Temperature exposure
Heated (30�C), air-equilibrated well water was
pumped into the holding tank at a flow of 1.8 l min-1,
leading to a temperature increase of approximately
1.25�C every 30 min. The temperature was allowed to
increase from 19 to ca. 26�C after which the heated
water was turned off and the water temperature
returned to pre-test values.
Hypoxia exposure
Nitrogen gas (N2) was introduced via gas diffuser
stones into the holding tank and partial oxygen
pressure (PO2) was measured every 20 min using an
oxygen electrode (E101 Cameron oxygen electrode,
Analytical Sensors, Inc, Sugar Land, TX, USA). The
flow of N2 was increased stepwise from 20 to
40 ml min-1 to decrease PO2 from the initial value
of 120 mm Hg down below 60 mm Hg before the N2
flow was turned off and the water in the tank slowly
returned to normoxia.
Chasing
The sturgeon were manually chased with a net for
15 min. Chasing as a general stressor has previously
been used as for shortnose sturgeon, Acipenser
brevirostrum (Lesueur) (Beyea et al., 2005) and a
5-min chasing session also has been used previously as
a stressor for green sturgeon (Lankford et al., 2005).
Feeding
After the three stimuli described above one of the
animals was lightly anaesthetized, placed on its back
and fed by gavage. The meal consisted of commercial
Silvercup trout pellets equivalent to 1% of the body
weight. This is a common method in laboratory based
feeding studies and makes it possible to control for
meal size and exact time of feeding. The postprandial
effects on Q and qCoMe were then followed contin-
uously for approximately 33 h.
Results
Validation of technique
Green sturgeon were ideal to use for validating this
new system. Surgery was fast and minimal and they
seemed unaffected by the implants. Signals were
strong and clear despite movements in the tanks.
During the chasing all the animals were very active,
Hydrobiologia
123

but no deterioration of the signal quality could be
seen, indicating a robust signal transmission.
Figure 3 shows a longer (60-min) section of
continuous recording of Q and qCoMe, in sturgeon
#1, 24 h after implantation. From the two traces of
raw data, a 30-s section is enlarged to verify the
quality of the signals. The left part of the lower
panels shows the unfiltered signal, while the right
shows the signal after filtering using a Triangular
Bartlett window (window width: 25 points). During
the exposures, adjustments in the setting were done
manually which resulted in some variations in the
duration of each step in the treatments. Hence, only
the response of one sturgeon to each stimulus is
presented in the figures (Figs. 4–7).
Temperature
Figure 4 shows the original recording of sturgeon #3
during the temperature challenge. Only a moderate
increase in Q, Vs and fH occurred when the temper-
ature were raised from 19 to 24�C. In contrast, qCoMe
increased by more than 150% when the temperature
increased above around 23�C. The response differed
among the three tested fishes with two animals
showing a decreased qCoMe (\50%) when the tem-
perature was increased above 25�C. Between 19 and
24�C, all three fish showed similar responses and
mean values (change in response at 24�C in percent
of response at 19�C) are presented in Table 2. The
fish body temperature was obtained from the implant
0
40
80
120
0
20
40
Blo
odF
low
(cm
s-1
)
Raw data Same trace Smoothed*
* Triangular (Bartlett), window width: 25 points 10 s
10 min
0
0
20
100
Blo
odF
low
(cm
s-1
)
A
BQ
qCoMe
Q
qCoMe
Fig. 3 Verification of signal quality from the two flow probes.
(A) Raw data for cardiac output (Q) and gastrointestinal blood
flow (qCoMe) during 60 min continuous recording in a resting
sturgeon under control conditions. (B) A section of the trace in
A corresponding to 30 s with raw data for cardiac output (Q)
and gastrointestinal blood flow (qCoMe) shown on the left and
the same trace after filtering using a Triangular Bartlett,
window (window width: 25 points) on the right
Hydrobiologia
123

and water temperature measured with an additional
thermometer showed no lag time between the
increase in water temperature and fish temperature.
Hypoxia
Figure 5 shows the data from sturgeon #2 during the
hypoxia challenge. In this animal, Q, Vs and fHincreased when PO2 got below 80 mm Hg. The
opposite was seen in qCoMe where the flow decreased
95% compared with the initial value. Also during the
hypoxia trial the responses differed among individual
sturgeon. In sturgeon #3, bradycardia appeared when
PO2 was dropped to approx. 50 mm Hg. Mean values
for all animals comparing the responses at PO2 of 120
and 60 mmHg are presented in Table 2.
Chasing
Figure 6 shows the response from sturgeon #2 to the
15 min net chasing. Q, Vs and fH showed an increase
when the chasing started. Q and Vs increased ca. 20%,
but again the largest effect was seen in qCoMe, which
decreased 95% compared with the initial value. A
comparison of the responses prior and during chasing
is presented as mean values for all three animals in
Table 2.
