
FAD binding properties of a cytosolic version of Escherichia coli NADH dehydrogenase-2
Upload
independentCategory
view
1download
0
Lysophosphatidylcholines prime the NADPH oxidase andstimulate multiple neutrophil functions through changesin cytosolic calcium
Christopher C. Silliman,*,†,‡ David J. Elzi,*,† Daniel R. Ambruso,*,† Rene J. Musters,‡
Christine Hamiel,‡ Ronald J. Harbeck,§ Andrew J. Paterson,*,† A. Jason Bjornsen,†
Travis H. Wyman,† Marguerite Kelher,‡ Kelly M. England,† Nathan McLaughlin-Malaxecheberria,†
Carlton C. Barnett,‡ Junichi Aiboshi,‡ and Anirban Bannerjee‡
*Bonfils Blood Center and Departments of †Pediatrics and ‡Surgery, University of Colorado School of Medicine,Denver; and §Department of Laboratory Medicine, National Jewish Center for Allergy and RespiratoryMedicine, Denver, Colorado
Abstract: A mixture of lysophosphatidylcholines(lyso-PCs) are generated during blood storage andare etiologic in models of acute lung injury. Wehypothesize that lyso-PCs stimulate polymorpho-nuclear neutrophils (PMNs) through Ca2�-depen-dent signaling. The lyso-PC mix (0.45–14.5 �M)and the individual lyso-PCs primed formyl-Met-Leu-Phe (fMLP) activation of the oxidase (1.8- to15.7-fold and 1.7- to 14.8-fold; P<0.05). Labeledlyso-PCs demonstrated a membrane associationwith PMNs and caused rapid increases in cytosolicCa2�. Receptor desensitization studies implicateda common receptor or a family of receptors for theobserved lyso-PC-mediated changes in PMN prim-ing, and cytosolic Ca2� functions were pertussistoxin-sensitive. Lyso-PCs caused rapid serine phos-phorylation of a 68-kD protein but did not activatemitogen-activated protein kinases or cause changesin tyrosine phosphorylation. With respect to alter-ations in PMN function, lyso-PCs caused PMN ad-herence, increased expression of CD11b and thefMLP receptor, reduced chemotaxis, provokedchanges in morphology, elicited degranulation,and augmented fMLP-induced azurophilic degran-ulation (P<0.05). Cytosolic Ca2� chelation inhib-ited lyso-PC-mediated priming of the oxidase,CD11b surface expression, changes in PMN mor-phology, and serine phosphorylation of the 68-kDprotein. In conclusion, lyso-PCs affect multiplePMN functions in a Ca2�-dependent manner thatinvolves the activation of a pertussis toxin-sensitiveG-protein. J. Leukoc. Biol. 73: 511–524; 2003.
Key Words: adhesion � chemotaxis � morphology
INTRODUCTION
Polymorphonuclear neutrophils (PMNs) are critical in hostdefense against microbial invaders [1, 2]. Although abundantin the circulation, PMNs exert their major microbicidal func-
tion in the tissues [3–7]. The process of PMN emigration fromthe circulation to the site of infection has been the subject ofnumerous papers and will not be extensively reviewed here[3–7]. PMN priming, an orderly part of emigration, is initiatedby the attraction and firm adhesion of PMNs to activatedvascular endothelium and continues until the pathogens areencountered and destroyed [8–12]. Priming is a process that isdistinct from oxidase activation and not only maximizes themicrobicidal function of PMNs in response to a subsequentstimulus but also causes a change in the avidity and the surfaceexpression of the �2 integrins and results in modest degranu-lation [13–15].
In previous studies, we have demonstrated the generation of aneffective PMN priming activity during routine storage of cellularblood components [16, 17]. This activity has been identified bygas chromatography/mass spectroscopy as a mixture of lysophos-phatidylcholines (lyso-PCs) in micromolar concentrations [16].Moreover, we have implicated these lipids in the pathogenesis oftransfusion-related acute lung injury (TRALI) in humans and havedemonstrated that lyso-PCs are etiologic in a two-event animalmodel of PMN-mediated TRALI [18, 19]. However, a controversyexists regarding the activity of lyso-PCs, as a number of investi-gators have asserted that lyso-PCs are inactive with respect toleukocytes and platelets [20–24]. We hypothesize that lyso-PCsdirectly prime the PMN oxidase and stimulate multiple PMNfunctions through signaling mechanisms that require activation ofa receptor linked to a pertussis toxin-sensitive G-protein andincreases in cytosolic Ca2�.
MATERIALS AND METHODS
MaterialsAll chemicals, unless otherwise specified, were purchased from Sigma Chem-ical Co. (St. Louis, MO). 1-O-Octadecyl-sn-glycero-3-phosphocholine (C18
Correspondence: Christopher C. Silliman, M.D., Ph.D., Associate MedicalDirector, Bonfils Blood Center, 717 Yosemite Circle, Denver, CO 80230.E-mail: [email protected]
Received April 10, 2002; revised November 21, 2002; accepted December18, 2002; doi: 10.1189/jlb.0402179.
Journal of Leukocyte Biology Volume 73, April 2003 511

lyso-PAF) was obtained from Biomol (King of Prussia, PA). A custom synthesisof fluorescein isothiocyanate (FITC)-labeled 1-O-dodecanoyl (lauroyl) lyso-PCwas procured from Avanti Polar Lipids (Alabaster, AL). Indo-1, acetoxymethylester (AM; indo-1), and 1,2-bis(O-aminophenyl-ethane-ethane)-N,N,N�,N�-tet-raacetic acid (BAPTA), AM, were purchased from Molecular Probes (Eugene,OR). Murine monoclonal antibodies (mAb) to human CD11b and to theformyl-Met-Leu-Phe receptor (fMLPr), plus the isotype controls, were pur-chased from Becton Dickinson (San Jose, CA). All solutions were made fromsterile water, U.S. Pharmacopeia (USP), purchased along with sterile 0.9%saline, USP, from Baxter Healthcare Corp. (Deerfield, IL). All buffers weremade from the following stock USP solutions: 10% CaCl2 (Fujisawa USA,Deerfield, IL); 23.4% NaCl, 20 Meq/ml KCl, and 50% MgSO4 (AmericanRegent Laboratories, Shirley, NY); and sodium phosphates (278 mg/ml mono-basic and 142 mg/ml dibasic) and 50% dextrose (Abbott Laboratories, NorthChicago, IL). All solutions were sterile-filtered with NalgeneTM MF75 seriesdisposable sterilization filter units purchased from Fischer Scientific Corp. (Pitts-burgh, PA). A Leica DRM-mechanized fluorescence microscope, equipped with amovable stage with a custom Zeiss 63X water-immersion lens, was purchased fromLeica Microsystems (Exton, PA). Four epifluorescence cubes (FITC, Cy-3, Cy-5,and 7-amino-4-methyl-3-coumarinyl acetic acid) were obtained from ChromaTechnology Corp. (Brattleboro, VT). A cooled charged-coupled device camera andSlidebookTM software for computer operation were purchased from The CookeCorporation (Tonawanda, NY) and Intelligent Imaging Innovations (Lakewood,CO), respectively. Western detection kits for active, dual-phosphorylated p38 andp42/44 mitogen-activated protein (MAP) kinases were purchased from Cell Sig-naling (Beverly, MA). Antibodies to phosphotyrosine, phosphoserine, and horse-radish peroxidase-linked donkey anti-mouse secondary antibodies were purchasedfrom Zymed Laboratories (San Francisco, CA) and Accurate Chemical Corporation(Westbury, NY), respectively. WEB 2170 and WEB 2347 were the kind gifts ofBoehringer Ingelheim Pharmaceuticals (Ridgefield, CT), and A-79981.0 was ob-tained from Dr. Dan Albert (Abbott Laboratories, Abbott Park, IL). Oleoyl-acetyl-glycerol (OAG), 5-hydroxyeicosatetrenoic acid (5-HETE), and leukotriene B4
(LTB4) were purchased from Biomol. 3H-1-O-Octadecyl-sn-glycerol PC (3H–C18
lyso-PAF) was purchased from Amersham Pharmacia Biotech (Piscataway, NJ).
Lyso-PC preparationThe individual lyso-PCs and those used in the mixture were solubilized in1.25–5% essentially fatty acid-free, globulin-free, human albumin with 3-minpulses using a bath sonicator, model W-220F (Heat Systems–Ultrasonics,Plainview, NY) set at 30% maximal voltage. To determine the role of albuminas a lipid carrier, the lyso-PC mix was dissolved in 1.25% albumin, 95%ethanol, or 0.9% saline for injection, USP. The lyso-PC mixture containedpurified, individual lyso-PCs in the following molar ratios as previously pub-lished [16]: 1-O-palmitoyl:24; 1-O-oleoyl:10; 1-O-stearoyl:10; 1-O-hexadecyl(C16) lyso-PAF:0.65; and 1-O-octadecyl (C18) lyso-PAF:0.35. Other purifiedlyso-PCs, including 1-O-lauroyl, 1-O-heptadecanoyl, and 1-O-myristoyl, wereobtained to test their ability to prime the PMN oxidase.
PMN isolation and oxidase primingPMNs were isolated from whole blood drawn from healthy donors under theauspices of a protocol approved by the Colorado Multiple Institutional ReviewBoard at the University of Colorado School of Medicine (Denver). The isolationused standard techniques including dextran sedimentation, ficoll-hypaquegradient centrifugation, and hypotonic lysis of contaminating red blood cells[16]. PMN priming assays were completed as follows: PMNs (3.75x105) wereincubated with vehicle controls (saline, 1.25–5% albumin or 1% ethanol),lyso-PCs (0.045–25 �M), or 2 �M platelet-activating factor (PAF) for 5 min at37°C. The lyso-PCs and the PAF were dissolved in saline, albumin, or ethanol.After the 5-min incubation, the PMNs were activated with 1 �M fMLP or 200ng/ml phorbol myristate acetate (PMA), and the maximal rate of superoxideanion was measured as the reduction of cytochrome c at 550 nm as described[25]. Priming activity was measured as the augmentation of the maximal rateof O2
– in response to fMLP or to PMA.
Cellular association of 3H–C18 lyso-PAF and 7-nitrobenz-2-oxa-1,3�-diazol-4-yl (NBD)-labeled1-O-lauroyl lyso-PCWhole blood (20 ml) was obtained from healthy donors and mixed for 5 min at37°C, and 10 ml was incubated for 15 min with 14.5 �M lyso-PCs with 2 nM
3H–C18 lyso-PAF tracer bound to albumin. The other 10 ml whole bloodunderwent identical separation, and the cells were used to obtain cell countsusing a Coulter blood Plus IV cell counter (Beckman Coulter, Houston, TX).Following incubation, the blood was spun at 3000 g for 5 min to isolateplatelet-rich plasma, which was separated from the red cells and the leuko-cytes, and the platelets were isolated from the plasma by centrifugation [26,27]. The remaining cells underwent dextran sedimentation, which isolated themajority of the red blood cells from the leukocytes, and the supernatant wasseparated into mononuclear and polymorphonuclear leukocytes by ficoll-hypaque gradient centrifugation. All samples were centrifuged to yield cellularpellets, and the numbers of counts were determined using a tritium scintilla-tion counter. For the leukocytes and platelets, all of the cells were counted andin the case of the leukocytes, 1.0 x 107 mononuclear cells and 2.5 x 107
polymorphonuclear cells. For the red blood cells, 5 x 107 cells were counted,and the number of counts was multiplied to represent the total number of redblood cells in 10 ml whole blood. Isolated PMNs were incubated with NBD-labeled 1-O-dodecanoyl-sn-glycero-PC (10 pM–100 �M) for 60 min at 4°C,fixed with 4% paraformaldehyde, and examined by flow cytometry. Controlsconsisted of PMNs incubated with unlabeled 1-O-lauroyl lyso-PC at the twohighest concentrations used and free NBD. These experiments were repeatedthree times on PMNs isolated from healthy donors.
