
Melittin disruption of raft and non-raft-forming biomimetic membranes: A study by quartz crystal...
7
Please cite this article in press as: P. Losada-Pérez, et al., Melittin disruption of raft and non-raft-forming biomimetic membranes: A study by quartz crystal microbalance with dissipation monitoring, Colloids Surf. B: Biointerfaces (2014), http://dx.doi.org/10.1016/j.colsurfb.2014.10.048 ARTICLE IN PRESS G Model COLSUB-6712; No. of Pages 7 Colloids and Surfaces B: Biointerfaces xxx (2014) xxx–xxx Contents lists available at ScienceDirect Colloids and Surfaces B: Biointerfaces j o ur nal ho me pa ge: www.elsevier.com/locate/colsurfb Melittin disruption of raft and non-raft-forming biomimetic membranes: A study by quartz crystal microbalance with dissipation monitoring P. Losada-Pérez a,b,∗ , M. Khorshid a , C. Hermans a , T. Robijns a , M. Peeters a , K.L. Jiménez-Monroy a , L.T.N. Truong c , P. Wagner a,b a Institute for Materials Research IMO, Hasselt University, Wetenschapspark 1, B-3590 Diepenbeek, Belgium b IMEC vzw, Division IMOMEC, Wetenschapspark 1, B-3590 Diepenbeek, Belgium c School of Engineering Physics, Hanoi University of Science and Technology 1, Dai Co Viet, Hanoi, Vietnam a r t i c l e i n f o Article history: Received 26 July 2014 Received in revised form 20 October 2014 Accepted 24 October 2014 Available online xxx Keywords: Melittin Lipid rafts Cholesterol Quartz crystal microbalance with dissipation a b s t r a c t In this work we examine the role of lateral phase separation in cholesterol-containing biomimetic mem- branes on the disrupting action of melittin using a label-free surface-sensitive technique, quartz crystal microbalance with dissipation monitoring (QCM-D). Melittin disruption mechanisms depend strongly on the geometry of the lipid layer; however, despite the interplay between layer geometry/thickness and melittin activity, results indicate that the presence of lipid heterogeneity and lateral phase separation greatly influences the disrupting efficiency of melittin. In homogeneous non-raft forming membranes with high cholesterol content, melittin spontaneous activity is strongly delayed compared to heteroge- neous raft-forming systems with the same amount of cholesterol. These results confirm the importance of lateral phase separation as a determinant factor in peptide activity. The information provided can be used for the design of more efficient antimicrobial peptides and the possibility of using a label-free approach for tailored-membranes and interactions with other types of peptides, such as amyloid peptides. © 2014 Elsevier B.V. All rights reserved. 1. Introduction Antimicrobial peptides (AMPs) are naturally occurring amphiphilic molecules that display cell lytic properties. Due to their high selectivity toward different types of cells, i.e., Gram-negative and Gram-positive bacteria, the knowledge of their biological activity is of major importance for potential applications as antiseptic agents and as alternatives for conven- tional antibiotics [1–3]. The general picture of the membrane perturbing effect of AMPs is that it induces membrane leakage via mechanisms including the formation of well-defined pores or the detergent mechanism [4–9]. One of the major differences between bacterial and eukaryotic cell membranes is the absence of cholesterol in the former and the presence in the latter. Choles- terol is known to increase membrane cohesion and mechanical stiffness in most homogeneous lipid systems [10,11] for instance, in phosphatidylcholine-based lipid model membranes, it has a ∗ Corresponding author at: Institute for Materials Research IMO, Hasselt Univer- sity, Wetenschapspark 1, B-3590 Diepenbeek, Belgium. Tel.: +32 496658831. E-mail address: [email protected] (P. Losada-Pérez). pronounced reducing effect on AMP-induced leakage [12]. The apparently better resistance of cholesterol containing mem- branes is a consequence of cholesterol condensing effect, which precludes deep peptide penetration [12,13]. The presence of membrane-stabilizing cholesterol has been shown to protect human erythrocytes from mangainin 2 and pardaxin [14–17]. Both bacterial and eukaryotic cell membranes are complex mix- tures of lipids whose physical properties vary depending on the composition of the membrane, in particular, on the phase of the constituent lipids. Liquid disordered–liquid ordered (L d –L o ) lateral phase separation in eukaryotic membranes, i.e., the formation of the so-called lipid rafts, has shown to play an important role in the localization or transport of different proteins [18]. Despite the ability of cholesterol to suppress the action of an AMP on a homogeneous lipid bilayer, recent works using fluorescence on release of vesicle entrapped dyes have shown that cholesterol does not have the same effect in heterogeneous lipid systems [19–22]. These studies revealed that membrane permeabilization by AMPs like -lysin occurs exclusively in the L d phase with L d –L o phase segregation and that the localization of the peptide into the L d phase results in greater membrane disruption than would be expected in the absence of lateral phase separation. Those http://dx.doi.org/10.1016/j.colsurfb.2014.10.048 0927-7765/© 2014 Elsevier B.V. All rights reserved.
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Melittin disruption of raft and non-raft-forming biomimetic membranes: A study by quartz crystal...

C
Mmm
PKa
b
c
a
ARRAA
KMLCQd
1
atGtatpvobotsi
s
h0
ARTICLE IN PRESSG ModelOLSUB-6712; No. of Pages 7
Colloids and Surfaces B: Biointerfaces xxx (2014) xxx–xxx
Contents lists available at ScienceDirect
Colloids and Surfaces B: Biointerfaces
j o ur nal ho me pa ge: www.elsev ier .com/ locate /co lsur fb
elittin disruption of raft and non-raft-forming biomimeticembranes: A study by quartz crystal microbalance with dissipationonitoring
. Losada-Péreza,b,∗, M. Khorshida, C. Hermansa, T. Robijnsa, M. Peetersa,.L. Jiménez-Monroya, L.T.N. Truongc, P. Wagnera,b
Institute for Materials Research IMO, Hasselt University, Wetenschapspark 1, B-3590 Diepenbeek, BelgiumIMEC vzw, Division IMOMEC, Wetenschapspark 1, B-3590 Diepenbeek, BelgiumSchool of Engineering Physics, Hanoi University of Science and Technology 1, Dai Co Viet, Hanoi, Vietnam
r t i c l e i n f o
rticle history:eceived 26 July 2014eceived in revised form 20 October 2014ccepted 24 October 2014vailable online xxx
eywords:
a b s t r a c t
In this work we examine the role of lateral phase separation in cholesterol-containing biomimetic mem-branes on the disrupting action of melittin using a label-free surface-sensitive technique, quartz crystalmicrobalance with dissipation monitoring (QCM-D). Melittin disruption mechanisms depend stronglyon the geometry of the lipid layer; however, despite the interplay between layer geometry/thickness andmelittin activity, results indicate that the presence of lipid heterogeneity and lateral phase separationgreatly influences the disrupting efficiency of melittin. In homogeneous non-raft forming membranes
elittinipid raftsholesteroluartz crystal microbalance withissipation
with high cholesterol content, melittin spontaneous activity is strongly delayed compared to heteroge-neous raft-forming systems with the same amount of cholesterol. These results confirm the importance oflateral phase separation as a determinant factor in peptide activity. The information provided can be usedfor the design of more efficient antimicrobial peptides and the possibility of using a label-free approachfor tailored-membranes and interactions with other types of peptides, such as amyloid peptides.