Tem
p(°
C)
20 min0
17
19
21
23
25
27
0
100
200
300%
VsqCoMe
Q
T
f H(m
in-1
)
0
20
40
60 fH
Fig. 4 Trace from a temperature (T) challenge obtained from
sturgeon #3. The temperature was increased from 19 to 26�C
during a period of approximately 2 h. When the temperature
increased moderate increases occurred in cardiac output (Q),
stroke volume (VS) and heart frequency (fH), while the
gastrointestinal blood flow (qCoMe) increased by more than
150%
15 min
0
50
100
150
200
0
20
40
60
50
75
100
125
0
fH
PO2
f H(m
in-1
)%
PO
2(m
mH
g)
VsqCoMe
Q
Fig. 5 Trace from a hypoxia challenge obtained from sturgeon
#2. Cardiac output (Q), stroke volume (VS) and heart frequency
(fH) increased and the gastrointestinal blood flow (qCoMe)
decreased when partial oxygen pressure (PO2) declined below
80 mm Hg
Hydrobiologia
123
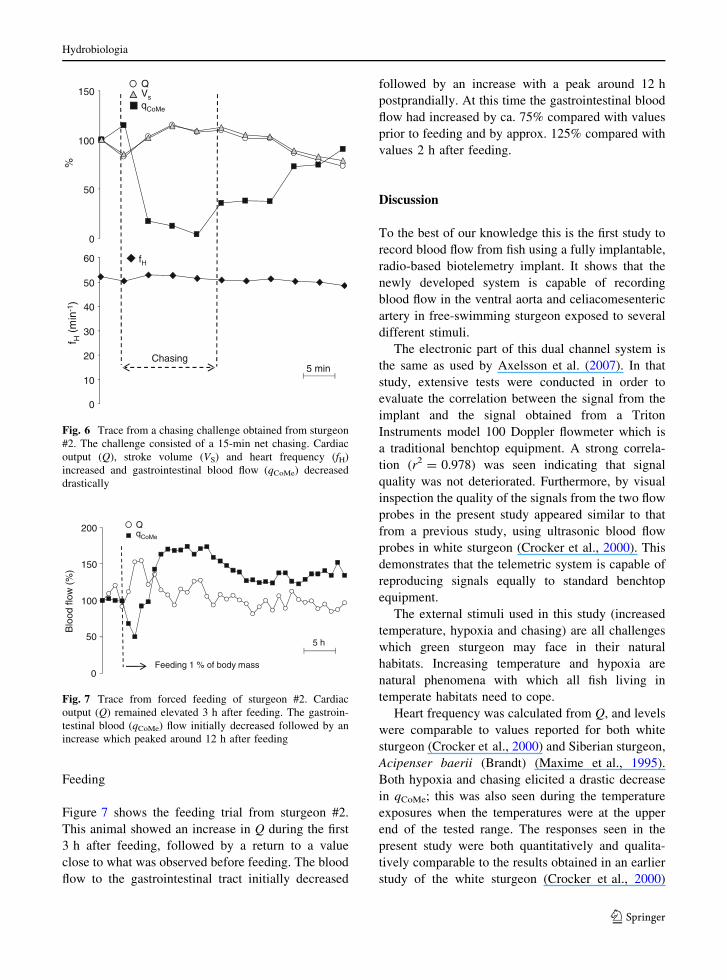
Feeding
Figure 7 shows the feeding trial from sturgeon #2.
This animal showed an increase in Q during the first
3 h after feeding, followed by a return to a value
close to what was observed before feeding. The blood
flow to the gastrointestinal tract initially decreased
followed by an increase with a peak around 12 h
postprandially. At this time the gastrointestinal blood
flow had increased by ca. 75% compared with values
prior to feeding and by approx. 125% compared with
values 2 h after feeding.
Discussion
To the best of our knowledge this is the first study to
record blood flow from fish using a fully implantable,
radio-based biotelemetry implant. It shows that the
newly developed system is capable of recording
blood flow in the ventral aorta and celiacomesenteric
artery in free-swimming sturgeon exposed to several
different stimuli.
The electronic part of this dual channel system is
the same as used by Axelsson et al. (2007). In that
study, extensive tests were conducted in order to
evaluate the correlation between the signal from the
implant and the signal obtained from a Triton
Instruments model 100 Doppler flowmeter which is
a traditional benchtop equipment. A strong correla-
tion (r2 = 0.978) was seen indicating that signal
quality was not deteriorated. Furthermore, by visual
inspection the quality of the signals from the two flow
probes in the present study appeared similar to that
from a previous study, using ultrasonic blood flow
probes in white sturgeon (Crocker et al., 2000). This
demonstrates that the telemetric system is capable of
reproducing signals equally to standard benchtop
equipment.
The external stimuli used in this study (increased
temperature, hypoxia and chasing) are all challenges
which green sturgeon may face in their natural
habitats. Increasing temperature and hypoxia are
natural phenomena with which all fish living in
temperate habitats need to cope.
Heart frequency was calculated from Q, and levels
were comparable to values reported for both white
sturgeon (Crocker et al., 2000) and Siberian sturgeon,
Acipenser baerii (Brandt) (Maxime et al., 1995).