Inhibition of the PAF receptor
PMNs were incubated with 400 �M WEB 2170, 5 �M A-79981.0, or 10 �MWEB 2347 PAF receptor antagonists for 5 min at 37°C with gentle agitation.These PMNs were then used in priming assays with 0.45–14.5 �M lyso-PCs or2 �M PAF as described previously [16]. These concentrations were deter-mined to have maximal inhibitory capacity by performing the concentration:response relationship for each of the compounds on PAF priming of thefMLP-activated respiratory burst in isolated PMNs (results not shown).
Changes in cytosolic Ca2� concentration
Isolated PMNs (2.5�107) were loaded with indo-1 and washed, and thechanges in cytosolic Ca2� for 2 � 106 PMNs were measured at 37°C over realtime as previously reported [25, 28].
Pertussis toxin inhibition
Isolated PMNs were exposed to 2 �g/ml pertussis toxin for 2 h at 37°C and7.5% CO2 with gentle rocking. Following incubation, the PMNs were primedwith lyso-PCs, PAF, or albumin control and were activated with PMA or fMLP,and the maximal amount of superoxide anion was measured as describedpreviously. In addition, the PAF-, fMLP-, and lyso-PC-mediated changes incytosolic Ca2� were measured over real time as described previously [25, 28].
MAP kinase activation and changes in tyrosineand serine phosphorylation
Isolated PMNs (1.25x106) were exposed to 4.5–14.5 �M lyso-PC for 15 s–5min, the PMNs were lysed using Laemmli sample buffer, and the proteins wereseparated using 5–15% sodium dodecyl sulfate-polyacrylamide gel electro-phoresis (SDS-PAGE), as described previously [29]. The proteins were trans-ferred to nitrocellulose and incubated with a rabbit polyclonal antibody tophosphorylated (Tyr180/Thr182) p38 MAP kinase or a rabbit antibody tophosphorylated (Thr202/Tyr204) p42/44 MAP kinase or to a mAb to phospho-tyrosine. PAF (2 �M) was used as a positive control, and 10 �M 1,2 diolein,a diacylglycerol, was used as a negative control. In addition, control immuno-blots for total p38 and p42/44 MAP kinase were completed, which demon-strated an identical band of immunoreactivity for all treatment groups (resultsnot shown).
PMN adhesion to fibrinogen-coated microtiterplates
Fibrinogen (50 �l, 1 mg/ml) was added to individual wells of a 96-wellmicrotiter plate and was allowed to dry overnight [30, 31]. Freshly isolatedPMNs (1�106) were placed into each well and allowed to settle for 30 min. Thesettled PMNs were then stimulated with buffer control, 200 ng/ml PMA, 1 �MfMLP, 2 �M PAF, or varying concentrations of the mixture of lyso-PCs (0.045�M–15 �M) and were allowed to incubate for 30 min at 37°C. Following
512 Journal of Leukocyte Biology Volume 73, April 2003 http://www.jleukbio.org

incubation, the plates were centrifuged inverted for 5 min at 200 g, thesupernatants were discarded, and 0.1% Triton X was added to each well. Fromthese lysates, the percentage of adherent PMNs was determined by measure-ment of the cellular elastase from firmly adherent PMNs divided by the cellularelastase from the total number of PMNs added to each well. The elastase wasmeasured spectrophotometrically as described previously [32]. It is importantto note that this assay may underestimate the number of firmly adherent PMNs,as do all assays that use granule constituents as a marker of PMN presenceincluding myeloperoxidase (MPO). To authenticate the results of this assay, weperformed analogous experiments using 51Cr-loaded PMNs as described pre-viously [32]. Briefly, PMNs were incubated with 1 �Ci 51Cr/2.5 x 107 PMNs for30 min at 37°C. The PMNs were washed, resuspended in warmKrebs-Ringers-phosphate with 2% dextrose, ph 7.35 (KRPD), and incubated in fibrinogen-coated plates exactly as described above. PMN adherence was determined byusing Triton X lysis of the identical number of 51Cr-loaded cells not placed intothe wells and calculating the percentage of counts from lysates of the variouswells from the fibrinogen-coated plates as previously reported [32].
Measurement of CD11b and fMLPr surfaceexpression
The surface expression of the �2-integrin subunit CD11b and the fMLPr wasmeasured using mAb to CD11b and the fMLPr and flow cytometry, as previ-ously reported [25].
PMN chemotaxis
Directed chemotaxis of isolated PMNs was determined in a Boyden chamber asdescribed previously [33]. Directed PMN chemotaxis to differing concentra-tions of lyso-PCs was compared with directed PMN migration to zymosan-activated, normal human serum (positive control) and to Hanks’ buffered salinewith 2% albumin, pH 7.4 (negative control). Results were expressed as thedistance migrated, in microns, of the leading front of PMNs [33].
Digital microscopy
Isolated PMNs were incubated with albumin or lyso-PCs (1.45–14.5 �M) for1–10 min, fixed with 4% paraformaldehyde, and smeared onto slides. Allmanipulations with these slides, unless otherwise indicated, occurred at roomtemperature. Slides were washed three times with phosphate-buffered saline(PBS), pH 7.4, for 10 min, and the PMNs were porated with 30% acetone–70%methanol for 3 min at –20°C. After washing with PBS for 10 min, the slideswere stained for 60 min with bis-benzimide and wheat germ agglutinin (WGA)and then washed, mounted in antiquenching media, and examined for lyso-PC-induced changes in cellular morphology. PMNs from four healthy donorswere treated and processed identically and analyzed as a replicate.
Determination of elastase, lactoferrin (LF), andMPO release in isolated PMNs
PMNs (1.25�106) were warmed to 37°C in a shaking water bath and treatedwith 5 �M cytochalasin B or buffer control for 5 min. The PMNs were thenincubated for 5 min with buffer, 4.5 �M lyso-PCs, or 2 �M PAF and thenactivated with buffer, 1 �M fMLP, or 200 ng/ml PMA. After a 5-min reactiontime, the PMNs were pelleted, and the supernatants were removed. Thesesupernatants were used to determine the amount of elastase, MPO, and LFreleased. Thus, the measurements of released proteases were performed on thesame samples with the same number of cells under virtually identical condi-tions.
Elastase release was determined spectrophotometrically by the reduction ofthe specific elastase substrate methoxysuccinyl-alanyl-alanyl-prolyl-valyl p-nitroanilide (AAPVNA) at 405 nm in duplicate [32]. To ensure the reductionof AAPVNA was secondary to elastase, identical wells containing 5 �M of thespecific elastase inhibitor methoxysuccinyl-alanyl-alanyl-prolyl-valyl chloro-methyl ketone were run in conjunction with each treatment. MPO release wasdetermined by the oxidation of o-dianisidine at 405 nm as described previously[34]. LF release was quantified by a modification of a competitive enzyme-linked immunosorbent assay as described previously [35, 36]. Elastase, MPO,and LF release is reported as the percentage of total cellular elastase, MPO,and LF, as determined by 0.1% Triton X-paired treatment of an identicalnumber of PMNs.
Cytosolic Ca2� chelation with BAPTA-AM
PMNs were incubated with 50 �M BAPTA or dimethyl sulfoxide (DMSO) for30 min at 37°C as described previously [25, 29]. Following incubation, thePMNs were washed and placed into warm KRPD buffer at the identicalconcentration (2.5x107 PMNs/ml). The BAPTA concentration was determinedby measuring BAPTA inhibition of cytosolic Ca2� flux over a range ofconcentrations [25, 29]. PMNs loaded with 50 �M BAPTA did not exhibit acytosolic Ca2� in response to 2 �M PAF or 1 �M fMLP, and PMNs loaded withlesser concentrations of BAPTA, 10–25 �M, exhibited an attenuated cytosolicCa2� flux in response to these agonists (results not shown) [29].
Statistical analysis
The means, standard deviations, and SEM were calculated using standardtechniques. Statistical differences among groups were determined by a pairedor an independent ANOVA followed by a Tukey post hoc analysis for multiplecomparisons. Statistical significance was determined at the P � 0.05 level.
RESULTS
Priming PMN oxidase
As a necessary, preliminary experiment to determine if thelyso-PC mixture contained contaminating PAF or PAF analogs,the lyso-PC mix and PAF were incubated with buffer or withtwo distinct secretory phospholipase A2 (sPLA2) enzymes, Najanaja venom and bovine pancreatic, for 30 min at 37°C, pH7.35. Following this incubation, the lipids were extracted,dried, and resuspended in 1.25% human albumin. As com-pared with buffer-treated control PAF or lyso-PCs, the activityof the lyso-PCs was unaffected, and the PAF activity wasdecreased by 75 � 11%. In addition, we used a mass spec-troscopy-based assay for PAF, acetylated PAF analogs, anddetermined that no acetylated compounds were present in twoseparate lots of the lyso-PC mixture [37–39].
Lyso-PC priming of the fMLP-activated respiratory burst wasevaluated over a range of concentrations from 0.01 to 25 �M(Table 1). As compared with albumin-treated controls, thepriming activity of the lyso-PC mix, found in stored blood,became statistically significant at 0.45 �M and reached arelative maximum at 14.5 �M. This priming activity at 14.5�M was even greater than PAF-primed positive controls (lyso-PCs: 15.3�2.0 vs. 2 �M; PAF: 12.1�1.6 nmol O2
–/3.75x105
PMNs/min, n�8, P�0.05). At all concentrations of thelyso-PC mix used (0.01–25 �M), there was no evidence ofPMN lysis, and cell viability following lyso-PC incubation was99% by trypan blue exclusion, similar to controls.
The priming activity of individual lyso-PC compounds foundin stored blood and of some additional purified lyso-PC com-pounds is shown in Table 1. The C16 and C18 lyso-PAFcompounds demonstrated significant, concentration-dependentpriming of the oxidase, as compared with albumin-treatedcontrols from 1 to 10 �M (P�0.05). At higher concentrations,15–25 �M, the priming activity of the lyso-PAF compoundswas not significantly increased compared with 10 �M (Table1). Two 1-O-acyl lyso-PC compounds, 1-O-oleoyl lyso-PC and1-O-stearoyl lyso-PC, primed the fMLP-activated respiratoryburst at concentrations from 1 to 10 �M, as compared withcontrols (P�0.05). However, at 10 �M, 1-O-oleoyl lyso-PCpriming was partially diminished as compared with the activityat 4.5 �M without evidence of diminished PMN viability. In
Silliman et al. Lyso-PCs affect multiple PMN functions 513

contrast, the priming activity of 1-O-stearoyl lyso-PC contin-ued to increase over the range of concentrations until 25 �M.In addition, the activity of 1-O-palmitoyl lyso-PC was differentfrom any of the other lyso-PC moieties tested, as it significantlyprimed the fMLP-activated PMN oxidase at 4.5 �M but not at1.0 �M (Table 1). At 25 �M, 1-O-palmitoyl lyso-PC signifi-cantly primed the PMN oxidase without loss of PMN viability;however, this priming activity was no higher than concentra-tions of 4.5–10 �M. The priming activity of three other purifiedlyso-PC compounds was also evaluated. 1-O-Myristoyl and1-O-heptadecanoyl lyso-PCs did not exhibit priming activity,and the latter compound adversely affected PMN viability(Table 1). The 1-O-lauroyl lyso-PC did show modest primingactivity at concentrations of 10 and 25 �M (Table 1).