© 2014 Elsevier B.V. All rights reserved.
. Introduction
Antimicrobial peptides (AMPs) are naturally occurringmphiphilic molecules that display cell lytic properties. Dueo their high selectivity toward different types of cells, i.e.,ram-negative and Gram-positive bacteria, the knowledge of
heir biological activity is of major importance for potentialpplications as antiseptic agents and as alternatives for conven-ional antibiotics [1–3]. The general picture of the membraneerturbing effect of AMPs is that it induces membrane leakageia mechanisms including the formation of well-defined poresr the detergent mechanism [4–9]. One of the major differencesetween bacterial and eukaryotic cell membranes is the absencef cholesterol in the former and the presence in the latter. Choles-
Please cite this article in press as: P. Losada-Pérez, et al., Mmembranes: A study by quartz crystal microbalance with dishttp://dx.doi.org/10.1016/j.colsurfb.2014.10.048
erol is known to increase membrane cohesion and mechanicaltiffness in most homogeneous lipid systems [10,11] for instance,n phosphatidylcholine-based lipid model membranes, it has a
∗ Corresponding author at: Institute for Materials Research IMO, Hasselt Univer-ity, Wetenschapspark 1, B-3590 Diepenbeek, Belgium. Tel.: +32 496658831.
E-mail address: [email protected] (P. Losada-Pérez).
ttp://dx.doi.org/10.1016/j.colsurfb.2014.10.048927-7765/© 2014 Elsevier B.V. All rights reserved.
pronounced reducing effect on AMP-induced leakage [12]. Theapparently better resistance of cholesterol containing mem-branes is a consequence of cholesterol condensing effect, whichprecludes deep peptide penetration [12,13]. The presence ofmembrane-stabilizing cholesterol has been shown to protecthuman erythrocytes from mangainin 2 and pardaxin [14–17].Both bacterial and eukaryotic cell membranes are complex mix-tures of lipids whose physical properties vary depending on thecomposition of the membrane, in particular, on the phase of theconstituent lipids. Liquid disordered–liquid ordered (Ld–Lo) lateralphase separation in eukaryotic membranes, i.e., the formation ofthe so-called lipid rafts, has shown to play an important role inthe localization or transport of different proteins [18]. Despitethe ability of cholesterol to suppress the action of an AMP on ahomogeneous lipid bilayer, recent works using fluorescence onrelease of vesicle entrapped dyes have shown that cholesteroldoes not have the same effect in heterogeneous lipid systems[19–22]. These studies revealed that membrane permeabilization
elittin disruption of raft and non-raft-forming biomimeticsipation monitoring, Colloids Surf. B: Biointerfaces (2014),
by AMPs like �-lysin occurs exclusively in the Ld phase with Ld–Lo
phase segregation and that the localization of the peptide intothe Ld phase results in greater membrane disruption than wouldbe expected in the absence of lateral phase separation. Those

ING ModelC
2 rface
rea
dotapsnQla
ttTlibma[toca(a
2
2
c7ahdutw(o
2
tsorrbshfwa(add
ARTICLEOLSUB-6712; No. of Pages 7
P. Losada-Pérez et al. / Colloids and Su
esults suggested that the phase separation naturally occurring inukaryotic membranes is likely to nullify the effect of cholesterolgainst membrane disruption by AMPs.
In phase-related lipid studies the use of label-free techniques isesirable in order to enable their characterization without the needf fluorescent labels, which may modify the structural properties. Inhis respect, quartz crystal microbalance with dissipation (QCM-D),
label-free surface sensitive technique, enables to explore sup-orted model membrane systems (supported layers of vesicles andupported lipid bilayers), which are also relevant for lipid-basedanobiotechnology applications [23–25]. Though less numerous,CM-D has been successfully used for phase transitions studies of
ipid model membranes [26,27], as well as for peptide–lipid inter-ctions [28–31].
In this work we examine the role of lateral phase separation onhe disrupting action of melittin using QCM-D. Melittin is a pep-ide naturally present in the venom of the honeybee Apis mellifera.he �-helical peptide consists out of 26 amino acids of which theargest part is hydrophobic (1–21), while near the C-terminus theres a hydrophilic sequence (22–26) [32]. Despite its cytotoxicity toe used as an antibiotic, melittin can be considered as a relevantodel peptide since it shares common features with AMPs such
s magainins and cecropins when bound to lipid bilayer interfaces2,33,34]. Our aim is to verify that the disrupting efficiency of melit-in is enhanced in raft-containing membranes, despite the presencef cholesterol. To this end, we chose dipalmitoylphosphatidyl-holine (DPPC), 1,2-dioleoyl-sn-glycero-3-phosphocholine (DOPC),nd cholesterol, since the phase boundaries of DPPC + cholesterolnon-raft forming) and DPPC + DOPC + cholesterol (raft-forming)re well mapped in phase diagrams [35–38].
. Experimental
.1. Materials
DPPC, cholesterol in powder, and DOPC in chloroform were pur-hased from Avanti Polar Lipids (Alabaster, AL). HEPES buffer (pH.4) consisting of 10 mM HEPES from Fisher Scientific (assay 99%)nd 150 mM NaCl from Sigma–Aldrich (assay ≥ 99.5%) was used forydration of the dried lipids. The quantities of lipids to reach theesired mixture concentrations were determined gravimeticallysing a Sartorius balance yielding a maximal mole fraction uncer-ainty of ±0.002. Melittin from honey bee venom (assay ≥ 85%)as obtained from Sigma and dissolved in buffer to 10 �g/mL or
3.5 �M), being above the minimum melittin concentration releasef 0.85 �g/mL (0.3 �M) [29,30].