Both hypoxia and chasing elicited a drastic decrease
in qCoMe; this was also seen during the temperature
exposures when the temperatures were at the upper
end of the tested range. The responses seen in the
present study were both quantitatively and qualita-
tively comparable to the results obtained in an earlier
study of the white sturgeon (Crocker et al., 2000)
Chasing5 min
0
50
100
150
0
10
20
30
40
50
60 fH
f H(m
in-1
)%
VsqCoMe
Q
Fig. 6 Trace from a chasing challenge obtained from sturgeon
#2. The challenge consisted of a 15-min net chasing. Cardiac
output (Q), stroke volume (VS) and heart frequency (fH)
increased and gastrointestinal blood flow (qCoMe) decreased
drastically
5 h
Feeding 1 % of body mass
Blo
odflo
w(%
)
0
100
200
50
150
qCoMe
Q
Fig. 7 Trace from forced feeding of sturgeon #2. Cardiac
output (Q) remained elevated 3 h after feeding. The gastroin-
testinal blood (qCoMe) flow initially decreased followed by an
increase which peaked around 12 h after feeding
Hydrobiologia
123

with the difference that the animals were swimming
freely without any physical connection to any
recording equipment in the present study.
A reduction in blood flow to the gastrointestinal
tract during exercise (induced swimming or chasing)
has been described in various species: for example,
Chinook salmon, Oncorhynchus tshawytscha
(Walbaum) (Thorarensen et al., 1993; Thorarensen
& Farrell, 2006) and rainbow trout, O. mykiss
(Walbaum), (Randall & Daxboeck, 1982). A decrease
in qCoMe during hypoxia has also been shown in
several species including European sea bass, Dicen-
trarchus labrax (L.), (Axelsson et al., 2002) and
Atlantic cod, Gadus morhua (L.) (Axelsson & Frit-
sche, 1991). All these studies support the hypothesis
that blood is distributed away from the stomach when
there is a need to optimize oxygen delivery, either due
to a reduction in available environmental oxygen
(hypoxia) or during exercise when there is an
increased demand from swimming muscles that needs
to be met by the cardiovascular system.
During hypoxia, a bradycardia was observed in
sturgeon #3 at the lowest partial oxygen pressure.
Hypoxic bradycardia is a well-documented response
in fish, including the closely related Adriatic sturgeon
where bradycardia developed at deeper levels of
hypoxia (Agnisola et al., 1996). Hypoxic bradycardia
is discussed in several reviews but is still not
completely understood (Randall, 1982; Farrell, 2007).
In the feeding test the sudden decrease in qCoMe
and increase in Q after feeding was most likely due to
stress caused by handling at the time of feeding.
Hyperemia in the gastrointestinal canal induced by
feeding is expected when activities such as gut
motility, intracellular biochemical activity and mem-
brane transport all increase after feeding (McCue,
2006). This has been described in several fish species
including sea raven, Hemitripterus americanus
(Gmelin), (Axelsson et al., 1989), red Irish lord,
Hemilepidotus hemilepidotus (Tilesius), (Axelsson
et al., 1989), Atlantic cod (Axelsson & Fritsche,
1991), European sea bass (Axelsson et al., 2002) and
Chinook salmon (Thorarensen et al., 1993). Due to
differences in temperature, amount of food given,
species-specific differences and effects of instrumen-
tation between the studies it is difficult to make direct
comparisons. However, in all cases feeding induces
an increase in blood flow to the gastrointestinal canal.
In some species this increase is provided by an
increase in cardiac output, while in other species
there is a redistribution of blood flow, leaving cardiac
output unchanged. The dynamics of the changes are
highly dependent on temperature and effects of
instrumentation. Axelsson et al. (2002) showed in
their study on the effects of feeding and hypoxia in
the European sea bass that instrumentation signifi-
cantly increased the gastric evacuation time. This is
an area where implantable telemetric systems will be
useful with the associated, increased possibility for a
successful recovery.
Few studies have successfully managed to link
natural behaviour with changes in blood flow due to
the difficulty in getting instrumented or in other ways
restrained fishes to behave naturally. The use of
biotelemetry in basic comparative physiology and
applied animal ecology could help scientists to
collect information that has previously been chal-
lenging to obtain and to open the possibility for new
types of physiological and ecophysiological studies
(Cooke et al., 2004).
Pros and cons with blood flow biotelemetry?
A perfect biotelemetry device should be very small
and fully implantable so that it will not affect the
bearer. The size of the biotelemetry implants, relative
to the animals to be studied, has been and is still one of
the largest problems when developing the systems.
Table 2 Response in cardiac output (Q), gastrointestinal blood flow (qCoMe), stroke volume (VS) and heart frequency (fH) to
increased temperature, hypoxia or chasing in green sturgeon (Acipenser medirostris)
Q qCoMe VS fH
Temperature 23.44 ± 16.48 45.28 ± 22.86 -5.78 ± 11.26 29.87 ± 2.99
Hypoxia -4.63 ± 16.80 -63.19 ± 15.10 3.57 ± 8.89 8.45 ± 9.79
Chasing 10.54 ± (5.37) -75.73 ± 11.80 7.22 ± 4.54 3.20 ± 1.67
Values are mean ± standard deviation from all three sturgeons, presented as percent change compared with the baseline response
(19�C, 120 mm Hg and before chasing, respectively) at peak response (24�C, 60 mm Hg and after chasing, respectively)
Hydrobiologia
123

The general recommendation is that the implant
should not exceed 2 to 5% of the body mass of the
animal, although the appropriate forms and mass
should be evaluated for each species because size and
morphology vary enormously (Jepsen et al., 2002).