In addition, to determine if the solvent for the priming agentaffected PMN function, PAF and the lyso-PC mix were dis-solved in higher concentrations of albumin, 2.5% and 5%,95% ethanol, and aqueous solutions without albumin. Higheralbumin concentrations augmented PAF priming but did notaffect lyso-PC priming as compared with lipid-priming agentssolubilized in the 1.25% albumin (Table 2). Dissolving thelipid stocks in ethanol, final concentrations of 0.1–1%, inhib-ited fMLP activation of the oxidase by 60 � 22% to 67 � 20%,
respectively, and no lipid priming was observed (Table 2).When PAF or lyso-PCs were added to aqueous solutions with-out albumin, the solutions were turbid and inhibited the fMLPactivation of the respiratory burst at all concentrations tested.Lipids dissolved in albumin did not affect viability of PMNs;however, when lyso-PCs were solubilized in ethanol or added toaqueous solutions without a protein carrier, PMN viability wascompromised (Table 2). Lyso-PC mediated changes in cytosolic Ca2�
As many priming agents elicit changes in cytosolic Ca2�, weinvestigated the ability of lyso-PCs to induce increases incytosolic Ca2� concentration of PMNs. Lyso-PCs, the mixtureand the individual compounds, elicited rapid cytosolic Ca2�
propagation in PMNs (Fig. 1). The lyso-PC mix (0.45–14.5�M) caused a concentration-dependent rise to 500–3500 nM(Fig. 1A); however, lyso-PC concentrations less than 45 nM didnot elicit a change in cytosolic Ca2�. At 14.5 �M, lyso-PCselicited a sustained, cytosolic Ca2� flux that lasted for up to 8min and eventually returned to baseline at 12–15 min withoutaffecting cellular integrity (Fig. 1A; results not shown).
The individual lyso-PC compounds also caused rapid, con-centration-dependent increases in cytosolic Ca2� (Fig. 1,B–F). In general, all of the lyso-PC compounds tested elicitedpropagation of cytosolic Ca2� at concentrations that primed the
TABLE 1. Concentration-Dependent Lyso-PC Priming of the fMLP-Activated Respiratory Burst and Their Effects on PMN Viability
A. Priming activity of lyso-PCs in 1.25% albumin1
Viability2Concentration (�M) 0.045 0.45 1.0 4.5 10 14.5 25
Lyso-PC mix 1.2 � 0.4 3.0 � 0.5* 5.5 � 0.6* 9.1 � 0.7* 12.4 � 1.6* 15.3 � 2.0* 13.0 � 2.5* 97 � 6%C16 Lyso-PAF 1.3 � 0.2 1.3 � 0.2 4.1 � 1.1* 14.8 � 3.8* 10.2 � 0.5* 98 � 3%C18 Lyso-PAF 1.2 � 0.1 2.1 � 0.4* 4.4 � 0.3* 10.2 � 1.1* 6.5 � 0.3* 99 � 1%1-O-Palmitoyl 0.9 � 0.3 1.1 � 0.1 1.4 � 0.1 6.8 � 1.3* 5.9 � 1.1* 4.1 � 0.6* 96 � 3%1-O-Oleoyl 1.8 � 0.2 2.3 � 0.2* 2.9 � 0.4* 6.6 � 1.0* 4.0 � 0.5* 4.4 � 1.2* 97 � 2%1-O-Stearoyl 1.7 � 0.2 1.9 � 0.2 2.9 � 0.5* 7.0 � 0.8* 13.7 � 1.6* 17.1 � 1.8* 98 � 1%1-O-Lauroyl 1.7 � 0.7 2.2 � 0.6* 2.1 � 0.4* 98 � 2%1-O-Myristoyl 1.7 � 0.3 1.4 � 0.3 1.2 � 0.5 99 � 1%1-O-Heptadecanoyl 2.0 � 0.6 1.4 � 0.6 0.2 � 0.1# 65 � 14%#
PAF 12.8 � 1.2* 99 � 2%1.25% Albumin 1.7 � 0.1 1.7 � 0.1 1.7 � 0.1 1.7 � 0.1 1.7 � 0.1 1.7 � 0.1 1.7 � 0.1 99 � 1%
1 Data are expressed as the mean � SEM of the maximal rate of O2– (nmol/3.75�106 PMNs/min) in response to activation with 1 �M fMLP.2 Viability is expressed as the mean � SEM of the percentage of trypan blue-negative PMNs that were incubated for 10 min with the highest concentration of
the respective lipid. The data represent the results from eight separate PMN isolations from healthy donors.* A statistical difference (P�0.05) as compared with 1.25% albumin-primed PMNs.# A significant decrease (P�0.05) as compared with 1.25% albumin-treated control PMNs.
TABLE 2. The Effects on Lyso-PC Priming of the fMLP-Activated Respiratory by Different Solvents
Lipid priming activity in different solutions1 Viability2
Albumin Aqueous Ethanol Albumin Aqueous Ethanol
1.25% 2.5% 5.0%
Vehicle 1.8 � 0.11 1.7 � 0.3 1.8 � 0.2 2.1 � 0.3 0.9 � 0.3 99 � 2% 99 � 2% 22 � 8%Lyso-PCs (4.5 �M) 6.1 � 0.8* 6.3 � 1.2* 6.3 � 1.2* 1.9 � 1.2 1.0 � 0.5 98 � 4% 75 � 15% 78 � 20%PAF (1 �M) 9.4 � 0.5* 9.9 � 0.5* 11.7 � 0.8* 1.8 � 0.8 0.9 � 0.2 99 � 2% 57 � 12% 63 � 16%
1 Data are expressed as the mean � SEM of the maximal rate of O2– (nmol/3.75�106 PMNs/min) in response to activation with 1 �M fMLP.2 Viability is expressed as the mean � SEM of the percentage of trypan blue-negative PMNs that were incubated for 10 min with the highest concentration of
the respective lipid.* A statistical difference (P�0.05) as compared with albumin-primed PMNs. The data represent the results from four separate PMN isolations from healthy
donors.
514 Journal of Leukocyte Biology Volume 73, April 2003 http://www.jleukbio.org
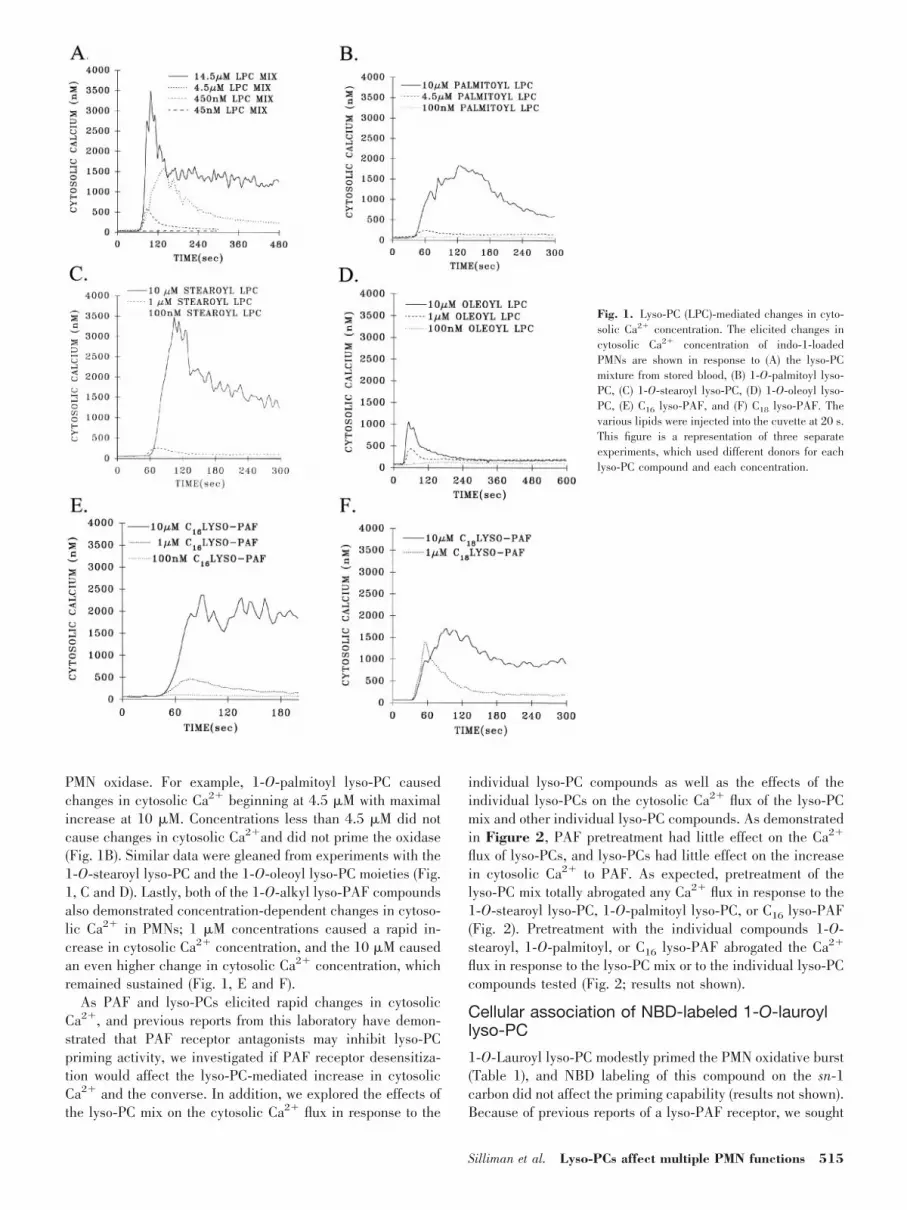
PMN oxidase. For example, 1-O-palmitoyl lyso-PC causedchanges in cytosolic Ca2� beginning at 4.5 �M with maximalincrease at 10 �M. Concentrations less than 4.5 �M did notcause changes in cytosolic Ca2�and did not prime the oxidase(Fig. 1B). Similar data were gleaned from experiments with the1-O-stearoyl lyso-PC and the 1-O-oleoyl lyso-PC moieties (Fig.1, C and D). Lastly, both of the 1-O-alkyl lyso-PAF compoundsalso demonstrated concentration-dependent changes in cytoso-lic Ca2� in PMNs; 1 �M concentrations caused a rapid in-crease in cytosolic Ca2� concentration, and the 10 �M causedan even higher change in cytosolic Ca2� concentration, whichremained sustained (Fig. 1, E and F).