.2. Vesicle preparation
The pure lipids or lipid mixtures were first dissolved in spec-roscopic grade chloroform from Analar Normapur (assay 99.3%tabilized with about 0.6% ethanol) and the solvent was then evap-rated under a mild flow of nitrogen in a round-bottom flask. Theesulting lipid film was kept under vacuum overnight to removeesidual solvent. Afterwards, the lipid was hydrated with HEPESuffer. Hydration to 1 mg/mL was carried out under continuoustirring in a temperature-controlled water bath at sufficientlyigh temperature (∼60 ◦C). Small unilamellar vesicles (SUVs) were
ormed by extrusion through a filter support (Avanti Polar Lipids)ith a pore size of 100 nm for 25 times. Vesicle effective sizes
nd polydispersities were determined by dynamic light scattering
Please cite this article in press as: P. Losada-Pérez, et al., Mmembranes: A study by quartz crystal microbalance with dishttp://dx.doi.org/10.1016/j.colsurfb.2014.10.048
Zeta Pals, Brookhaven Instruments Corporation). The obtainedverage diameters and polydispersities of the samples used areisplayed in Table 1 of the supplementary material. The vesicleispersions were diluted to 0.1 mg/mL prior to use in the QCM-D
PRESSs B: Biointerfaces xxx (2014) xxx–xxx
experiments. Non-raft forming mixtures of DPPC + cholesterolwere made with the following weight fractions wchol = 0, 0.07, 0.17,0.24, DOPC + cholesterol with weigh fraction wchol = 0.24, whileraft-forming mixtures were DPPC and DOPC (1:1) with the sameweight fractions of cholesterol.
2.3. Quartz crystal microbalance with dissipation monitoring
Quartz crystal microbalance with dissipation monitoring (QCM-D) is an acoustic surface-sensitive technique based on the inversepiezoelectric effect. The application of an AC voltage over the sensorelectrodes causes the piezoelectric quartz crystal to oscillate at itsacoustic resonance frequency. The result of the vibration motionof the quartz crystal is a transverse acoustic wave that propagatesacross the crystal, reflecting back into the crystal at the surface.When the AC voltage is turned off, the oscillation amplitude decaysexponentially, this decay is recorded and the frequency (f) and theenergy dissipation factor (D) of different overtones are extracted[23]. The dissipation D is the ratio between the dissipated energyduring one vibration cycle and the total kinetic and potential energyof the crystal at that moment.
When molecules adsorb to an oscillating quartz crystal, water(or other liquid) couples to the adsorbed material as an additionaldynamic mass via direct hydration and/or entrapment within theadsorbed film. Thus, the layer is sensed as a viscoelastic hydro-gel composed of the molecules and the coupled water together.For the current measurements, we have used QCM-D on a Q-senseE4 instrument (Gothenborg, Sweden) monitoring the frequencyshift �f and the dissipation change �D. AT-cut quartz crystalswith Au and SiO2 coating (diameter 14 mm, thickness 0.3 mm, sur-face roughness 3 nm and resonant frequency 4.95 MHz) were used.The Au-coated quartz sensors were cleaned with a 5:1:1 mixtureof Milli-Q water (conductivity of 0.055 S cm−1 at 25 ◦C), ammoniaand hydrogen peroxide and were UV-ozone treated with a DigitalPSD series UV-ozone system from Novascan for 15 min, followedby rinsing in water and drying with N2. The SiO2-coated quartzsensors were cleaned in a solution of sodium dodecyl sulfate (2%SDS) for 30 min and UV-ozone treated for 1 h, followed by rinsing inwater and drying with N2. The changes in �f and in �D were moni-tored at five different overtones (from 3rd to 11th, the fundamentalfrequency is rather unstable since this is the one that reaches thefarthest out to the edge of the sensor and may be disturbed by the O-ring). For all the samples, Au and SiO2 substrates were used to assessthe substrate dependence on layer formation and the experimentswere carried out at 50 ◦C and at 25 ◦C. The temperature stability atconstant temperature is ±0.02 K. The measurements were done inflow mode with a flow rate of 50 �L/min. First, a baseline with pureHEPES buffer was established and afterwards lipid vesicles wereinjected over the sensor chip. After reaching a stable vesicle layeror lipid bilayer, buffer rinsing was carried out to prove the stabilityof the layer and melittin at 10 �g/mL in HEPES buffer was inserted.
3. Results and discussion
The results obtained are divided into two main parts: (i) vesicledeposition and lipid layer formation and (ii) melittin interactionswith the lipid layer. For the former, the influence of parameterssuch as substrate nature (SiO2, Au), temperature of deposition andamount of cholesterol is evaluated. For the latter, the action onmelittin is explained in terms of the geometry, i.e., supported layerof vesicles SLV or supported lipid bilayer SLB, the thickness and thecomposition of the lipid layer formed.
