
Metabolic Integra1on and Organ Specializa1on - Di.Univr.It
66
Metabolic Integra/on and Organ Specializa/on What principles underlie the integra1on of catabolism and energy produc1on with anabolism and energy consump1on? How is metabolism integrated in complex organisms with mul1ple organ systems?
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Metabolic Integra1on and Organ Specializa1on - Di.Univr.It

MetabolicIntegra/onandOrganSpecializa/on
Whatprinciplesunderlietheintegra1onofcatabolismandenergyproduc1onwithanabolismandenergyconsump1on?Howismetabolismintegratedincomplexorganismswithmul1pleorgansystems?

TheWashington,D.C.,Metromap.Thecoordinatedflowofpassengersalongdifferenttransitlinesisanaptmetaphorformetabolicregula1on.

Severalpathwaysarecatabolicandservetogeneratechemicalenergyusefultothecell;othersareanabolicandusethisenergytodrivethesynthesisofessen/albiomolecules.Despitetheiropposingpurposes,thesereac/onstypicallyoccuratthesame0measnutrientmoleculesarebrokendowntoprovidethebuildingblocksandenergyforongoingbiosynthesis.Cellsmaintainadynamicsteadystatethroughprocessesthatinvolveconsiderablemetabolicflux.Themetabolismthattakesplaceinjustasinglecellissocomplexthatitdefiesdetailedquan/ta/vedescrip/on.However,anapprecia/onofoverallrela/onshipscanbeachievedbysteppingbackandconsideringintermediarymetabolismatasystemsleveloforganiza/on.


HowIsMetabolismIntegratedinaMul1cellularOrganism?
Incomplexmul/cellularorganisms,organsystemscarryoutspecificphysiologicalfunc/ons.Eachorganexpressesarepertoireofmetabolicpathwaysthatisconsistentwithitsphysiologicalpurpose.Suchspecializa1ondependsoncoordina1onofmetabolicresponsibili/esamongorgans.Essen/allyallcellsinanimalshavethesetofenzymescommontothecentralpathwaysofintermediarymetabolism,especiallytheenzymesinvolvedintheforma/onofATPandthesynthesisofglycogenandlipidreserves.Nevertheless,organsdifferinthemetabolicfuelstheypreferassubstratesforenergyproduc/on.ImportantdifferencesalsooccurinthewaysATPisusedtofulfilltheorgans’specializedmetabolicfunc/ons.

Wewillconsiderthemetabolicinterac/onsamongthemajororgansystemsfoundinhumans:ü brain,ü skeletalmuscle,ü heart,ü adipose1ssue,ü andliverThemajorfueldepotsinanimalsareglycogeninliverandmuscle;triacylglycerols(fats)storedinadipose/ssue;andprotein,mostofwhichisinskeletalmuscle.Theorderofpreferencefortheuseofthesefuelsis:Glycogen>triacylglycerol>protein.Nevertheless,the/ssuesofthebodyworktogethertomaintainenergyhomeostasis(calorichomeostasis),definedasaconstantavailabilityoffuelsintheblood.

Metabolicrela1onshipsamongthemajorhumanorgans.

TheMajorOrganSystemsHaveSpecializedMetabolicRoles

BrainTworemarkablemetabolicfeatures:First,ithasaveryhighrespiratorymetabolism.Inres/ngadulthumans,20%oftheoxygenconsumedisusedbythebrain,eventhoughitcons/tutesonly2%orsoofbodymass.(thislevelofO2consump/onisindependentofmentalac/vity,con/nuingevenduringsleep.Second,thebrainhasnofuelreserves—noglycogen,noexpendableprotein,orfat.Normally,thebrainusesonlyglucoseasafuelandistotallydependentonthebloodforacon/nuousincomingsupply.Interrup/onofglucosesupplyforevenbriefperiodsof/me(asinastroke)canleadtoirreversiblelossesinbrainfunc/on.