As PAF and lyso-PCs elicited rapid changes in cytosolicCa2�, and previous reports from this laboratory have demon-strated that PAF receptor antagonists may inhibit lyso-PCpriming activity, we investigated if PAF receptor desensitiza-tion would affect the lyso-PC-mediated increase in cytosolicCa2� and the converse. In addition, we explored the effects ofthe lyso-PC mix on the cytosolic Ca2� flux in response to the
individual lyso-PC compounds as well as the effects of theindividual lyso-PCs on the cytosolic Ca2� flux of the lyso-PCmix and other individual lyso-PC compounds. As demonstratedin Figure 2, PAF pretreatment had little effect on the Ca2�
flux of lyso-PCs, and lyso-PCs had little effect on the increasein cytosolic Ca2� to PAF. As expected, pretreatment of thelyso-PC mix totally abrogated any Ca2� flux in response to the1-O-stearoyl lyso-PC, 1-O-palmitoyl lyso-PC, or C16 lyso-PAF(Fig. 2). Pretreatment with the individual compounds 1-O-stearoyl, 1-O-palmitoyl, or C16 lyso-PAF abrogated the Ca2�
flux in response to the lyso-PC mix or to the individual lyso-PCcompounds tested (Fig. 2; results not shown).
Cellular association of NBD-labeled 1-O-lauroyllyso-PC
1-O-Lauroyl lyso-PC modestly primed the PMN oxidative burst(Table 1), and NBD labeling of this compound on the sn-1carbon did not affect the priming capability (results not shown).Because of previous reports of a lyso-PAF receptor, we sought
Fig. 1. Lyso-PC (LPC)-mediated changes in cyto-solic Ca2� concentration. The elicited changes incytosolic Ca2� concentration of indo-1-loadedPMNs are shown in response to (A) the lyso-PCmixture from stored blood, (B) 1-O-palmitoyl lyso-PC, (C) 1-O-stearoyl lyso-PC, (D) 1-O-oleoyl lyso-PC, (E) C16 lyso-PAF, and (F) C18 lyso-PAF. Thevarious lipids were injected into the cuvette at 20 s.This figure is a representation of three separateexperiments, which used different donors for eachlyso-PC compound and each concentration.
Silliman et al. Lyso-PCs affect multiple PMN functions 515

to determine if there was a cellular association of lyso-PCs withintact cells by flow cytometry and digital microscopy [22] (Fig.3). PMNs were incubated with differing concentrations (104–1014 M) of NBD-labeled 1-O-lauroyl lyso-PC for 60 min at4°C with 100 �M-unlabeled 1-O-lauroyl lyso-PC and free NBDas controls. Albumin-treated PMNs and PMNs treated with 100�M-unlabeled 1-O-lauroyl lyso-PC did not demonstrate anydifference in cellular mean fluorescence intensity (MFI;9.9�0.4 vs. 10.2�0.5). As compared with controls, incubationof PMNs with NBD-labeled 1-O-lauroyl lyso-PC demonstrateda shift in the MFI from 10.2 � 0.5 to 604 � 47 at 100 �M and99% of the cells marked for the labeled lyso-PC ligand. As theconcentration of labeled 1-O-lauroyl lyso-PC was decreased,there was a reduction in the shift with return to baselinefluorescence at 1 nM (Fig. 3; results no shown). Moreover,samples of the identical PMNs used in the flow cytometryexperiments were also examined by digital microscopy. PMNsevidence a brighter green color in the cellular membrane whenconcentrations of NBD-labeled 1-O-lauroyl lyso-PC were
added, especially in contrast to the control PMNs treated with100 �M-unlabeled 1-O-lauroyl lyso-PC (Fig. 3).
To ensure that the labeling of PMNs with lyso-PCs was nota result of nonspecific lipid—membrane interactions—we usedtwo separate strategies. The first incubated human whole bloodwith 3H–C18 lyso-PAF for 15 min at 37°C followed by isolationof the different cellular fractions, including platelets, red bloodcells, mononuclear leukocytes, and polymorphonuclear leuko-cytes. These cellular fractions were then assayed for 3H countsto determine if a selective lyso-PAF association might exist.These results demonstrated that the majority of the 3Hcounts was in the acellular plasma (97.4�1.2%). Of thecell-associated counts, the platelet fraction had 70.5 � 4%,the mononuclear leukocytes had 15.5 � 1.4%, and thepolymorphonuclear leukocytes, which include the eosino-phils and basophils, had 14.0 � 1.3%. The red blood cellshad less than 0.01% of the total cellular-associated counts(Table 3). Second, when unlabeled 1-O-lauroyl lyso-PC wasadded concomitantly with the NBD-labeled 1-O-lauroyl
Fig. 2. Receptor desensitization. The changes incytosolic Ca2� were determined in indo-1-loadedPMNs in a dual-wavelength spectrofluorimeter overreal time. PAF (2 �M) and lyso-PCs (4.5 �M)elicited changes in cytosolic Ca2� concentrationdespite pretreatment with one another (A and B).Pretreatment of PMNs with the lyso-PC mix (4.5�M) inhibited the Ca2� response of C16 lyso-PAF(C). Pretreatment with 1 �M 1-O-palmitoyl (D), 1�M 1-O-stearoyl (E), or 1 �M C16 lyso-PAF (F)inhibited the Ca2� response to the lyso-PC (L-PC)mix or C16 lyso-PAF. The figure represents the datafrom at least three different experiments using dif-ferent, healthy donors for each lyso-PC or PAFtreatment.
516 Journal of Leukocyte Biology Volume 73, April 2003 http://www.jleukbio.org

lyso-PC, the magnitude of the shift in MFI was decreased ina concentration-dependent manner, and the green color wasalso decreased by digital microscopy (Fig. 3B; results notshown).
PAF receptor antagonism and pertussis toxininhibition
Our previous studies documented inhibition of lyso-PC priming bya PAF receptor antagonist WEB 2170 [16]. Additional studieswere completed using WEB 2170 (400 �M), WEB 2347 (10 �M),a highly selective PAF receptor antagonist, and A-79981.0 (4�M), a PAF receptor antagonist unrelated to those of the WEBseries [40–42]. WEB 2170 and A-79981.0 partially inhibited thelyso-PC mix (4.5 �M) priming of the fMLP-activated respiratoryburst by 49.6 � 12% and 42.3 � 8% and PAF, by 92 � 7% and82 � 6%, respectively (n�7, P�0.05). However, pretreatment ofPMN with WEB 2347 inhibited PAF priming of the oxidase by98 � 3% but did not show a significant diminution in lyso-PC
priming of the fMLP-activated respiratory burst (n�5, P�0.05).As WEB 2170 partially inhibited lyso-PC priming, we investi-gated its effects on other lipids, which rapidly prime the PMNoxidase, including LTB4, OAG, and 5-HETE. Surprisingly, WEB2170 inhibited priming of the fMLP-activated respiratory burst ofLTB4 by 75.5 � 5% and OAG, by 48 � 8% and did not affect5-HETE priming of the fMLP-activated respiratory burst, 8 � 6%(n�6, P�0.05 for OAG and LTB4; Table 4).
As a lyso-PC receptor has been reported on HL-60 cells andmature granulocytes, we hypothesized that this receptor may be
Fig. 3. Cellular association of NBD-labeled 1-O-lauroyl lyso-PC. Albumin (Alb)-treated PMNs were incubated with differingconcentrations of NBD-labeled 1-O-lauroyl lyso-PC for 60 minat 4°C. The PMNs were fixed and examined by digital micros-copy (A, upper) and flow cytometry (A, lower) and digitalmicroscopy (B). (A) The green cellular fluorescence, visualizedby digital microscopy, which is directly related to the concen-tration of labeled lyso-PC added as quantitated by flow cytom-etry in the lower portion of the panel. (B) The results of PMNsincubated for 60 min with 104 M NBD-labeled 1-O-lauroyllyso-PC alone (left) or the identical PMNs incubated concom-itantly with 104 M-unlabeled 1-O-lauroyl lyso-PC � 104 M
NBD-labeled 1-O-lauroyl lyso-PC. This figure is representative of three separate experiments using healthy donors.
TABLE 3. Plasma and Cellular Association of 3H-C16
Lyso-PAF Incubations of Whole Blood
A. Distribution of 3H–C16 lyso-PAFSource % Counts
Plasma 97.4 � 1.2*Blood cells 2.5 � 0.4
B. Cellular distribution of 3H–C16 lyso-PAFCell type % Counts
Platelets 70.5 � 4.0Mononuclear leukocytes 15.5 � 1.4Granulocytes 14.0 � 1.3Red blood cells �0.02
* Data are expressed as the mean � SEM. The data presented are the resultof two separate experiments on whole blood from different, healthy donors.
TABLE 4. Lipid Receptor and Pertussis ToxinInhibition of Lyso-PC Priming
A. PAF receptor antagonismControl WEB 2170 WEB 2374 A-79981.0
fMLP 1.8 � 0.2* 1.6 � 0.3 1.8 � 0.3 1.7 � 0.34.5 �M Lyso-
PCs/fMLP 9.4 � 1.2† 5.4 � 1.3# 8.9 � 1.0 5.5 � 1.5#
14.5 �M Lyso-PCs/fMLP 13.5 � 1.7† 10.9 � 1.2# 14.1 � 1.1 11.4 � 1.6#
PAF/fMLP 12.5 � 2.1† 2.0 � 0.6# 1.9 � 0.3# 2.5 � 1.0#
5-HETE/fMLP 2.4 � 0.6† 2.3 � 0.5OAG/fMLP 4.2 � 0.6† 2.5 � 0.3#
LTB4/fMLP 5.0 � 1.0† 2.4 � 0.3#
B. Pertussis toxin inhibitionControl Pertussis toxin
PMA 5.6 � 0.9 5.8 � 1.2Lyso-PCs/PMA 8.6 � 1.5† 6.6 � 1.5#
PAF/PMA 10.5 � 2.0 10.9 � 2.1
* The data (nmol/3.75�105 PMNs/min) are expressed as the mean � SEM
(n�5). Abbreviations used: fMLP, 1 �M; Lyso-PCs, 4.5 �M; PAF, 2 �M;5-HETE, 1 �M; OAG, 1 �M; LTB4, 100 nM; PMA, 200 ng/ml. † P � 0.05as compared with albumin-primed PMNs activated with fMLP; # P � 0.05 ascompared with lipid-primed PMNs activated with fMLP (A) or with PMA (B).