elittin disruption of raft and non-raft-forming biomimeticsipation monitoring, Colloids Surf. B: Biointerfaces (2014),
3.1. Lipid layer formation
Lipid layer formation depends on the vesicle–vesicle inter-actions and vesicles–substrate interactions that influence the
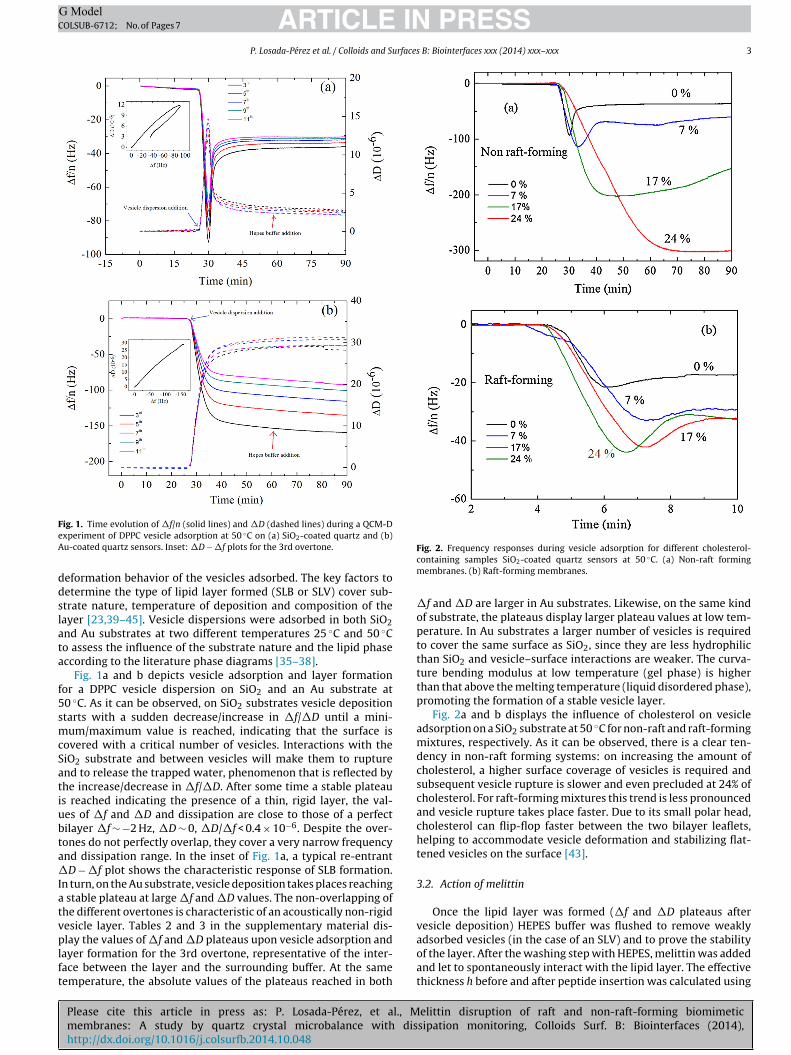
ARTICLE IN PRESSG ModelCOLSUB-6712; No. of Pages 7
P. Losada-Pérez et al. / Colloids and Surfaces B: Biointerfaces xxx (2014) xxx–xxx 3
FeA
ddslata
f5smcSatiubta�Iatvplft
ig. 1. Time evolution of �f/n (solid lines) and �D (dashed lines) during a QCM-Dxperiment of DPPC vesicle adsorption at 50 ◦C on (a) SiO2-coated quartz and (b)u-coated quartz sensors. Inset: �D − �f plots for the 3rd overtone.
eformation behavior of the vesicles adsorbed. The key factors toetermine the type of lipid layer formed (SLB or SLV) cover sub-trate nature, temperature of deposition and composition of theayer [23,39–45]. Vesicle dispersions were adsorbed in both SiO2nd Au substrates at two different temperatures 25 ◦C and 50 ◦Co assess the influence of the substrate nature and the lipid phaseccording to the literature phase diagrams [35–38].
Fig. 1a and b depicts vesicle adsorption and layer formationor a DPPC vesicle dispersion on SiO2 and an Au substrate at0 ◦C. As it can be observed, on SiO2 substrates vesicle depositiontarts with a sudden decrease/increase in �f/�D until a mini-um/maximum value is reached, indicating that the surface is
overed with a critical number of vesicles. Interactions with theiO2 substrate and between vesicles will make them to rupturend to release the trapped water, phenomenon that is reflected byhe increase/decrease in �f/�D. After some time a stable plateaus reached indicating the presence of a thin, rigid layer, the val-es of �f and �D and dissipation are close to those of a perfectilayer �f ∼ −2 Hz, �D ∼ 0, �D/�f < 0.4 × 10−6. Despite the over-ones do not perfectly overlap, they cover a very narrow frequencynd dissipation range. In the inset of Fig. 1a, a typical re-entrantD − �f plot shows the characteristic response of SLB formation.
n turn, on the Au substrate, vesicle deposition takes places reaching stable plateau at large �f and �D values. The non-overlapping ofhe different overtones is characteristic of an acoustically non-rigidesicle layer. Tables 2 and 3 in the supplementary material dis-
Please cite this article in press as: P. Losada-Pérez, et al., Mmembranes: A study by quartz crystal microbalance with dishttp://dx.doi.org/10.1016/j.colsurfb.2014.10.048
lay the values of �f and �D plateaus upon vesicle adsorption andayer formation for the 3rd overtone, representative of the inter-ace between the layer and the surrounding buffer. At the sameemperature, the absolute values of the plateaus reached in both
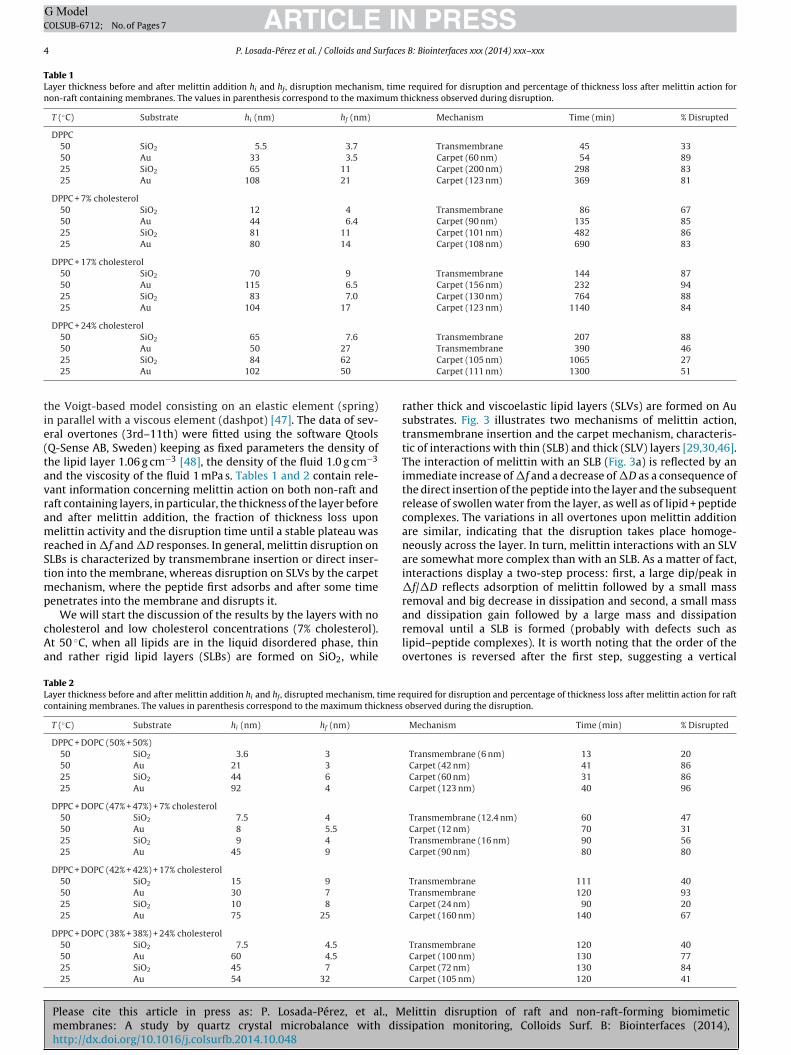
Fig. 2. Frequency responses during vesicle adsorption for different cholesterol-containing samples SiO2-coated quartz sensors at 50 ◦C. (a) Non-raft formingmembranes. (b) Raft-forming membranes.