ThebrainusesglucosetocarryoutATPsynthesisviacellularrespira/on.HighratesofATPproduc/onarenecessarytopowertheplasmamembraneNa,K-ATPasesothatthemembranepoten/alessen/alfortransmissionofnerveimpulsesismaintained.Duringprolongedfas/ngorstarva/on,thebody’sglycogenreservesaredepleted.Undersuchcondi/ons,thebrainadaptstouseβ-hydroxybutyrate,conver/ngittoacetyl-CoAforenergyproduc/onviathecitricacidcycle.β-Hydroxybutyrateisformedfromfa`yacidsintheliver.Theadapta/onofthebraintouseβ-hydroxybutyratefromfatsparesproteinfromdegrada/onun/llipidreservesareexhausted.


Thebrain’sotherpoten/alsourceoffuelduringstarva/onisglucoseobtainedfromgluconeogenesisintheliverusingthecarbonskeletonsofaminoacidsderivedfrommuscleproteinbreakdown.

13
Metabolic Profile of Brain
Glucose is fuel for human brain -> consumes 120g/day -> 60-70 % of utilization of glucose in starvation -> ketone bodies can replace glucose

Muscle
Skeletalmuscleisresponsibleforabout30%oftheO2consumedbythehumanbodyatrest.Duringperiodsofmaximalexer/on,skeletalmusclecanaccountformorethan90%ofthetotalmetabolism.Metabolismisprimarilydedicatedtotheproduc/onofATPasthesourceofenergyforcontrac1onandrelaxa1on.

Musclecontrac/onoccurswhenamotornerveimpulsecausesCa2releasefromspecializedendomembranecompartments(thetransversetubulesandsarcoplasmicre1culum).Ca2floodsthesarcoplasm(thetermdeno/ngthecytosoliccompartmentofmusclecells),whereitbindstotroponinC,aregulatoryprotein,ini/a/ngaseriesofeventsthatculminateintheslidingofmyosinthickfilamentsalongac/nthinfilaments.ThismechanicalmovementisdrivenbyenergyreleaseduponhydrolysisofATP.

Thenetresultisthatthemuscleshortens.Relaxa/onoccurswhentheCa2ionsarepumpedbackintothesarcoplasmicre/culumbytheac/onofaCa2-transpor/ngmembraneATPase.TwoCa2ionsaretranslocatedperATPhydrolyzed.TheamountofATPusedduringrelaxa1onisalmostasmuchasthatconsumedduringcontrac1on.

Thestructureofaskeletalmusclecell,showingthemannerinwhichtransversetubulesenablethesarcolemmalmembranetoextendintotheinteriorofthefiber.T-tubulesandsarcoplasmicre/culum(SR)membranesarejuxtaposedatstructurestermedtriadjunc/ons(inset).


Becausemusclecontrac/onisaprocessthatoccursupondemand,musclemetabolismisdesignedforademandresponse.Muscleatrestusesfa`yacids,glucose,orketonebodiesasfuelandproducesATPviaoxida/vephosphoryla/on.Res/ngmusclealsocontainsabout2%glycogenandabout0.08%phosphocrea1nebyweight.WhenATPisusedduringmusclecontrac/on,theADPformedcanbereconvertedtoATPbycrea1nekinasefromphosphocrea/ne:

Musclephosphocrea/necangenerateenoughATPtopowerabout4secofexer/on.Duringstrenuousexer/on,suchasa100-metersprint,oncethephosphocrea/neisdepleted,musclereliessolelyonitsglycogenreserves,makingtheATPforcontrac/onviaglycolysis.Incontrastwiththecitricacidcycleandoxida/vephosphoryla/onpathways,glycolysisiscapableofexplosiveburstsofac1vity,andthefluxofglucose-6-phosphatethroughthispathwaycanincrease2000-foldalmostinstantaneously.Thetriggersforthisac/va/onareCa2andthe“fightorflight”hormoneepinefrine