Silliman et al. Lyso-PCs affect multiple PMN functions 517
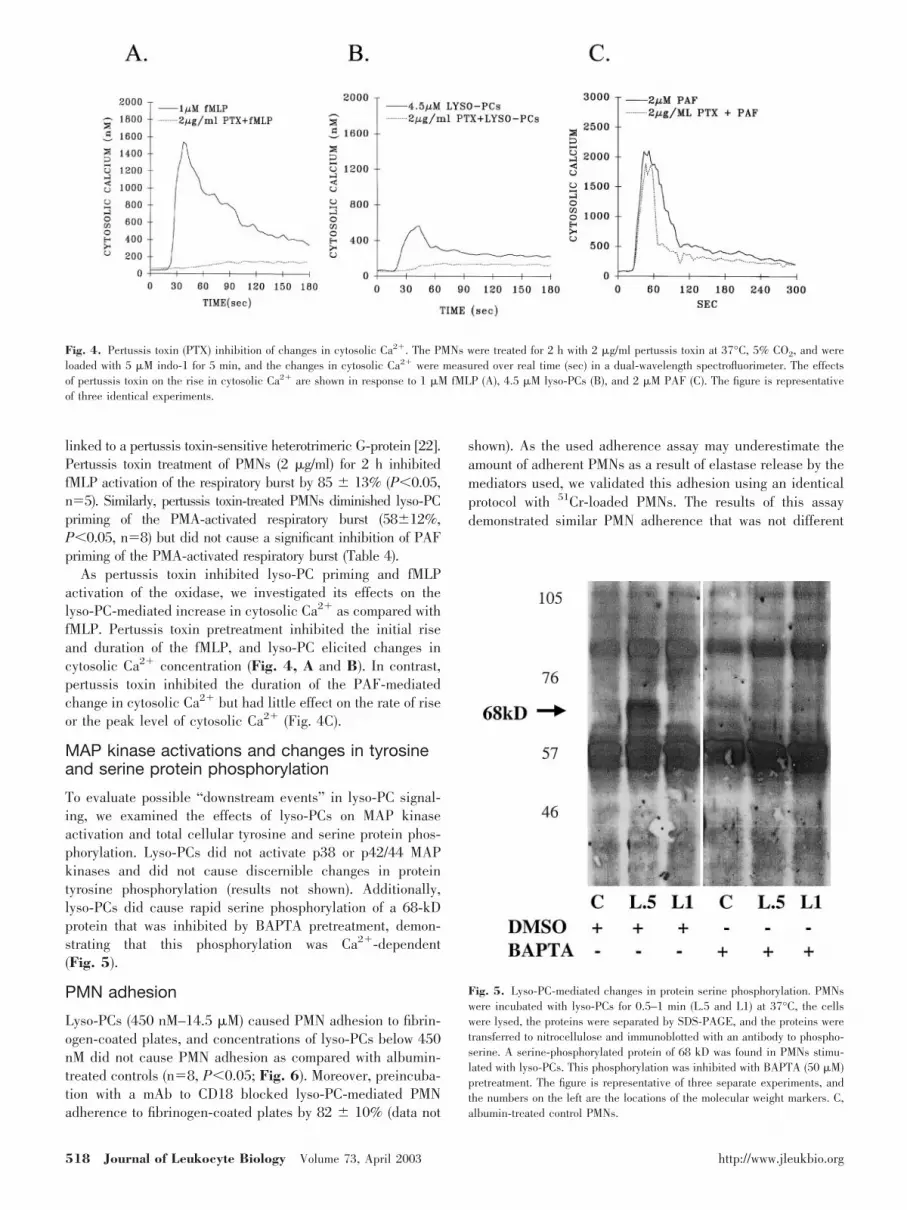
linked to a pertussis toxin-sensitive heterotrimeric G-protein [22].Pertussis toxin treatment of PMNs (2 �g/ml) for 2 h inhibitedfMLP activation of the respiratory burst by 85 � 13% (P�0.05,n�5). Similarly, pertussis toxin-treated PMNs diminished lyso-PCpriming of the PMA-activated respiratory burst (58�12%,P�0.05, n�8) but did not cause a significant inhibition of PAFpriming of the PMA-activated respiratory burst (Table 4).
As pertussis toxin inhibited lyso-PC priming and fMLPactivation of the oxidase, we investigated its effects on thelyso-PC-mediated increase in cytosolic Ca2� as compared withfMLP. Pertussis toxin pretreatment inhibited the initial riseand duration of the fMLP, and lyso-PC elicited changes incytosolic Ca2� concentration (Fig. 4, A and B). In contrast,pertussis toxin inhibited the duration of the PAF-mediatedchange in cytosolic Ca2� but had little effect on the rate of riseor the peak level of cytosolic Ca2� (Fig. 4C).
MAP kinase activations and changes in tyrosineand serine protein phosphorylation
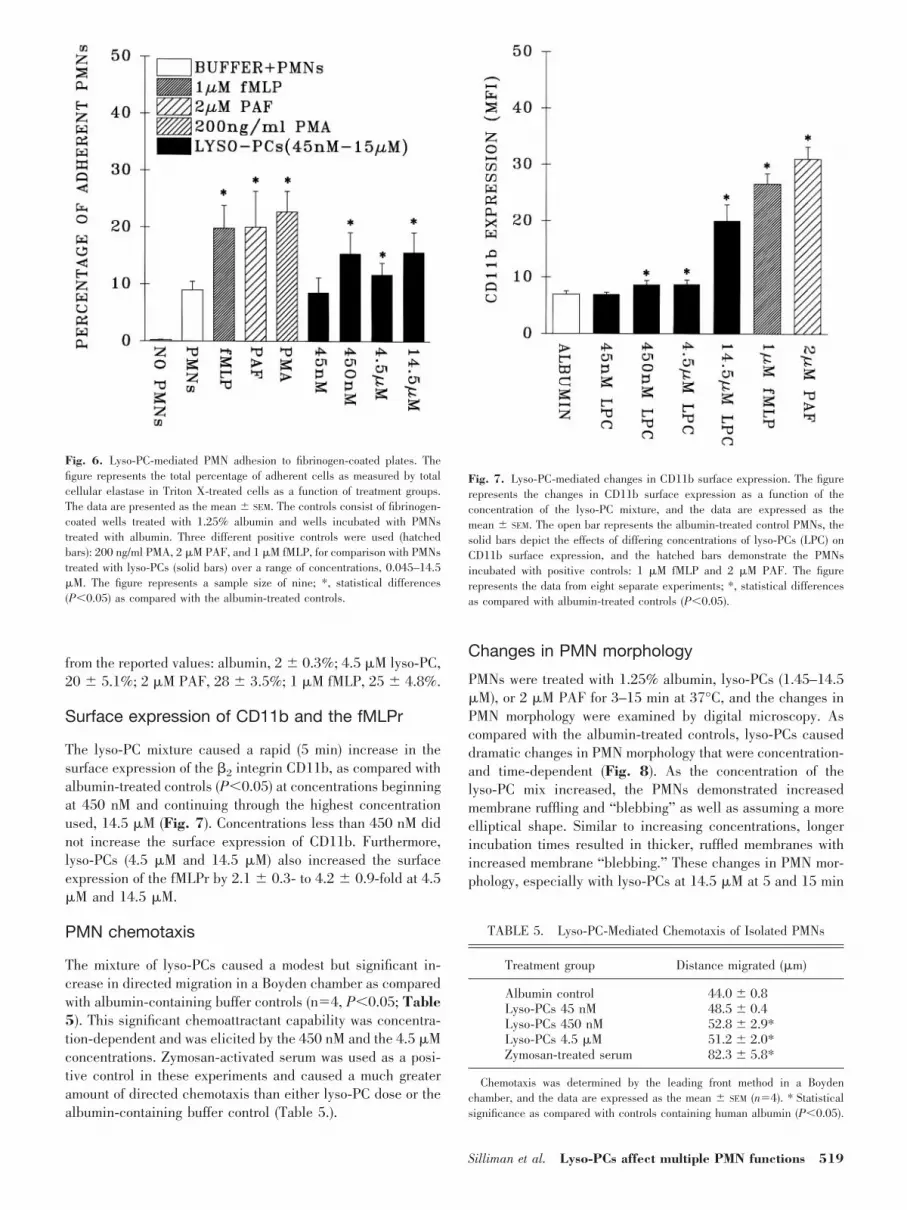
To evaluate possible “downstream events” in lyso-PC signal-ing, we examined the effects of lyso-PCs on MAP kinaseactivation and total cellular tyrosine and serine protein phos-phorylation. Lyso-PCs did not activate p38 or p42/44 MAPkinases and did not cause discernible changes in proteintyrosine phosphorylation (results not shown). Additionally,lyso-PCs did cause rapid serine phosphorylation of a 68-kDprotein that was inhibited by BAPTA pretreatment, demon-strating that this phosphorylation was Ca2�-dependent(Fig. 5).
PMN adhesion
Lyso-PCs (450 nM–14.5 �M) caused PMN adhesion to fibrin-ogen-coated plates, and concentrations of lyso-PCs below 450nM did not cause PMN adhesion as compared with albumin-treated controls (n�8, P�0.05; Fig. 6). Moreover, preincuba-tion with a mAb to CD18 blocked lyso-PC-mediated PMNadherence to fibrinogen-coated plates by 82 � 10% (data not
shown). As the used adherence assay may underestimate theamount of adherent PMNs as a result of elastase release by themediators used, we validated this adhesion using an identicalprotocol with 51Cr-loaded PMNs. The results of this assaydemonstrated similar PMN adherence that was not different
Fig. 4. Pertussis toxin (PTX) inhibition of changes in cytosolic Ca2�. The PMNs were treated for 2 h with 2 �g/ml pertussis toxin at 37°C, 5% CO2, and wereloaded with 5 �M indo-1 for 5 min, and the changes in cytosolic Ca2� were measured over real time (sec) in a dual-wavelength spectrofluorimeter. The effectsof pertussis toxin on the rise in cytosolic Ca2� are shown in response to 1 �M fMLP (A), 4.5 �M lyso-PCs (B), and 2 �M PAF (C). The figure is representativeof three identical experiments.
Fig. 5. Lyso-PC-mediated changes in protein serine phosphorylation. PMNswere incubated with lyso-PCs for 0.5–1 min (L.5 and L1) at 37°C, the cellswere lysed, the proteins were separated by SDS-PAGE, and the proteins weretransferred to nitrocellulose and immunoblotted with an antibody to phospho-serine. A serine-phosphorylated protein of 68 kD was found in PMNs stimu-lated with lyso-PCs. This phosphorylation was inhibited with BAPTA (50 �M)pretreatment. The figure is representative of three separate experiments, andthe numbers on the left are the locations of the molecular weight markers. C,albumin-treated control PMNs.
518 Journal of Leukocyte Biology Volume 73, April 2003 http://www.jleukbio.org
from the reported values: albumin, 2 � 0.3%; 4.5 �M lyso-PC,20 � 5.1%; 2 �M PAF, 28 � 3.5%; 1 �M fMLP, 25 � 4.8%.
Surface expression of CD11b and the fMLPr
The lyso-PC mixture caused a rapid (5 min) increase in thesurface expression of the �2 integrin CD11b, as compared withalbumin-treated controls (P�0.05) at concentrations beginningat 450 nM and continuing through the highest concentrationused, 14.5 �M (Fig. 7). Concentrations less than 450 nM didnot increase the surface expression of CD11b. Furthermore,lyso-PCs (4.5 �M and 14.5 �M) also increased the surfaceexpression of the fMLPr by 2.1 � 0.3- to 4.2 � 0.9-fold at 4.5�M and 14.5 �M.
PMN chemotaxis
The mixture of lyso-PCs caused a modest but significant in-crease in directed migration in a Boyden chamber as comparedwith albumin-containing buffer controls (n�4, P�0.05; Table5). This significant chemoattractant capability was concentra-tion-dependent and was elicited by the 450 nM and the 4.5 �Mconcentrations. Zymosan-activated serum was used as a posi-tive control in these experiments and caused a much greateramount of directed chemotaxis than either lyso-PC dose or thealbumin-containing buffer control (Table 5.).