�f and �D are larger in Au substrates. Likewise, on the same kindof substrate, the plateaus display larger plateau values at low tem-perature. In Au substrates a larger number of vesicles is requiredto cover the same surface as SiO2, since they are less hydrophilicthan SiO2 and vesicle–surface interactions are weaker. The curva-ture bending modulus at low temperature (gel phase) is higherthan that above the melting temperature (liquid disordered phase),promoting the formation of a stable vesicle layer.
Fig. 2a and b displays the influence of cholesterol on vesicleadsorption on a SiO2 substrate at 50 ◦C for non-raft and raft-formingmixtures, respectively. As it can be observed, there is a clear ten-dency in non-raft forming systems: on increasing the amount ofcholesterol, a higher surface coverage of vesicles is required andsubsequent vesicle rupture is slower and even precluded at 24% ofcholesterol. For raft-forming mixtures this trend is less pronouncedand vesicle rupture takes place faster. Due to its small polar head,cholesterol can flip-flop faster between the two bilayer leaflets,helping to accommodate vesicle deformation and stabilizing flat-tened vesicles on the surface [43].
3.2. Action of melittin
Once the lipid layer was formed (�f and �D plateaus aftervesicle deposition) HEPES buffer was flushed to remove weakly
elittin disruption of raft and non-raft-forming biomimeticsipation monitoring, Colloids Surf. B: Biointerfaces (2014),
adsorbed vesicles (in the case of an SLV) and to prove the stabilityof the layer. After the washing step with HEPES, melittin was addedand let to spontaneously interact with the lipid layer. The effectivethickness h before and after peptide insertion was calculated using
ARTICLE IN PRESSG ModelCOLSUB-6712; No. of Pages 7
4 P. Losada-Pérez et al. / Colloids and Surfaces B: Biointerfaces xxx (2014) xxx–xxx
Table 1Layer thickness before and after melittin addition hi and hf , disruption mechanism, time required for disruption and percentage of thickness loss after melittin action fornon-raft containing membranes. The values in parenthesis correspond to the maximum thickness observed during disruption.
T (◦C) Substrate hi (nm) hf (nm) Mechanism Time (min) % Disrupted
DPPC50 SiO2 5.5 3.7 Transmembrane 45 3350 Au 33 3.5 Carpet (60 nm) 54 8925 SiO2 65 11 Carpet (200 nm) 298 8325 Au 108 21 Carpet (123 nm) 369 81
DPPC + 7% cholesterol50 SiO2 12 4 Transmembrane 86 6750 Au 44 6.4 Carpet (90 nm) 135 8525 SiO2 81 11 Carpet (101 nm) 482 8625 Au 80 14 Carpet (108 nm) 690 83
DPPC + 17% cholesterol50 SiO2 70 9 Transmembrane 144 8750 Au 115 6.5 Carpet (156 nm) 232 9425 SiO2 83 7.0 Carpet (130 nm) 764 8825 Au 104 17 Carpet (123 nm) 1140 84
DPPC + 24% cholesterol50 SiO2 65 7.6 Transmembrane 207 88
tie(tavramrStmp
cAa
TLc
50 Au 50 27
25 SiO2 84 62
25 Au 102 50
he Voigt-based model consisting on an elastic element (spring)n parallel with a viscous element (dashpot) [47]. The data of sev-ral overtones (3rd–11th) were fitted using the software QtoolsQ-Sense AB, Sweden) keeping as fixed parameters the density ofhe lipid layer 1.06 g cm−3 [48], the density of the fluid 1.0 g cm−3
nd the viscosity of the fluid 1 mPa s. Tables 1 and 2 contain rele-ant information concerning melittin action on both non-raft andaft containing layers, in particular, the thickness of the layer beforend after melittin addition, the fraction of thickness loss uponelittin activity and the disruption time until a stable plateau was
eached in �f and �D responses. In general, melittin disruption onLBs is characterized by transmembrane insertion or direct inser-ion into the membrane, whereas disruption on SLVs by the carpet
echanism, where the peptide first adsorbs and after some timeenetrates into the membrane and disrupts it.
Please cite this article in press as: P. Losada-Pérez, et al., Mmembranes: A study by quartz crystal microbalance with dishttp://dx.doi.org/10.1016/j.colsurfb.2014.10.048
We will start the discussion of the results by the layers with noholesterol and low cholesterol concentrations (7% cholesterol).t 50 ◦C, when all lipids are in the liquid disordered phase, thinnd rather rigid lipid layers (SLBs) are formed on SiO2, while
able 2ayer thickness before and after melittin addition hi and hf , disrupted mechanism, time reontaining membranes. The values in parenthesis correspond to the maximum thickness
T (◦C) Substrate hi (nm) hf (nm)
DPPC + DOPC (50% + 50%)50 SiO2 3.6 3
50 Au 21 3
25 SiO2 44 6
25 Au 92 4
DPPC + DOPC (47% + 47%) + 7% cholesterol50 SiO2 7.5 4
50 Au 8 5.5
25 SiO2 9 4
25 Au 45 9
DPPC + DOPC (42% + 42%) + 17% cholesterol50 SiO2 15 9
50 Au 30 7
25 SiO2 10 8
25 Au 75 25
DPPC + DOPC (38% + 38%) + 24% cholesterol50 SiO2 7.5 4.5
50 Au 60 4.5
25 SiO2 45 7
25 Au 54 32
Transmembrane 390 46Carpet (105 nm) 1065 27Carpet (111 nm) 1300 51
rather thick and viscoelastic lipid layers (SLVs) are formed on Ausubstrates. Fig. 3 illustrates two mechanisms of melittin action,transmembrane insertion and the carpet mechanism, characteris-tic of interactions with thin (SLB) and thick (SLV) layers [29,30,46].The interaction of melittin with an SLB (Fig. 3a) is reflected by animmediate increase of �f and a decrease of �D as a consequence ofthe direct insertion of the peptide into the layer and the subsequentrelease of swollen water from the layer, as well as of lipid + peptidecomplexes. The variations in all overtones upon melittin additionare similar, indicating that the disruption takes place homoge-neously across the layer. In turn, melittin interactions with an SLVare somewhat more complex than with an SLB. As a matter of fact,interactions display a two-step process: first, a large dip/peak in�f/�D reflects adsorption of melittin followed by a small massremoval and big decrease in dissipation and second, a small mass
elittin disruption of raft and non-raft-forming biomimeticsipation monitoring, Colloids Surf. B: Biointerfaces (2014),
and dissipation gain followed by a large mass and dissipationremoval until a SLB is formed (probably with defects such aslipid–peptide complexes). It is worth noting that the order of theovertones is reversed after the first step, suggesting a vertical
quired for disruption and percentage of thickness loss after melittin action for raft observed during the disruption.