Musclefa1gueistheinabilityofamuscletomaintainpoweroutput.Duringmaximumexer/on,theonsetoffa/guetakesonly20secondsorso.Fa/gueisnottheresultofexhaus/onoftheglycogenreserves,norisitaconsequenceoflactateaccumula/oninthemuscle.ItiscausedbyadeclineinintramuscularpHasprotonsaregeneratedduringglycolysis.(Theoverallconversionofglucoseto2lactateinglycolysisisaccompaniedbythereleaseof2H+)ThepHmayfallaslowas6.4.ItislikelythatthedeclineinPFKac1vityatlowpHleadstoaloweredfluxofhexosethroughglycolysisandinadequateATPlevels,causingafeelingoffa1gue.

Duringfas/ngorexcessiveac/vity,skeletalmuscleproteinisdegradedtoaa,sothattheircarbonskeletonscanbeusedasfuel.Manyoftheskeletonsareconvertedtopyr,whichcanbetransaminatedintoAlaforexportviathecircula/on.Alaiscarriedtotheliver,whichdeaminatesitbackintopyruvatesothatitcanserveasasubstrateforgluconeogenesis.Althoughmuscleproteincanbemobilizedasanenergysource,itisnotefficientforanorganismtoconsumeitsmuscleandloweritsoverallfitnessforsurvival.Muscleproteinrepresentsafueloflastresort.

23
Metabolic Profile of Muscles
Major fuels are glucose, fatty acids, and ketone bodies -> has a large storage of glycogen -> about ¾ of all glycogen stored in muscles -> glucose is preferred fuel for burst of activity -> production of lactate (anaerobe) -> fatty acid major fuel in resting muscles and in heart muscle (aerobe)

Heart
Incontrastwiththeintermi`entworkofskeletalmuscle,theac/vityofheartmuscleisconstantandrhythmic.Therangeofac/vityinheartismuchlessthanthatinmuscle.Consequently,theheartfunc/onsasacompletelyaerobicorganand,assuch,isveryrichinmitochondria(halfthecytoplasmicvolumeofheartmusclecellsisoccupiedbymitochondria).Undernormalworkingcondi/ons,theheartprefersfaYyacidsasfuel,oxidizingacetyl-CoAviathecitricacidcycleandproducingATPforcontrac/onviaoxida/vephosphoryla/on.Heart/ssuehasminimalenergyreserves:asmallamountofphosphocrea/neandlimitedquan//esofglycogen.Asaresult,theheartmustbecon1nuallynourishedwithO2andfreefa`yacids,glucose,orketonebodiesasfuel.

AdiposeTissueAdipose/ssueisanamorphous/ssuethatiswidelydistributedinthebody—aroundbloodvessels,intheabdominalcavityandmammaryglands,andmostprevalently,asdepositsundertheskin.
Longconsideredonlyastoragedepotforfat,adipose/ssueisnowappreciatedasanendocrineorganresponsibleforsecre/onofavarietyofhormonesthatgovernea1ngbehaviorandcalorichomeostasis.

HumanFatDistribu1on

Itconsistsprincipallyofcellsknownasadipocytesthatnolongerreplicate,butcanincreaseinnumberasadipocyteprecursorcellsdivide,andobeseindividualstendtohavemoreofthem.65%oftheweightofadipose/ssueistriacylglycerolthatisstoredinadipocytes,essen/allyasoildroplets.Theaverage70-kgmanhasenoughcaloricreservestoredasfattosustaina6000kJ/dayrateofenergyproduc1onfor3months,whichisadequateforsurvival

Despitetheirroleasenergystoragedepots,adipocyteshaveahighrateofmetabolicac/vity,synthesizingandbreakingdowntriacylglycerol.Adipocytesac/velycarryoutcellularrespira/on,transformingglucosetoenergyviaglycolysis,thecitricacidcycle,andoxida/vephosphoryla/on.Ifglucoselevelsinthedietarehigh,glucoseisconvertedtoacetyl-CoAforfa`yacidsynthesis.However,undermostcondi/ons,freefa`yacidsfortriacylglycerolsynthesisareobtainedfromtheliver.Becauseadipocyteslackglycerolkinase,theycannotrecycleglyceroloftriacylglycerolbutdependonglycoly/cconversionofglucosetodihydroxyacetone-3-phosphate(DHAP)andthereduc/onofDHAPtoglycerol-3-phosphatefortriacylglycerolbiosynthesis.AdipocytesalsorequireglucosetofeedthepentosephosphatepathwayforNADPHproduc1on.