The mass of our implant was \2% of the body mass
and the implant fitted easily within the abdominal
cavity of the sturgeon. Our system is also the first fully
implantable system capable of measuring blood flow
in fish. A fully implantable system minimizes the risk
of infection and expulsion (Schulz, 2003) and also
maximizes the likelihood that the studied fish will
behave naturally and be treated normally by sur-
rounding fish (Connors et al., 2002). Even the
presence of a small, protruding antenna has been
shown to elicit aggressive attacks from other individ-
uals of Atlantic salmon, Salmo salar (L.), smolts
(Connors et al., 2002).
Long-term effects of implants are often evaluated
using dummy implants (Connors et al., 2002; Jepsen
et al., 2002; Schulz, 2003). Since the primary goal of
the current study was to look at the functionality of
this newly developed blood flow telemetric system in
fish, this was not considered necessary at this stage.
However, the long-term effects of implantation will
be studied in more detail in future studies.
Until now the commercially available biotelemetry
equipment used in fish has been transmitters designed
to measure fH and electromyograms (EMG). EMG
telemetry has been an essential tool in studies that
have helped us, e.g. to understand better the migra-
tion and spawning ecology of sockeye salmon
Oncorhynchus nerka (Walbaum) (Hinch & Rand,
1998; Hinch & Bratty, 2000; Hinch et al., 2002;
Healey et al., 2003).
Estimating metabolic rate from blood flow
Metabolic rate (VO2) is the basis of life and the single
most important variable in a living system. Metabolic
rate is a function of cardiac output and arterio-venous
oxygen content. Several attempts have been made to
obtain information about VO2 in free-swimming fish,
based on fH. This was first described by Priede and
Tytler (1977) and has thereafter frequently been
used in other studies although with mixed results
(Armstrong et al., 1989; Lucas & Armstrong, 1991;
Lucas et al., 1991; Lucas, 1994; Armstrong, 1998;
Lefrancois et al., 1998). Because stroke volume and
heart rate can change independently, the use of heart
rate as an indicator of cardiac output has to be
considered carefully. For a somewhat reliable estimate
of VO2 from fH, one must make a laboratory calibra-
tion to obtain fH/VO2 correlation curves which can be
used in a field situation. It has been shown that the use
of fH as an indicator of metabolic rate is limited to
stable environments and that the VO2 values obtained
in the laboratory from confined animals may compare
poorly with those from fishes in their natural environ-
ments (Claireaux et al., 1995; Thorarensen et al.,
1996; Webber et al., 1998). In their study on Atlantic
cod, Webber et al. (1998) showed that, if the fH/VO2
correlation-curves are based on the recovery period,
VO2 can be overestimated by as much as 100%. For a
more detailed discussion describing the limitations of
fH as a predictor of VO2, see reviews by Thorarensen
et al. (1996) and Butler et al. (2004).
Q is a more reliable variable for prediction of VO2
because both fH and VS are considered. Therefore, a
biotelemetric system that simultaneous measures both
blood flow and heart rate would minimize the
problems associated with fH-based VO2 estimates
(Thorarensen et al., 1996; Webber et al., 1998;
Altimiras & Larsen, 2000). However, problems also
can arise from estimating VO2 from Q (Brodeur
et al., 2001), and it is important to stress that
correlations among Q, fH and VO2 may differ
between species and environments (Thorarensen
et al., 1996; Armstrong, 1998; Lefrancois et al.,
1998; Webber et al., 1998; Altimiras & Larsen,
2000; Brodeur et al., 2001).
Heart rate, obtained with biotelemetry devices, has
also been used to estimate variables such as activity
and digestion (Armstrong, 1986), although this
method also has been criticized for being biased. In
a study conducted on European sea bass, it was
shown that following stress or during digestion, fish
metabolic turn-over rates temporarily reached values
close to active VO2 (Lefrancois et al., 1998). Taking
into account the described problems with laboratory-
derived correlation curves based on fH values, with
the expected tachycardia response to digestion,
exercise and stress, the difficulty to separate the
relevant factors in a uncontrolled situation is obvious.
By using two separate blood flows it will be possible
not only to measure changes in both fH and VS but
also to measure regional blood flows (e.g. the blood
allocated to the gastrointestinal tract).
Hydrobiologia
123

The effect of welfare on data quality
Another important and interesting aspect of using
biotelemetric equipment instead of hardwired animals
is the possibilities for prolonged recovery after
surgery and reduced stress from handling. The time
needed for a fish to recover from surgery depends on
species, individuals and environment (Fontenot &
Neiffer, 2004). Most studies only allow a recovery
period of 12–48 h after surgery before onset of the
experiment. However, several studies have shown
elevated fH levels for much longer and have con-
cluded that 48 h of recovery after surgery may be too
short (Webber et al., 1998; Altimiras & Larsen, 2000;
Campbell et al., 2005). Webber et al. (1998) describe
a 2–7-day period for physiological recovery in
Atlantic cod. Altimiras & Larsen (2000) showed
how the metabolic scope decreased when rainbow
trout recovered for up till 2 days after surgery.