Mechanism Time (min) % Disrupted
Transmembrane (6 nm) 13 20Carpet (42 nm) 41 86Carpet (60 nm) 31 86Carpet (123 nm) 40 96
Transmembrane (12.4 nm) 60 47Carpet (12 nm) 70 31Transmembrane (16 nm) 90 56Carpet (90 nm) 80 80
Transmembrane 111 40Transmembrane 120 93Carpet (24 nm) 90 20Carpet (160 nm) 140 67
Transmembrane 120 40Carpet (100 nm) 130 77Carpet (72 nm) 130 84Carpet (105 nm) 120 41

ARTICLE IN PRESSG ModelCOLSUB-6712; No. of Pages 7
P. Losada-Pérez et al. / Colloids and Surfaces B: Biointerfaces xxx (2014) xxx–xxx 5
F ◦
as
irmipracAcS
ig. 3. Melittin disrupting mechanisms at 50 C. (a) Transmembrane insertion into SLB of DPPC on SiO2 substrate. (b) Carpet mechanism into a SLV of DPPC on a Auubstrate. Insets: Thickness of the lipid layer before and after melittin addition.
nhomogeneous mass removal dominated process [29]. Theseesponses are characteristic of the so-called carpet mechanism:elittin first adsorbs onto the lipid layer and after some time it
nserts into the membrane and perforates it, releasing water andeptide–lipid complexes until an SLB is eventually formed. Theesulting layers are imperfect SLBs with ∼3–4 nm in both SiO2nd Au, most likely characterized by defects from lipid–melittin
Please cite this article in press as: P. Losada-Pérez, et al., Mmembranes: A study by quartz crystal microbalance with dishttp://dx.doi.org/10.1016/j.colsurfb.2014.10.048
omplexes released. At 25 ◦C, SLVs are formed both on SiO2 and onu substrates, being thicker than their 50 ◦C counterparts. In raftontaining membranes, SLVs are disrupted giving rise to a finalLB (∼4–7 nm) as a result of melittin action, while slightly thicker
Fig. 5. Time dependence of melittin disrupting activity on cholesterol content for
Fig. 4. Melittin disrupting mechanisms at 25 ◦C on Au substrate (a) DPPC + 24%cholesterol (b) DPPC + DOPC (1:1) + 24% cholesterol. Insets: Thickness of the lipidlayer before and after melittin addition.
layers ∼11 nm are obtained for non-raft containing ones. Theoverall thickness loss is similar in both non-raft and raft containinglipid layers. It is worth noting that for layers with similar thickness,it takes longer time for melittin to insert and disrupt non-raftforming than raft-forming (see Tables 1 and 2). In the former,coexistence of liquid-disordered (DOPC) and gel phases (DPPC),melittin selectively targets and inserts into DOPC-rich domains,
elittin disruption of raft and non-raft-forming biomimeticsipation monitoring, Colloids Surf. B: Biointerfaces (2014),
while in non-raft forming all lipids are in the gel, compact phasemaking more difficult for melittin to act.
At high cholesterol concentrations (17% and 24% cholesterol),non-raft-containing vesicles form slightly thicker layers than
non-raft forming (a) and raft-forming (b) biomimetic membranes at 50 ◦C.
ARTICLE IN PRESSG ModelCOLSUB-6712; No. of Pages 7
6 P. Losada-Pérez et al. / Colloids and Surfaces B: Biointerfaces xxx (2014) xxx–xxx
nt for
ratarbaF5as
tToraactimcFms
4
mmsgcswtglaato
mwi
[[
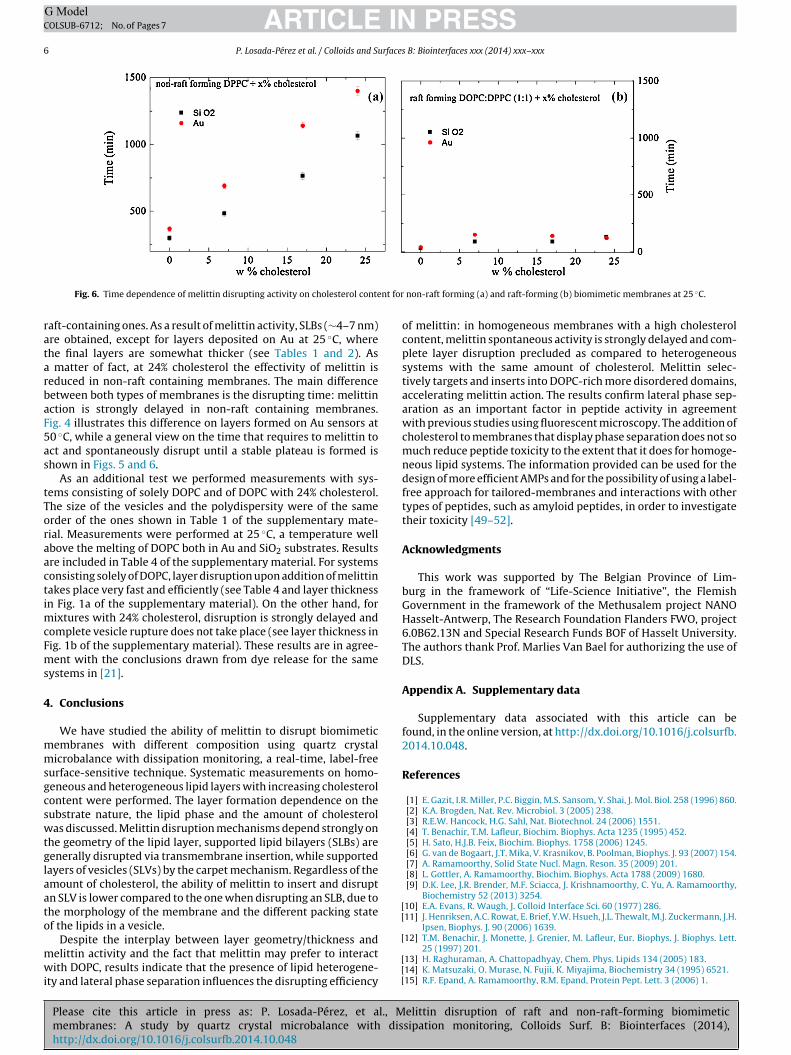
Fig. 6. Time dependence of melittin disrupting activity on cholesterol conte
aft-containing ones. As a result of melittin activity, SLBs (∼4–7 nm)re obtained, except for layers deposited on Au at 25 ◦C, wherehe final layers are somewhat thicker (see Tables 1 and 2). As
matter of fact, at 24% cholesterol the effectivity of melittin iseduced in non-raft containing membranes. The main differenceetween both types of membranes is the disrupting time: melittinction is strongly delayed in non-raft containing membranes.ig. 4 illustrates this difference on layers formed on Au sensors at0 ◦C, while a general view on the time that requires to melittin toct and spontaneously disrupt until a stable plateau is formed ishown in Figs. 5 and 6.