“BrownFat”Aspecializedtypeofadipose/ssuethatisaboundantinnewbornsandhiberna/nganimals.Theabundanceofmitochondria,whicharerichincytochromes,isresponsibleforthebrowncolorofthisfat.thesemitochondriaareveryac/veinelectrontransport–drivenprotontransloca/on,butthesepar/cularmitochondriacontainintheirinnermembranesaprotein,thermogenin(uncouplingprotein1),thatcreatesapassiveprotonchannel,permilngtheH+ionstoreenterthemitochondrialmatrixwithoutgenera/ngATP

30
Metabolic Profile of Adipose tissue
Triacylglycerols are stored in tissue -> enormous reservoir of metabolic fuel -> needs glucose to synthesis TAG; -> glucose level determines if fatty acids are released into blood
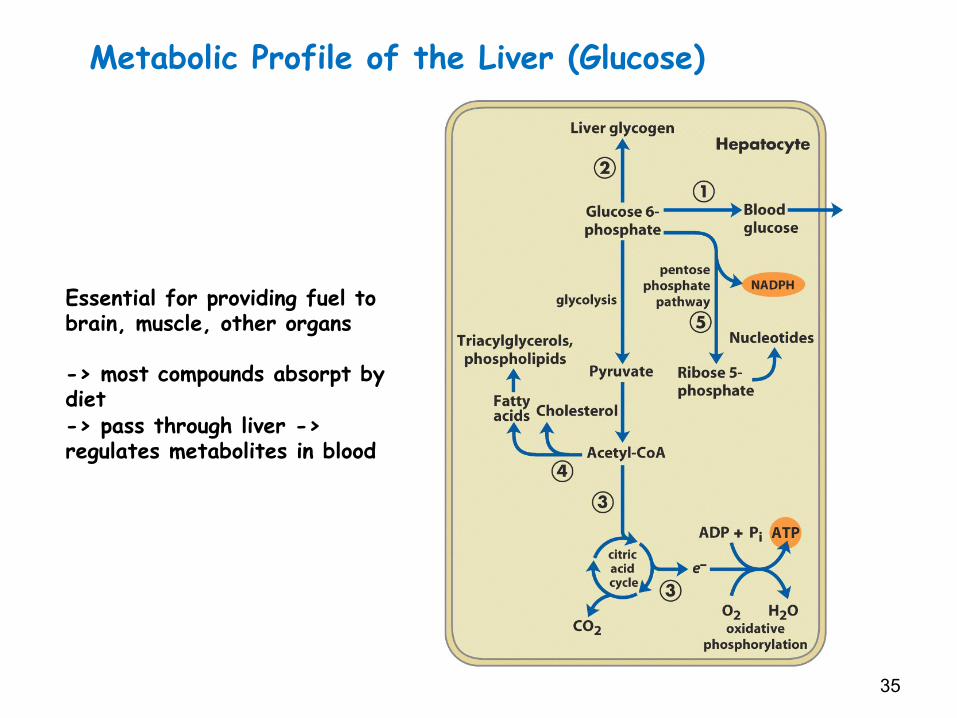
LiverLiverservesasmajormetabolicprocessingcenterinvertebrates.Exceptfordietarytriacylglycerols,whicharemetabolizedprincipallybyadipose/ssue,mostoftheincomingnutrientsthatpassthroughtheintes/naltractareroutedviatheportalveintotheliverforprocessinganddistribu/on.Muchoftheliver’sac/vitycentersaroundconversionsinvolvingglucose-6-phosphate