Changes in PMN morphology
PMNs were treated with 1.25% albumin, lyso-PCs (1.45–14.5�M), or 2 �M PAF for 3–15 min at 37°C, and the changes inPMN morphology were examined by digital microscopy. Ascompared with the albumin-treated controls, lyso-PCs causeddramatic changes in PMN morphology that were concentration-and time-dependent (Fig. 8). As the concentration of thelyso-PC mix increased, the PMNs demonstrated increasedmembrane ruffling and “blebbing” as well as assuming a moreelliptical shape. Similar to increasing concentrations, longerincubation times resulted in thicker, ruffled membranes withincreased membrane “blebbing.” These changes in PMN mor-phology, especially with lyso-PCs at 14.5 �M at 5 and 15 min
Fig. 6. Lyso-PC-mediated PMN adhesion to fibrinogen-coated plates. Thefigure represents the total percentage of adherent cells as measured by totalcellular elastase in Triton X-treated cells as a function of treatment groups.The data are presented as the mean � SEM. The controls consist of fibrinogen-coated wells treated with 1.25% albumin and wells incubated with PMNstreated with albumin. Three different positive controls were used (hatchedbars): 200 ng/ml PMA, 2 �M PAF, and 1 �M fMLP, for comparison with PMNstreated with lyso-PCs (solid bars) over a range of concentrations, 0.045–14.5�M. The figure represents a sample size of nine; *, statistical differences(P�0.05) as compared with the albumin-treated controls.
Fig. 7. Lyso-PC-mediated changes in CD11b surface expression. The figurerepresents the changes in CD11b surface expression as a function of theconcentration of the lyso-PC mixture, and the data are expressed as themean � SEM. The open bar represents the albumin-treated control PMNs, thesolid bars depict the effects of differing concentrations of lyso-PCs (LPC) onCD11b surface expression, and the hatched bars demonstrate the PMNsincubated with positive controls: 1 �M fMLP and 2 �M PAF. The figurerepresents the data from eight separate experiments; *, statistical differencesas compared with albumin-treated controls (P�0.05).
TABLE 5. Lyso-PC-Mediated Chemotaxis of Isolated PMNs
Treatment group Distance migrated (�m)
Albumin control 44.0 � 0.8Lyso-PCs 45 nM 48.5 � 0.4Lyso-PCs 450 nM 52.8 � 2.9*Lyso-PCs 4.5 �M 51.2 � 2.0*Zymosan-treated serum 82.3 � 5.8*
Chemotaxis was determined by the leading front method in a Boydenchamber, and the data are expressed as the mean � SEM (n�4). * Statisticalsignificance as compared with controls containing human albumin (P�0.05).
Silliman et al. Lyso-PCs affect multiple PMN functions 519

of incubation, were very similar to the changes in PMN shapeelicited by 2 �M PAF.
PMN degranulation
Lyso-PCs caused a direct increase in elastase release as com-pared with buffer-treated, control PMNs, similar to the amountof elastase released in response to 2 �M PAF, 1 �M fMLP, and200 ng/ml PMA (Fig. 9A). However, pretreatment of PMNswith cytochalasin B did not augment the amount of lyso-PC-induced elastase release but did augment the amounts ofelastase released by PAF, fMLP, and PMA. Furthermore, PAFand lyso-PC pretreatment did increase the amount of elastasereleased in response to fMLP, i.e., primed the fMLP response,as compared with PMNs treated with PAF, fMLP, or lyso-PCsalone (n�7, P�0.05).
The results obtained for MPO release from isolated PMNswere similar to that from elastase release (Fig. 9B). Lyso-PCselicited significant amounts of MPO release, as compared withalbumin-pretreated controls, similar to the amount of MPOreleased by 2 �M PAF or 200 ng/ml PMA (Fig. 4B). Moreover,cytochalasin B pretreatment did not significantly augment theamount of MPO released by lyso-PCs but did increase theamount released by PAF, PMA, and 1 �M fMLP (n�7,P�0.05). In addition, lyso-PC pretreatment significantly in-creased the fMLP-mediated degranulation of MPO as com-pared with fMLP or lyso-PCs alone (P�0.05). PAF priming offMLP-mediated MPO release was not different from the amountof MPO released by PAF itself.
LF release from specific granules was also quantified, andwe demonstrated that lyso-PC elicited a direct, reproduciblerelease of LF from PMNs, similar to PAF-, fMLP-, and PMA-mediated LF release and significantly greater than buffer-treated controls (n�7, P�0.05; Fig. 9C). Pretreatment withcytochalasin B significantly enhanced the amount of LF releaseto fMLP, PAF, and PMA. In addition, lyso-PC and PAF pre-
Fig. 8. Lyso-PC-mediated changes in PMN morphology. PMNs were incu-bated with albumin (control) lyso-PCs (1.45–14.5 �M) and PAF (2 �M) for3–15 min at 37°C. Immediately at the end of the incubation period, the PMNswere fixed with paraformaldehyde and then doubly stained with a blue nuclearstain (bis-benzamide) and a green membrane stain (WGA). The PMNs wereexamined at 100x (original magnification), and the bar in the first control panelis 5 �m (original magnification). This figure is representative of five separateexperiments.
Fig. 9. Lyso-PC-mediated release of PMN granule constituents. (A–C) Elastase, MPO, and LF release from isolated PMNs as a function of treatment group, andthe data are represented as the mean � SEM. The open bars are DMSO-pretreated controls (pre-TX), the solid bars represent PMNs pretreated with 5 �Mcytochalasin B for 30 min at 37°C, and the hatched bars represent PMNs primed with 4.5 �M lyso-PC (LPC) mix or 2 �M PAF for 5 min followed by activationwith 1 �M fMLP. *, Statistical significance as compared with albumin-treated, control PMNs (P�0.05). �, Statistical significance (P�0.05) in cytochalasinB-pretreated PMNs as compared with paired DMSO-pretreated PMNs. #, Statistical significance (P�0.05) of lyso-PC- or PAF-pretreated PMNs activated with fMLPas compared with lyso-PC-, PAF-, or fMLP-treated PMNs. The figure represents the results of seven separate experiments.
520 Journal of Leukocyte Biology Volume 73, April 2003 http://www.jleukbio.org

treatment did not enhance the amount of fMLP-induced LFrelease.
BAPTA inhibition of lyso-PC activity
As changes in cytosolic Ca2� were elicited by lyso-PC primingof the PMN oxidase, we examined the effects of cytosolic Ca2�
chelation with BAPTA (50 �M), which buffers cytosolic Ca2�,rendering it biologically unavailable, regardless of the Ca2�
source [42]. PMNs were pretreated with 50 �M BAPTA orDMSO, primed with 1.25% albumin, lyso-PCs (4.5 �M or 14.5�M), or PAF (2 �M) for 5 min, and then activated with 200ng/ml PMA. BAPTA inhibited the lyso-PC priming by 85 �6% and 88 � 7%, respectively, and inhibited PAF priming by67 � 12%. In addition, BAPTA chelation inhibited lyso-PC-mediated increases in CD11b expression by 52 � 11% andchanges in PMN morphology (results not shown).
DISCUSSION
Although previous data from a number of laboratories havedocumented that lyso-PCs, especially lyso-PAF, do not stimu-late PMNs, this study has demonstrated that albumin wasrequired as a carrier, and concentrations �450 nM must beused for PMN stimulation [20–24, 43]. Moreover, the additionof these compounds to fresh, human plasma resulted in primingof the PMN oxidase by 1.7 � 0.2-fold as compared withalbumin-pretreated controls [16]. Recent studies have demon-strated that lyso-PCs not only rapidly prime human PMNs butalso rapidly prime PMNs from healthy rats [19]. Thus, lyso-PCs, solubilized in albumin solutions, effectively prime humanand rodent PMNs at physiologically relevant concentrations inthe context of blood transfusions [16].
The initial observations that a mixture of lyso-PCs primesthe reduced nicotinamide adenine dinucleotide phosphate(NADPH) oxidase have been extended by documentation of arange of priming concentrations (three logs); by demonstrationof the activity of individual lyso-PC compounds and identifi-cation of the requirement of albumin as a carrier for PMNstimulation, identical to human plasma; and by explanation ofconflicting reports that deemed lyso-PCs inactive as a result ofthe use of suboptimal concentrations or the deletion of thealbumin carrier [16, 19–24, 43–47]. These experiments alsodemonstrated that PAF required an albumin carrier for activ-ity, as reported previously, and that the activity of lyso-PCs wasnot affected by incubation with sPLA2, an enzyme that inacti-vates the activity of PAF [48]. With respect to the primingactivity of the individual lyso-PC compounds, the 1-O-alkyllyso-PAF moieties and the 1-O-stearoyl lyso-PC showed in-creasing augmentation of the fMLP-activated respiratory burstat concentrations greater than 4.5 �M, similar to the lyso-PCmixture. The other two acyl compounds, 1-O-palmitoyl and1-O-oleoyl, did cause rapid priming of the NADPH oxidase,but their activity did not increase above the levels demon-strated at 4.5 �M. In addition, the concentrations of thelyso-PCs, the individual species and the mix, which inducedchanges in cytosolic Ca2�, parallel those concentrations thatprime the NADPH oxidase. Thus, the lyso-PC concentrations,
which elicited a rapid rise in cytosolic Ca2�, also primed theNADPH oxidase. It is also important to note that the highestconcentrations of lyso-PCs used in these experiments, includ-ing the lyso-PC mix and the individual species, did not com-promise PMN cellular integrity. Furthermore, the activity of thelyso-PC mixture was similar to the activity of the three mosteffective, individual lyso-PC compounds, 1-O-stearoyl lyso-PC, C16 lyso-PAF, and C18 lyso-PAF; however, the two lyso-PAF compounds comprise only 100–400 nM of the totalconcentration of the lyso-PC mixture at concentrations rangingfrom 4.5 to 15 �M, and the 1-O-stearoyl lyso-PC accounts foran additional 1–3.5 �M of the total. The 1-O-oleoyl and1-O-palmitoyl lyso-PC compounds did display activity thatshould not be discounted, as there may be some additive orsynergistic activity among the lyso-PCs that comprise the mix-ture and influence its relative activity, and further experimen-tation is required to elucidate the relative participation of theindividual species that comprise the mixture.
Lyso-PCs demonstrated selective association with bloodcells, mononuclear leukocytes, polymorphonuclear leukocytes,and platelets, which are affected by lipid stimuli. If such a cellmembrane association were nonspecific, there should havebeen 3H–C18 lyso-PAF associated with red blood cells. More-over, one would have expected that more of the total radioac-tive lyso-PC tracer would have become cell-associated, butvirtually 97% was free in the plasma bound to an albumincarrier. A concentration-dependent membrane association ofan active NBD-labeled 1-O-lauroyl lyso-PC provided support-ive evidence of the existence of a lyso-PC receptor on the PMNsurface [22]. In addition, receptor desensitization studiesshowed that when PMNs were desensitized to PAF, lyso-PCscould still cause an increase in cytosolic Ca2�, and whenPMNs were desensitized to lyso-PCs, the PAF response wasalso not affected. Such results are not surprising, as we havepreviously shown that lyso-PCs could activate the oxidase ofPAF-primed but not vehicle-treated PMNs, and PAF couldactivate the oxidase of lyso-PC-primed PMNs but not vehicle-treated controls [49]. Moreover, pertussis toxin dramaticallyaffected lyso-PC-induced changes in cytosolic Ca2� and prim-ing of the oxidase, whereas pertussis toxin had little effect onPAF-mediated changes in cytosolic Ca2� and PMN priming.Taken together with WEB 2347 data, these results suggest thatPAF and lyso-PCs activate PMNs through disparate receptors,and the observed activity was not a result of contamination ofthe purified lipid by acetylated PAF analogs.