As an additional test we performed measurements with sys-ems consisting of solely DOPC and of DOPC with 24% cholesterol.he size of the vesicles and the polydispersity were of the samerder of the ones shown in Table 1 of the supplementary mate-ial. Measurements were performed at 25 ◦C, a temperature wellbove the melting of DOPC both in Au and SiO2 substrates. Resultsre included in Table 4 of the supplementary material. For systemsonsisting solely of DOPC, layer disruption upon addition of melittinakes place very fast and efficiently (see Table 4 and layer thicknessn Fig. 1a of the supplementary material). On the other hand, for
ixtures with 24% cholesterol, disruption is strongly delayed andomplete vesicle rupture does not take place (see layer thickness inig. 1b of the supplementary material). These results are in agree-ent with the conclusions drawn from dye release for the same
ystems in [21].
. Conclusions
We have studied the ability of melittin to disrupt biomimeticembranes with different composition using quartz crystalicrobalance with dissipation monitoring, a real-time, label-free
urface-sensitive technique. Systematic measurements on homo-eneous and heterogeneous lipid layers with increasing cholesterolontent were performed. The layer formation dependence on theubstrate nature, the lipid phase and the amount of cholesterolas discussed. Melittin disruption mechanisms depend strongly on
he geometry of the lipid layer, supported lipid bilayers (SLBs) areenerally disrupted via transmembrane insertion, while supportedayers of vesicles (SLVs) by the carpet mechanism. Regardless of themount of cholesterol, the ability of melittin to insert and disruptn SLV is lower compared to the one when disrupting an SLB, due tohe morphology of the membrane and the different packing statef the lipids in a vesicle.
Please cite this article in press as: P. Losada-Pérez, et al., Mmembranes: A study by quartz crystal microbalance with dishttp://dx.doi.org/10.1016/j.colsurfb.2014.10.048
Despite the interplay between layer geometry/thickness andelittin activity and the fact that melittin may prefer to interactith DOPC, results indicate that the presence of lipid heterogene-
ty and lateral phase separation influences the disrupting efficiency
[
[[[
non-raft forming (a) and raft-forming (b) biomimetic membranes at 25 ◦C.
of melittin: in homogeneous membranes with a high cholesterolcontent, melittin spontaneous activity is strongly delayed and com-plete layer disruption precluded as compared to heterogeneoussystems with the same amount of cholesterol. Melittin selec-tively targets and inserts into DOPC-rich more disordered domains,accelerating melittin action. The results confirm lateral phase sep-aration as an important factor in peptide activity in agreementwith previous studies using fluorescent microscopy. The addition ofcholesterol to membranes that display phase separation does not somuch reduce peptide toxicity to the extent that it does for homoge-neous lipid systems. The information provided can be used for thedesign of more efficient AMPs and for the possibility of using a label-free approach for tailored-membranes and interactions with othertypes of peptides, such as amyloid peptides, in order to investigatetheir toxicity [49–52].
Acknowledgments
This work was supported by The Belgian Province of Lim-burg in the framework of “Life-Science Initiative”, the FlemishGovernment in the framework of the Methusalem project NANOHasselt-Antwerp, The Research Foundation Flanders FWO, project6.0B62.13N and Special Research Funds BOF of Hasselt University.The authors thank Prof. Marlies Van Bael for authorizing the use ofDLS.
Appendix A. Supplementary data
Supplementary data associated with this article can befound, in the online version, at http://dx.doi.org/10.1016/j.colsurfb.2014.10.048.
References
[1] E. Gazit, I.R. Miller, P.C. Biggin, M.S. Sansom, Y. Shai, J. Mol. Biol. 258 (1996) 860.[2] K.A. Brogden, Nat. Rev. Microbiol. 3 (2005) 238.[3] R.E.W. Hancock, H.G. Sahl, Nat. Biotechnol. 24 (2006) 1551.[4] T. Benachir, T.M. Lafleur, Biochim. Biophys. Acta 1235 (1995) 452.[5] H. Sato, H.J.B. Feix, Biochim. Biophys. 1758 (2006) 1245.[6] G. van de Bogaart, J.T. Mika, V. Krasnikov, B. Poolman, Biophys. J. 93 (2007) 154.[7] A. Ramamoorthy, Solid State Nucl. Magn. Reson. 35 (2009) 201.[8] L. Gottler, A. Ramamoorthy, Biochim. Biophys. Acta 1788 (2009) 1680.[9] D.K. Lee, J.R. Brender, M.F. Sciacca, J. Krishnamoorthy, C. Yu, A. Ramamoorthy,
Biochemistry 52 (2013) 3254.10] E.A. Evans, R. Waugh, J. Colloid Interface Sci. 60 (1977) 286.11] J. Henriksen, A.C. Rowat, E. Brief, Y.W. Hsueh, J.L. Thewalt, M.J. Zuckermann, J.H.
Ipsen, Biophys. J. 90 (2006) 1639.
elittin disruption of raft and non-raft-forming biomimeticsipation monitoring, Colloids Surf. B: Biointerfaces (2014),
12] T.M. Benachir, J. Monette, J. Grenier, M. Lafleur, Eur. Biophys. J. Biophys. Lett.25 (1997) 201.