Glucose-6-Pcanbeconvertedtoglycogen,releasedasbloodglucose,usedtogenerateNADPHandpentosesviathepentosephosphatecycle,orcatabolizedtoacetyl-CoAforfa`yacidsynthesisorforenergyproduc/onviaoxida/vephosphoryla/on.Mostoftheliverglucose-6-Parisesfromdietarycarbohydrate,fromdegrada/onofglycogenreserves,orfrommusclelactatethatentersthegluconeogenicpathway

Theliverplaysanimportantregulatoryroleinmetabolismbybufferingthelevelofbloodglucose.Liverhastwoenzymesforglucosephosphoryla/on:hexokinaseandglucokinaseTheliverisamajorcenterforfaYyacidturnover.Demandformetabolicenergyhighàtriacylglycerolsarebrokendownandfa`yacidsaredegradedinthelivertoacetyl-CoAtoformketonebodies,whichareexportedtotheheart,brain,andother/ssues.Energydemandslowàfa`yacidsareincorporatedintotriacylglycerolsandcarriedtoadipose/ssuefordeposi/onasfat.Cholesterolisalsosynthesizedintheliverfromtwo-carbonunitsderivedfromacetyl-CoA.

Livercanuseaminoacidsasmetabolicfuels.Aminoacidsarefirstconvertedtotheircorrespondingα-ketoacidsbyaminotransferases.Theaminogroupisexcretedanerincorpora/onintoureaintheureacycle.Thecarbonskeletonsofglucogenicaminoacidscanbeusedforglucosesynthesis,whereasthoseofketogenicaminoacidsappearinketonebodies.Liveristheprincipaldetoxifica1onorgan.Theendoplasmicre/culumoflivercellsisrichinenzymesthatconvertbiologicallyac/vesubstancessuchashormones,poisons,anddrugsintolessharmfulby-products.Liverdiseaseleadstoseriousmetabolicderangements,par/cularlyinaminoacidmetabolism.Incirrhosis,theliverbecomesdefec/veinconver/ngNH4toureaforexcre/on,andbloodlevelsofNH4rise.Ammoniaistoxictothecentralnervoussystem,andcomaensues.
35
Metabolic Profile of the Liver (Glucose)
Essential for providing fuel to brain, muscle, other organs -> most compounds absorpt by diet -> pass through liver -> regulates metabolites in blood

36
Metabolic Activities of the Liver (Amino Acids)
α-Ketoacids (derived from amino acid degradation) -> liver’s own fuel

37
Metabolic Activities of the Liver (Fatty Acids)
cannot use acetoacetate as fuel -> almost no transferase to generate acetyl-CoA

38
Food Intake, Starvation, and Obesity
Normal Starved-Fed Cycle: 1. Postabsorptive state -> after a
meal
2. Early fasting state -> during the night
3. Refed state -> after breakfast
-> Major goal is to maintain blood-glucose level

39
Blood-Glucose

40
1. Postabsorptive state
Glucose + Amino acids -> transport from intestine to blood
Dietary lipids transported -> lymphatic system -> blood
Glucose stimulates -> secretion of insulin
Insulin:
-> signals fed state
-> stimulates storage of fuels and synthesis of proteins
-> high level -> glucose enters muscle + adipose tissue (synthesis of TAG)
-> stimulates glycogen synthesis in muscle + liver
-> suppresses gluconeogenesis by the liver
-> accelerates glycolysis in liver -> increases synthesis of fatty acids
-> accelerates uptake of blood glucose into liver -> glucose 6-phosphate more rapidly formed than level of blood glucose rises -> built up of glycogen stores