Therefore, short recovery periods might lead to
misinterpretations and false conclusions. One of the
main reasons for the commonly used short recovery
time is the high risk of infection after an extensive
surgery (Fontenot & Neiffer, 2004). Much of histor-
ical data might be biased by the poor health of
the studied fishes (Altimiras & Larsen, 2000;
Huntingford et al., 2006; Johansen et al., 2006). In
a study using bioelectric potential recordings to
measure fH, rainbow trout were handled and then left
to recover for at least 3 days. The basal heart rates of
these trout were much lower than those presented in
previous studies using traditional methods involving
surgical procedures (Altimiras & Larsen, 2000).
Long-term measurements using biotelemetry would
help to verify or reject proposed basal values for
physiological parameters. The focus in this study was
to test the system and to evaluate the quality of the
data obtained by comparing the results with data from
studies using traditional bench-test equipment on
hardwired animals. The aspects of recovery time on
the recorded variables and the possible differences
in response to the various challenges should be
examined in a future study.
Future applications for the system
It is clear that the biotelemetry system can record
cardiac output and gastrointestinal blood flow from
free-swimming sturgeon (at least in a laboratory
environment) and that the responses obtained with
this system compare well with results from earlier
studies. In this study the aim was primarily to
evaluate the functionality of the system and there was
no possibility to look at the long-term effects in the
measured variables if the animals were allowed full
recovery, something which is impossible with tradi-
tional recording methods. With the increased use of
fish models in most biological disciplines (Powers,
1989), the need for animal welfare guidelines asso-
ciated with animal models in research has emerged
(Harms, 2005; Johansen et al., 2006). The telemetry
system can also help the fish culture industry. With
increasing commercial values of farmed fish, it is of
both ethical and economical interests to keep healthy,
unstressed animals (Conte, 2004; Ashley, 2007). If
fish are stressed and the responses, including loss of
appetite, impaired growth and muscle wasting are
prolonged, the consequences for the fish farmers can
be devastating (Huntingford et al., 2006). Developing
a management protocol to reduce stress and its
consequences is a shared goal in aquaculture (Conte,
2004). Today there are several ways to monitor
cultured fish’s welfare, looking at both environmental
variables, such as levels of oxygen and carbon
dioxide, and physiological variables. Most physio-
logical variables are monitored by visual cues such as
changes in coloration or behaviours of the fish
(Huntingford et al., 2006; Ashley, 2007). Although
trials have been conducted using biotelemetry and
include measurements of muscular activity, gill
ventilation rate and heart rate, questions concerning
how culture conditions affect the fish’s health remain
unanswered (Baras & Lagardere, 1995). Each year
the aquaculture industry loses massive amounts of
money on problems related to stress and the farmers
are requesting new ways of monitoring the health of
the fish (Conte, 2004; Ashley, 2007).
With this novel biotelemetry system, a possibility
to conduct long-term monitoring in cultured fish
emerges. By monitoring both total cardiac output and
gastrointestinal blood flow together with temperature
within an aquaculture setting, it would be possible to
devise systems and operating procedures to minimize
stressful situations, optimizing fish growth rates and
production.
Another area where biotelemetric studies may
increase our understanding is the effects of climate
changes on fish populations. It has been shown that
Hydrobiologia
123

populations of some marine fishes are moving
northwards as ocean temperatures increase (Roessig
et al., 2004; Perry et al., 2005). Migration towards
colder northern water is possible in many marine
environments but may not be a solution for freshwa-
ter species, living in lakes and river systems.
Although Ficke et al. (2007) documented stream
fishes, moving to higher elevations, some populations
will have to either adapt or adjust to the changes
instead. If this is not possible, their scope for activity
and reproduction will be reduced, potentially jeopar-
dizing their survival. The seasonal differences and
shifting environments caused by global warming
alters both the physiology and behaviour of many
freshwater species (review by Farrell, 1997). The
effects of climate-induced changes are only possible
to study partially in the laboratory; it is important to
get information from animals in their natural envi-
ronment and biotelemetric systems are the only
possible way to obtain physiological data in this
situation. Because temperature changes directly
affect the metabolism of fish (and all other ectother-
mic animals) and because blood flow is a much better
estimate of metabolism compared with heart rate, the
biotelemetry system described in this paper could
be a useful tool in assessing effects of temperature
changes in fish, caused by either natural seasonal
variations or anthropogenic effects (Randall &
Brauner, 1991).
Conclusion
The first evaluation of this novel biotelemetric system
shows that it is capable of replicating results obtained
using traditional benchtop equipment. We show that
blood flow measurements are possible to conduct
with animals swimming in their ‘‘home’’ tanks
instead of in confined cages. To evaluate the advan-
tages of this system as compared with traditional
methods, further studies are needed.