The cytosolic Ca2� desensitization studies demonstratedthat the individual lyso-PC compounds, 1-O-alkyl and 1-O-acyl, could inhibit the activity of the lyso-PC mix and otherindividual moieties. These results implicate a receptor thatrecognizes 1-O-alky and 1-O-acyl lyso-PC analogs, much likethe PAF receptor and chemokine receptors [50–52]. In addi-tion, pertussis toxin inhibition of the lyso-PC priming impli-cates a receptor-linked heterotrimeric G-protein in the signal-ing pathway. Downstream from the pertussis toxin-sensitiveG-protein, lyso-PCs caused rapid, concentration-dependent in-creases in cytosolic Ca2� concentration that were linked to anincrease in serine phosphorylation of a 68-kD protein. Chela-tion of cytosolic Ca2� in intact PMNs abrogated lyso-PC-mediated effects in PMNs, including priming the PMA-acti-
Silliman et al. Lyso-PCs affect multiple PMN functions 521

vated respiratory burst, increased surface expression ofCD11b, changes in PMN morphology, and serine phosphory-lation of a 68-kD protein. However, unlike most chemoattrac-tants, lyso-PCs did not cause changes in whole-cell proteintyrosine phosphorylation or activation of MAP kinases.
The lyso-PC mixture directly caused changes in multiplePMN functions. At concentrations �0.45 �M, lyso-PCs elic-ited firm adhesion of PMNs to the RGD ligands inherent tofibrinogen, without significant differences among the effectivelyso-PC concentrations (0.45–14.5 �M) [53]. As the adhesionassay used subjects the adherent PMNs to 200 g, only firmlyadherent PMNs remain attached to the Arg-Gly-Asp ligandsvia �2 integrins [32]. Such a stringent assay does not invitecomparisons with other adhesion assays that subject adherentPMNs to substantially less detachment force. The lyso-PCmixture also directly caused concentration-dependent in-creases in surface expression of CD11b and the fMLPr, similarto a number of PMN-priming agents [15, 25, 54, 55]. Moreover,the lyso-PC-mediated changes in chemotaxis were modest ascompared with zymosan-activated serum but were statisticallysignificant as compared with the albumin-treated controls.Furthermore, lyso-PC-induced changes in cellular morphologywere similar to those of PAF, and such morphologic changesare typical of chemotactic agents that cause increases in F-actin [56–58]. In addition, the lyso-PC mix also elicited rapid(5 min) degranulation of elastase, MPO, and LF, similar toother agonists including PAF. The lyso-PC mixture also aug-mented the amount of elastase and MPO released by fMLP;however, cytochalasin B pretreatment did not significantlyaffect lyso-PC-mediated degranulation, the significance ofwhich is not entirely clear. Thus, lyso-PC treatment resulted inthe direct release of the contents from azurophilic and specificgranules and was able to augment the fMLP-mediated releaseof contents from primary granules.
Recent work has delineated the importance of lyso-PCs andother lysophospholipids in diverse organisms from yeast tohumans [59]. A “subfamily of orphan receptors recognize lyso-PCs and other lysophospholipids and are G-protein-linkedcellular receptors present on fibroblasts, ovarian cancer cells,and leukocytes” [59]. Although their roles in human diseaseare poorly characterized, there appears to be a linkage amonglyso-PCs, their receptors, and autoimmunity [60, 61]. This lineof research is in its infancy, and much remains unknown at thepresent time; however, some years ago, a receptor that recog-nized two types of alkyl lyso-PAF, the C16 and C18, waslocalized on the surface of PMNs [22, 59]. Although the pres-ence of this receptor may provide an attractive candidate forthe lyso-PC stimulation of PMNs, these studies were marred bythe use of ethanol as the lipid solvent and lyso-PC concentra-tions not exceeding 100 nM [22]. Thus, it was of little surprisethat the authors were unable to find lyso-PC-mediated changesin PMN function despite receptor occupancy [22]. Moreover,there are no data regarding the activation of PMNs through thisreceptor, its avidity for the 1-O-acyl lyso-PC compounds, norits antagonism by PAF receptor antagonists [22].
In conclusion, lyso-PCs affect multiple PMN functions in aconcentration-dependent manner. The relatively high concen-trations used appear to have physiologic significance, becauseof the millimolar concentrations generated during the routine
storage of blood [16]. Moreover, lyso-PCs have been reported tocause activation of vascular endothelium, an increase in adhe-sion molecule expression, and adherence of PMNs to theseactivated cells via a protein kinase C-dependent mechanism[62–65]. Thus, previous work has documented the activity oflyso-PCs in other primary cells or cell lines [62–65]. Recentfindings from our laboratory have documented that lyso-PCsmay serve as one of two insults in a two-event animal model ofacute lung injury, and lyso-PCs have been implicated in clin-ical cases of transfusion-related, acute lung injury [18, 19].Thus, lyso-PCs may not only affect individual cells in vivo orin vitro, e.g., PMNs or vascular endothelium, but also have thecapacity to influence the physiology of the lung.
ACKNOWLEDGMENTS
This work was supported by Bonfils Blood Center, The MargeryWilson Transfusion Medicine Award, The Stacy True MemorialTrust, a Clinical Associate Physician Award (M01-RR00069)from the General Clinical Research Centers Program, NationalCenters for Research Resources, NIH, a grant from The Na-tional Blood Foundation, a Transfusion Medicine AcademicAward #K07-HL02036, NHLBI, NIH, and grant #HL59355from NHLBI, NIH. The authors thank Keith Refior and PatrickMurphy from the Department of Information Services, BonfilsBlood Center, for preparation of the color figures in this manu-script.
REFERENCES
1. Babior, B. M., Kipnes, R. S., Curnutte, J. T. (1973) Biological defensemechanisms. The production by leukocytes of superoxide, a potentialbactericidal agent. J. Clin. Invest. 52, 741–744.
2. Sbarra, A. J., Karnovsky, M. L. (1959) The biological basis of phagocytosisI. Metabolic changes during the ingestion of particles of polymorphonu-clear leukocytes. J. Biol. Chem. 234, 1355–1362.
3. Albelda, S. M., Smith, C. W., Ward, P. A. (1994) Adhesion molecules andinflammatory injury. FASEB J. 8, 504–512.
4. Carlos, T. M., Harlan, J. M. (1994) Leukocyte-endothelial adhesion mol-ecules. Blood 84, 2068–2101.
5. Granger, D. N., Kubes, P. (1994) The microcirculation and inflammation:modulation of leukocyte- endothelial cell adhesion. J. Leukoc. Biol. 55,662–675.
6. Newman, P. J. (1997) The biology of PECAM-1. J. Clin. Invest. 99, 3–8.7. Springer, T. A. (1990) Adhesion receptors of the immune system. Nature
346, 425–434.8. Aida, Y., Pabst, M. J. (1991) Neutrophil responses to lipopolysaccharide.
Effect of adherence on triggering and priming of the respiratory burst.J. Immunol. 146, 1271–1276.
9. Hynes, R. O. (1992) Integrins: versatility, modulation, and signaling incell adhesion. Cell 69, 11–25.
10. Jaconi, M. E., Theler, J. M., Schlegel, W., Appel, R. D., Wright, S. D.,Lew, P. D. (1991) Multiple elevations of cytosolic-free Ca2� in humanneutrophils: initiation by adherence receptors of the integrin family. J. CellBiol. 112, 1249–1257.
11. Lofgren, R., Ng-Sikorski, J., Sjolander, A., Andersson, T. (1993) Beta 2integrin engagement triggers actin polymerization and phosphatidylinosi-tol trisphosphate formation in non-adherent human neutrophils. J. CellBiol. 123, 1597–1605.
12. Grundy, J. E., Lawson, K. M., MacCormac, L. P., Fletcher, J. M., Yong,K. L. (1998) Cytomegalovirus-infected endothelial cells recruit neutro-phils by the secretion of C-X-C chemokines and transmit virus by directneutrophil-endothelial cell contact and during neutrophil transendothelialmigration. J. Infect. Dis. 177, 1465–1474.
522 Journal of Leukocyte Biology Volume 73, April 2003 http://www.jleukbio.org

13. Guthrie, L. A., McPhail, L. C., Henson, P. M., Johnston Jr., R. B. (1984)Priming of neutrophils for enhanced release of oxygen metabolites bybacterial lipopolysaccharide. Evidence for increased activity of the super-oxide-producing enzyme. J. Exp. Med. 160, 1656–1671.
14. Ingraham, L. M., Lafuze, J. E., Boxer, L. A., Baehner, R. L. (1987) In vitroand in vivo effects of treatment by platelet-activating factor on N-formyl-met-leu-phe-mediated responses of polymorphonuclear leucocytes. Br. J.Haematol. 66, 219–225.
15. Vercellotti, G. M., Yin, H. Q., Gustafson, K. S., Nelson, R. D., Jacob, H. S.(1988) Platelet-activating factor primes neutrophil responses to agonists:role in promoting neutrophil-mediated endothelial damage. Blood 71,1100–1107.
16. Silliman, C. C., Clay, K. L., Thurman, G. W., Johnson, C. A., Ambruso,D. R. (1994) Partial characterization of lipids that develop during theroutine storage of blood and prime the neutrophil NADPH oxidase. J. Lab.Clin. Med. 124, 684–694.
17. Silliman, C. C., Dickey, W. O., Paterson, A. J., Thurman, G. W., Clay,K. L., Johnson, C. A., Ambruso, D. R. (1996) Analysis of the primingactivity of lipids generated during routine storage of platelet concentrates.Transfusion 36, 133–139.
18. Silliman, C. C., Paterson, A. J., Dickey, W. O., Stroneck, D. F., Popovsky,M. A., Caldwell, S. A., Ambruso, D. R. (1997) The association of biolog-ically active lipids with the development of transfusion-related acute lunginjury: a retrospective study. Transfusion 37, 719–726.
19. Silliman, C. C., Voelkel, N. F., Allard, J. D., Elzi, D. J., Tuder, R. M.,Johnson, J. L., Ambruso, D. R. (1998) Plasma and lipids from storedpacked red blood cells cause acute lung injury in an animal model. J. Clin.Invest. 101, 1458–1467.
20. Arnoux, B., Denjean, A., Page, C. P., Nolibe, D., Morley, J., Benveniste,J. (1988) Accumulation of platelets and eosinophils in baboon lung afterpaf-acether challenge. Inhibition by ketotifen. Am. Rev. Respir. Dis. 137,855–860.
21. Humphrey, D. M., Hanahan, D. J., Pinckard, R. N. (1982) Induction ofleukocytic infiltrates in rabbit skin by acetyl glyceryl ether phosphoryl-choline. Lab. Invest. 47, 227–234.
22. Korth, R. M. (1997) Specific binding sites for 1-O-alkyl-sn-glyceryl-3-phosphorylcholine on intact human blood neutrophils. Int. Arch. AllergyImmunol. 113, 460–464.