41
Insulin Secretion –Stimulated by Glucose Uptake

42
Postabsorptive State -> after a Meal

43
2. Early Fasting State
Blood-glucose level drops after several hours after the meal -> decrease in insulin secretion -> rise in glucagon secretion
Low blood-glucose level -> stimulates glucagon secretion of α-cells of the pancreas
Glucagon:
-> signals starved state
-> mobilizes glycogen stores (break down)
-> inhibits glycogen synthesis
-> main target organ is liver
-> inhibits fatty acid synthesis
-> stimulates gluconeogenesis in liver
-> large amount of glucose in liver released to blood stream -> maintain blood-glucose level
Muscle + Liver use fatty acids as fuel when blood-glucose level drops

44
Early Fasting State -> During the Night

45
3. Refed State Fat is processed in same way as normal fed state
First -> Liver does not absorb glucose from blood (diet)
Liver still synthesizes glucose to refill liver’s glycogen stores
When liver has refilled glycogen stores + blood-glucose level still rises -> liver synthesizes fatty acids from excess glucose

46
Prolonged Starvation Well-fed 70 kg human -> fuel reserves about 161,000 kcal
-> energy needed for a 24 h period -> 1600 kcal - 6000 kcal
-> sufficient reserves for starvation up to 1 – 3 months
-> however glucose reserves are exhausted in 1 day
Even under starvation -> blood-glucose level must be above 40 mg/100 ml

47
First priority -> provide sufficient glucose to brain and other tissues that are dependent on it
Second priority -> preserve protein -> shift from utilization of glucose to utilization of fatty acids + ketone bodies
-> mobilization of TAG in adipose tissues + gluconeogenesis by liver -> muscle shift from glucose to fatty acids as fuel
After 3 days of starvation -> liver forms large amounts of ketone bodies (shortage of oxaloacetate) -> released into blood -> brain and heart start to use ketone bodies as fuel
After several weeks of starvation -> ketone bodies major fuel of brain
After depletion of TAG stores -> proteins degradation accelerates -> death due to loss of heart, liver, and kidney function
Prolonged Starvation

The fasting state computing
Presentation copyright © 2002 David A Bender and some images copyright © 2002 Taylor & Francis Ltd
glycogen
glucose
`
triacyglycerol
hormone-sensi1velipase
glycerol
faYyacidsfaYyacids
protein
aminoacids
aminoacidsglucose
ketonebodies
The fasting state
F

Starvation
Starvation in the 3rdworld
Starvation in the rich-world

50
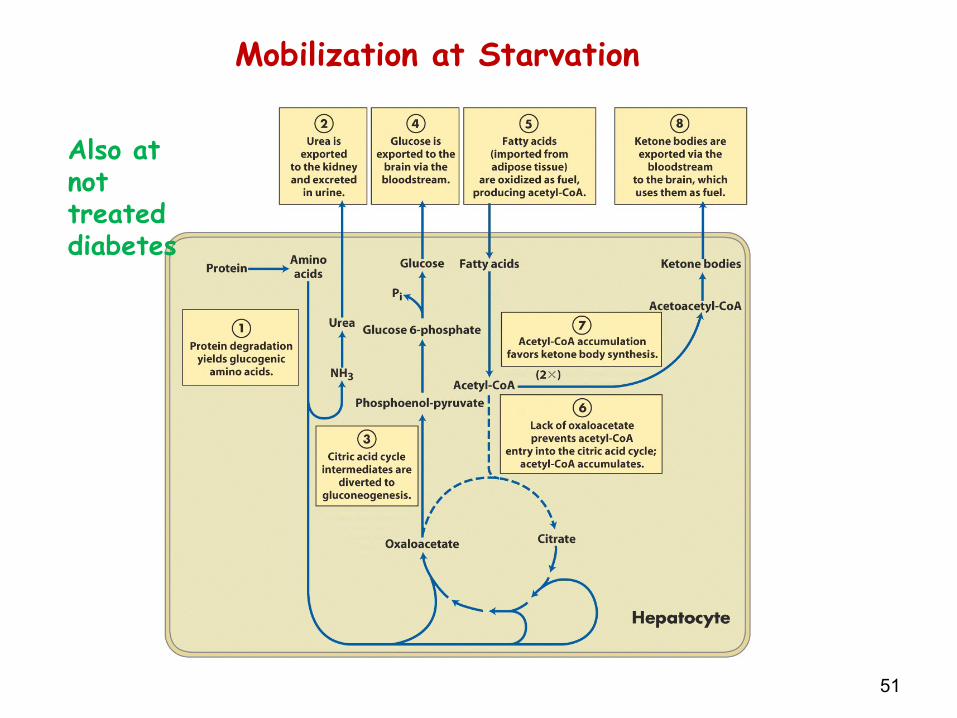
51
Mobilization at Starvation
Also at not treated diabetes