Acknowledgements This research was supported by
Goteborg University Research Platform on Integrative
Physiology (GRIP), the Swedish Science Research Council
Grants B-DR 09856-304 and 621-2002-3869 (M. Axelsson),
the Helge Ax:son Jonsson foundation and the Knut & Alice
Wallenberg foundation. We gratefully acknowledge the
assistance of W. Wesley Dowd, Christa M. Woodley and the
rest of the staff at Department of Wildlife, Fish and
Conservation Biology, University of California, for adjusting
their time plans to make this study possible. K. Pitsillides owns
EndoSomatic Technologies, LLC, which developed the
biotelemetry system described in this manuscript and for
more information on the biotelemetry system visit
www.endosomatic.com.
References
Agnisola, C., D. J. Mckenzie, E. W. Taylor, C. L. Bolis &
B. Tota, 1996. Cardiac performance in relation to oxygen
supply varies with dietary lipid composition in sturgeon.
American Journal of Physiology 40: R417–R425.
Allen, H. V., J. W. Knutti & J. D. Meindl, 1978. Integrated
circuits for a bidirectional implantable pulsed Doppler
ultrasonic blood flowmeter. IEEE Journal of Solid-State
Circuits SC-13: 853–863.
Allen, H. V., J. W. Knutti & J. D. Meindl, 1979. Totally
implantable directional Doppler flowmeter. Biotelemetry
and Patient Monitoring 6: 118–132.
Altimiras, J. & E. Larsen, 2000. Non-invasive recording of
heart rate and ventilation rate in rainbow trout during rest
and swimming. Fish go wireless!. Journal of Fish Biology
57: 197–209.
Armstrong, J. D., 1986. Heart-rate as an indicator of activity,
metabolic-rate, food-intake and digestion in pike,
Esox-Lucius. Journal of Fish Biology 29: 207–221.
Armstrong, J. D., 1998. Relationships between heart rate and
metabolic rate of pike: integration of existing data. Jour-
nal of Fish Biology 52: 362–368.
Armstrong, J. D., M. C. Lucas, I. G. Priede & L. Devera, 1989.
An acoustic telemetry system for monitoring the heart-
rate of pike, Esox-Lucius L., and other fish in their nat-
ural-environment. Journal of Experimental Biology 143:
549–552.
Ashley, P. J., 2007. Fish welfare: current issues in aquaculture.
Applied Animal Behaviour Science 104: 199–235.
Axelsson, M., J. Altimiras & G. Claireaux, 2002. Post-prandial
blood flow to the gastrointestinal tract is not compromised
during hypoxia in the sea bass Dicentrarchus labrax.
Journal of Experimental Biology 205: 2891–2896.
Axelsson, M., Q. Dang, K. Pitsillides, S. Munns, J. Hicks &
G. S. Kassab, 2007. A novel, fully implantable, multi-
channel biotelemetry system for measurement of blood
flow, pressure, ECG, and temperature. Journal of Applied
Physiology 102: 1220–1228.
Axelsson, M., W. R. Driedzic, A. P. Farrell & S. Nilsson, 1989.
Regulation of cardiac-output and gut blood-flow in the
Sea Raven, Hemitripterus-americanus. Fish Physiology
and Biochemistry 6: 315–326.
Axelsson, M. & R. Fritsche, 1991. Effects of exercise, hypoxia
and feeding on the gastrointestinal blood-flow in the
Atlantic Cod Gadus morhua. Journal of Experimental
Biology 158: 181–198.
Baras, E. & J. P. Lagardere, 1995. Fish telemetry in aquacul-
ture: review and perspectives. Aquaculture International
3: 77–102.
Beyea, M. M., T. J. Benfey & J. D. Kieffer, 2005. Hematology
and stress physiology of juvenile diploid and triploid
Hydrobiologia
123

shortnose sturgeon (Acipenser brevirostrum). Fish Physi-
ology and Biochemistry 31: 303–313.
Brodeur, J. C., D. G. Dixon & R. S. Mckinley, 2001. Assess-
ment of cardiac output as a predictor of metabolic rate in
rainbow trout. Journal of Fish Biology 58: 439–452.
Butcher, R. L. & W. H. Wildgoose, 2001. BSAVA Manual of
Ornamental Fish, British Small Animal Veterinary Asso-
ciation; 2 Rev edition (19 Dec 2001).
Butler, P. J., J. A. Green, I. L. Boyd & J. R. Speakman, 2004.
Measuring metabolic rate in the field: the pros and cons of
the doubly labelled water and heart rate methods. Func-
tional Ecology 18: 168–183.
Campbell, H. A., V. S. Bishop, D. A. Davies & S. Egginton,
2005. Recording long-term heart rate in Paranototheniaangustata using an electronic datalogger. Journal of Fish
Biology 67: 1150–1156.
Cathignol, D., J. Y. Chapelon, B. Lavandier & B. Fourcade,
1976. Detail description of an implantable directional
Doppler flowmeter. Biotelemetry 3: 117–128.
Claireaux, G., D. Webber, S. Kerr & R. Boutilier, 1995.
Physiology and behaviour of free-swimming Atlantic cod
(Gadus morhua) facing fluctuating temperature condi-
tions. Journal of Experimental Biology 198: 49–60.
Connors, K. B., D. Scruton, J. A. Brown & R. S. Mckinley,
2002. The effects of surgically-implanted dummy radio
transmitters on the behaviour of wild Atlantic salmon
smolts. Hydrobiologia 483: 231–237.
Conte, F. S., 2004. Stress and the welfare of cultured fish.