23. O’Flaherty, J. T., Wykle, R. L., Miller, C. H., Lewis, J. C., Waite, M., Bass,D. A., McCall, C. E., DeChatelet, L. R. (1981) 1-O-Alkyl-sn-glyceryl-3-phosphorylcholines: a novel class of neutrophil stimulants. Am. J. Pathol.103, 70–79.
24. Quinn, M. T., Parthasarathy, S., Steinberg, D. (1988) Lysophosphatidyl-choline: a chemotactic factor for human monocytes and its potential rolein atherogenesis. Proc. Natl. Acad. Sci. USA 85, 2805–2809.
25. Elzi, D. J., Hiester, A. A., Silliman, C. C. (1997) Receptor-mediatedcalcium entry is required for maximal effects of platelet activating factorprimed responses in human neutrophils. Biochem. Biophys. Res. Commun.240, 763–765.
26. Technical Manual, 14th ed. Bethesda, MD, American Association ofBlood Banks, 737–750.
27. Silliman, C. C., Cusack, N. A., Swanson, N. J., Ghaffarifar, S., Ambruso,D. R. (1995) Platelets and neutrophils from healthy term neonates exhibitincreased levels of immunoglobulins. Pediatr. Res. 38, 993–997.
28. Grynkiewicz, G., Poenie, M., Tsien, R. Y. (1985) A new generation ofCa2� indicators with greatly improved fluorescence properties. J. Biol.Chem. 260, 3440–3450.
29. Elzi, D. J., Bjornsen, A. J., MacKenzie, T., Wyman, T. H., Silliman, C. C.(2001) Ionomycin causes activation of p38 and p42/44 mitogen-activatedprotein kinases in human neutrophils. Am. J. Physiol. Cell Physiol. 281,C350–C360.
30. Hanlon, W. A., Stolk, J., Davies, P., Humes, J. L., Mumford, R.,Bonney, R. J. (1991) rTNF alpha facilitates human polymorphonuclearleukocyte adherence to fibrinogen matrices with mobilization of spe-cific and tertiary but not azurophilic granule markers. J. Leukoc. Biol.50, 43– 48.
31. Metzner, B., Heger, M., Hofmann, C., Czech, W., Norgauer, J. (1997)Evidence for the involvement of phosphatidylinositol 4,5-bisphosphate 3-kinase in CD18-mediated adhesion of human neutrophils to fibrinogen.Biochem. Biophys. Res. Commun. 232, 719–723.
32. Barnett Jr., C. C., Moore, E. E., Mierau, G. W., Partrick, D. A., Biffl, W. L.,Elzi, D. J., Silliman, C. C. (1998) ICAM-1-CD18 interaction mediatesneutrophil cytotoxicity through protease release. Am. J. Physiol. 274,C1634–C1644.
33. Ward, P. A., Mazera, E. G. (1980) Leukocyte chemotaxis. In Manual ofClinical Immunology (N. R. Rose, H. Friedman, eds.), Washington, DC,American Society for Microbiology, 261–266.
34. Ambruso, D. R., Bentwood, B., Henson, P. M., Johnston Jr., R. B. (1984)Oxidative metabolism of cord blood neutrophils: relationship to contentand degranulation of cytoplasmic granules. Pediatr. Res. 18, 1148–1153.
35. Peterson, V. M., Ambruso, D. R., Emmett, M., Bartle, E. J. (1988)Inhibition of colony-stimulating factor (CSF) production by postburn se-rum: negative feedback inhibition mediated by lactoferrin. J. Trauma 28,1533–1540.
36. Voller, A., Bidwell, D. E., Bartlett, A. (1976) Enzyme immunoassays indiagnostic medicine. Theory and practice. Bull. World Health Organ. 53,55–65.
37. Clay, K. L., Murphy, R. C., Andres, J. L., Lynch, J., Henson, P. M. (1984)Structure elucidation of platelet activating factor derived from humanneutrophils. Biochem. Biophys. Res. Commun. 121, 815–825.
38. Clay, K. L., Johnson, C., Worthen, G. S. (1991) Biosynthesis of plateletactivating factor and 1-O-acyl analogues by endothelial cells. Biochim.Biophys. Acta 1094, 43–50.
39. Silliman, C. C., Johnson, C. A., Clay, K. L., Thurman, G. W., Ambruso,D. R. (1993) Compounds biologically similar to platelet activating factorare present in stored blood components. Lipids 28, 415–418.
40. Heuer, H. O., Casals-Stenzel, J., Muacevic, G., Weber, K. H. (1990)Pharmacologic activity of bepafant (WEB 2170), a new and selectivehetrazepinoic antagonist of platelet activating factor. J. Pharmacol. Exp.Ther. 255, 962–968.
41. Heuer, H. O. (1991) WEB 2347: pharmacology of a new very potent andlong acting hetrazepinoic PAF-antagonist and its action in repeatedlysensitized guinea-pigs. J. Lipid Mediat. 4, 39–44.
42. Sheppard, G. S., Pireh, D., Carrera Jr., G. M., Bures, M. G., Heyman,H. R., Steinman, D. H., Davidsen, S. K., Phillips, J. G., Guinn, D. E., May,P. D. (1994) 3-(2-(3-Pyridinyl)thiazolidin-4-oyl)indoles, a novel series ofplatelet activating factor antagonists. J. Med. Chem. 37, 2011–2032.
43. Korth, R. M., Hirafuji, M., Benveniste, J., Russo-Marie, F. (1995)Human umbilical vein endothelial cells: specific binding of platelet-activating factor and cytosolic calcium flux. Biochem. Pharmacol. 49,1793–1799.
44. Corey, S. J., Rosoff, P. M. (1991) Unsaturated fatty acids and lipoxygenaseproducts regulate phagocytic NADPH oxidase activity by a nondetergentmechanism. J. Lab. Clin. Med. 118, 343–351.
45. Englberger, W., Bitter-Suermann, D., Hadding, U. (1987) Influence oflysophospholipids and PAF on the oxidative burst of PMNL. Int. J. Im-munopharmacol. 9, 275–282.
46. Ginsburg, I., Ward, P. A., Varani, J. (1989) Lysophosphatides enhancesuperoxide responses of stimulated human neutrophils. Inflammation 13,163–174.
47. Savage, J. E., Theron, A. J., Anderson, R. (1993) Activation of neutrophilmembrane-associated oxidative metabolism by ultraviolet radiation. J. In-vest. Dermatol. 101, 532–536.
48. Worthen, G. S., Seccombe, J. F., Clay, K. L., Guthrie, L. A., Johnston Jr.,R. B. (1988) The priming of neutrophils by lipopolysaccharide for pro-duction of intracellular platelet-activating factor. Potential role in medi-ation of enhanced superoxide secretion. J. Immunol. 140, 3553–3559.
49. Aiboshi, J., Moore, E. E., Ciesla, D. J., Silliman, C. C. (2001) Bloodtransfusion and the two-insult model of post-injury multiple organ failure.Shock 15, 302–306.
50. Baggiolini, M., Moser, B. (1997) Blocking chemokine receptors. J. Exp.Med. 186, 1189–1191.
51. Gong, J. H., Uguccioni, M., Dewald, B., Baggiolini, M., Clark-Lewis, I.(1996) RANTES and MCP-3 antagonists bind multiple chemokine recep-tors. J. Biol. Chem. 271, 10521–10527.
52. Pinckard, R. N., Showell, H. J., Castillo, R., Lear, C., Breslow, R.,McManus, L. M., Woodard, D. S., Ludwig, J. C. (1992) Differentialresponsiveness of human neutrophils to the autocrine actions of 1-O-alkyl-homologs and 1-acyl analogs of platelet-activating factor. J. Immunol.148, 3528–3535.
53. Puzon-McLaughlin, W., Kamata, T., Takada, Y. (2000) Multiple discon-tinuous ligand-mimetic antibody binding sites define a ligand bindingpocket in integrin alpha(IIb)beta(3). J. Biol. Chem. 275, 7795–7802.
54. Gay, J. C., Beckman, J. K., Brash, A. R., Oates, J. A., Lukens, J. N. (1984)Enhancement of chemotactic factor-stimulated neutrophil oxidative me-tabolism by leukotriene B4. Blood 64, 780–785.
55. Lobos, E. A., Sharon, P., Stenson, W. F. (1987) Chemotactic activity ininflammatory bowel disease. Role of leukotriene B4. Dig. Dis. Sci. 32,1380–1388.
56. Bochsler, P. N., Neilsen, N. R., Dean, D. F., Slauson, D. O. (1992)Stimulus-dependent actin polymerization in bovine neutrophils. Inflam-mation 16, 383–392.
57. Coates, T. D., Watts, R. G., Hartman, R., Howard, T. H. (1992) Relation-
Silliman et al. Lyso-PCs affect multiple PMN functions 523

ship of F-actin distribution to development of polar shape in humanpolymorphonuclear neutrophils. J. Cell Biol. 117, 765–774.
58. Shirley, M. A., Reidhead, C. T., Murphy, R. C. (1992) Chemotactic LTB4metabolites produced by hepatocytes in the presence of ethanol. Biochem.Biophys. Res. Commun. 185, 604–610.
59. Hla, T., Lee, M. J., Ancellin, N., Paik, J. H., Kluk, M. J. (2001) Lyso-phospholipids–receptor revelations. Science 294, 1875–1878.
60. Le, L. Q., Kabarowski, J. H., Weng, Z., Satterthwaite, A. B., Harvill, E. T.,Jensen, E. R., Miller, J. F., Witte, O. N. (2001) Mice lacking the orphanG protein-coupled receptor G2A develop a late-onset autoimmune syn-drome. Immunity 14, 561–571.
61. Zhu, K., Baudhuin, L. M., Hong, G., Williams, F. S., Cristina, K. L.,Kabarowski, J. H., Witte, O. N., Xu, Y. (2001) Sphingosylphosphorylcho-line and lysophosphatidylcholine are ligands for the G protein-coupledreceptor GPR4. J. Biol. Chem. 276, 41325–41335.
62. Bergmann, S. R., Ferguson Jr., T. B., Sobel, B. E. (1981) Effects ofamphiphiles on erythrocytes, coronary arteries, and perfused hearts.Am. J. Physiol. 240, H229–H237.
63. Kume, N., Cybulsky, M. I., Gimbrone Jr., M. A. (1992) Lysophosphati-dylcholine, a component of atherogenic lipoproteins, induces mononuclearleukocyte adhesion molecules in cultured human and rabbit arterialendothelial cells. J. Clin. Invest. 90, 1138–1144.
64. Kume, N., Gimbrone Jr., M. A. (1994) Lysophosphatidylcholine transcrip-tionally induces growth factor gene expression in cultured human endo-thelial cells. J. Clin. Invest. 93, 907–911.
65. Sugiyama, S., Kugiyama, K., Ohgushi, M., Fujimoto, K., Yasue, H. (1994)Lysophosphatidylcholine in oxidized low-density lipoprotein increasesendothelial susceptibility to polymorphonuclear leukocyte-induced endo-thelial dysfunction in porcine coronary arteries. Role of protein kinase C.Circ. Res. 74, 565–575.
524 Journal of Leukocyte Biology Volume 73, April 2003 http://www.jleukbio.org
Copyright © 2022 FDOKUMEN




