52
Diabetes Mellitus – Insulin Insufficiency Characterized by: -> high blood-glucose level -> Glucose overproduced by liver
-> glucose underutilized by other organs
-> shift in fuel usage from carbohydrates to fats -> keton bodies (shortage of oxaloacetate)
-> high level of keton bodies -> kidney cannot balance pH any more -> lowered pH in blood and dehydration -> coma
Type I diabetes: insulin-dependent diabetes (requires insulin to live)
caused by autoimmune destruction of β-cells
begins before age 20
-> insulin absent -> glucagon present
-> person in biochemical starvation mode + high blood-glucose level
-> entry of glucose into cells is blocked
-> glucose excreted into urine -> also water excreted -> feel hungry + thirsty
Type II diabetes: insulin-independent diabetes
have a normal-high level of insulin in blood -> unresponsive to hormone
develops in middle-aged, obese people

53
Obesity
In the U. S. -> about 70% of adults are suffering from obesity (2009)
Risk factor for: Diabetes + Cardiovascular diseases
Cause of Obesity -> more food consumed than needed -> storage of energy as fat
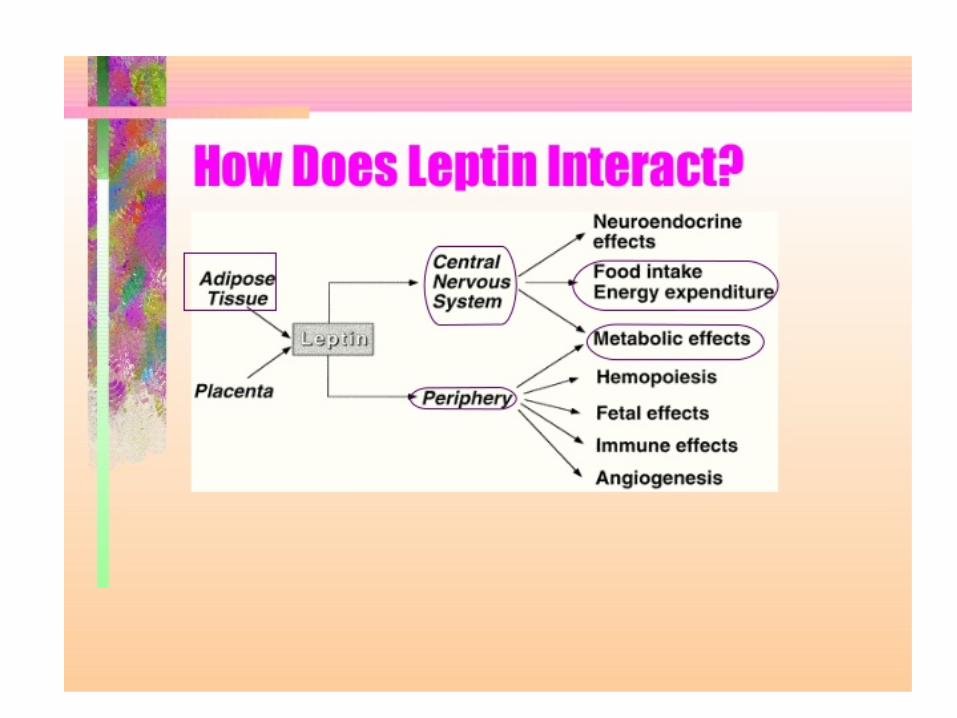
Two important signals for “caloric homeostasis” and “appetite” control -> insulin + leptin
Mouse lacking leptin or Leptin receptor