Applied Animal Behaviour Science 86: 205–223.
Cooke, S. J., S. G. Hinch, M. Wikelski, R. D. Andrews,
L. J. Kuchel, T. G. Wolcott & P. J. Butler, 2004. Biote-
lemetry: a mechanistic approach to ecology. Trends in
Ecology & Evolution 19: 334–343.
Crocker, C. E., A. P. Farrell, A. K. Gamperi & J. J. Cech, 2000.
Cardiorespiratory responses of white sturgeon to envi-
ronmental hypercapnia. American Journal of Physiology
279: R617–R628.
Farrell, A. P., 1997. Effects of Temperature on Cardiovascular
Performance. Cambridge, Cambridge University Press.
Farrell, A. P., 2007. Tribute to P. L. Lutz: a message from the
heart—why hypoxic bradycardia in fishes? Journal of
Experimental Biology 210: 1715–1725.
Ficke, A. D., C. A. Myrick & L. J. Hansen, 2007. Potential
impacts of global climate change on freshwater fisheries.
Reviews in Fish Biology and Fisheries 17: 581–613.
Fontenot, D. K. & D. L. Neiffer, 2004. Wound management in
teleost fish: biology of the healing process, evaluation,
and treatment. Veterinary Clinics of North America:
Exotic Animal Practice 7: 57–86.
Franklin, D., N. W. Wason & R. L. Van Citters, 1964. Blood
velocity telemetered from unrestrained animals. Nature
203: 528–530.
Fryer, T. B., H. Sandler, W. Freund, E. P. Mccutcheon & I.
Carlson, 1975. A multichannel implantable system for
flow and ECG measurements. Journal of Applied Physi-
ology 39: 318–326.
Fuller, S. L. & T. M. Gordon, 1948. The radio inductograph, a
device for recording physiological activity in unrestrained
animals. Science 108: 287.
Harms, C. A., 2005. Surgery in fish research: common proce-
dures and postoperative care. Lab Animal 34: 28–34.
Healey, M. C., R. Lake & S. G. Hinch, 2003. Energy expen-
ditures during reproduction by sockeye salmon
(Oncorhynchus nerka). Behaviour 140: 161–182.
Hinch, S. G. & J. Bratty, 2000. Effects of swim speed and
activity pattern on success of adult sockeye salmon
migration through an area of difficult passage. Transac-
tions of the American Fisheries Society 129: 598–606.
Hinch, S. G. & P. S. Rand, 1998. Swim speeds and energy use
of upriver-migrating sockeye salmon (Oncorhynchusnerka): role of local environment and fish characteristics.
Canadian Journal of Fisheries and Aquatic Sciences 55:
1821–1831.
Hinch, S. G., E. M. Standen, M. C. Healey & A. P. Farrell,
2002. Swimming patterns and behaviour of upriver-
migrating adult pink (Oncorhynchus gorbuscha) and
sockeye (O. nerka) salmon as assessed by EMG telemetry
in the Fraser River, British Columbia, Canada. Hydrobi-
ologia 483: 147–160.
Huntingford, F. A., C. Adams, V. A. Braithwaite, S. Kadri,
T. G. Pottinger, P. Sandoe & J. F. Turnbull, 2006. Current
issues in fish welfare. Journal of Fish Biology 68: 332–372.
Jepsen, N., A. Koed, E. B. Thorstad & E. Baras, 2002. Surgical
implantation of telemetry transmitters in fish: how much
have we learned? Hydrobiologia 483: 239–248.
Johansen, R., J. R. Needham, D. J. Colquhoun, T. T. Poppe &
A. J. Smith, 2006. Guidelines for health and welfare
monitoring of fish used in research. Laboratory Animals
40: 323–340.
Kaufman, R. C., A. G. Houck & J. J. Cech, 2006. Effects of
temperature and carbon dioxide on green sturgeon blood-
oxygen equilibria. Environmental Biology of Fishes 76:
119–127.
Kong, W., J. Huang, D. L. Rollins, R. E. Ideker & W. M.
Smith, 2007. A semi-implantable multichannel telemetry
system for continuous electrical, mechanical and hemo-
dynamical recordings in animal cardiac research.
Physiological Measurement 28: 249–257.
Lankford, S. E., T. E. Adams & J. J. Cech, 2003. Time of day
and water temperature modify the physiological stress
response in green sturgeon, Acipenser medirostris. Com-
parative Biochemistry and Physiology Part A-Molecular
& Integrative Physiology 135: 291–302.
Lankford, S. E., T. E. Adams, R. A. Miller & J. J. Cech, 2005.
The cost of chronic stress: impacts of a nonhabituating
stress response on metabolic variables and swimming
performance in sturgeon. Physiological and Biochemical
Zoology 78: 599–609.
Lefrancois, C., G. Claireaux & J. P. Lagardere, 1998. Heart
rate telemetry to study environmental influences on fish
metabolic expenditure. Hydrobiologia 372: 215–224.
Lucas, M. C., 1994. Heart-rate as an indicator of metabolic-rate
and activity in adult Atlantic Salmon, Salmo salar. Jour-
nal of Fish Biology 44: 889–903.