13] H. Raghuraman, A. Chattopadhyay, Chem. Phys. Lipids 134 (2005) 183.14] K. Matsuzaki, O. Murase, N. Fujii, K. Miyajima, Biochemistry 34 (1995) 6521.15] R.F. Epand, A. Ramamoorthy, R.M. Epand, Protein Pept. Lett. 3 (2006) 1.

ING ModelC
rfaces
[
[
[[[
[
[[[
[[[
[
[[[[[
[
[[[[[[[[[[[[
[
[
[
ARTICLEOLSUB-6712; No. of Pages 7
P. Losada-Pérez et al. / Colloids and Su
16] A. Bhunia, P.N. Domadia, J. Torres, K.J. Hallock, A. Ramamoorthy, S. Bhattachar-jya, J. Biol. Chem. 285 (2010) 3883.
17] A. Ramamoorthy, D.K. Lee, T. Narasimhaswamy, R.V.P. Nanga, Biochim. Biophys.Acta 1798 (2010) 223.
18] K. Simons, E. Ikonen, Nature 387 (1997) 569.19] A. Pokorny, P.F.F. Almeida, Biochemistry 44 (2005) 9538.20] A. Pokorny, L.E. Yandek, A.I. Elegbede, A. Hinderliter, P.F.F. Almeida, Biophys. J.
91 (2006) 2184.21] A.J. McHenry, M.F.M. Sciacca, J.R. Brender, A. Ramamoorthy, Biochim. Biophys.
Acta 1818 (2012) 3019.22] J.R. Brender, A.J. McHenry, A. Ramamoorthy, Front. Inmunol. 3 (2012) 1.23] C.A. Keller, B. Kasemo, Biophys. J. 75 (1998) 1397.24] T. Hanik, M. Snejdarkova, L. Sokolikova, E. Meszar, R. Krivanek, V. Tvarozek, L.
Novotny, J. Wang, Sens. Actuators B 57 (1999) 201.25] S. Mashaghi, T. Jadidi, G. Koenderink, A. Mashaghi, J. Mol. Sci. 14 (2013) 4242.26] G. Ohlsson, A. Tigerström, F. Höök, B. Kasemo, Soft Matter 7 (2011) 10479.27] P. Losada-Pérez, K.L. Jiménez-Monroy, B. van Grinsven, J. Leys, S.D. Janssens, M.
Peeters, C. Glorieux, J. Thoen, K. Haenen, W. De Ceuninck, P. Wagner, Phys. Stat.Solidi A 211 (2014) 1377.
28] K.F. Wang, R. Nagarajan, C.M. Mello, T.A. Camesano, J. Phys. Chem. B 115 (2011)15228.
Please cite this article in press as: P. Losada-Pérez, et al., Mmembranes: A study by quartz crystal microbalance with dishttp://dx.doi.org/10.1016/j.colsurfb.2014.10.048
29] N. Lu, K. Yang, B. Yuan, Y.Q. Ma, J. Phys. Chem. B 116 (2012) 9432.30] N. Lu, K. Yang, J.L. Li, B. Yuan, Y.Q. Ma, Biochim. Biophys. Acta 1828 (2013) 1918.31] K.F. Wang, R. Nagarajan, T.A. Camesano, Colloids Surf. B 116 (2014) 472.32] H. Vogel, F. Jähnig, Biophys. J. 50 (1986) 573.33] B. Bechinger, B.K. Lohner, Biophys. Acta 1758 (2006) 1529.
[[
[
PRESS B: Biointerfaces xxx (2014) xxx–xxx 7
34] P. Wessman, A.A. Strömstedt, M. Malmsten, K. Edwards, Biophys. J. 95 (2008)4324.
35] M.R. Vist, J.H. Davis, Biochemistry 29 (1990) 451.36] S.L. Veatch, S.L. Keller, Biophys. J. 85 (2003) 3074.37] J.H. Davis, J.J. Clair, J. Juhasz, Biophys. J. 96 (2009) 521.38] D. Marsh, Biochim. Biophys. Acta 1798 (2010) 688.39] E. Reimbult, F. Höök, B. Kasemo, Phys. Rev. E 66 (2002) 051905.40] E. Reimbult, F. Höök, B. Kasemo, Langmuir 19 (2003) 1681.41] R. Richter, A. Mukhopadhyay, A. Brisson, Biophys. J. 85 (2003) 3035.42] B. Seantier, C. Breffa, O. Félix, G. Decher, J. Phys. Chem. B 109 (2005) 21755.43] M. Sundh, S. Svedhem, D. Sutherland, Phys. Chem. Chem. Phys. 12 (2010) 453.44] T. Zhu, F. Xu, B. Yuan, C. Ren, Z. Jiang, Y. Ma, Colloids Surf. B 89 (2012) 228.45] Y. Jing, H. Trefna, M. Persson, B. Kasemo, S. Svedhem, Soft Matter 10 (2014) 187.46] G.A. McCubbin, D. Praporski, S. Piantavigna, D. Knappe, R. Hoffmann, J.H. Bowie,
F. Separovic, L.L. Martin, Eur. Biophys. J. 40 (2011) 437.47] T.H. Courtney, in: L. Grove (Ed.), Mechanical Behavior of Materials, Waveland
Press, Illinois, 2005.48] A.P. Serro, A. Carapeto, G. Paiva, J.P.S. Farinha, R. Colac o, B. Saramago, Surf.
Interface Anal. 44 (2012) 426.49] S.A. Kotler, P. Walsh, J.R. Brender, A. Ramamoorthy, Chem. Soc. Rev. 43 (2014)
6692.
elittin disruption of raft and non-raft-forming biomimeticsipation monitoring, Colloids Surf. B: Biointerfaces (2014),
50] J.R. Brender, S. Salamekh, A. Ramamoorthy, Acc. Chem. Res. 116 (2012) 3650.51] J.R. Brender, U.H.N. Dürr, D. Heyl, M.B. Budarapu, A. Ramamoorthy, Biochim.
Biophys. Acta 1768 (2007) 2026.52] J.R. Brender, D.L. Heyl, S. Samisetti, S.A. Kotler, J.M. Osborne, R.R. Pesaru, A.
Ramamoorthy, Phys. Chem. Chem. Phys. 15 (2013) 8908.




