54

55
The Role of Leptin and Insulin on Weight Control
Leptin (dal greco leptos, snello) is a hormone that is produced in direct proportion to fat mass (adipocytes)
Leptina regola il senso di SAZIETA’ Grelina stimola
il senso di appetito

56
High Levels of Leptin and Insulin are a Signal for “caloric homeostasis”

Leptin: A Key Player In Weight Regulation

What is Leptin? • A peptide hormone which is coded for by the obese gene
(ob) • Influences the quantity of food consumed relative to the
amount of energy expended – When leptin levels are high, appetite is reduced and
energy expenditure is increased • Leptin has been found in gastric epithelium, placenta
and adipose tissue – Most abundant in white adipose tissue

La Leptina è un ormone proteico che regola il peso corporeo, il metabolismo e le funzioni riproduttive. Essa è codificata dal gene dell’obesità (ob)
La leptina è espressa prevalentemente dagli adipociti. Piccole quantità vengono prodotte nello stomaco e nella placenta. I recettori specifici per la leptina sono espressi nell’ipotalamo, nei linfociti T e nelle cellule endoteliali

La leptina arriva nel Sistema Nervoso Centrale (SNC) attraverso la barriera ematoencefalica mediante un meccanismo di trasporto mediato da specifici recettori.
Questo segnale di natura ormonale ha lo scopo di informare il SNC sullo stato di riserve energetiche dell’individuo.
Agisce regolando l’apporto alimentare attraverso l’inibizione della sintesi e del rilascio del neuropeptide Y (NPY), sostanza stimolante l’appetito.

61
The Role of Leptin and Insulin on Weight Control
Leptin (dal greco leptos, snello) is a hormone that is produced in direct proportion to fat mass (adipocytes)
Leptina regola il senso di SAZIETA’ Grelina stimola
il senso di appetito

Regulating Food Intake and Energy Expenditure
• Leptin binds to its receptor which is expressed primarily in the brains hypothalamus region
• In turn the hypothalamus modulates food intake and energy expenditure
• When low leptin levels are detected, the body is warned of limited energy supplies
• If high leptin levels are detected, the hypothalamus senses the body as being overweight – This then trigger the body to eat less and expend more
energy • When energy intake and output are equal, leptin reflects the
amount of triglyceride stored in the bodies adipose tissue

Metabolic Effects of Leptin
• Decreases intracellular lipid concentration through reduction of fatty acid and triglyceride synthesis and a concomitant increase in lipid oxidation
• It has been postulated that leptin inhibits acetyl-CoA carboxylase , enzyme involved in the committed step of fatty acid synthesis
• This inhibition leads to decrease in malonyl-CoA levels – Together the inhibition of acetyl-CoA to malonyl-CoA encourages the
mobilization of fatty acids from storage sites and simultaneously discourages synthesis

Experimentation on Humans • Few experiments done at this point • Leptin is said to circulate freely or attached to a binding protein
– It has been found that obese individuals have more circulating bound leptin than lean individuals
• The greater the initial level, the more it declines with dieting • Levels tend to vary greatly from person to person • Typically females have more leptin than males
– Adipose tissue accounts for 20-25% of weight in females and only 15-20% in males
• In general the greater the body mass and percent body fat, the higher the levels – People suffering from obesity have extremely high levels

Possible Reasons For Increased Leptin In Obese Individuals
• Differences in the fat production rate of leptin – Some obese people may make leptin at greater rate to compensate for
faulty signaling process or action • Resistance to leptin at its site of action
– If resistance is partial, not complete, more leptin may be required for action
• A combination of both could influence eating behaviors and energy use to cause obesity
• All these possibilities indicate that obese individuals are in a state of percieved starvation




