Lucas, M. C. & J. D. Armstrong, 1991. Estimation of meal
energy-intake from heart-rate records of pike, Esox-
Lucius L. Journal of Fish Biology 38: 317–319.
Lucas, M. C., I. G. Priede, J. D. Armstrong, A. N. Z. Gindy &
L. Devera, 1991. Direct measurements of metabolism,
activity and feeding-behavior of pike, Esox-Lucius L, in
the wild, by the use of heart-rate telemetry. Journal of Fish
Biology 39: 325–345.
Hydrobiologia
123

Marey, M., 1896. Phenomenon of flight in the animal kingdom.
Smithsonian Annual Report. Washington, DC: US Gov.
Print Office 226–285.
Maxime, V., G. Nonnotte, C. Peyraud, P. Williot &
J. P. Truchot, 1995. Circulatory and respiratory effects of
an hypoxic stress in the siberian sturgeon. Respiration
Physiology 100: 203–212.
McCue, M. D., 2006. Specific dynamic action: a century of
investigation. Comparative Biochemistry and Physiology
Part A-Molecular & Integrative Physiology 144: 381–394.
Perry, A. L., P. J. Low, J. R. Ellis & J. D. Reynolds, 2005.
Climate change and distribution shifts in marine fishes.
Science 308: 1912–1915.
Powers, D. A., 1989. Fish as model systems. Science 246:
352–358.
Priede, I. G. & P. Tytler, 1977. Heart rate as a measure of
metabolic rate in teleost fishes; Salmo gairdneri, Salmotrutta and Gadus morhua. Journal of Fish Biology 10:
231–242.
Rader, R. D., J. P. Meehan & J. K. C. Henricksen, 1975. An
implantable blood pressure and flow transmitter. IEEE
Transactions on Biomedical Engineering BME 20: 37–43.
Randall, D. J. (1982) The control of respiration and circulation
in fish during exercise and hypoxia. Journal of Experi-
mental Biology 100: 275–288.
Randall, D. & C. Brauner, 1991. Effects of environmental-
factors on exercise in fish. Journal of Experimental Biol-
ogy 160: 113–126.
Randall, D. J. & C. Daxboeck (1982) Cardiovascular changes
in the rainbow trout (Salmo gairdneri Richardson) during
exercise. Canadian Journal of Zoology 60: 1135–1140.
Roessig, J. M., C. M. Woodley, J. J. Cech & L. J. Hansen,
2004. Effects of global climate change on marine and
estuarine fishes and fisheries. Reviews in Fish Biology and
Fisheries 14: 251–275.
Schulz, U. H., 2003. Effects of surgically implanted dummy
transmitters on the South American catfish Jundia
(Rhamdia quelen). Brazilian Journal of Biology 63:
345–348.
Spelman, F. A., C. A. Astley, E. V. Golanov, J. J. Cupal,
A. R. Henkins, E. Fonzo, T. G. Susor, G. Mcmorrow,
D. M. Bowden & O. A. Smith, 1991. A system to acquire
and record physiological and behavioral data remotely
from nonhuman primates. IEEE Transactions on Bio-
medical Engineering BME 38–12: 1175–1185.
Thorarensen, H. & A. P. Farrell, 2006. Postprandial intestinal
blood flow, metabolic rates, and exercise in Chinook
salmon (Oncorhynchus tshawytscha). Physiological and
Biochemical Zoology 79: 688–694.
Thorarensen, H., P. E. Gallaugher & A. P. Farrell, 1996. The
limitations of heart rate as a predictor of metabolic rate in
fish. Journal of Fish Biology 49: 226–236.
Thorarensen, H., P. E. Gallaugher, A. K. Kiessling &
A. P. Farrell, 1993. Intestinal blood-flow in swimming
chinook Salmon Oncorhynchus tshawytscha and the
effects of Hematocrit on blood-flow distribution. Journal
of Experimental Biology 179: 115–129.
Van Eenennaam, J. P., M. A. H. Webb, X. Deng, S. I. Doroshov,
R. B. Mayfield, J. J. Cech, D. C. Hillemeier & T. E.
Willson, 2001. Artificial spawning and larval rearing of
klamath River green sturgeon. Transactions of the Amer-
ican Fisheries Society 130: 159–165.
Webber, D. M., R. G. Boutilier & S. R. Kerr, 1998. Cardiac
output as a predictor of metabolic rate in cod Gadusmorhua. Journal of Experimental Biology 201: 2779–
2789.
Winters, S. R., 1921. Diagnostics by wireless. Scientific
American 124: 465.
Yonezawa, Y., T. Nakayama, I. Ninomiya & W. M. Caldwell,
1992. Radio telemetry directional ultrasonic blood flow-
meter for use with unrestrained animals. Medical &
Biological Engineering & Computing 30: 659–665.
Yonezawa, Y., J. C. Schadt, W. M. Caldwell & A. W. Hahn,
1989. A miniaturized ultrasonic flowmeter and telemetry
transmitter for chronic animal blood flow measurements.
Biomedical Sciences Instrumentation 25: 107–111.
Hydrobiologia
123
Copyright © 2022 FDOKUMEN




















